Interactive effects of ozone and drought stress on plants: A review
GAOFeng1,2, LIPin1, FENGZhao-Zhong1,2,*, 1State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China2University of Chinese Academy of Sciences, Beijing 100049, China 通讯作者:* 通信作者Author for correspondence (E-mail:fzz@rcees.ac.cn) 收稿日期:2016-06-12 接受日期:2016-12-16 网络出版日期:2017-02-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院“****”项目和国家自然科学基金(31500396)
关键词:臭氧;干旱;交互;植物;响应 Abstract Ground-level ozone (O3) and drought are two key factors limiting plant growth. O3 can enter into the plant tissue through the stomata, then causing the formation of reactive oxygen species (ROS) which inspires programmed cell death. Drought usually induces the accumulation of ROS due to damage to antioxidant systems of plants. The effects of two kinds of stress on plants are similar due to the accumulation of ROS, resulting in reduced photosynthesis rate and physiological metabolism, eventually decreased plant growth and biomass. Nevertheless, O3 and drought interacts synergistically to accumulate detrimental effects or antagonistically to reduce harmful effects. Actually, it is complex interactive process between O3 and drought. On the one hand, O3 triggers stomatal sluggishness or even dysfunction, which exacerbates water transpiration of leaves, water loss from plants and further O3 phytotoxicity. On the other hand, drought induces stomatal closure, and thus protecting plants against the O3 influx and evaporation of water. However, prolonged drought could limit the uptake of CO2 and thus result in reduced plant growth. The response of plants to both O3 and drought not only depends on the occurring sequence and duration of any factor but also rely on the difference in physiological metabolism of the plant itself. The interactive effects of O3 and drought on stomatal characteristics, photosynthetic carbon mechanism, antioxidant response and growth development are reviewed in this paper and the aspects to be further studied are also suggested.
0, 10%, 25%, 50%的年降水量减少 0, 10%, 25%, 50% Reductions in total annual precipitation
模型模拟胁迫下生长 Simulation growth by model under stress
Retzlaff et al., 2000
Pinus halepensis
2
OTC
69
50%对照灌溉水量 Half the water supplied to control
叶片抗氧化酶类 Antioxidative enzymes in needles
Alonso et al., 2001
Pinus halepensis
4
GC
300
以叶片水势和气体交换临界值确定灌水量 Characterized by very low leaf water potential and gas exchange values
叶片生理 Physiological response of needles
Manes et al., 2001
植物类型 Plant type
植物种类 Plants species
树龄(年) Tree-age (year)
实验方法Experimental method
O3处理 O3 treatment (nmol·mol-1)
干旱处理 Drought treatment
研究内容 Research content
参考文献 Reference
常绿乔木 Evergreen trees
Pinus halepensis
3
GC
200
100%-50%缓慢水分散失(中度); 以黎明前叶片水势确定灌溉水量(重度) From 100% to 50% of the water loss by evapotranspiration (mild); watered by pre-dawn needle water potential (intensity)
叶片酶(PEPC) Phosphoenolpyruvatecarboxylase in needles
Fontaine et al., 2003
Pinus halepensis
2
OTC
AA + 40
50%对照灌溉水量 Half the water supplied to control
生理生化补偿过程 Compensation processes in physiology and biochemistry
Inclan et al., 2005
Quercus ilex
3
气候室 Climatic chambers
250
20 mL水每间隔一周灌溉 20 mL of water per plant every week
生理响应 Physiological responses
Vitale et al., 2008
Quercus ilex ssp. ilex, 长角豆 Ceratonia siliqua
1
OTC
37; 57
50%对照灌溉次数 Half the watered times to control
生理生化 Physiology and biochemistry
Ribas et al., 2005
Quercus ilex ssp. ilex, Q. ilex ssp. ballota
1-2
OTC
31; 49
50%对照灌溉水量 Half the water supplied to control
气体交换、生长和生物量 Gas exchange, growth and biomass
Alonso et al., 2014
西黄松 Pinus ponderosa
6-8
NE
40-70
NE
O3和碳吸收 O3 and carbon uptake
Panek & Goldstein, 2001
西黄松 Pinus ponderosa
40
NE
58-60, 64-67, 80
黎明前木质部水势< -1.2 MPa Xylem potentials <-1.2 MPa at predawn
碳获取 Carbon acquisition
Grulke et al., 2002
西黄松 Pinus ponderosa
5-7
NE
53
NE (2年不同气候) NE (Climatic variation in two years)
O3暴露度量参数 O3 exposure metrics parameters
Panek et al., 2002
西黄松 Pinus ponderosa
7
NE
AA
NE (4个不同地点) NE (Four sites)
吸收模型参数 Parameters for O3 uptake modeling
Panek, 2004
落叶乔木Deciduous trees
海南蒲桃 Syzygium hainanense, 糖胶树 Alstonia scholaris
当年 Current year
OTC
75
40%-50%土壤相对含水量 40%-50% Relative soil water content
AbramsMD, KubiskeME, MostollerSA (1994). Relating wet and dry year ecophysiology to leaf structure in contrastingtemperate tree species. Ecology, 75, 123-133. [本文引用: 1]
[2]
AlonsoR, ElviraS, CastilloFJ, GimenoBS (2001). Interactive effects of ozone and drought stress on pigments and activities of antioxidative enzymes in Pinus halepensis. Plant, Cell & Environment, 24, 905-916. [本文引用: 6]
[3]
AlonsoR, ElviraS, González-FernándezI, CalveteH, García- GómezH, BermejoV (2014). Drought stress does not protect Quercus ilex L. from ozone effects: Results from a comparative study of two subspecies differing in ozone sensitivity. Plant Biology, 16, 375-384. [本文引用: 1]
[4]
AnjumF, YaseenM, RasulE, WahidA, AnjumS (2003). Water stress in barley (Hordeum vulgare L.). II. Effect on chemical composition and chlorophyll contents. Pakistan Journal of Agricultural Sciences, 40(1-2), 45-49. [本文引用: 1]
[5]
AsadaK (1997). The role of ascorbate peroxidase and monodehydroascorbate reductase in H2O2 scavenging in plants. Oxidatives Stress and the Molecular Biology of Antioxidant Defenses, 34, 715-735. [本文引用: 1]
[6]
AshmoreMR (2005). Assessing the future global impacts of ozone on vegetation. Plant, Cell & Environment, 28, 949-964. [本文引用: 1]
[7]
AvneryS, MauzerallDL, LiuJF, HorowitzLW (2011). Global crop yield reductions due to surface ozone exposure: 2. Year 2030 potential crop production losses and economic damage under two scenarios of O3 pollution. Atmospheric Environment, 45, 2297-2309. [本文引用: 1]
[8]
BiehlerK, FockH (1996). Evidence for the contribution of the Mehler-peroxidase reaction in dissipating excess electrons in drought-stressed wheat. Plant Physiology, 112, 265-272. [本文引用: 1]
[9]
BiswasDK, JiangGM (2011). Differential drought-induced modulation of ozone tolerance in winter wheat species. Journal of Experimental Botany, 62, 4153-4162. [本文引用: 6]
