
Biomass estimation models and allocation patterns of 14 shrub species in Mountain Luya, Shanxi, China
LUOYong-Kai通讯作者:
责任编辑:
收稿日期:2016-04-11
接受日期:2016-07-23
网络出版日期:2017-01-10
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Methods We used plant height, basal diameter, canopy diameter and their combination as variables to establish the optimal biomass estimation models for each shrub species. In addition, we used the ratios of leaf, stem, root to total biomass, and belowground to aboveground biomass to explore the difference of biomass allocation patterns of 14 shrub species.
Important findings Most of biomass estimation models could be well expressed by the exponential and linear functions. Biomass for shorter shrub species with more stems could be better estimated by canopy area; biomass for taller shrub species with less stems could be better estimated by the sum of the square of total base diameter multiply stem height; and biomass for the rest shrub species could be better estimated by canopy volume. The averaged value for these shrub species was 0.61, 0.17, 0.48, and 0.35 for R/S, leaf biomass fraction, stem biomass fraction, and root biomass fraction, respectively. Except for leaf biomass fraction, R/S, stem biomass fraction, and root biomass fraction for shrubs with thorn was significantly greater than that for shrubs without thorn.
Keywords:
-->0
PDF (520KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(Bloom et al., 1985; 刘长成等, 2009)。灌木作为陆地生态系统一种重要的生活型, 以较强的抗逆性及特殊的形态特征在全球陆地广泛分布, 它不仅在保持水土、防风固沙等方面有着不可替代的作用, 而且是陆地生态系统物质循环和能量流动的重要组成部分(俞海生等, 2003)。灌木生物量作为灌木群落和生态系统研究的重要指标, 它不仅是灌木与乔木幼苗竞争能力的反映, 而且反映了灌丛生态系统对周围环境的适应。刈割法是研究灌木生物量的方法之一, 但这种方法费事费力, 破坏性较大。通过建立生物量和灌木形态指标之间的关系来估测灌木的生物量, 是现在常用的估算灌木生物量的方法。关于灌木生物量模型, 国外如Whittaker (1982)对灌丛的净生产力和生物量有过研究, Salis等(2006)对巴西稀树草原灌木地上生物量建立过模型。国内较早见于姜凤岐和卢凤勇(1982)对小叶锦鸡儿(Caragana microphylla)等少数几种灌木的地上生物量建立过生物量方程。之后, 关于灌木生物量模型的研究陆续出现(陈遐林等, 2002; 张海清等, 2005; 蔡哲等, 2006; 方精云等, 2006;
生物量分配是表征植物生理过程的重要指标(Enquist & Niklas, 2002), 主要受植物的遗传特性、环境条件、个体大小、植物功能型的影响, 因而在不同环境与生长压力下植物有不同的生长特性与物质分配规律(程栋梁, 2007; 李旭东等, 2012; Pooter et al., 2012)。
目前关于生物量分配的研究多集中于乔木, 而有关灌木生物量分配的研究较少。雷蕾(2012)对祁连山高山灌丛生物量分配有过研究, 刘兴良等(2006)探讨过川滇高山栎(Quercus aquifolioides)灌丛地上生物量分配的特征。相比于乔木, 灌木所处的环境往往较为干旱、贫瘠。为适应所处环境, 灌木往往会呈现带刺、根深等特殊的形态特征(易海燕, 2010)。因此, 刺不仅是植物防御敌害的生存策略, 而且是植物对干旱环境适应的一种形态特征。而生长在较为干旱环境中的带刺与不带刺灌木之间生物量分配是否有差异, 带刺灌木是否改变了生物量分配策略的研究目前较为少见。
芦芽山国家自然保护区位于山西省忻州市, 保护区内植被类型丰富, 森林覆盖率达到58.8%, 灌木种类较多, 但关于当地灌木生物量估测及生物量分配方面的研究鲜有报道。为准确评估保护区内的森林资源, 为保护区内灌丛生态系统生物量的持续调查提供依据, 本研究通过调查保护区内14种常见灌木的形态因子与生物量, 建立了保护区内14种常见灌木的最优生物量估算模型; 此外, 还对这些灌木种各器官间的生物量分配规律及带刺与不带刺灌木种之间的生物量分配差异进行了探讨。这些研究一方面为保护区估算灌木群落生物量提供了准确、简洁的方法, 另一方面有助于完善对灌木生物量分配规律的认识。
1 材料和方法
1.1 研究区概况
