Effects of nitrogen addition on soil respiration of Rhododendron simsii shrubland in the subtropical mountainous areas of China
ZHANGQiang1,2, LIJia-Xiang3, XIEZong-Qiang1,*, 1State Key Laboratory of Vegetation and Environment Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2University of Chinese Academy of Sciences, Beijing 100049, Chinaand 3Faculty of Forestry, Central South University of Forestry and Technology, Changsha 410004, China 通讯作者:* 通信作者Author for correspondence (E-mail: xie@ibcas.ac.cn) 责任编辑:ZHANGQiangLIJia-XiangXIEZong-Qiang 收稿日期:2015-08-17 接受日期:2016-01-17 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略性先导科技专项(XDA- 05050302)
关键词:土壤呼吸的温度敏感性(Q10);土壤温度;土壤含水量;根系生物量;凋落物生物量 Abstract Aims As the second largest C flux between the atmosphere and terrestrial ecosystems, soil respiration plays a vital role in regulating atmosphere CO2 concentration. Therefore, understanding the response of soil respiration to the increasing nitrogen deposition is urgently needed for prediction of future climate change. However, it is still unclear how nitrogen deposition influences soil respiration of shrubland in subtropical China. Our objectives were to explore the effects of different levels of nitrogen fertilization on soil respiration, root biomass increment, and litter biomass, and to analyze the relationships between soil respiration and soil temperature and moisture. Methods From January 2013 to September 2014, we conducted a short-term simulated nitrogen deposition experiment in the Rhododendron simsii shrubland of Dawei Mountain, located in Hunan Province, southern China. Four levels of nitrogen addition treatments (each level with three replicates) were established: control (CK, no nitrogen addition), low nitrogen addition (LN, 2 g·m-2·a-1), medium nitrogen addition (MN, 5 g·m-2·a-1) and high nitrogen addition (HN, 10 g·m-2·a-1). Soil respiration was measured by LI-8100 soil CO2 efflux system. At the same time, we measured root biomass increment and litter biomass in each plot. Important findings Soil respiration exhibited a strong seasonal pattern, with the highest rates found in summer and the lowest rates in winter. Annual accumulative soil respiration rate in the CK, LN, MN and HN was (2.37 ± 0.39), (2.79 ± 0.42), (2.26 ± 0.38) and (2.30 ± 0.36) kg CO2·m-2, respectively. Annual mean soil respiration rate in the CK, LN, MN and HN was (1.71 ± 0.28), (2.01 ± 0.30), (1.63 ± 0.27) and (1.66 ± 0.26) μmol CO2·m-2·s-1, respectively, and it was 17.25% higher in the LN treatment compared with CK (p = 0.06). The root biomass increment was increased by LN, MN, and HN treatments by 18.36%, 36.49% and 61.63%, respectively, compared to CK. The litter biomass was increased by LN, MN, and HN treatments by 35.87%, 22.17% and 15.35%, respectively, compared with CK. Soil respiration exhibited a significant exponential relationship with soil temperature (p < 0.01, R2 is 0.77 to 0.82) and a significant linear relationship with soil moisture at the depth of 5 cm (p < 0.05, R2 is 0.10 to 0.15). The temperature sensitivity (Q10) value of CK, LN, MN and HN plots was 3.96, 3.60, 3.71 and 3.51, respectively. These results suggested that nitrogen addition promoted plant growth and decreased the temperature sensitivity of soil respiration. The increase of root biomass under N addition may be an important reason for the change of soil respiration in the study area.
在2013年1月-11月和2014年3月-9月, CK、LN、MN和HN处理下土壤呼吸的月平均速率均呈现出明显的季节动态, 且变化趋势相同。夏季7、8月份土壤呼吸速率达到最高(最大值分别为3.17、3.49、2.71和3.09 μmol CO2·m-2·s-1), 此时各处理之间的差异较明显; 冬季11月-1月份土壤呼吸速率达到最低(最小值分别为0.44、0.65、0.54和0.55 μmol CO2·m-2·s-1), 此时各处理之间的差异较小(图3)。LN处理样地的各月土壤呼吸速率均高于其他样地。 显示原图|下载原图ZIP|生成PPT 图3不同氮添加浓度下土壤呼吸的季节动态(平均值±标准误差)。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。 -->Fig. 3Seasonal variations of soil respiration under different nitrogen treatments (mean ± SE). CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively. -->
根据对照样地2013年的土壤呼吸值计算, 研究区域年均土壤呼吸速率为(1.71 ± 0.28) μmol CO2·m-2·s-1, 相当于每年释放CO2 (2.37 ± 0.39) kg·m-2。4种氮处理中, LN处理的土壤呼吸速率在2013年生长季和2014年生长季均高于对照样地, CO2排放量分别增加了15.51%和11.25% (图4)。MN 和HN处理的土壤呼吸比对照样地低。进行重复测量的方差分析显示, 氮处理对土壤呼吸的影响在2013年全年和2013年生长季均接近显著水平(p值分别为0.06和0.08), 在2014年生长季不显著(p = 0.27)。测定时间对土壤呼吸产生显著影响(p < 0.01), 而测定时间和氮添加的交互作用并不显著(2013和2014年生长季的p值分别为0.73和0.58)。 显示原图|下载原图ZIP|生成PPT 图4不同氮添加浓度下的生长季土壤呼吸速率(平均值±标准误差)。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。 -->Fig. 4Soil respiration rate in the growing season under different nitrogen treatments (mean ± SE). CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively. -->
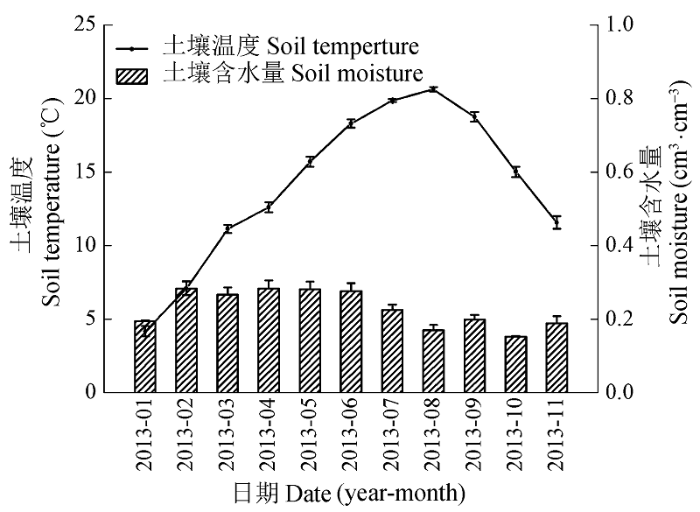
2.4 土壤呼吸与土壤温度和土壤含水量的关系
不同氮处理下, 杜鹃灌丛的土壤呼吸与5 cm深土壤温度均呈极显著的指数相关关系(p < 0.01)(图5), 土壤温度可以解释土壤呼吸变化的76.53%- 82.21%, CK、LN、MN和HN处理下的Q10值分别为3.96、3.60、3.71和3.51。与对照样地相比, 施氮降低了Q10。土壤呼吸与5 cm深土壤含水量呈显著线性相关关系(p < 0.05)(图6), 土壤含水量可以解释土壤呼吸变化的9.93%-15.21%。与土壤温度相比, 土壤含水量对土壤呼吸变化的解释率较低。氮处理下土壤温度与土壤呼吸的相关性与对照相比有所下降, 而土壤含水量与土壤呼吸的相关性与对照相比有上升的趋势。 显示原图|下载原图ZIP|生成PPT 图5不同氮添加水平下5 cm深度土壤呼吸与土壤温度的关系。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。Q10, 土壤呼吸的温度敏感性。 -->Fig. 5Relationships between soil respiration rate and soil temperature at 5 cm soil depth under different nitrogen treatments. CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively. Q10, temperature sensitivity. -->
