 , 张鸣谦1, 杨骁婧1,3, 任苗苗1, 张守玉1, 金宁1, 刘燕霞1, 鲁红旭1, 许建强2,4, 徐卫平1,2,3
, 张鸣谦1, 杨骁婧1,3, 任苗苗1, 张守玉1, 金宁1, 刘燕霞1, 鲁红旭1, 许建强2,4, 徐卫平1,2,3
1. 大连理工大学海洋科学与技术学院, 盘锦 124221;
2. 大连理工大学盘锦产业技术研究院, 盘锦 124221;
3. 工业生态与环境工程教育部重点实验室, 大连 116024;
4. 大连理工大学生命科学与药学学院, 盘锦 124221
收稿日期: 2021-04-07; 修回日期: 2021-05-24; 录用日期: 2021-05-24
基金项目: 国家自然科学基金(No.31670767);中央高校基本科研业务费专项资金(No.DUT21LK29,DUT20LK36);营口市企业博士双创计划项目(No.202005,202112)
作者简介: 路延(1996—), 男, E-mail: lyllyying@mail.dlut.edu.cn
通讯作者(责任作者): 徐卫平, E-mail: Weiping.Xu@dlut.edu.cn
摘要:黑水虻处理是近年来发展起来的一种餐厨垃圾的有效资源化方法, 但由于黑水虻的生理特性, 油脂类物质经常不能被彻底降解和去除, 残渣仍有酸败气味, 需堆肥化二次处理.为了提高油脂类成分的降解效率, 提升黑水虻富集油脂的资源化能力, 本研究拟以含钙矿物和微生物为主要辅助手段, 通过向餐厨垃圾中添加湿重比例为0~10%的CaCO3、0~10%的Ca(OH)2、5%CaCO3和菌液的混合物及6种不同菌液, 研究含钙矿物和微生物对于油脂减量化和虫油转化效率的影响.结果发现, 向餐厨垃圾中添加含钙矿物, 能够显著调节和缓冲黑水虻处理过程中的环境pH值, 但对增加油脂的去除效率和黑水虻油脂的转化能力没有显著作用.而且添加7%~10% 的CaCO3对于餐厨垃圾的减量化率和黑水虻的虫产率有负作用, 添加5%~10%的Ca(OH)2会导致显著的幼虫死亡率.同时添加CaCO3和菌液对于油脂的去除不利.仅添加微生物菌液是提高垃圾油脂减量化率和黑水虻油脂富集率的最有效方法.枯草芽孢杆菌NJZ菌株、地衣芽孢杆菌NY菌株、植物乳酸杆菌RS72菌株和粪肠球菌L102菌株, 是提高黑水虻油脂转化效率的有效菌株.添加微生物后, 黑水虻的虫油产率由对照组的10.4%提升至12.8%~14.4%, 虫油富集率由对照组的93.9%提升至116%~130%.虫油富集率超过100%, 说明餐厨垃圾中碳水化合物、蛋白质等非油脂成分的一部分转化为黑水虻的体脂, 而微生物的辅助降解可能是强化这一过程的主要因素.但即使在接种微生物以后, 虫沙中仍然有29%~44%的油脂在处理结束时剩余, 说明餐厨垃圾中的油脂较难被彻底降解和利用.未来研究中, 可以继续筛选高效降解油脂的微生物, 与黑水虻共同处理餐厨垃圾, 实现油脂成分的资源化全利用.
关键词:黑水虻餐厨垃圾油脂Ca微生物强化资源化
Effects of calcium mineral and microbial inoculum on the lipid conversion efficiency during food waste treatment by black soldier fly larvae
LU Yan1,2
, ZHANG Mingqian1, YANG Xiaojing1,3, REN Miaomiao1, ZHANG Shouyu1, JIN Ning1, LIU Yanxia1, LU Hongxu1, XU Jianqiang2,4, XU Weiping1,2,31. School of Marine Science and Technology, Dalian University of Technology, Panjin 124221;
2. Panjin Industrial Technology Research Institute, Dalian University of Technology, Panjin 124221;
3. Key Laboratory of Industrial Ecology and Environmental Engineering, Ministry of Education, Dalian 116024;
4. School of Life Science and Pharmacy, Dalian University of Technology, Panjin 124221
Received 7 April 2021; received in revised from 24 May 2021; accepted 24 May 2021
Abstract: The food waste treatment by black soldier fly larvae (BSFL) is an effective method for nutrient recycle, which has been developed rapidly in recent years. However, due to the physiological limitation of BSFL, lipid substances in the food waste generally can not be fully degraded, and the residue frass still has rancidity odor and needs to be further composted. In order to improve the degradation efficiency of lipid component and elevate the lipid conversion rate of BSFL, calcium minerals and microorganisms were utilized in this study. Generally, 0~10% CaCO3, 0~10% Ca(OH)2, 5% CaCO3 with bacterial culture, and pure bacterial cultures were added to the food waste during the BSFL treatment. The effects of calcium minerals and microbial inoculum on lipid reduction and conversion rates were studied. The results showed that addition of calcium minerals to food waste could significantly adjust and buffer the environmental pH value, however limited effects were observed on the lipid removal and conversion efficiency. The addition of 7% ~10% CaCO3 resulted in negative effect on the food waste reduction and BSFL yield. The addition of 5%~10% Ca(OH)2 could lead to significant larval mortality. Moreover, the addition of both CaCO3 and bacterial inoculum resulted in unreliable results of lipid removal. Addition of microbial culture alone is the most effective way to improve the lipid reduction and BSFL lipid enrichment rate. Bacillus subtilis NJZ, Bacillus licheniformis NY, Lactobacillus plantarum RS72 and Enterococcus faecalis L102 were effective strains for improving the lipid conversion efficiency. Following the microbial inoculation, the BSFL lipid yield increased from 10.4% to 12.8%~14.4%, and the lipid enrichment rate elevated from 93.9% to 116%~130%. The lipid enrichment rates were more than 100%, indicating that part of the non-lipid components, e.g. carbohydrate and protein in the food waste were transformed into the larval body fat, and the microbial assistance might be the main factor that strengthen this effect. However, even with the microbial inoculum, there are still 29%~44% of the lipid in the food waste remained in the frass by the end of the treatment, indicating that the lipid component is difficult to be thoroughly degraded or recycled. Thus, it is highly recommended to screen microorganisms with high lipid degradation ability, in order to use them associated with BSFL, to achieve complete lipid conversion during food waste treatment by the BSFL.