山西芦芽山国家级自然保护区位于山西省吕梁山脉北端, 地处宁武、五寨、岢岚三县的交界处, 111.83°-112.09° E, 38.59°-38.75° N, 区内最高海拔为主峰芦芽山2787 m, 最低海拔1346 m, 年平均气温4-7 ℃, 年降水量500-600 mm, 无霜期90-120天。该区土壤分布复杂, 区域东部是黄土母质发育而成的灰褐土, 西部以褐土为主。随海拔高度的升高, 土壤呈现垂直地带性, 依次为山地褐土、山地淋溶褐土、棕色森林土和亚高山草甸土。芦芽山植被具有明显的垂直地带性, 从山麓到山顶依次为森林草原带、落叶阔叶林带、针阔叶混交林带、寒温性针叶林带、亚高山灌丛草甸带。芦芽山东麓西麓基带不同, 东麓基带为森林草原带, 西麓基带为落叶阔叶林带。沿海拔梯度, 保护区内优势的灌丛群落主要有沙棘(Hippophae rhamnoides)灌丛、山刺玫(Rosa davurica)灌丛、绣线菊(Spiraea spp.)灌丛、虎榛子(Ostryopsis davidiana)灌丛、蒙古荚蒾(Viburnum mongolicum)灌丛、金花忍冬(Lonicera chrysantha)灌丛、银露梅(Potentilla glabra)灌丛和鬼箭锦鸡儿(Caragana jubata)灌丛(张金屯, 1989)。1.2 样品采集方法
所有样品均采自于2014年7月, 选取芦芽山自然保护区内14种常见灌木作为研究对象, 即沙棘、三裂绣线菊(Spiraea trilobata)、土庄绣线菊(S. pubescens)、黄刺玫(Rosa xanthina)、虎榛子、榛(Corylus heterophylla)、金花忍冬、蒙古荚蒾、唐古特忍冬(Lonicera tangutica)、复盆子(Rubus idaeus)、银露梅、山刺玫、刺果茶藨子(Ribes burejense)和鬼箭锦鸡儿, 14种灌木取样点的群落类型都为灌丛群落。14种灌木采样调查基本信息见表1。MacDicken (1997)①(① MacDicken KG (1997). A guide to monitoring carbon storage in forestry and agroforestry projects. Forest Carbon Monitoring Program, Winrock International Institute for Agricultural Development.)认为对于区域性生物量表的建立需要30-100株样木, 如果每个径级的样木均匀分布的话, 30株样木是最合适的。因此我们在每种灌木分布较为集中的区域从小到大调查30株样木。其中鬼箭锦鸡儿为贴地生长, 银露梅和三裂绣线菊地上分枝数极多, 难以调查其基径, 只调查其冠幅(取最大直径和最小直径的平均值)和株高, 其余11种灌木调查株高、冠幅以及每株灌木地上所有分枝的基径用以求得每株的总基径。测量完毕后将植株整株挖出, 深度为根系分布所达范围。将植株分根、茎、叶三部分储存, 所有样品带回实验室在65 ℃烘箱内烘干至恒质量, 测得相应的干质量。模型参数的实测区间和相应的生物量见附录I。Table 1
表1
表1芦芽山14种灌木调查基本信息表
Table 1The basic information of 14 shrub species in Mountain Luya
| 物种 Species | 经度 Longitude (E) | 纬度 Latitude (N) | 海拔 Elevation (m) | 科 Family | 属 Genus | 是否带刺 Thorn |
|---|---|---|---|---|---|---|
| 沙棘 Hippophae rhamnoides | 112.01° | 38.68° | 1 499.8 | 胡颓子科 Elaeagnaceae | 沙棘属 Hippophae | 是 Yes |
| 三裂绣线菊 Spiraea trilobata | 111.99° | 38.69° | 1 535.6 | 蔷薇科 Rosaceae | 绣线菊属 Spiraea | 否 No |
| 黄刺玫 Rosa xanthina | 111.99° | 38.69° | 1 535.6 | 蔷薇科 Rosaceae | 蔷薇属 Rosa | 是 Yes |
| 虎榛子 Ostryopsis davidiana | 111.99° | 38.69° | 1 535.6 | 桦木科 Betulaceae | 虎榛子属 Ostryopsis | 否 No |
| 榛 Corylus heterophylla | 111.99° | 38.69° | 1 535.6 | 桦木科 Betulaceae | 榛属 Corylus | 否 No |
| 土庄绣线菊 Spiraea pubescens | 111.98° | 38.72° | 1 659.9 | 蔷薇科 Rosaceae | 绣线菊属 Spiraea | 否 No |
| 金花忍冬 Lonicera chrysantha | 111.92° | 38.65° | 1 778.4 | 忍冬科 Caprifoliaceae | 忍冬属 Lonicera | 否 No |
| 蒙古荚蒾 Viburnum mongolicum | 111.92° | 38.65° | 1 780.0 | 忍冬科 Caprifoliaceae | 荚迷属 Viburnum | 否 No |
| 唐古特忍冬 Lonicera tangutica | 111.92° | 38.65° | 1 780.0 | 忍冬科 Caprifoliaceae | 忍冬属 Lonicera | 否 No |
| 复盆子 Rubus idaeus | 111.92° | 38.65° | 1 780.0 | 蔷薇科 Rosaceae | 悬钩子属 Rubus | 是 Yes |
| 银露梅 Potentilla glabra | 111.96° | 38.72° | 1 812.2 | 蔷薇科 Rosaceae | 委陵菜属 Potentilla | 否 No |
| 山刺玫 Rosa davurica | 111.95° | 38.71° | 1 874.3 | 蔷薇科 Rosaceae | 蔷薇属 Rosa | 是 Yes |
| 刺果茶藨子 Ribes burejense | 111.95° | 38.71° | 1 891.1 | 虎耳草科 Saxifragaceae | 茶藨子属 Ribes | 是 Yes |
| 鬼箭锦鸡儿 Caragana jubata | 111.86° | 38.73° | 2 602.2 | 豆科 Leguminosae | 锦鸡儿属 Caragana | 是 Yes |
新窗口打开
Appendix I
附录I
附录I模型参数的实测区间
Appendix IMeasurement intervals of model parameters
 |
新窗口打开
1.3 数据处理分析