显示原图|下载原图ZIP|生成PPT 图6不同氮添加水平下5 cm深度土壤呼吸与土壤含水量的关系。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。 -->Fig. 6Relationships between soil respiration rate and soil moisture at 5 cm soil depth under different nitrogen treatments. CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively. -->
研究结果显示, 不同氮处理下的土壤呼吸呈现出相似的季节变化模式, 在夏季达到最大, 冬季降到最低, 这与在中国亚热带其他地区得到的研究结果相符(Tang et al., 2006; Mo et al., 2007)。土壤呼吸的季节变化主要受到温度等气候因子和植物生长的调控影响(韩广轩和周广胜, 2009)。2013年在对照样地测得的杜鹃灌丛年CO2释放量为(2.37 ± 0.39) kg·m-2, 低于Deng等(2010)在中国南亚热带森林测得的值((3.53 ± 0.03) kg·m-2), 但高于中国暖温带森林的年CO2释放量((1.50 ± 0.03) kg·m-2, Sun et al., 2014)。试验结果显示, 低氮处理对土壤呼吸有促进作用, 而中氮和高氮处理则有轻微的抑制效应。综合前人的研究可知, 研究区域和施氮浓度不同, 土壤呼吸的响应也不同, 这受到土壤本身养分状况的影响(Mo et al., 2007)。在氮含量丰富的环境中氮添加会抑制土壤呼吸, 而在氮限制的地区氮添加则会促进土壤呼吸(Hyvonen et al., 2007; Cusack et al., 2011)。 一般来说, 将土壤呼吸分为自养呼吸(根系和根际微生物呼吸)和异养呼吸(微生物和土壤动物呼吸)两个组分(Sotta et al., 2004), 土壤总呼吸对氮添加的反应是自养呼吸、异养呼吸反应的综合效应。在全球范围内的森林生态系统中, 自养呼吸约占土壤总呼吸的45.8% (Hanson et al., 2000)。Zhou等(2014)对来自不同生态系统的295个研究结果进行整合分析, 结果显示自养呼吸对氮添加的反应与根系生物量密切相关(R2 = 0.803)。Rodeghiero和Cescatti (2006)对来自7种不同森林生态系统的数据进行研究, 也发现土壤呼吸与根系生物量之间存在着线性回归关系。所以, 氮添加对根系生物量的影响将会很大程度上影响植物根呼吸和与之相关的根际微生物呼吸。本试验中, LN、MN和HN处理样地的根系生物量年增量与对照样地相比, 分别高出了18.36%、36.49%和61.63%。可以看出, 随着氮添加浓度的升高, 根系生物量年增量呈现出递增的趋势, 因此会导致植物自养呼吸也随之上升。土壤异养呼吸来源于土壤中有机物质的分解, 主要依赖于分解底物的质量和数量、胞外酶活性、微生物生物量和微生物活性(Ryan & Law, 2005)。Janssens等(2010)和Zhou等(2014)利用大量研究结果进行的整合分析显示, 氮添加与异养呼吸呈负相关关系, 特别是在生产力高、氮素不受限制的生态系统中, 这种负相关性更明显。Egerton-Warburton和Allen (2000)在加利福尼亚沿海灌丛林中进行的试验表明, 氮肥会降低菌根感染率和存活率。Rühling和Tyler (1991)观察到氮添加会使菌根真菌几乎停止产生子实体。研究发现, 在目前的氮添加浓度(通常大于5 kg·hm-2·a-1)下, 凋落物的分解经常是被抑制的(Knorr et al., 2005), 主要是通过腐生性营养的微生物群落组成的改变(Gallo et al., 2004)和酶活性的改变(Frey et al., 2004)。在本研究中, 作为微生物分解的底物, 凋落物在氮处理样地中的生物量要高于对照样地(图3), 分别比对照样地高出35.87%、22.17%和15.35%。但是, 氮添加在使地上部分凋落物增加的同时, 也可能导致微生物生物量和活性的下降, 从而在一定程度上限制凋落物的分解(Burton et al., 2004), 这也许会使得异养呼吸有所降低。另外, 有研究表明, 随着施氮时间的增加, 氮添加对凋落物分解的抑制作用也会增强(Fang et al., 2007)。综上可知, 自养呼吸和异养呼吸对氮添加的不同反应综合导致了土壤总呼吸对氮添加的响应。
3.3 不同氮处理下土壤呼吸与土壤温度和土壤含水量的关系
在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008)。土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子。在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量。当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005)。 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006)。本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009)。这可能与本研究区域海拔较高有关。罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感。本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似。一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低。呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010)。同时氮添加也会降低微生物酶活性(Ma et al., 2014)。这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低。但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010)。这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加。总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响。 致谢 感谢中南林业科技大学林学院的老师和学生在样品采集、样品处理和野外监测工作中给予的帮助。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
CraineJM, WedinDA, ReichPB (2001). Grassland species effects on soil CO2 flux track the effects of elevated CO2 and nitrogen .New Phytologist, 150, 425-434. [本文引用: 1]
[6]
CusackDF, SilverWL, TornMS, McDowellWH (2011). Effects of nitrogen additions on above- and below-ground carbon dynamics in two tropical forests .Biogeochemistry, 104, 203-225. [本文引用: 1]
[7]
DavidsonEA, JanssensIA (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change .Nature, 440, 165-173. [本文引用: 2]
[8]
DenmanKL, BrasseurG, ChidthaisongA, CiaisP, CoxPM, DickinsonRE, HauglustaineD, HeinzeC, HollandE, JacobD, LohmannU, amachandran S, da Silva Dias PL, Wofsy SC, Zhang X (2007). Couplings between changes in the climate system and biogeochemistry . In: Solomon S, Qin D, Manning M, Marquis M, Averyt K, Tignor MMB, Miller HLR, Chen Z eds. Climate Change 2007, The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. RCambridge University Press, Cambridge, UK. [本文引用: 1]
[9]
DengQ, ZhouG, LiuJ, LiuS, DuanH, ZhangD (2010). Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China .Biogeosciences, 7, 315-328. [本文引用: 5]
[10]
DilustroJJ, CollinsB, DuncanL, CrawfordC (2005). Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests .Forest Ecology and Management, 204, 85-95. [本文引用: 1]
[11]
DuanHL, LiuJX, DengQ, ChenXM, ZhangDQ (2009). Effects of elevated CO2 and N deposition on plant biomass accumulation and allocation in subtropical forest ecosystems: A mesocosm study .Journal of Plant Ecology (Chinese Version), 33, 570-579. (in Chinese with English abstract)[段洪浪, 刘菊秀, 邓琦, 陈小梅, 张德强 (2009). CO2浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响 . 植物生态学报, 33, 570-579.] [本文引用: 1]
[12]
Egerton-WarburtonL, AllenE (2000). Shifts in the diversity of arbuscular mycorrhizal fungi along an anthropogenic nitrogen gradient .Ecological Applications, 10, 484-496.
