Composition of plant life forms of subtropical shrubland in China and its correlation with temperature and precipitation
LIJia-Xiang1,2, XIONGGao-Ming1, XUWen-Ting1, LIYue-Lin3, LUZhi-Jun4, ZHAOChang-Ming1, XIEZong-Qiang1,*, 1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2College of Forest, Central South University of Forestry & Technology, Changsha 410004, China;3South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, Chinaand 4Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan 430074, China 通讯作者:* 通信作者Author for correspondence (E-mail: xie@ibcas.ac.cn) 责任编辑:LIJia-XiangXIONGGao-MingXUWen-TingLIYue-LinLUZhi-JunZHAOChang-MingXIEZong-Qiang 收稿日期:2016-06-28 接受日期:2016-11-11 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略性先导科技专项(XDA05050302)
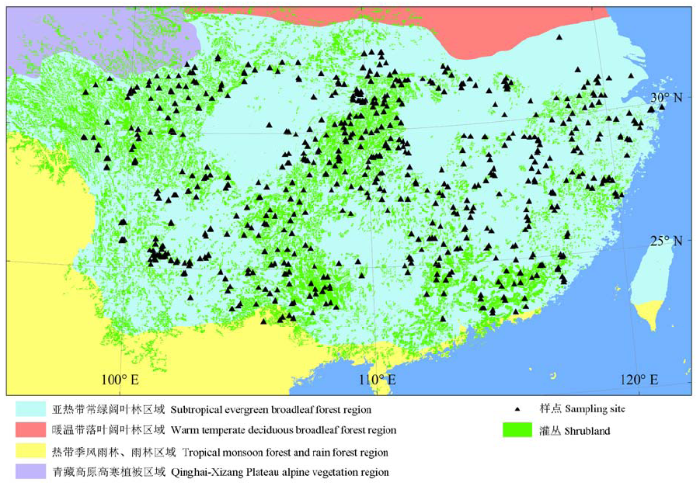
关键词:植物生活型谱;地理格局;敏感性;环境梯度;RDA排序 Abstract Aims Shrubland is one of the most important vegetation types in subtropical region, but studies on its life-form composition and geographic patterns are scarce. In this paper, we addressed the following questions: what are the characteristics of the composition of plant life forms of subtropical shrublands in China? And what are the relation-ships between the life form compositions, geographic patterns of subtropical shrublands and hydrothermal factors? Methods A total of 688 plots, designed with the stratified random sampling method, were systematically set and investigated in subtropical shrublands in China. Life forms were identified based on Raunkiaer’s (1934) life form systems. Life form spectrums were compiled at each sampling site and within the whole shrubland, respectively. Partial correlation analysis was applied to reveal the latitudinal, longitudinal and altitudinal patterns of each life form. Pearson’s correlation analysis was adopted to evaluate the relationships between each life form and hydrothermal factors. Further, redundancy analysis (RDA) was used to distinguish the effect of temperature and precipitation on geographical variations of life forms. Important findings (1) In subtropical shrublands in China, 56.37% of the total species were woody species, among which 33.88% were deciduous species and 22.49% were evergreen species. The remaining 43.63% of the total species were herbaceous species, dominated by 35.15% perennials. Generally, phaenerophytes (Ph) was the most dominant component, while the second dominant one was hemicryptophytes (H). The life form spectrum showed a pattern of Ph (55.74%) > H (31.03%) > T (therophytes 8.80%) > Cr (cryptophytes 4.08%) > Ch (chamaephytes 1.34%). (2) Plant life forms mainly exhibited longitudinal and altitudinal patterns. With the increase of longitude, the percentage of Ph tended to increase while H, Cr and T tended to decrease. Furthermore, an increase in altitude would lead to an increase in H and Cr but a decrease in Ph, Ch and T. (3) Among all the investigated climatic factors, the mean temperature of the warmest month (MWT) and the mean precipitation of the coldest month (MCP) had the greatest impact on Ph, H, Cr and T. With an increase of MWT and MCP, Ph would increase while H, Cr and T would decrease. Taken together, MWT and MCP could explain 41.97% of the variation of life form spectrum, among which 28.35% was attributed to their joint-effect. The results suggested that Ph and H were mainly influenced by both temperature and precipitation, and that Cr was more sensitive to temperature gradients. Besides, T was more sensitive to precipitation, and Ch showed little fluctuation along hydrothermal gradients. In conclusion, MWT and MCP are the most important hydrothermic factors that determine the geographic patterns of life forms in subtropical shrublands in China.
