Soil water use strategy of dominant species in typical natural and planted shrubs in loess hilly region
LÜTing1, ZHAOXi-Ning2,3,*,, GAOXiao-Dong2,3, PANYan-Hui4 1College of Water Resources and Architectural Engineering, Northwest A & F University, Yangling, Shaanxi 712100, China2Institute of Soil and Water Conservation, Northwest A & F University, Yangling, Shaanxi 712100, China3Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling, Shaanxi 712100, China4Key Laboratory of Western China’s Environmental Systems of Ministry of Education, Lanzhou University, Lanzhou 730000, China 通讯作者:* 通信作者Author for correspondence (E-mail:xiningz@aliyun.com; zxnsbs@gmail.com)* 通信作者Author for correspondence (E-mail:xiningz@aliyun.com; zxnsbs@gmail.com) 收稿日期:2016-08-1 接受日期:2016-12-25 网络出版日期:2017-02-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
关键词:氧稳定同位素;水分来源;MixSIR模型;细裂叶莲蒿;柠条锦鸡儿 Abstract AimsArtemisia gmelinii is a dominant specie naturally established after abandonment of cultivated lands in the Loess Plateau, and Caragana korshinskii is one of the main planted shrub species to control soil erosion. Improved understanding of water use strategies of these two species is of great significance to evaluate the sustainable development of the Loess Plateau under the trend of climate warming and increasing drought events. Methods Stable oxygen-18 isotope was used to determine seasonal variations in the water sources of native A. gmelinii communities established after abandonment of cultivated lands for 7 and 30 years and planted C. korshinskii after 30 years. The contributions of soil water from different depths to water uptake were estimated by the MixSIR Bayesian mixing model. The geometric mean regression method was used to fit the line of precipitation to get the local meteoric water line (LWML). Important findings The stable hydrogen isotope rate (δD) and stable oxygen isotope rate (δ18O) of soil water and xylem water plotted to the right side of the LWML, indicating that the isotopic compositions of soil water were enriched due to evaporation. The native A. gmelinii communities established after abandonment of cultivated lands for 7 years and planted C. korshinskii after 30 years showed plasticity in switching water sources from different soil layers, extracting water from shallow soil (0-40 cm) when soil water was available, but deeper soil (40-80 cm) when shallow soil water was dry. In