[13]
ElserJJ, AndersenT, BaronJS, BergstroemA-K, JanssonM, KyleM, NydickKR, StegerL, HessenDO (2009). Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition .Science, 326, 835-837. [本文引用: 1]
[14]
FangH, MoJM, PengSL, LiZA, WangH (2007). Cumulative effects of nitrogen additions on litter decomposition in three tropical forests in southern China .Plant and Soil, 297, 233-242. [本文引用: 1]
[15]
ForsterP, RamaswamyV, ArtaxoP, BerntsenT, BettsR, FaheyDW, HaywoodJ, LeanJ, LoweDC, MyhreG, NgangaJ, PrinnR, RagaG, Schulz M, van Dorland R (2007). Changes in atmospheric constituents and in radiative forcing . In: Solomon S, Qin D, Manning M, Marquis M, Averyt K, Tignor MMB, Miller HLR, Chen Z eds. Climate Change 2007, The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. [本文引用: 1]
[16]
FreySD, KnorrM, ParrentJL, SimpsonRT (2004). Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests .Forest Ecology and Management, 196, 159-171. [本文引用: 1]
[17]
GalloM, AmonetteR, LauberC, SinsabaughRL, ZakDR (2004). Microbial community structure and oxidative enzyme activity in nitrogen-amended north temperate forest soils .Microbial Ecology, 48, 218-229. [本文引用: 1]
[18]
HanGX, ZhouGS (2009). Review of spatial and temporal variations of soil respiration and driving mechanisms .Chinese Journal of Plant Ecology, 33, 197-205. (in Chinese with English abstract)[韩广轩, 周广胜 (2009). 土壤呼吸作用时空动态变化及其影响机制研究与展望 . 植物生态学报, 33, 197-205.] [本文引用: 1]
[19]
HansonPJ, EdwardsNT, GartenCT, AndrewsJA (2000). Separating root and soil microbial contributions to soil respiration: A review of methods and observations .Biogeochemistry, 48, 115-146. [本文引用: 1]
HögbergP, FanHB, QuistM, BinkleyD, TammCO (2006). Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest .Global Change Biology, 12, 489-499. [本文引用: 1]
[22]
HuHF, WangZH, GH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Eco- logy (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 1]
[23]
HyvonenR, AgrenGI, LinderS, PerssonT, CotrufoMF, EkbladA, FreemanM, GrelleA, JanssensIA, JarvisPG, KellomakiS, LindrothA, LoustauD, LundmarkT, NorbyRJ, OrenR, PilegaardK, RyanMG, SigurdssonBD, StromgrenM, van OijenM, WallinG (2007). The likely impact of elevated CO2, nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review .New Phytologist, 173, 463-480. [本文引用: 1]
[24]
HyvonenR, PerssonT, AnderssonS, OlssonB, AgrenGI, LinderS (2008). Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe .Biogeochemistry, 89, 121-137. [本文引用: 1]
[25]
IPCC (Intergovernmental Panel on Climate Change) (2013). Contribution of working group 1 to the fifth assessment report of the intergovernmental panel on climate change . In: Stocker TF, Qin DH, Plattner GK, Tignor MMB, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM eds. Climate Change 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK. [本文引用: 2]
KnorrM, FreySD, CurtisPS (2005). Nitrogen additions and litter decomposition: A meta-analysis .Ecology, 86, 3252-3257.
[28]
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed .Ecology, 89, 371-379. [本文引用: 2]
[29]
LiJX, ZhangX, XieZQ, LuCF, TuXY, XunY (2015). Community composition and structure ofRhododendron simsii shrubland in the Dawei Mountain, Hunan Province . Biodiversity Science, 23, 815-823. (in Chinese with English abstract)[李家湘, 张旭, 谢宗强, 卢从发, 涂向阳, 寻院 (2015). 湖南大围山杜鹃灌丛的群落组成及结构特征 . 生物多样性, 23, 815-823.] [本文引用: 1]
[30]
LiuX, DuanL, MoJ, DuE, ShenJ, LuX, ZhangY, ZhouX, HeC, ZhangF (2011). Nitrogen deposition and its ecological impact in China: An overview .Environmental Pollution, 159, 2251-2264.
[31]
LueCQ, TianHQ (2007). Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data . Journal of Geophysical Research-Atmospheres, 112, D22S05, doi: 10.1029/2006JD007990. [本文引用: 2]
[32]
LuoL, ShengGZ, XieZQ, ZhouLG (2011). Components of soil respiration and its temperature sensitivity in four types of forests along an elevational gradient in Shennongjia, China .Chinese Journal of Plant Ecology, 35, 722-730. (in Chinese with English abstract)[罗璐, 申国珍, 谢宗强, 周利光 (2011). 神农架海拔梯度上4种典型森林的土壤呼吸组分及其对温度的敏感性 . 植物生态学报, 35, 722-730.] [本文引用: 1]
[33]
LuoYQ, HuiDF, ZhangDQ (2006). Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: A meta-analysis .Ecology, 87, 53-63. [本文引用: 1]
[34]
MaYC, ZhuB, SunZZ, ZhaoC, YangY, PiaoSL (2014). The effects of simulated nitrogen deposition on extracellular enzyme activities of litter and soil among different-aged stands of larch .Journal of Plant Ecology, 7, 240-249. [本文引用: 1]
[35]
MaskellLC, SmartSM, BullockJM, ThompsonK, StevensCJ (2010). Nitrogen deposition causes widespread loss of species richness in British habitats .Global Change Biology, 16, 671-679. [本文引用: 1]
[36]
MelilloJM, SteudlerPA, AberJD, NewkirkK, LuxH, BowlesFP, CatricalaC, MagillA, AhrensT, MorrisseauS (2002). Soil warming and carbon-cycle feedbacks to the climate system .Science, 298, 2173-2176. [本文引用: 1]
[37]
MoJM, BrownS, XueJH, FangYT, LiZA (2006).Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China . Plant and Soil, 282, 135-151. [本文引用: 3]
[38]
MoJM, ZhangW, ZhuWX, FangYT, LiDJ, ZhaoP (2007). Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China .Plant and Soil, 296, 125-135. [本文引用: 2]
[39]
MoJM, ZhangW, ZhuWX, GundersenP, FangYT, LiDJ, WangH (2008). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China .Global Change Biology, 14, 403-412. [本文引用: 1]
[40]
MoscatelliMC, LagornarsinoA, de AngelisP, GregoS (2008). Short and mediumterm contrasting effects of nitrogen fertilization on C and N cycling in a poplar plantation soil .Forest Ecology and Management, 255, 447-454. [本文引用: 1]
[41]
PendallE, BridghamS, HansonPJ, HungateB, KicklighterDW, JohnsonDW, LawBE, LuoYQ, MegonigalJP, OlsrudM, RyanMG, WanSQ (2004). Belowground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods, and models .New Phytologist, 162, 311-322.