亚热带灌丛中, 木本(乔木、灌木、亚灌木和木质藤本)和草本植物(草质藤本和草本)分别占总种数的56.37%和43.63% (表1)。其中落叶木本和常绿木本植物分别占33.88%和22.49%。乔木习性的树种占总种数的15.14%; 灌木树种占37.15%; 藤本植物占4.08%, 其中不乏大型木质藤本, 如崖豆藤属(Mille- ttia)、瓜馥木属(Fissistigma)、羊蹄甲属(Bauhinia)等。草本物种以多年生草本最丰富, 占总种数的35.15% (包括草质藤本2.09%)。植物生活型以高位芽植物为主, 占总种数的54.75%; 其次为地面芽植物, 占31.03%; 一年生植物、隐芽植物和地上芽植物分别占8.80%、4.08%和1.34% (图2)。 Table 1 表1 表1亚热带灌丛维管植物生长型 Table 1Growth forms of vascular plants in subtropical shrublands
生长型 Growth form
常绿 Evergreen
落叶 Deciduous
多年生 Perennial
一年生 Annual
合计 Total
乔木 Tree
128 (6.87)
154 (8.27)
282 (15.14)
灌木 Shrub
268 (14.39)
411 (22.06)
679 (36.45)
亚灌木 Sub-shrub
13 (0.70)
13 (0.70)
木质藤本 Woody vine
23 (1.23)
53 (2.84)
76 (4.08)
草质藤本 Herbaceous vine
31 (1.66)
8 (0.43)
39 (2.09)
草本植物 Herb
624 (33.49)
150 (8.05)
774 (41.54)
合计 Total
419 (22.49)
631 (33.87)
655 (35.15)
158 (8.48)
1 863 (100.00)
The numbers in parentheses indicate the proportion (%) of each growth form in total species.括号内数字为各生长型所占总种数的比例(%)。 新窗口打开 显示原图|下载原图ZIP|生成PPT 图2亚热带灌丛植物生活型谱。Ch、Cr、H、Ph和T分别代表地上芽植物、隐芽植物、地面芽植物、高位芽植物和一年生植物。 -->Fig. 2The spectrum of life-forms of shrublands across sub- tropical region. Ch, Cr, H, Ph and T represents chamaephytes, cryptophytes, hemicryptophytes, phanerophytes, and therophy- tes, respectively. -->
3.2 生活型与地理因子的相关性
灌丛中生活型与纬度、经度和海拔之间不同程度地相关, 总体上各生活型与经度和海拔相关性均显著大于纬度(表2)。其中, 高位芽植物与经度显著正相关(r = 0.42, p < 0.001), 与纬度(r = -0.13, p < 0.001)和海拔(r = -0.18, p < 0.001)显著负相关; 地上芽植物和一年生植物与经度和海拔均呈显著负相关关系(p < 0.001), 但前者未表现出显著的纬度相关性(p > 0.05), 后者则与纬度显著正相关(p < 0.001); 地面芽植物与经度显著负相关(p < 0.001), 而与纬度(p < 0.05)和海拔(p < 0.001)显著正相关; 然而, 隐芽植物仅与海拔显著正相关(r = 0.39, p < 0.001), 而与纬度和经度没有显著相关关系(p < 0.05)。尽管高位芽植物、地面芽植物和一年生植物也都表现出一定的纬度相关性, 但其相关系数(依次为-0.13、0.08和0.15)却显著低于经度(0.42、-0.27和-0.33)和海拔(-0.18、0.25和-0.17)。 Table 2 表2 表2各生活型比例与经纬度和海拔的偏相关系数 Table 2Partial correlation coefficients between each life-form and latitude, longitude and altitude
生活型 Life-form
纬度 Latitude (°)
经度 Longitude (°)
海拔 Altitude (m)
高位芽植物 Phanerophyte
-0.13***
0.42***
-0.18***
地上芽植物 Chamaephyte
-0.05
-0.26***
-0.25***
地面芽植物 Hemicryptophyte
0.08*
-0.27***
0.25***
隐芽植物 Cryptophyte
0.00
0.02
0.39***
一年生植物 Therophyte
0.15***
-0.33***
-0.17***
***, ** and * represent significant level p < 0.001, p < 0.01 and p < 0.05, respectively.***、**和*分别表示回归显著性水平p < 0.001、p < 0.01和p < 0.05。 新窗口打开 RDA排序(图3A)也显示出经度和海拔是影响灌丛生活型组成变异的主要地理因子, 排序图从左到右主要反映随经度减少和海拔升高, 高位芽植物比例逐渐减少, 地面芽植物、隐芽植物、一年生植物逐渐增加的地理格局; 地上芽植物位于排序图的中心点附近, 说明其不随地理梯度发生较大变异。 显示原图|下载原图ZIP|生成PPT 图3生活型谱与地理变量(A)和主要水热因子(B)的冗余分析(RDA)排序图。Ch、Cr、H、Ph和T分别代表地上芽植物、隐芽植物、地面芽植物、高位芽植物和一年生植物。MCP和MWT为平均最冷月降水量和最热月平均气温。 -->Fig. 3Redundancy analysis (RDA) ordination biplots between life-form spectrum and geographical factors (A) and the main hydrothermal factors (B). Ch, Cr, H, Ph and T represents chamaephytes, cryptophytes, hemicryptophytes, phanerophytes, and therophytes, respectively. MCP and MWT are mean precipitation of the coldest month and mean temperature of the warmest month, respectively. -->
3.3 水热因子对灌丛生活型组成的影响