用SPSS 20.0软件进行生物量模型的拟合。选取总基径(Dt)、株高(H)、冠幅(C)、冠幅面积(A = π(C/2)2)、冠幅体积(V = A × H)、总基径平方与株高乘积(Dt2H)为变量, 对各器官、地上和总生物量进行模型拟合。模型采用以下3种类型:W = a + bX (1)
W = a + blnX (2)
W = aXb (3)
其中W表示预测变量(本文中指各类生物量), X表示拟合变量(如株高、冠幅等), a、b表示拟合参数。根据决定系数(R2)、估计值的标准误差(SEE)以及平均预估误差(MPE)的大小及回归检验显著水平(p值)来评价模型的优劣(曾伟生和唐守正, 2011)。从中选出拟合度最好、相关最密切的数学模型作为生物量的估算模型。生物量分叶生物量W叶、茎生物量W茎、根生物量W根、地上生物量W地上和总生物量W总。
生物量分配测定指标包括: 根冠比(地下生物量/地上生物量)、叶质比(叶生物量/总生物量)、茎质比(茎生物量/总生物量)和根质比(根生物量/总生物量)。分别根据灌木是否带刺对14种灌木进行分类, 将物种作为随机变量, 对不同种类灌木的根冠比、叶质比、茎质比和根质比采用单因素方差分析比较其差异, 所有的数据处理和分析在Excel 2010和SPSS 20.0中完成。
2 结果
2.1 生物量模型的拟合
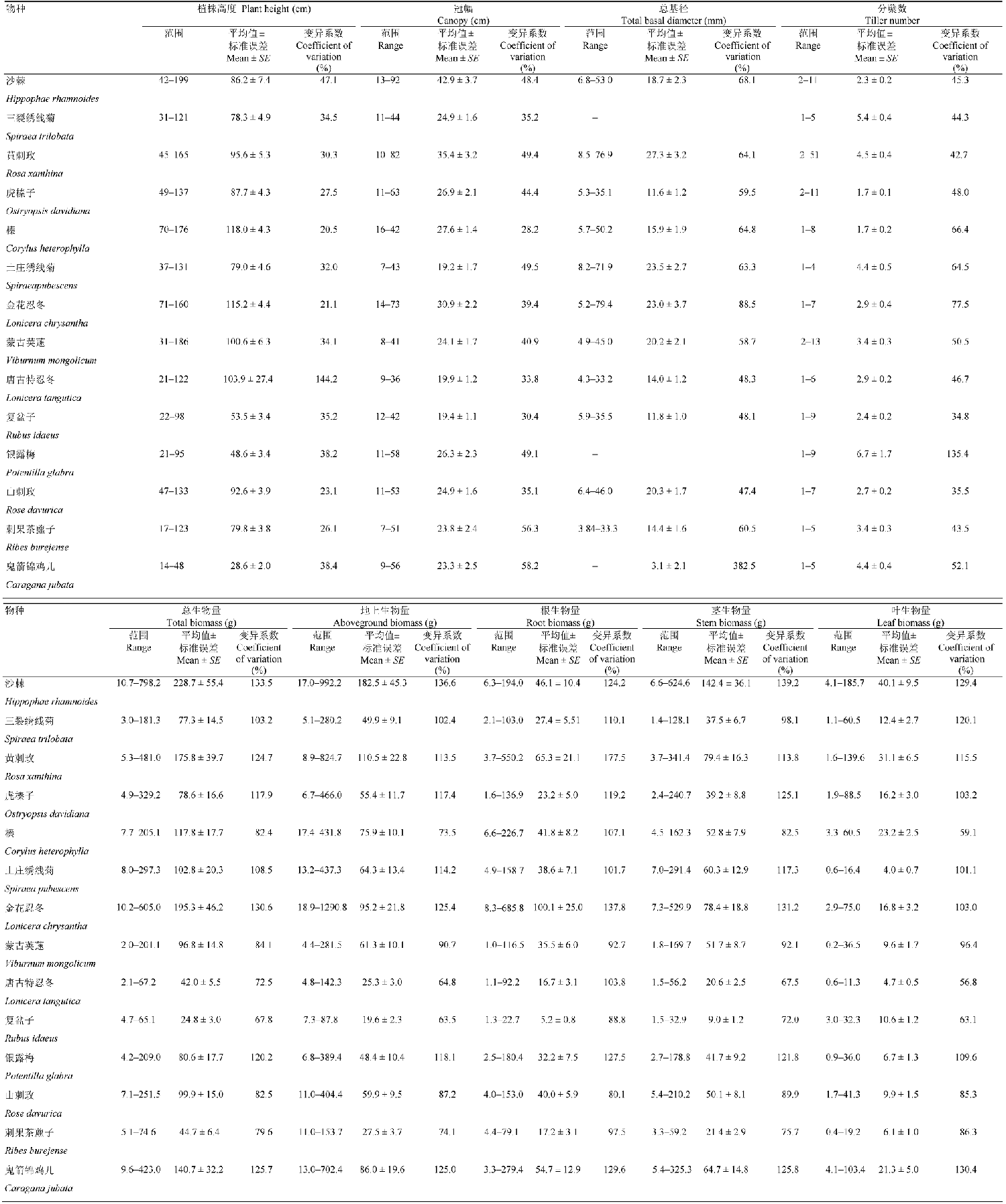
经回归分析, 建立了14种灌木3个器官、地上及总生物量的最优生物量拟合模型(表2)。除复盆子的叶生物量模型外, 14种灌木的最优生物量模型均为幂函数或线性函数。冠幅面积(A)、 冠幅体积(V)、总基径平方与树高乘积(Dt2H)是生物量方程的最佳预测变量。所有灌木生物量的拟合方程都表现出较好的拟合效果(p < 0.001; MPE < 25.1%), 并且除三裂绣线菊和刺果茶藨子外, 其余12种灌木的生物量拟合方程R2都在0.7以上。从模型的筛选结果来看, 沙棘、金花忍冬的所有生物量方程以及山刺玫、榛、唐古特忍冬的大部分生物量方程其预测变量都是Dt2H, 银露梅、鬼箭锦鸡儿所有生物量方程预测变量都为A, 其他种生物量方程最佳预测变量大多为V。图1以不同种的总生物量为例, 对3种预测变量、两种方程形式构成的6种主要生物量方程进行展示。Table 2
表2
表214种灌木总生物量(WTotal)、地上生物量(WAboveground)、根生物量(WRoot)、茎生物量(WStem)、叶生物量(WLeaf)的最佳拟合模型(n = 30)
Table 2The best estimation models for total biomass (WTotal), aboveground biomass (WAboveground), root biomass (WRoot) , stem biomass (WStem), and leaf biomass (WLeaf) of 14 shrub species (n = 30)
| 物种名 Species | 最佳方程 Best model | 最佳变量 Best variable | 参数a Parameter variable a | 参数b Parameter variable b | 平均预估误差Mean prediction error (MPE) (%) | 估计值的标准误差 Standard error of estimate (SEE) | R2 | p |
|---|---|---|---|---|---|---|---|---|
| 沙棘 Hippophae rhamnoides | WTotal = aDt2H + b | Dt2H | 0.002 0 | 52.34 | 15.48 | 94.78 | 0.91 | <0.001 |
| WAboveground = aDt2H + b | Dt2H | 0.002 0 | 40.77 | 17.42 | 85.18 | 0.90 | <0.001 | |
| WRoot = aDt2H + b | Dt2H | 0.000 4 | 11.58 | 12.24 | 15.12 | 0.93 | <0.001 | |
| WStem = aDt2H + b | Dt2H | 0.001 0 | 30.06 | 18.10 | 69.05 | 0.89 | <0.001 | |
| WLeaf = aDt2H + b | Dt2H | 0.000 4 | 10.71 | 17.27 | 18.55 | 0.88 | <0.001 | |
| 三裂绣线菊 Spiraea trilobata | WTotal = aVb | V | 0.049 0 | 0.66 | 0.40 | 0.83 | 0.45 | <0.001 |
| WAboveground = aVb | V | 0.022 0 | 0.69 | 0.64 | 0.85 | 0.46 | <0.001 | |
| WRoot = aVb | V | 0.030 0 | 0.60 | 1.17 | 0.86 | 0.39 | <0.001 | |
| WStem = aVb | V | 0.012 0 | 0.72 | 0.95 | 0.96 | 0.42 | <0.001 | |
| WLeaf = aVb | V | 0.005 0 | 0.70 | 2.57 | 0.85 | 0.46 | <0.001 | |
| 黄刺玫 Rosa xanthina | WTotal = aVb | V | 0.005 0 | 0.88 | 0.07 | 0.34 | 0.90 | <0.001 |
| WAboveground = aVb | V | 0.003 0 | 0.88 | 0.11 | 0.32 | 0.93 | <0.001 | |
| WRoot = aVb | V | 0.005 0 | 0.77 | 0.29 | 0.50 | 0.80 | <0.001 | |
| WStem= aVb | V | 0.002 0 | 0.89 | 0.16 | 0.35 | 0.92 | <0.001 | |
| WLeaf = aVb | V | 0.001 0 | 0.86 | 0.40 | 0.33 | 0.92 | <0.001 | |
| 虎榛子 Ostryopsis davidiana | WTotal = aV + b | V | 0.001 0 | 9.28 | 13.66 | 29.28 | 0.90 | <0.001 |
| WAboveground = aV + b | V | 0.000 8 | 6.25 | 12.87 | 19.43 | 0.91 | <0.001 | |
| WRoot = aV + b | V | 0.000 2 | 3.03 | 16.66 | 10.56 | 0.86 | <0.001 | |
