Effects of environmental factors on gross caloric values of three life-forms aquatic plants on the Qinghai-Xizang Plateau, China
WANGYang, MIAOLin-Lin, YUDan, LIUChun-Hua*,, WANGZhong*, College of Life Sciences, Wuhan University, Wuhan 430072, China 通讯作者:* 通信作者Author for correspondence (E-mail:liuchh@163.com, wangzhong@whu.edu.cn)* 通信作者Author for correspondence (E-mail:liuchh@163.com, wangzhong@whu.edu.cn) 收稿日期:2016-01-22 接受日期:2016-07-19 网络出版日期:2017-02-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
关键词:热值;水生植物;青藏高原;环境影响 Abstract Aims Gross caloric value (GCV) reflected plants’ capability of converting solar energy. It provided a reliable indicator of plants’ adaptations to environments in perspective of energy conversion and fixation. The aims of this study were (1) to illustrate the characteristics of GCV of aquatic plants on the Qinghai-Xizang Plateau, (2) to explore the geographical and environmental patterns and (3) to discuss the underlying mechanisms in forming the patterns. Methods In July and August 2015, we collected 533 samples of aquatic plants’ leaves in 143 field sites on the Qinghai-Xizang Plateau, and measured their GCV using SDACM-4000 oxygen bomb calorimeter. Together with mean annual temperature (MAT) of climatic factor and properties of water body, this study compared the differences of GCV among submerged, emergent and floating-leaved plants by analysis of variance (ANOVA) and Tukey’s HSD. We further regressed GCV of submerged and emergent plants against geographical and climatic factors and properties of water body by simple regression to explore the relative effects of environmental factors on GCV. Important findings On the Qinghai-Xizang Plateau, the mean GCV was (15.95 ± 3.90) kJ·g-1. Among the three life forms, the rank of GCV was the emergent plants (18.10 kJ·g-1) > the floating-leaved plants (16.77 kJ·g-1) > the submerged plants (14.31 kJ·g-1). With an increasing latitude, the GCV of emergent and submerged plants increased. Only GCV of emergent plants decreased with increasing altitude and temperature. The GCV of emergent and submerged plants increased with increased water salinity. Dissolved oxygen had significant negative effects on emergent plants, while pH value had no significant effects.