各生活型比例均不同程度地受水热因子的显著影响(表3)。从相关系数大小来看, 高位芽植物对温度和降水变化的响应最为强烈, 其次是地面芽植物和隐芽植物, 而地上芽植物受温度和降水的影响最小。7个气候因子中, MWT和MCP为影响最大的温度和降水指标, 其与高位芽植物、地面芽植物、隐芽植物和一年生植物的相关系数在同类因子中均最大。高位芽植物与MWT和MCP显著正相关(相关系数均为0.65), 地面芽植物、隐芽植物和一年生植物与MWT和MCP显著负相关, 地面芽植物与MWT的相关性(-0.60)略大于MCP (-0.54), 隐芽植物与MWT更为相关(-0.55), 而一年生植物与MCP相关性更大(-0.36)。然而, 温度(MAT和MCT)和降水(MCP)对地上芽植物的影响趋势相反, 前者与其正相关, 后者与其负相关。 Table 3 表3 表3各水热因子与生活型的Pearson相关系数 Table 3Pearson’s coefficients between hydrothermic factors and each life-form
气候指标 Climate factor
Ph
Ch
H
Cr
T
年平均气温 Mean annual temperature (MAT, ℃)
0.56***
0.14***
-0.54***
-0.50***
-0.13***
最冷月平均气温 Mean temperature of the coldest month (MCT, ℃)
0.42***
0.17***
-0.43***
-0.40***
-0.08*
最热月平均气温 Mean temperature of the warmest month (MWT, ℃)
0.65***
0.07
-0.60***
-0.55***
-0.19***
年降水量 Mean annual precipitation (MAP, mm)
0.60***
-0.03
-0.51***
-0.37***
-0.31***
最冷月降水量 Mean precipitation of the coldest month (MCP, mm)
0.65***
-0.10**
-0.54***
-0.37***
-0.36***
最热月降水量 Mean precipitation of the warmest month (MWP, mm)
-0.25***
0.05
0.18***
0.25***
0.13***
生长季降水量 Mean precipitation of growing season (MGP, mm)
0.59***
0.02
-0.53***
-0.37***
-0.28***
Ch, Cr, H, Ph and T represents chamaephytes, cryptophytes, hemicryptophytes, phanerophytes, and therophytes, respectively. ***, ** and * represent significant level p < 0.001, p < 0.01 and p < 0.05, respectively.Ch、Cr、H、Ph和T分别代表地上芽植物、隐芽植物、地面芽植物、高位芽植物和一年生植物。***、**和*分别表示回归显著性水平p < 0.001, p < 0.01和p < 0.05。 新窗口打开 RDA排序(图3B)轴1从左到右反映温度和降水逐渐增加的水热趋势, 占排序总解释量的98.21%, 可代表排序的主要环境梯度, 可解释生活型组成变异的41.38%。MWT和MCP间的夹角小于45°, 说明温度和降水共同决定了灌丛生活型的组成变异。从左到右随着MWT和MCP增加, 高位芽植物逐渐增多, 地面芽植物逐渐减少; 一年生植物和隐芽植物分别位于MCP和MWT的反向延长线上, 与MCP和MWT负相关; 地上芽植物位于排序图的中心位置, 受温度和降水的影响微弱。同时, 排序共解释了亚热带灌丛生活型组成变异的41.97%, 主要来源于MCP和MWT的共同贡献(28.35%), 各自独立贡献仅为7.60%和6.03% (图4)。 显示原图|下载原图ZIP|生成PPT 图4最热月平均气温(MWT)和最冷月降水量(MCP)对灌丛生活型组成贡献的变差分解。 -->Fig. 4Variation partitioning results of the contribution of mean temperature of the warmest month (MWT) and mean precipitation of the coldest month (MCP) to the life-form composition in shrublands. -->
AmjadMS, ArshadM, SadafHM, Durr-E-Shahwar, AkrimF, ArshadA (2016). Floristic composition, biological spectrum and conservation status of the vegetation in Nikyal valley, Azad Jammu and Kashmir .Asian Pacific Journal of Tropical Disease, 6, 63-69. [本文引用: 1]
[2]
BatalhaMA, MartinsFR (2004). Floristic, frequency, and vegetation life-form spectra of a Cerrado site .Brazilian Journal of Biology, 64, 201-209. [本文引用: 1]
[3]
BorcardD, GilletF, LegendreP (2011). Numerical Ecology with R . Springer, New York. [本文引用: 1]
[4]
ChenS, BaiY, LinG, HanX (2005). Variations in life-form composition and foliar carbon isotope discrimination among eight plant communities under different soil moisture conditions in the Xilin River Basin, Inner Mongolia, China .Ecological Research, 20, 167-176. [本文引用: 2]
[5]
ChenWJ (2006). Floristic Phytogeography of Evergreen Broad-leaved Forest (EBLF) in Mid-Subtropical China . PhD dissertation, East China Normal University, Shanghai. (in Chinese with English abstract)[陈卫娟 (2006). 中亚热带常绿阔叶林植物区系地理研究 . 博士学位论文, 华东师范大学, 上海.] [本文引用: 1]
[6]
GaoXM, ChenLZ (1998). The revision of plant life-form system and an analysis of the life-form spectrum of forest plants in the warm temperate zone of China .Acta Botanica Sinica, 40, 553-559. (in Chinese with English abstract)[高贤明, 陈灵芝 (1998). 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析 . 植物学报, 40, 553-559.] [本文引用: 2]