| WStem = aV + b | V | 0.000 6 | 2.93 | 16.70 | 17.86 | 0.87 | <0.001 | |
| WLeaf = aV + b | V | 0.000 2 | 3.32 | 9.14 | 4.03 | 0.94 | <0.001 | |
| 榛 Corylus heterophylla | WTotal = a(Dt2H) b | Dt2H | 0.145 0 | 0.64 | 0.10 | 0.32 | 0.86 | <0.001 |
| WAboveground = aVb | V | 0.000 1 | 1.15 | 0.17 | 0.34 | 0.85 | <0.001 | |
| WRoot = a(Dt2H) b | Dt2H | 0.032 0 | 0.68 | 0.30 | 0.34 | 0.86 | <0.001 | |
| WStem = a(Dt2H) b | Dt2H | 0.031 0 | 0.71 | 0.28 | 0.40 | 0.82 | <0.001 | |
| WLeaf = aVb | V | 0.000 9 | 0.90 | 0.48 | 0.30 | 0.81 | <0.001 | |
| 土庄绣线菊 Spiraea pubescens | WTotal = aV + b | V | 0.001 0 | 18.90 | 13.81 | 38.03 | 0.89 | <0.001 |
| WAboveground = aV + b | V | 0.002 0 | 8.02 | 12.84 | 22.11 | 0.91 | <0.001 | |
| WRoot = aVb | V | 0.020 0 | 0.74 | 0.44 | 0.45 | 0.78 | <0.001 | |
| WStem = aV + b | V | 0.002 0 | 6.18 | 12.95 | 20.92 | 0.92 | <0.001 | |
| WLeaf = a(Dt2H) b | Dt2H | 0.011 0 | 0.53 | 4.33 | 0.46 | 0.71 | <0.001 | |
| 金花忍冬 Lonicera chrysantha | WTotal = a(Dt2H) b | Dt2H | 0.365 0 | 0.56 | 0.08 | 0.42 | 0.82 | <0.001 |
| WAboveground = a(Dt2H) b | Dt2H | 0.233 0 | 0.53 | 0.16 | 0.40 | 0.83 | <0.001 | |
| WRoot = aDt2H + b | Dt2H | 0.000 5 | 43.22 | 25.05 | 67.14 | 0.76 | <0.001 | |
| WStem = a(Dt2H)b | Dt2H | 0.136 0 | 0.56 | 0.20 | 0.41 | 0.83 | <0.001 | |
| WLeaf = aDt2H + b | Dt2H | 0.000 1 | 9.08 | 18.23 | 8.19 | 0.78 | <0.001 | |
| 蒙古荚蒾 Viburnum mongolicum | WTotal = aVb | V | 0.006 0 | 0.88 | 0.12 | 0.32 | 0.92 | <0.001 |
| WAboveground = aVb | V | 0.003 0 | 0.90 | 0.21 | 0.34 | 0.92 | <0.001 | |
| WRoot = aVb | V | 0.002 0 | 0.84 | 0.70 | 0.66 | 0.72 | <0.001 | |
| WStem= aVb | V | 0.001 0 | 0.91 | 0.34 | 0.47 | 0.91 | <0.001 | |
| WLeaf = aVb | V | 0.000 3 | 0.89 | 1.94 | 0.50 | 0.85 | <0.001 | |
| 唐古特忍冬 Lonicera tangutica | WTotal = a(Dt2H) b | Dt2H | 0.200 0 | 0.54 | 0.30 | 0.34 | 0.80 | <0.001 |
| WAboveground = aVb | V | 0.007 0 | 0.80 | 0.49 | 0.33 | 0.84 | <0.001 | |
| WRoot= aDt2H + b | Dt2H | 0.000 4 | 6.22 | 11.48 | 5.13 | 0.73 | <0.001 | |
| 物种名 Species | 最佳方程 Best model | 最佳变量 Best variable | 参数a Parameter variable a | 参数b Parameter variable b | 平均预估误差Mean prediction error (MPE) (%) | 估计值的标准误差Standard error of estimate (SEE) | R2 | p |
| 唐古特忍冬 Lonicera tangutica | WStem = a(Dt2H) b | Dt2H | 0.063 0 | 0.59 | 0.74 | 0.41 | 0.77 | <0.001 |
| WLeaf = aVb | V | 0.005 0 | 0.68 | 2.30 | 0.29 | 0.81 | <0.001 | |
| 复盆子 Rubus idaeus | WTotal = aV + b | V | 0.000 6 | 12.52 | 10.18 | 6.76 | 0.84 | <0.001 |
| WAboveground = aV + b | V | 0.000 4 | 10.71 | 10.63 | 5.59 | 0.80 | <0.001 | |
| WRoot = aV + b | V | 0.000 0 | 1.81 | 13.92 | 1.92 | 0.83 | <0.001 | |
| WStem = aV + b | V | 0.000 2 | 4.59 | 14.17 | 3.43 | 0.73 | <0.001 | |
| WLeaf = alnV + b | V | 14.166 0 | -69.23 | 11.41 | 3.24 | 0.77 | <0.001 | |
| 银露梅 Potentilla glabra | WTotal = aA + b | A | 0.131 0 | -6.74 | 19.50 | 42.11 | 0.82 | <0.001 |
