Anatomical characteristics of xylem in tree rings and its relationship with environments
ZHULiang-Jun1, LIZong-Shan2, WANGXiao-Chun1,*, 1Center for Ecological Research, Northeast Forestry University, Harbin 150040, China2State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Science, Chinese Academy of Sciences, Beijing 100085, China 通讯作者:* 通信作者Author for correspondence (E-mail:wangx@nefu.edu.cn) 收稿日期:2016-06-12 接受日期:2017-01-3 网络出版日期:2017-02-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41471168和31370463)、中央高校基本科研业务费专项资金(2572016AA32和2572014DA05)和****和创新团队发展计划(IRT-15R09)项目
关键词:细胞年表;木质部解剖;水力结构;全球变化;塑性响应;树轮 Abstract The anatomical traits of xylem are the characteristics of tree rings at the cellular and subcellular scales, and are often reflection of environmental signals. Studying the relationships between anatomical traits of xylem and environmental change not only provide physiological explanations to the statistics in dendroclimatology, but can also provide a new vision for studying the adaptation process and response strategies of tree growth to climate change. In this paper, with the relationships between the anatomical characteristics of xylem in tree-rings (cell chronology) and climate change as a main thread, we first outline the basic principles and mechanisms of wood anatomical features to record environmental signals, and expounded the basic methods involved in the process of xylem anatomy. Secondly, we discuss the relationship between the anatomical features of xylem and climate factors. We then propose the following as possible directions of future research based on the existing knowledge gap in the topical area: (1) to explore the temporal and spatial variations in the anatomical characteristics of xylem in tree-rings along radial and tangential directions and the relationships with environmental changes; (2) to explore the threshold of tree growth response to environmental plasticity and adaptation processes; (3) to assess the synergistic and antagonistic effects as well as the formation mechanisms of climate response among different tree-ring proxies, and to determine the specific roles and contributions of major climatic factors during different periods of tree-ring formation.
显示原图|下载原图ZIP|生成PPT 图3低降水与高温导致树木形成窄轮的部分响应机制。红色、蓝色和黑色区域分别表示生长季前、生长季及共同的过程; 高降水与低温导致树木形成宽轮的机制与其相反。改自Fritts (1976)。 -->Fig. 3The partial response mechanisms of narrowing in tree-rings caused by low precipitation and high temperatures. The areas marked by blue, red and black indicate the processes occurring prior to the growing season, during the growing season and the entire time, respectively. The effects of high precipitation and low temperatures are in reverse, that is, ring width will increase. Modified from Fritts (1976). -->
AbrantesJ, CampeloF, García-GonzálezI, NabaisC (2013). Environmental control of vessel traits in Quercus ilex under Mediterranean climate: Relating xylem anatomy to function. Trees, 27, 655-662. [本文引用: 5]
[2]
AloniR (2001). Foliar and axial aspects of vascular differentia- tion: Hypotheses and evidence. Journal of Plant Growth Regulation, 20, 22-34. [本文引用: 1]
[3]
ArbellayE, StoffelM, BollschweilerM (2010). Wood ana- tomical analysis of Alnus incana and Betula pendula injured by a debris-flow event. Tree Physiology, 30, 1290-1298. [本文引用: 3]
[4]
BarbarouxC, BrédaN (2002). Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. Tree Physiology, 22, 1201-1210. [本文引用: 2]
[5]
BattipagliaG, de MiccoV, BrandWA, LinkeP, AronneG, SaurerM, CherubiniP (2010). Variations of vessel diameter and δ13C in false rings of Arbutus unedo L. reflect different environmental conditions. New Phytologist, 188, 1099-1112. [本文引用: 5]
[6]
BriffaK, OsbornT, SchweingruberF (2004). Large-scale temperature inferences from tree rings: A review. Global and Planetary Change, 40, 11-26. [本文引用: 2]
[7]
CampeloF, NabaisC, CarvalhoA, VieiraJ (2016). tracheideR —An R package to standardize tracheidograms. Dendrochronologia, 37, 64-68. [本文引用: 1]
[8]
CampeloF, NabaisC, GutiérrezE, FreitasH, García-GonzálezI (2010). Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees, 24, 463-470. [本文引用: 6]
[9]
CampeloF, VieiraJ, BattipagliaG, de LuisM, NabaisC, FreitasH, CherubiniP (2015). Which matters most for the formation of intra-annual density fluctuations in Pinus pinaster: Age or size? Trees, 29, 237-245. [本文引用: 3]
[10]
CarlquistS, HoekmanDA (1985). Ecological wood anatomy of the woody southern Californian flora. IAWA Journal, 6, 319-347. [本文引用: 1]
[11]
CarrerM, vonAG, CastagneriD, PetitG (2015). Distilling allometric and environmental information from time series of conduit size: The standardization issue and its relationship to tree hydraulic architecture. Tree Physiology, 35, 27-33. [本文引用: 5]
[12]
CarvalhoA, NabaisC, VieiraJ, RossiS, CampeloF (2015). Plastic response of tracheids in Pinus pinaster in a water- limited environment: Adjusting lumen size instead of wall thickness. PLOS ONE, 10, e0136305. doi: 10.1371/journal. pone.0136305. [本文引用: 3]
[13]
CastagneriD, PetitG, CarrerM (2015). Divergent climate response on hydraulic-related xylem anatomical traits of Picea abies along a 900-m altitudinal gradient. Tree Physiology, 35, 1378-1387. [本文引用: 1]
[14]
CorcueraL, CamareroJJ, Gil-PelegrínE (2004). Effects of a severe drought on growth and wood anatomical properties of Quercus faginea. IAWA Journal, 25, 185-204. [本文引用: 1]
[15]
D’ArrigoR, FrankD, JacobyG, PedersonN (2001). Spatial response to major volcanic events in or about AD 536, 934 and 1258: Frost rings and other dendrochronological evidence from Mongolia and Northern Siberia: comment on RB Stothers, ‘Volcanic dry fogs, climate cooling, and plague pandemics in Europe and the Middle East’. Climatic Change, 49, 239-246. [本文引用: 2]
[16]
de LuisM, NovakK, RaventósJ, GričarJ, PrislanP, ČufarK (2011). Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia, 29, 163-169. [本文引用: 2]
[17]
de MiccoV, CampeloF, LuisMD, BrauningA, GrabnerM, BattipagliaG (2016). Intra-annual density fluctuations in tree rings: How, when, where, and why? IAWA Journal, 37, 232-259. [本文引用: 1]
[18]
DenneM (1971). Temperature and tracheid development in Pinus sylvestris seedlings. Journal of Experimental Botany, 22, 362-370. [本文引用: 1]
[19]
DeslauriersA, MorinH (2005). Intra-annual tracheid production in balsam fir stems and the effect of meteorological variables. Trees, 19, 402-408. [本文引用: 1]
[20]
DeslauriersA, MorinH, BeginY (2003). Cellular phenology of annual ring formation of Abies balsamea in the Québec boreal forest (Canada). Canadian Journal of Forest Research, 33, 190-200.
[21]
DeSotoL, de la CruzM, FontiP (2011). Intra-annual patterns of tracheid size in the Mediterranean tree Juniperus thurifera as an indicator of seasonal water stress. Canadian Journal of Forest Research, 41, 1280-1294. [本文引用: 1]
[22]
EilmannB, ZweifelR, BuchmannN, PannatierEG, RiglingA (2011). Drought alters timing, quantity, and quality of wood formation in Scots pine. Journal of Experimental Botany, 62, 2763-2771. [本文引用: 5]
[23]
EsperJ, BüntgenU, FrankDC, NievergeltD, LiebholdA (2007). 1200 years of regular outbreaks in alpine insects. Proceedings of the Royal Society of London B: Biological Sciences, 274, 671-679. [本文引用: 1]
[24]
EsperJ, CookER, SchweingruberFH (2002). Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science, 295, 2250-2253. [本文引用: 2]
[25]
FebruaryEC (1994). Rainfall reconstruction using wood charcoal from two archaeological sites in south Africa. Quaternary Research, 42, 100-107. [本文引用: 2]
[26]
FontiP, BryukhanovaMV, MyglanVS, KirdyanovAV, NaumovaOV, VaganovEA (2013). Temperature-induced responses of xylem structure of Larix sibirica (Pinaceae) from the Russian Altay. American Journal of Botany, 100, 1332-1343. [本文引用: 3]
[27]
FontiP, García-GonzálezI (2004). Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytologist, 163, 77-86. [本文引用: 9]
[28]
FontiP, von ArxG, García-GonzálezI, EilmannB, Sass- KlaassenU, GärtnerH, EcksteinD (2010). Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytologist, 185, 42-53. [本文引用: 23]
[29]
FrittsHC (1976). Tree Rings and Climate . Elsevier, Amsterdam, the Netherland. [本文引用: 7]
[30]
García-GonzálezI, FontiP (2006). Selecting earlywood vessels to maximize their environmental signal. Tree Physiology, 26, 1289-1296. [本文引用: 2]
[31]
García-GonzálezI, FontiP (2008). Ensuring a representative sample of earlywood vessels for dendroecological studies: An example from two ring-porous species. Trees, 22, 237-244. [本文引用: 1]
[32]