[7]
GuoK, ZhengD, LiBS (1998). The characteristics of plant life form spectra in the Karakorum-Kunlun mountains Acta Phytoecologica Sinica, 22, 51-59. (in Chinese with English abstract)[郭柯, 郑度, 李渤生 (1998). 喀喇昆仑山——昆仑山地区植物的生活型组成 . 植物生态学报, 22, 51-59.] [本文引用: 3]
[8]
GuoQS, JiangH, WangB, YanH (1999). The quantitive classfication and spatial distribution pattern of life form spectra of the plants in major Chinese forest communities .Acta Ecologica Sinica, 19, 573-577. (in Chinese with English abstract)[郭泉水, 江洪, 王兵, 阎洪 (1999). 中国主要森林群落植物生活型谱的数量分类及空间分布格局的研究 . 生态学报, 19, 573-577.] [本文引用: 8]
[9]
GuoZX, WangZM, SongKS, ZhangB, LiF, LiuDW (2007). Correlations between forest vegetation NDVI and wa- ter/thermal condition in Northeast China forest regions in 1982-2003 . Chinese Journal of Ecology, 26, 1930-1936. (in Chinese with English abstract)[国志兴, 王宗明, 宋开山, 张柏, 李方, 刘殿伟 (2007). 1982-2003年东北林区森林植被NDVI与水热条件的相关分析 . 生态学杂志, 26, 1930-1936.] [本文引用: 1]
[10]
HedwallPO, SkoglundJ, LinderS (2015). Interactions with successional stage and nutrient status determines the life-form-specific effects of increased soil temperature on boreal forest floor vegetation .Ecology & Evolution, 5, 948-960. [本文引用: 2]
[11]
HijmansRJ, CameronSE, ParraJL, JonesPG, JarvisA (2005). Very high resolution interpolated climate surfaces for global land areas . International Journal of Climatology, 25, 1965-1978. [本文引用: 1]
[12]
HolmS (1979). A simple sequentially rejective multiple test procedure .Scandinavian Journal of Statistics, 6, 65-70. [本文引用: 1]
[13]
HuZH, YuMJ, XuXH, FuHL (2004). Castanopsis eyrei community characteristics in Gutianshan Natural Reserve, Zhejiang Province .Chinese Journal of Ecology, 23, 15-18. (in Chinese with English abstract)[胡正华, 于明坚, 徐学红, 付海龙 (2004). 浙江古田山自然保护区甜槠群落特征研究 . 生态学杂志, 23, 15-18.] [本文引用: 2]
[14]
JiangH (1994). Study on life-form spectrum of plant community in Dongling mountain .Acta Botanica Sinica, 36, 884-894. (in Chinese with English abstract)[江洪 (1994). 东灵山植物生活型谱的比较研究 . 植物学报, 36, 884-894.] [本文引用: 1]
[15]
LegendreP, OksanenJ, ter BraakCJF (2011). Testing the significance of canonical axes in redundancy analysis .Methods in Ecology and Evolution, 2, 269-277. [本文引用: 1]
[16]
LeiNF, SuZX, SongHX, ZhangY (2002). A comparative study on life-form spectra of evergreen broad-leaved forest in different successions in Jinyun mountain .Chinese Journal of Applied Ecology, 13, 267-270. (in Chinese with English abstract)[雷泞菲, 苏智先, 宋会兴, 张焱 (2002). 缙云山常绿阔叶林不同演替阶段植物生活型谱比较研究 . 应用生态学报, 13, 267-270.] [本文引用: 6]
[17]
LiJX (2016). Biodiversity Pattern and Ecosystem Productivity in Shrublands Across the Subtropical Region in China . PhD dissertation, Institute of Botany, Chinese Academy of Sciences, University of the Chinese Academy of Sciences, Beijing. (in Chinese with English abstract)[李家湘 (2016). 亚热带灌丛生物多样性及其与生产力关系 . 博士学位论文, 中国科学院大学, 中国科学院植物研究所, 北京.] [本文引用: 2]
[18]
LiuSJ, SuZX, ZhangJX, HuJY (2003). Perspectives of the research on life form in land plant communities .Journal of Sichuan Teachers College (Natural Science), 24, 155-159. (in Chinese with English abstract)[刘守江, 苏智先, 张璟霞, 胡进耀 (2003). 陆地植物群落生活型研究进展 . 四川师范大学学报(自然科学版), 24, 155-159.] [本文引用: 1]
[19]