[42]
PengSS, PiaoSL, WangT, SunJY, ShenZH (2009). Temperature sensitivity of soil respiration in different ecosystems in China .Soil Biology & Biochemistry, 41, 1008-1014. [本文引用: 2]
RaichJW, PotterCS, BhagawatiD (2002). Interannual varia- bility in global soil respiration, 1980-1994 .Global Change Biology, 8, 800-812.
[45]
RodeghieroM, CescattiA (2006). Indirect partitioning of soil respiration in a series of evergreen forest ecosystems .Plant and Soil, 284, 7-22.
[46]
RühlingÅ, TylerG (1991). Effects of simulated nitrogen deposition to the forest floor on the macrofungal flora of a beech forest .Ambio, 20, 261-263. [本文引用: 1]
SchlesingerWH, AndrewsJA (2000). Soil respiration and the global carbon cycle .Biogeochemistry, 48, 7-20. [本文引用: 1]
[49]
SottaED, MeirP, MalhiY, NobreAD, HodnettM, GraceJ (2004). Soil CO2 efflux in a tropical forest in the central Amazon .Global Change Biology, 10, 601-617. [本文引用: 1]
[50]
SunZZ, LiuLL, MaYC, YinGD, ZhaoC, ZhangY, PiaoSL (2014). The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil .Agricultural and Forest Meteorology, 197, 103-110. [本文引用: 2]
[51]
TangXL, LiuSG, ZhouGY, ZhangDQ, ZhouCY (2006). Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China .Global Change Biology, 12, 546-560. [本文引用: 1]
[52]
TarnocaiC, CanadellJG, SchuurEAG, KuhryP, MazhitovaG, ZimovS (2009). Soil organic carbon pools in the northern circumpolar permafrost region . Global Biogeochemical Cycles, 23, GB2023, doi: 10.1029/2008GB003327. [本文引用: 1]
[53]
ThomasRQ, CanhamCD, WeathersKC, GoodaleCL (2010). Increased tree carbon storage in response to nitrogen deposition in the US .Nature Geoscience, 3, 13-17. [本文引用: 1]
[54]
TuLH, HuTX, ZhangJ, LiXW, HuHL, LiuL, XiaoYL (2013). Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem .Soil Biology & Biochemistry, 58, 255-264. [本文引用: 1]
[55]
XuWH, WanSQ (2008). Water- and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China .Soil Biology & Biochemistry, 40, 679-687. [本文引用: 1]
[56]
ZhengSW, TangM, ZouJH, MuCL (2007). Summary of research on shrub biomass in China .Journal of Chengdu University (Natural Science Edition), 26, 189-192. (in Chinese with English abstract)[郑绍伟, 唐敏, 邹俊辉, 慕长龙 (2007). 灌木群落及生物量研究综述 . 成都大学学报(自然科学版), 26, 189-192.] [本文引用: 1]
[57]
ZhengXH, FuCB, XuXK, YanXD, HuangY, HanSH, HuF, ChenGX (2002). The Asian nitrogen cycle case study .AMBIO, 31, 79-87. [本文引用: 1]
[58]
ZhengZM, YuGR, FuYL, WangYS, SunXM, WangYH (2009). Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content: A trans-China based case study .Soil Biology & Biochemistry, 41, 1531-1540.
[59]
ZhouLY, ZhouXH, ZhangBC, LuM, LuoYQ, LiuLL, LiB (2014). Different responses of soil respiration and its components to nitrogen addition among biomes: A meta- analysis .Global Change Biology, 20, 2332-2343.
Nitrogen saturation in northern forest ecosystems 1 1989
Effects of nitrogen additions on above- and below-ground carbon dynamics in two tropical forests 1 2011
... 研究结果显示, 不同氮处理下的土壤呼吸呈现出相似的季节变化模式, 在夏季达到最大, 冬季降到最低, 这与在中国亚热带其他地区得到的研究结果相符(Tang et al., 2006; Mo et al., 2007).土壤呼吸的季节变化主要受到温度等气候因子和植物生长的调控影响(韩广轩和周广胜, 2009).2013年在对照样地测得的杜鹃灌丛年CO2释放量为(2.37 ± 0.39) kg·m-2, 低于Deng等(2010)在中国南亚热带森林测得的值((3.53 ± 0.03) kg·m-2), 但高于中国暖温带森林的年CO2释放量((1.50 ± 0.03) kg·m-2, Sun et al., 2014).试验结果显示, 低氮处理对土壤呼吸有促进作用, 而中氮和高氮处理则有轻微的抑制效应.综合前人的研究可知, 研究区域和施氮浓度不同, 土壤呼吸的响应也不同, 这受到土壤本身养分状况的影响(Mo et al., 2007).在氮含量丰富的环境中氮添加会抑制土壤呼吸, 而在氮限制的地区氮添加则会促进土壤呼吸(Hyvonen et al., 2007; Cusack et al., 2011). ...
Temperature sensitivity of soil carbon decomposition and feedbacks to climate change 2 2006
... 陆地生态系统的大部分碳储存于土壤中.已有的研究显示, 全球植物和土壤的碳储量分别为560 Pg (Forster et al., 2007)和>3300 Pg (Tarnocai et al., 2009).土壤呼吸是CO2从陆地生态系统返回到大气中的主要途径(Schlesinger & Andrews, 2000).CO2的主要来源是根系和根际微生物呼吸、土壤微生物和土壤动物呼吸.作为大气和陆地生态系统之间的第二大碳交换途径, 土壤呼吸每年向大气中释放的碳量高达68-98 Pg (Raich et al., 2002; Bond- Lamberty & Thomson, 2010), 在调节大气CO2浓度和地球的气候变化方面发挥着重要作用.因此, 土壤呼吸的轻微变化就会对全球的气候和环境产生剧烈的影响(Davidson & Janssens, 2006).导致全球变化的主要因子(如氮沉降的增加和CO2浓度的上升)在很大程度上影响了土壤呼吸速率(Bowden et al., 2004; Deng et al., 2010), 因此, 理解全球变化下土壤呼吸的反应机制对于预测未来的气候变化十分 必要. ...
