Estimation of biomass allocation and carbon density of Rhododendron simsii shrubland in the subtropical mountainous areas of China
ZHANGQiang1,2, LIJia-Xiang3, XUWen-Ting1, XIONGGao-Ming1, XIEZong-Qiang1,*, 1State Key Laboratory of Vegetation and Environment Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2University of Chi- nese Academy of Sciences, Beijing 100049, China3Faculty of Forestry, Central South University of Forestry and Technology, Changsha 410004, China 通讯作者:* 通信作者Author for correspondence (E-mail: xie@ibcas.ac.cn) 责任编辑:ZHANGQiangLIJia-XiangXUWen-TingXIONGGao-MingXIEZong-Qiang 收稿日期:2016-05-17 接受日期:2016-09-21 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略性先导科技专项 (XDA05050302)和国家科技基础性专项(Y5220B1001)
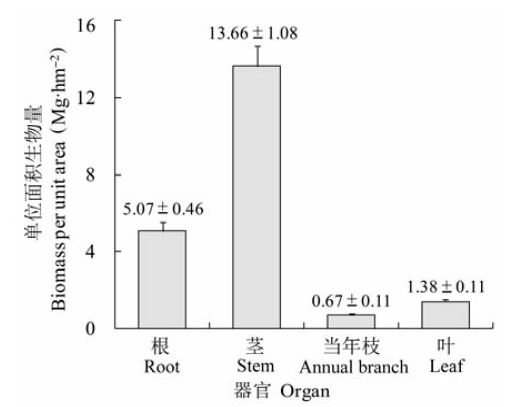
关键词:回归模型;根冠比;养分归还;地上生物量;地下生物量;含碳率 Abstract Aims As an important potential carbon sink, shrubland ecosystem plays a vital role in global carbon balance and climate regulation. Our objectives were to derive appropriate regression models for shrub biomass estimation, and to reveal the biomass allocation pattern and carbon density in Rhododendron simsii shrubland. Methods We conducted investigations in 27 plots, and developed biomass regression models for shrub species to estimate shrub biomass. The biomass of herb and litterfall were obtained through harvesting. Plant samples were collected from each plot to measure carbon content in different organs. Important findings The results showed that the power and linear models were the most appropriate equation forms. The D and D2H (where D was the basal diameter (cm) and H was the shrub height (m)) were good predictors for organ biomass and total biomass of shrubs. All of the biomass models reached extremely significant level, and could be used to estimate shrub biomass with high accuracy. It was more difficult to predict leaf and annual branch biomass than stem biomass, because leaf and annual branch were susceptible to herbivores and inter-plant competition. The mean biomass of the shrub layer was 20.78 Mg·hm-2, in which Rhododendron simsii and Symplocos paniculata biomass accounted for 93.63%. Influenced by both environment and species characteristics, the biomass of the shrub layer organs was in the order of stem > root > leaf > annual branch. The root:shoot ratio of the shrub layer was 0.32, which was less than other shrubs in subtropical regions. The relative higher aboveground biomass allocation reflected the adaptation of plants to the warm and humid environment for more photosynthesis. The mean total community biomass was 26.26 Mg·hm-2, in which shrub layer, herb layer and litter layer accounted for 79.14%, 7.62% and 13.25%, respectively. Litter biomass was relatively high, which suggested that this community had high nutrient return. There were significant correlations among aboveground biomass, belowground biomass and total biomass of shrub layer and herb layer. The mean biomass carbon density of the community was 11.70 Mg·hm-2 and the carbon content ratio was 44.55%. The carbon density was usually obtained using the conversion coefficient of 0.5 in previous studies, which could overestimate carbon density by 12.22%.