MahdaviP, AkhaniH, van der MaarelE (2012). Species diversity and life-form patterns in steppe vegetation along a 3000 m altitudinal gradient in the Alborz Mountains, Iran .Folia Geobotanica, 48, 7-22. [本文引用: 4]
[20]
OdlandA (2009). Interpretation of altitudinal gradients in south central norway based on vascular plants as environmental indicators .Ecological Indicators, 9, 409-421. [本文引用: 1]
[21]
PavónNP, HumbertoH, VíctorR (2000). Distribution of plant life forms along altitudinal gradient in the semi-arid valley of zapotitlán, méxico .Journal of Vegetation Science, 11, 39-42. [本文引用: 2]
[22]
QiCJ, XiaoYT (1990). Vegetation of Hunan. Hunan Science and Technology Press, Changsha. (in Chinese)[祁承经, 肖育檀 (1990). 湖南植被. 湖南科学技术出版社, 长沙.] [本文引用: 1]
RajuVS, GopalP, SuthariS (2014). Environmental assessment of climate of a habitat through floristic life-form spectra, a case study of Warangal north forest division, Telangana, India .Journal of Natural Sciences, 2, 77-93. [本文引用: 4]
[25]
RaunkiaerC (1934). The Life Forms of Plants and Statistical Plant Geography. Clarendon Press, Oxford, UK. [本文引用: 4]
[26]
SongYC (2004). Vegetation Ecology. East China Normal University Press, Shanghai. (in Chinese)[宋永昌 (2004). 植被生态学. 华东师范大学出版社, 上海.] [本文引用: 2]
[27]
SwaineMD, BeckerP (1999). Woody life-form composition and association on rainfall and soil fertility gradients in Ghana .Plant Ecology, 145, 167-173. [本文引用: 1]
[28]
The Editorial Committee of Vegetation Map of China, Chinese Academy of Sciences (2007). Vegetation Map of the People’s Republic of China 1:1000000. Geological Publishing House, Beijing. (in Chinese)[中国科学院中国植被图编辑委员会 (2007). 中华人民共和国植被图1:1000000. 地质出版社, 北京.]
[29]
VashisthaRK, RawatN, ChaturvediAK, NautiyalBP, PrasadP, NautiyalMC (2011). Characteristics of life-form and growth-form of plant species in an alpine ecosystem of Northwest Himalaya .Journal of Forestry Research, 22, 501-506. [本文引用: 2]
[30]
WangG, ZhouG, YangL, LiZ (2003). Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slope of Qilianshan Mountains, Gansu, China .Plant Ecology, 165, 169-181. [本文引用: 3]
[31]
WangMT (1987). Study on life-forms of evergreen broad- leaved forest in subtropics of China .Journal of Ecology, 6, 21-23. (in Chinese with English abstract)[王梅峒 (1987). 中国亚热带常绿阔叶林生活型的研究 . 生态学杂志, 6, 21-23.] [本文引用: 5]
[32]
WangX, HuWQ, LiJX, HuangSN (2011). Preliminary studies on montane elfin forest community characteristics in mountain Shikengkong of Nanling in Guangdong Province .Journal of Zhejiang Forestry Science and Technology, 31, 12-17. (in Chinese with English abstract)[王旭, 胡文强, 李家湘, 黄世能 (2011). 广东南岭石坑崆山顶矮林群落结构特征 . 浙江林业科技, 31, 12-17.] [本文引用: 1]
[33]
WangYH, XuX, ZhangDJ, ZhangHY (2016). Effects of climate and habitat heterogeneity on the distribution pattern of plant life form in North China . Journal of Anhui Agriculture Science, 44(16), 9-13. (in Chinese with English abstract)[王艳红, 徐翔, 张东杰, 张化勇 (2016). 气候和生境异质性对华北地区植物生活型分布格局的影响 . 安徽农业科学, 44(16), 9-13.] [本文引用: 3]
[34]
WhittakerRH (1970). Communities and Ecosystems . Mac- millan Company, New York. [本文引用: 1]
[35]
WuZY (1980). Vegetation of China. Science Press, Beijing. (in Chinese)[吴征镒 (1980). 中国植被. 科学出版社, 北京.] [本文引用: 1]
[36]