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Couplings between changes in the climate system and biogeochemistry 1 2007
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China 5 2010
... 陆地生态系统的大部分碳储存于土壤中.已有的研究显示, 全球植物和土壤的碳储量分别为560 Pg (Forster et al., 2007)和>3300 Pg (Tarnocai et al., 2009).土壤呼吸是CO2从陆地生态系统返回到大气中的主要途径(Schlesinger & Andrews, 2000).CO2的主要来源是根系和根际微生物呼吸、土壤微生物和土壤动物呼吸.作为大气和陆地生态系统之间的第二大碳交换途径, 土壤呼吸每年向大气中释放的碳量高达68-98 Pg (Raich et al., 2002; Bond- Lamberty & Thomson, 2010), 在调节大气CO2浓度和地球的气候变化方面发挥着重要作用.因此, 土壤呼吸的轻微变化就会对全球的气候和环境产生剧烈的影响(Davidson & Janssens, 2006).导致全球变化的主要因子(如氮沉降的增加和CO2浓度的上升)在很大程度上影响了土壤呼吸速率(Bowden et al., 2004; Deng et al., 2010), 因此, 理解全球变化下土壤呼吸的反应机制对于预测未来的气候变化十分 必要. ...
... 在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008).土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子.在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量.当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005). ...
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
... ; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
... ).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests 1 2005
... 在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008).土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子.在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量.当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005). ...
CO2浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响 1 2009
... 在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008).土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子.在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量.当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005). ...
Shifts in the diversity of arbuscular mycorrhizal fungi along an anthropogenic nitrogen gradient 2000
Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition 1 2009
Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest 1 2006
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
The likely impact of elevated CO2, nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review 1 2007
... 研究结果显示, 不同氮处理下的土壤呼吸呈现出相似的季节变化模式, 在夏季达到最大, 冬季降到最低, 这与在中国亚热带其他地区得到的研究结果相符(Tang et al., 2006; Mo et al., 2007).土壤呼吸的季节变化主要受到温度等气候因子和植物生长的调控影响(韩广轩和周广胜, 2009).2013年在对照样地测得的杜鹃灌丛年CO2释放量为(2.37 ± 0.39) kg·m-2, 低于Deng等(2010)在中国南亚热带森林测得的值((3.53 ± 0.03) kg·m-2), 但高于中国暖温带森林的年CO2释放量((1.50 ± 0.03) kg·m-2, Sun et al., 2014).试验结果显示, 低氮处理对土壤呼吸有促进作用, 而中氮和高氮处理则有轻微的抑制效应.综合前人的研究可知, 研究区域和施氮浓度不同, 土壤呼吸的响应也不同, 这受到土壤本身养分状况的影响(Mo et al., 2007).在氮含量丰富的环境中氮添加会抑制土壤呼吸, 而在氮限制的地区氮添加则会促进土壤呼吸(Hyvonen et al., 2007; Cusack et al., 2011). ...
Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe 1 2008
Contribution of working group 1 to the fifth assessment report of the intergovernmental panel on climate change 2 2013
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Reduction of forest soil respiration in response to nitrogen deposition 1 2010
... 一般来说, 将土壤呼吸分为自养呼吸(根系和根际微生物呼吸)和异养呼吸(微生物和土壤动物呼吸)两个组分(Sotta et al., 2004), 土壤总呼吸对氮添加的反应是自养呼吸、异养呼吸反应的综合效应.在全球范围内的森林生态系统中, 自养呼吸约占土壤总呼吸的45.8% (Hanson et al., 2000).Zhou等(2014)对来自不同生态系统的295个研究结果进行整合分析, 结果显示自养呼吸对氮添加的反应与根系生物量密切相关(R2 = 0.803).Rodeghiero和Cescatti (2006)对来自7种不同森林生态系统的数据进行研究, 也发现土壤呼吸与根系生物量之间存在着线性回归关系.所以, 氮添加对根系生物量的影响将会很大程度上影响植物根呼吸和与之相关的根际微生物呼吸.本试验中, LN、MN和HN处理样地的根系生物量年增量与对照样地相比, 分别高出了18.36%、36.49%和61.63%.可以看出, 随着氮添加浓度的升高, 根系生物量年增量呈现出递增的趋势, 因此会导致植物自养呼吸也随之上升.土壤异养呼吸来源于土壤中有机物质的分解, 主要依赖于分解底物的质量和数量、胞外酶活性、微生物生物量和微生物活性(Ryan & Law, 2005).Janssens等(2010)和Zhou等(2014)利用大量研究结果进行的整合分析显示, 氮添加与异养呼吸呈负相关关系, 特别是在生产力高、氮素不受限制的生态系统中, 这种负相关性更明显.Egerton-Warburton和Allen (2000)在加利福尼亚沿海灌丛林中进行的试验表明, 氮肥会降低菌根感染率和存活率.Rühling和Tyler (1991)观察到氮添加会使菌根真菌几乎停止产生子实体.研究发现, 在目前的氮添加浓度(通常大于5 kg·hm-2·a-1)下, 凋落物的分解经常是被抑制的(Knorr et al., 2005), 主要是通过腐生性营养的微生物群落组成的改变(Gallo et al., 2004)和酶活性的改变(Frey et al., 2004).在本研究中, 作为微生物分解的底物, 凋落物在氮处理样地中的生物量要高于对照样地(图3), 分别比对照样地高出35.87%、22.17%和15.35%.但是, 氮添加在使地上部分凋落物增加的同时, 也可能导致微生物生物量和活性的下降, 从而在一定程度上限制凋落物的分解(Burton et al., 2004), 这也许会使得异养呼吸有所降低.另外, 有研究表明, 随着施氮时间的增加, 氮添加对凋落物分解的抑制作用也会增强(Fang et al., 2007).综上可知, 自养呼吸和异养呼吸对氮添加的不同反应综合导致了土壤总呼吸对氮添加的响应. ...
Nitrogen additions and litter decomposition: A meta-analysis 2005
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed 2 2008
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Nitrogen deposition and its ecological impact in China: An overview 2011
Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data 2 2007
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: A meta-analysis 1 2006
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
The effects of simulated nitrogen deposition on extracellular enzyme activities of litter and soil among different-aged stands of larch 1 2014
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Nitrogen deposition causes widespread loss of species richness in British habitats 1 2010
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Soil warming and carbon-cycle feedbacks to the climate system 1 2002
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China 3 2006
... 研究结果显示, 不同氮处理下的土壤呼吸呈现出相似的季节变化模式, 在夏季达到最大, 冬季降到最低, 这与在中国亚热带其他地区得到的研究结果相符(Tang et al., 2006; Mo et al., 2007).土壤呼吸的季节变化主要受到温度等气候因子和植物生长的调控影响(韩广轩和周广胜, 2009).2013年在对照样地测得的杜鹃灌丛年CO2释放量为(2.37 ± 0.39) kg·m-2, 低于Deng等(2010)在中国南亚热带森林测得的值((3.53 ± 0.03) kg·m-2), 但高于中国暖温带森林的年CO2释放量((1.50 ± 0.03) kg·m-2, Sun et al., 2014).试验结果显示, 低氮处理对土壤呼吸有促进作用, 而中氮和高氮处理则有轻微的抑制效应.综合前人的研究可知, 研究区域和施氮浓度不同, 土壤呼吸的响应也不同, 这受到土壤本身养分状况的影响(Mo et al., 2007).在氮含量丰富的环境中氮添加会抑制土壤呼吸, 而在氮限制的地区氮添加则会促进土壤呼吸(Hyvonen et al., 2007; Cusack et al., 2011). ...