Keywords:regression model;root/shoot ratio;nutrient return;aboveground biomass;belowground biomass;carbon content ratio -->0 PDF (467KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 张蔷, 李家湘, 徐文婷, 熊高明, 谢宗强. 中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算. 植物生态学报, 2017, 41(1): 43-52 https://doi.org/10.17521/cjpe.2016.0174 ZHANGQiang, LIJia-Xiang, XUWen-Ting, XIONGGao-Ming, XIEZong-Qiang. Estimation of biomass allocation and carbon density of Rhododendron simsii shrubland in the subtropical mountainous areas of China. Chinese Journal of Plant Ecology, 2017, 41(1): 43-52 https://doi.org/10.17521/cjpe.2016.0174 灌丛生态系统作为一种分布广泛的陆地生态系统类型, 在全球碳循环和气候调节中起着重要的作用(方精云等, 2007)。在全球变暖的影响下, 灌丛的分布范围有所扩大(Sturm et al., 2001), 并引起了北半球局部区域碳储量的变化(Goodale & Davidson, 2002)。我国灌丛分布面积69.2 × 104 km2, 占国土面积的7.3% (中华人民共和国环保部和中国科学院, 2015), 是一个重要的潜在碳汇, 其年均碳汇占中国植被年均总碳汇的14.6%-22.6% (方精云等, 2007)。但目前对灌丛生态系统生产力和碳汇的研究相对缺乏, 尤其缺少地下根系部分和凋落物的研究(Vourlitis et al., 2007), 导致对灌丛碳储量的估算存在极大的不确定性。 生物量是量化生态系统碳循环和植被碳储量的重要指标, 不仅反映了生态系统生产力水平, 而且反映了生态系统功能的强弱(方精云和陈安平, 2001), 对其进行测定是评估生态系统碳汇功能的重要前提。生物量在植物中的分配策略是植物对环境长期适应的结果, 受到外界环境、物种组成及植株大小等因素的影响(Poorter et al., 2012), 对植物的生长、发育和繁殖有巨大的作用(Cairns et al., 1997)。由于不同层片和不同植物器官中碳含量的差异, 研究生物量的分配规律对提高生态系统碳储量的评估精度具有重要价值。 中国灌丛约有一半分布在亚热带区域, 已有的对其生物量的报道主要集中在地上部分(张光富和宋永昌, 2001; 胡会峰等, 2006; 李轩然等, 2006), 对于地下部分生物量和地上、地下生物量相关关系研究较少。对灌丛碳密度的估算多采用转换因子法(胡会峰等, 2006), 缺乏实际测量, 导致估算结果与实际值相差较大。杜鹃(Rhododendron simsii)作为分布广泛的物种, 在我国亚热带大部分地区均有生长, 以其为优势的群落是亚热带最为常见的山地灌丛类型(吴征镒, 1980)。本文以中国中亚热带山地杜鹃灌丛为对象, 通过生物量回归模型来研究其群落生物量分配规律和碳密度特征, 以期为估算灌丛碳储量提供依据。
表2反映了研究区域杜鹃灌丛群落灌木层个体数量和生物量的种类分配情况。其中, 杜鹃的个体数量占整个灌木层个体数量的87.54%, 其生物量占灌木层生物量的79.61%; 湖南白檀的个体数量占灌木层的6.99%, 其生物量占灌木层的14.02%; 剩余的17种灌木物种的个体数量占灌木层的5.48%, 其生物量占灌木层的6.37%。优势物种杜鹃和湖南白檀的个体数量及其生物量均占到本研究群落灌木层的近95%。 Table 2 表2 表2杜鹃灌丛灌木层个体密度和生物量的种类分配 Table 2Individual density and biomass allocation of shrub layer in different species
对27个样方的灌木层和草本层的地上生物量与地下生物量和总生物量进行相关分析, 结果如图3所示。可以看出, 灌木层和草本层的地上生物量与地下生物量和总生物量之间都存在极显著的相关关系(p < 0.001), 决定系数R2平均达到0.912。说明灌木层和草本层生物量在地上、地下分配上具有相对稳定的分配规律, 这种相关关系可用于由已知的地上生物量来推算地下生物量和总生物量。 显示原图|下载原图ZIP|生成PPT 图3杜鹃灌丛灌木层和草本层地上/地下生物量之间的相关关系。 -->Fig. 3The relationship between above- and belowground biomass of the shrub layer and the herb layer in Rhododendron shrubland. -->
AraújoTM, HiguchiN, de Carvalho JúniorJADC (1999). Comparison of formulae for biomass content determina- tion in a tropical rain forest site in the state of Pará, Brazil .Forest Ecology and Management, 117, 43-52. [本文引用: 1]
[2]
BasukiT, van LaakeP, SkidmoreA, HussinY (2009). Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests .Forest Ecology and Management, 257, 1684-1694. [本文引用: 1]
[3]
BloomAJ, ChapinFS, MooneyHA (1985). Resource limitation in plants—An economic analogy .Annual Review of Ecology and Systematics, 16, 363-392. [本文引用: 1]
[4]
CairnsMA, BrownS, HelmerEH, BaumgardnerGA (1997). Root biomass allocation in the world’s upland forests .Oecologia, 111, 1-11. [本文引用: 1]
[5]