XieZQ, TangZY (2015). Technical specification of field investigation and laboratory analysis for carbon sequestration in shrubland ecosystems. In: Compilation Group of Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems ed. Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems. Science and Technology Press, Beijing. 145-191. (in Chinese)[谢宗强, 唐志尧 (2015). 灌丛生态系统固碳研究的野外调查与室内分析技术规范. 见: 生态系统固碳观测与调查技术规范编写组编. 生态系统固碳观测与调查技术规范. 科学技术出版社, 北京. 145-191.] [本文引用: 1]
[37]
YuSL, ChenLZ, MaKP (2000). Life-form spectrum of vascular plants of Quercus mongolica communities in Northeast China . Scientia Silvae Sinicae, 36, 118-121. (in Chinese with English abstract)[于顺利, 陈灵芝, 马克平 (2000). 东北地区蒙古栎群落生活型谱比较 . 林业科学, 36, 118-121.] [本文引用: 1]
[38]
ZhangGF, SongYC (2001). Studies on species composition, structure and physiognomy of shrub communities in Tiantong Region, Zhejiang Province .Guihaia, 21, 201-207. (in Chinese with English abstract)[张光富, 宋永昌 (2001). 浙江天童灌丛群落的种类组成、结构及外貌特征 . 广西植物, 21, 201-207.] [本文引用: 3]
[39]
ZhaoJ (1995). Natural Geography of China. Higher Education Press, Beijing. (in Chinese)[赵济 (1995). 中国自然地理. 高等教育出版社, 北京.] [本文引用: 1]
Floristic composition, biological spectrum and conservation status of the vegetation in Nikyal valley, Azad Jammu and Kashmir 1 2016
... 生活型是植物通过对环境条件长期适应, 在其生理、结构, 尤其是外部形态上的一种具体表现(生态适应性), 其组成是群落层片结构划分的主要依据, 群落中优势植物生活型决定了群落的外貌形态(高贤明和陈灵芝, 1998; 宋永昌, 2004; Raju et al., 2014).常用的植物生活型系统是Raunkiaer分类系统(Raunkiaer, 1934).该系统认为植物在不良季节休眠芽所在的位置是表达其适应不利季节的最好方式(Pavón et al., 2000; Odland, 2009).由于植物长期对区域综合自然环境条件的趋同适应, 植物生活型谱可以较清楚地反映出当地的气候特征, 并与植被类型有着密切的联系.不同地区相同群落类型由于环境条件的差异形成不同的植物生活型谱(王梅峒, 1987; 郭泉水等, 1999; Vashistha et al., 2011; Amjad et al., 2016); 同一地区处于不同演替阶段的植物群落, 生活型谱也有差异(张光富和宋永昌, 2001; 雷泞菲等, 2002; Hedwall et al., 2015).因此, 分析植被生活型组成特点及其地理格局有助于我们认识区域气候环境特点及其地域分异规律, 理解植物对环境的适应策略(郭柯等, 1998; Chen et al., 2005; Raju et al., 2014). ...
Floristic, frequency, and vegetation life-form spectra of a Cerrado site 1 2004
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
Numerical Ecology with R 1 2011
... 为了分析温度和降水对各生活型组成的影响, 采用Pearson’s相关系数评估单个水热因子与各生活型的关系, 运用双尾t检验检验相关系数的显著性, 所有p值用Holm法(Holm, 1979)进行校正.同时采用RDA排序分析来分析亚热带灌丛生活型组成在水热梯度上的变化规律和主要水热因子, 排序分析以及水热因子与排序轴之间的关系均用Monte-Carlo置换999次进行显著性检验(p < 0.001)(Legendre et al., 2011).用基于偏RDA分析的变差分解来衡量温度和降水用对亚热带灌丛生活型组成变异的相对贡献量(Borcard et al., 2011).因同类水热因子间存在较强的共线性, 在RDA排序和偏RDA分析时, 仅选择了影响最大的温度和降水因子参与分析. ...
Variations in life-form composition and foliar carbon isotope discrimination among eight plant communities under different soil moisture conditions in the Xilin River Basin, Inner Mongolia, China 2 2005
... 生活型是植物通过对环境条件长期适应, 在其生理、结构, 尤其是外部形态上的一种具体表现(生态适应性), 其组成是群落层片结构划分的主要依据, 群落中优势植物生活型决定了群落的外貌形态(高贤明和陈灵芝, 1998; 宋永昌, 2004; Raju et al., 2014).常用的植物生活型系统是Raunkiaer分类系统(Raunkiaer, 1934).该系统认为植物在不良季节休眠芽所在的位置是表达其适应不利季节的最好方式(Pavón et al., 2000; Odland, 2009).由于植物长期对区域综合自然环境条件的趋同适应, 植物生活型谱可以较清楚地反映出当地的气候特征, 并与植被类型有着密切的联系.不同地区相同群落类型由于环境条件的差异形成不同的植物生活型谱(王梅峒, 1987; 郭泉水等, 1999; Vashistha et al., 2011; Amjad et al., 2016); 同一地区处于不同演替阶段的植物群落, 生活型谱也有差异(张光富和宋永昌, 2001; 雷泞菲等, 2002; Hedwall et al., 2015).因此, 分析植被生活型组成特点及其地理格局有助于我们认识区域气候环境特点及其地域分异规律, 理解植物对环境的适应策略(郭柯等, 1998; Chen et al., 2005; Raju et al., 2014). ...