Keywords: black soldier fly larvaefood wastelipidCamicrobial inoculumnutrient recycle
1 引言(Introduction)餐厨垃圾在城市固体垃圾中占比超过60%, 是城市固体垃圾中有机垃圾的主要组成部分(Zhang et al., 2010;邓俊, 2019).据统计, 仅2011年一年全世界产出了近2×108 t城市固体垃圾(Amoo et al., 2013), 我国2016年城市固体垃圾的日产量高达52×104 t, 位居世界第二(Aniekan et al., 2016).我国城市生活垃圾处理能力达到75.8×104 t·d-1(2015年底), 较2010年增加了30.1×104 t·d-1 (国家发展和改革委员会, 2016).餐厨垃圾的化学组成包括纤维素、淀粉、蛋白质、无机盐和脂类等(谢炜平等, 2008), 其中, 米面类食物的残余、动植物油、蔬菜、鱼刺、骨头等有机物质极易滋生细菌病菌, 腐败变质, 产生二氧化碳和甲烷(Rasit et al., 2019)等气体.因此, 亟需找到安全高效的处理方法, 实现餐厨垃圾的资源化回收利用.
黑水虻(Hermetia illucens L.)又称亮斑扁角水虻, 是在南北纬40°之间广泛分布的一种营腐性昆虫(James, 1935;Callan, 1974).已有研究表明, 黑水虻能够减量化和资源化餐厨垃圾.例如, Zheng等(2012)研究发现, 餐厨垃圾经黑水虻处理后, 垃圾减量化率达61.8%, 幼虫产率达到6.5%;Lalander等(2019)研究表明, 餐厨垃圾经黑水虻处理后, 垃圾减量化率为55.3%, 虫产率为13.9%.然而, 根据黑水虻的生理习性(郭孝结, 2016), 其对蛋白质和糖类物质的处理能力较强, 对脂类物质的处理能力较弱.我国餐厨垃圾中油脂含量经常较高, 粗油脂率可达9%~20%(谢炜平等, 2008).餐厨垃圾经黑水虻处理后, 油脂经常不能被彻底去除, 残渣有油脂酸败的气味, 需要二次堆肥才能彻底去除油脂(Xiao et al., 2018).如何在黑水虻处理餐厨垃圾的过程中, 提高油脂的降解效率, 强化黑水虻的油脂资源化率, 成为黑水虻处理研究的重要课题.
本课题组在前期研究中发现, 黑水虻处理餐厨垃圾的过程中, 环境pH值较长时间处于4.0~5.5的酸性范围(姜慧敏等, 2020).而黑水虻的最适生长环境pH值范围为6~10(Ma et al., 2018).如果调节环境pH值到中性至弱碱性环境, 则可能增强黑水虻的生长代谢, 并且有助于油脂类物质的碱性水解.Na离子虽然具有碱性调节作用, 但本课题组前期工作中发现, Na离子的调节能力有限, 经NaOH调节后的餐厨垃圾, 在处理过程中环境pH值仍以酸性范围为主.而Ca离子作为一种弱碱离子, 可能中和环境中的脂肪酸, 生成脂肪酸钙, 从而起到比Na离子更好的中和酸性pH值的作用.而且在无机盐离子中, 黑水虻幼虫仅对Ca、Mn和Fe等阳性离子有积累作用(Proc et al., 2020), 且幼虫对Ca离子的积累能力最强, 晚期幼虫体内含钙量是早期幼虫的2.26倍, 是3种阳离子中最高的, 说明Ca离子对黑水虻具有安全性和必须性.因此, 能否通过添加含钙矿物, 调节环境pH值, 促进黑水虻生长, 强化黑水虻对油脂的转化和利用, 值得深入研究.
除添加无机盐外, 接种微生物可能是另一种提高黑水虻油脂转化效率的方法.在以往的研究中, Xiao等(2018)发现向畜禽粪便中添加枯草芽孢杆菌可增强餐厨垃圾的转化效率, Somroo等(2019)发现向豆腐渣底物中添加乳酸杆菌可以增加黑水的虻产率和体脂含量, 而Wong等(2020)指出向椰子胚乳残渣中添加酿酒酵母能够提高黑水虻的产率和脂肪含量.因此, 向餐厨垃圾中添加适宜微生物, 有可能是强化黑水虻油脂转化效率的可行路径, 亟需进行深入研究.
基于此, 本研究拟以含钙矿物和微生物为主要辅助手段, 通过向餐厨垃圾中添加不同比例或种类的CaCO3、Ca(OH)2和菌液, 研究含钙矿物和微生物对油脂减量化效率和虫油转化率的影响, 以期能够发现高效资源化餐厨垃圾油脂成分的方法.
2 材料与方法(Materials and methods)2.1 实验仪器与试剂实验仪器:多功能碎菜机(型号280, 永康市铂欧五金制品)、高通量组织研磨仪(SCIENTZ-48, 宁波新芝)、pH计(FE38型, 上海梅特勒-托利多)、涡旋震荡混匀器(型号MX-F, SCILOGEX公司)、数显游标卡尺(CR2032, 永康市飞尔工贸)、四分位分析天平(ME204E, 上海梅特勒-托利多)、高速离心机(5417C, 德国艾本德)、烘箱(DKM610C, 武汉雅马拓仪器).
试剂药品:胰蛋白胨大豆肉汤培养基(TSB), 购自杭州微生物试剂公司;轻质CaCO3、Ca(OH)2等化学药品均为分析纯药品, 购自上海阿拉丁生化科技股份有限公司.实验微生物菌种为6株实验室自主分离并保存的食品发酵菌株, 菌株经16S rRNA基因鉴定确定种属.所用菌株包括枯草芽孢杆菌NJZ株(Bacillus subtilis NJZ)、地衣芽孢杆菌NY株(Bacillus licheniformis NY)、植物乳酸杆菌RS72株(Lactobacillus plantarum RS72)、鼠李糖乳酸杆菌RS54株(Lactobacillus rhamnosus RS54)、鲁梅利杆菌L131株(Rummeliibacillus stabekisii L131)及耐久粪肠球菌L102株(Enterococcus faecalis L102).实验中, 将冷冻保存的菌种保存液以1∶100的比例接种至TSB培养基中, 于37 ℃过夜振荡培养, 获得新鲜菌液, 于4 ℃保存, 12 h内使用.