[10]
BohlerS, BagardM, OufirM, PlanchonS, HoffmannL, JolivetY, HausmanJ-F, DizengremelP, RenautJ (2007). A DIGE analysis of developing poplar leaves subjected to ozone reveals major changes in carbon metabolism. Proteomics, 7, 1584-1599. [本文引用: 2]
[11]
BohlerS, SergeantK, HoffmannL, DizengremelP, HausmanJ-F, RenautJ, JolivetY (2011). A difference gel electrophoresis study on thylakoids isolated from poplar leaves reveals a negative impact of ozone exposure on membrane proteins. Journal of Proteome Research, 10, 3003-3011. [本文引用: 1]
[12]
BohlerS, SergeantK, JolivetY, HoffmannL, HausmanJ-F, DizengremelP, RenautJ (2013). A physiological and proteomic study of poplar leaves during ozone exposure combined with mild drought. Proteomics, 13, 1737-1754. [本文引用: 6]
[13]
BohlerS, SergeantK, LefevreI, JolivetY, HoffmannL, RenautJ, DizengremelP, HausmanJ-F (2010). Differential impact of chronic ozone exposure on expanding and fully expanded poplar leaves. Tree Physiology, 30, 1415-1432. [本文引用: 1]
[14]
BotaJ, MedranoH, FlexasJ (2004). Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New phytologist, 162, 671-681. [本文引用: 1]
[15]
BrédaN, HucR, GranierA, DreyerE (2006). Temperate forest trees and stands under severe drought: A review of eco- physiological responses, adaptation processes and long- term consequences. Annals of Forest Science, 63, 625-644. [本文引用: 1]
[16]
BrendleyBW, PellEJ (1998). Ozone-induced changes in biosynthesis of Rubisco and associated compensation to stress in foliage of hybrid poplar. Tree Physiology, 18, 81-90. [本文引用: 1]
[17]
BroadmeadowM (1998). Ozone and forest trees. New Phytologist, 139, 123-125. [本文引用: 1]
[18]
BucklandSM, PriceAH, HendryGAF (1991). The role of ascorbate in drought-treated Cochlearia atlantica Pobed. and Armeria maritima (Mill.) Willd. New Phytologist, 119, 155-160. [本文引用: 1]
[19]
ChangSC, LeeCT (2006). Ozone variations through vehicle emissions reductions based on air quality monitoring data in Taipei City, Taiwan, from 1994 to 2003. Atmospheric Environment, 40, 3513-3526. [本文引用: 1]
[20]
ChenZ, GallieDR (2005). Increasing tolerance to ozone by elevating foliar ascorbic acid confers greater protection against ozone than increasing avoidance. Plant Physiol- ogy, 138, 1673-1689. [本文引用: 2]
[21]
CiaisP, ReichsteinM, ViovyN, GranierA, OgeeJ, AllardV, AubinetM, BuchmannN, BernhoferC, CarraraA, ChevallierF, de NobletN, FriendAD, FriedlingsteinP, GrunwaldT, HeineschB, KeronenP, KnohlA, KrinnerG, LoustauD, MancaG, MatteucciG, MigliettaF, OurcivalJM, PapaleD, PilegaardK, RambalS, SeufertG, SoussanaJF, SanzMJ, SchulzeED, VesalaT, ValentiniR (2005). Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature, 437, 529-533.
[22]
ComteDL (1998). Weather highlights around the world. Weatherwise, 51(2), 26-31. [本文引用: 1]
[23]
ConklinPL, BarthC (2004). Ascorbic acid, a familiar small molecule intertwined in the response of plants to ozone, pathogens, and the onset of senescence. Plant, Cell & Environment, 27, 959-970. [本文引用: 1]
CreissenGP, MullineauxPM (2002). The molecular biology of the ascorbate-glutathione cycle in higher plants. In: Inzé D, Montagu MV eds. Oxidative Stress in Plants. Taylor & Francis, Abingdon, UK. 247-270. [本文引用: 1]
[26]
DaiA (2011). Drought under global warming: A review. Climate Change, 2(1), 45-65. [本文引用: 1]
[27]
DesotgiuR, PollastriniM, CascioC, GerosaG, MarzuoliR, BussottiF (2012). Chlorophyll a fluorescence analysis along a vertical gradient of the crown in a poplar (Oxford clone) subjected to ozone and water stress. Tree Physiology, 32, 976-986.
[28]
D’HaeseD, VandermeirenK, AsardH, HoremansN (2005). Other factors than apoplastic ascorbate contribute to the differential ozone tolerance of two clones of Trifolium repens L. Plant, Cell & Environment, 28, 623-632. [本文引用: 1]
[29]
DizengremelP (2001). Effects of ozone on the carbon metabolism of forest trees. Plant Physiology and Biochemistry, 39, 729-742. [本文引用: 1]
[30]
DizengremelP, JolivetY, TuzetA, RanieriA, Le ThiecD (2013). Integrative leaf-level phytotoxic ozone dose assessment for forest risk modelling. Developments in Environmental Science, 13, 267-288. [本文引用: 1]
[31]
DizengremelP, Le ThiecD, BagardM, JolivetY (2008). Ozone risk assessment for plants: Central role of metabolism-dependent changes in reducing power. Environmental Pollution, 156, 11-15. [本文引用: 2]
[32]
DizengremelP, Le ThiecD, Hasenfratz-SauderMP, VaultierMN, BagardM, JolivetY (2009). Metabolic-dependent changes in plant cell redox power after ozone exposure. Plant Biology, 11(Suppl. 1), 35-42. [本文引用: 1]
[33]
DumontJ, SpicherF, MontpiedP, DizengremelP, JolivetY, Le ThiecD (2013). Effects of ozone on stomatal responses to environmental parameters (blue light, red light, CO2 and vapour pressure deficit) in three Populus deltoides × Populus nigra genotypes. Environmental Pollution, 173, 85-96. [本文引用: 1]
[34]
EdwardsIP, ZakDR (2011). Fungal community composition and function after long-term exposure of northern forests to elevated atmospheric CO2 and tropospheric O3. Global Change Biology, 17, 2184-2195. [本文引用: 1]
[35]
EltayebAE, KawanoN, BadawiGH, KaminakaH, SanekataT, MorishimaI, ShibaharaT, InanagaS, TanakaK (2006). Enhanced tolerance to ozone and drought stresses in transgenic tobacco overexpressing dehydroascorbate reductase in cytosol. Physiologia Plantarum, 127, 57-65. [本文引用: 1]
[36]
ErnstD, JürgensenM, BahnwegG, HellerW, Müller-StarckG (2012). Common links of molecular biology with biochemistry and physiology in plants under ozone and pathogen attack. In: Matyssek R, Schnyder H, Oßwald W, Ernst D, Munch CJ, Pretzsch H eds. Growth and Defence in Plants: Resource Allocation at Multiple Scales. Springer, Berlin. 29-51. [本文引用: 1]
[37]
FedinaIS, PopovaAV (1996). Photosynthesis, photorespiration and proline accumulation in water-stressed pea leaves. Photosynthetica, 32, 213-220. [本文引用: 1]
[38]
FengYW, ShimizuH (2005). Effects of ozone and/or water stress on the growth of Abies veitchii seedlings. Phyton- Annales Rei Botanicae, 45, 591-594.