contrast, A. gmelinii growing in site after cultivation abandonment for 30 years mainly relied on water from the surface soil (0-10 cm) throughout the growing season. Our results suggest that the ability of A. gmelinii to compete for soil water reduces with aging of the community while the planted C. korshinskii will have competitive advantage under the condition of increasing frequency of drought events in the future.
Keywords:oxygen stable isotopes;water-use source;MixSIR model;Artemisia gmelinii; Caragana korshinskii -->0 PDF (629KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 吕婷, 赵西宁, 高晓东, 潘燕辉. 黄土丘陵区典型天然灌丛和人工灌丛优势植物土壤水分利用策略. 植物生态学报, 2017, 41(2): 175-185 https://doi.org/10.17521/cjpe.2016.0253 LÜTing, ZHAOXi-Ning, GAOXiao-Dong, PANYan-Hui. Soil water use strategy of dominant species in typical natural and planted shrubs in loess hilly region. Chinese Journal of Plant Ecology, 2017, 41(2): 175-185 https://doi.org/10.17521/cjpe.2016.0253 细裂叶莲蒿(Artemisia gmelinii), 俗称铁杆蒿或万年蒿, 为多年生半灌木草本植物, 是黄土丘陵区植被自然恢复过程中重要的植物群落优势种, 也是黄土丘陵沟壑区天然草地的主要建群种(周萍等, 2008)。柠条锦鸡儿(Caragana korshinskii)属豆科锦鸡儿属植物, 其适应性强, 根系极为发达, 是黄土丘陵区水土保持和生态防护首选的灌木(宁婷, 2014)。近期研究表明黄土高原气候朝暖干化趋势发展, 极端气候事件发生频率的增大引起了降水量减少和降水年内分配不均(张宝庆等, 2012), 这将导致黄土丘陵区水文循环过程发生变化。天然植被和人工植被是SPAC循环过程主要的调控者(赵文智和程国栋, 2001; 赵西宁等, 2005), 研究黄土丘陵区天然灌丛优势种细裂叶莲蒿和主要人工灌木柠条锦鸡儿的土壤水分利用策略对评价气候变化环境下该区域生态建设可持续性具有重要意义。 氢氧稳定同位素示踪技术是评估植物水分利用策略的一种有效和精确的手段(Ehleringer & Dawson, 1992)。研究表明, 植物根系在获取土壤水分及向木质部运输过程中不发生同位素分馏(Brunel et al., 1995), 因此通过比较植物茎水与各潜在水源的氢氧稳定同位素值就可以确定植物的水分来源(Dawson et al., 2002)。二源或三源线性混合模型可快速估算各水源对植物的贡献率, 当超过3个水分来源时, 二源或三源线性模型不能提供唯一解, 基于同位素质量平衡原理, Phillips和Gregg (2003)给出了IsoSource混合模型来确定多源贡献的范围, IsoSource混合模型在识别作物的水分来源方面得到了广泛应用(Wang et al., 2010; 郑肖然等, 2015)。但是IsoSource模型没有融合来源的不确定性分析, 不能提供更严谨的模型估算。近几年来, 科学家们开发了基于贝叶斯统计的混合模型软件包MixSIR (Moore & Semmens, 2008)。相比之下, 贝叶斯混合模型在量化植物水源上更具优势, 它可以集合所有可能的多种来源的同位素特征值及同位素数据的不确定度, 并能融合先验信息, 使模型估计更为严谨准确(Leng et al., 2013; Yang et al., 2015a, 2015b)。 本文主要研究目的是通过氧稳定同位素和MixSIR模型相结合的方法探讨黄土丘陵区典型天然灌丛细裂叶莲蒿(退耕7年和30年)和柠条锦鸡儿(退耕30年)的土壤水分利用策略。通过分析典型植物吸水深度的季节性变化, 探讨黄土高原丘陵植物的水分适应机制, 为黄土丘陵区退化生态系统的植被恢复、植物资源的有效利用提供决策依据。
1 材料和方法
1.1 试验地点和物种
研究区位于陕西省清涧县店则沟镇园则沟小流域(37.23° N, 110.35° E)。 该区为典型的黄土丘陵沟壑区, 具有显著的温带大陆性季风气候, 年平均气温8.6 ℃, 月平均最低气温-6.5 ℃ (1月), 月平均最高气温22.8 ℃ (7月), 年降水量505 mm, 季节分配不均, 70%的降水集中在7-9月。土壤主要为黄绵土, 属于粉质壤土。研究区内不同土层土壤理化性质不同, 土壤颗粒均以粉粒为主(表1), 具有较强的入渗能力, 土壤总有机碳(TOC)和全氮(total N)含量在0-20 cm土层最高。植被组成简单, 群落郁闭度低, 分布稀疏, 以不同年限的细裂叶莲蒿(天然灌丛优势建群种)和柠条锦鸡儿(人工灌木林主要建群种)为主。 Table 1 表1 表1不同土层的颗粒大小及土壤养分(平均值±标准偏差) Table 1Particle size distribution and major nutrients in different soil layers (mean ± SD)