2.2 黑水虻幼虫的准备黑水虻虫卵购自贵州毕节养殖场.收到黑水虻虫卵后, 配置孵化基质进行虫卵孵化, 孵化基质使用干燥的豆粕∶玉米粉∶麸皮(质量比6∶3∶1), 加入去离子水, 调节含水率到65%, 500 g孵化基质撒入25 g虫卵, 平铺放入4 L的长方形塑料盒后, 盒上覆盖2层无纺布, 用夹子固定, 在25 ℃室温下进行孵化.6 d后, 得到肉眼可见的小幼虫, 使用10目筛网, 将幼虫分离备用.
2.3 餐厨垃圾底物的准备餐厨垃圾取自校园食堂, 为午餐后的剩余食物垃圾, 包含米饭、面条、豆腐、鸡蛋、蔬菜、肉、鱼等餐厨废物.稻壳为水稻脱壳后的产物, 购自辽宁省盘锦市的农家店铺.餐厨垃圾经多功能碎菜机搅拌, 切碎为细小且均匀的颗粒物, 通过烘干法测得其含水率(n=3)为79%, 通过元素分析仪分析, 测得其C/N比(n=3)为21.3.随后向每100 g餐厨垃圾中添加0.308 g尿素, 调节其C/N比至约16∶1, 将调节好的餐厨垃圾于4 ℃保存, 12 h内使用.
2.4 实验设计实验共分为4个大组, 分别是添加轻质CaCO3的A组, 添加Ca(OH)2的B组, 同时添加轻质CaCO3和微生物的C组, 以及仅添加微生物的D组, 具体添加比例如表 1所示.其中, A、B、C、D各组均添加了质量为餐厨垃圾湿重10%的含钙矿物和稻壳的混合物, A组添加了0~10%的CaCO3, 不足10%的质量部分以稻壳补齐;B组添加了0~10%的Ca(OH)2, 不足10%的部分以稻壳补齐;C组添加了5%的CaCO3和5%的稻壳;D组添加了10%的稻壳.为保证餐厨垃圾的额外添加物总量一致, 使用了稻壳添加物, 在A、B组中, 稻壳是含钙矿物的替代物, 在C、D组中, 稻壳是液体菌液的固相载体.在菌液方面, A、B组添加了餐厨垃圾湿重10%的无菌TSB培养基, C、D组添加了餐厨垃圾湿重10%的过夜菌液, 且C1~C6和D1~D6小组的菌种如表 1所示.实验中, 每个条件为2个平行, 每个样品平行检测2次, 数据分析中, 每个条件含4个重复(n=4, 为2个样品×2次检测).
表 1(Table 1)
| 表 1 实验设计与分组 Table 1 Experimental design and groups | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 1 实验设计与分组 Table 1 Experimental design and groups
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2.5 垃圾处理过程黑水虻处理餐厨垃圾的实验在室内环境下进行, 室温(25±3) ℃, 日照时间12~14 h.使用容积为1750 mL的长方形塑料饭盒(177 mm×145 mm×82 mm)进行实验.按照每300 g(湿重)厨余垃圾, 配置30 g含钙矿物及稻壳, 30 mL TSB培养基(A、B组)或过夜菌液(C、D组), 以及700只黑水虻幼虫的方法进行处理.黑水虻幼虫先经手工数出100只称重, 重复3次, 3次均值为0.3929 g/100只虫, 则700只虫称重为2.75 g.在第0 d, 根据实验分组, 向每个容器中添加餐厨垃圾, 含钙矿物、稻壳及TSB培养基或菌液.用无菌金属棒充分搅拌和混合均匀后, 添加700只6日龄的黑水虻幼虫.餐厨垃圾处理共持续12 d, 在第0、3、6、9、12 d分别采集餐厨或虫沙样品(处理剩余物)进行pH值检测, 采集幼虫样品, 进行虫长和虫重的测量, 测量后放回盒中.在第12 d, 手工分离虫沙和幼虫, 分别称重, 冷冻保存.对采集到的第0、12 d的餐厨垃圾和虫沙样品, 以及第12 d的幼虫样品, 进一步测量含水率和含油率.
2.6 理化性质检测及转化效率评价方法虫沙和幼虫样品的含水率使用烘干法测量, 样品在105 ℃下的烘箱中烘干至恒重(约需2 h), 烘干过程中减少的质量比率为含水率, 剩余的质量比率为干物质率.烘干的虫沙和幼虫的油脂率, 通过石油醚浸提法测量(Li et al., 2011).虫沙样品经混合处理, 幼虫样品经剪碎处理后, 按照1∶10的质量比例加入石油醚, 室温浸提12 h, 回收石油醚, 并重复浸提2次, 合并3次浸提液, 蒸干石油醚, 得到粗油脂, 粗油脂质量占样品干重的比率为油脂率.幼虫体长和体重使用游标卡尺和四分位天平测量, 且体重测量中, 每次称量10只幼虫, 称重质量除以10后, 得到单只幼虫体重.餐厨及虫沙样品的pH值使用pH计测量, 每次称取2 g样品, 加入20 mL去离子水, 涡旋振荡2 min, 室温静置30 min, 使用FE38型pH计读取上清液的pH值.垃圾处理效率和油脂转化效率的综合评价, 使用式(1)~(7)计算.
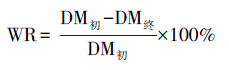 | (1) |
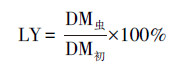 | (2) |
 | (3) |
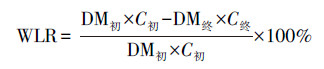 | (4) |
 | (5) |
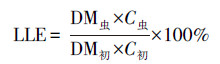 | (6) |
 | (7) |
2.7 统计分析方法数据制图使用SigmaPlot 14.0软件, 使用均值±标准差的方法作图.使用R软件的multcomp程序包完成显著性差异分析, 使用单因素方差分析方法(ANOVA)分析A、B、C、D大组内各小组间的差异, 辅助以glht和cld命令比较各小组大小.使用单因素协方差分析方法(ANCOVA)处理以时间为自变量的数据, 设定时间为协变量.将P <0.05的情况定义为差异显著.