[39]
FengZZ, SunJS, WanWX, HuEZ, CalatayudV (2014). Evidence of widespread ozone-induced visible injury on plants in Beijing, China. Environmental Pollution, 193, 296-301. [本文引用: 1]
[40]
FengZZ, KobayashiK (2009). Assessing the impacts of current and future concentrations of surface ozone on crop yield with meta-analysis. Atmospheric Environment, 43, 1510-1519. [本文引用: 1]
[41]
FlexasJ, BotaJ, EscalonaJM, SampolB, MedranoH (2002). Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Functional Plant Biology, 29, 461-471. [本文引用: 3]
[42]
FontaineV, CabaneM, DizengremelP (2003). Regulation of phosphoenolpyruvate carboxylase in Pinus halepensis needles submitted to ozone and water stress. Physiologia Plantarum, 117, 445-452. [本文引用: 1]
[43]
FoyerCH, NoctorG (2000). Oxygen processing in photosynthesis: Regulation and signalling. New Phytologist, 146, 359-388. [本文引用: 1]
[44]
FuJM, HuangBR (2001). Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environmental and Experimental Botany, 45, 105-114. [本文引用: 1]
[45]
GerantD, PodorM, GrieuP, AfifD, CornuS, MorabitoD, BanvoyJ, RobinC, DizengremelP (1996). Carbon metabolism enzyme activities and carbon partitioning in Pinus halepensis Mill. exposed to mild drought and ozone. Journal of Plant Physiology, 148, 142-147. [本文引用: 1]
[46]
GrulkeNE, JohnsonR, EsperanzaA, JonesD, NguyenT, PoschS, TauszM (2003a). Canopy transpiration of Jeffrey pine in mesic and xeric microsites: O3 uptake and injury response. Trees, 17, 292-298. [本文引用: 1]
[47]
GrulkeNE, JohnsonR, MonscheinS, NikolovaP, TauszM (2003b). Variation in morphological and biochemical O3 injury attributes of mature Jeffrey pine within canopies and between microsites. Tree Physiology, 23, 923-929.
[48]
GrulkeNE, PreislerHK, RoseC, KirschJ, BaldumanL (2002). O3 uptake and drought stress effects on carbon acquisition of ponderosa pine in natural stands. New Phytologist, 154, 621-631. [本文引用: 1]
[49]
GrünhageL, JagerHJ (2003). From critical levels to critical loads for ozone: A discussion of a new experimental and modelling approach for establishing flux-response relationships for agricultural crops and native plant species. Environmental Pollution, 125, 99-110. [本文引用: 1]
[50]
GuidiL, NaliC, LorenziniG, FilippiF, SoldatiniGF (2001). Effect of chronic ozone fumigation on the photosynthetic process of poplar clones showing different sensitivity. Environmental Pollution, 113, 245-254. [本文引用: 1]
[51]
GuptaP, DuplessisS, WhiteH, KarnoskyDF, MartinF, PodilaGK (2005). Gene expression patterns of trembling aspen trees following long-term exposure to interacting elevated CO2 and tropospheric O3. New Phytologist, 167, 129-142. [本文引用: 1]
[52]
HabererK, HerbingerK, AlexouM, RennenbergH, TauszM (2008). Effects of drought and canopy ozone exposure on antioxidants in fine roots of mature European beech (Fagus sylvatica). Tree Physiology, 28, 713-719.
[53]
HaoYT, LinM, XueL, WangZY, LinJT, LiangZY, SunBC, TianMT (2014). Effects of ozone stress and drought stress on photosynthesis characteristics of Syzygium hainanense and Alstonia scholaris seedlings. Journal of Anhui Agricultural University, 41, 193-197. (in Chinese with English abstract)[郝云亭, 林敏, 薛立, 王志云, 林婧庭, 梁梓毅, 孙冰超, 田茂涛 (2014). 臭氧与干旱胁迫对海南蒲桃和盆架子幼苗光合生理的影响 . 安徽农业大学学报, 41, 193-197.]
[54]
HatchM, SlackC, BullT (1969). Light-induced changes in the content of some enzymes of the C4-dicarboxylic acid pathway of photosynthesis and its effect on other characteristics of photosynthesis. Phytochemistry, 8, 697-706. [本文引用: 1]
[55]
HeathRL (1994). Alterations of plant metabolism by ozone exposure. In: Alscher RG, Wellburn AR eds. Plant Responses to the Gaseous Environment: Molecular, Metabolic and Physiological aspects. Springer Netherlands, Dordrecht. 121-145. [本文引用: 2]
[56]
HeathRL (2008). Modification of the biochemical pathways of plants induced by ozone: What are the varied routes to change? Environmental Pollution, 155, 453-463. [本文引用: 1]
[57]
HeathRL, TaylorGE (1997). Physiological processes and plant responses to ozone exposure. In: Sandermann H, Wellburn AR, Heath RL eds. Forest Decline and Ozone: A Comparison of Controlled Chamber and Field Experiments. Springer, Berlin. 317-368.
[58]
HeberU, KaiserW, LuweM, KindermannG, Veljovic- JavonovicS, YinZ, PfanzH, SlovikS (1997). Air pollution, photosynthesis and forest decline: Interactions and consequences. In: Schulze E-D, Caldwell MM eds. Ecophysiology of Photosynthesis. Springer, Berlin. 279-296. [本文引用: 1]
[59]
HerbingerK, TauszM, WonischA, SojaG, SorgerA, GrillD (2002). Complex interactive effects of drought and ozone stress on the antioxidant defence systems of two wheat cultivars. Plant Physiology and Biochemistry, 40, 691-696.
[60]
HerbingerK, ThenC, LowM, HabererK, AlexousM, KochN, RemeleK, HeerdtC, GrillD, RennenbergH, HaberleKH, MatyssekR, TauszM, WieserG (2005). Tree age dependence and within-canopy variation of leaf gas exchange and antioxidative defence in Fagus sylvatica under experimental free-air ozone exposure. Environmental Pollution, 137, 476-482. [本文引用: 1]
[61]
HoshikaY, CarrieroG, FengZZ, ZhangYL, PaolettiE (2014). Determinants of stomatal sluggishness in ozone-exposed deciduous tree species. Science of the Total Environment, 481, 453-458. [本文引用: 3]
[62]
HoshikaY, OmasaK, PaolettiE (2013). Both ozone exposure and soil water stress are able to induce stomatal sluggishness. Environmental and Experimental Botany, 88, 19-23.