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
Interactions with successional stage and nutrient status determines the life-form-specific effects of increased soil temperature on boreal forest floor vegetation 2 2015
... 生活型是植物通过对环境条件长期适应, 在其生理、结构, 尤其是外部形态上的一种具体表现(生态适应性), 其组成是群落层片结构划分的主要依据, 群落中优势植物生活型决定了群落的外貌形态(高贤明和陈灵芝, 1998; 宋永昌, 2004; Raju et al., 2014).常用的植物生活型系统是Raunkiaer分类系统(Raunkiaer, 1934).该系统认为植物在不良季节休眠芽所在的位置是表达其适应不利季节的最好方式(Pavón et al., 2000; Odland, 2009).由于植物长期对区域综合自然环境条件的趋同适应, 植物生活型谱可以较清楚地反映出当地的气候特征, 并与植被类型有着密切的联系.不同地区相同群落类型由于环境条件的差异形成不同的植物生活型谱(王梅峒, 1987; 郭泉水等, 1999; Vashistha et al., 2011; Amjad et al., 2016); 同一地区处于不同演替阶段的植物群落, 生活型谱也有差异(张光富和宋永昌, 2001; 雷泞菲等, 2002; Hedwall et al., 2015).因此, 分析植被生活型组成特点及其地理格局有助于我们认识区域气候环境特点及其地域分异规律, 理解植物对环境的适应策略(郭柯等, 1998; Chen et al., 2005; Raju et al., 2014). ...
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
Very high resolution interpolated climate surfaces for global land areas 1 2005
Environmental assessment of climate of a habitat through floristic life-form spectra, a case study of Warangal north forest division, Telangana, India 4 2014
... 生活型是植物通过对环境条件长期适应, 在其生理、结构, 尤其是外部形态上的一种具体表现(生态适应性), 其组成是群落层片结构划分的主要依据, 群落中优势植物生活型决定了群落的外貌形态(高贤明和陈灵芝, 1998; 宋永昌, 2004; Raju et al., 2014).常用的植物生活型系统是Raunkiaer分类系统(Raunkiaer, 1934).该系统认为植物在不良季节休眠芽所在的位置是表达其适应不利季节的最好方式(Pavón et al., 2000; Odland, 2009).由于植物长期对区域综合自然环境条件的趋同适应, 植物生活型谱可以较清楚地反映出当地的气候特征, 并与植被类型有着密切的联系.不同地区相同群落类型由于环境条件的差异形成不同的植物生活型谱(王梅峒, 1987; 郭泉水等, 1999; Vashistha et al., 2011; Amjad et al., 2016); 同一地区处于不同演替阶段的植物群落, 生活型谱也有差异(张光富和宋永昌, 2001; 雷泞菲等, 2002; Hedwall et al., 2015).因此, 分析植被生活型组成特点及其地理格局有助于我们认识区域气候环境特点及其地域分异规律, 理解植物对环境的适应策略(郭柯等, 1998; Chen et al., 2005; Raju et al., 2014). ...
... ; Raju et al., 2014). ...
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
... ; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
4 1934
... 生活型是植物通过对环境条件长期适应, 在其生理、结构, 尤其是外部形态上的一种具体表现(生态适应性), 其组成是群落层片结构划分的主要依据, 群落中优势植物生活型决定了群落的外貌形态(高贤明和陈灵芝, 1998; 宋永昌, 2004; Raju et al., 2014).常用的植物生活型系统是Raunkiaer分类系统(Raunkiaer, 1934).该系统认为植物在不良季节休眠芽所在的位置是表达其适应不利季节的最好方式(Pavón et al., 2000; Odland, 2009).由于植物长期对区域综合自然环境条件的趋同适应, 植物生活型谱可以较清楚地反映出当地的气候特征, 并与植被类型有着密切的联系.不同地区相同群落类型由于环境条件的差异形成不同的植物生活型谱(王梅峒, 1987; 郭泉水等, 1999; Vashistha et al., 2011; Amjad et al., 2016); 同一地区处于不同演替阶段的植物群落, 生活型谱也有差异(张光富和宋永昌, 2001; 雷泞菲等, 2002; Hedwall et al., 2015).因此, 分析植被生活型组成特点及其地理格局有助于我们认识区域气候环境特点及其地域分异规律, 理解植物对环境的适应策略(郭柯等, 1998; Chen et al., 2005; Raju et al., 2014). ...
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slope of Qilianshan Mountains, Gansu, China 3 2003
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
... ; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
... ; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
中国亚热带常绿阔叶林生活型的研究 5 1987
... 生活型是植物通过对环境条件长期适应, 在其生理、结构, 尤其是外部形态上的一种具体表现(生态适应性), 其组成是群落层片结构划分的主要依据, 群落中优势植物生活型决定了群落的外貌形态(高贤明和陈灵芝, 1998; 宋永昌, 2004; Raju et al., 2014).常用的植物生活型系统是Raunkiaer分类系统(Raunkiaer, 1934).该系统认为植物在不良季节休眠芽所在的位置是表达其适应不利季节的最好方式(Pavón et al., 2000; Odland, 2009).由于植物长期对区域综合自然环境条件的趋同适应, 植物生活型谱可以较清楚地反映出当地的气候特征, 并与植被类型有着密切的联系.不同地区相同群落类型由于环境条件的差异形成不同的植物生活型谱(王梅峒, 1987; 郭泉水等, 1999; Vashistha et al., 2011; Amjad et al., 2016); 同一地区处于不同演替阶段的植物群落, 生活型谱也有差异(张光富和宋永昌, 2001; 雷泞菲等, 2002; Hedwall et al., 2015).因此, 分析植被生活型组成特点及其地理格局有助于我们认识区域气候环境特点及其地域分异规律, 理解植物对环境的适应策略(郭柯等, 1998; Chen et al., 2005; Raju et al., 2014). ...
