Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China
LIJia-Xiang1,2, XUWen-Ting1, XIONGGao-Ming1, WANGYang1, ZHAOChang-Ming1, LUZhi-Jun3, LIYue-Lin4, XIEZong-Qiang1,*, 1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2College of Forest, Central South University of Forestry & Technology, Changsha 410004, China;3Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan 430074, Chinaand 4South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 通讯作者:* 通信作者Author for correspondence (E-mail: xie@ibcas.ac.cn) 责任编辑:LIJia-XiangXUWen-TingXIONGGao-MingWANGYangZHAOChang-MingLUZhi-JunLIYue-LinXIEZong-Qiang 收稿日期:2016-07-28 接受日期:2016-12-25 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略性先导科技专项 (XDA05050302)
关键词:热带亚热带;植物功能性状;植物生活型;生态化学计量 Abstract Aims Understanding the changes in N and P concentration in plant organs along the environmental gradients can provide meaningful information to reveal the underline mechanisms for the geochemical cycles and adaptation strategies of plants to the changing environment. In this paper, we aimed to answer: (1) How did the N and P concentration in leaves of evergreen and deciduous woody plants change along the environmental gradients? (2) What were the main factors regulating the N and P concentration in leaves of woody plants in the shrublands across southern China? Methods Using a stratified random sampling method, we sampled 193 dominant woody plants in 462 sites of 12 provinces in southern China. Leaf samples of dominant woody plants, including 91 evergreen and 102 deciduous shrubs, and soil samples at each site were collected. N and P concentration of the leaves and soils were measured after lapping and sieving. Kruskal-Wallis and Nemenyi tests were applied to quantify the difference among the organs and life-forms. For each life-form, the binary linear regression was used to estimate the relationships between leaf log [N] and log [P] concentration and mean annual air temperature (MAT), mean annual precipitation (MAP) and log soil total [N], [P]. The effects of climate, soil and plant life-form on leaf chemical traits were modeled through the general linear models (GLMs) and F-tests. Important findings 1) The geometric means of leaf N and P concentrations of the dominant woody plants were 16.57 mg·g-1 and 1.02 mg·g-1, respectively. The N and P concentration in leaves (17.91 mg·g-1, 1.14 mg·g-1) of deciduous woody plants was higher than those of evergreen woody plants (15.19 mg·g-1, 0.89 mg·g-1). The dependent of leaf P concentration on environmental (climate and soil) appeared more variable than N concentration. 2) Leaf N and P in evergreen woody plants decreased with MAT and but increased with MAP, whereas those in deciduous woody plants showed opposite trends. With increase in MAP, leaf P concentration decreased for both evergreen and deciduous woody plants. 3) Soil N concentration had no significant effect on both evergreen and deciduous woody plants. However, leaf P concentration of the tow increased significantly with soil P concentration. (4) GLMs showed that plant growth form explained 7.6% and 14.4% of variation in leaf N and P, respectively. MAP and soil P concentration contributed 0.8% and 16.4% of the variation in leaf P, respectively. These results suggested that leaf N was mainly influenced by plant growth form, while leaf P concentration was driven by soil, plant life-form, and climate at our study sites.