HuangBR, FuJM (2000). Photosynthesis, respiration, and carbon allocation of two cool-season perennial grasses in response to surface soil drying. Plant and Soil, 227(1-2), 17-26. [本文引用: 1]
[65]
HuseynovaIM, AliyevaDR, AliyevJA (2014). Subcellular localization and responses of superoxide dismutase isoforms in local wheat varieties subjected to continuous soil drought. Plant Physiology and Biochemistry, 81, 54-60. [本文引用: 1]
[66]
ICPVegetation (2014). Ozone pollution: A hidden threat to food security. Programme Coordination Centre for the ICP Vegetation. In: Mills G, Harmens H eds. Programme Coordination Centre for the ICP Vegetation. NERC/Centre for Ecology and Hydrology, Bangor, UK. 116. [本文引用: 3]
[67]
InclanR, GimenoBS, DizengremelP, SanchezM (2005). Compensation processes of Aleppo pine (Pinus halepensis Mill.) to ozone exposure and drought stress. Environmental Pollution, 137, 517-524. [本文引用: 2]
[68]
IPCC (Intergovernmental Panel on Climate Change) (2013). Climate Change 2013: The Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK and New York, USA. 1552. [本文引用: 4]
[69]
IyerNJ, TangY, MahalingamR (2013). Physiological, biochemical and molecular responses to a combination of drought and ozone in Medicago truncatula. Plant, Cell & Environment, 36, 706-720. [本文引用: 1]
[70]
KarlssonPE, MedinEL, WickstromH, SelldenG, WallinG, OttossonS, SkarbyL (1995). Ozone and drought stress— Interactive effects on the growth and physiology of Norway spruce (Picea abies (L) Karst). Water, Air, & Soil Pollution, 85, 1325-1330. [本文引用: 1]
[71]
KarlssonPE, PleijelH, KarlssonGP, MedinEL, SkarbyL (2000). Simulations of stomatal conductance and ozone uptake to Norway spruce saplings in open-top chambers. Environmental Pollution, 109, 443-451. [本文引用: 1]
[72]
KarnoskyDF, PregitzerKS, ZakDR, KubiskeME, HendreyGR, WeinsteinD, NosalM, PercyKE (2005). Scaling ozone responses of forest trees to the ecosystem level in a changing climate. Plant, Cell & Environment, 28, 965-981. [本文引用: 1]
[73]
KarnoskyDF, WernerH, HolopainenT, PercyK, OksanenT, OksanenE, HeerdtC, FabianP, NagyJ, HeilmanW, CoxR, NelsonN, MatyssekR (2007). Free-air exposure systems to scale up ozone research to mature trees. Plant Biology, 9, 181-190. [本文引用: 1]
[74]
KayaMD, OkcuG, AtakM, CikiliY, KolsariciO (2006). Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). European Journal of Agronomy, 24, 291-295.
[75]
KhanS, SojaG (2003). Yield responses of wheat to ozone exposure as modified by drought-induced differences in ozone uptake. Water, Air, & Soil Pollution, 147, 299-315.
[76]
KivimäenpääM, SutinenS, KarlssonPE, SelldenG (2003). Cell structural changes in the needles of Norway spruce exposed to long-term ozone and drought. Annals of Botany, 92, 779-793.
[77]
KivimäenpääM, SutinenS, MedinEL, KarlssonPE, SelldenG (2001). Diurnal changes in microscopic structures of mesophyll cells of Norway spruce, Picea abies (L.) Karst., and the effects of ozone and drought. Annals of Botany, 88, 119-130.
[78]
KouTJ, ChangHQ, ZhangLH, XuXF, GuoDY, ZhouWL, ZhuJG, MiaoYF (2009). Effects of ground-level O3 pollution on the terrestrial ecosystem . Ecology and Environmental Sciences, 18, 704-710. (in Chinese with English abstract) [寇太记, 常会庆, 张联合, 徐晓峰, 郭大勇, 周文利, 朱建国, 苗艳芳 (2009). 近地层O3污染对陆地生态系统的影响 . 生态环境学报, 18, 704-710.] [本文引用: 2]
[79]
KronfußG, PolleA, TauszM, HavranekWM, WieserG (1998). Effects of ozone and mild drought stress on gas exchange, antioxidants and chloroplast pigments in current-year needles of young Norway spruce [Picea abies (L.) Karst]. Trees, 12, 482-489.
[80]
KuehnAR, GrillS, BaumgartenM, AnkerstDP, MatyssekR (2015). Daily growth of European beech (Fagus sylvatica L.) on moist sites is affected by short-term drought rather than ozone uptake. Trees, 29, 1501-1519. [本文引用: 1]
Le ThiecD, DixonM, GarrecJP (1994). The effects of slightly elevated ozone concentrations and mild drought stress on the physiology and growth of Norway Spruce, Picea abies (L.) Karst. and beech, Fagus sylvatica L., in open-top chambers. New Phytologist, 128, 671-678. [本文引用: 1]
[83]
LiL, ChenCH, HuangC, HuangHY, ZhangGF, WangYJ, ChenMH, WangHL, ChenYR, StreetsDG, FuJM (2011). Ozone sensitivity analysis with the MM5-CMAQ modeling system for Shanghai. Journal of Environmental Sciences, 23, 1150-1157.
[84]
LiL, ManningWJ, TongL, WangXK (2015). Chronic drought stress reduced but not protected Shantung maple (Acer truncatum Bunge) from adverse effects of ozone (O3) on growth and physiology in the suburb of Beijing, China. Environmental Pollution, 201, 34-41.
[85]
LiQJ, LuGC, XueL, TianMT, LinM, LinJT (2015). Effects of ozone and drought on fluorescence physiology of seedlings of three afforestation tree species in South China. Journal of South China Agricultural University, 36, 91-95. (in Chinese with English abstract)[李秋静, 卢广超, 薛立, 田茂涛, 林敏, 林婧庭 (2011). 臭氧与干旱胁迫对华南地区3种绿化树种 . 华南农业大学学报, 36, 91-95.]
[86]
LöwM, HerbingerK, NunnAJ, HaeberleKH, LeuchnerM, HeerdtC, WernerH, WipflerP, PretzschH, TauszM, MatyssekR (2006). Extraordinary drought of 2003 overrules ozone impact on adult beech trees (Fagus sylvatica). Trees, 20, 539-548. [本文引用: 1]
[87]
LutzC, AneggS, GerantD, Alaoui-SosseB, GerardJ, DizengremelP (2000). Beech trees exposed to high CO2 and to simulated summer ozone levels: Effects on photosynthesis, chloroplast components and leaf enzyme activity. Physiologia Plantarum, 109, 252-259. [本文引用: 1]
[88]
LuweMWF, TakahamaU, HeberU (1993). Role of ascorbate in detoxifying ozone in the apoplast of spinach (Spinacia oleracea L.) leaves. Plant Physiology, 101, 969-976. [本文引用: 1]
[89]
Maier-MaerckerU (1998). Image analysis of the stomatal cell walls of Picea abies (L.) Karst. in pure and ozone-enriched air. Trees, 12, 181-185. [本文引用: 1]
[90]
ManderscheidR, JägerH-J, SchoenebergerM (1991). Dose- response relationships of ozone effects on foliar levels of antioxidants, soluble polyamines and peroxidase activity of Pinus taeda (L.): Assessment of the usefulness as early ozone indicators. Angewandte Botanik, 29, 188.