GärtnerH, CherubiniP, FontiP, von ArxG, SchneiderL, NievergeltD, VerstegeA, BastA, SchweingruberFH, BüntgenU (2015). A technical perspective in modern tree-ring research—How to overcome dendroecological and wood anatomical challenges. Journal of Visualized Experiments, 97, e52337. doi: 10.3791/52337. [本文引用: 4]
[33]
GärtnerH, LucchinettiS, SchweingruberFH (2014). New perspectives for wood anatomical analysis in dendrosci- ences: The GSL1-microtome. Dendrochronologia, 32, 47-51. [本文引用: 5]
[34]
GärtnerH, NievergeltD (2010). The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia, 28, 85-92. [本文引用: 5]
[35]
Gea-IzquierdoG, FontiP, CherubiniP, Martín-BenitoD, ChaarH, CañellasI (2012). Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability. Tree Physiology, 32, 401-423. [本文引用: 9]
[36]
GiantomasiMA, JunentFAR, VillagraPE, SrurAM (2009). Annual variation and influence of climate on the ring width and wood hydrosystem of Prosopis flexuosa DC. trees using image analysis. Trees, 23, 117-126. [本文引用: 7]
[37]
GrabnerM (2005). Functional Tree Ring Analysis: Wood as an Information Source to Understand Physiological, Environ- mental and Technological Questions . PhD dissertation, University of Natural Resources and Life Sciences, Vienna. [本文引用: 1]
[38]
GričarJ, de LuisM, HafnerP, LevaničT (2013). Anatomical characteristics and hydrologic signals in tree-rings of oaks (Quercus robur L.). Trees, 27, 1669-1680. [本文引用: 6]
[39]
GričarJ, ZupančičM, ČufarK, KochG, SchmittU, OvenP (2006). Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). Annals of Botany, 97, 943-951. [本文引用: 2]
[40]
GruberA, StroblS, VeitB, OberhuberW (2010). Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiology, 30, 490-501. [本文引用: 3]
[41]
HackeUG, SperryJS, WheelerJK, CastroL (2006). Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiology, 26, 689-701. [本文引用: 2]
[42]
KozlowskiTT, PallardySG (1997). Growth Control in Woody Plants. Academic Press, New York. [本文引用: 1]
[43]
LealS, SousaVB, KnapicS, LouzadaJL, PereiraH (2011). Vessel size and number are contributors to define wood density in cork oak. European Journal of Forest Research, 130, 1023-1029. [本文引用: 2]
[44]
LeikolaM (1969). The influence of environmental factors on the diameter growth of forest trees: Auxanometric study. Acta Forestalia Fennica, 92, 59-88. [本文引用: 1]
[45]
LiangW, HeinrichI, SimardS, HelleG, LiñánID, HeinkenT (2013). Climate signals derived from cell anatomy of Scots pine in NE Germany. Tree Physiology, 33, 833-844. [本文引用: 15]
[46]
LiuYJ, ZhuLJ, SuJJ, WangXC (2015). Impact of decreasing precipitation on Larix gmelinii radial growth in Maoershan, Xiaoxing’an Mountain, China. Acta Ecologica Sinica, 35, 4527-4537.(in Chinese with English abstract) [刘玉佳, 朱良军, 苏金娟, 王晓春 (2015). 模拟降水减少对帽儿山地区兴安落叶松径向生长的影响 . 生态学报, 35, 4527-4537.] [本文引用: 1]
[47]
LoepfeL, Martinez-VilaltaJ, PinolJ, MencucciniM (2007). The relevance of xylem network structure for plant hydraulic efficiency and safety. Journal of Theoretical Biology, 247, 788-803. [本文引用: 2]
[48]
Martin-BenitoD, BeeckmanH, CanellasI (2013). Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. European Journal of Forest Research, 132, 33-45. [本文引用: 4]
[49]
MatisonsR, ElfertsD, BrūmelisG (2012). Changes in climatic signals of English oak tree-ring width and cross-section area of earlywood vessels in Latvia during the period 1900-2009. Forest Ecology and Management, 279, 34-44. [本文引用: 6]
[50]
MencucciniM (2003). The ecological significance of long-distance water transport: Short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms. Plant, Cell & Environment, 26, 163-182. [本文引用: 2]
NovakK, LuísMD, RaventósJ, ČufarK (2013). Climatic signals in tree-ring widths and wood structure of Pinus halepensis in contrasted environmental conditions. Trees, 27, 927-936. [本文引用: 4]
[53]
OlanoJM, ArzacA, García-CervigónAI, ArxG, RozasV (2013). New star on the stage: Amount of ray parenchyma in tree rings shows a link to climate. New Phytologist, 198, 486-495. [本文引用: 3]
[54]
OlanoJM, EugenioM, García-CervigónAI, FolchM, RozasV (2012). Quantitative tracheid anatomy reveals a complex environmental control of wood structure in continental Mediterranean climate. International Journal of Plant Sciences, 173, 137-149. [本文引用: 2]
[55]
PanyushkinaIP, HughesMK, VaganovEA, MunroMA (2003). Summer temperature in northeastern Siberia since 1642 reconstructed from tracheid dimensions and cell numbers of Larix cajanderi. Canadian Journal of Forest Research, 33, 1905-1914.
[56]
Pérez-De-LisG, RossiS, Vázquez-RuizRA, RozasV, García- GonzálezI (2016). Do changes in spring phenology affect earlywood vessels? Perspective from the xylogenesis monitoring of two sympatric ring-porous oaks. New Phytologist, 209, 521-530.
[57]
PetersRL, GroenendijkP, VlamM, ZuidemaPA (2015). Detecting long-term growth trends using tree rings: A critical evaluation of methods. Global Change Biology, 21, 2040-2054. [本文引用: 2]
[58]
PiermatteiA, CrivellaroA, CarrerM, UrbinatiC (2014). The “blue ring”: Anatomy and formation hypothesis of a new tree-ring anomaly in conifers. Trees, 29, 455-461. [本文引用: 1]
[59]
PritzkowC, HeinrichI, GruddH, HelleG (2014). Relationship between wood anatomy, tree-ring widths and wood density of Pinus sylvestris L. and climate at high latitudes in northern Sweden. Dendrochronologia, 32, 295-302. [本文引用: 2]
[60]
PritzkowC, WaznyT, HeußnerKU, SłowińskiM, BieberA, LiñánID (2016). Minimum winter temperature reconstruc- tion from average earlywood vessel area of european European oak (Quercus robur) in N-Poland. Palaeoge- ography Palaeoclimatology Palaeoecology, 449, 520-530. [本文引用: 2]
[61]
RiglingA, BrühlhartH, BräkerOU, ForsterT, SchweingruberFH (2003). Effects of irrigation on diameter growth and vertical resin duct production in Pinus sylvestris L. on dry sites in the central Alps, Switzerland. Forest Ecology & Management, 175, 285-296. [本文引用: 2]
[62]
RitaA, CherubiniP, LeonardiS, TodaroL, BorghettiM (2015). Functional adjustments of xylem anatomy to climatic variability: Insights from long-term Ilex aquifolium tree-ring series. Tree Physiology, 35, 817-828. [本文引用: 3]
[63]
Rodríguez-GarcíaA, MartínJA, LópezR, MutkeS, PinillosF, GilL (2015). Influence of climate variables on resin yield and secretory structures in tapped Pinus pinaster Ait. in central Spain. Agricultural & Forest Meteorology, 202, 83-93. [本文引用: 2]
[64]
RossiS, DeslauriersA, GriçarJ, SeoJW, RathgeberCB, AnfodilloT, MorinH, LevanicT, OvenP, JalkanenR (2008). Critical temperatures for xylogenesis in conifers of cold climates. Global Ecology and Biogeography, 17, 696-707. [本文引用: 1]
[65]
RossiS, MorinH, DeslauriersA, PlourdePY (2011). Predicting xylem phenology in black spruce under climate warming. Global Change Biology, 17, 614-625. [本文引用: 1]
[66]
SassU, EcksteinD (1995). The variability of vessel size in beech (Fagus sylvatica L.) and its ecophysiological interpretation. Trees, 9, 247-252. [本文引用: 3]
[67]
SchmittU, MöllerR, EcksteinD (2000). Seasonal wood formation dynamics of beech (Fagus sylvatica L.) and black locust (Robinia pseudoacacia L.) as determined by the “pinning” technique. Angewandte Botanik, 74, 10-16. [本文引用: 1]
[68]
SchochWH, HellerI, SchweingruberFH, KienastF (2004). Wood Anatomy of Central European Species .(. Cited: 2016-09-16.URL [本文引用: 2]
[69]
SchumeH, GrabnerM, EckmüllnerO (2004). The influence of an altered groundwater regime on vessel properties of hybrid poplar. Trees, 18, 184-194. [本文引用: 2]
[70]
SmithKT, SutherlandEK (1999). Fire-scar formation and compartmentalization in oak. Canadian Journal of Forest Research, 29, 166-171. [本文引用: 4]
[71]
SperryJS, HackeUG, PittermannJ (2006). Size and function in conifer tracheids and angiosperm vessels. American Journal of Botany, 93, 1490-1500. [本文引用: 2]
[72]
StoffelM, BollschweilerM (2008). Tree-ring analysis in natural hazards research? An overview. Natural Hazards and Earth System Science, 8, 187-202. [本文引用: 1]
[73]
SuzukiM, YodaK, SuzukiH (1996). Phenological comparison of the onset of vessel formation between ring-porous and diffuse-porous deciduous trees in a Japanese temperate forest. IAWA Journal, 17, 431-444. [本文引用: 5]
[74]
TyreeMT, SperryJS (1989). Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Biology, 40, 19-36. [本文引用: 1]
[75]
TyreeMT, ZimmermannMH (2013). Xylem Structure and the Ascent of Sap . Springer, Berlin. [本文引用: 1]
[76]
UrsacheR, NieminenK, HelariuttaY (2013). Genetic and hormonal regulation of cambial development. Physiologia Plantarum, 147, 36-45. [本文引用: 1]
[77]
VaganovEA (1990). The Tracheidogram Method in Tree-Ring Analysis and Its Application. Methods of Dendrochro- nology: Applications in the Environmental Science . Kluwer Academic Publishers, Dordrecht, the Netherlands.63-75.