| WAboveground = aA + b | A | 0.077 0 | -2.81 | 19.75 | 25.62 | 0.81 | <0.001 | |
| WRoot = aA + b | A | 0.054 0 | -3.93 | 22.54 | 19.43 | 0.78 | <0.001 | |
| WStem = aA + b | A | 0.068 0 | -3.26 | 20.73 | 23.16 | 0.80 | <0.001 | |
| WLeaf = aA + b | A | 0.010 0 | 0.44 | 20.89 | 3.76 | 0.74 | <0.001 | |
| 山刺玫 Rosa davurica | WTotal = a(Dt2H) b | Dt2H | 0.061 0 | 0.69 | 0.11 | 0.29 | 0.88 | <0.001 |
| WAboveground = a(Dt2H) b | Dt2H | 0.042 0 | 0.67 | 0.22 | 0.35 | 0.83 | <0.001 | |
| WRoot = a(Dt2H) b | Dt2H | 0.023 0 | 0.70 | 0.37 | 0.40 | 0.80 | <0.001 | |
| WStem = a(Dt2H) b | Dt2H | 0.025 0 | 0.70 | 0.27 | 0.36 | 0.84 | <0.001 | |
| WLeaf = aA+b | A | 0.020 0 | -0.87 | 7.35 | 1.94 | 0.95 | <0.001 | |
| 刺果茶藨子 Ribes burejense | WTotal = aVb | V | 0.550 0 | 0.41 | 0.36 | 0.43 | 0.64 | <0.001 |
| WAboveground = aV + b | V | 0.000 4 | 12.39 | 16.40 | 12.07 | 0.66 | <0.001 | |
| WRoot = aAb | A | 0.938 0 | 0.46 | 1.39 | 0.64 | 0.47 | <0.001 | |
| WStem = aV + b | V | 0.000 2 | 9.22 | 15.79 | 9.06 | 0.70 | <0.001 | |
| WLeaf = aVb | V | 0.016 0 | 0.55 | 3.89 | 0.63 | 0.60 | <0.001 | |
| 鬼箭锦鸡儿 Caragana jubata | WTotal = aA + b | A | 0.268 0 | -10.27 | 12.92 | 48.69 | 0.93 | <0.001 |
| WAboveground = aA + b | A | 0.159 0 | -3.65 | 16.51 | 38.04 | 0.88 | <0.001 | |
| WRoot = aA + b | A | 0.110 0 | -6.63 | 11.13 | 16.30 | 0.95 | <0.001 | |
| WStem = aA + b | A | 0.120 0 | -2.83 | 16.74 | 29.02 | 0.88 | <0.001 | |
| WLeaf = aA + b | A | 0.040 0 | -0.82 | 21.53 | 12.27 | 0.81 | <0.001 |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1芦芽山6种灌木总生物量最佳拟合模型。A, 刺玫。B, 蒙古荚蒾。C, 刺果茶藨子。D, 沙棘。E, 虎榛子。F, 鬼箭锦鸡儿。
-->Fig. 1The best biomass estimation model of six shrub species in Mountain Luya. A, Rosa davurica. B, Viburnum mongolicum. C, Ribes burejense. D, Hippophae rhamnoides. E, Ostryopsis davidiana. F, Caragana jubata.
-->
2.2 生物量分配
表3统计了芦芽山14种灌木的根冠比、叶质比、茎质比和根质比的基本特征(范围、均值和中位数)。整体而言, 芦芽山14种常见灌木的平均根冠比为0.61 (0.31-1.05), 平均叶质比为0.17 (0.05-0.44), 平均茎质比为0.48 (0.36-0.57), 平均根质比为0.35 (0.2-0.48)。其中, 对根冠比而言, 除金花忍冬(1.05)外, 其他13种灌木的平均根冠比都小于1。此外, 平均叶质比表现出最大的种间差异, 叶质比最大的复盆子(0.44)高出叶质比最小的土庄绣线菊(0.05)将近8倍; 平均茎质比的种间差异最小, 而平均根质比的种间差异居中。Table 3
表3
表314种灌木的根冠比、叶质比、茎质比及根质比的统计特征
Table 3The descriptive statistics of root to shoot mass ratio (R/S), leaf to total mass ratio (LMF), stem to total mass ratio (SMF), and root to total mass ratio (RMF) of 14 shrub species
| 物种 Species | 根冠比 R/S | 叶质比 LMF | 茎质比 SMF | 根质比 RMF | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 范围 Range | 平均值± 标准偏差 Mean ± SD | 中位值 Median | 范围 Range | 平均值± 标准偏差 Mean ± SD | 中位值 Median | 范围 Range | 平均值± 标准偏差 Mean ± SD | 中位值 Median | 范围 Range | 平均值± 标准偏差 Mean ± SD | 中位值 Median | ||||
| 沙棘 Hippophae rhamnoides | 0.06-0.59 | 0.31 ± 0.14 | 0.29 | 0.07-0.30 | 0.20 ± 0.06 | 0.20 | 0.39-0.79 | 0.57 ± 0.11 | 0.54 | 0.06-0.37 | 0.23 ± 0.08 | 0.22 | |||