[91]
ManesF, DonatoE, VitaleM (2001). Physiological response of Pinus halepensis needles under ozone and water stress conditions. Physiologia Plantarum, 113, 249-257. [本文引用: 1]
[92]
MatyssekR, InnesJL (1999). Ozone—A risk factor for trees and forests in Europe? Water, Air, & Soil Pollution, 116, 199-226.
[93]
MatyssekR, KozovitsAR, SchnitzlerJ-P, PretzschH, DielerJ, WieserG (2014). Forest trees under air pollution as a factor of climate change. In: Tausz M, Grulke N eds. Trees in a Changing Environment: Ecophysiology, Adaptation, and Future Survival. Springer, Dordrecht, The Netherlands. 117-163. [本文引用: 3]
[94]
MatyssekR, Le ThiecD, LöwM, DizengremelP, NunnAJ, HäberleKH (2006). Interactions between drought and O3 stress in forest trees. Plant Biology, 8, 11-17. [本文引用: 1]
[95]
MatyssekR, SandermannH (2003). Impact of ozone on trees: An ecophysiological perspective. Progress in Botany, 64, 349-404. [本文引用: 1]
[96]
MatyssekR, WieserG, CalfapietraC, de VriesW, DizengremelP, ErnstD, JolivetY, MikkelsenTN, MohrenGMJ, Le ThiecD, TuovinenJP, WeatherallA, PaolettiE (2012). Forests under climate change and air pollution: Gaps in understanding and future directions for research. Environmental Pollution, 160, 57-65. [本文引用: 2]
McLaughlinSB, NosalM, WullschlegerSD, SunG (2007). Interactive effects of ozone and climate on tree growth and water use in a southern Appalachian forest in the USA. New Phytologist, 174, 109-124.
[99]
MillerPR, McBride JR (1999). Oxidant air pollution impacts in the montane forests of southern California—A case study of the San Bernardino Mountains—Introduction. In: Miller PR, McBride JR eds. Oxidant Air Pollution Impacts in the Montane Forests of Southern California. Springer, New York. [本文引用: 2]
[100]
MillsG, HayesF, SimpsonD, EmbersonL, NorrisD, HarmensH, BukerP (2011). Evidence of widespread effects of ozone on crops and (semi-) natural vegetation in Europe (1990-2006) in relation to AOT40- and flux-based risk maps. Global Change Biology, 17, 592-613.
[101]
Ministry of Water Resources, China (2006-2010). Bulletin of Flood and Drought Disasters in China . . Cited: 2016-6.(in Chinese)[中国水利部 (2006-2010). 中国水旱灾害公报 . . Cited: 2016-6.URL [本文引用: 1]
[102]
MittlerR, ZilinskasBA (1994). Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. Plant Journal, 5, 397-405. [本文引用: 2]
[103]
MonakhovaOF, ChernyadèvII (2002). Protective role of kartolin-4 in wheat plants exposed to soil draught. Applied Biochemistry and Microbiology, 38, 373-380. [本文引用: 1]
[104]
MorganPB, AinsworthEA, LongSP (2003). How does elevated ozone impact soybean? A meta-analysis of photosynthesis, growth and yield. Plant, Cell & Environment, 26, 1317-1328. [本文引用: 1]
[105]
NaliC, PaolettiE, MarabottiniR, Della RoccaG, LorenziniG, PaolacciAR, CiaffiM, BadianiM (2004). Ecophysiological and biochemical, strategies of response to ozone in Mediterranean evergreen broadleaf species. Atmospheric Environment, 38, 2247-2257.
[106]
National Bureau of Statistics of China (2007-2012). China Statistical Yearbook. China Statistics Press, Beijing. (in Chinese)[国家统计局 (2007-2012). 中国统计年鉴. 中国统计出版社, 北京.] [本文引用: 1]
[107]
NikolovaPS, AndersenCP, BlaschkeH, MatyssekR, HaeberleK-H (2010). Belowground effects of enhanced tropospheric ozone and drought in a beech/spruce forest (Fagus sylvatica L./Picea abies (L.) Karst.). Environmental Pollution, 158, 1071-1078. [本文引用: 1]
[108]
NoctorG, FoyerCH (1998). Ascorbate and glutathione: Keeping active oxygen under control. Annual Review of Plant Physiology and Plant Molecular Biology, 49, 249-279. [本文引用: 1]
[109]
NonamiH (1998). Plant water relations and control of cell elongation at low water potentials. Journal of Plant Research, 111, 373-382. [本文引用: 1]
[110]
OksanenE (2003). Responses of selected birch (Betula pendula Roth) clones to ozone change over time. Plant, Cell & Environment, 26, 875-886. [本文引用: 1]
[111]
PääkkönenE, SeppänenS, HolopainenT, KokkoH, KärenlampiS, KärenlampiL, KangasjärviJ (1998). Induction of genes for the stress proteins PR-10 and PAL in relation to growth, visible injuries and stomatal conductance in birch (Betula pendula) clones exposed to ozone and/or drought. New Phytologist, 138, 295-305. [本文引用: 1]
[112]
PandeyD, GoswamiC, KumarB, JainS (2001). Hormonal regulation of photosynthetic enzymes in cotton under water stress. Photosynthetica, 38, 403-407.