根据1:100万中国植被图(中国科学院中国植被图编辑委员会, 2007)所记载的我国南方灌丛分布范围, 采用分层随机抽样方法布设了462个样点, 样点之间间隔20 km以上。记录每个样点的经度和纬度信息, 对每个样点的灌丛群落优势木本植物, 分种选择3-5株收集叶片的混合样品, 所有植物样品带回实验室烘干、磨碎、过60目筛待测。同时, 每个样点用土钻采集0-10 cm表土层土壤样品3个(采样点间隔15 m), 将样品阴干后均匀混合、除根、研磨, 并过100目的分样筛待测(谢宗强和唐志尧, 2015)。所有调查和采集在2011-2013年的7-9月进行, 共采集优势木本植物53科122属193种(含种下等级), 包括91种常绿木本植物和102种落叶木本植物(附录I)。 Appendix I(1) 附录I(1) 附录I(1)中国南方灌丛优势木本植物名录及生活型 Appendix I(1)Plant catalogue and life form of dominant woody plants in shrublands across southern China
科名 Family
属名 Genera
种名 Species
生活型 Life form
八角枫科 Alangiaceae
八角枫属 Alangium
瓜木 Alangium platanifoliam
落叶 Deciduous
漆树科 Anacardiaceae
黄栌属 Cotinus
黄栌 Cotinus coggygria
落叶 Deciduous
漆树科 Anacardiaceae
盐肤木属 Rhus
盐肤木 Rhus chinensis
落叶 Deciduous
漆树科 Anacardiaceae
漆属 Toxicodendron
野漆 Toxicodendron succedaneum
落叶 Deciduous
番荔枝科 Annonaceae
假鹰爪属 Desmos
假鹰爪 Desmos chinensis
常绿 Evergreen
番荔枝科 Annonaceae
暗罗属 Polyalthia
细基丸 Polyalthia cerasoides
常绿 Evergreen
番荔枝科 Annonaceae
紫玉盘属 Uvaria
紫玉盘 Uvaria microcarpa
常绿 Evergreen
冬青科 Aquifoliaceae
冬青属 Ilex
秤星树 Ilex asprella
落叶 Deciduous
冬青科 Aquifoliaceae
冬青属 Ilex
刺叶冬青 Ilex bioritsensis
常绿 Evergreen
冬青科 Aquifoliaceae
冬青属 Ilex
冬青 Ilex chinensis
常绿 Evergreen
冬青科 Aquifoliaceae
冬青属 Ilex
大叶冬青 Ilex latifolia
常绿 Evergreen
冬青科 Aquifoliaceae
冬青属 Ilex
具柄冬青 Ilex pedunculosa
常绿 Evergreen
五加科 Araliaceae
楤木属 Aralia
楤木 Aralia chinensis
落叶 Deciduous
禾本科 Gramineae
箭竹属 Fargesia
箭竹 Fargesia spathacea
常绿 Evergreen
禾本科 Gramineae
刚竹属 Phyllostachys
篌竹 Phyllostachys nidularia
常绿 Evergreen
禾本科 Gramineae
刚竹属 Phyllostachys
金竹 Phyllostachys sulphurea
常绿 Evergreen
桦木科 Betulaceae
鹅耳枥属 Carpinus
鹅耳枥 Carpinus turczaninowii
落叶 Deciduous
紫葳科 Bignoniaceae
菜豆树属 Radermachera
菜豆树 Radermachera sinica
常绿 Evergreen
紫草科 Boraginaceae
基及树属 Carmona
基及树 Carmona micarophylla
常绿 Evergreen
忍冬科 Caprifoliaceae
六道木属 Abelia
糯米条 Abelia chinensis
落叶 Deciduous
忍冬科 Caprifoliaceae
荚蒾属 Viburnum
金佛山荚蒾 Viburnum chinshanense
常绿 Evergreen
忍冬科 Caprifoliaceae
荚蒾属 Viburnum
荚蒾 Viburnum dilatatum
落叶 Deciduous
忍冬科 Caprifoliaceae
荚蒾属 Viburnum
红荚蒾 Viburnum erubescens
常绿 Evergreen
忍冬科 Caprifoliaceae
荚蒾属 Viburnum
球核荚蒾 Viburnum propinquum
常绿 Evergreen
忍冬科 Caprifoliaceae
荚蒾属 Viburnum
烟管荚蒾 Viburnum utile
常绿 Evergreen
忍冬科 Caprifoliaceae
锦带花属 Weigela
半边月 Weigela japonica var. sinica
落叶 Deciduous
卫矛科 Celastraceae
南蛇藤属 Celastrus
南蛇藤 Celastrus orbiculatus
落叶 Deciduous
卫矛科 Celastraceae
美登木属 Maytenus
刺茶美登木 Maytenus variabilis
常绿 Evergreen
牛栓藤科 Connaraceae
红叶藤属 Rourea
红叶藤 Rourea minor
常绿 Evergreen
马桑科 Coriariaceae
马桑属 Coriaria
马桑 Coriaria nepalensis
落叶 Deciduous
山茱萸科 Cornaceae
鞘柄木属 Toricellia
角叶鞘柄木 Toricellia angulata
落叶 Deciduous
柿树科 Ebenaceae
柿树属 Diospyros
野柿 Diospyros kaki var. silvestris
落叶 Deciduous
杜鹃花科 Ericaceae
珍珠花属 Lyonia
小果珍珠花 Lyonia ovalifolia var. elliptica
落叶 Deciduous
杜鹃花科 Ericaceae
杜鹃属 Rhododendron
满山红 Rhododendron mariesii
落叶 Deciduous
杜鹃花科 Ericaceae
杜鹃属 Rhododendron
杜鹃 Rhododendron simsii
落叶 Deciduous
杜鹃花科 Ericaceae
越桔属 Vaccinium
南烛 Vaccinium bracteatum
常绿 Evergreen
杜鹃花科 Ericaceae
越桔属 Vaccinium
短尾越桔 Vaccinium carlesii
常绿 Evergreen
杜鹃花科 Ericaceae
越桔属 Vaccinium
江南越桔 Vaccinium mandarinorum
常绿 Evergreen
大戟科 Euphorbiaceae
山麻杆属 Alchornea
红背山麻杆 Alchornea trewioides
落叶 Deciduous
科名 Family
属名 Genera
种名 Species
生活型 Life form
大戟科 Euphorbiaceae
重阳木属 Bischofia
重阳木 Bischofia polycarpa
落叶 Deciduous
大戟科 Euphorbiaceae
黑面神属 Breynia
黑面神 Breynia fruticosa
常绿 Evergreen
大戟科 Euphorbiaceae
算盘子属 Glochidion
算盘子 Glochidion puberum
落叶 Deciduous
大戟科 Euphorbiaceae
野桐属 Mallotus
白背叶 Mallotus apelta
落叶 Deciduous
