Distribution of biomass in relation to environments in shrublands of temperate China
YANGXian1, GUOYan-Pei1, MOHHAMOTAnwar2, LIUHong-Yan1, MAWen-Hong3, YUShun-Li4, TANGZhi-Yao1,*, 1College of Urban and Environmental Science, Peking University, Beijing 100871, China2Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Ürümqi 830011, China;3College of Life Sciences, Inner Mongolia University, Hohhot 010021, China4State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 通讯作者:* 通信作者Author for correspondence (E-mail: zytang@urban.pku.edu.cn) 责任编辑:YANGXianGUOYan-PeiMOHHAMOTAnwarLIUHong-YanMAWen-HongYUShun-LiTANGZhi-Yao 收稿日期:2016-06-13 接受日期:2016-06-23 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略先导性科技专项(XDA05050301和XDA05050700)和科技部科技基础性工作专项(2011FY110300)
关键词:灌丛;生物量密度;地下-地上生物量比;叶-枝生物量比 Abstract Aims Shrubland is one of the most widely distributed vegetation types in northern China. Previous studies on pattern and dynamics of plant biomass have been focused on forest and grassland ecosystems, while relevant knowledge on shrubland ecosystems is lacking. It is important to include shrublands in northern China to improve the accuracy in estimating the terrestrial ecosystem biomass in China. Methods Based on investigations and samplings from 433 shrubland sites, we explored the distribution and allocation patterns of biomass in relation to climatic and soil nutrient factors of shrublands of temperate China. Important findings The average shrubland biomass density in northern China is 12.5 t·hm-2. It decreases significantly from temperate deciduous shrubland in northeast to desert shrubland in northwest. The average biomass density of temperate deciduous shrubland, alpine shrubland, and desert shrubland is 14.4, 28.8, and 5.0 t·hm-2, respectively. Within temperate deciduous shrublands, plant biomass is lower in North China than in Northeast China. The average aboveground and belowground biomass density of shrub layer is 4.5 and 5.4 t·hm-2, respectively; while that of grass layer is 0.8 and 1.8 t·hm-2, respectively. Environmental factors affect biomass allocation across different plant organs. The belowground-aboveground biomass ratio of shrub exhibits no significant changes with environmental variables. The leaf-stem ratio increases with annual precipitation, and leaf biomass is low in arid region.
Keywords:shrubland biome;biomass density;belowground-aboveground biomass ratio;leaf-stem biomass ratio -->0 PDF (739KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 杨弦, 郭焱培, 安尼瓦尔·买买提, 刘鸿雁, 马文红, 于顺利, 唐志尧. 中国北方温带灌丛生物量的分布及其与环境的关系. 植物生态学报, 2017, 41(1): 22-30 https://doi.org/10.17521/cjpe.2016.0199 YANGXian, GUOYan-Pei, MOHHAMOTAnwar, LIUHong-Yan, MAWen-Hong, YUShun-Li, TANGZhi-Yao. Distribution of biomass in relation to environments in shrublands of temperate China. Chinese Journal of Plant Ecology, 2017, 41(1): 22-30 https://doi.org/10.17521/cjpe.2016.0199 灌丛生态系统是我国重要的植被碳库(胡会峰等, 2006)。