| 三裂绣线菊 Spiraea trilobata | 0.32-1.24 | 0.57 ± 0.21 | 0.51 | 0.04-0.33 | 0.17 ± 0.09 | 0.15 | 0.25-0.67 | 0.48 ± 0.12 | 0.48 | 0.24-0.55 | 0.35 ± 0.07 | 0.34 | |||
| 黄刺玫 Rosa xanthina | 0.19-2.00 | 0.55 ± 0.34 | 0.45 | 0.09-0.29 | 0.19 ± 0.05 | 0.18 | 0.24-0.67 | 0.48 ± 0.09 | 0.47 | 0.16-0.67 | 0.33 ± 0.11 | 0.31 | |||
| 虎榛子 Ostryopsis davidiana | 0.16-0.97 | 0.42 ± 0.15 | 0.40 | 0.09-0.45 | 0.24 ± 0.08 | 0.23 | 0.35-0.58 | 0.47 ± 0.06 | 0.48 | 0.14-0.49 | 0.29 ± 0.07 | 0.29 | |||
| 榛 Corylus heterophylla | 0.28-1.24 | 0.53 ± 0.22 | 0.47 | 0.10-0.34 | 0.24 ± 0.07 | 0.23 | 0.26-0.56 | 0.43 ± 0.07 | 0.42 | 0.22-0.55 | 0.34 ± 0.08 | 0.32 | |||
| 土庄绣线菊 Spiraea pubescens | 0.45-1.42 | 0.72 ± 0.27 | 0.63 | 0.01-0.10 | 0.05 ± 0.03 | 0.05 | 0.37-0.67 | 0.54 ± 0.08 | 0.57 | 0.31-0.59 | 0.41 ± 0.08 | 0.39 | |||
| 金花忍冬 Lonicera chrysantha | 0.24-2.42 | 1.05 ± 0.54 | 0.98 | 0.05-0.26 | 0.10 ± 0.04 | 0.10 | 0.20-0.72 | 0.42 ± 0.13 | 0.39 | 0.19-0.71 | 0.48 ± 0.13 | 0.49 | |||
| 蒙古荚蒾 Viburnum mongolicum | 0.16-2.06 | 0.67 ± 0.45 | 0.54 | 0.04-0.23 | 0.10 ± 0.04 | 0.10 | 0.28-0.76 | 0.53 ± 0.11 | 0.54 | 0.14-0.67 | 0.37 ± 0.13 | 0.35 | |||
| 唐古特忍冬 Lonicera tangutica | 0.12-1.84 | 0.67 ± 0.38 | 0.54 | 0.06-0.24 | 0.13 ± 0.04 | 0.12 | 0.28-0.65 | 0.50 ± 0.10 | 0.52 | 0.11-0.65 | 0.37 ± 0.12 | 0.35 | |||
| 复盆子 Rubus idaeus | 0.11-0.54 | 0.26 ± 0.10 | 0.23 | 0.26-0.67 | 0.44 ± 0.10 | 0.43 | 0.17-0.61 | 0.36 ± 0.12 | 0.35 | 0.10-0.35 | 0.20 ± 0.06 | 0.19 | |||
| 银露梅 Potentilla glabra | 0.29-2.54 | 0.76 ± 0.46 | 0.64 | 0.04-0.21 | 0.11 ± 0.05 | 0.09 | 0.25-0.67 | 0.49 ± 0.11 | 0.51 | 0.22-0.72 | 0.41 ± 0.12 | 0.39 | |||
| 山刺玫 Rosa davurica | 0.36-1.29 | 0.71 ± 0.24 | 0.67 | 0.04-0.19 | 0.11 ± 0.04 | 0.10 | 0.37-0.62 | 0.49 ± 0.07 | 0.48 | 0.26-0.56 | 0.40 ± 0.08 | 0.4 | |||
| 刺果茶藨子 Ribes burejense | 0.27-1.89 | 0.69 ± 0.44 | 0.52 | 0.02-0.35 | 0.13 ± 0.07 | 0.11 | 0.24-0.67 | 0.49 ± 0.12 | 0.52 | 0.21-0.65 | 0.38 ± 0.13 | 0.34 | |||
| 鬼箭锦鸡儿 Caragana jubata | 0.16-1.54 | 0.60 ± 0.30 | 0.58 | 0.08-0.34 | 0.18 ± 0.07 | 0.17 | 0.26-0.68 | 0.47 ± 0.09 | 0.46 | 0.14-0.61 | 0.36 ± 0.10 | 0.37 | |||
| 整体 Total | 0.61 ± 0.38 | 0.17 ± 0.11 | 0.48 ± 0.11 | 0.35 ± 0.12 | |||||||||||
新窗口打开
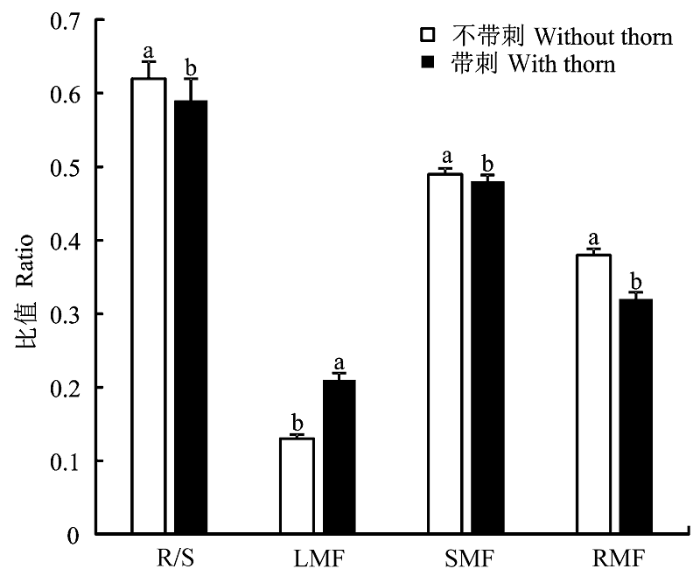
依照灌木是否带刺, 我们将14种灌木分为带刺(6种)与不带刺(8种)灌木两类。单因素方差分析表明, 不带刺灌木的根冠比、茎质比和根质比显著大于带刺灌木的根冠比、茎质比和根质比, 而其叶质比则显著小于带刺灌木的叶质比(图2)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2带刺与不带刺灌木之间叶质比(L/T)、茎质比(S/T)、根质比(R/T)、根冠比(R/S)的比较(平均值±标准误差)。用单因素方差分析(Tukey post hoc)检验不同类别之间的差异; 显著性差异(p < 0.05)用不同字母标记。
-->Fig. 2Comparisons of leaf biomass fraction (L/T), stem biomass fraction (S/T), root biomass fraction (R/T), root to shoot mass ratio (R/S) between shrubs with and without thorn (mean ± SE). Differences between each group were tested using a one-way ANOVA with a Tukey post hoc test of significance; significance different at p < 0.05 was indicated by different letters.