[78]
VaganovEA (1996).Recording of Warming in Current Century by Tracheids of the Annual Tree Rings . Doklady Akademii Nauk, Moscow, Russia. [本文引用: 11]
[79]
VaganovEA, AnchukaitisKJ, EvansMN (2011). How well understood are the processes that create dendroclimatic records? A mechanistic model of the climatic control on conifer tree-ring growth dynamics. Dendroclimatology, 11, 37-75. [本文引用: 5]
[80]
VaganovEA, HughesMK, ShashkinAV (2006). Growth Dynamics of Conifer Tree Rings . Springer, Berlin. [本文引用: 14]
[81]
Venegas-GonzálezA, von ArxG, ChagasMP, Tomazello FilhoM (2015). Plasticity in xylem anatomical traits of two tropical species in response to intra-seasonal climate variability. Trees, 29, 423-435. [本文引用: 1]
[82]
VieiraJ, CampeloF, RossiS, CarvalhoA, FreitasH, NabaisC (2015). Adjustment capacity of maritime pine cambial activity in drought-prone environments. PLOS ONE, 10, e0126223. doi:10.1371/journal.pone.0126223. [本文引用: 2]
[83]
von ArxG, CarrerM (2014). ROXAS—A new tool to build centuries-long tracheid-lumen chronologies in conifers. Dendrochronologia, 32, 290-293. [本文引用: 1]
[84]
WangLL, PayetteS, BéginY (2002). Relationships between anatomical and densitometric characteristics of black spruce and summer temperature at tree line in northern Quebec. Canadian Journal of Forest Research, 32, 477-486. [本文引用: 4]
[85]
WertzEL, St GeorgeS, ZeleznikJD (2013). Vessel anomalies in Quercus macrocarpa tree rings associated with recent floods along the Red River of the North, United States. Water Resources Research, 49, 630-634. [本文引用: 4]
[86]
WimmerR (2002). Wood anatomical features in tree-rings as indicators of environmental change. Dendrochronologia, 20, 21-36. [本文引用: 1]
[87]
WimmerR, GrabnerM (1997). Effects of climate on vertical resin duct density and radial growth of norway spruce [Picea abies (L.) Karst.]. Trees, 11, 271-276. [本文引用: 1]
[88]
WimmerR, GrabnerM (2000). A comparison of tree-ring features in Picea abies as correlated with climate. IAWA Journal, 21, 403-416.
[89]
WimmerR, VetterRE (1999). Tree-ring analysis biological, methodological and environmental aspects. In: Wimmer R, Grabner M, Strumia G eds. Significance of Vertical Ducts in the Tree Rings of Spruce. CABI Publishing, London. 1107-1110. [本文引用: 2]
[90]
WoodLJ, SmithDJ, HartleyID (2016). Predicting softwood quality attributes from climate data in interior British Columbia, Canada. Forest Ecology and Management, 361, 81-89. [本文引用: 4]
[91]
XuJM, LvJX, BaoFC, EvansR, DownesG, HuangRF, ZhaoYK (2012). Cellulose microfibril angle variation in Picea crassifolia tree rings improves climate signals on the Tibetan Plateau. Trees, 26, 1007-1016. [本文引用: 2]
[92]
YangB, QinC, WangJ, HeM, MelvinTM, OsbornTJ, BriffaKR (2014). A 3,500-year tree-ring record of annual pre- cipitation on the northeastern Tibetan Plateau . Proceedings of the National Academy of Sciences of the United States of America, 111, 2903-2908. [本文引用: 1]
[93]
YasueK, FunadaR, KobayashiO, OhtaniJ (2000). The effects of tracheid dimensions on variations in maximum density of Picea glehnii and relationships to climatic factors. Trees, 14, 223-229. [本文引用: 4]
[94]
ZimrnennannM (1983). Xylem Structure and the Ascent of Sap . Springer, Berlin, Germany. [本文引用: 2]
Environmental control of vessel traits in Quercus ilex under Mediterranean climate: Relating xylem anatomy to function. 5 2013
... 树轮解剖是基于树轮年代学与木材解剖学方法, 以获得定年年轮序列上细胞尺度解剖特征(如导管大小、密度、细胞壁厚度和组织含量等), 并通过其时间(年内或年际尺度)序列分析树木生长与环境变化关系的一种多学科方法(Fonti et al., 2010).探究树轮解剖特征与环境因子的潜在关系是解决上述问题的关键, 也是当前树轮生态学研究的一个重要内容(Fonti et al., 2010).木质部解剖特征是树轮在细胞尺度上的分量, 许多解剖特征已被证实能准确记录与传统代用指标不同或更强的环境信息(Campelo et al., 2010; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 近年来, 利用SilviScan-3设备可快速分析测定年轮宽度、细胞数量、径向及切向直径、细胞壁厚度、木材密度及微纤丝角度等多项宏微观年轮特征.虽然此设备价格昂贵, 但因其测量指标多、快速精准且不需要单独进行图像分析, 已逐渐受研究者们青睐(Xu et al., 2012; Abrantes et al., 2013; von Arx & Carrer, 2014; Wood et al., 2016).需要强调的是, 无论采用哪种方法, 准确交叉定年均是精准获取树轮木质部显微解剖特征的前提. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Foliar and axial aspects of vascular differentia- tion: Hypotheses and evidence. 1 2001
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
Wood ana- tomical analysis of Alnus incana and Betula pendula injured by a debris-flow event. 3 2010
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. 2 2002
Variations of vessel diameter and δ13C in false rings of Arbutus unedo L. reflect different environmental conditions. 5 2010
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
Large-scale temperature inferences from tree rings: A review. 2 2004
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... ; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
tracheideR —An R package to standardize tracheidograms. 1 2016
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. 6 2010
... 树轮解剖是基于树轮年代学与木材解剖学方法, 以获得定年年轮序列上细胞尺度解剖特征(如导管大小、密度、细胞壁厚度和组织含量等), 并通过其时间(年内或年际尺度)序列分析树木生长与环境变化关系的一种多学科方法(Fonti et al., 2010).探究树轮解剖特征与环境因子的潜在关系是解决上述问题的关键, 也是当前树轮生态学研究的一个重要内容(Fonti et al., 2010).木质部解剖特征是树轮在细胞尺度上的分量, 许多解剖特征已被证实能准确记录与传统代用指标不同或更强的环境信息(Campelo et al., 2010; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 样品预处理(切片或样芯)完毕即可利用超高分辨率扫描仪(仅适用于打磨法)或高像素数码相机将清晰的切片或样芯表面转换为数字化图像.随后, 利用专业图像分析软件或辅助测量软件(如WinCELL、Image-Pro Plus、ROXAS及MiVnt图像分析系统等)自动或半自动化读取及测量木质部解剖特征(Fonti et al., 2010; Liang et al., 2013; Venegas- González et al., 2015).借助辅助测量软件目标控制属性预设好提取目标的范围(如阔叶树导管大小及长宽比范围等), 自动捕捉图像中细胞或腔隙等解剖结构.结合人工辅助检查纠错, 便可高效且相对准确地完成测量(Campelo et al., 2010; von Arx & Carrer, 2014; Gärtner et al., 2015). ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 树轮木质部解剖使树木生长与环境变化关系的研究在时间尺度上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010).木质部解剖特征(如导管或管胞的尺寸、密度及壁厚等)记录着与传统树轮指标不同的气候信号(Campelo et al., 2010; Liang et al., 2013; Venegas-González et al., 2015), 是对传统树木年轮学的继承与发展.树轮木质部解剖为从生理学上解释年轮-气候响应关系奠定了基础, 为研究全球变化背景下物种的适应过程与响应策略提供了新视野, 并为科学评估未来气候变化对植被的影响提供了依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
Which matters most for the formation of intra-annual density fluctuations in Pinus pinaster: Age or size? 3 2015
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Ecological wood anatomy of the woody southern Californian flora. 1 1985
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Distilling allometric and environmental information from time series of conduit size: The standardization issue and its relationship to tree hydraulic architecture. 5 2015
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... ; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... ., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... ; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Plastic response of tracheids in Pinus pinaster in a water- limited environment: Adjusting lumen size instead of wall thickness. 3 2015
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... ; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
Divergent climate response on hydraulic-related xylem anatomical traits of Picea abies along a 900-m altitudinal gradient. 1 2015
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
Effects of a severe drought on growth and wood anatomical properties of Quercus faginea. 1 2004
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Spatial response to major volcanic events in or about AD 536, 934 and 1258: Frost rings and other dendrochronological evidence from Mongolia and Northern Siberia: comment on RB Stothers, ‘Volcanic dry fogs, climate cooling, and plague pandemics in Europe and the Middle East’. 2 2001
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. 2 2011
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
Intra-annual density fluctuations in tree rings: How, when, where, and why? 1 2016
Temperature and tracheid development in Pinus sylvestris seedlings. 1 1971
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
Intra-annual tracheid production in balsam fir stems and the effect of meteorological variables. 1 2005
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
Cellular phenology of annual ring formation of Abies balsamea in the Québec boreal forest (Canada). 2003
Intra-annual patterns of tracheid size in the Mediterranean tree Juniperus thurifera as an indicator of seasonal water stress. 1 2011
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
Drought alters timing, quantity, and quality of wood formation in Scots pine. 5 2011
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
1200 years of regular outbreaks in alpine insects. 1 2007
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. 2 2002
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... ; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
Rainfall reconstruction using wood charcoal from two archaeological sites in south Africa. 2 1994
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Temperature-induced responses of xylem structure of Larix sibirica (Pinaceae) from the Russian Altay. 3 2013
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... )或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Suitability of chestnut earlywood vessel chronologies for ecological studies. 9 2004
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... ; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... ).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... )、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. 23 2010
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... 树轮解剖是基于树轮年代学与木材解剖学方法, 以获得定年年轮序列上细胞尺度解剖特征(如导管大小、密度、细胞壁厚度和组织含量等), 并通过其时间(年内或年际尺度)序列分析树木生长与环境变化关系的一种多学科方法(Fonti et al., 2010).探究树轮解剖特征与环境因子的潜在关系是解决上述问题的关键, 也是当前树轮生态学研究的一个重要内容(Fonti et al., 2010).木质部解剖特征是树轮在细胞尺度上的分量, 许多解剖特征已被证实能准确记录与传统代用指标不同或更强的环境信息(Campelo et al., 2010; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... ; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... ).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... ; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... 维管植物形成层为双向活动模式, 可向内形成木质部, 向外形成韧皮部, 其活动受基因(Ursache et al., 2013)和环境(Rossi et al., 2008)共同控制.环境(气候)变化能够通过影响植物体内的各种生理进程, 直接或间接地影响形成层活动(树轮解剖特征的形成), 使树体的结构和功能(如年轮宽度或密度等宏观特征)发生改变, 以适应当前环境(图1).如生长季前或生长季的低降水或高温(干旱胁迫)能直接或间接地影响多种生理过程, 使部分树木管胞直径变小且数量减少, 最终形成窄轮(Fritts, 1976).正是这种塑性响应关系, 确保了树轮解剖特征能够准确记录周边环境(气候)的高频变化(Fonti et al., 2010).需要强调的是, 这种塑性响应关系存在一定的阈值范围, 环境变化超出该阈值将造成树体发生不可逆伤害甚至死亡(Fonti et al., 2010), 如严重的霜冻灾害.此外, 树轮微观解剖特征能够随外界环境变化而塑性改变, 当这种改变能够适应现有气候时, 植物能够正常生长; 而当这种改变不足以适应现有气候时, 树木将会出现衰退或死亡(图1). ...