研究灌丛的生物量分布格局和分配是对我国估算整个陆地生态系统生物量的重要补充。然而, 由于研究数据的缺乏, 开展大尺度的灌丛生物量研究面临着巨大的困难。相对于森林和草地的生物量研究来说, 对我国灌丛的相应研究较少。国内对灌丛生物量的研究开始于20世纪80年代(姜凤岐和卢凤勇, 1982), 后来的一系列研究估算了不同类型灌丛的生物量密度(贺金生等, 1997; 罗天祥等, 1999; 陈遐林等, 2002; 刘国华等, 2003a, 2003b)。现有的灌丛生物量研究往往着眼于特定地区或特定类型的灌丛, 难以从区域尺度上反映灌丛生物量的分布格局。 植物为了适应不同的环境条件, 可能将资源分配到不同的器官, 植物各器官的生物量比值, 如地下-地上生物量比或根冠比反映了植物生物量在不同器官中的分配策略。根据植物生长的类经济学模型及平衡生长假说, 植物将资源分配到各个器官以使得生物量生产最大化, 当某一资源最为限制植物生长时, 植物将倾向于把更多生物量分配给获取该资源的器官(Bloom et al., 1985; Chapin et al., 1987)。例如在干旱或养分贫瘠的土壤中, 植物会增加根系的生长来加强对水和氮磷的吸收, 从而导致更高的地下-地上生物量比(Subbarao et al., 1995; Turner & Haygarth, 2001)。随着植物密度的增加, 植物对光的竞争增强会使得叶生物量减少而茎生物量增加, 即叶-枝比减小, 这可能是因为茎的生长可以使得叶处于群落上层, 从而获得更多的光(Poorter et al., 2012)。Markesteijn和Poorter (2009)发现, 降水对热带森林生态系统地下-地上生物量比具有重要的影响, 一般而言, 干旱地区的森林具有较大的地下生物量, 而湿润地区的森林地下生物量较小, 因此其地下-地上生物量比随着降水增加而减小。王娓等(2008)认为中国北方草地地下-地上生物量比随温度变化不明显, 随降水增加而降低; 但Yang等(2010)认为中国草地的地下-地上生物量比与温度降水都没有显著关系。与之相比, 对灌丛生物量分配的研究依旧主要关注特定地区和特定类型的灌丛(王勇军等, 2010; 雷蕾等, 2011; 高巧等, 2014; 钟泽兵等, 2014), 缺乏大尺度研究。 灌丛广泛分布于我国北方地区, 占据了大量森林和草地无法发育的生境。研究北方灌丛的生物量分布格局和分配策略能够加深对环境胁迫下的植物群落状况的认识。中国北方灌丛中的灌木植物基本为落叶灌木, 将其作为研究对象能排除常绿和落叶生活型差异造成的误差; 另一方面, 干旱是中国北方植物生长的主要限制因子(Bai et al., 2008), 将中国北方作为研究区域有助于深入理解水分对灌丛植被生物量极其分配策略的影响。因此, 我们以中国北方所有的灌丛作为研究对象, 估算了灌丛生物量在中国北方的分布格局以及气候和土壤因子对灌丛生物量分配的影响。
中国北方灌丛植被平均生物量为12.5 t·hm-2, 其中灌木层地上、地下平均生物量分别为4.5 (±0.3)和5.4 (±0.4) t·hm-2, 草本层地上、地下平均生物量分别为0.8 (±0.0)和1.8 (±0.1) t·hm-2。不同区域生物量变异较大, 东部温带落叶灌丛生物量高于西部的荒漠灌丛, 东部温带落叶灌丛生物量一般高于10 t·hm-2, 而西部荒漠区低于5.0 t·hm-2 (图2)。其中温带落叶灌丛、亚高山落叶阔叶灌丛、荒漠灌丛3种植被型平均生物量分别为14.4、28.8和5.0 t·hm-2, 其中平均地上生物量分别为6.0、13.0和2.5 t·hm-2; 平均生物量分别为8.4、15.8和2.5 t·hm-2。3种灌丛类型平均凋落物量分别为2.6 (±0.2)、2.2 (±0.5)和1.4(±0.2) t·hm-2 (表1)。 Table 1 表1 表1中国北方主要灌丛植被型的生物量密度 Table 1Biomass density of major shrubland types of Northern China
灌丛类型 Shrubland type
温带落叶灌丛 Temperate deciduous shrublands (mean ± SE)
亚高山落叶灌丛 Alpine shrublands (mean ± SE)
荒漠灌丛 Desert shrublands (mean ± SE)
所有 Overall (mean ± SE)
样地数 No. of study site
321
4
108
433
生物量 Biomass (t·hm-2)
14.4
28.8
5.0
12.5
地上生物量 Aboveground biomass
6.0
13.0
2.5
5.3
灌木层 Shrub layer
5.2 ± 0.3
12.2 ± 4.3
2.0 ± 0.3
4.5 ± 0.3
草本层 Herb layer
0.8 ± 0.0
0.8 ± 0.2
0.4 ± 0.1
0.8 ± 0.0
地下生物量 Belowground biomass
8.4
15.8
2.5
7.2
灌木层 Shrub layer
6.5 ± 0.4
13.2 ± 6.4
2.0 ± 0.5
5.4 ± 0.4
草本层 Herb layer
1.9 ± 0.1
2.6 ± 0.3
0.5 ± 0.1
1.8 ± 0.1
凋落物量 Litterfall (t·hm-2)
2.6 ± 0.2
2.2 ± 0.5
1.4 ± 0.2
2.5 ± 0.2
SE, standard error.SE, 标准误差。 新窗口打开 显示原图|下载原图ZIP|生成PPT 图2中国北方温带灌丛植被总生物量空间分布格局。 -->Fig. 2Spatial distribution of vegetation biomass in shrubland of the northern China. -->