... ., 2014).试验结果显示, 低氮处理对土壤呼吸有促进作用, 而中氮和高氮处理则有轻微的抑制效应.综合前人的研究可知, 研究区域和施氮浓度不同, 土壤呼吸的响应也不同, 这受到土壤本身养分状况的影响(Mo et al., 2007).在氮含量丰富的环境中氮添加会抑制土壤呼吸, 而在氮限制的地区氮添加则会促进土壤呼吸(Hyvonen et al., 2007; Cusack et al., 2011). ...
... 在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008).土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子.在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量.当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005). ...
Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China 2 2007
... 在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008).土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子.在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量.当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005). ...
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China 1 2008
Short and mediumterm contrasting effects of nitrogen fertilization on C and N cycling in a poplar plantation soil 1 2008
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Belowground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods, and models 2004
Temperature sensitivity of soil respiration in different ecosystems in China 2 2009
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
... 陆地生态系统的大部分碳储存于土壤中.已有的研究显示, 全球植物和土壤的碳储量分别为560 Pg (Forster et al., 2007)和>3300 Pg (Tarnocai et al., 2009).土壤呼吸是CO2从陆地生态系统返回到大气中的主要途径(Schlesinger & Andrews, 2000).CO2的主要来源是根系和根际微生物呼吸、土壤微生物和土壤动物呼吸.作为大气和陆地生态系统之间的第二大碳交换途径, 土壤呼吸每年向大气中释放的碳量高达68-98 Pg (Raich et al., 2002; Bond- Lamberty & Thomson, 2010), 在调节大气CO2浓度和地球的气候变化方面发挥着重要作用.因此, 土壤呼吸的轻微变化就会对全球的气候和环境产生剧烈的影响(Davidson & Janssens, 2006).导致全球变化的主要因子(如氮沉降的增加和CO2浓度的上升)在很大程度上影响了土壤呼吸速率(Bowden et al., 2004; Deng et al., 2010), 因此, 理解全球变化下土壤呼吸的反应机制对于预测未来的气候变化十分 必要. ...
Interannual varia- bility in global soil respiration, 1980-1994 2002
Indirect partitioning of soil respiration in a series of evergreen forest ecosystems 2006
Effects of simulated nitrogen deposition to the forest floor on the macrofungal flora of a beech forest 1 1991
... 一般来说, 将土壤呼吸分为自养呼吸(根系和根际微生物呼吸)和异养呼吸(微生物和土壤动物呼吸)两个组分(Sotta et al., 2004), 土壤总呼吸对氮添加的反应是自养呼吸、异养呼吸反应的综合效应.在全球范围内的森林生态系统中, 自养呼吸约占土壤总呼吸的45.8% (Hanson et al., 2000).Zhou等(2014)对来自不同生态系统的295个研究结果进行整合分析, 结果显示自养呼吸对氮添加的反应与根系生物量密切相关(R2 = 0.803).Rodeghiero和Cescatti (2006)对来自7种不同森林生态系统的数据进行研究, 也发现土壤呼吸与根系生物量之间存在着线性回归关系.所以, 氮添加对根系生物量的影响将会很大程度上影响植物根呼吸和与之相关的根际微生物呼吸.本试验中, LN、MN和HN处理样地的根系生物量年增量与对照样地相比, 分别高出了18.36%、36.49%和61.63%.可以看出, 随着氮添加浓度的升高, 根系生物量年增量呈现出递增的趋势, 因此会导致植物自养呼吸也随之上升.土壤异养呼吸来源于土壤中有机物质的分解, 主要依赖于分解底物的质量和数量、胞外酶活性、微生物生物量和微生物活性(Ryan & Law, 2005).Janssens等(2010)和Zhou等(2014)利用大量研究结果进行的整合分析显示, 氮添加与异养呼吸呈负相关关系, 特别是在生产力高、氮素不受限制的生态系统中, 这种负相关性更明显.Egerton-Warburton和Allen (2000)在加利福尼亚沿海灌丛林中进行的试验表明, 氮肥会降低菌根感染率和存活率.Rühling和Tyler (1991)观察到氮添加会使菌根真菌几乎停止产生子实体.研究发现, 在目前的氮添加浓度(通常大于5 kg·hm-2·a-1)下, 凋落物的分解经常是被抑制的(Knorr et al., 2005), 主要是通过腐生性营养的微生物群落组成的改变(Gallo et al., 2004)和酶活性的改变(Frey et al., 2004).在本研究中, 作为微生物分解的底物, 凋落物在氮处理样地中的生物量要高于对照样地(图3), 分别比对照样地高出35.87%、22.17%和15.35%.但是, 氮添加在使地上部分凋落物增加的同时, 也可能导致微生物生物量和活性的下降, 从而在一定程度上限制凋落物的分解(Burton et al., 2004), 这也许会使得异养呼吸有所降低.另外, 有研究表明, 随着施氮时间的增加, 氮添加对凋落物分解的抑制作用也会增强(Fang et al., 2007).综上可知, 自养呼吸和异养呼吸对氮添加的不同反应综合导致了土壤总呼吸对氮添加的响应. ...
Interpreting, measuring, and modeling soil respiration 1 2005
... 陆地生态系统的大部分碳储存于土壤中.已有的研究显示, 全球植物和土壤的碳储量分别为560 Pg (Forster et al., 2007)和>3300 Pg (Tarnocai et al., 2009).土壤呼吸是CO2从陆地生态系统返回到大气中的主要途径(Schlesinger & Andrews, 2000).CO2的主要来源是根系和根际微生物呼吸、土壤微生物和土壤动物呼吸.作为大气和陆地生态系统之间的第二大碳交换途径, 土壤呼吸每年向大气中释放的碳量高达68-98 Pg (Raich et al., 2002; Bond- Lamberty & Thomson, 2010), 在调节大气CO2浓度和地球的气候变化方面发挥着重要作用.因此, 土壤呼吸的轻微变化就会对全球的气候和环境产生剧烈的影响(Davidson & Janssens, 2006).导致全球变化的主要因子(如氮沉降的增加和CO2浓度的上升)在很大程度上影响了土壤呼吸速率(Bowden et al., 2004; Deng et al., 2010), 因此, 理解全球变化下土壤呼吸的反应机制对于预测未来的气候变化十分 必要. ...