青藏高原水生植物热值为(15.95 ± 3.90) kJ·g-1, 总体数据的CV为0.24。从不同生活型来看, 挺水型植物的热值最高, 为(18.10 ± 4.03) kJ·g-1, 浮叶型植物次之, 为(16.77 ± 1.52) kJ·g-1, 二者无明显差异, 但都显著高于沉水型植物热值((14.31 ± 3.03) kJ·g-1)(表1)。 Table 1 表1 表1青藏高原3种不同生活型水生植物热值特征 Table 1Characteristics of gross caloric values of three life-forms of aquatic plants on the Qinghai-Xizang Plateau
生活型 Life-form
样品量 No. of samples
热值 Gross caloric values (kJ·g-1)
平均值 Mean
标准偏差 Standard deviation
变异系数 Coefficient of variation
沉水型 Submerged
295
14.31a
3.03
0.21
浮叶型 Floating-leaved
20
16.77b
1.52
0.09
挺水型 Emergent
218
18.10b
4.03
0.22
总计 Total
533
15.95
3.90
0.24
Different small letters indicate significant differences between different life forms of aquatic plants (p < 0.05).不同小写字母表示不同生活型之间热值差异显著(p < 0.05)。 新窗口打开
2.2 水生植物热值的地理格局
挺水型和沉水型两种生活型水生植物热值表现出了一致的地理格局(图2)。随海拔升高, 挺水型植物热值显著降低(p < 0.001, 图2D), 沉水型植物也有降低的趋势, 但未达到显著水平(p = 0.088, 图2A)。随经度和纬度升高, 两种生活型水生植物热值则均表现出显著增加的趋势(p < 0.001, 图2B、2C、2E、2F)。 显示原图|下载原图ZIP|生成PPT 图2两种生活型水生植物的热值随海拔、经度和纬度的变化格局。A-C, 沉水型植物, n = 295。D-F, 挺水型植物, n = 218。 -->Fig. 2Geographic patterns of gross caloric values of submerged plants and emergent plants along altitudinal, longitudinal and latitudinal gradients. A-C, Submerged plants, n = 295. D-F, Emergent plants, n = 218. -->
2.3 环境因子对水生植物热值的影响
水体盐度对挺水型和沉水型两种生活型植物热值有显著影响(p < 0.05), 热值随水体盐度的升高而升高; 溶氧量只与挺水型植物的热值呈负相关关系(p = 0.010), 对沉水型植物影响不显著(p = 0.075); 而pH值则对两种生活型植物的热值都没有表现出显著的影响(p > 0.1)(图3)。 显示原图|下载原图ZIP|生成PPT 图3水体盐度、溶氧量和pH值对两种生活型植物热值的影响。A-C, 沉水型植物, n = 295。D-F, 挺水型植物, n = 218。 -->Fig. 3Effects of waterbody properties on gross caloric value of submerged plants and emergent plants. A-C, Submerged plants, n = 295. D-F, Emergent plants, n = 218. -->
气温对沉水型植物的热值影响不显著(R2 = 0.011, p = 0.070), 但与挺水型植物热值呈明显的负相关关系(R2 = 0.018, p = 0.045)(图4)。 显示原图|下载原图ZIP|生成PPT 图4年平均气温(MAT)对两种生活型水生植物热值的影响。A, 沉水型植物, n = 295。B, 挺水型植物, n = 218。 -->Fig. 4Effects of annual mean air temperature (MAT) on gross caloric values of submerged and emergent plants. A, Submerged plants, n = 295. B, Emergent plants, n = 218. -->
BaoYJ, LiZH (2003). Dynamic of caloric values of major species in Leymus chinensis steppe of Inner Mongolia. Acta Ecologica Sinica, 23, 606-613. (in Chinese with English abstract)[鲍雅静, 李政海 (2003). 内蒙古羊草草原群落主要植物的热值动态 . 生态学报, 23, 606-613.] [本文引用: 2]