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... ; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... .目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... )(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... 样品预处理(切片或样芯)完毕即可利用超高分辨率扫描仪(仅适用于打磨法)或高像素数码相机将清晰的切片或样芯表面转换为数字化图像.随后, 利用专业图像分析软件或辅助测量软件(如WinCELL、Image-Pro Plus、ROXAS及MiVnt图像分析系统等)自动或半自动化读取及测量木质部解剖特征(Fonti et al., 2010; Liang et al., 2013; Venegas- González et al., 2015).借助辅助测量软件目标控制属性预设好提取目标的范围(如阔叶树导管大小及长宽比范围等), 自动捕捉图像中细胞或腔隙等解剖结构.结合人工辅助检查纠错, 便可高效且相对准确地完成测量(Campelo et al., 2010; von Arx & Carrer, 2014; Gärtner et al., 2015). ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
... 树轮木质部解剖使树木生长与环境变化关系的研究在时间尺度上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010).木质部解剖特征(如导管或管胞的尺寸、密度及壁厚等)记录着与传统树轮指标不同的气候信号(Campelo et al., 2010; Liang et al., 2013; Venegas-González et al., 2015), 是对传统树木年轮学的继承与发展.树轮木质部解剖为从生理学上解释年轮-气候响应关系奠定了基础, 为研究全球变化背景下物种的适应过程与响应策略提供了新视野, 并为科学评估未来气候变化对植被的影响提供了依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
... ; Fonti et al., 2010; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
Tree Rings and Climate 7 1976
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... ).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... ).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... 树轮解剖是基于树轮年代学与木材解剖学方法, 以获得定年年轮序列上细胞尺度解剖特征(如导管大小、密度、细胞壁厚度和组织含量等), 并通过其时间(年内或年际尺度)序列分析树木生长与环境变化关系的一种多学科方法(Fonti et al., 2010).探究树轮解剖特征与环境因子的潜在关系是解决上述问题的关键, 也是当前树轮生态学研究的一个重要内容(Fonti et al., 2010).木质部解剖特征是树轮在细胞尺度上的分量, 许多解剖特征已被证实能准确记录与传统代用指标不同或更强的环境信息(Campelo et al., 2010; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... 维管植物形成层为双向活动模式, 可向内形成木质部, 向外形成韧皮部, 其活动受基因(Ursache et al., 2013)和环境(Rossi et al., 2008)共同控制.环境(气候)变化能够通过影响植物体内的各种生理进程, 直接或间接地影响形成层活动(树轮解剖特征的形成), 使树体的结构和功能(如年轮宽度或密度等宏观特征)发生改变, 以适应当前环境(图1).如生长季前或生长季的低降水或高温(干旱胁迫)能直接或间接地影响多种生理过程, 使部分树木管胞直径变小且数量减少, 最终形成窄轮(Fritts, 1976).正是这种塑性响应关系, 确保了树轮解剖特征能够准确记录周边环境(气候)的高频变化(Fonti et al., 2010).需要强调的是, 这种塑性响应关系存在一定的阈值范围, 环境变化超出该阈值将造成树体发生不可逆伤害甚至死亡(Fonti et al., 2010), 如严重的霜冻灾害.此外, 树轮微观解剖特征能够随外界环境变化而塑性改变, 当这种改变能够适应现有气候时, 植物能够正常生长; 而当这种改变不足以适应现有气候时, 树木将会出现衰退或死亡(图1). ...
... The partial response mechanisms of narrowing in tree-rings caused by low precipitation and high temperatures. The areas marked by blue, red and black indicate the processes occurring prior to the growing season, during the growing season and the entire time, respectively. The effects of high precipitation and low temperatures are in reverse, that is, ring width will increase. Modified from Fritts (1976). ...
... 树轮木质部解剖使树木生长与环境变化关系的研究在时间尺度上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010).木质部解剖特征(如导管或管胞的尺寸、密度及壁厚等)记录着与传统树轮指标不同的气候信号(Campelo et al., 2010; Liang et al., 2013; Venegas-González et al., 2015), 是对传统树木年轮学的继承与发展.树轮木质部解剖为从生理学上解释年轮-气候响应关系奠定了基础, 为研究全球变化背景下物种的适应过程与响应策略提供了新视野, 并为科学评估未来气候变化对植被的影响提供了依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
Selecting earlywood vessels to maximize their environmental signal. 2 2006
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... )早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
Ensuring a representative sample of earlywood vessels for dendroecological studies: An example from two ring-porous species. 1 2008
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
A technical perspective in modern tree-ring research—How to overcome dendroecological and wood anatomical challenges. 4 2015
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... 样品预处理(切片或样芯)完毕即可利用超高分辨率扫描仪(仅适用于打磨法)或高像素数码相机将清晰的切片或样芯表面转换为数字化图像.随后, 利用专业图像分析软件或辅助测量软件(如WinCELL、Image-Pro Plus、ROXAS及MiVnt图像分析系统等)自动或半自动化读取及测量木质部解剖特征(Fonti et al., 2010; Liang et al., 2013; Venegas- González et al., 2015).借助辅助测量软件目标控制属性预设好提取目标的范围(如阔叶树导管大小及长宽比范围等), 自动捕捉图像中细胞或腔隙等解剖结构.结合人工辅助检查纠错, 便可高效且相对准确地完成测量(Campelo et al., 2010; von Arx & Carrer, 2014; Gärtner et al., 2015). ...
New perspectives for wood anatomical analysis in dendrosci- ences: The GSL1-microtome. 5 2014
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. 5 2010
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability. 9 2012
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... ; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... ), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... ).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
Annual variation and influence of climate on the ring width and wood hydrosystem of Prosopis flexuosa DC. trees using image analysis. 7 2009
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Functional Tree Ring Analysis: Wood as an Information Source to Understand Physiological, Environ- mental and Technological Questions 1 2005
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
Anatomical characteristics and hydrologic signals in tree-rings of oaks (Quercus robur L.). 6 2013
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... ; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... ; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). 2 2006
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. 3 2010
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
Scaling of angiosperm xylem structure with safety and efficiency. 2 2006
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
1 1997
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
Vessel size and number are contributors to define wood density in cork oak. 2 2011
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ; Leal et al., 2011)的导管密度几乎恒定. ...
The influence of environmental factors on the diameter growth of forest trees: Auxanometric study. 1 1969
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
Climate signals derived from cell anatomy of Scots pine in NE Germany. 15 2013
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... 树轮解剖是基于树轮年代学与木材解剖学方法, 以获得定年年轮序列上细胞尺度解剖特征(如导管大小、密度、细胞壁厚度和组织含量等), 并通过其时间(年内或年际尺度)序列分析树木生长与环境变化关系的一种多学科方法(Fonti et al., 2010).探究树轮解剖特征与环境因子的潜在关系是解决上述问题的关键, 也是当前树轮生态学研究的一个重要内容(Fonti et al., 2010).木质部解剖特征是树轮在细胞尺度上的分量, 许多解剖特征已被证实能准确记录与传统代用指标不同或更强的环境信息(Campelo et al., 2010; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... ); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... ; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... ; Liang et al., 2013; Venegas- González et al., 2015). ...