... 植物生活型的地理格局(即地带性规律), 反映了植物群落物种组成在经度、纬度或海拔梯度上的变化规律, 是水分、热量及水热综合作用的结果(刘守江等, 2003).目前, 大部分关于植物生活型的研究多在热带、亚热带、温带以及干旱半干旱地区地带性植被中开展(王梅峒, 1987; 高贤明和陈灵芝, 1998; Wang et al., 2003; 胡正华等, 2004; Raju et al., 2014; Hedwall et al., 2015).众多研究发现, 随着纬度和海拔的增加, 高位芽植物和一年生植物比例逐渐减小, 地面芽植物、地下芽植物相应地增多(Raunkiaer, 1934; 郭泉水等, 1999; Wang et al., 2003; Mahdavi et al., 2012); 亦有一年生植物随海拔梯度逐渐增加的报道(Pavón et al., 2000).在同一植被区域内, 由于植被类型和环境因子差异不大, 生活型谱在纬度和海拔梯度上没有显著差异, 但随着植被演替阶段的正向更替, 高位芽植物的比例呈递增趋势, 地面芽植物、地下芽和一年生植物有所减少(雷泞菲等, 2002).造成这种差异的主要原因与各生活型植物对水热梯度的响应方式(郭柯等, 1998; Swaine & Becker, 1999; Chen et al., 2005; Raju et al., 2014), 以及与各研究区域内受限制的环境梯度(郭泉水等, 1999; Mahdavi et al., 2012)有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...
... )有关.以往基于局域群落或较大范围的全部维管植物生活型的研究不仅反映了区域环境特点, 而且得到生活型谱随环境梯度变化的定性结果(王梅峒, 1987; 郭泉水等, 1999; Wang et al., 2003; Batalha & Martins, 2004), 但特定生态系统在较大地理范围内的定量研究结果较为少见, 尤其是湿润地区非地带性优势植被类型(如灌丛)的资料极为缺乏. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT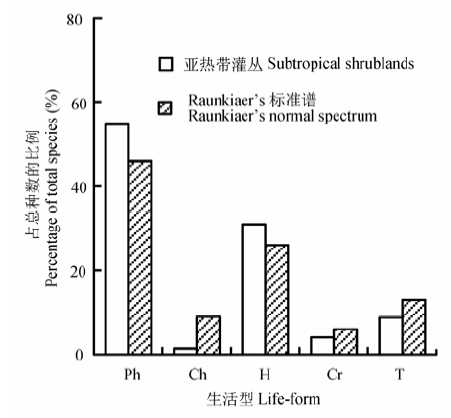 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT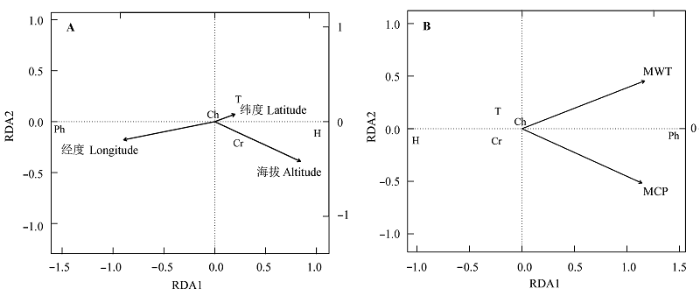 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT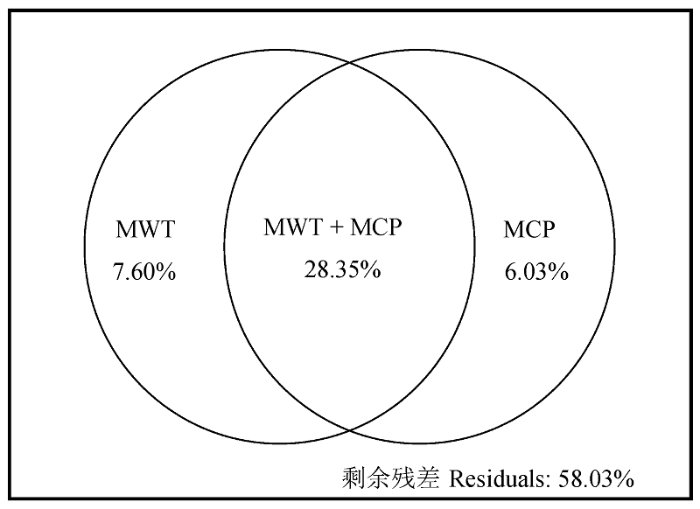 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}