[2]
BaoYJ, LiZH, HanXG, SongGB, YangXH, LüHY (2006). Plant caloric value and its bio-ecological attributes. Chinese Journal of Ecology, 25, 1095-1103. (in Chinese with English abstract)[鲍雅静, 李政海, 韩兴国, 宋国宝, 杨晓慧, 吕海燕 (2006). 植物热值及其生物生态学属性 . 生态学杂志, 25, 1095-1103] [本文引用: 1]
[3]
BiYF, CheWG (2004). Studies on the calorific value of Medicago population. Acta Agrestia Sinica, 10, 265-269. (in Chinese with English abstract)[毕玉芬, 车伟光 (2004). 几种苜蓿属植物植株热值研究 . 草地学报, 10, 265-269.] [本文引用: 2]
[4]
ChenML, ShangguanZP (2009). Caloric value and nutrient characteristics of dominant plant species of six typical vegetation communities in Ziwuling forest area of the Loess Plateau. Scientia Silvae Sinicae, 45(3), 140-144. (in Chinese with English abstract)[陈美玲, 上官周平 (2009). 黄土高原子午岭林区6个典型群落优势种的热值和养分特征 . 林业科学, 45(3), 140-144.] [本文引用: 1]
[5]
FangY, XuSM, SunG (2004). Primary production of the aquatic ecosystem of the Nanhu lake in Changchun II—Periphyton and macrophyte. Journal of Jilin Agricultural University, 26(1), 46-49. (in Chinese with English abstract)[房岩, 徐淑敏, 孙刚 (2004). 长春南湖水生生态系统的初级生产II——附生藻类与大型水生植物 . 吉林农业大学学报, 26(1), 46-49.] [本文引用: 2]
[6]
GaoK, HanGD, YuYQ, ZhangLJ (2015). Characterization of caloric values in 14 plant species in Hunshandake Sandy Land. Chinese Journal of Ecology, 34, 341-346. (in Chinese with English abstract)[高凯, 韩国栋, 于永奇, 张丽娟 (2015). 浑善达克沙地14种植物热值特征 . 生态学杂志, 34, 341-346.] [本文引用: 2]
GolleyFB (1969). Caloric value of wet tropical forest vegetation. Ecology, 50, 517-519. [本文引用: 2]
[9]
GuoJX, WangRD, BaoGZ (2001). Caloric value of northeast Aneurolepidium chinense grassland species. Acta Phytoecologica Sinica, 25, 746-750. (in Chinese with English abstract)[郭继勋, 王若丹, 包国章 (2001). 东北羊草草原主要植物热值 . 植物生态学报, 25, 746-750.] [本文引用: 1]
[10]
HaoCY, LiuP (2006). The caloric value of the dominant plant species of a Heptacodium miconioides forest at Bei Mountain, Zhejiang Province, China. Acta Ecologica Sinica, 26, 1709-1717. (in Chinese with English abstract)[郝朝运, 刘鹏 (2006). 浙江北山七子花群落主要植物叶热值 . 生态学报, 26, 1709-1717.] [本文引用: 1]
[11]
HeJN, KangWX, WangD (2015). The plant calorific value in the Chinese fir (Cunninghamia lanceolata) plantations at different ages. Acta Ecologica Sinica, 35, 449-459. (in Chinese with English abstract)[何介南, 康文星, 王东 (2015). 不同年龄阶段杉木人工林植物热值分析 . 生态学报, 35, 449-459.] [本文引用: 1]