3 结果与讨论(Results and discussion)3.1 环境pH的变化规律环境pH值的检测结果显示, 含钙矿物对餐厨垃圾处理过程中的pH值变化有较大的缓冲或调节作用, 而微生物的调节作用不明显(图 1).在处理的第0 d, A组的pH值为5.0~5.8, B组的pH值为5.0~10.4, C组的pH值为6.5~6.8, D组的pH值为5.0~5.3.可以看出, Ca(OH)2的添加(B组)对环境pH值的调节最为显著(p <0.001), 可将初始餐厨垃圾的酸碱度由酸性调节至碱性, CaCO3的(A组)调节作用其次.而同时添加CaCO3和菌液(C组), 餐厨垃圾的初始pH值可被调节至中性, 而仅添加菌液(D组), 餐厨垃圾的初始pH值为明显的酸性.在12 d的处理过程中, A组中的A2~A5组的变化曲线比A1组平缓, 组间没有显著差异(p=0.157), 整体呈现由酸性pH值向碱性pH值变化的趋势(图 1a).B组的不同小组差异显著(p<0.001), 在pH值上, 呈现B1<B2<B3<B4=B5的特点(图 1b).C组和D组的不同小组间差异均不显著, 环境pH值呈现第3~9 d由5.1逐渐升高到8.3, 第9~12 d由8.3逐渐下降到6.6的趋势(图 1c~1d).
图 1(Fig. 1)
 |
| 图 1 餐厨垃圾黑水虻处理过程中环境pH值的变化规律 Fig. 1Changing of environmental pH values during the kitchen waste larval treatment |
不论是在处理的初始阶段还是过程之中, CaCO3和Ca(OH)2都展现出较好的pH值缓冲或调节能力, CaCO3的缓冲能力更强, 添加不同CaCO3的小组在整个过程中pH值的变化曲线更平缓(图 1a), 而Ca(OH)2的酸碱调节能力更强, 环境pH值被调节至5~11(图 1b).根据黑水虻的生长特性, 环境pH值维持在6~10之间较为有利于黑水虻的生长存活(Ma et al., 2018; Meneguz et al., 2018), 这一条件对应于本研究2%~10% 的CaCO3和2%~5%的Ca(OH)2的范围, 说明上述条件可能有利于黑水虻的生长代谢, 且CaCO3比Ca(OH)2具有更宽的安全使用范围.相比于含钙矿物, 微生物是通过影响有机质的分解速率来改变环境pH值的.本研究中, 微生物仅对处理早期的环境pH值有影响, 即D组中第3 d的环境pH值有显著差异, 而微生物对中后期的环境pH值几乎无影响.与已有研究相比, 本课题组姜慧敏等(2020)前期研究发现, 餐厨垃圾在处理的前2~6 d, 环境pH值保持在4.0~5.5的显著弱酸性范围, 本研究的对照组A1小组重现了此研究结果, 并且通过添加含钙矿物发现, 2%~10%的CaCO3和2%~5%的Ca(OH)2能够调节初始阶段的环境pH值至5~7的范围, 可能因此有利于黑水虻的生长代谢.而在pH值变化的一般规律上, Ma等(2018)、Meneguz等(2018)、Kawasaki等(2020)和本研究都阐释了黑水虻处理餐厨垃圾的过程, 是环境pH值不断升高并由酸性向碱性转化的过程, 这与有机质的不断分解, 有机酸由初期生成、到中期消耗和末期耗尽的规律相吻合.
3.2 黑水虻的生长发育比较黑水虻的体长和体重的增长趋势在A、B、C、D 4个大组间存在显著差异(图 2).总的来看, A组和D组的生长情况较好, C组的生长情况为中等, B组的生长情况较差.根据处理结束时黑水虻幼虫的体长和体重(表 2)分析:A组中A2和A3组与A1组无差异, A4和A5组的体长和体重低于A1组, 说明添加CaCO3在2%~5%的范围对黑水虻生长发育无影响, 而7%~10%的范围对黑水虻生长不利.B组的黑水虻出现大量死亡, 在实验结束时, 仅B1组和B2组有黑水虻存活, B3、B4、B5组无幼虫存活, 且B2组的体重高于B1组, 说明Ca(OH)2添加量为2%时, 对黑水虻幼虫有利, 而添加量≥5%时, 对黑水虻幼虫有致死作用.这可能是由于添加的Ca(OH)2吸水结块, 黑水虻幼虫难以正常进食导致的.C组的各小组间, 体长无差异, 体重有差异, C5组最小, 且C组各小组的体长体重显著小于A1对照组, 说明同时添加5%的CaCO3和过夜菌液对黑水虻的生长发育不利.D组各组间, 体长有差异, D4组最短, 体重无差异, 且与A1对照组比较无差异, 且D大组总体的体重高于A、B、C大组, 说明添加微生物的条件相对于添加含钙矿物, 是有利于黑水虻个体增重的优势条件.