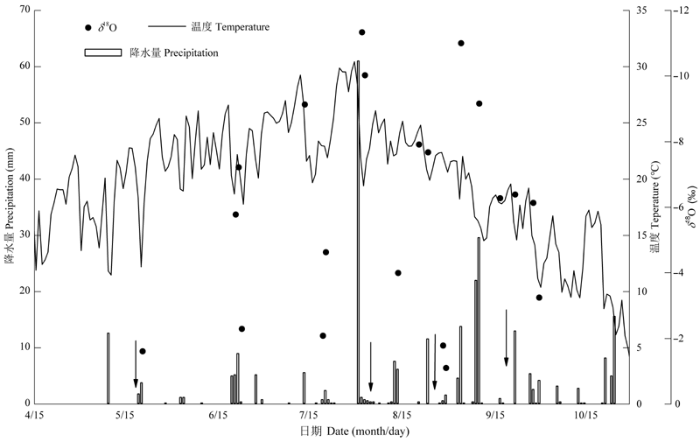
2015年5-9月降水量、降水δ18O及平均每日温度随时间的变化如图1所示。2015年研究区内5-9月总降水量为283.6 mm, 降水主要集中在8-9月, 占全年总降水量的70.9%, 且80%以上的降水事件降水量小于5 mm, 最大单次降水量为8月1日的61 mm。降水δ18O值在-11.33‰- -1.61‰范围内变化, 波动范围较大, 并呈现大降水事件同位素组成较贫化、小降水事件同位素富集的现象。 显示原图|下载原图ZIP|生成PPT 图12015年5-9月降水量、降水氧稳定同位素比率(δ18O)值及平均每日温度变化。实心圆点代表δ18O平均值, 向下的箭头代表采样日期。 -->Fig. 1Temporal variations of precipitation amount, isotopic compositions and daily temperature during May to September in 2015. The circles represent the means of stable oxygen isotope rate (δ18O), and the downward arrows represent sampling dates. -->
图2显示了土壤水分、植物茎水和降水的δD-δ18O的线性相关关系。对黄土丘陵区降水样品的氢氧稳定同位素组成进行回归分析, 得到当地大气降水线(LMWL: δD = 6.35δ18O + 4.85, R2 = 0.866, p < 0.001), 相对于全球降水线(Rozanski et al., 1993)(GMWL: δD = 8δ18O + 10), LMWL有较低的斜率和截距, 表明当地降水受蒸发影响, 这与黄土丘陵区延安降水线方程基本一致。在δD-δ18O分布图上, 土壤水分线(SWL: δD = 3.65δ18O-34.57, R2 = 0.658, p < 0.01)基本绘制在当地降水线的右下方, 斜率和截距都显著低于降水线, 表明土壤水分经历了强烈的二次蒸发。植物茎水的δD值和δ18O值均落在土壤水氢氧同位素分布区间, 表明了7年细裂叶莲蒿、30年细裂叶莲蒿和30年柠条锦鸡儿主要利用不同土层土壤水分, 并且利用深度存在明显差异。 显示原图|下载原图ZIP|生成PPT 图22015年研究区大气降水(空心正方形)、土壤水分和植物水的δD-δ18O的关系。当地大气降水线用虚线表示, 土壤水线用短点划线表示。δ18O, 氧稳定同位素比率; δD, 氢稳定同位素比率。 -->Fig. 2Values of δD as a function of δ18O for precipitation (open square), soil water and xylem water in 2015 at the experimental site. The local precipitation line is indicated by a dashed line, soil water line is indicated by a short dash-dotted line. δ18O, stable oxygen isotope rate; δD, stable hydrogen isotope rate. -->
2.2 土壤含水量和土壤水18O同位素分布
从图3中可以看出, 7年和30年细裂叶莲蒿土壤水分随降水的变化, 不同月份之间表现出相似的规律性, 5月为细裂叶莲蒿的生长初期, 由于采样之前有连续9天的无雨期, 导致表层土壤含水量低, 40-120 cm土层土壤水分高于0-40 cm土层土壤水分, 而到了细裂叶莲蒿生长旺盛期(8月初), 随着降雨量的增加, 0-40 cm土壤水分得以恢复至较高水平, 40 cm以下土层土壤水分急剧下降, 直至接近土壤凋萎湿度7%。到8月25日, 30年细裂叶莲蒿0-40 cm土层土壤含水率下降到7%, 而40-120 cm土层土壤水分基本不变。9月在细裂叶莲蒿生长后期, 在降水的影响下, 土壤水分含量迅速升高, 0-40 cm土层土壤水分恢复较快, 80-120 cm土层土壤水分仍然没有得到恢复。30年柠条锦鸡儿土壤水分的月变化类似于细裂叶莲蒿, 8月初的大降雨短期提高了0-40 cm土层的土壤水分, 随后在强烈的蒸散作用下土壤含水量逐渐减小, 9月份0-40 cm土层的土壤水分得到恢复, 8月份和9月份80-120 cm土层的土壤含水量维持在较低水平(5.78%), 均低于土壤凋萎湿度。 显示原图|下载原图ZIP|生成PPT 图3不同深度土壤质量含水量和土壤水分的氧稳定同位素比率(δ18O) (平均值±标准误差, n = 4)。A、D, 退耕7年的细裂叶莲蒿。B、E, 退耕30年的细裂叶莲蒿。C、F, 退耕30年的柠条锦鸡儿。 -->Fig. 3Soil water content and stable oxygen isotope rate (δ18O) of soil water at different depths (mean ± SE, n = 4). A, D, Artemisia gmelinii from site abandoned from cultivation 7 years ago. B, E, Artemisia gmelinii from site abandoned from cultivation 30 years ago. C, F, Caragana korshinskii from site abandoned from cultivation 30 years ago. -->