[12]
HeX, BaoWK, GuB, ZhengWJ, LengL (2007). The characteristic of gross caloric value of higher plants in China. Ecology and Environment, 16, 973-981. (in Chinese with English abstract)[何晓, 包维楷, 辜彬, 郑帷婕, 冷俐 (2007). 中国高等植物干质量热值特点 . 生态环境, 16, 973-981.] [本文引用: 2]
[13]
Howard-WilliamsC (1974). Nutritional quality and calorific value of Amazonian forest litter. Amazoniana, 5(1), 67-75. [本文引用: 1]
[14]
Institute ofBotany, Chinese Academy ofSciences (1988). Xizang Vegetation. Science Press, Beijing. (in Chinese)[中国科学院植物研究所 (1988). 西藏植被. 科学出版社, 北京.] [本文引用: 1]
LiYD, WuZM, ZengQB, ZhouGY, ChenBF (1996). Caloric values of main species in a tropical mountain rain forest at Jianfengling, Hainan Island. Acta Phytoecologica Sinica, 20, 1-10 (in Chinese with English abstract)[李意德,吴仲民, 曾庆波, 周光益, 陈步峰 (1996). 尖峰岭热带山地雨林主要种类能量背景值测定分析 . 植物生态学报, 20, 1-10.] [本文引用: 1]
[17]
LinGH, LinP (1988). Studies on energy in two mangrove communities, Bruguiera sexangula and Kandelia candel, in China. Acta Phytocological and Geobotanica Sinica, 12, 31-39. (in Chinese with English abstract)[林光辉, 林鹏 (1988). 海莲、秋茄两种红树群落能量的研究 . 植物生态学与地植物学学报, 12, 31-39.] [本文引用: 1]
[18]
LinH, CaoM (2008). Plant energy storage strategy and caloric value. Ecological Modelling, 217, 132-138. [本文引用: 1]
[19]
LinP, LinGH (1991). Study on the caloric value and ash content of some mangrove species in China. Acta Phytocologica and Geobotanica Sinica, 15, 114-120. (in Chinese with English abstract)[林鹏, 林光辉 (1991). 几种红树植物的热值和灰分含量研究 . 植物生态学与地植物学学报, 15, 114-120.] [本文引用: 4]
[20]
LinP, WangWQ (1999). Changes in the caloric values of Kandelia candel seedlings under salt stress. Acta Phytoecologica Sinica, 23, 466-470. (in Chinese with English abstract)[林鹏, 王文卿 (1999). 盐胁迫下红树植物秋茄(Kandelia candel)热值变化的研究 . 植物生态学报, 23, 466-470.] [本文引用: 1]
[21]
LinYM, GuoQR, YeGF, LinL, LinP (2004a). Characteristics of matter and energy of some Casuarinaceae species in Dongshan County, Fujian Province. Acta Ecologica Sinica, 24, 2217-2224. (in Chinese with English abstract)[林益明, 郭启荣, 叶功富, 林玲, 林鹏 (2004). 福建东山几种木麻黄的物质与能量特征 . 生态学报, 24, 2217-2224.] [本文引用: 2]
[22]