ChunML, XieZQ, ZhaoCM, FanDY, XuXW, PingL (2009). Litter production and nutrient characteristic of Abies far-gesii natural forest in Shennongjia Nature Reserve, Hubei, China . Chinese Journal of Plant Ecology, 33, 492-498. (in Chinese with English abstract)[春敏莉, 谢宗强, 赵常明, 樊大勇, 徐新武, 平亮 (2009). 神农架巴山冷杉天然林凋落量及养分特征 . 植物生态学报, 33, 492-498.] [本文引用: 1]
[6]
FangJY, ChenAP (2001). Dynamic forest biomass carbon pools in China and their significance .Acta Botanica Sinica, 43, 967-973. (in Chinese with English abstract)[方精云, 陈安平 (2001). 中国森林植被碳库的动态变化及其意义 . 植物学报, 43, 967-973.] [本文引用: 1]
[7]
FangJY, GuoZD, PiaoSL, ChenAP (2007). Estimation of carbon sink of terrestrial vegetation in 1981-2000 .Science in China: Series D, 37, 804-812. (in Chinese)[方精云, 郭兆迪, 朴世龙, 陈安平 (2007). 1981-2000年中国陆地植被碳汇的估算 . 中国科学D辑, 37, 804-812.] [本文引用: 2]
[8]
GaoQ, YangXC, YinCY, LiuQ (2014). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province .Chinese Journal of Plant Ecology, 38, 355-365. (in Chinese with English abstract)[高巧, 阳小成, 尹春英, 刘庆 (2014). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算 . 植物生态学报, 38, 355-365.] [本文引用: 1]
[9]
GoodaleCL, DavidsonEA (2002). Carbon cycle: Uncertain sinks in the shrubs .Nature, 418, 593-594. [本文引用: 1]
[10]
HeJS, WangQB, HuD (1997). Studies on the biomass of topical shrubland and their regeneration capacity after cutting .Acta Phytoecologica Sinica, 21, 512-520. (in Chinese with English abstract)[贺金生, 王其兵, 胡东 (1997). 长江三峡地区典型灌丛的生物量及其再生能力 . 植物生态学报, 21, 512-520.] [本文引用: 1]
[11]
HouL (2009). Carbon Balance in Natural Secondary Pinus tabulaeformis Forest at Huoditang Forest Zone in the Qinling Mountains. PhD dissertation, Northwest A & F University, Yangling, Shaanxi. 30-32. (in Chinese with English abstract)[侯琳 (2009). 秦岭火地塘林区天然次生油松林碳平衡研究 . 博士学位论文, 西北农林科技大学, 陕西杨凌. 30-32.] [本文引用: 1]
[12]
HuHF, WangZH, LiuGH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Ecology (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 4]
[13]
KetteringsQM, CoeR, van NoordwijkM, PalmCA (2001). Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests .Forest Ecology and Management, 146, 199-209. [本文引用: 1]
[14]
LiJX (2015). Biodiversity Pattern and Ecosystem Productivity in Shrubland Across the Subtropical Region in China. PhD dissertation, Institute of Botany, Chinese Academy of Sciences, Beijing. 76-84. (in Chinese with English abstract).[李家湘 (2015). 亚热带灌丛生物多样性及其与生产力的关系 . 博士学位论文, 中国科学院植物研究所, 北京. 76-84.] [本文引用: 3]
[15]
LiJX, ZhangX, XieZQ, LuCF, TuXY, XunY (2015). Community composition and structure of Rhododendron simsii shrubland in the Dawei Mountain, Hunan Province . Biodiversity Science, 23, 815-823. (in Chinese with English abstract)[李家湘, 张旭, 谢宗强, 卢从发, 涂向阳, 寻院 (2015). 湖南大围山杜鹃灌丛的群落组成及结构特征 . 生物多样性, 23, 815-823.] [本文引用: 2]
[16]
LiXR, LiuJQ, ChenYR, HuLL, YangFT (2006). Aboveground biomass of three conifers in Qianyanzhou plantation .Chinese Journal of Applied Ecology, 17, 1382-1388. (in Chinese with English abstract)[李轩然, 刘琪璟, 陈永瑞, 胡理乐, 杨风亭 (2006). 千烟洲人工林主要树种地上生物量的估算 . 应用生态学报, 17, 1382-1388.] [本文引用: 1]
[17]