7年细裂叶莲蒿、30年细裂叶莲蒿和30年柠条锦鸡儿样地土壤水分δ18O随采样时间和采样深度而变化(图3D、3E、3F)。0-10 cm土层δ18O受近期贫化的降水同位素值和蒸发富集的影响而高度变化, 显著不同于其他土层(p < 0.01)。与此相反, 除7年细裂叶莲蒿样地外, 30年细裂叶莲蒿和30年柠条锦鸡儿样地40 cm以下土层土壤水分的δ18O月份间无显著性差异(30年细裂叶莲蒿样地n = 4, F < 1.16, p > 0.112; 柠条锦鸡儿样地n = 4, F < 5.419, p > 0.012)。7年细裂叶莲蒿、30年细裂叶莲蒿和柠条锦鸡儿样地80 cm以下土壤水分同位素值相对一致, 分别为-7.79‰、-7.14‰和-8.13‰。
2.3 植物茎18O同位素与土壤水分利用来源
图2显示所有植物木质部同位素值都分布在土壤水氢氧同位素值分布区间, 表明植物水源可能主要来源于不同土层的土壤水分。7年细裂叶莲蒿和30年柠条锦鸡儿的木质部同位素值主要在0-10 cm、10-40 cm和40-80 cm土层土壤水同位素值区间波动, 而30年细裂叶莲蒿木质部同位素值几乎分布在0-10 cm土层土壤水同位素值周围。植物种类、采样时间和植物种类×采样时间的交互作用对木质部同位素值均有显著影响(p < 0.01)。从表3可以看到, 不同植物木质部同位素在相同采样日期, 同一植物木质部同位素值在不同采样日期差异显著, 30年细裂叶莲蒿木质部δ18O值显著高于30年柠条锦鸡儿, 表明它吸收更多的表层土壤水。 Table 3 表3 表3退耕7年的细裂叶莲蒿、退耕30年的细裂叶莲蒿和退耕30年的柠条锦鸡儿在不同采样日期内木质部氧稳定同位素比率(δ18O) (平均值±标准偏差) Table 3Stable oxygen isotope rate (δ18O) of plant xylem values for the five sample collection periods for Artemisia gmelinii from site abandoned from cultivation 7 years ago, Artemisia gmelinii and planted Caragana korshinskii from site abandoned from cultivation 30 years ago (mean ± SD)
植物种类(退耕年限) Plant species (years since cultivation abandonment)
采样时间 Sample date
2015-5-19
2015-8-5
2015-8-25
2015-9-19
细裂叶莲蒿 Artemisia gmelinii (7 a)
-4.37 ± 1.84aA
-9.85 ± 0.97aB
-
-8.00 ± 1.60aC
细裂叶莲蒿 Artemisia gmelinii (30 a)
-6.39 ± 1.11bA
-9.37 ± 0.49aB
-6.65 ± 0.44aA
-7.02 ± 1.01abA
柠条锦鸡儿 Caragana korshinskii (30 a)
-
-7.35 ± 0.59bA
-4.71 ± 0.67bB
-6.18 ± 0.70bC
Different uppercase letters represent significant level of the same species among the different sampling dates (independent-sample t- test and LSD-t are used to test). Different lowercase letters represent significant level of different species within the same sampling date.不同大写字母代表相同植物种类在不同采样日期的显著性差异(样本数组只有两个时进行独立样本t检验, 样本数量>2时, 进行LSD两两比较), 不同小写字母代表不同植物种类在同一采样日期的显著性差异。 新窗口打开 用MixSIR模型计算植物对不同水分来源(0- 10 cm、10-40 cm、40-80 cm和80-120 cm土层)的吸水比例, 从图4可以看出, 不同植物的吸水比例随生长进程显示出季节性差异。7年细裂叶莲蒿的吸水模式表现出很强的可塑性, 水源由生长初期(5月19日)的40-80 cm转移到生长中期(8月5日)的0-10 cm, 生长末期(9月19日)再次吸收40-80 cm土层的水分, 吸水比例高达88.0%。与7年细裂叶莲蒿相反, 30年细裂叶莲蒿在整个生长季节主要吸收0-10 cm土层的土壤水分。 显示原图|下载原图ZIP|生成PPT 图4不同植物对4个潜在水源利用比例的季节变化(平均值±标准偏差)。A, 退耕7年的细裂叶莲蒿。B, 退耕30年的细裂叶莲蒿。C, 退耕30年的柠条锦鸡儿。 -->Fig. 4Seasonal variations in fraction of uptake from four potential soil water sources for different plants (mean ± SD). A, Artemisia gmelinii from site abandoned from cultivation 7 years ago. B, Artemisia gmelinii from site abandoned from cultivation 30 years ago. C, Caragana korshinskii from site abandoned from cultivation 30 years ago. -->