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... ; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... 样品预处理(切片或样芯)完毕即可利用超高分辨率扫描仪(仅适用于打磨法)或高像素数码相机将清晰的切片或样芯表面转换为数字化图像.随后, 利用专业图像分析软件或辅助测量软件(如WinCELL、Image-Pro Plus、ROXAS及MiVnt图像分析系统等)自动或半自动化读取及测量木质部解剖特征(Fonti et al., 2010; Liang et al., 2013; Venegas- González et al., 2015).借助辅助测量软件目标控制属性预设好提取目标的范围(如阔叶树导管大小及长宽比范围等), 自动捕捉图像中细胞或腔隙等解剖结构.结合人工辅助检查纠错, 便可高效且相对准确地完成测量(Campelo et al., 2010; von Arx & Carrer, 2014; Gärtner et al., 2015). ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
... 树轮木质部解剖使树木生长与环境变化关系的研究在时间尺度上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010).木质部解剖特征(如导管或管胞的尺寸、密度及壁厚等)记录着与传统树轮指标不同的气候信号(Campelo et al., 2010; Liang et al., 2013; Venegas-González et al., 2015), 是对传统树木年轮学的继承与发展.树轮木质部解剖为从生理学上解释年轮-气候响应关系奠定了基础, 为研究全球变化背景下物种的适应过程与响应策略提供了新视野, 并为科学评估未来气候变化对植被的影响提供了依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
... ; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
模拟降水减少对帽儿山地区兴安落叶松径向生长的影响 1 2015
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
The relevance of xylem network structure for plant hydraulic efficiency and safety. 2 2007
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. 4 2013
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... ; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Changes in climatic signals of English oak tree-ring width and cross-section area of earlywood vessels in Latvia during the period 1900-2009. 6 2012
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... ; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
The ecological significance of long-distance water transport: Short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms. 2 2003
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Highly variable northern hemisphere temperatures reconstructed from low- and high-resolution proxy data. 1 2005
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
Climatic signals in tree-ring widths and wood structure of Pinus halepensis in contrasted environmental conditions. 4 2013
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
New star on the stage: Amount of ray parenchyma in tree rings shows a link to climate. 3 2013
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Quantitative tracheid anatomy reveals a complex environmental control of wood structure in continental Mediterranean climate. 2 2012
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Summer temperature in northeastern Siberia since 1642 reconstructed from tracheid dimensions and cell numbers of Larix cajanderi. 2003
Do changes in spring phenology affect earlywood vessels? Perspective from the xylogenesis monitoring of two sympatric ring-porous oaks. 2016
Detecting long-term growth trends using tree rings: A critical evaluation of methods. 2 2015
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... ; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
The “blue ring”: Anatomy and formation hypothesis of a new tree-ring anomaly in conifers. 1 2014
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Relationship between wood anatomy, tree-ring widths and wood density of Pinus sylvestris L. and climate at high latitudes in northern Sweden. 2 2014
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
Minimum winter temperature reconstruc- tion from average earlywood vessel area of european European oak (Quercus robur) in N-Poland. 2 2016
Effects of irrigation on diameter growth and vertical resin duct production in Pinus sylvestris L. on dry sites in the central Alps, Switzerland. 2 2003
Critical temperatures for xylogenesis in conifers of cold climates. 1 2008
... 维管植物形成层为双向活动模式, 可向内形成木质部, 向外形成韧皮部, 其活动受基因(Ursache et al., 2013)和环境(Rossi et al., 2008)共同控制.环境(气候)变化能够通过影响植物体内的各种生理进程, 直接或间接地影响形成层活动(树轮解剖特征的形成), 使树体的结构和功能(如年轮宽度或密度等宏观特征)发生改变, 以适应当前环境(图1).如生长季前或生长季的低降水或高温(干旱胁迫)能直接或间接地影响多种生理过程, 使部分树木管胞直径变小且数量减少, 最终形成窄轮(Fritts, 1976).正是这种塑性响应关系, 确保了树轮解剖特征能够准确记录周边环境(气候)的高频变化(Fonti et al., 2010).需要强调的是, 这种塑性响应关系存在一定的阈值范围, 环境变化超出该阈值将造成树体发生不可逆伤害甚至死亡(Fonti et al., 2010), 如严重的霜冻灾害.此外, 树轮微观解剖特征能够随外界环境变化而塑性改变, 当这种改变能够适应现有气候时, 植物能够正常生长; 而当这种改变不足以适应现有气候时, 树木将会出现衰退或死亡(图1). ...
Predicting xylem phenology in black spruce under climate warming. 1 2011
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
The variability of vessel size in beech (Fagus sylvatica L.) and its ecophysiological interpretation. 3 1995
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Seasonal wood formation dynamics of beech (Fagus sylvatica L.) and black locust (Robinia pseudoacacia L.) as determined by the “pinning” technique. 1 2000
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
The influence of an altered groundwater regime on vessel properties of hybrid poplar. 2 2004
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Fire-scar formation and compartmentalization in oak. 4 1999
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
Size and function in conifer tracheids and angiosperm vessels. 2 2006
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Vulnerability of xylem to cavitation and embolism. 1 1989
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
Xylem Structure and the Ascent of Sap 1 2013
... 维管植物形成层为双向活动模式, 可向内形成木质部, 向外形成韧皮部, 其活动受基因(Ursache et al., 2013)和环境(Rossi et al., 2008)共同控制.环境(气候)变化能够通过影响植物体内的各种生理进程, 直接或间接地影响形成层活动(树轮解剖特征的形成), 使树体的结构和功能(如年轮宽度或密度等宏观特征)发生改变, 以适应当前环境(图1).如生长季前或生长季的低降水或高温(干旱胁迫)能直接或间接地影响多种生理过程, 使部分树木管胞直径变小且数量减少, 最终形成窄轮(Fritts, 1976).正是这种塑性响应关系, 确保了树轮解剖特征能够准确记录周边环境(气候)的高频变化(Fonti et al., 2010).需要强调的是, 这种塑性响应关系存在一定的阈值范围, 环境变化超出该阈值将造成树体发生不可逆伤害甚至死亡(Fonti et al., 2010), 如严重的霜冻灾害.此外, 树轮微观解剖特征能够随外界环境变化而塑性改变, 当这种改变能够适应现有气候时, 植物能够正常生长; 而当这种改变不足以适应现有气候时, 树木将会出现衰退或死亡(图1). ...
Genetic and hormonal regulation of cambial development. 1 2013
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
The Tracheidogram Method in Tree-Ring Analysis and Its Application. Methods of Dendrochro- nology: Applications in the Environmental Science 1990
Recording of Warming in Current Century by Tracheids of the Annual Tree Rings 11 1996
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... ; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
How well understood are the processes that create dendroclimatic records? A mechanistic model of the climatic control on conifer tree-ring growth dynamics. 5 2011
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... ), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
... 温度可通过影响多种生理过程直接或间接影响木质部管胞特征(图3)(Vaganov et al., 2006, 2011; Novak et al., 2013); 其中最直接的影响就是通过控制形成层活动调整树木的生长周期.生长季开始时, 外界环境温度的略微波动将导致形成层活动开始时间发生明显改变(Leikola, 1969), 只有当环境温度高于形成层活动的最低阈值温度时, 树木生长才可能被触发(Denne, 1971).模型预测发现温度上升1 ℃可导致Pinus thunbergii形成层活动开始时间提前4-5天, 结束期推迟4-6天, 生长季延长8-11天(Rossi et al., 2011).形成层细胞产生速率在很大程度上由同期温度决定, 如Deslauriers等(2003)发现北方针叶林内香脂冷杉(Abies balsamea)年轮形成各时期管胞数量变化剧烈, 早材形成时期可短至1周, 晚材形成周期短至5-10天.低温可缩短形成层活动周期、降低细胞分裂速率, 导致木质部晚材的比例降低(Gričar et al., 2006); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
... ); 高温则会延长生长季, 增加树轮中晚材的比例(Vaganov et al., 2006).温度对树木管胞解剖特征的影响会随研究区域发生变化, 高纬度地区树木形成层细胞增大阶段细胞伸长的时间明显短于低纬度地区植物(Vaganov et al., 2006), 其对外界温度变化响应更迅速(Deslauriers & Morin, 2005).影响早材管胞直径的初夏温度介于5-14 ℃, 接近或超出该界限温度时, 温度对管胞直径的影响减弱, 如Vaganov (1996)发现北方树线附近欧洲落叶松早材内管胞直径会因长期的夏季升温而显著增加, 而Denne (1971)则发现将欧洲赤松幼苗的培养温度从17.5 ℃增加到27.5 ℃, 管胞直径仅增加10%.另外, 管胞相关特征也能准确记录外界的温度变化(Vaganov et al., 2011; Liang et al., 2013).例如Panyushkina等(2003)研究发现西伯利亚东北部落叶松(Larix cajanderi), 早晚材管胞直径均与6月温度正相关, 晚材管胞直径(壁厚)与7月温度负(正)相关; 管胞数量与5月温度负相关, 与6月温度正相关; 管胞直径及数量可被用来重建1642年以来的夏季温度.一般来说, 早材内的管胞特征(如大小、数量、细胞壁厚度等)多受早春与前一年冬季温度的影响, 晚材内的管胞特征常与晚夏温度有关, 而早晚材过渡区的管胞特征多受春末夏初温度的影响(Novak et al., 2013). ...
... 树轮解剖是基于树轮年代学与木材解剖学方法, 以获得定年年轮序列上细胞尺度解剖特征(如导管大小、密度、细胞壁厚度和组织含量等), 并通过其时间(年内或年际尺度)序列分析树木生长与环境变化关系的一种多学科方法(Fonti et al., 2010).探究树轮解剖特征与环境因子的潜在关系是解决上述问题的关键, 也是当前树轮生态学研究的一个重要内容(Fonti et al., 2010).木质部解剖特征是树轮在细胞尺度上的分量, 许多解剖特征已被证实能准确记录与传统代用指标不同或更强的环境信息(Campelo et al., 2010; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas-González et al., 2015); 尤其是在非限制性地点, 木质部解剖特征具有巨大的应用潜能(Liang et al., 2013).树轮解剖使传统树轮研究在时间上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010), 开展此类研究既有利于进一步揭示树轮-气候响应关系, 为树轮气候学机制与机理研究奠定基础, 为传统年轮-气候关系及古气候重建研究开辟新天地, 又可为研究全球气候变化情境下物种的适应过程与响应策略提供新视野, 为科学评估未来气候变化对植被的影响提供依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).因此, 本文综述了树轮木质部解剖特征记录气候或环境信息的机理、木质部解剖特征研究的手段与方法、解剖特征的选取与标准化、主要解剖特征与气候变化关系以及未来展望等内容,以求为进一步开展树轮木质部解剖研究提供参考. ...
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 样品及取样过程中的质量控制是树轮解剖研究成败的关键.普通生长芯(直径一般为4.35、5.15或12 mm)、微生长芯(直径 1.9 mm)或圆盘均可用于分析.树轮木质部解剖特征相比传统年轮代用指标往往具有更高的变异性; 林分条件(如竞争和土壤条件)或个体特征(基因)等潜在差异均可能导致同一采样点不同个体之间甚至同一个体不同样本间的树轮解剖特征产生较大的变异(Gea-Izquierdo et al., 2012; Gričar et al., 2013).此外, 树轮木质部解剖特征获取耗时费力, 树轮解剖相关的研究复本量常低于传统树轮宽度研究(Gärtner & Nievergelt, 2010).因此, 多数研究表明细胞年表的统计特征往往低于宽度或密度等传统代用指标年表(Fonti & García- González, 2004; Gea-Izquierdo et al., 2012; Fonti et al., 2013; Liang et al., 2013; Venegas-González et al., 2015), 尽管这些统计学参数并不代表年表记录环境信息的强弱(Wimmer, 2002; Fonti & García- González, 2004; García-González & Fonti, 2008).部分年表质量统计特征已不太适合被用来描述细胞年表质量(Fonti & García-González, 2004; Giantomasi et al., 2009; Gričar et al., 2013; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... ; Venegas-González et al., 2015), 例如许多细胞年表样本总体代表性(EPS)均未达到阈值0.85.因此, 适当增加样本量、合理筛选相关性较高的样芯用于分析, 减少因年龄、样本量、微环境或干扰等因素而导致的变异, 有利于从树轮解剖特征中提取更准确的环境信号(Gea- Izquierdo et al., 2012; Gričar et al., 2013). ...