LinYM, LiZB, ChenYY, LinP (2001). Caloric value in leaves of some bamboo species in the bamboo garden of Hua’an County, Fujian. Chinese Bulletin of Botany, 18, 356-362. (in Chinese with English abstract)[林益明, 黎中宝, 陈奕源, 林鹏 (2001). 福建华安竹园一些竹类植物叶的热值研究 . 植物学报, 18, 356-362.] [本文引用: 1]
[23]
LinYM, LinP, WangT (2000). Caloric values and ash contents of some mangrove woods. Chinese Journal of Applied Ecology, 11, 181-184. (in Chinese with English abstract)[林益明, 林鹏, 王通 (2000). 几种红树植物木材热值和灰分含量的研究 . 应用生态学报, 11, 181-184.] [本文引用: 1]
[24]
LinYM, WangZC, KeLN, LinP, TanZQ, DingYL (2003). Monthly changes in the caloric values of the leaves of four shrubby and four tree-dwelling Palmae species. Acta Ecologica Sinica, 23, 1117-1124. (in Chinese with English abstract)[林益明, 王湛昌, 柯莉娜, 林鹏, 谭忠奇, 丁印龙 (2003). 四种灌木状与四种乔木状棕榈植物热值的月变化 . 生态学报, 23, 1117-1124.] [本文引用: 1]
[25]
LinYM, XiangP, LinP (2004b). Caloric values of propagules and leaves at the different development stages of mangrove species at Futian, Shenzhen. Marine Science, 28(2), 43-48. (in Chinese with English abstract)[林益明, 向平, 林鹏 (2004b). 深圳福田几种红树植物繁殖体与不同发育阶段叶片热值研究 . 海洋科学, 28(2), 43-48.] [本文引用: 3]
[26]
LongRJ, XuCL, HuZZ, DingWG, ZhangYS, KangTF (1993). Calorific value and its seasonal dynamics of fodder shrub species on Tianzhu alpine grasslands. Chinese Journal of Ecology, 12(5), 13-16. (in Chinese with English abstract)[龙瑞军, 徐长林, 胡自治, 丁文广, 张映生, 康天福 (1993). 天祝高山草原15种饲用灌木的热值及季节动态 . 生态学杂志, 12(5), 13-16.] [本文引用: 1]
[27]
QiaoXJ, CaoM, LinH (2007). Caloric value allocation of dominant species in four secondary forests at different ages in Xishuangbanna, Southwest China . Journal of Plant Ecology (Chinese Version), 31, 326-332. (in Chinese with English abstract)[乔秀娟, 曹敏, 林华 (2007). 西双版纳不同林龄次生植物群落优势树种的热值 . 植物生态学报, 31, 326-332.] [本文引用: 1]
[28]
RenH, PengSL (1999). The characteristics of ecological energetics of the forest ecosystem in the successional process in Dinghushan, Guangdong, China. Acta Ecologica Sinica, 19, 817-822. (in Chinese with English abstract)[任海, 彭少麟 (1999). 鼎湖山森林生态系统演替过程中的能量生态特征 . 生态学报, 19, 817-822.] [本文引用: 2]
[29]
RenH, PengSL, LiuHX, CaoHL, HuangZL (2004). The caloric value of main plant species at Dinghushan, Guangdong, China. Acta Phytoecologica Sinica, 23, 148-154. (in Chinese with English abstract)[任海, 彭少麟, 刘鸿先, 曹洪麟, 黄忠良 (2004). 鼎湖山植物群落及其主要植物的热值研究 . 植物生态学报, 23, 148-154.] [本文引用: 3]