AsbjornsenH, ShepherdG, HelmersM, MoraG (2008). Seasonal patterns in depth of water uptake under contrasting annual and perennial systems in the corn belt region of the midwestern US. Plant and Soil, 308, 69-92. [本文引用: 1]
[2]
BrunelJP, WalkerGR, Kennett-SmithAK (1995). Field validation of isotopic procedures for determining sources of water used by plants in a semi-arid environment . Journal of Hydrology, 167, 351-368. [本文引用: 1]
[3]
CaiM, HanRL, JinSJ, GuoY, LiangZS (2012). Effect of soul drought on growth and water consumption of two subshrub species in Loess Plateau . Journal of Northwest Forestry University, 27, 26-32. (in Chinese with English abstract)[蔡马, 韩蕊莲, 靳淑静, 郭颖, 梁宗锁 (2012). 土壤干旱对黄土高原2种半灌木植物生长与耗水规律的影响 . 西北林学院学报, 27, 26-32.] [本文引用: 1]
[4]
ChenJ, XuQ, GaoDQ, SongAY, HaoYG, MaYB (2017). Differential water use strategies among selected rare and endangered species in West Ordos Desert of China. Journal of Plant Ecology, 10, in press. doi: 10.1093/jpe/rtw-059. [本文引用: 1]
[5]
ChenXL, ChenYN, ChenYP (2014). Relationship among water use of different plants in Heihe River riparian forests. Chinese Journal of Eco-Agriculture, 22, 927-979. (in Chinese with English abstract)[陈小丽, 陈亚宁, 陈亚鹏 (2014). 黑河下游荒漠河岸林植物水分利用关系研究 . 中国生态农业学报, 22, 972-979.] [本文引用: 1]
[6]
ChengXL, AnSQ, LiB, ChenGQ, LinGH, LiuYH, LuoYQ, LiuSR (2006). Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China. Plant Ecology, 184, 1-12. [本文引用: 1]
[7]
DaiY, ZhengXJ, TangLS, LiY (2014). Dynamics of water usage in Haloxylon ammodendron in the southern edge of the Gurbantünggüt Desert . Chinese Journal of Plant Ecology, 38, 1214-1225. (in Chinese with English abstract)[戴岳, 郑新军, 唐立松, 李彦 (2014). 古尔班通古特沙漠南缘梭梭水分利用动态 . 植物生态学报, 38, 1214-1225.] [本文引用: 1]
[8]
DaiY, ZhengXJ, TangLS, LiY (2015). Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantünggüt Desert. Plant and Soil, 389, 73-87. [本文引用: 2]
[9]
DawsonTE, MambelliS, PlamboeckAH, TemplerP, TuKP (2002). Stable isotopes in plant ecology. Annual Review of Ecology and Systematics, 33, 507-559. [本文引用: 1]
[10]
EhleringerJR, DawsonTE (1992). Water uptake by plants: Perspectives from stable isotope composition. Plant, Cell & Environment, 15, 1073-1082. [本文引用: 1]
[11]