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 树轮木质部解剖研究中样品制备方法主要分为显微切片法和直接打磨法两大类(表2).显微切片法是树轮解剖学研究中最传统的方法(Fonti et al., 2010; Gärtner et al., 2014), 具有分辨率高、清晰度好、切片便于携带保存且能制成连续切片等优点(Fonti et al., 2010), 适用于小至细胞壁厚度、纹孔大小及木纤维厚度等亚细胞特征的量化.传统切片(微生长芯)制备太过复杂, 严重制约了树轮解剖学的发展(Gärtner et al., 2014, 2015).近年来, 随着树轮解剖学研究的逐渐增多, 部分专业用于制作树轮木质部大芯切片(普通生长芯)的切片机及配套软件被相继研发出来(Gärtner & Nievergelt, 2010; Gärtner et al., 2014), 明显简化了样品预处理的过程(表2), 例如运用GSL1-切片机可显著提高树轮木质部显微切片制作与解剖特征解读的效率(Gärtner & Niever- gelt, 2010; Gärtner et al., 2014).对于相对较大的木质部解剖结构(如阔叶树导管或针叶树管胞), 还可利用数字化成像技术从经打磨的样芯表面直接捕捉获得解剖特征, 从而提高工作效率(Giantomasi et al., 2009; Liang et al., 2013; Venegas- González et al., 2015; 刘玉佳等, 2015).因打磨易造成薄细胞壁断裂或细胞腔堵满灰尘, 降低样品表面的分辨率, 故采用直接打磨法捕捉木质部解剖结构时, 样品须经一系列特殊的表面处理, 以提高分辨率(表2)(Fonti et al., 2010; Gärtner et al., 2015; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... ; Venegas-González et al., 2015).如样品打磨后需用高压水(气)枪清洗样品表面及管腔内的杂物或灰尘, 确保样品表面洁净且管腔内无异物填充(Gärtner & Nievergelt, 2010; Matisons et al., 2012; Gärtner et al., 2015); 低分辨率样品需预先用深色墨水沾拭样品表面(表面暗化), 待墨迹干透再用明亮物质(如白色的粉笔、橡皮泥或蜡等)擦试样品表面(细胞腔亮化), 以增加样品细胞壁及腔隙间的对比度(Gärtner & Nievergelt, 2010; Gärtner et al., 2014; Venegas- González et al., 2015).此外, 为确保捕捉的准确性, 避免因自动捕捉错误而进行的频繁手动修改测量, 必须保证样品的年轮边界清楚, 且整个样芯的表面在一个平面内.故样品预处理质量(主要指打磨法)直接决定解剖特征解读的效率及准确性. ...
... 样品预处理(切片或样芯)完毕即可利用超高分辨率扫描仪(仅适用于打磨法)或高像素数码相机将清晰的切片或样芯表面转换为数字化图像.随后, 利用专业图像分析软件或辅助测量软件(如WinCELL、Image-Pro Plus、ROXAS及MiVnt图像分析系统等)自动或半自动化读取及测量木质部解剖特征(Fonti et al., 2010; Liang et al., 2013; Venegas- González et al., 2015).借助辅助测量软件目标控制属性预设好提取目标的范围(如阔叶树导管大小及长宽比范围等), 自动捕捉图像中细胞或腔隙等解剖结构.结合人工辅助检查纠错, 便可高效且相对准确地完成测量(Campelo et al., 2010; von Arx & Carrer, 2014; Gärtner et al., 2015). ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 树轮木质部解剖使树木生长与环境变化关系的研究在时间尺度上由年际尺度精确到月或季节尺度, 空间上由整个生长轮分辨到细胞或亚细胞尺度(Fonti et al., 2010).木质部解剖特征(如导管或管胞的尺寸、密度及壁厚等)记录着与传统树轮指标不同的气候信号(Campelo et al., 2010; Liang et al., 2013; Venegas-González et al., 2015), 是对传统树木年轮学的继承与发展.树轮木质部解剖为从生理学上解释年轮-气候响应关系奠定了基础, 为研究全球变化背景下物种的适应过程与响应策略提供了新视野, 并为科学评估未来气候变化对植被的影响提供了依据(Fritts, 1976; Fonti et al., 2010; Liang et al., 2013).考虑到树轮解剖学研究现状及其在全球变化研究中的广阔前景, 未来可在以下几个重要方面开展研究: ...
Plasticity in xylem anatomical traits of two tropical species in response to intra-seasonal climate variability. 1 2015
... 管胞是裸子植物木质部解剖研究中最常见解剖结构, 其在年轮内排列整齐且规则(Fonti et al., 2010; Campelo et al., 2016).管胞随形成时间在年轮内呈规律性变化且同一年内形成的管胞常被作为整体研究.管胞径向直径大小与在年轮内的相对位置的函数被称为“管胞图像(Tracheidograms)”.个体间或年际间管胞数量常存在差异(Vaganov et al., 2006; Carvalho et al., 2015; Vieira et al., 2015), 为比较不同年轮间的管胞或相同时期形成的管胞特征常需要对管胞图形进行管胞数量标准化即管胞图像标准化(Vaganov et al., 2006; Carvalho et al., 2015; Campelo et al., 2016).Vaganov (1990)最先提出利用某一常数来标准化所有年份管胞数目的标准化方法, 在不改变细胞尺寸变化曲线形状即纵坐标(径向细胞大小)不变的情况下, “压缩”或“延伸”初始管胞图像的横坐标(细胞数量)(Vaganov, 1990).该方法是目前公认的管胞图像标准化方法(Campelo et al., 2016), 适用于管胞数量变异小的情形.当管胞数量年际间变异较大时, 如地中海地区海岸松(Pinus pinaster)连续两年间管胞数量差异可达到20个以上(Carvalho et al., 2015; Vieira et al., 2015), 使用该标准化方法往往会低估观测值或损失部分显著的气候信号(Campelo et al., 2016).此后, 有****将Vaganov方法中的常数调整为相应年份内各径向管胞数量的均值, 允许不同年份拥有各自数量的细胞去标准化(DeSoto et al., 2011).上述两种方法均无法确定年轮中密度波动(IADFs)的相对比例, 这在地中海地区生长的针阔叶树中较为常见(Battipaglia et al., 2010; de Luis et al., 2011; Campelo et al., 2015; Carvalho et al., 2015).近年来, ****们还提出了基于年轮中管胞相对位置(距早材起始端距离)来进行管胞标准化的方法(Campelo et al., 2016).该方法适用于环境条件差异较大或管胞数量变异较高的环境, 且保持了准确的年内变化信息, 还提高了前两种方法在评估IADFs较大或管胞数量年际变异较高的年轮内晚材评估的准确性, 有利于更好地估测树木的生产力与固碳能力(Campelo et al., 2016).此外, 阔叶树中散孔材导管具有与针叶树管胞相似的特征, 也可利用管胞图像法标准化散孔材导管解剖特征(Grabner, 2005).阔叶树木质部解剖特征往往分布无规律、排列不规则, 其大小过渡急或不呈梯度, 如环孔材导管的大小或密度等(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013).针对类似排列不规则且呈非梯度性变化的解剖特征, 常以年轮内某部分解剖特征为对象开展研究, 不能用类似的管胞图像法来进行标准化. ...
Adjustment capacity of maritime pine cambial activity in drought-prone environments. 2 2015
... 样品预处理(切片或样芯)完毕即可利用超高分辨率扫描仪(仅适用于打磨法)或高像素数码相机将清晰的切片或样芯表面转换为数字化图像.随后, 利用专业图像分析软件或辅助测量软件(如WinCELL、Image-Pro Plus、ROXAS及MiVnt图像分析系统等)自动或半自动化读取及测量木质部解剖特征(Fonti et al., 2010; Liang et al., 2013; Venegas- González et al., 2015).借助辅助测量软件目标控制属性预设好提取目标的范围(如阔叶树导管大小及长宽比范围等), 自动捕捉图像中细胞或腔隙等解剖结构.结合人工辅助检查纠错, 便可高效且相对准确地完成测量(Campelo et al., 2010; von Arx & Carrer, 2014; Gärtner et al., 2015). ...
... 近年来, 利用SilviScan-3设备可快速分析测定年轮宽度、细胞数量、径向及切向直径、细胞壁厚度、木材密度及微纤丝角度等多项宏微观年轮特征.虽然此设备价格昂贵, 但因其测量指标多、快速精准且不需要单独进行图像分析, 已逐渐受研究者们青睐(Xu et al., 2012; Abrantes et al., 2013; von Arx & Carrer, 2014; Wood et al., 2016).需要强调的是, 无论采用哪种方法, 准确交叉定年均是精准获取树轮木质部显微解剖特征的前提. ...
ROXAS—A new tool to build centuries-long tracheid-lumen chronologies in conifers. 1 2014
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Relationships between anatomical and densitometric characteristics of black spruce and summer temperature at tree line in northern Quebec. 4 2002
... 根据外部环境事件是否具有连续性, 可将环境对树轮解剖特征的影响分为不连续的环境事件影响和连续环境事件影响两类(Fonti et al., 2010).不连续环境事件往往危害较大, 常在年轮中留下某些特殊的“痕迹”.****们常利用这些“痕迹”重建特殊的环境事件, 如林火(Smith & Sutherland, 1999)、虫灾(Esper et al., 2007)、干旱(Martin-Benito et al., 2013)、洪灾(Arbellay et al., 2010; Wertz et al., 2013)及霜冻事件(D’Arrigo et al., 2001).然而, 连续的环境变化(如温度或降水等)往往能被连续的树轮解剖特征序列(即细胞年表)所记录(Battipaglia et al., 2010; Matisons et al., 2012; Liang et al., 2013).多个树轮木质部解剖特征被证实能够准确记录周边的环境变化且具备气候重建的潜力(February, 1994; Fonti et al., 2010; Abrantes et al., 2013; Liang et al., 2013; Venegas- González et al., 2015). ...