-->
3 讨论
3.1 生物量模型
曾珍英等(2005)在对灌木生物量的研究中发现, 用单个变量来模拟生物量回归方程是不理想的, 只有各个变量的适当组合才能比较准确地估算灌木生物量。卢振龙和龚孝生(2009)研究认为不同形态指标的组合代表了一定的物理意义, 例如树高(H)和树干直径(D)的乘积(H × D)可理解为树干纵断面周长, CH可以理解为投影纵断面积, 而树干直径平方和树高的乘积(D2H)可近似地理解为树干纵断面积。本研究表明, 不同灌木种的最优生物量估算模型不同, 而相同物种的各个生物量估测模型不论是方程的形式还是预测变量几乎都是一致的, 这与种与种之间的差异有关。A、V、Dt2H是拟合生物量方程的3个最佳测量指标。其中, 鬼箭锦鸡儿和银露梅的预测变量为A, 其外部形态低矮(两种灌木平均高度40 cm), 且分枝较多(两种灌木平均分蘖数4.8); 而沙棘、榛、山刺玫、金花忍冬其预测变量为Dt2H, 其外部形态直立且分枝数较少(平均树高和分蘖数分别为99 cm和3.5); 而其他灌木的最佳拟合变量多为V。曾慧卿等(2006)研究表明冠幅和高度的乘积能较为精确地估算檵木(Loropetalum chinense)的地上生物量, 同时也体现了檵木地上形态近似圆柱形的形态特征。由实际经验来看, 基径和树高相对于冠幅来说是两个测量误差较小、拟合方程效果较好的变量, 因此, 在预测生物量上常用这两个指标作为拟合变量(林伟等, 2010)。本研究中生物量分析所得模型均建立在研究区内的植株调查的基础上, 对于灌木种株高、基径调查范围以外或区域以外的应用需通过验证。3.2 生物量分配
在植物的整个生长发育过程中, 生长、繁殖和防御等各种功能对有限的资源始终存在着竞争问题。植物如何协调各种功能之间资源的分配, 即植物在生长过程中如何将有限的资源分配到根、茎、叶各器官中反映了植物的生存策略(张大勇, 2010)。在以往对生物量分配的研究中, 更多地集中在对乔木物种的研究, 对灌木种的研究较少(Pooter et al., 2012)。本研究中, 芦芽山的14种灌木除金花忍冬根冠比大于1外, 其余灌木种根冠比都小于1, 表明芦芽山灌木更多将光合产物分配到地上部分。这与Hilbert和Canadell (1995)的研究结果不同, 其发现地中海地区10种常绿灌木的平均根冠比为2.06。此外, 芦芽山14种灌木的平均叶质比为0.17, 茎质比为0.48, 都大于Hilbert和Canadell (1995)的中灌木的平均叶质比(0.07)和平均茎质比(0.31), 表明不同生活型(落叶与常绿)的灌木种在资源分配上的策略不同。刺是植物在与环境的相互作用过程中逐渐进化形成的一种性状特征, 是植物适应环境的一种外在表现。植物的刺有防止动物侵害、抵御干旱以及促进种子传播等功能(贺猛和米楷, 2009)。前人研究多表明干旱环境条件下的灌木种往往会出现带刺、根深等特殊的形态特征(易海燕, 2000), 说明带刺灌木更能在干旱的条件下生存。在我们调查的14种灌木中, 有6种灌木带刺, 可见刺是芦芽山常见灌木适应当地环境的一种性状特征。因此, 我们将芦芽山灌木按照带刺/不带刺分为两类, 发现不带刺灌木和带刺灌木在生物量分配上(根冠比、叶质比、茎质比和根质比)存在显著差异, 尤其是叶质比, 带刺灌木显著小于不带刺灌木, 这可能是带刺灌木对干旱环境适应的一种表现。这也与Pooter等(2012)的研究表明随着水分利用率的增加叶质比逐渐降低的结果相一致。
致谢 感谢山西芦芽山国家级自然保护区郭建荣副局长、宫树龙主任在野外调查中给予的协助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . . |
| [3] | . |
| [4] | . . |
| [5] | . |
| [6] | . . |
| [7] | . . |
| [8] | . In: |
| [9] | . . |
| [10] | . |
| [11] | . . |
| [12] | . . |
| [13] | . . |
| [14] | . . |
| [15] | . . |
| [16] | . |
| [17] | . |
| [18] | . |
| [19] | . . |
| [20] | . . |
| [21] | . . |
| [22] | . . |
| [23] | . . |
| [24] | . . |
| [25] | |
| [26] | . . |
| [27] | . . |
1
1985
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
千烟洲实验区几种灌木生物量估算模型的研究
1
2006
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
植物生物量分配模式与生长速率的相关规律研究
1
2007
... 生物量分配是表征植物生理过程的重要指标(
山西太岳山典型灌木林生物量及生产力研究
1
2002
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
Global allocation rules for patterns of biomass portioning in seed plants
1
2002
... 生物量分配是表征植物生理过程的重要指标(
北京东灵山三种温带森林生态系统的碳循环
1
2006
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
山东省常见带刺植物研究
1
2009
... 刺是植物在与环境的相互作用过程中逐渐进化形成的一种性状特征, 是植物适应环境的一种外在表现.植物的刺有防止动物侵害、抵御干旱以及促进种子传播等功能(
Biomass partitioning and resource allocation of plants from mediterranean-type ecosystems: Possible responses to elevated atmospheric CO2
1995
小叶锦鸡儿灌丛地上生物量的预测模式
1982
祁连山高山灌丛生物量沿海拔梯度分配特征研究
2012
黄土高原典型草原地根冠比的季节动态及其影响因素
1
2012
... 生物量分配是表征植物生理过程的重要指标(
井冈山自然保护区12种常见灌木生物量的估测模型
2
2010