LiY, ZhangJG, DuanAG, XiangCW (2010). Selection of biomass estimation models for Chinese fir plantation .Chinese Journal of Applied Ecology, 21, 3036-3046. (in Chinese with English abstract)[李燕, 张建国, 段爱国, 相聪伟 (2010). 杉木人工林生物量估算模型的选择 . 应用生态学报, 21, 3036-3046.] [本文引用: 1]
[18]
LiYL, GongHD (2015). Characteristics of community structure and biomass distribution of Rhododendron adenogynum shrub in Laojunshan Mountain . Journal of West China Forestry Science, 44, 121-124. (in Chinese with English abstract)[李云龙, 巩合德 (2015). 云南老君山腺房杜鹃灌丛群落特征及生物量分配特征研究 . 西部林业科学, 44, 121-124.]
[19]
LüXT, TangJW, HeYC, DuanWG, SongJP, XuHL, ZhuSZ (2007). Biomass and its allocation in tropical seasonal rain forest in Xishuangbanna, southwest China .Journal of Plant Ecology (Chinese Version), 31, 11-22. (in Chinese with English abstract)[吕晓涛, 唐建维, 何有才, 段文贵, 宋军平, 许海龙, 朱胜忠 (2007). 西双版纳热带季节雨林的生物量及其分配特征 . 植物生态学报, 31, 11-22.] [本文引用: 1]
[20]
MariessiiA, BetuleE, ToshihicokoH (1991). Growth patterns of tree height and stem diameter in populations of Abies veitchi. Journal of Ecology, 79, 1085-1095. [本文引用: 1]
[21]
McCarthyMC, EnquistBJ (2007). Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation .Functional Ecology, 21, 713-720. [本文引用: 1]
[22]
McConnaughayK, ColemanJ (1999). Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients .Ecology, 80, 2581-2593. [本文引用: 1]
MokanyK, RaisonRJ, ProkushkinAS (2006). Critical analysis of root:shoot ratios in terrestrial biomes .Global Change Biology, 12, 84-96. [本文引用: 1]
[25]
MontesN, GauquelinT, BadriW, BertaudiereV, ZaouiEH (2000). A non-destructive method for estimating above- ground forest biomass in threatened woodlands .Forest Ecology and Management, 130, 37-46. [本文引用: 1]
[26]
NávarJ (2009). Allometric equations for tree species and carbon stocks for forests of northwestern Mexico .Forest Ecology and Management, 257, 427-434. [本文引用: 1]
[27]
PoorterH, NiklasKJ, ReichPB, OleksynJ, PootP, MommerL (2012). Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control .New Phytologist, 193, 30-50. [本文引用: 3]
[28]
SpainAV (1984). Litterfall and the standing crop of litter in three topical Australian rain forests .Journal of Ecology, 72, 947-961. [本文引用: 1]
[29]
SturmM, RacineC, TapeK (2001). Climate change: Increasing shrub abundance in the Arctic .Nature, 411, 546-547. [本文引用: 1]
[30]
VourlitisGL, ZorbaG, PasquiniSC, MustardR (2007). Carbon and nitrogen storage in soil and litter of southern Californian semi-arid shrublands .Journal of arid Environments, 70, 164-173. [本文引用: 1]
[31]
WangJS, ZhangCY, FanXH, ZhaoYZ (2011). Biomass allo- cation patterns and allometric models of Abies nephrolepis Maxim . Acta Ecologica Sinica, 31, 3918-3927. (in Chinese with English abstract)[汪金松, 张春雨, 范秀华, 赵亚洲 (2011). 臭冷杉生物量分配格局及异速生长模型 . 生态学报, 31, 3918-3927.] [本文引用: 1]
[32]
WangL (2009). Study of Biomass and Its Models of Main Shrub Community Type in Northwest Sichuan . Master de- gree dissertation, Sichuan Agricultural University, Ya’an, Sichuan. (in Chinese with English abstract)[王玲 (2009). 川西北地区主要灌丛类型生物量及其模型的研究 . 硕士学位论文, 四川农业大学, 四川雅安.] [本文引用: 3]