灌木层和草本层植物地下生物量(图3B、3D、3F)均高于地上生物量(图3A、3C、3E)。东部温带落叶灌丛灌木层植物地上(>5.2 vs. <2.0 t·hm-2)和地下生物量(>6.5 vs. <2.0 t·hm-2)均略高于西部荒漠区(图3A、3B)。草本层生物量在东西部差异较大, 草本层地上和地下生物量都小于0.2 t·hm-2的灌丛大部分出现在西部荒漠地区, 而草本层地上生物量超过0.9 t·hm-2、地下生物量超过1.0 t·hm-2的灌丛大部分出现在东部(图3C、3D)。草本层生物量远低于灌木植物, 但草本层在空间分布上的差异加大了东西部地上、地下总生物量的差异(图3E、3F)。 显示原图|下载原图ZIP|生成PPT 图3中国北方灌丛不同组分生物量分布。A, 灌木层地上生物量。B, 灌木层地下生物量。C, 草本层地上生物量。D, 草本层地下生物量。E, 总地上生物量。F, 总地下生物量。 -->Fig. 3Spatial distributions of above and belowground biomass in shrubland of the northern China. A, Aboveground biomass of shrub layer. B, Belowground biomass of shrub layer. C, Aboveground biomass of herb layer. D, Belowground biomass of herb layer. E, Total aboveground biomass. F, Total belowground biomass. -->
2.2 中国北方灌丛地下-地上及叶-枝生物量分配格局
由于亚高山落叶阔叶灌丛调查样地数量少, 因此只对温带落叶灌丛和荒漠灌丛灌木的地下-地上生物量比及叶-枝生物量比进行了分析。温带落叶灌丛灌木的地下-地上和叶-枝生物量比变异比荒漠灌丛大。温带落叶灌丛灌木的平均地下-地上生物量比为1.33 (±0.06), 平均叶-枝生物量比为0.47 (±0.02), 荒漠灌丛的平均地下-地上生物量比为1.22 (±0.07), 平均叶-枝生物量比为0.47 (±0.04)(图4)。 显示原图|下载原图ZIP|生成PPT 图4中国北方温带落叶灌丛和荒漠灌丛灌木地下-地上生物量比和叶-枝生物量比频度分布, A, 温带落叶灌丛地下-地上生物量比。B, 温带落叶灌丛叶-枝生物量比。C, 荒漠灌丛地下-地上生物量比。D, 荒漠灌丛叶-枝生物量比。 -->Fig. 4Frequency distributions of belowground:aboveground biomass ratio (BGB:AGB) and leaf:stem biomass ratio of shrubs in the temperate deciduous shrubland and dessert shrubland of the northern China. A, Belowground:aboveground biomass ratio in the temperate deciduous shrubland. B, Leaf:stem biomass ratio in the temperate deciduous shrubland. C, Belowground: aboveground biomass ratio in the dessert shrubland. D, Leaf:stem biomass ratio in the dessert shrubland. -->
中国北方灌丛灌木地上-地下生物量比随年平均气温、年降水量增加而显著减少(年平均气温: p < 0.001, 年降水量: p = 0.002), 但随土壤氮和磷含量的变化无显著变化规律(p > 0.05)。灌木层叶-枝生物量比随年平均气温和土壤磷增加而显著降低(p < 0.001), 但随年降水的增加趋势较弱(p = 0.024), 随土壤磷无显著的变化趋势(p > 0.05)(图5)。 显示原图|下载原图ZIP|生成PPT 图5中国北方灌丛地下-地上生物量比(A-D)和叶-枝生物量比(E-H)与环境的关系。点线: p > 0.01; 短划线: 0.001< p < 0.01; 实线: p < 0.001。 -->Fig. 5Distribution of belowground:aboveground biomass ratio (A-D) and leaf:stem biomass ratio (E-H) in shrubland of the northern China in relation to environmental variables. Fitted lines show significant relationships (dotted line: p > 0.01; dashed line: 0.001< p < 0.01; solid line: p < 0.001). AGB, aboveground biomass; BGB, belowground biomass. MAT, mean annual temperature; AP, annual precipitation; STN, soil total nitrogen concentration; STP, soil total phosphorus concentration. -->
BaiY, WuJ, XingQ, PanQ, HuangJ, YangD, HanX (2008). Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau .Ecology, 89, 2140-2153. [本文引用: 1]
[2]
BloomAJ, ChapinFS, MooneyHA (1985). Resource limitation in plants—An economic analogy . Annual review of Ecology and Systematics, 363-392. [本文引用: 2]
ChenXL, MaQY, KangFF, CaoWQ, ZhangGH, ChenZW (2002). Studies on the biomass and productivity of typical shrubs in Taiyue Mountain, Shanxi Province .Forest Research, 15, 304-309. (in Chinese with English abstract)[陈遐林, 马钦彦, 康峰峰, 曹文强, 张国华, 陈宗伟 (2002). 山西太岳山典型灌木林生物量及生产力研究 . 林业科学研究, 15, 304-309.] [本文引用: 1]