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
... 曾珍英等(2005)在对灌木生物量的研究中发现, 用单个变量来模拟生物量回归方程是不理想的, 只有各个变量的适当组合才能比较准确地估算灌木生物量.卢振龙和龚孝生(2009)研究认为不同形态指标的组合代表了一定的物理意义, 例如树高(H)和树干直径(D)的乘积(H × D)可理解为树干纵断面周长, CH可以理解为投影纵断面积, 而树干直径平方和树高的乘积(D2H)可近似地理解为树干纵断面积.本研究表明, 不同灌木种的最优生物量估算模型不同, 而相同物种的各个生物量估测模型不论是方程的形式还是预测变量几乎都是一致的, 这与种与种之间的差异有关.A、V、Dt2H是拟合生物量方程的3个最佳测量指标.其中, 鬼箭锦鸡儿和银露梅的预测变量为A, 其外部形态低矮(两种灌木平均高度40 cm), 且分枝较多(两种灌木平均分蘖数4.8); 而沙棘、榛、山刺玫、金花忍冬其预测变量为Dt2H, 其外部形态直立且分枝数较少(平均树高和分蘖数分别为99 cm和3.5); 而其他灌木的最佳拟合变量多为V.曾慧卿等(2006)研究表明冠幅和高度的乘积能较为精确地估算檵木(Loropetalum chinense)的地上生物量, 同时也体现了檵木地上形态近似圆柱形的形态特征.由实际经验来看, 基径和树高相对于冠幅来说是两个测量误差较小、拟合方程效果较好的变量, 因此, 在预测生物量上常用这两个指标作为拟合变量(
贵州普定喀斯特次生林乔灌层地上生物量
1
2009
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
巴郎山川滇高山栎灌丛地上生物量及其对海拔梯度的响应
2006
灌木生物量测定的研究进展
2009
Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control
2
2012
... 生物量分配是表征植物生理过程的重要指标(
... 在植物的整个生长发育过程中, 生长、繁殖和防御等各种功能对有限的资源始终存在着竞争问题.植物如何协调各种功能之间资源的分配, 即植物在生长过程中如何将有限的资源分配到根、茎、叶各器官中反映了植物的生存策略(
Estimating the aboveground biomass and wood volume of savanna woodlands in Brazil’s Pantanal wetlands based on allometric correlations
2006
Net production relations of shrubs in the Great Smoky Mountains
1982
岷江上游山地森林-干旱河谷交错带不同植被类型土壤微生物及土壤酶活性的研究
2
2010
... 目前关于生物量分配的研究多集中于乔木, 而有关灌木生物量分配的研究较少.雷蕾(2012)对祁连山高山灌丛生物量分配有过研究, 刘兴良等(2006)探讨过川滇高山栎(Quercus aquifolioides)灌丛地上生物量分配的特征.相比于乔木, 灌木所处的环境往往较为干旱、贫瘠.为适应所处环境, 灌木往往会呈现带刺、根深等特殊的形态特征(
... 刺是植物在与环境的相互作用过程中逐渐进化形成的一种性状特征, 是植物适应环境的一种外在表现.植物的刺有防止动物侵害、抵御干旱以及促进种子传播等功能(
灌木林主要生态作用的探讨
1
2003
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
基于冠幅及植株高度的檵木生物量回归模型
4
2006
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
...
... ).通过数理分析得到的最佳生物量方程不仅为灌木生物量的连续监测提供了可能, 而且可用于对类似环境灌木生物量的估算(
...
红壤丘陵区林下灌木生物量估算模型的建立及其应用
1
2007
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
江西千烟洲几种灌木生物量模型的研究
2005
立木生物量方程的优度评价和精度分析
1
2011
... 其中W表示预测变量(本文中指各类生物量), X表示拟合变量(如株高、冠幅等), a、b表示拟合参数.根据决定系数(R2)、估计值的标准误差(SEE)以及平均预估误差(MPE)的大小及回归检验显著水平(p值)来评价模型的优劣(
1
2010
... 在植物的整个生长发育过程中, 生长、繁殖和防御等各种功能对有限的资源始终存在着竞争问题.植物如何协调各种功能之间资源的分配, 即植物在生长过程中如何将有限的资源分配到根、茎、叶各器官中反映了植物的生存策略(
千烟洲试验站几种常见灌木生物量估测
1
2005
... 生物量是生态系统生产力的主要组成成分, 对生态系统功能体现和群落结构形成都起着重要作用(
山西芦芽山植被垂直带的划分
1
1989
... 山西芦芽山国家级自然保护区位于山西省吕梁山脉北端, 地处宁武、五寨、岢岚三县的交界处, 111.83°-112.09° E, 38.59°-38.75° N, 区内最高海拔为主峰芦芽山2787 m, 最低海拔1346 m, 年平均气温4-7 ℃, 年降水量500-600 mm, 无霜期90-120天.该区土壤分布复杂, 区域东部是黄土母质发育而成的灰褐土, 西部以褐土为主.随海拔高度的升高, 土壤呈现垂直地带性, 依次为山地褐土、山地淋溶褐土、棕色森林土和亚高山草甸土.芦芽山植被具有明显的垂直地带性, 从山麓到山顶依次为森林草原带、落叶阔叶林带、针阔叶混交林带、寒温性针叶林带、亚高山灌丛草甸带.芦芽山东麓西麓基带不同, 东麓基带为森林草原带, 西麓基带为落叶阔叶林带.沿海拔梯度, 保护区内优势的灌丛群落主要有沙棘(Hippophae rhamnoides)灌丛、山刺玫(Rosa davurica)灌丛、绣线菊(Spiraea spp.)灌丛、虎榛子(Ostryopsis davidiana)灌丛、蒙古荚蒾(Viburnum mongolicum)灌丛、金花忍冬(Lonicera chrysantha)灌丛、银露梅(Potentilla glabra)灌丛和鬼箭锦鸡儿(Caragana jubata)灌丛(


{kind=link}
{kind=link}
{kind=link}
{kind=link}