图 2(Fig. 2)
 |
| 图 2 黑水虻幼虫的体长(a~d)和体重(e~h)的生长变化曲线 Fig. 2Profiles of larval body length(a~d) and weight development (e~h) |
表 2(Table 2)
| 表 2 处理结束时黑水虻幼虫体长体重结果和垃圾转化效率的综合评价 Table 2 Evaluation of larval body length, body weight and waste conversion efficiency | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 2 处理结束时黑水虻幼虫体长体重结果和垃圾转化效率的综合评价 Table 2 Evaluation of larval body length, body weight and waste conversion efficiency
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
基于黑水虻体长和体重的增长曲线和终点值, 可以看出仅添加菌液的条件(D组)对黑水虻的生长最为有利, 其次是添加2%~5% CaCO3的小组(A2, A3)和2% Ca(OH)2的小组(B2组).同时添加CaCO3和菌液(C组)反而减弱了由CaCO3和菌液各自产生的促生长作用.然而根据经验判断, C组的幼虫较小, 可能是由于生长时间不足导致的, 即CaCO3和菌液的同时添加减弱了黑水虻的生长速度, 如果延长处理时间, 黑水虻还有可能继续发育.而Ca(OH)2小组对黑水虻的存活最为危险, 添加剂量≥5%时, 将导致显著的死亡现象.实际上, 本研究的原始设计还包括添加5%Ca(OH)2和菌液的6个小组, 但由于实验中多数小组在实验结束时幼虫死亡, 所以此结果并未纳入实验结果分析中.这也再次证明了Ca(OH)2的致死作用及较窄的应用剂量范围.在已有的研究中, Lopes等(2020)报道了使用有机废弃物饲养的黑水虻, 幼虫终体重为(0.171±0.012) g, Ma等(2018)报道了使用麸皮、玉米粉混合的人工饲料饲养黑水虻, 幼虫终体重为0.16~0.21 g.本研究中2%~5% CaCO3的小组(A2, A3)、2% Ca(OH)2小组(B2)和菌液小组(D组)与上述研究结果接近, 说明是有利于黑水虻生长和增重的可行条件.
3.3 餐厨垃圾处理效率的比较处理结束时, 垃圾减量化率、虫产率和生物转化率在A、B、C、D 4个大组之间存在显著差异(表 2), 总的来说, 垃圾减量化率和虫产率A、D组较高, C组较小, B组由于有幼虫死亡的现象, 有3个小组的参数接近0.在减量化率方面, A1、A2、A3小组无差别, A4、A5小组显著低于A1组, 说明CaCO3添加量为2%~5%时对垃圾减量化无影响, 而达到7%~10%时会显著降低餐厨垃圾的减量化效率.B2与B1小组无差别, 说明添加2%的Ca(OH)2对垃圾减量化无影响, 而B3、B4、B5小组由于黑水虻死亡, 所以垃圾减量化率几乎为零.C大组的减量化率显著低于A1小组, 说明同时添加CaCO3和菌液降低了垃圾的减量化率.D大组中, D3小组的减量化率显著高于A1组, D4小组的减量化率显著低于A1组.说明添加植物乳酸杆菌RS72株的菌液(C3), 对提高餐厨垃圾的减量化率有显著效果.虫产率的分析结果与垃圾减量化率的结果基本趋势一致, 即当处理小组的垃圾减量化率较大时, 虫产率也较高, 说明黑水虻幼虫的取食和生长代谢是垃圾减量化的主要动力.而且D大组中, D2、D3、D5小组的虫产率显著高于A1小组, 说明添加地衣芽孢杆菌NY株(D2)、植物乳酸杆菌RS72株(D3)和鲁梅利杆菌L131株(D5), 对于提高黑水虻幼虫产率具有显著作用.而D大组的虫产率总体高于A、B、C大组, 说明添加菌液比添加含钙矿物对于提高黑水虻幼虫产率更为有利.而在垃圾减量化率、虫产率和生物转化率这3个参数中, 生物转化率的指示作用可能最差.原因是C组的垃圾减量化率和虫产率最小, 但生物转化率最大, 这说明生物转化率参数受到垃圾减量化率的显著影响, C组可能是因为垃圾减量化率小, 而生物转化率较大.
不同处理条件下, 餐厨垃圾处理后的质量分配律体现了虫沙剩余率、虫产率和代谢损耗率的分配规律(图 3).总的来说, A1、A2、A3、B1、B2、D1、D2、D3、D5、D6属于质量分配规律相似的小组, 特点是虫沙剩余率较少, 为43%~52%, 虫产率较高, 为21%~29%, 代谢消耗率较高, 为21%~29%.
图 3(Fig. 3)
 |
| 图 3 黑水虻处理餐厨垃圾后的虫沙、幼虫和代谢损耗的质量分配规律 (虚线位于32%, 代表在总物质中含钙矿物和稻壳的质量占比为32%, 所有质量均以干基计) Fig. 3The mass distribution of frass, larvae and metabolic loss following black soldier fly larvae treatment |
姜慧敏等(2020)在前期研究中发现, 餐厨垃圾在黑水虻转化后, 减量化率可以达到70%~80%, 本研究中餐厨垃圾的减量化率降为50%左右.原因可能是本研究使用了含Ca矿物、稻壳及菌液为餐厨垃圾添加剂, 其中, 含钙矿物和稻壳的添加比例达到厨余垃圾湿重的10%, 干重的47.6%, 即为总干重的32%(图 3中的虚线).如果扣除32%的Ca矿和稻壳的质量占比, 本轮餐厨垃圾的减量化率仍然接近70%~80%, 与前期研究结果相似.虫产率方面, Lalander等(2019)报道过城市垃圾处理的虫产率为4.1%~15.2%, Gold等(2019)报道过餐厨垃圾的虫产率为15.3%~22.7%, 本研究空白小组(A1)的虫产率达到25%, 添加某些菌液后(D2、D3、D5组), 虫产率可达27%~29%, 说明空白组餐厨垃圾的处理条件和添加菌液后的条件, 可以达到较为高效的餐厨垃圾昆虫转化效果.
综合减量化率和虫产率的结果分析, 含钙矿物的添加并未显著提升餐厨垃圾的减量化率和虫产率, 相反, 仅添加菌液, 具体为添加地衣芽孢杆菌NY株、植物乳酸杆菌RS72株和鲁梅利杆菌L131株的菌液(D2、D3、D5组), 可以显著增强黑水虻的虫产率.分析这一现象的原因, 可能是所添加的菌液通过增强餐厨垃圾的降解效率或者促进黑水虻的生长代谢, 强化了黑水虻的餐厨垃圾转化作用.相似的研究成果在黑水虻处理粪便和食物垃圾中也有报道, 例如, Xiao等(2018)研究发现, 向鸡粪中添加枯草芽孢杆菌后, 可以将黑水虻处理的减量化率由35.8%提升至40.5%, 虫产率由8.04%提高到9.32%;Somroo等(2019)研究表明, 向豆腐渣中添加布赫内氏乳酸杆菌, 可以将黑水虻处理的减量化率由49.0%提高到55.7%, 虫产率由5.0%提高到6.9%;Wong等(2020)报道, 向椰子胚乳残渣中添加酿酒酵母菌, 可以将虫产率由9.5%提升至11.5%.可见, 向黑水虻处理底物中添加某些微生物, 可以促进垃圾减量化和虫体的生长转化.进一步的结合本研究的pH值变化和虫产率结果可以看出, 含钙矿物虽然具有调节环境pH值的能力, 但黑水虻并没有因为环境pH值的缓冲而显著提升垃圾处理能力或生长效率, 相比之下, 菌液对环境pH值的调节能力有限, 但可能因为增强有机质的降解或黑水虻的生长, 从而对黑水虻代谢产生了正向促进作用.