EllsworthPZ, WilliamsDG (2007). Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant and Soil, 291, 93-107. [本文引用: 1]
[12]
GaoXD (2013). Catchment-Scale Available Soil Moisture Spatial-Temporal Variability in the Hilly Areas of the Loess Plateau . PhD dissertation, Graduate University of Chinese Academy of Sciences and Ministry of Education Research Center of Soil and Water Conservation and Ecological Environment, Yangling, Shaanxi. 35-64. (in Chinese with English abstract)[高晓东 (2013). 黄土丘陵区小流域土壤有效水时空变异与动态模拟研究 . 博士学位论文, 中国科学院研究生院教育部水土保持与生态环境研究中心, 陕西杨凌. 35-64.] [本文引用: 1]
[13]
HuXM, ChenJM, WanHE, ZhaoYY, XuXB, GuXJ, LiWJ (2006). Effect of short-term watering on photosynthetic characteristics and chlorophyll fluorescence of Artemisia sacrorum under long-term drought. Acta Agrestia Sinica, 14, 236-241. (in Chinese with English abstract)[胡相明, 程积民, 万惠娥, 赵艳云, 徐宣斌, 古晓林, 李维军 (2006). 短期施水对长期干旱细裂叶莲蒿光合作用和叶绿素荧光特征的影响 . 草地学报, 14, 236-241.] [本文引用: 1]
[14]
LengX, CuiJ, ZhangS, ZhangW, LiuY, LiuS, AnS (2013). Differential water uptake among plant species in humid alpine meadows. Journal of Vegetation Science, 24, 138-147. [本文引用: 2]
[15]
LiuW, WangP, LiJ, LiuW, LiH (2014). Plasticity of source water acquisition in epiphytic, transitional and terrestrial growth phases of Ficus tinctoria. Ecohydrology, 7, 1524-1533. [本文引用: 1]
[16]
MengXJ, WenXF, ZhangXY, HanJY, SunXM, LiXB (2012). Potential impacts of organic contaminant on δ18O and δD in leaf and xylem water detected by isotope ratio infrared spectroscopy. Chinese Journal of Eco-Agricul- ture, 20, 1359-1365. (in Chinese with English abstract)[孟宪菁, 温学发, 张心昱, 韩佳音, 孙晓敏, 李晓波 (2012). 有机物对红外光谱技术测定植物叶片和茎秆水δ18O和δD的影响 . 中国生态农业学报, 20, 1359-1365.] [本文引用: 1]
[17]
MooreJW, SemmensBX (2008). Incorporating uncertainty and prior information into stable isotope mixing models . Ecology Letters, 11, 470-480. [本文引用: 2]
[18]
NingT (2014). Study on Suitable Initial Planting Density of Caragana korshinskii Plantation in Semi-Arid Loess Hilly Region . PhD dissertation, Graduate University of Chinese Academy of Sciences and Ministry of Education Research Center of Soil and Water Conservation and Ecological Environment, Yangling, Shaanxi. 1-9. (in Chinese with English abstract)[宁婷 (2014). 半干旱黄土丘陵区人工柠条锦鸡儿林合理初植密度研究 . 博士学位论文, 中国科学院研究生院教育部水土保持与生态环境研究中心, 陕西杨凌. 1-9.] [本文引用: 1]
[19]
PhillipsDL, GreggJW (2003). Source partitioning using stable isotopes: Coping with too many sources. Oecologia, 136, 261-269. [本文引用: 1]