[5]
GaoQ, YangXC, YinCY, LiuQ (2014). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province, China .Chinese Journal of Plant Ecology, 38, 355-365. (in Chinese with English abstract)[高巧, 阳小成, 尹春英, 刘庆 (2014). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算 . 植物生态学报, 38, 355-365.] [本文引用: 1]
[6]
HeJS, WangQB, HuD (1997). Studies on the biomass of typical shrubland and their regeneration capacity after cutting .Acta Phytoecologica Sinica, 21, 512-520. (in Chinese with English abstract)[贺金生, 王其兵, 胡东 (1997). 长江三峡地区典型灌丛的生物量及其再生能力 . 植物生态学报, 21, 512-520.] [本文引用: 1]
[7]
HijmansRJ, CameronSE, ParraJL, JonesPG, JarvisA (2005). Very high resolution interpolated climate surfaces for global land areas .International Journal of Climatology, 25, 1965-1978. [本文引用: 1]
[8]
HuHF, WangZH, LiuGH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Ecology (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 1]
[9]
JiangFQ, LuFY (1982). The model of estimating above-ground biomass of Caragana microphylla brush . Acta Ecologica Sinica, 2, 103-110. (in Chinese with English abstract)[姜凤岐, 卢凤勇 (1982). 小叶锦鸡儿灌丛地上生物量的预测模式 . 生态学报, 2, 103-110.] [本文引用: 1]
[10]
LeiL, LiuXD, WangSL, LiY, ZhangXL (2011). Assignment rule of alpine shrubs biomass and its relationships to environmental factors in Qilian Mountains .Ecology and Environmental Sciences, 20, 1602-1607. (in Chinese with English abstract)[雷蕾, 刘贤德, 王顺利, 李毅, 张学龙 (2011). 祁连山高山灌丛生物量分配规律及其与环境因子的关系 . 生态环境学报, 20, 1602-1607.] [本文引用: 1]
[11]
LiuGH, ZhangJY, ZhangYX, ZhouJY, GuanWB, MaKM, FuBJ (2003a). Distribution regulation of aboveground biomass of three main shrub types in the dry valley of Minjiang River .Journal of Mountain Science, 21, 24-32. (in Chinese with English abstract)[刘国华, 张洁瑜, 张育新, 周建云, 关文彬, 马克明, 傅伯杰 (2003a). 岷江干旱河谷三种主要灌丛地上生物量的分布规律 . 山地学报, 21, 24-32.] [本文引用: 1]
[12]
LiuGH, MaKM, FuBJ, GuanWB, KangYX, ZhouJY, LiuSL (2003b). Aboveground biomass of main shrubs in dry valley of Minjiang River .Acta Ecologica Sinica, 23, 1757-1764. (in Chinese with English abstract)[刘国华, 马克明, 傅伯杰, 关文彬, 康永祥, 周建云, 刘世梁 (2003b). 岷江干旱河谷主要灌丛类型地上生物量研究 . 生态学报, 23, 1757-1764.] [本文引用: 1]
[13]
LuoTX, LiWH, LuoJ, WangQJ (1999). A comparative study on biological production of major vegetation types on the Tibetan Plateau .Acta Ecologica Sinica, 19, 823-831. (in Chinese with English abstract)[罗天祥, 李文华, 罗辑, 王启基 (1999). 青藏高原主要植被类型生物生产量的比较研究 . 生态学报, 19, 823-831.] [本文引用: 1]
[14]
MarkesteijnL, PoorterL (2009). Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance .Journal of Ecology, 97, 311-325. [本文引用: 1]
[15]
McCarthyMC, EnquistBJ (2007). Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation .Functional Ecology, 21, 713-720.