3.4 脂肪转化能力的比较经油脂含量测定, 初始餐厨垃圾含油率为11.06%±0.60%.处理结束时, 虫沙中的含油率和总体油脂减量化率如表 3所示.A组中, A2小组的含油率和油脂减量化率与A1小组无差异, 其他小组的油脂减量化率均小于A1小组, 说明添加CaCO3对增加油脂减量化率无显著作用.B组中, B2小组的虫沙含油率高于B1小组, 油脂减量化率低于B1小组, 说明添加Ca(OH)2对于增加油脂减量化无效果.C大组中, 虫沙的含油率总体等于或高于A1对照组, 油脂减量化率等于或低于A1对照组, 说明同时添加CaCO3和菌液对于促进厨余垃圾的油脂减量化无显著作用.D组中, D2小组的虫沙含油率低于A1小组, 油脂减量化率高于A1小组, 说明添加地衣芽孢杆菌NY株, 对于提高油脂的减量化率起到显著作用, 同时, D1、D3、D4、D6小组的油脂减量化率与A1对照小组无差异, D5小组的油脂减量化率低于A1小组, 说明只有添加适宜菌液, 才能对餐厨垃圾的油脂减量化起到显著作用.
表 3(Table 3)
| 表 3 处理结束时虫沙和幼虫的油脂含量与油脂转化效率的综合评价 Table 3 Evaluation of lipid content and lipid conversion efficiency of frass and larvae after food waste black soldier fly larvae treatment | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 3 处理结束时虫沙和幼虫的油脂含量与油脂转化效率的综合评价 Table 3 Evaluation of lipid content and lipid conversion efficiency of frass and larvae after food waste black soldier fly larvae treatment
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
在虫的油脂含量、虫油产率和虫油富集率方面(表 3), A大组中, 虽然A4小组的虫油含量略高于A1小组, 但虫油产率和虫油富集率低于A1小组, 总的来看, 添加CaCO3对于提高黑水虻的虫油产率和虫油富集率无作用.B大组结果与A大组相似, 添加Ca(OH)2对于促进黑水虻的油脂富集无作用.C大组的虫体油脂含量高于A1组, 但虫油产率和虫油富集率等于或低于A1组, 说明同时添加CaCO3和菌液可以提高黑水虻幼虫的体脂含量, 但由于幼虫体重较小, 该条件对于提高总的虫油产率和虫油富集率作用有限.D大组的D1、D2、D3、D6小组的虫体油脂率、虫油产率和虫油富集率均显著高于A1组, 而D4和D5组显著低于A1组, 说明向餐厨垃圾中添加枯草芽孢杆菌NJZ株(D1)、地衣芽孢杆菌NY株(D2)、植物乳酸杆菌RS72株(D3)和耐久性粪肠球菌L102株(D6), 对于提高黑水虻幼虫富集油脂的能力有显著作用.
以初始餐厨垃圾中的油脂总量为基准, 处理结束时虫沙中油脂总量和黑水虻幼虫中油脂总量相对于初始餐厨垃圾中油脂总量的比值如图 4所示.总的来看, D大组的虫沙油脂剩余率低, 虫体油脂富集率高, 而A、C大组处于中间水平, B大组的油脂剩余率高, 虫体油脂富集率低.D大组中, D1、D2、D3、D6小组的虫体富集油脂的效率最好, 表现为虫沙中油脂剩余率为28.9%~44.1%, 虫体中油脂富集率为115.9%~130.4%.另一方面, A、C、D大组内虫沙和黑水虻幼虫的总油脂量已经超过了餐厨垃圾的初始总油脂量, 说明黑水虻体内的脂肪一部分来自于餐厨垃圾的油脂, 一部分来自于非油脂物质的代谢转化.
图 4(Fig. 4)
 |
| 图 4 黑水虻处理餐厨垃圾结束时油脂的分配规律 (图中柱形图表示虫沙和虫体总油量相对于初始餐厨垃圾中总油量的比率(图中虚线位于100%, 代表餐厨垃圾的初始总油量为100%, 所有质量均以干基计) Fig. 4The lipid distribution profile after food waste treatment by black soldier fly larvae |
从虫沙中油脂的剩余含量来看, 黑水虻处理后, 餐厨垃圾的油脂率由11%最低降到6.81%, 虫沙中仍然有6.8%~15.4%的油脂残余, 而且也依然带有油脂酸败的味道, 说明在处理结束时, 油脂成分未被彻底降解和去除, 黑水虻分解油脂成分的能力有限.从A、B、C、D大组的油脂减量化率比较可以看出, 含钙矿物的添加对于提高油脂的去除效率没有起到显著作用, 而微生物的添加则可能有利于油脂的降解和去除.本研究中仅有一株地衣芽孢杆菌NY株(D2)对于提高油脂去除效率有显著作用, 所以未来工作中有必要进一步筛选促进油脂降解和利用的微生物, 将其与黑水虻共同用于餐厨垃圾的降解和处理, 从而提高油脂成分的转化和降解效率.在已有报道中, 关于黑水虻处理前后油脂减量化率的研究十分有限, Zheng等(2012)曾报道餐厨垃圾经黑水虻处理后, 总垃圾的减量化率达61.8%, 虫产率达到6.5%, 但垃圾中油脂成分的减量化率未见报道;Lalander等(2019)报道了餐厨垃圾经黑水虻处理后, 垃圾减量化率为55.3%, 虫产率为13.9%, 但餐厨和虫沙中的油脂成分含量未见分析.据笔者所知, 本研究首次报道了黑水虻处理后, 餐厨垃圾中油脂的减量化率参数, 在未接种微生物的条件下, 油脂减量化率达到约63%, 接种微生物后, 达到47%~71%, 此结果可作为以后研究的参照.