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Vessel anomalies in Quercus macrocarpa tree rings associated with recent floods along the Red River of the North, United States. 4 2013
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
Predicting softwood quality attributes from climate data in interior British Columbia, Canada. 4 2016
... 木质部解剖特征会因其在树轮内的位置不同, 在径向(时间)和切向(空间)上呈现出较高的变异性, 如导管或管胞的大小及密度等(图2).不同位置或尺寸的解剖特征记录的环境信号也不尽相同(García- González & Fonti, 2006; Campelo et al., 2010; Fonti et al., 2010; Carrer et al., 2015), 如García-González和Fonti (2006)研究发现欧洲栗(Castanea sativa)早材中最大导管面积与3月温度相关性较强, 小导管面积与6月温度相关性更高, 而综合大、小导管面积则与温度不相关.故选定测量对象数量太少, 将不能完全反映相应时期的环境信号, 过多则会混入其他时期形成的细胞从而导致环境信号被遮蔽(García-González & Fonti, 2006, 2008).此外, 解剖特征记录的气候信号还会因研究区域、树种、生境或解剖特征的不同而不同(Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Liang et al., 2013; Martin-Benito et al., 2013; Castagneri et al.,2015; Venegas-González et al., 2015).树木种类及木质部解剖参数较多且年内空间分布复杂, 故选取适宜的树种以及解剖参数是研究木质部解剖特征与环境变化关系的关键.本文作者将文献和维基百科中查到的常用木质部解剖参数及特征总结于表1.目前, 许多树轮木质部解剖特征(细胞年表)已被证实能够准确记录亚年际尺度(日、月或季节)的气候变化(Fonti et al., 2010).针叶树研究主要集中在管胞密度、大小及细胞壁厚度(Olano et al., 2012; Liang et al., 2013; Martin- Benito et al., 2013; Pritzkow et al., 2014)、纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)、木质部年内密度波动(intra-annual density fluctuations, IADFs; 指正常生长季的生长过程中断引起局部木材密度的突然变化, 也称“伪轮”)(Campelo et al., 2015; Carvalho et al., 2015)以及树脂道(Novak et al., 2013)等方面; 而阔叶树种研究多关注于环孔材或半环孔材早材导管的大小及密度(Gea-Izquierdo et al., 2012; Matisons et al., 2012; Gričar et al., 2013), 也有就部分散孔材导管开展的探索性研究, 如水青冈属(Fagus)(Sass & Eckstein, 1995)、桦木属(Betula)(Arbellay et al., 2010)、杨属(Populus)(Schume et al., 2004)、Prosopis flexuosa (Giantomasi et al., 2009)及Arbutus unedo (Battipaglia et al., 2010)等. ...
... 近年来, 利用SilviScan-3设备可快速分析测定年轮宽度、细胞数量、径向及切向直径、细胞壁厚度、木材密度及微纤丝角度等多项宏微观年轮特征.虽然此设备价格昂贵, 但因其测量指标多、快速精准且不需要单独进行图像分析, 已逐渐受研究者们青睐(Xu et al., 2012; Abrantes et al., 2013; von Arx & Carrer, 2014; Wood et al., 2016).需要强调的是, 无论采用哪种方法, 准确交叉定年均是精准获取树轮木质部显微解剖特征的前提. ...
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
... 另外, 森林火灾会导致部分形成层被烧死, 而形成特殊的“猫脸痕” (Smith & Sutherland, 1999); 洪水等特殊的气候事件会在木质部解剖结构中留下特殊痕迹, 即“洪灾轮”, 根据树轮解剖结构中“洪灾轮”的发生情况可重建过去洪水的发生历史(Arbe- llay et al., 2010; Wertz et al., 2013); 高海拔地区或北方林线, 寒秋或夏末温度的急剧变冷会造成年轮晚材细胞壁变薄, 形成所谓的“亮轮” (Wang et al., 2002); 生长季内的异常低温事件(温度降至0 ℃以下)会导致形成层分化出的未成熟木质部细胞无法完成次生壁加厚及木质化过程, 细胞脱水还会使未正常发育的细胞因挤压而变形, 在年轮细胞结构上出现明显的异常标志即“霜轮” (D’Arrigo et al., 2001).上述“痕迹”均能准确记录周围的环境信号, 并被用作年轮气候分析.此外, 未木质化或未完全木质化的木质部细胞会被固绿染成蓝色, 这些特殊的“蓝轮”可被用作分析木质化过程与气候要素的关系(Piermattei et al., 2014); 树轮木质部纤维素微纤丝角度(Xu et al., 2012)、射线薄壁细胞数量(Olano et al., 2013)等解剖参数, 也被证实能够准确记录周围的环境信号. ...
Cellulose microfibril angle variation in Picea crassifolia tree rings improves climate signals on the Tibetan Plateau. 2 2012
... 树轮具有定年准确、连续性强、分辨率高、易于获取和复本量大等优点, 其宽度、密度以及同位素含量等能够准确记录周围环境的变化(Fritts, 1976).全球变化背景下, 树轮是研究树木生长与气候变化关系的重要资料, 更是准确评估气候变化情境以及物种响应策略与适应性的关键(Fonti et al., 2010), 已被广泛用于全球变化以及古气候研究(Esper et al., 2002; Briffa et al., 2004; Moberg et al., 2005; Yang et al., 2014).树轮是众多环境因子在不同时段内综合作用的结果, 很难确定某一特定环境因子在特定时期内对树木生长的影响(de Luis et al., 2011).树轮气候学研究中常用表征年际变化的指标(如宽度、密度以及同位素含量等)与亚年际尺度(候、月或季节)的气候因子做响应分析或重建亚年尺度上的古气候; 但这种统计学上的年轮-气候“响应”关系, 常因缺乏内在机制或生理学解释而受质疑(Fritts, 1976).此外, 传统树轮气候学研究强调限制地点采样, 这些区域树木往往年龄较大且具有更强的气候代表性(Fritts, 1976; Esper et al., 2002; Briffa et al., 2004; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
... ; Yang et al., 2014); 但随着树轮研究工作的逐渐增多, 理想的树轮研究场所愈来愈少, 研究工作不得不在一些响应不太敏感的非限制区开展, 树轮研究遇到了前所未有的瓶颈(Liang et al., 2013).树轮中记载了大量的气候或环境信息, 传统树轮代用指标中记录的环境信号仅占一小部分, 如何充分挖掘树轮中记录的环境信号也是目前亟待解决的问题. ...
A 3,500-year tree-ring record of annual pre- cipitation on the northeastern Tibetan Plateau 1 2014
... 树轮解剖特征中也存在因年龄或竞争等因素而造成的变异(Matisons et al., 2012; Gričar et al., 2013; Peters et al., 2015; Venegas-González et al., 2015), 常需选择理想去趋势方法, 剔除解剖特征序列中的噪音(Fonti et al., 2010; Liang et al., 2013; Peters et al., 2015).针对木质部解剖特征序列的去趋势, 实践中往往直接运用传统方法去趋势(Fonti & García- González, 2004; Giantomasi et al., 2009; Matisons et al., 2012; Gričar et al., 2013; Venegas-González et al., 2015).针叶树管胞、阔叶树导管、IADFs、射线薄壁细胞数量、树脂道数量及微纤丝角解剖特征均可采用常规方法去趋势(Xu et al., 2012; Fonti et al., 2013; Olano et al., 2013; Novak et al., 2013; Campelo et al., 2015).树轮解剖学研究中常用的去趋势方法有线性回归(Yasue et al., 2000)、样条函数(Fonti & García-González, 2004; Campelo et al., 2010; Gričar et al., 2013)、自回归模型(Olano et al., 2012)或负指数函数(Fonti et al., 2013; Martin-Benito et al., 2013)等.去趋势方法不同, 去趋势效果往往也不一样(Carrer et al., 2015; Peters et al., 2015), 但无论采用哪种方法都会在剔除干扰信号的同时, 损失部分低频或高频的气候信号(Peters et al., 2015), 例如Liang等(2013)研究发现德国东北部欧洲赤松(Pinus sylvestris)早材内细胞腔的平均面积(原始值)与日平均最低温度显著正相关, 而利用样条函数剔除低频趋势后, 这种显著相关也随之消失.树种特性、去趋势目的、解剖特征类型以及木材性质等差异, 均可能影响去趋势方法的选择(Abrantes et al., 2013; Carrer et al., 2015; Peters et al., 2015), 故选用去趋势函数对木质部解剖特征序列进行去趋势与标准化时应谨慎(Carrer et al., 2015; Peters et al., 2015).另外, 部分研究发现树轮解剖特征所呈现的年龄趋势仅表现在树木生长的幼龄阶段(10-30年)(Gea- Izquierdo et al., 2012; Liang et al., 2013; Pritzkow et al., 2014; Carrer et al., 2015).实际研究中尽可能多地保留气候信号远比略微延长序列长度有意义, 故可将包含幼龄趋势的部分区段直接删除后再用于树轮气候学分析, 巧妙地避免去趋势带来的弊端(Gea-Izquierdo et al., 2012; Gričar et al., 2013; Liang et al., 2013; Venegas-González et al., 2015). ...