[30]
SinghAK (1980). Energy dynamics in a savanna ecosystem in India. Japanese Journal of Ecology, 30, 295-305. [本文引用: 1]
SuZX, ZhongZC (1994). Studies on the energy of Neosinocalamus affinis clonal population. Acta Ecologica Sinica, 14, 142-148. (in Chinese with English abstract)[苏智先, 钟章成 (1994). 慈竹克隆种群能量动态研究 . 生态学报, 14, 142-148.] [本文引用: 1]
[33]
SunGF, ZhengZM, WangZQ (1993). Dynamics of calorific values of rice. Chinese Journal of Ecology, 2, 218-220. (in Chinese with English abstract)[孙国夫, 郑志明, 王兆骞 (1993). 水稻热值的动态变化研究 . 生态学杂志, 2, 218-220.] [本文引用: 2]
[34]
WangD (2003).The Geography of Aquatic Vascular Plants of Qinghai-Xizang Plateau . PhD dissertation, Wuhan University, Wuhan. (in Chinese with English abstract)[王东 (2003). 青藏高原水生植物地理研究 . 博士学位论文, 武汉大学, 武汉.] [本文引用: 1]
[35]
WangSM, DouHS (1998). Lakes in China. Science Press, Beijing. (in Chinese)[王苏民, 窦鸿身 (1998). 中国湖泊志. 科学出版社, 北京.] [本文引用: 1]
[36]
WangZ, XiaCX, YuD, WuZG (2015). Low-temperature induced leaf elements accumulation in aquatic macrophytes across Tibetan Plateau. Ecological Engineering, 75, 1-8. [本文引用: 2]
[37]
XingW, WuHP, HaoBB, LiuGH (2013). Stoichiometric characteristics and responses of submerged macrophytes to eutrophication in lakes along the middle and lower reaches of the Yangtze River. Ecological Engineering, 54(5), 16-21. [本文引用: 1]
[38]
XuYR, ZhangWJ, FengZW, ZhangJL (2003). Caloric values, elemental contents and correlations between them of some plants on sea beach salinity soil in Tianjin, China. Acta Ecologica Sinica, 23, 450-455. (in Chinese with English abstract)[徐永荣, 张万钧, 冯宗炜, 张金龙 (2003). 天津滨海盐渍土上几种植物的热值和元素含量及其相关性 . 生态学报, 23, 450-455.] [本文引用: 1]
[39]
YangFT, HeHJ (1983). A preliminary study on caloric value of common plants in alpine meadow. Acta Phytoecological and Geobotanica Sinica, 7, 280-288. (in Chinese with English abstract)[杨福囤, 何海菊 (1983). 高寒草甸地区常见植物热值的初步研究 . 植物生态学与地植物学丛刊, 7, 280-288.] [本文引用: 1]
[40]
YangGF, XingK, LiG, LiuHY, PanYZ, ZhangL (2015). Caloric values, element contents (carbon, nitrogen, and phosphorus) of Zostera marina and their correlation. Oceanologia et Limnologia Sinica, 46(1), 212-220. (in Chinese with English abstract)[杨贵福, 邢坤, 李耕, 刘海映, 潘玉州, 张力 (2015). 大叶藻(Zostera marina)的热值和元素含量(碳、氮、磷)及其相关性 . 海洋与湖沼, 46(1), 212-220.] [本文引用: 3]