大戟科 Euphorbiaceae
野桐属 Mallotus
野桐 Mallotus japonicus var. floccosus
落叶 Deciduous
大戟科 Euphorbiaceae
叶下珠属 Phyllanthus
余甘子 Phyllanthus emblica
落叶 Deciduous
大戟科 Euphorbiaceae
乌桕属 Sapium
乌桕 Sapium sebiferum
落叶 Deciduous
壳斗科 Fagaceae
栗属 Castanea
茅栗 Castanea seguinii
落叶 Deciduous
壳斗科 Fagaceae
栲属 Castanopsis
苦槠 Castanopsis sclerophylla
常绿 Evergreen
壳斗科 Fagaceae
青冈属 Cyclobalanopsis
青冈 Cyclobalanopsis glauca
常绿 Evergreen
壳斗科 Fagaceae
青冈属 Cyclobalanopsis
细叶青冈 Cyclobalanopsis gracilis
常绿 Evergreen
壳斗科 Fagaceae
青冈属 Cyclobalanopsis
多脉青冈 Cyclobalanopsis multinervis
常绿 Evergreen
壳斗科 Fagaceae
青冈属 Cyclobalanopsis
曼青冈 Cyclobalanopsis oxyodon
常绿 Evergreen
壳斗科 Fagaceae
石栎属 Lithocarpus
石栎 Lithocarpus glaber
常绿 Evergreen
壳斗科 Fagaceae
栎属 Quercus
麻栎 Quercus acutissima
落叶 Deciduous
壳斗科 Fagaceae
栎属 Quercus
槲栎 Quercus aliena
落叶 Deciduous
壳斗科 Fagaceae
栎属 Quercus
小叶栎 Quercus chenii
落叶 Deciduous
壳斗科 Fagaceae
栎属 Quercus
白栎 Quercus fabri
落叶 Deciduous
壳斗科 Fagaceae
栎属 Quercus
短柄枹栎 Quercus serrata var. brevipetiolata
落叶 Deciduous
壳斗科 Fagaceae
栎属 Quercus
栓皮栎 Quercus variabilis
落叶 Deciduous
大风子科 Flacourtiaceae
山桂花属 Bennettiodendron
山桂花 Bennettiodendron leprosipes
常绿 Evergreen
大风子科 Flacourtiaceae
刺篱木属 Flacourtia
刺篱木 Flacourtia indica
落叶 Deciduous
大风子科 Flacourtiaceae
箣柊属 Scolopia
珍珠箣柊 Scolopia henryi
常绿 Evergreen
藤黄科 Guttiferae
黄牛木属 Cratoxylum
黄牛木 Cratoxylum cochinchinensis
常绿 Evergreen
藤黄科 Guttiferae
金丝桃属 Hypericum
金丝桃 Hypericum monogynum
落叶 Deciduous
金缕梅科 Hamamelidaceae
蜡瓣花属 Corylopsis
蜡瓣花 Corylopsis sinensis
落叶 Deciduous
金缕梅科 Hamamelidaceae
金缕梅属 Hamamelis
金缕梅 Hamamelis mollis
落叶 Deciduous
金缕梅科 Hamamelidaceae
枫香树属 Liquidambar
枫香树 Liquidambar formosana
落叶 Deciduous
金缕梅科 Hamamelidaceae
檵木属 Loropetalum
檵木 Loropetalum chinense
常绿 Evergreen
胡桃科 Juglandaceae
化香树属 Platycarya
化香树 Platycarya strobilacea
落叶 Deciduous
樟科 Lauraceae
山胡椒属 Lindera
乌药 Lindera aggregata
常绿 Evergreen
樟科 Lauraceae
山胡椒属 Lindera
香叶树 Lindera communis
常绿 Evergreen
樟科 Lauraceae
山胡椒属 Lindera
香叶子 Lindera fragrans
常绿 Evergreen
樟科 Lauraceae
山胡椒属 Lindera
山胡椒 Lindera glauca
落叶 Deciduous
樟科 Lauraceae
木姜子属 Litsea
山鸡椒 Litsea cubeba
落叶 Deciduous
樟科 Lauraceae
木姜子属 Litsea
潺槁木姜子 Litsea glutinosa
常绿 Evergreen
樟科 Lauraceae
木姜子属 Litsea
毛叶木姜子 Litsea mollis
落叶 Deciduous
樟科 Lauraceae
木姜子属 Litsea
木姜子 Litsea pungens
落叶 Deciduous
樟科 Lauraceae
木姜子属 Litsea
豺皮樟 Litsea rotundifolia var. oblongifolia
常绿 Evergreen
樟科 Lauraceae
木姜子属 Litsea
钝叶木姜子 Litsea veitchiana
常绿 Evergreen
樟科 Lauraceae
润楠属 Machilus
黄绒润楠 Machilus grijsii
常绿 Evergreen
樟科 Lauraceae
润楠属 Machilus
绒毛润楠 Machilus velutina
常绿 Evergreen
豆科 Leguminosae
金合欢属 Acacia
金合欢 Acacia farnesiana
落叶 Deciduous
豆科 Leguminosae
合欢属 Albizia
合欢 Albizia julibrissin
落叶 Deciduous
豆科 Leguminosae
合欢属 Albizia
山槐 Albizia kalkora
落叶 Deciduous
豆科 Leguminosae
紫穗槐属 Amorpha
紫穗槐 Amorpha fruticosa
落叶 Deciduous
豆科 Leguminosae
羊蹄甲属 Bauhinia
龙须藤 Bauhinia championii
常绿 Evergreen
豆科 Leguminosae
羊蹄甲属 Bauhinia
羊蹄甲 Bauhinia purpurea
落叶 Deciduous
豆科 Leguminosae
云实属 Caesalpinia
云实 Caesalpinia decapetala
落叶 Deciduous
豆科 Leguminosae
杭子梢属 Campylotropis
杭子梢 Campylotropis macrocarpa
落叶 Deciduous
豆科 Leguminosae
黄檀属 Dalbergia
黄檀 Dalbergia hupeana
落叶 Deciduous
豆科 Leguminosae
黄檀属 Dalbergia
象鼻藤 Dalbergia mimosoides
落叶 Deciduous
豆科 Leguminosae
木蓝属 Indigofera
苏木蓝 Indigofera carlesii
落叶 Deciduous
豆科 Leguminosae
木蓝属 Indigofera
马棘 Indigofera pseudotinctoria
落叶 Deciduous
新窗口打开 Appendix I(2) 附录I(2) 附录I(2)中国南方灌丛优势木本植物名录及生活型 Appendix I(2)Plant catalogue and life form of dominant woody plants in shrublands across southern China