The effects of tracheid dimensions on variations in maximum density of Picea glehnii and relationships to climatic factors. 4 2000
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... ).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 导管是双子叶植物木质部轴向输送水分的最主要结构, 其相关特征与水分密切相关(Zimrnennann, 1983; Fonti et al., 2010).许多研究表明导管特征能够准确记录周围的水分信号, 早在1994年February就曾用南非考古遗址木炭年轮内导管直径重建了过去的降水变化(February, 1994).外界(土壤)水分的可利用性也能直接或间接影响导管特征, 调整水分策略以适应外界环境(Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).一般来说, 干旱胁迫(降水不足或高温)环境下, 木质部的导管往往会尺寸变小、变量减少(Eilmann et al., 2011; Vaganov et al., 2011); 而水分供应充足时, 木质部形成的导管往往面积更大、数量更多(Tyree & Sperry, 1989; Vaganov et al., 2011).干旱地区植物导管的大小或数量均低于湿润地区(Rita et al., 2015), 如Campelo等(2010)研究发现Quercus ilex导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
... 导管面积与多月降水量极显著正相关, 与温度相关性不显著, 从而指出导管面积主要受生长季水分可利用性控制, 冬季冻结事件和夏季干旱可能会导致较小的导管产生, 以应对空穴化和栓塞.导管直径与其面临栓塞与空穴化风险成正比(Zimrnennann, 1983; Sperry et al., 2006).在干旱期, 较大栓塞与空穴化风险可能危及植物的输水系统, 严重影响植物生存(Tyree & Sperry, 1989); 小而少的导管可降低水分输导效率, 确保其水力安全(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006), 如干旱条件下, Quercus canariensis通过减少早材内导管数量和直径(降低水力导度), 保存土壤水分且避免极端负水势的发生, 以适应夏季干旱的地中海气候(Gea-Izquierdo et al., 2012).干旱生境多强调水力安全性, 而湿润生境多强调水力效率性(Giantomasi et al., 2009; Venegas- González et al., 2015).植物还能够通过增加导管的密度或改变导管的数量与分布等结构来提高植物的导水率, 以规避空穴化的风险(Hacke et al., 2006; Loepfe et al., 2007).管腔密度增加也是提高水力效率的重要途径, 它可适当规避植物空穴化的风险, 但会增加空穴化或栓塞的蔓延(Carlquist & Hoekman, 1985).许多研究表明导管密度与水分关系密切且能够记录显著的水分信号(Corcuera et al., 2004; Schume et al., 2004).导管大小常与密度负相关(Abrantes et al., 2013; Rita et al., 2015), 导管变小的同时, 常伴随着密度的提高(Hacke et al., 2006; Loepfe et al., 2007), 例如干旱区Ilex aquifolium导管大小与各季节降水显著正相关, 而导管密度与各季节降水显著负相关(Rita et al., 2015).也有研究发现导管密度并不能像导管面积那样准确记录外界环境信号(Sass & Eckstein, 1995; Campelo et al., 2010; Leal et al., 2011), 如半环孔材Quercus suber和散孔材Fagus sylvatica (Sass & Eckstein, 1995; Leal et al., 2011)的导管密度几乎恒定. ...
Xylem Structure and the Ascent of Sap 2 1983
... 维管植物木质部的主要功能是输导水分与支撑植物体(Zimrnennann, 1983).本文仅以木质部水分输导组织为例, 阐述基于管孔特征对环境变化的响应机理.长时间尺度上, 植物为适应外界的环境变化, 不断改变其结构和功能即进化.木质部水分输导系统经长期进化(适应及响应), 主要形成了无孔材、环孔材、散孔材和半环孔材4类树种(图2), 这是水分输导组织响应环境变化的典型代表(Zimrn- ennann, 1983).材性差异也决定了树体对外界环境(尤其是水分)响应或适应策略差异.如环孔材树干水分传输和存贮能力明显高于散孔材, 而其却更易遭受栓塞的威胁(Tyree & Zimmermann, 2013).短时间尺度上, 水分输导组织特征(如排列、大小、数量以及细胞壁厚度等)也会随外界环境变化而呈现出年内或年际差异(Wimmer, 2002; Giantomasi et al., 2009; Venegas-González et al., 2015).管腔直径能够直接影响木质部的导水能力, 根据哈根-泊肃叶定律(Hagen-Poiseuille’s law), 管腔半径增加相应输水效率呈4次方增加, 因此管腔直径略微下降或增加将导致水力效率大幅减少或增加(Zimrnennann, 1983).土壤水分可利用性能够在年际或季节水平上影响木材的形成(Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011; Wertz et al., 2013).水分胁迫(降水不足或高温)能通过多种途径直接或间接地导致导管或管胞细胞变小或数量减小(Eilmann et al., 2011; Vaganov et al., 2011), 如影响木质部同化物的积累、控制细胞的分裂、扩大和分化等(图3) (Battipaglia et al., 2010; Gruber et al., 2010).相反, 充足的水分供应既能够直接提高细胞膨压及细胞扩张速率导致形成较大的管腔(Tyree & Sperry, 1989; Vaganov et al., 2011), 又能间接地通过降低生长素浓度来减缓细胞分化或延长树木生长周期, 允许更多及更大的导管或管胞细胞形成(Kozlowski & Pallardy, 1997; Aloni, 2001; Vaganov et al., 2011).值得注意的是, 较大导管或管胞直径在增加水力效率与气孔导度、促进光合碳增益的同时, 也使植物面临更大的栓塞与空穴化风险(Tyree & Sperry, 1989; Mencuccini, 2003; Sperry et al., 2006).水分往往是干旱条件下促进树木生长的关键, 而湿润条件下过多的水分也会抑制树木生长.例如湿润条件下生长的松树形成的年轮较窄且晚材比例更低, 抑制时期每个年轮仅由1-2列早材细胞及1列较小且细胞壁较薄的晚材细胞构成(Vaganov et al., 2011).因此, 在全球变化背景下, 依据树轮木质部解剖特征对环境(气候)变化的塑性响应特性开展树轮解剖学研究, 是研究植被与全球变化关系的重要手段(Wimmer, 2002; Stoffel & Bollschweiler, 2008; Fonti et al., 2010). ...
... 管胞是裸子植物木质部最主要的组成部分, 是运输水分和矿物质的主要通道(表1).许多研究表明管胞特征与水分条件关系密切(Fonti et al., 2010; Gea-Izquierdo et al., 2012), 如Martin-Benito等(2013)发现前一年秋季和当年春、夏季的降水对地中海森林中欧洲黑松(Pinus nigra)和欧洲赤松径向生长十分有利, 尤其是对早材管胞直径增大及细胞壁加厚至关重要; 当年夏季水分利用率较低会导致欧洲赤松管胞数量减少、腔径增加及细胞壁变薄, 晚材比例降低; 当年夏季的降水量及标准化降水蒸发指数(standardized precipitation evapotranspiration index, SPEI)与黑松的管胞数量及壁厚正相关, 而当年秋季的降水量及SPEI与黑松的管胞数量及壁厚负相关.土壤水分既能直接影响木质部细胞的分裂、扩大以及分化, 还会间接影响树木生长期长短, 导致针叶树管胞特征发生改变, 最终影响年轮宽窄(图3)(Battipaglia et al., 2010; Gruber et al., 2010; Eilmann et al., 2011; Vaganov et al., 2011) , 如在不利的水分条件下, 欧洲赤松细胞伸长与细胞壁加厚阶段持续时间明显缩短, 木质部细胞活动提早结束约4周, 致使窄轮发生(Gruber et al., 2010); 而在灌溉条件下, 欧洲赤松早晚材转变时间比干旱条件下推迟2-4周, 细胞停止活动推迟约5周, 径向增量明显高于干旱条件下的欧洲赤松(Eilmann et al., 2011).针叶树管胞与阔叶树导管对水分环境响应策略一样, 也遵循水力效率性和安全性的权衡(Zimrnen- nann, 1983; Sperry et al., 2006). ...

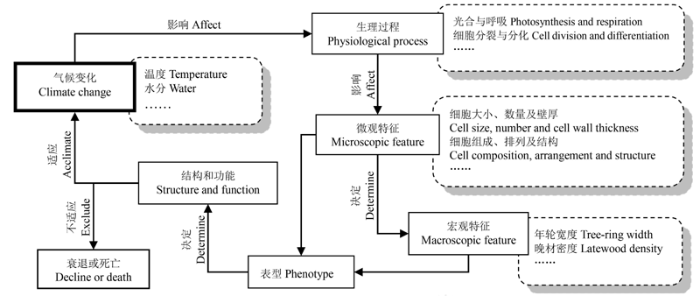 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT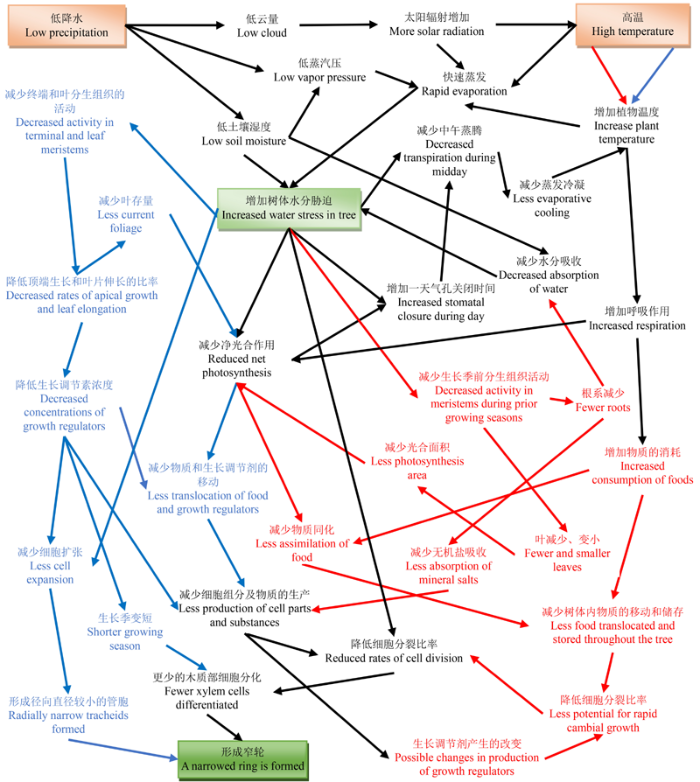 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}