[41]
YangSC, LiYB, LinP (2003). Change of leaf caloric value from Avicennia marina and Aegiceras corniculatum mangrove plants under cold stress. Journal of Applied Oceanography, 22(1), 46-52. (in Chinese with English abstract)[杨盛昌, 李云波, 林鹏 (2003). 冷胁迫下红树植物白骨壤和桐花树叶片热值的变化 . 应用海洋学报, 22(1), 46-52.] [本文引用: 1]
[42]
YouWH (1995). A study on phytoplankton and its energy production in Dianshan Lake. Transactions of Oceanology and Limnology, 1, 47-53. (in Chinese with English abstract)[由文辉 (1995). 淀山湖的浮游植物及其能量生产 . 海洋湖沼通报, 1, 47-53.] [本文引用: 1]
[43]
YouWH, SongYC (1995). A study on energy in vascular aquatic macrophytes communities in Dianshan Lake. Acta Phytoecologica Sinica, 19, 208-216. (in Chinese with English abstract)[由文辉, 宋永昌 (1995). 淀山湖水生维管束植物群落能量的研究 . 植物生态学报, 19, 208-216.] [本文引用: 6]
[44]
YuYW, HuZZ, ZhangDG, XuCL (2000). Calorific value and its seasonal dynamics of Dasiphora (Potentlia) fruticosa in alpine area of Jinqiang River of Tianzhu County. Pratacultural Science, 17(2), 1-4. (in Chinese with English abstract)[于应文, 胡自治, 张德罡, 徐长林 (2000). 天祝金强河高寒地区金露梅的热值及其季节动态 . 草业科学, 17(2), 1-4.] [本文引用: 1]
[45]
ZhangSB, ZhangJL, SlikJWF, CaoKF (2012). Leaf element concentrations of terrestrial plants across China are influenced by taxonomy and the environment. Global Ecology & Biogeography, 21, 809-818. [本文引用: 2]
[46]
ZuYG (1990). Introduction to Energy Ecology. Jilin Science & Technology Publishing House, Changchun. (in Chinese)[祖元刚 (1990). 能量生态学导论. 吉林科学技术出版社, 长春.] [本文引用: 1]
[47]
ZuYG, ZhangHY (1986). Several technical problems in the measurement of plants’ caloric value. Chinese Journal of Ecology, 12(1), 1-4. (in Chinese)[祖元刚, 张宏一 (1986). 植物热值测定中的若干技术问题 . 生态学杂志, 12(1), 1-4.] [本文引用: 1]
Stoichiometric characteristics and responses of submerged macrophytes to eutrophication in lakes along the middle and lower reaches of the Yangtze River. 1 2013
 , 王忠
, 王忠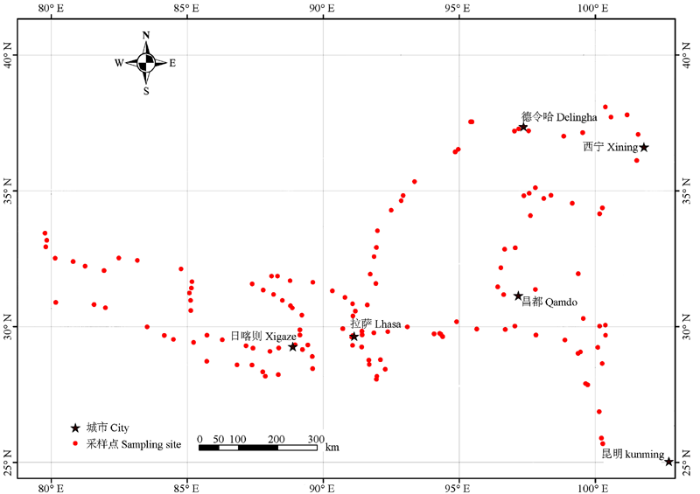 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT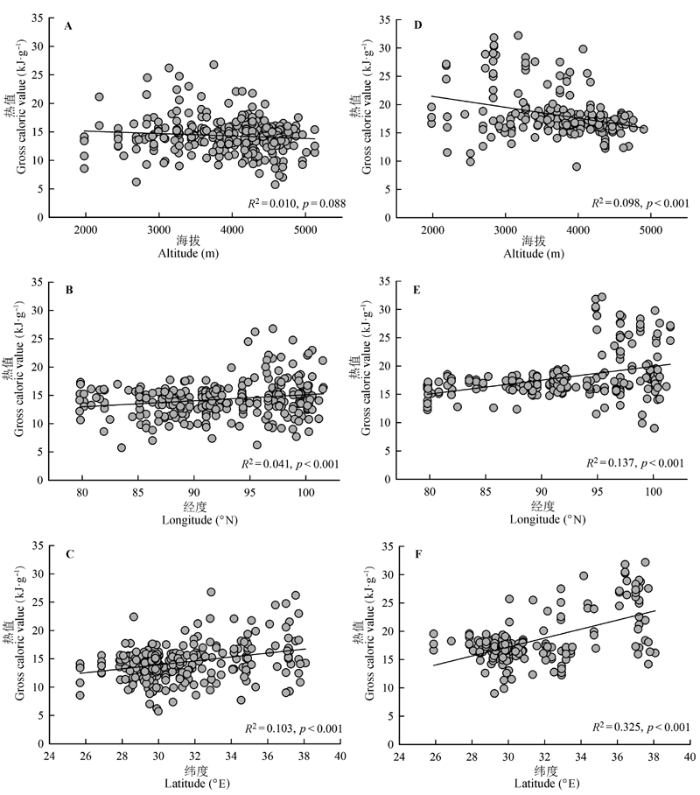 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT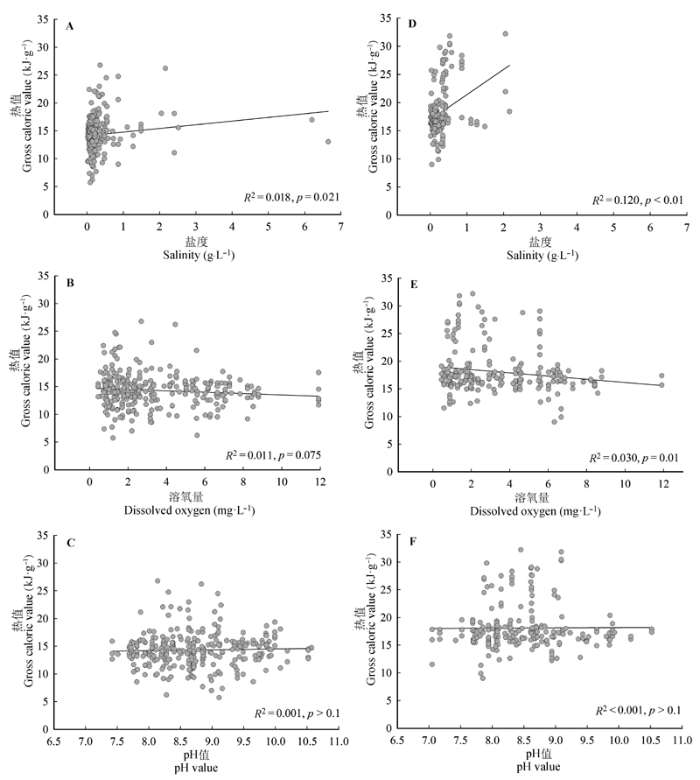 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT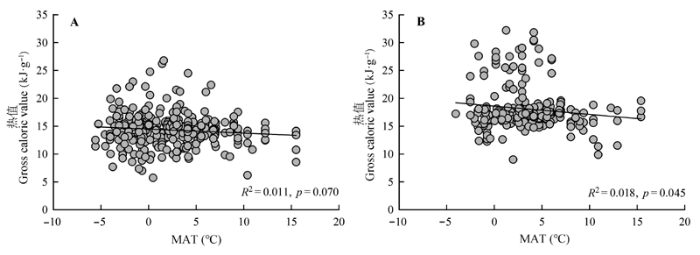 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}