基于原始数据, 按种统计叶N、P含量的算术平均值, 并进行正态性检验, 叶N、P含量呈非正态分布(偏度>1.6), 但符合对数正态分布, 因此在统计各生活型(常绿和落叶)和所有木本叶片N含量、P含量时采用几何平均数(Han et al., 2005)。由于N、P含量数据不符合正态分布, 生活型间的N、P含量比较时采用Kruskal-Wallis (K-W)检验, 并用Nemenyi进行两两比较(李单凤等, 2015)。 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014)。同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014)。所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行。
3 结果
3.1 叶片N、P含量特征
所有木本植物叶的N、P含量的几何均值分别为16.57 mg·g-1和1.02 mg·g-1, 其中N的范围为5.06- 43.78 mg·g-1, P的范围为0.18-4.04 mg·g-1 (表1)。落叶木本植物和常绿木本植物的叶N、P含量存在显著差异, 其中落叶木本植物叶N、P含量的几何均值分别较常绿木本植物叶高15.19%和21.93%。从变异系数来看, 所有木本和各生活型叶的P含量变异大于N, 落叶木本植物叶N含量的变异略高于常绿木本植物, 而常绿木本植物叶P含量的变异大于落叶木本植物(表1)。 Table 1 表1 表1中国南方灌丛优势木本植物叶的氮磷含量 Table 1Leaf N and P concentration of dominant woody plants in shrublands across southern China
生活型 Life-form
N含量 N content (mg·g-1)
P含量 P content (mg·g-1)
n
GM
Range
CV
n
GM
Range
CV
常绿木本植物 Evergreen woody plants
91
15.19a
7.68-33.00
0.31
91
0.89a
0.37-3.81
0.52
落叶木本植物 Deciduous woody plants
102
17.91b
5.06-43.78
0.35
102
1.14b
0.18-4.04
0.47
所有木本植物 All woody plants
193
16.57
5.06-43.78
0.35
193
1.02
0.18-4.04
0.50
GM, n, Range and CV were geometric mean values, statistical samples, variation range and coefficient of variation, respectively. “a” and “b” indicated significant difference (p < 0.05) between evergreen and deciduous woody plants.GM、n、Range和CV分别为几何平均数、样本数、范围和变异系数; a和b表示常绿和落叶木本器官元素含量差异显著(p < 0.05)。 新窗口打开
3.2 叶片N、P随环境梯度的变化
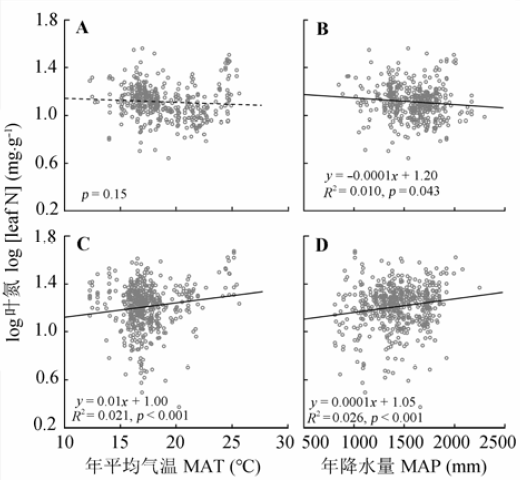
常绿和落叶木本植物叶N、P含量对气候因子的响应方式如下: 随着MAT和MAP的升高, 常绿木本叶片N、P含量均有下降趋势, 落叶木本叶片N、P含量呈增加趋势(图1, 图2); 然而, 常绿和落叶木本植物叶P含量随MAP的增加均呈递减趋势(图2D)。同时, 气候对落叶木本植物叶N含量的影响(图1C; 图1D)大于常绿木本植物(图1A; 图1B); 而对常绿木本植物叶片P含量的影响(图2A; 图2B)高于落叶木本植物(图2C; 图2D)。 显示原图|下载原图ZIP|生成PPT 图1年平均气温和年降水量对常绿(A、B)和落叶木本(C、D)植物叶氮含量的影响。 -->Fig. 1Effects of mean annual temperature (MAT) and mean annual precipitation (MAP) on leaf N concentration of evergreen (A, B) and deciduous woody plants (C, D). -->
显示原图|下载原图ZIP|生成PPT 图2年平均气温和年降水量对常绿(A、B)和落叶木本(C、D)植物叶磷含量的影响。 -->Fig. 2Effects of mean annual temperature (MAT) and mean annual precipitation (MAP) on leaf P concentrations of evergreen (A, B) and deciduous woody plants (C, D). -->
土壤养分对常绿和落叶木本植物叶N、P含量的影响趋于一致: 随土壤N含量的增加, 两者N含量没有显著变化趋势(图3A, 3C, p > 0.05); 随着土壤P含量的增加, 两类植物叶的P含量均显著增加, 且影响力相当(图3B, r2 = 0.176, 斜率0.26; 图3D, r2 = 0.170, 斜率0.29)。 显示原图|下载原图ZIP|生成PPT 图3土壤氮、磷含量对常绿(A、B)和落叶木本(C、D)植物叶氮、磷含量的影响。 -->Fig. 3Effects of soil N, P concentration on leaf N, P concentration of evergreen (A, B) and deciduous woody plants (C, D). -->
3.3 生活型、气候和土壤养分对木本植物叶N、P的影响
GLM回归结果(表2)显示, 对于叶N含量, 只有生活型和土壤P含量在最优模型中, 生活型可解释7.6%的N变异, 土壤P含量解释了2.3%的变异; 对于叶P含量, 在最优模型中包括了生活型、MAP、土壤N含量和土壤P含量, 分别解释了14.4%、0.8%、0.9%和16.4%的P含量变异。 Table 2 表2 表2木本植物叶氮磷含量的广义回归结果 Table 2Summary of general linear models for leaf N and P content of woody plants
因子 Factor
叶片氮含量 Leaf N content
叶片磷含量 Leaf P content
DF
MS
F
SS%
DF
MS
F
SS%
生活型 Life form
1
2.08
78.51
7.6
1
5.79
199.75
14.4
年平均气温 MAT (℃)
1
0.07
2.68
1
0.00
0.05
年降水量 MAP (mm)
1
0.09
3.24
1
0.31
10.52
0.8
土壤氮含量 Soil N content (mg·g-1)
1
0.03
1.22
1
0.37
12.91
0.9
土壤磷含量 Soil P content (mg·g-1)
1
0.63
23.58
2.3
1
6.57
226.60
16.4
DF, MS, F and SS% represent freedom degree, mean sum of squares, F value and interpretation quantity of each factor. F values in bold indicate p < 0.05. MAT, mean annual temperature; MAP, mean annual precipitation.DF、MS、F和SS%分别表示自由度、平均残差平方和、F值和各因子的解释量; 加粗的F值表示p < 0.05。 新窗口打开