Soil respiration and the global carbon cycle 1 2000
... 一般来说, 将土壤呼吸分为自养呼吸(根系和根际微生物呼吸)和异养呼吸(微生物和土壤动物呼吸)两个组分(Sotta et al., 2004), 土壤总呼吸对氮添加的反应是自养呼吸、异养呼吸反应的综合效应.在全球范围内的森林生态系统中, 自养呼吸约占土壤总呼吸的45.8% (Hanson et al., 2000).Zhou等(2014)对来自不同生态系统的295个研究结果进行整合分析, 结果显示自养呼吸对氮添加的反应与根系生物量密切相关(R2 = 0.803).Rodeghiero和Cescatti (2006)对来自7种不同森林生态系统的数据进行研究, 也发现土壤呼吸与根系生物量之间存在着线性回归关系.所以, 氮添加对根系生物量的影响将会很大程度上影响植物根呼吸和与之相关的根际微生物呼吸.本试验中, LN、MN和HN处理样地的根系生物量年增量与对照样地相比, 分别高出了18.36%、36.49%和61.63%.可以看出, 随着氮添加浓度的升高, 根系生物量年增量呈现出递增的趋势, 因此会导致植物自养呼吸也随之上升.土壤异养呼吸来源于土壤中有机物质的分解, 主要依赖于分解底物的质量和数量、胞外酶活性、微生物生物量和微生物活性(Ryan & Law, 2005).Janssens等(2010)和Zhou等(2014)利用大量研究结果进行的整合分析显示, 氮添加与异养呼吸呈负相关关系, 特别是在生产力高、氮素不受限制的生态系统中, 这种负相关性更明显.Egerton-Warburton和Allen (2000)在加利福尼亚沿海灌丛林中进行的试验表明, 氮肥会降低菌根感染率和存活率.Rühling和Tyler (1991)观察到氮添加会使菌根真菌几乎停止产生子实体.研究发现, 在目前的氮添加浓度(通常大于5 kg·hm-2·a-1)下, 凋落物的分解经常是被抑制的(Knorr et al., 2005), 主要是通过腐生性营养的微生物群落组成的改变(Gallo et al., 2004)和酶活性的改变(Frey et al., 2004).在本研究中, 作为微生物分解的底物, 凋落物在氮处理样地中的生物量要高于对照样地(图3), 分别比对照样地高出35.87%、22.17%和15.35%.但是, 氮添加在使地上部分凋落物增加的同时, 也可能导致微生物生物量和活性的下降, 从而在一定程度上限制凋落物的分解(Burton et al., 2004), 这也许会使得异养呼吸有所降低.另外, 有研究表明, 随着施氮时间的增加, 氮添加对凋落物分解的抑制作用也会增强(Fang et al., 2007).综上可知, 自养呼吸和异养呼吸对氮添加的不同反应综合导致了土壤总呼吸对氮添加的响应. ...
Soil CO2 efflux in a tropical forest in the central Amazon 1 2004
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil 2 2014
... 研究结果显示, 不同氮处理下的土壤呼吸呈现出相似的季节变化模式, 在夏季达到最大, 冬季降到最低, 这与在中国亚热带其他地区得到的研究结果相符(Tang et al., 2006; Mo et al., 2007).土壤呼吸的季节变化主要受到温度等气候因子和植物生长的调控影响(韩广轩和周广胜, 2009).2013年在对照样地测得的杜鹃灌丛年CO2释放量为(2.37 ± 0.39) kg·m-2, 低于Deng等(2010)在中国南亚热带森林测得的值((3.53 ± 0.03) kg·m-2), 但高于中国暖温带森林的年CO2释放量((1.50 ± 0.03) kg·m-2, Sun et al., 2014).试验结果显示, 低氮处理对土壤呼吸有促进作用, 而中氮和高氮处理则有轻微的抑制效应.综合前人的研究可知, 研究区域和施氮浓度不同, 土壤呼吸的响应也不同, 这受到土壤本身养分状况的影响(Mo et al., 2007).在氮含量丰富的环境中氮添加会抑制土壤呼吸, 而在氮限制的地区氮添加则会促进土壤呼吸(Hyvonen et al., 2007; Cusack et al., 2011). ...
... 在本研究区域, 土壤呼吸与土壤温度(R2为 0.77-0.82, p < 0.001)和土壤水分(R2为0.10-0.15, p < 0.05)显著正相关, 这一结果与已有的报道一致(Tang et al., 2006; Mo et al., 2007, 2008).土壤水分与土壤呼吸的相关性要小于土壤温度, 说明本区域土壤温度是影响土壤呼吸变化的主导因素, 这与研究地的中亚热带季风湿润气候类型有关, 本地区温度的季节变化明显, 降水丰富, 土壤含水量较高, 不是土壤呼吸的主要限制因子.在氮处理下土壤呼吸与土壤含水量的相关性与对照相比有所上升, 我们推测这是由于氮添加对植物生长的促进会加强植物对土壤水分的消耗(段洪浪等, 2009), 同时氮添加会增加叶片扩散导度和气孔导度(Deng et al., 2010), 加剧了水分的散失, 从而降低土壤含水量.当土壤水分含量降低时, 土壤含水量与土壤呼吸的相关性会更高(Dilustro et al., 2005). ...
Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China 1 2006
... 陆地生态系统的大部分碳储存于土壤中.已有的研究显示, 全球植物和土壤的碳储量分别为560 Pg (Forster et al., 2007)和>3300 Pg (Tarnocai et al., 2009).土壤呼吸是CO2从陆地生态系统返回到大气中的主要途径(Schlesinger & Andrews, 2000).CO2的主要来源是根系和根际微生物呼吸、土壤微生物和土壤动物呼吸.作为大气和陆地生态系统之间的第二大碳交换途径, 土壤呼吸每年向大气中释放的碳量高达68-98 Pg (Raich et al., 2002; Bond- Lamberty & Thomson, 2010), 在调节大气CO2浓度和地球的气候变化方面发挥着重要作用.因此, 土壤呼吸的轻微变化就会对全球的气候和环境产生剧烈的影响(Davidson & Janssens, 2006).导致全球变化的主要因子(如氮沉降的增加和CO2浓度的上升)在很大程度上影响了土壤呼吸速率(Bowden et al., 2004; Deng et al., 2010), 因此, 理解全球变化下土壤呼吸的反应机制对于预测未来的气候变化十分 必要. ...
Soil organic carbon pools in the northern circumpolar permafrost region 1 2009
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
Increased tree carbon storage in response to nitrogen deposition in the US 1 2010
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem 1 2013
Water- and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China 1 2008
... 由于化石燃料的燃烧和施肥等人类活动的影响, 大气氮沉降(NH+4-N和NO-3-N)在过去的一个世纪急剧增加(IPCC, 2013).在亚洲, 由于工农业的快速发展, 活性氮的使用和释放从1961年的14 Tg·a-1增加到2000年的68 Tg·a-1, 预计到2030年将达到105 Tg·a-1 (Zheng et al., 2002; Denman et al., 2007).中国的大气氮沉降也经历了快速的增长, 而且有不断发展的趋势(Liu et al., 2011), 一些亚热带地区森林的氮沉降量达到了30-73 kg·hm-2·a-1 (Mo et al., 2006).低水平的大气氮沉降可以刺激植物的生长, 增加植物对大气中碳的吸收, 从而缓解全球气候变化(Pregitzer et al., 2008; Thomas et al., 2010).但是过度的大气氮沉降会产生许多负面的生态学影响, 比如土壤酸化(Maskell et al., 2010)和生物多样性的丧失(Högberg et al., 2006).大气和土壤中的氮素积累已在很大程度上改变了区域和全球环境, 影响了陆地生态系统的碳循环(Luo et al., 2006), 从而对未来的气候变化产生影响(Melillo et al., 2002). ...