[20]
RozanskiK, Araguás-AraguásL, GonfiantiniR (1993). Isotopic patterns in modern global precipitation. Climate Change in Continental Isotopic Records, 78, 1-36. [本文引用: 1]
[21]
WangP, SongX, HanD, ZhangY, LiuX (2010). A study of root water uptake of crops indicated by hydrogen and oxygen stable isotopes: A case in Shanxi Province, China. Agricultural Water Management, 97, 475-482. [本文引用: 1]
[22]
XuGQ, LiY (2009). Roots distribution of three desert shrubs and their response to precipitation under co-occurring conditions. Acta Ecologica Sinica, 29, 130-137. (in Chinese with English abstract)[徐贵青, 李彦 (2009). 共生条件下三种荒漠灌木的根系分布特征及其对降水的响应 . 生态学报, 29, 130-137.] [本文引用: 1]
[23]
XuQ, LiHB, ChenJQ, ChengXL, LiuSR, ANSQ (2011). Water use patterns of three species in subalpine forest, Southwest China: The deuterium isotope approach. Ecohydrology, 4, 236-244. [本文引用: 1]
[24]
YangB, WenX, SunX (2015a). Irrigation depth far exceeds water uptake depth in an oasis cropland in the middle reaches of Heihe River Basin. Scientific Reports, 5, 12506. doi: 10.1038/srep15206. [本文引用: 2]
[25]
YangB, WenX, SunX (2015b). Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region. Agricultural and Forest Meteorology, 201, 218-228. [本文引用: 2]
[26]
YinL, ZhaoLJ, RuanYF, XiaoHL, ChengGD, ZhouMX, WangF, LiCZ (2012). Study of the replenishment sources of typical ecosystems water and dominant plant water in the lower reaches of the Heihe, China. Journal of Glaciology and Geocryology, 34, 1478-1486. (in Chinese with English abstract)[尹力, 赵良菊, 阮云峰, 肖洪浪, 程国栋, 周茅先, 王芳, 李彩芝 (2012). 黑河下游典型生态系统水分补给源及优势植物水分来源研究 . 冰川冻土, 34, 1478-1486.] [本文引用: 1]
[27]
ZhangBQ (2014). Study on Spatiotemporal Variability of Drought and Rainwater Harvesting Potential on the Chinese Loess Plateau . PhD dissertation, Northwest A&F University, Yangling, Shaanxi. 106-111. (in Chinese with English abstract)[张宝庆 (2014). 黄土高原干旱时空变异及雨水资源化潜力研究 . 博士学位论文, 西北农林科技大学, 陕西杨凌. 106-111.] [本文引用: 1]
[28]
ZhangBQ, WuPT, ZhaoXN, GaoXD (2012). Study on regional drought assessment based on variable infiltration capacity model and palmer drought severity index. Journal of Hydraulic Engineering, 43, 926-934. (in Chinese with English abstract)[张宝庆, 吴普特, 赵西宁, 高晓东 (2012). 基于可变下渗容量模型和Palmer干旱指数的区域干旱化评价研究 . 水利学报, 43, 926-934.] [本文引用: 1]
[29]