[16]
PoorterH, NiklasKJ, ReichPB, OleksynJ, PootP, MommerL (2012). Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control .New Phytologist, 193, 30-50. [本文引用: 1]
[17]
R CoreTeam (2015). R: A language and environment for statistical computing . Vienna, Austria. . Cited by 2015-12-10.URL [本文引用: 1]
[18]
SubbaraoGV, JohansenC, SlinkardAE, Nageswara RaoRC, SaxenaNP, ChauhanYS, LawnRJ (1995). Strategies for improving drought resistance in grain legumes . Critical Reviews in Plant Sciences, 14, 469-523. [本文引用: 1]
[19]
The Editorial Committee of Vegetation Map of China, Chinese Academy of Sciences (2007). Vegetation Map of the People’s Republic of China 1:1000000. Geological Publishing House, Beijing. (in Chinese)[中国科学院中国植被图编辑委员会 (2007). 中华人民共和国植被图1:1000000. 地质出版社, 北京.] [本文引用: 2]
WangW, PengSS, FangJY (2008). Biomass distribution of natural grasslands and it response to climate change in north China .Arid Zone Research, 25, 90-97. (in Chinese with English abstract)[王娓, 彭书时, 方精云 (2008). 中国北方天然草地的生物量分配及其对气候的响应 . 干旱区研究, 25, 90-97.]
[22]
WangYJ, HuangCD, ZhangJ, YangWQ, WangXS (2010). Species diversity biomass and their relationship of shrubberies in an arid valley of the Minjiang River .Arid Zone Research, 27, 567-572. (in Chinese with English abstract)[王勇军, 黄从德, 张健, 杨万勤, 王宪帅 (2010). 岷江干旱河谷灌丛物种多样性, 生物量及其关系 . 干旱区研究, 27, 567-572.] [本文引用: 1]