相对于虫沙中油脂成分的减量化研究, 黑水虻体内的脂肪含量及虫油转化率的研究则比较丰富.Li等(2011)报道在用猪粪、牛粪、鸡粪养殖的黑水虻幼虫中, 幼虫体内的粗脂肪含量达到29.1%~30.1%, 虫油总产率达到3.83%~9.87%.Surendra等(2016)报道的餐厨垃圾所饲养的黑水虻预蛹中, 粗脂肪的含量达到31.8%.Zheng等(2012)报道厨余垃圾经黑水虻处理后, 幼虫体内的粗脂肪含量为39.2%, 虫油总产率约为2.4%.可见, 黑水虻体内油脂的含量与饲养底物有关, 厨余垃圾底物可能比畜禽粪便垃圾具有更高的养分含量(Lalander et al., 2019), 从而使得黑水虻幼虫体内脂肪含量更高.本研究中, 空白组的黑水虻体脂含量达到41%, 与Zheng等(2012)的研究接近, 而在接种微生物以后, 体脂含量进一步提升至44%~53%, 说明微生物对于黑水虻体脂的积累产生了显著的促进作用.这一现象与另外2项利用微生物促进黑水虻转化作用的研究相似:Somroo等(2019)比较了接种和不接种赫内氏乳酸杆菌的豆腐残渣的处理效果, 结果发现, 接种菌液后黑水虻幼虫体内的脂肪含量由26.1%上升到30%;Wong等(2020)比较了接种不同剂量酿酒酵母的椰子残渣的处理效果, 结果发现, 接种了1%的酵母菌剂后, 黑水虻幼虫体内的油脂含量由42.5%提高到49.4%.本研究在向餐厨垃圾中接种枯草芽孢杆菌NJZ菌株、地衣芽孢杆菌NY菌株、植物乳酸杆菌RS72、粪肠球菌L102菌株以后, 黑水虻幼虫体内的脂肪含量由40.7%提高到44.1%~52.7%, 再次说明接种适宜微生物能够强化黑水虻幼虫的体脂含量.
有趣的是, 根据Somroo等(2019)和Wong等(2020)所报道的数据, 可以估算出黑水虻在豆腐残渣处理中的虫油产率约为1.3%~2.1%, 虫油富集率约为6.2%~9.9%, 在椰子残渣处理中的虫油产率约为4.3%~5.7%, 虫油富集率约为16.3%~21.8.这两项数据都明显低于本研究中餐厨垃圾接种适宜微生物后的虫油产率(12.8%~13.8%)和虫油富集率(115.9%~130.4%).然而, 本研究的底物含油率并不占优势, 豆腐残渣的含油率为20.86%(干基)(Somroo et al., 2019), 椰子残渣的含油率为26%(干基)(Wong et al., 2020), 本研究餐厨垃圾的含油率为11.06%(干基), 虽然黑水虻幼虫可能因为餐厨垃圾的营养条件产生更好的油脂转化效率, 但造成油脂富集率巨大差异的主要原因可能是虫料比.本研究向300 g(湿重)餐厨垃圾中加入了700只幼虫, 虫料的湿重比约为7∶3(700∶300), 而将垃圾质量换算为干重, 虫料比为7∶0.63(700∶63), 即11.1∶1, 而上述2项研究中均使用了500 g(干重)底物加入1000只幼虫的比例, 即虫料比为2∶1(1000∶500), 可见本研究中使用的虫料比为上数两项研究的5~6倍, 这可能是本研究虫油产率和虫油富集率高于上述两项研究的主要原因.
在虫油产率方面, 从本研究中A、B、C、D大组的比较可见, 添加2%~5%的CaCO3对于提高虫油产率无影响, 但添加7%~10%的CaCO3或者2%~10%的Ca(OH)2却能显著降低虫油产率, 对虫油积累不利.而添加适宜微生物是提高虫油产率最有效的方法.使用枯草芽孢杆菌NJZ菌株、地衣芽孢杆菌NY菌株、植物乳酸杆菌RS72、粪肠球菌L102菌株以后, 虫油产率可达12.8%~14.4%, 高于已报道的多数黑水虻转化垃圾生产油脂或生物柴油的研究(Li et al., 2011;Zheng et al., 2012;Somroo et al., 2019;Wong et al., 2020), 是利用黑水虻转化餐厨垃圾生产生物柴油的优势方法.使用餐厨垃圾底物、接种适宜微生物、应用较高的虫料比, 可能是本研究中虫油产率较高的主要原因.而且, 本研究中添加微生物的条件下, 虫油富集率已经超过100%, 说明黑水虻将餐厨垃圾中碳水化合物、蛋白质等非油脂成分转化为了自身体脂, 而微生物的添加可能对强化这一过程产生了主要作用. 但即使在接种微生物以后, 虫沙中仍然有29%~44%的油脂在处理结束时剩余, 说明餐厨垃圾中的油脂较难被彻底降解和利用.未来研究中, 可以继续筛选高效降解油脂的微生物, 与黑水虻共同处理餐厨垃圾, 目标实现油脂成分的资源化全利用.
4 结论(Conclusions)向餐厨垃圾中添加含钙矿物, 能够显著调节和缓冲黑水虻处理过程中的环境pH值, 但对增加油脂的去除效率和黑水虻油脂的富集能力没有显著作用, 而且添加7%~10%的CaCO3对于餐厨垃圾的减量化率和黑水虻的虫产率有负作用, 添加5%~10%的Ca(OH)2会导致显著的幼虫死亡率.同时添加CaCO3和菌液对于油脂的去除不利, 仅添加微生物菌液是提高垃圾油脂减量化率和黑水虻油脂富集率的最有效方法.枯草芽孢杆菌NJZ菌株、地衣芽孢杆菌NY菌株、植物乳酸杆菌RS72菌株、粪肠球菌L102菌株, 是提高黑水虻油脂转化效率的有效菌株.添加微生物后, 黑水虻的虫油产率由对照组的10.4%提升至12.8%~14.4%, 虫油富集率由对照组的93.9%提升至116%~130%, 说明餐厨垃圾中碳水化合物、蛋白质等非油脂成分的一部分转化为黑水虻的体脂, 而微生物可能是强化这一过程的主要因素.