[113]
PanekJA (2004). Ozone uptake, water loss and carbon exchange dynamics in annually drought-stressed Pinus ponderosa forests: Measured trends and parameters for uptake modeling. Tree Physiology, 24, 277-290. [本文引用: 3]
[114]
PanekJA, GoldsteinAH (2001). Response of stomatal conductance to drought in ponderosa pine: Implications for carbon and ozone uptake. Tree Physiology, 21, 337-344. [本文引用: 2]
[115]
PanekJA, KurpiusMR, GoldsteinAH (2002). An evaluation of ozone exposure metrics for a seasonally drought-stressed ponderosa pine ecosystem. Environmental Pollution, 117, 93-100. [本文引用: 1]
[116]
PaolettiE (2006). Impact of ozone on Mediterranean forests: A review. Environmental Pollution, 144, 463-474. [本文引用: 2]
[117]
PaolettiE, GrulkeNE (2010). Ozone exposure and stomatal sluggishness in different plant physiognomic classes. Environmental Pollution, 158, 2664-2671. [本文引用: 1]
[118]
ParryMAJ, AndralojcPJ, KhanS, LeaPJ, KeysAJ (2002). Rubisco activity: Effects of drought stress. Annals of Botany, 89, 833-839. [本文引用: 1]
[119]
PasqualiniS, BatiniP, EderliL, PorcedduA, PiccioniC, de MarchisF, AntonielliM (2001). Effects of short-term ozone fumigation on tobacco plants: Response of the scavenging system and expression of the glutathione reductase. Plant, Cell & Environment, 24, 245-252. [本文引用: 1]
[120]
PearsonM, MansfieldT (1993). Interacting effects of ozone and water stress on the stomatal resistance of beech (Fagus sylvatica L.). New Phytologist, 123, 351-358. [本文引用: 2]
[121]
PellEJ, SinnJP, EckardtN, JohansenCV, WinnerWE, MooneyHA (1993). Response of radish to multiple stresses. 2. Influence of season and genotype on plant- response to ozone and soil-moisture deficit. New Phytologist, 123, 153-163. [本文引用: 3]
[122]
PellouxJ, JolivetY, FontaineV, BanvoyJ, DizengremelP (2001). Changes in Rubisco and Rubisco activase gene expression and polypeptide content in Pinus halepensis M. subjected to ozone and drought. Plant, Cell & Environment, 24, 123-131. [本文引用: 1]
[123]
PignocchiC, FletcherJM, WilkinsonJE, BarnesJD, FoyerCH (2003). The function of ascorbate oxidase in tobacco. Plant Physiology, 132, 1631-1641.
[124]
PirkerKF, ReichenauerTG, PascualEC, KieferS, SojaG, GoodmanBA (2003). Steady state levels of free radicals in tomato fruit exposed to drought and ozone stress in a field experiment. Plant Physiology and Biochemistry, 41, 921-927. [本文引用: 1]
[125]
PollastriniM, DesotgiuR, CaminF, ZillerL, GerosaG, MarzuoliR, BussottiF (2014). Severe drought events increase the sensitivity to ozone on poplar clones. Environmental and Experimental Botany, 100, 94-104.
[126]
PollastriniM, DesotgiuR, CaminF, ZillerL, MarzuoliR, GerosaG, BussottiF (2013). Intra-annual pattern of photosynthesis, growth and stable isotope partitioning in a poplar clone subjected to ozone and water stress. Water, Air, & Soil Pollution, 224, 1761. [本文引用: 2]
[127]
PollastriniM, DesotgiuR, CascioC, BussottiF, CherubiniP, SaurerM, GerosaG, MarzuoliR (2010). Growth and physiological responses to ozone and mild drought stress of tree species with different ecological requirements. Trees, 24, 695-704. [本文引用: 2]
[128]
PottersG, de GaraL, AsardH, HoremansN (2002). Ascorbate and glutathione: Guardians of the cell cycle, partners in crime? Plant Physiology and Biochemistry, 40, 537-548.
[129]
PretzschH, DielerJ (2011). The dependency of the size-growth relationship of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.) in forest stands on long-term site conditions, drought events, and ozone stress. Trees-Structure and Function, 25, 355-369. [本文引用: 1]
[130]
QiuHJ, CaoMM, HaoJQ, WangYL, WangYM (2013). Analysis the relationship between drought frequency and scale of China in 1950-2010. Scientia Geographica Sinica, 33, 576-580. (in Chinese)[邱海军, 曹明明, 郝俊卿, 王雁林, 王彦民 (2013). 1950~2010年中国干旱灾情频率-规模关系分析 . 地理科学, 33, 576-580.] [本文引用: 1]
[131]
QuartacciMF, Navari-IzzoF (1992). Water-stress and free-radical mediated changes in sunflower seedlings. Journal of Plant Physiology, 139, 621-625.
[132]
RanaG, KaterjiN, MastrorilliM (2012). Method for automatic determination of soybean actual evapotranspiration under open top chambers (OTC) subjected to effects of water stress and air ozone concentration. Environmental Monitoring and Assessment, 184, 6377-6394. [本文引用: 1]
[133]
RaoMV, KochJR, DavisKR (2000). Ozone: A tool for probing programmed cell death in plants. Plant Molecular Biology, 44, 345-358. [本文引用: 1]
[134]
RenautJ, BohlerS, HausmanJ-F, HoffmannL, SergeantK, AhsanN, JolivetY, DizengremelP (2009). The impact of atmospheric composition on plants: A case study of ozone and poplar. Mass Spectrometry Reviews, 28, 495-516. [本文引用: 2]
[135]
RetzlaffWA, ArthurMA, GrulkeNE, WeinsteinDA, GollandsB (2000). Use of a single-tree simulation model to predict effects of ozone and drought on growth of a white fir tree. Tree Physiology, 20, 195-202. [本文引用: 1]
[136]
RibasA, PenuelasJ, ElviraS, GimenoBS (2005). Contrasting effects of ozone under different water supplies in two Mediterranean tree species. Atmospheric Environment, 39, 685-693. [本文引用: 1]
[137]
SadrasVO, MilroySP (1996). Soil-water thresholds for the responses of leaf expansion and gas exchange: A review. Field Crops Research, 47, 253-266. [本文引用: 1]
[138]
SandermannH (1996). Ozone and plant health. Annual Review of Phytopathology, 34, 347-366. [本文引用: 1]
[139]
SanmartinM, DrogoudiPD, LyonsT, PaterakiI, BarnesJ, KanellisAK (2003). Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. Planta, 216, 918-928. [本文引用: 1]
[140]
SchubertSD, SuarezMJ, PegionPJ, KosterRD, BacmeisterJT (2004). On the cause of the 1930s Dust Bowl. Science, 303, 1855-1859. [本文引用: 1]
[141]
SchupplerU, HePH, JohnPCL, MunnsR (1998). Effect of water stress on cell division and cell-division-cycle 2-like cell-cycle kinase activity in wheat leaves. Plant Physiology, 117, 667-678.
[142]
SergeantK, SpiessN, RenautJ, WilhelmE, HausmanJF (2011). One dry summer: A leaf proteome study on the response of oak to drought exposure. Journal of Proteomics, 74, 1385-1395. [本文引用: 1]
[143]
SharmaPN, MalikCP (1993). Photosynthetic responses of groundnut to moisture stress. Photosynthetica, 29, 157-160. [本文引用: 2]
[144]
ShimizuH, FengYW (2007). Ozone and/or water stresses could have influenced the Betula ermanii Cham. forest decline observed at Oku-Nikko, Japan. Environmental Monitoring and Assessment, 128, 109-119. [本文引用: 1]
[145]
ShowmanRE (1991). A comparison of ozone injury to vegetation during moist and drought years. Journal of the Air & Waste Management Association, 41, 63-64. [本文引用: 1]
[146]
ŠirceljH, TauszM, GrillD, BaticF (2005). Biochemical responses in leaves of two apple tree cultivars subjected to progressing drought. Journal of Plant Physiology, 162, 1308-1318. [本文引用: 1]
[147]
SitchS, CoxPM, CollinsWJ, HuntingfordC (2007). Indirect radiative forcing of climate change through ozone effects on the land-carbon sink. Nature, 448, 791-794. [本文引用: 1]
[148]
SkärbyL, Ro-PoulsenH, WellburnFAM, SheppardLJ (1998). Impacts of ozone on forests: A European perspective. New Phytologist, 139, 109-122. [本文引用: 1]
[149]
SofoA, TuzioAC, DichioB, XiloyannisC (2005). Influence of water deficit and rewatering on the components of the ascorbate-glutathione cycle in four interspecific Prunus hybrids. Plant Science, 169, 403-412.