AertsR, Chapin FSIII (2000). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns .Advance of Ecology Research, 30, 1-67. [本文引用: 1]
[2]
ÅgrenGI (2008). Stoichiometry and nutrition of plant growth in natural communities .Annual Review of Ecology, Evolution, and Systematics, 39, 153-170. [本文引用: 2]
[3]
ÅgrenGI, WeihM (2012). Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype .New Phytologist, 194, 944-952. [本文引用: 1]
[4]
ChenY, HanW, TangL, TangZ, FangJ (2013). Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form .Ecography, 36, 178-184. [本文引用: 5]
[5]
CornelissenJHC, CornwellWE, AustinA (2014). The tree of life in ecosystems: Evolution of plant effects on carbon and nutrient cycling .Journal of Ecology, 102, 269-274. [本文引用: 2]
[6]
Dı?azS, CabidoM (2001). Vive la différence: Plant functional diversity matters to ecosystem processes .Trends in Ecology & Evolution, 16, 646-655. [本文引用: 1]
[7]
DingF, LianPY, ZengDH (2011). Characteristics of plant leaf nitrogen and phosphorus stoichiometry in relation to soil nitrogen and phosphorus concentrations in Songnen Plain meadow .Chinese Journal of Ecology, 30, 77-81. (in Chinese with English abstract)[丁凡, 廉培勇, 曾德慧 (2011). 松嫩平原草甸三种植物叶N、P化学计量特征及其与土壤N、P浓度的关系 . 生态学杂志, 30, 77-81.] [本文引用: 2]
[8]
EdwardsEJ, ChateletDS, SackL, DonoghueMJ, CornwellW (2014). Leaf life span and the leaf economic spectrum in the context of whole plant architecture .Journal of Ecology, 102, 328-336. [本文引用: 1]
[9]
ElserJJ, BrackenME, ClelandEE, GrunerDS, HarpoleWS, HillebrandH, NgaiJT, SeabloomEW, ShurinJB, SmithJE (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems .Ecology Letters, 10, 1135-1142. [本文引用: 1]
[10]
GanL, ChenFS, HuXF, TianQX, GeG, ZhanSX (2008). Leaf N and P concentrations and their stoichiometric ratios of different functional groups of plants in Nanchang City . Chinese Journal of Ecology, 27, 344-348. (in Chinese with English abstract)[甘露, 陈伏生, 胡小飞, 田秋香, 葛刚, 詹书侠 (2008). 南昌市不同植物类群叶片氮磷浓度及其化学计量比 . 生态学杂志, 27, 344-348.] [本文引用: 1]
[11]
GüsewellS (2004). N:P ratios in terrestrial plants: Variation and functional significance .New Phytologist, 164, 243-266. [本文引用: 1]
[12]
HanW, FangJ, GuoD, ZhangY (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China .New Phytologist, 168, 377-385. [本文引用: 3]
[13]
HanWX, WuY, TangLY, ChenYH, LiLP, HeJS, FangJY (2009). Leaf carbon, nitrogen and phosphorus stoichio- metry across plant species in Beijing and its periphery .Acta Scientiarum Naturalium Universitatis Pekinensis, 45, 855-860. (in Chinese with English abstract)[韩文轩, 吴漪, 汤璐瑛, 陈雅涵, 李利平, 贺金生, 方精云 (2009). 北京及周边地区植物叶的碳氮磷元素计量特征 . 北京大学学报(自然科学版), 45, 855-860.] [本文引用: 3]
[14]
HedinLO (2004). Global organization of terrestrial plant-nutrient interactions .Proceedings of the National Academy of Sciences of the United States of America, 101, 10849-10850. [本文引用: 1]
[15]
HeJS, FangJ, WangZ, GuoD, FlynnDF, GengZ (2006). Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China .Oecologia, 149, 115-122. [本文引用: 2]
[16]
HeJS, WangL, FlynnDF, WangX, MaW, FangJ (2008). Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes .Oecologia, 155, 301-310. [本文引用: 4]
[17]
HongJ, WangX, WuJ (2014). Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the northern Tibetan Plateau .PLOS ONE, 9, e109052. doi: 10.1371/journal.pone.0109052. [本文引用: 3]
[18]
HuHF, WangZH, LiuGH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Ecology (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 1]
[19]