ZhaoWZ, ChengGD (2001). Comments on a number of issues of eco-hydrological processes in arid areas. Chinese Sci- ence Bulletin, 46, 1851-1857. (in Chinese with English abstract)[赵文智, 程国栋 (2001). 干旱区生态水文过程研究若干问题评述 . 科学通报, 46, 1851-1857.] [本文引用: 1]
[30]
ZhaoXN, WuPT, WangWZ, FengH (2005). Research ad- vance on eco-environmental water requirement. Advances in Water Science, 16, 617-622. (in Chinese with English abstract)[赵西宁, 吴普特, 王万忠, 冯浩 (2005). 生态环境需水研究进展 . 水科学进展, 16, 617-622.] [本文引用: 1]
[31]
ZhengXR, ZhaoGQ, LiXY, LiL, WuHW, ZhangSY, ZhangZH (2015). Application of stable hydrogen isotope in study of water sources for Caragana microphylla bushland in Nei Mongol. Chinese Journal of Plant Ecology, 39, 184-196. (in Chinese with English abstract)[郑肖然, 赵国琴, 李小雁, 李柳, 吴华武, 张思毅, 张志华 (2015). 氢同位素在内蒙古小叶锦鸡儿灌丛水分来源研究中的应用 . 植物生态学报, 39, 184-196.] [本文引用: 1]
[32]
ZhouH, ZhaoW, ZhengX, LiS (2015). Root distribution of Nitraria sibirica with seasonally varying water sources in a desert habitat. Journal of Plant Research, 128, 613-622. [本文引用: 1]
[33]
ZhouH, ZhengXJ, TangLS, LiY (2013). Differences and similarities between water sources of Tamarix ramosissima, Nitraria sibirica and Reaumuria soongorica in the southeastern Junggar Basin. Chinese Journal of Plant Ecology, 37, 665-673. (in Chinese with English abstract)[周海, 郑新军, 唐立松, 李彦 (2013). 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同 . 植物生态学报, 37, 665-673.] [本文引用: 1]
[34]
ZhouP, LiuGB, HouXL (2008). Study on vegetation and soil nutrient characters of Artemisia sacrorum communities in hilly-gully region of the Loess Plateau. Acta Prataculturae Sinica, 17, 9-18. (in Chinese with English abstract)[周萍, 刘国彬, 侯喜禄 (2008). 黄土丘陵区铁杆蒿群落植被特性及土壤养分特征研究 . 草业学报, 17, 9-18] [本文引用: 1]
[35]
ZhuL, QiYS, XuX (2014). Water sources of Medicago sativa grown in different slope positions in Yanchi County of Ningxia. Chinese Journal of Plant Ecology, 38, 1226-1240. (in Chinese with English abstract)[朱林, 祁亚淑, 许兴 (2014). 宁夏盐池不同坡位旱地紫苜蓿水分来源 . 植物生态学报, 38, 1226-1240.] [本文引用: 1]
Seasonal patterns in depth of water uptake under contrasting annual and perennial systems in the corn belt region of the midwestern US. 1 2008
 , 高晓东
, 高晓东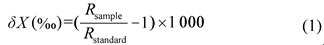
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT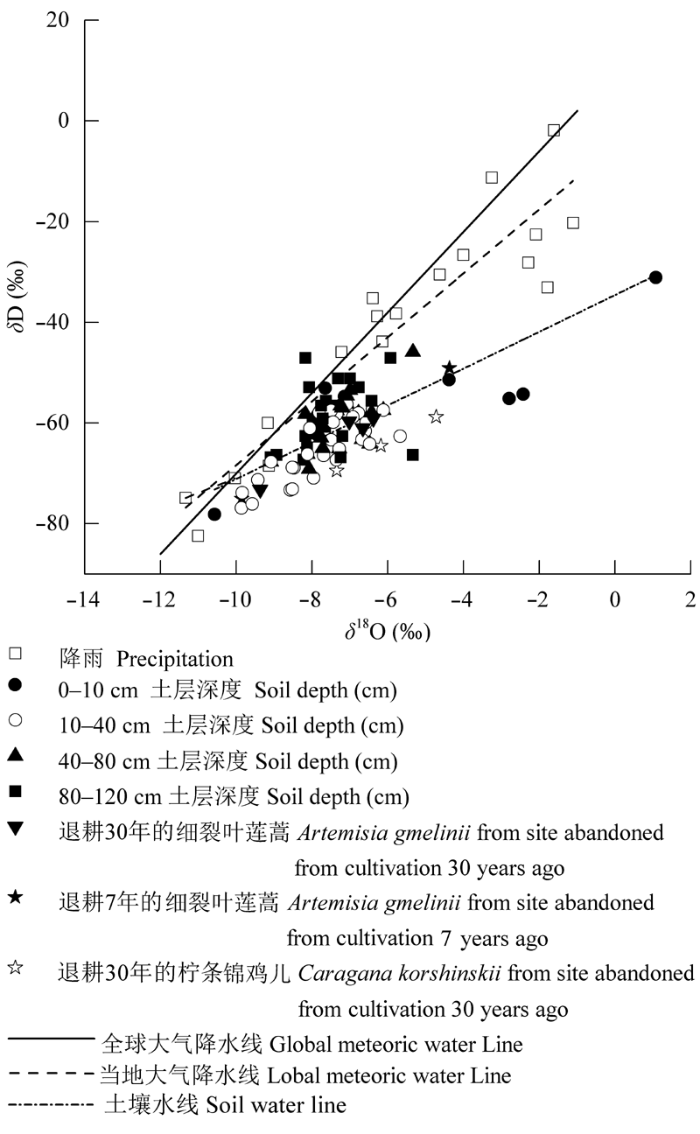 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT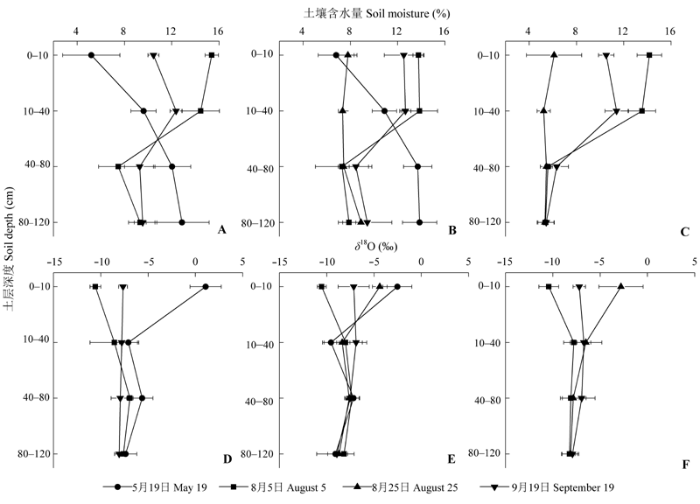 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT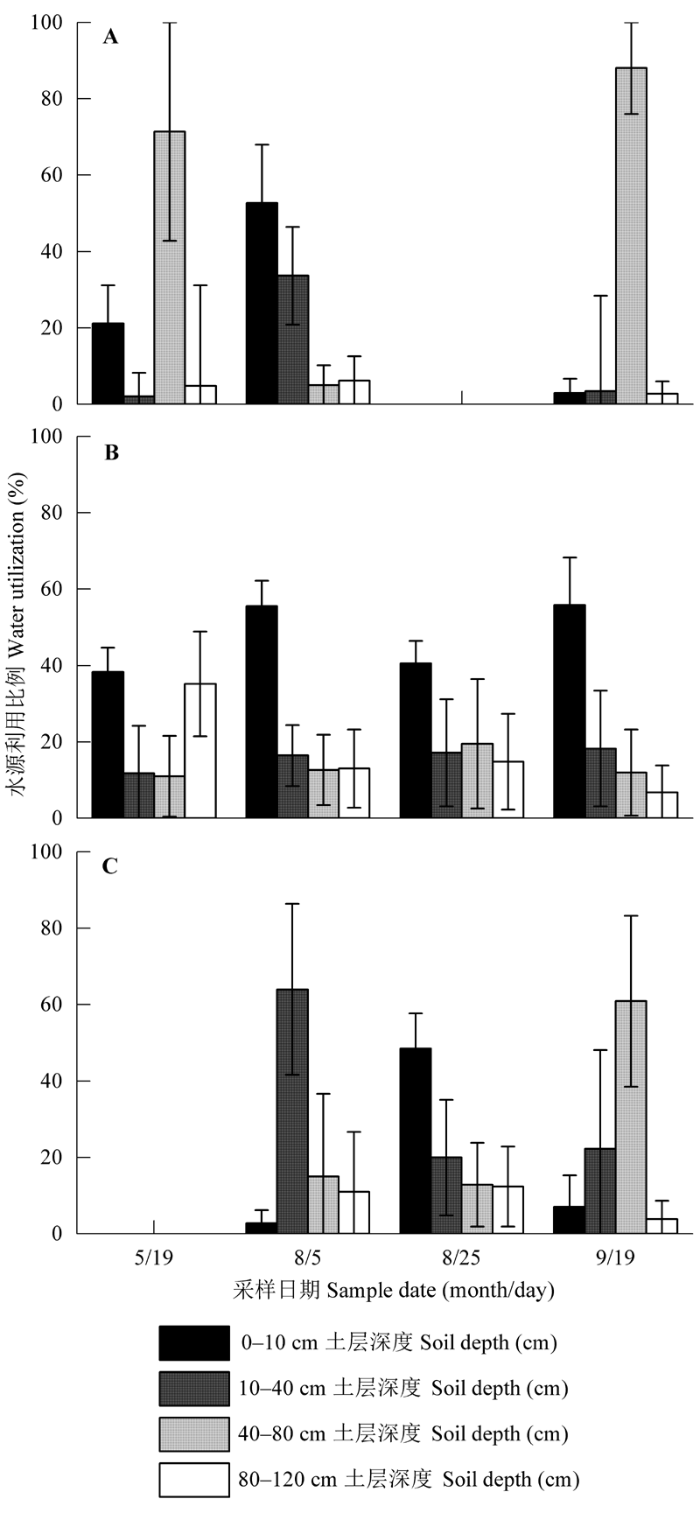 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}