[150]
SunMX, ZuCL, XuJN (2004). Advances research on the impact of drought on plant: A review. Journal of Anhui Agricultural Sciences, 32, 365-367, 384. (in Chinese)[孙梅霞, 祖朝龙, 徐经年 (2004). 干旱对植物影响的研究进展 . 安徽农业科学, 32, 365-367, 384.] [本文引用: 2]
[151]
TalbiS, Romero-PuertasMC, HernandezA, TerronL, Fer- chichiA, SandalioLM (2015). Drought tolerance in a Saharian plant Oudneya africana: Role of antioxidant defences. Environmental and Experimental Botany, 111, 114-126. [本文引用: 1]
TezaraW, MitchellV, DriscollS, LawlorD (1999). Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature, 401, 914-917. [本文引用: 1]
[154]
TianHQ, XuXF, SongX (2007). Drought impacts on terrestrial ecosystem productivity. Journal of Plant Ecology ( Chinese Version), 31, 231-241. (in Chinese with English abstract)[田汉勤, 徐小锋, 宋霞 (2007). 干旱对陆地生态系统生产力的影响 . 植物生态学报, 31, 231-241.] [本文引用: 1]
[155]
TietzS, WildA (1991). Investigations on the phosphoenolpyruvate carboxylase activity of spruce needles relative to the occurrence of novel forest decline. Journal of Plant Physiology, 137, 327-331. [本文引用: 1]
[156]
TorsethaugenG, PellEJ, AssmannSM (1999). Ozone inhibits guard cell K+ channels implicated in stomatal opening. Proceedings of the National Academy of Sciences of the United States of America, 96, 13577-13582. [本文引用: 1]
[157]
VitaleM, SalvatoriE, LoretoF, FaresS, ManesF (2008). Physiological responses of Quercus ilex leaves to water stress and acute ozone exposure under controlled conditions. Water, Air, & Soil Pollution, 189, 113-125. [本文引用: 1]
[158]
WaggS, MillsG, HayesF, WilkinsonS, CooperD, DaviesWJ (2012). Reduced soil water availability did not protect two competing grassland species from the negative effects of increasing background ozone. Environmental Pollution, 165, 91-99.
[159]
WallinG, KarlssonPE, SelldenG, OttossonS, MedinEL, PleijelH, SkarbyL (2002). Impact of four years exposure to different levels of ozone, phosphorus and drought on chlorophyll, mineral nutrients, and stem volume of Norway spruce, Picea abies. Physiologia Plantarum, 114, 192-206.
[160]
WangXL, ChenQC (1974). Plants as “atmospheric pollution monitoring alarm”. The Plant Journal, (4), 29-31. (in Chinese)[王勋陵, 陈庆诚 (1974). 利用植物作为大气污染“监测警报器” . 植物学杂志, (4), 29-31.]
[161]
WatanabeM, YonekuraT, HondaY, YoshidomeM, NakajiT, IzutaT (2005). Effects of ozone and soil water stress, singly and in combination, on leaf antioxidative systems of Fagus crenata seedlings. Journal of Agricultural Meteorology, 60, 1105-1108. [本文引用: 2]
[162]
WellburnFAM, LauKK, MillingPMK, WellburnAR (1996). Drought and air pollution affect nitrogen cycling and free radical scavenging in Pinus halepensis (Mill). Journal of Experimental Botany, 47, 1361-1367.
[163]
WenZ, WangL, WangXK, LiL, CuiJ (2014). Combined effects of O3 and drought on leaf stomata of Acer truncatum. Chinese Journal of Ecology, 33, 560-566. (in Chinese with English abstract)[文志, 王丽, 王效科, 李丽, 崔健 (2014). O3和干旱胁迫对元宝枫叶片气孔特征的复合影响 . 生态学杂志, 33, 560-566.] [本文引用: 2]
[164]
WilkinsonS, DaviesWJ (2009). Ozone suppresses soil drying- and abscisic acid (ABA)-induced stomatal closure via an ethylene-dependent mechanism. Plant, Cell & Environment, 32, 949-959. [本文引用: 1]
[165]
WilkinsonS, DaviesWJ (2010). Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant, Cell & Environment, 33, 510-525. [本文引用: 2]
[166]
WittigVE, AinsworthEA, LongSP (2007). To what extent do current and projected increases in surface ozone affect photosynthesis and stomatal conductance of trees? A meta- analytic review of the last 3 decades of experiments. Plant, Cell & Environment, 30, 1150-1162. [本文引用: 2]
[167]
WittigVE, AinsworthEA, NaiduSL, KarnoskyDF, LongSP (2009). Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: A quantitative meta-analysis. Global Change Biology, 15, 396-424.
[168]
XuH, BiswasDK, LiWD, ChenSB, ZhangL, JiangGM, LiYG (2007). Photosynthesis and yield responses of ozone- polluted winter wheat to drought. Photosynthetica, 45, 582-588. [本文引用: 1]
[169]
XuH, YangJC, ChenSB, JiangGB, LiKY (2007). Review of plant responses to ozone pollution. Journal of Plant Ecology ( Chinese Version), 31, 1205-1213. (in Chinese with English abstract)[许宏, 杨景成, 陈圣宾, 蒋高明, 李永庚 (2007). 植物的臭氧污染胁迫效应研究进展 . 植物生态学报, 31, 1205-1213.] [本文引用: 1]
[170]
YangF, MiaoLF, XuXiao, LiCY (2007). Progress in research of plant responses to drought stress. Chinese Journal of Applied and Environmental Biology, 13, 586-591. (in Chinese with English abstract)[杨帆, 苗灵凤, 胥晓, 李春阳 (2007). 植物对干旱胁迫的响应研究进展 . 应用与环境生物学报, 13, 586-591.] [本文引用: 1]
[171]
YaoYQ, LiuXP, LiZZ, MaXF, RennenbergH, WangX, LiHC (2013). Drought-induced H2O2 accumulation in subsidiary cells is involved in regulatory signaling of stomatal closure in maize leaves. Planta, 238, 217-227.
[172]
YeLH, BaoHY, WangZY, LieGW, ChenHY, ZhangXP, ChenX, KeH, TianXQ, TanJD (2014). Effects of ozone and drought on biomass allocation of four seedlings in South China. Advanced Materials Research, 864-867, 2478-2484.