[33]
WeinerJ (2004). Allocation, plasticity and allometry in plants .Perspectives in Plant Ecology, Evolution and Systematics, 6, 207-215. [本文引用: 1]
[34]
WeiXP, ZhaoCM, WangGX, ChenBM, ChengDL (2005). Estimation of above- and below-ground biomass of dominant desert plant species in an oasis-desert ecotone of Minqin, China .Acta Phytoecologica Sinica, 29, 878-883. (in Chinese with English abstract)[魏小平, 赵长明, 王根轩, 陈宝明, 程栋梁 (2005). 民勤荒漠绿洲过渡带优势植物地上和地下生物量的估测模型 . 植物生态学报, 29, 878-883.] [本文引用: 1]
[35]
WoodwellGM, WhitakerRH, ReinersWA, LikensGE, DelwichCC, BotkinDB (1978). Biota and the world carbon budget .Science, 199, 141-146. [本文引用: 1]
[36]
WuZY (1980). Vegetation of China. Science Press, Beijing. (in Chinese).[吴征镒 (1980). 中国植被. 科学出版社, 北京.] [本文引用: 2]
[37]
ZhangGF, SongYC (2001). Studies on the biomass of Castanopsis sclerophylla + Quercus fabri shrubland in Tiantong Region, Zhejiang Province . Journal of Wuhan Botanical Research, 19, 101-106. (in Chinese with English abstract)[张光富, 宋永昌 (2001). 浙江天童苦槠+白栎灌丛群落的生物量研究 . 武汉植物学研究, 19, 101-106.] [本文引用: 1]
[38]
ZhangQ, LiJX, XieZQ (2017). Effects of nitrogen addition on soil respiration of Rhododendron simsii shrubland in the subtropical mountainous areas of China . Chinese Journal of Plant Ecology, 41, 95-104. (in Chinese with English abstract)[张蔷, 李家湘, 谢宗强 (2017). 氮添加对亚热带山地杜鹃灌丛土壤呼吸的影响 . 植物生态学报, 41, 95-104.] [本文引用: 1]
[39]
ZhangYR, OuyangX, LiYL, LiuSZ, ZhangDQ, ZhouGY (2013). Shrub community characteristics and quantitative calculation of theirs biomass in southern China .Journal of Central South University of Forestry & Technology, 33(9), 71-79. (in Chinese with English abstract)[张亚茹, 欧阳旭, 李跃林, 刘世忠, 张德强, 周国逸 (2013). 我国南亚热带灌丛群落特征及生物量的定量计算 . 中南林业科技大学学报, 33(9), 71-79.] [本文引用: 1]
[40]
ZhengSW, TangM, ZouJH, MuCL (2007). Summary of research on shrub biomass in China .Journal of Chengdu University (Natural Science Edition), 26, 189-192. (in Chinese with English abstract)[郑绍伟, 唐敏, 邹俊辉, 慕长龙 (2007). 灌木群落及生物量研究综述 . 成都大学学报(自然科学版), 26, 189-192.] [本文引用: 3]
[41]
ZianisD, MencucciniM (2004). On simplifying allometric analyses of forest biomass .Forest Ecology and Management, 187, 311-332. [本文引用: 1]
Comparison of formulae for biomass content determina- tion in a tropical rain forest site in the state of Pará, Brazil 1 1999
... 利用树木易测因子建立生长方程来推算生物量, 此方法简单、迅速且破坏性小, 同时数据可以进行外推, 使生物量的跟踪调查研究成为可能, 受到广泛的关注和应用(Araújo et al., 1999; Montes et al., 2000).本试验中所有的生长方程都达到了极显著水平(p < 0.001), 对生物量变化的解释程度平均为90.3%.由此可知, 灌木生物量可以通过生长方程推算得出, 且结果具有较高的准确度.在本研究中, 方程拟合程度最好的自变量为D2H和D, 这与川西北地区主要灌丛类型生物量模型的研究结果(王玲, 2009)一致.建立生物量生长方程最常用的单一自变量是树干直径(Zianis & Mencuccini, 2004), 特别是对于高大乔木, 树径易于测量且准确度高, 而株高不易测量且误差较大, 株高的引入会使模型增加新的变异(吕晓涛等, 2007; 汪金松等, 2011).但是灌丛株高的测量较为方便, 虽然引入株高有时并不会增加方程对生物量变化的解释程度, 但是可以增加方程外推时的适用性(Ketterings et al., 2001).本研究中最佳生物量估测模型的函数类型以幂函数为主, 这与前人的研究结果一致(Basuki et al., 2009; Návar, 2009; 李燕等, 2010), 幂函数能较为真实地反映灌木生物量随株高、基径的变化趋势(郑绍伟等, 2007). ...
Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests 1 2009
... 利用树木易测因子建立生长方程来推算生物量, 此方法简单、迅速且破坏性小, 同时数据可以进行外推, 使生物量的跟踪调查研究成为可能, 受到广泛的关注和应用(Araújo et al., 1999; Montes et al., 2000).本试验中所有的生长方程都达到了极显著水平(p < 0.001), 对生物量变化的解释程度平均为90.3%.由此可知, 灌木生物量可以通过生长方程推算得出, 且结果具有较高的准确度.在本研究中, 方程拟合程度最好的自变量为D2H和D, 这与川西北地区主要灌丛类型生物量模型的研究结果(王玲, 2009)一致.建立生物量生长方程最常用的单一自变量是树干直径(Zianis & Mencuccini, 2004), 特别是对于高大乔木, 树径易于测量且准确度高, 而株高不易测量且误差较大, 株高的引入会使模型增加新的变异(吕晓涛等, 2007; 汪金松等, 2011).但是灌丛株高的测量较为方便, 虽然引入株高有时并不会增加方程对生物量变化的解释程度, 但是可以增加方程外推时的适用性(Ketterings et al., 2001).本研究中最佳生物量估测模型的函数类型以幂函数为主, 这与前人的研究结果一致(Basuki et al., 2009; Návar, 2009; 李燕等, 2010), 幂函数能较为真实地反映灌木生物量随株高、基径的变化趋势(郑绍伟等, 2007). ...
Resource limitation in plants—An economic analogy 1 1985

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT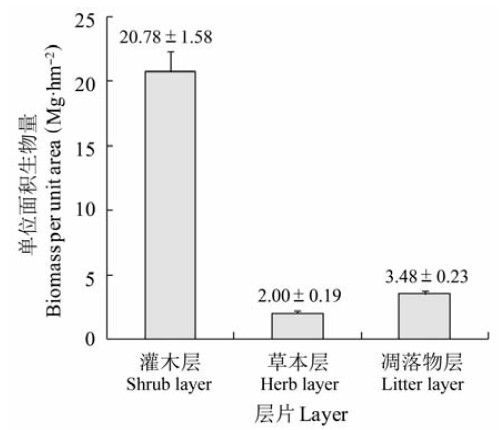 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT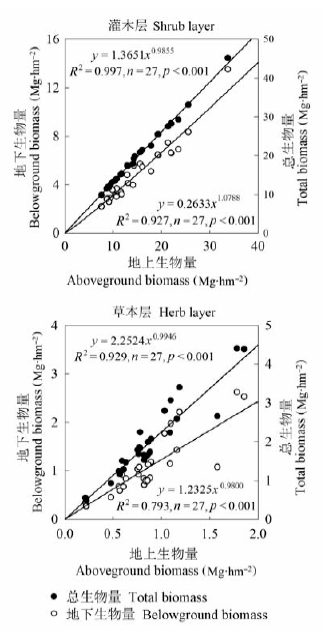 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}