KerkhoffAJ, EnquistBJ, ElserJJ, FaganWF (2005). Plant allometry, stoichiometry and the temperature-dependence of primary productivity .Global Ecology and Biogeo- graphy, 14, 585-598. [本文引用: 1]
[20]
KoerselmanW, MeulemanAFM (1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation .Journal of Applied Ecology, 33, 1441-1450. [本文引用: 1]
[21]
LiDF, YuSL, WangGX, FangWW (2015). Environmental heterogeneity and mechanism of stoichiometry properties of vegetative organs in dominant shrub communities across the Loess Plateau .Chinese Journal of Plant Ecology, 39, 453-465. (in Chinese with English abstract)[李单凤, 于顺利, 王国勋, 方伟伟 (2015). 黄土高原优势灌丛营养器官化学计量特征的环境分异和机制 . 植物生态学报, 39, 453-465.] [本文引用: 3]
[22]
LinZB, YanPY, YangZJ, WanXH, ChenGS (2011). Leaf nitrogen and phosphorus stoichiometry across 101 woody species in Wanmulin, Fujian Province .Journal of Subtropical Resources & Environment, 6, 32-38. (in Chinese with English abstract)[林志斌, 严平勇, 杨智杰, 万晓华, 陈光水 (2011). 福建万木林101种常见木本植物叶N, P化学计量学特征 . 亚热带资源与环境学报, 6, 32-38.] [本文引用: 1]
[23]
LiuC, WangY, WangN, WangGX (2012). Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial eco-systems: A review .Chinese Journal of Plant Ecology, 36, 1205-1216. (in Chinese with English abstract)[刘超, 王洋, 王楠, 王根轩 (2012). 陆地生态系统植被氮磷化学计量研究进展 . 植物生态学报, 36, 1205-1216.] [本文引用: 1]
[24]
McGroddyME, DaufresneT, HedinLO (2004). Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios .Ecology, 85, 2390-2401. [本文引用: 2]
[25]
NottinghamAT, TurnerBL, StottAW, TannerEVJ (2015). Nitrogen and phosphorus constrain labile and stable carbon turnover in lowland tropical forest soils .Soil Biology & Biochemistry, 80, 26-33. [本文引用: 1]
[26]
PanF, ZhangW, LiuS, LiD, WangK (2015). Leaf N:P stoichiometry across plant functional groups in the karst region of southwestern China .Trees, 29, 883-892. [本文引用: 3]
[27]
PetterG, WagnerK, WanekW, DelgadoEJS, ZotzG, CabralJS, KreftH (2016). Functional leaf traits of vascular epiphytes: Vertical trends within the forest, intra-and interspecific trait variability, and taxonomic signals .Functional Ecology, 30, 188-198. [本文引用: 1]
[28]
PiaoSL, FangJY, CiaisP, PeylinP, HuangY, SitchS, WangT (2009). The carbon balance of terrestrial ecosystems in China .Nature, 458, 1009-1013. [本文引用: 1]
[29]
ReichPB, OleksynJ (2004). Global patterns of plant leaf N and P in relation to temperature and latitude .Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006. [本文引用: 8]
[30]
SardansJ, PeñuelasJ (2013). Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood .Global Ecology and Biogeography, 22, 494-507. [本文引用: 2]
[31]
ReichPB (2005). Global biogeography of plant chemistry: Filling in the blanks .New Phytologist, 168, 263-266. [本文引用: 1]
[32]
RenSJ, YuGR, TaoB, WangSQ (2007). Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC .Environmental Science, 28, 2665-2673. (in Chinese with English abstract)[任书杰, 于贵瑞, 陶波, 王绍强 (2007). 中国东部南北样带654种植物叶氮和磷的化学计量学特征研究 . 环境科学, 28, 2665-2673.] [本文引用: 3]
[33]
SternerRW, ElserJJ (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton. [本文引用: 2]
[34]
The Editorial Committee of Vegetation Map of China, Chinese Academy ofSciences (2007). Vegetation Map of the People’s Republic of China 1:1000000. Geological Publishing House, Beijing. (in Chinese)[中国科学院中国植被图编辑委员会 (2007). 中华人民共和国植被图1:1000000. 地质出版社, 北京.] [本文引用: 3]
[35]
WangJY, WangSQ, LiRL, YanJH, ShaLQ, HanSJ (2011). C:N:P stoichiometric characteristics of four forest types’ dominant tree species in China . Chinese Journal of Plant Ecology, 35, 587-595. (in Chinese with English abstract)[王晶苑, 王绍强, 李纫兰, 闫俊华, 沙丽清, 韩士杰 (2011). 中国四种森林类型主要优势植物的C:N:P化学计量学特征 . 植物生态学报, 35, 587-595.] [本文引用: 1]