参考文献
| Amoo O M, Fagbenle R L. 2013. Renewable municipal solid waste pathways for energy generation and sustainable development in the Nigerian context[J]. International Journal of Energy and Environmental Engineering, 4(1): 1-17. DOI:10.1186/2251-6832-4-1 |
| Aniekan I, Ikechukwu O. 2016. Review of municipal solid waste management technologies and its practices in China and germany[J]. International Journal of Technology Enhancements and Emerging Engineering Research, 5(4): 2347-2352. |
| Callan E. 1974. Hermetia illucems (L.) (Diptera: Stratiomyidae), a cosmopolitan American species long established in Australia and New Zealand[J]. Entomol Mon Mag, 109: 232-234. |
| 邓俊. 2019. 餐厨垃圾无害化处理与资源化利用现状及发展趋势[J]. 环境工程技术学报, 9(6): 637-642. |
| Gold M, Cassar C M, Zurbrügg C, et al. 2019. Biowaste treatment with black soldier fly larvae: Increasing performance through the formulation of biowastes based on protein and carbohydrates[J]. Waste Management, 102: 319-329. |
| 郭孝结. 2016. 黑水虻幼虫营养配方优化及应用[D]. 南昌: 南昌大学. 76 |
| James M T. 1935. The genus Hermetia in the United States (Dipt., Stratiomyidae)[J]. Bulletin of the Brooklyn Entomological Society, 30: 165-170. |
| Kawasaki K, Kawasaki T, Hirayasu H, et al. 2020. Evaluation of fertilizer value of residues obtained after processing household organic waste with black soldier Fly Larvae (Hermetia illucens)[J]. Sustainability (Basel, Switzerland), 12(12): 4920. |
| 姜慧敏, 王文霞, 任苗苗, 等. 2020. 黑水虻转化厨余垃圾过程中病原菌灭活规律的研究与综合评价[J]. 环境科学学报, 40(3): 273-284. |
| Lalander C, Diener S, Zurbrugg C, et al. 2019. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens)[J]. Journal of Cleaner Production, 208: 211-219. DOI:10.1016/j.jclepro.2018.10.017 |
| Li Q, Zheng L, Hao C, et al. 2011. From organic waste to biodiesel: Black soldier fly, Hermetia illucens, makes it feasible[J]. Fuel, 90(4): 1545-1548. DOI:10.1016/j.fuel.2010.11.016 |
| Liu C, Wang C, Yao H. 2019. Comprehensive resource utilization of waste using the black soldier fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae)[J]. Animals, 9(6): 349. DOI:10.3390/ani9060349 |
| Lopes I G, Lalander C, Vidotti R M, et al. 2020. Reduction of bacteria in relation to feeding regimes when treating aquaculture waste in fly larvae composting[J]. Frontiers in Microbiology, 11: 1616. DOI:10.3389/fmicb.2020.01616 |
| Ma J, Lei Y, Rehman K U, et al. 2018. Dynamic effects of initial pH of substrate on biological growth and metamorphosis of black soldier fly (Diptera: Stratiomyidae)[J]. Environmental Entomology, 47(1): 159-165. DOI:10.1093/ee/nvx186 |
| Meneguz M, Gasco L, Tomberlin J K, et al. 2018. Impact of pH and feeding system on black soldier fly (Hermetia illucens, L; Diptera: Stratiomyidae) larval development[J]. PloS One, 13(8): e202591. |
| Proc K, Bulak P, Wicek D, et al. 2020. Hermetia illucens exhibits bioaccumulative potential for 15 different elements - Implications for feed and food production[J]. Science of the Total Environment, 723: 138125. DOI:10.1016/j.scitotenv.2020.138125 |
| Rasit N, Hwe Fern L, Wan A K G W. 2019. Production and characterization of eco enzyme produced from tomato and orange wastes and its influence on the aquaculture sludge[J]. International Journal of Civil Engineering and Technology, 10(3): 976-980. |
| Somroo A A, Rehman K U, Zheng L, et al. 2019. Influence of Lactobacillus buchneri on soybean curd residue co-conversion by black soldier fly larvae (Hermetia illucens) for food and feedstock production[J]. Waste Management, 86(MAR.): 114-122. |
| Surendra K C, Olivier R, Tomberlin J K, et al. 2016. Bioconversion of organic wastes into biodiesel and animal feed via insect farming[J]. Renewable energy, 98: 197-202. DOI:10.1016/j.renene.2016.03.022 |
| Wong C Y, Mohd Aris M N, Daud H, et al. 2020. In-Situ yeast fermentation to enhance bioconversion of coconut endosperm waste into larval biomass of Hermetia illucens: statistical augmentation of larval lipid content[J]. Sustainability (Basel, Switzerland), 12(4): 1558. |
| Xiao X, Mazza L, Yu Y, et al. 2018. Efficient co-conversion process of chicken manure into protein feed and organic fertilizer by Hermetia illucens L.(Diptera: Stratiomyidae) larvae and functional bacteria[J]. Journal of Environmental Management, 217(JUL.1): 668-676. |
| 谢炜平, 梁彦杰, 何德文, 等. 2008. 餐厨垃圾资源化技术现状及研究进展[J]. 环境卫生工程, 2: 43-45. |
| Zhang D Q, Tan S K, Gersberg R M. 2010. Municipal solid waste management in China: Status, problems and challenges[J]. Journal of Environmental Management, 91(8): 1623-1633. DOI:10.1016/j.jenvman.2010.03.012 |
| Zheng L, Li Q, Zhang J, et al. 2012. Double the biodiesel yield: Rearing black soldier fly larvae, Hermetia illucens, on solid residual fraction of restaurant waste after grease extraction for biodiesel production[J]. Renewable Energy, 41: 75-79. DOI:10.1016/j.renene.2011.10.004 |