[23]
YangY, FangJ, JiC, HanW (2009). Above- and belowground biomass allocation in Tibetan grasslands .Journal of Vegetation Science, 20, 177-184.
YangX, TangZ, JiC, LiuH, MaW, MohhamotA, ShiZ, SunW, WangT, WangX (2014). Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across Northern China .Scientific Reports, 4, 5448. [本文引用: 1]
[26]
ZhongZB, YangLC, LiuHC, SongWZ, LiF, ZhouGY (2014). The main shrubs aboveground biomass and effect factors in Yushu, Qinghai, China .Mountain Research, 32, 678-684. (in Chinese with English abstract)[钟泽兵, 杨路存, 刘何春, 宋文珠, 李璠, 周国英 (2014). 青海玉树地区主要灌丛类型地上生物量及其影响因素 . 山地学报, 32, 678-684.] [本文引用: 1]
Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau 1 2008

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT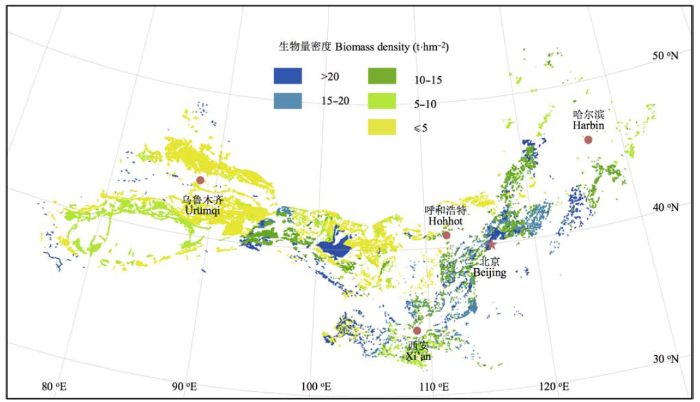 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT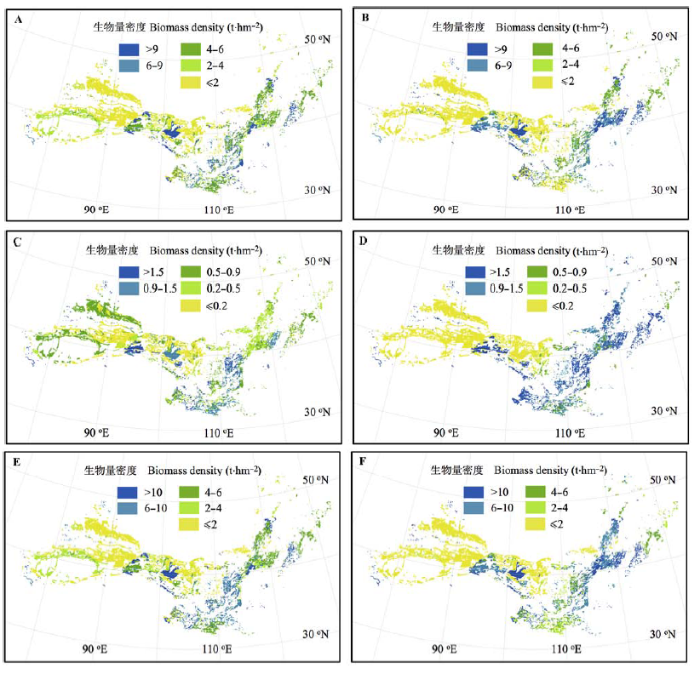 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT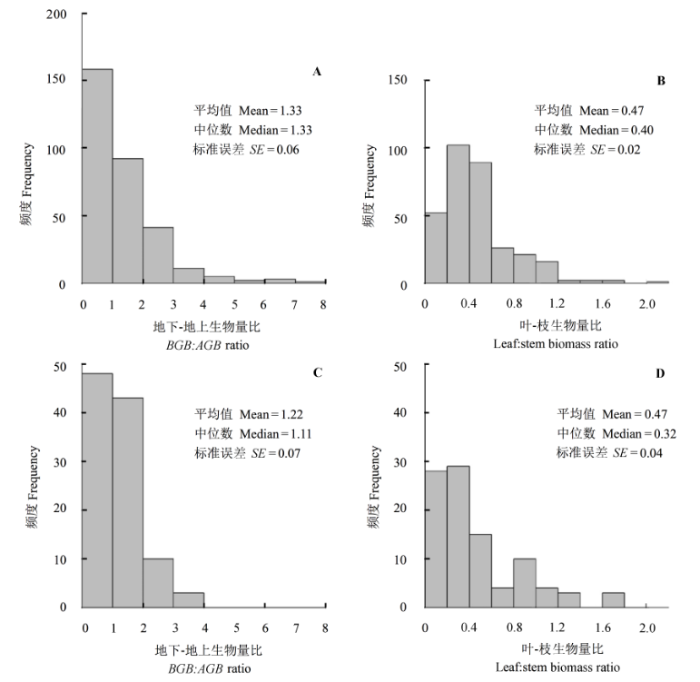 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT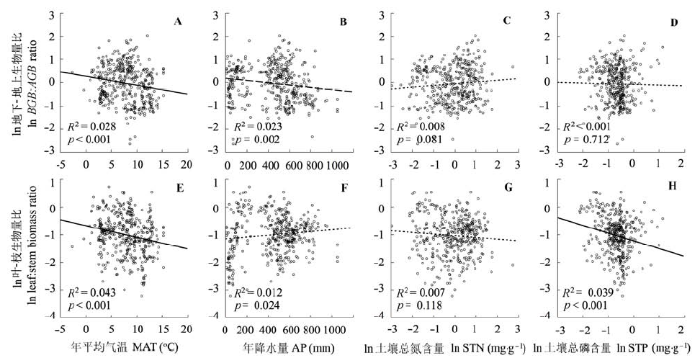 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}