[36]
WangT, YangYH, MaWH (2008). Storage, patterns and environmental controls of soil phosphorus in China .Acta Scientiarum Naturalium Universitatis Pekinensis, 44, 549-556. (in Chinese with English abstract)[汪涛, 杨元合, 马文红 (2008). 中国土壤磷库的大小、分布及其影响因素 . 北京大学学报(自然科学版), 44, 549-556.] [本文引用: 1]
[37]
WuTG, YuMK, WangGG, DongY, ChengXR (2012). Leaf nitrogen and phosphorus stoichiometry across forty-two woody species in southeast China .Biochemical Systematics & Ecology, 44, 255-263. [本文引用: 1]
[38]
XieZQ, TangZY (2015). Technical specification of field investigation and laboratory analysis for carbon sequestration in shrubland ecosystems . In: Compilation Group of Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems ed. Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems. Science Press, Beijing. 145-191. (in Chinese)[谢宗强, 唐志尧 (2015). 灌丛生态系统固碳研究的野外调查与室内分析技术规范 . 见: 生态系统固碳观测与调查技术规范编写组编 (2015). 生态系统固碳观测与调查技术规范. 科学出版社, 北京. 145-191.] [本文引用: 2]
[39]
YangX, TangZ, JiC, LiuH, MaW, MohhamotA, ShiZ, SunW, WangT, WangX, WuX, YuS, YueM, ZhengC (2014). Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across northern China .Scientific Reports, 4, 5448. [本文引用: 4]
[40]
ZengDH, ChenGS (2005). Ecological stoichiometry: A science to explore the complexity of living systems .Acta Phytoecologica Sinica, 29, 1007-1019. (in Chinese with English abstract)[曾德慧, 陈广生 (2005). 生态化学计量学——复杂生命系统奥秘的探索 . 植物生态学报, 29, 1007-1019.] [本文引用: 1]
[41]
ZhangLX (2003). Plant N:P Stoichiometry: The Differentiation Rules and Preliminary Validation in Field Experiments of High Plants of China . PhD dissertation, Institute of Botany, Chinese Academy of Science, Beijing. (in Chinese with English abstract)[张丽霞 (2003). 植物N:P计量化学: 中国高等植物的分异规律与野外实验初步验证 . 博士学位论文, 中国科学院植物研究所, 北京.] [本文引用: 1]
[42]
ZhaoN, HeN, WangQ, ZhangX, WangR, XuZ, YuG (2014). The altitudinal patterns of leaf C:N:P stoichi- ometry are regulated by plant growth form, climate and soil on Changbai Mountain, China .PLOS ONE, 9: e95196. doi: 10.1371/journal.pone.0095196. [本文引用: 1]
[43]
ZhengDN, WangXS, XieSD, DuanL, ChenDS (2014). Simulation of atmospheric nitrogen deposition in China in 2010 .China Environmental Science, 34, 1089-1097. (in Chinese with English abstract)[郑丹楠, 王雪松, 谢绍东, 段雷, 陈东升 (2014). 2010年中国大气氮沉降特征分析 . 中国环境科学, 34, 1089-1097.] [本文引用: 1]
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns 1 2000
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
Stoichiometry and nutrition of plant growth in natural communities 2 2008
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
... ), 支持生长速率假说(Ågren, 2008). ...
Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype 1 2012
... 植物的营养元素含量是植物在一定生境下吸收营养元素的能力, 也是植物与环境之间相互作用的结果.在全球或国家尺度上, 已有研究表明植物叶N、P含量随MAT增高和MAP增加逐渐减少(Güsewell, 2004; Hedin, 2004; McGroddy et al., 2004; He et al., 2006; Ågren & Weih, 2012; Chen et al., 2013).然而, 中国南方灌丛中常绿和落叶木本植物叶N、P含量与气候因子的关系不同(图1, 图2), 其中常绿木本植物叶N、P含量与Han等(2005)与Reich和Oleksyn (2004)的研究结果存在一样的温度趋势, 结果支持温度-植物生理假说: 叶片N、P含量的增加可补偿低温下代谢速率的下降, 因此随温度的增加, 叶片N、P含量呈降低趋势(Reich & Oleksyn, 2004).同时, 较高的叶片N、P含量也是植物适应干旱条件的反映(Yang et al., 2014), 也可能是较多的降水可增强土壤N、P的淋溶作用, 土壤养分的可获得性降低, 从而导致常绿木本植物叶N、P含量呈降低的趋势.然而, 落叶木本植物叶N、P含量随MAT和MAP的升高而呈增加趋势, 可能与较高的温度和较多的降水会加快养分周转, 增加土壤养分的可获得性有关; 再者, 在温暖湿润的环境中, 落叶木本植物生长较快, 较高的叶N、P含量有助于满足植物快速生长的养分需求, 因此落叶木本植物叶N、P含量随年平均气温和年降水量的增加呈增加趋势. ...
Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form 5 2013
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the northern Tibetan Plateau 3 2014
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...
中国主要灌丛植被碳储量 1 2006
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
Plant allometry, stoichiometry and the temperature-dependence of primary productivity 1 2005
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation 1 1996
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
黄土高原优势灌丛营养器官化学计量特征的环境分异和机制 3 2015
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
Functional leaf traits of vascular epiphytes: Vertical trends within the forest, intra-and interspecific trait variability, and taxonomic signals 1 2016
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
The carbon balance of terrestrial ecosystems in China 1 2009
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
Global patterns of plant leaf N and P in relation to temperature and latitude 8 2004
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
... ), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood 2 2013
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
Global biogeography of plant chemistry: Filling in the blanks 1 2005
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
中国东部南北样带654种植物叶氮和磷的化学计量学特征研究 3 2007
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...
... 植物的营养元素含量是植物在一定生境下吸收营养元素的能力, 也是植物与环境之间相互作用的结果.在全球或国家尺度上, 已有研究表明植物叶N、P含量随MAT增高和MAP增加逐渐减少(Güsewell, 2004; Hedin, 2004; McGroddy et al., 2004; He et al., 2006; Ågren & Weih, 2012; Chen et al., 2013).然而, 中国南方灌丛中常绿和落叶木本植物叶N、P含量与气候因子的关系不同(图1, 图2), 其中常绿木本植物叶N、P含量与Han等(2005)与Reich和Oleksyn (2004)的研究结果存在一样的温度趋势, 结果支持温度-植物生理假说: 叶片N、P含量的增加可补偿低温下代谢速率的下降, 因此随温度的增加, 叶片N、P含量呈降低趋势(Reich & Oleksyn, 2004).同时, 较高的叶片N、P含量也是植物适应干旱条件的反映(Yang et al., 2014), 也可能是较多的降水可增强土壤N、P的淋溶作用, 土壤养分的可获得性降低, 从而导致常绿木本植物叶N、P含量呈降低的趋势.然而, 落叶木本植物叶N、P含量随MAT和MAP的升高而呈增加趋势, 可能与较高的温度和较多的降水会加快养分周转, 增加土壤养分的可获得性有关; 再者, 在温暖湿润的环境中, 落叶木本植物生长较快, 较高的叶N、P含量有助于满足植物快速生长的养分需求, 因此落叶木本植物叶N、P含量随年平均气温和年降水量的增加呈增加趋势. ...
生态化学计量学——复杂生命系统奥秘的探索 1 2005
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
植物N:P计量化学: 中国高等植物的分异规律与野外实验初步验证 1 2003
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
The altitudinal patterns of leaf C:N:P stoichi- ometry are regulated by plant growth form, climate and soil on Changbai Mountain, China 1 2014
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT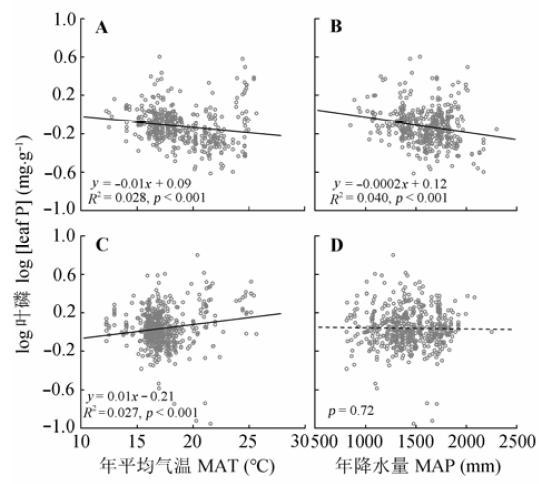 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT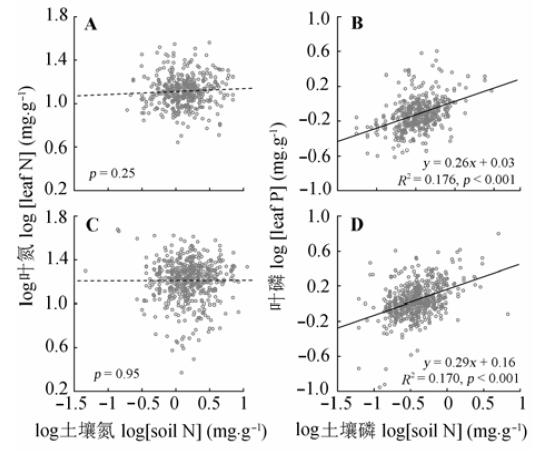 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}