The Asian nitrogen cycle case study 1 2002
... 作为反映土壤呼吸与土壤温度间关系的重要指标, Q10值反映了土壤呼吸的温度敏感性, 对于量化和预测未来气候变化下的全球碳循环具有重要意义(Davidson & Janssens, 2006).本试验对照样地的Q10值为3.96, 高于前人测得的灌丛生态系统的Q10值(2.49 ± 0.54, Peng et al., 2009).这可能与本研究区域海拔较高有关.罗璐等(2011)对同一地区不同海拔梯度上的4种典型森林的土壤呼吸的研究显示, 海拔越高, 土壤呼吸对温度变化的响应越敏感.本研究中, 氮添加使得Q10值与对照相比有所降低, 这与前人的研究结果(Mo et al., 2008; Tu et al., 2013; Sun et al., 2014)相似.一个可能的原因是氮添加通过形成难利用有机质增强了土壤有机碳的稳定性, 氮与土壤有机质结合会产生高度耐微生物酶降解的化合物(Janssens et al., 2010), 使得微生物异养呼吸的底物可利用性降低.呼吸底物的可利用性在土壤呼吸对土壤温度的响应中起关键作用, 当底物供应下降时, 土壤呼吸的温度敏感性也会下降(Deng et al., 2010).同时氮添加也会降低微生物酶活性(Ma et al., 2014).这些改变会降低分解作用对温度变化的敏感性, 从而导致土壤呼吸的Q10降低.但是也有研究得出了相反的结论, 即氮添加增强了土壤呼吸温度敏感性(Pendall et al., 2004; Deng et al., 2010).这也许同样与底物供应有关, 在养分贫瘠的地区, 施氮促进植物生长, 增加了碳向根系的分配(Deng et al., 2010), 使得施氮对土壤有机质含量的促进作用大于其抑制作用, 从而增加了底物供应, 使Q10值增加.总的来说, 氮沉降对Q10的影响是个复杂的过程, 受到生态系统类型、环境因子和土壤养分含量等多种因素的影响(Zheng et al., 2009), 要综合考虑各因素的交互作用, 才能准确判断全球变化背景下氮沉降对Q10的影响. ...
Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content: A trans-China based case study 2009
Different responses of soil respiration and its components to nitrogen addition among biomes: A meta- analysis 2014

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT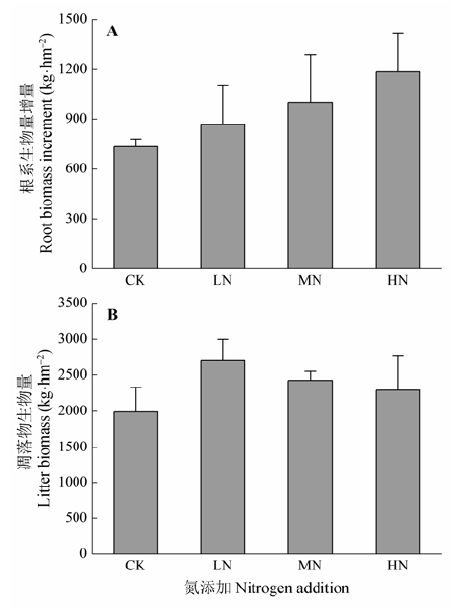 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT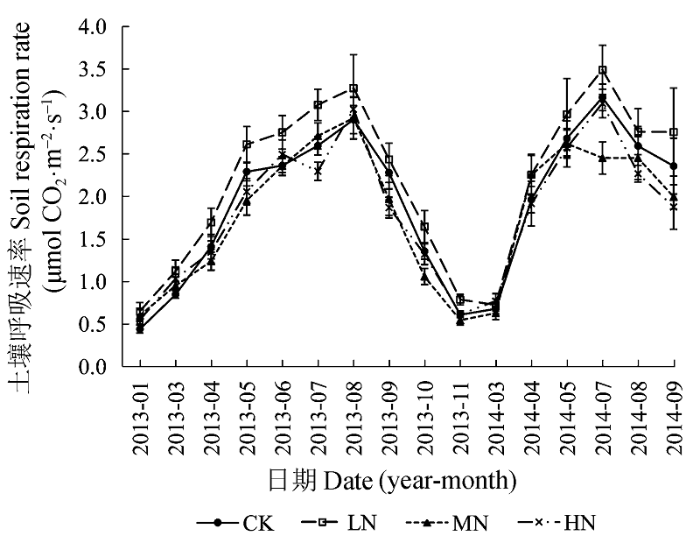 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT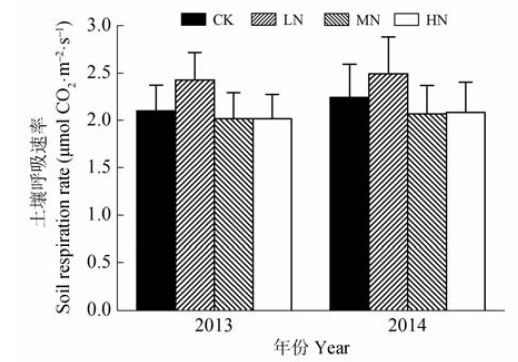 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT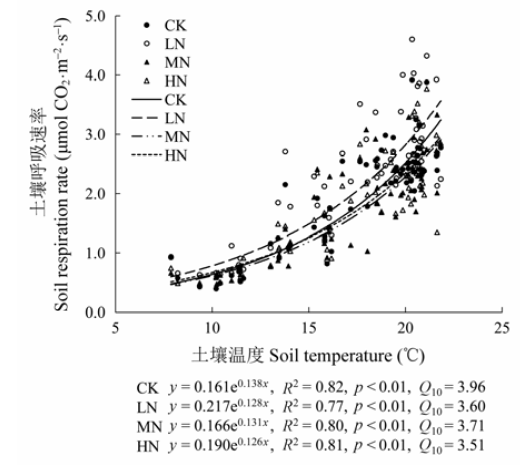 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT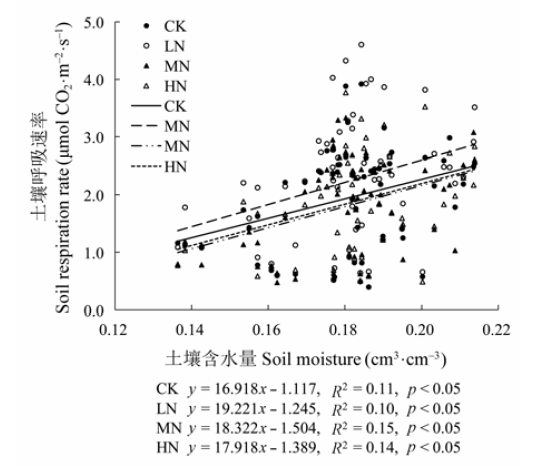 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}