[173]
YonekuraTHY, OksanenE, YoshidomeM, WatanabeM, Funada RKT, IzutaT (2001b). The influences of ozone and soil water stress, singly and in combination, on leaf gas exchange rates, leaf ultrastructural characteristics and annual ring width of Fagus crenata seedlings. Journal of Japan Society for Atmospheric Environment, 36, 333-351.
[174]
YonekuraT, DokiyaY, FukamiM, IzutaT (2001a). Effects of ozone and/or soil water stress on growth and photosynthesis of Fagus crenata seedlings. Water, Air, & Soil Pollution, 130, 965-970.
[175]
YonekuraT, YoshidomeM, WatanabeM, HondaY, OgiwaraI, IzutaT (2004). Carry-over effects of ozone and water stress on leaf phenological characteristics and bud frost hardiness of Fagus crenata seedlings. Trees, 18, 581-588. [本文引用: 1]
[176]
ZhangJX, KirkhamMB (1994). Drought-stress-induced chang- es in activities of superoxide-dismutase, catalase, and peroxidase in wheat species. Plant & Cell Physiology, 35, 785-791. [本文引用: 1]
[177]
ZhangJX, KirkhamMB (1996). Enzymatic responses of the ascorbate-glutathione cycle to drought in sorghum and sunflower plants. Plant Science, 113, 139-147. [本文引用: 1]
[178]
ZhaoMS, RunningSW (2010). Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science, 329, 940-943.
Relating wet and dry year ecophysiology to leaf structure in contrastingtemperate tree species. 1 1994
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Interactive effects of ozone and drought stress on pigments and activities of antioxidative enzymes in Pinus halepensis. 6 2001
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
... 抗坏血酸-谷胱甘肽(AsA-GSH)循环通过调节氧化还原反应防御ROS的伤害, 是植物细胞中主要的抗氧化途径.植物会通过抗氧化物质的结合消除ROS的毒害, 例如存在于叶绿体、细胞质、线粒体和过氧化物酶体中的一些抗氧化物质, 如抗坏血酸盐、谷胱甘肽、维生素a、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)以及AsA-GSH循环途径中抗坏血酸过氧化物酶(APX)、单脱水抗坏血酸还原酶(MDAR)、脱氢抗坏血酸还原酶(DHAR)和谷胱甘肽还原酶(GR)等构成细胞器中重要的抗氧化防御体系, 防止过氧化氢(H2O2)的产生(Potters et al., 2002).Iyer等(2013)证实由于还原型AsA和GSH的增加, O3和干旱复合胁迫下ROS的水平与无胁迫对照相近.干旱增加了Fagus crenata叶片的GSH含量, 提高了抗性机制, 减轻了植物受到的O3伤害(Watanabe et al., 2005).O3暴露下植物的APX、CAT、SOD等抗氧化酶类的活性和(或)丰度是增加的(Alonso et al., 2001), 且干旱条件下植物的GR和SOD也是增加的(Alonso et al., 2001; Huseynova et al., 2014), 而与O3和干旱的单独胁迫相比, 两者交互作用下抗氧化酶类反而是降低的, 从而推断两种胁迫的累积作用破坏了植物的防御体系(Wellburn et al., 1996; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
... ), 且干旱条件下植物的GR和SOD也是增加的(Alonso et al., 2001; Huseynova et al., 2014), 而与O3和干旱的单独胁迫相比, 两者交互作用下抗氧化酶类反而是降低的, 从而推断两种胁迫的累积作用破坏了植物的防御体系(Wellburn et al., 1996; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
... ; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Drought stress does not protect Quercus ilex L. from ozone effects: Results from a comparative study of two subspecies differing in ozone sensitivity. 1 2014
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
Water stress in barley (Hordeum vulgare L.). II. Effect on chemical composition and chlorophyll contents. 1 2003
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
The role of ascorbate peroxidase and monodehydroascorbate reductase in H2O2 scavenging in plants. 1 1997
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Assessing the future global impacts of ozone on vegetation. 1 2005
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Global crop yield reductions due to surface ozone exposure: 2. Year 2030 potential crop production losses and economic damage under two scenarios of O3 pollution. 1 2011
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Evidence for the contribution of the Mehler-peroxidase reaction in dissipating excess electrons in drought-stressed wheat. 1 1996
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Differential drought-induced modulation of ozone tolerance in winter wheat species. 6 2011
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
... 的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
... ; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
... 和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
A DIGE analysis of developing poplar leaves subjected to ozone reveals major changes in carbon metabolism. 2 2007
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
A difference gel electrophoresis study on thylakoids isolated from poplar leaves reveals a negative impact of ozone exposure on membrane proteins. 1 2011
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
A physiological and proteomic study of poplar leaves during ozone exposure combined with mild drought. 6 2013
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
... 单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
... O3和干旱对不同植物的器官、组织和细胞具有不同程度的氧化伤害, 当胁迫达到一定程度时, 首先伤害敏感植物的叶片, 进而限制整株植物的生长.两种胁迫下叶片的表观伤害症状则各有特征.O3引起叶片坏疽斑块和不规则的褪绿、萎黄斑点, 而干旱引起叶片同质和渐进的变色, 同时伴有叶片的发育不良和卷曲下垂(Bohler et al., 2013).O3和干旱交互作用下的植物则并不表现为一致的加重伤害.与单独胁迫下的植物相比, 双重胁迫可能减弱(Showman, 1991; Iyer et al., 2013)或者加重(Bohler et al., 2013)叶片受伤害的表型特征.因此, 叶片的不同伤害症状表现可能是由于病斑的产生与胁迫诱导ROS大量生成并引发植物自身调节抗氧化系统有关, 不一定是气孔吸收O3所致(Grulke et al., 2003a).这也表明O3和干旱的相互作用对植物的影响在表观上和形态学上可能是相似的或累积的.但即使O3和(或)干旱胁迫下植物未表现出明显可见的叶片伤害症状, 其生长最后也可能受到限制(Shimizu & Feng, 2007). ...
... )或者加重(Bohler et al., 2013)叶片受伤害的表型特征.因此, 叶片的不同伤害症状表现可能是由于病斑的产生与胁迫诱导ROS大量生成并引发植物自身调节抗氧化系统有关, 不一定是气孔吸收O3所致(Grulke et al., 2003a).这也表明O3和干旱的相互作用对植物的影响在表观上和形态学上可能是相似的或累积的.但即使O3和(或)干旱胁迫下植物未表现出明显可见的叶片伤害症状, 其生长最后也可能受到限制(Shimizu & Feng, 2007). ...
Differential impact of chronic ozone exposure on expanding and fully expanded poplar leaves. 1 2010
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? 1 2004
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Temperate forest trees and stands under severe drought: A review of eco- physiological responses, adaptation processes and long- term consequences. 1 2006
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Ozone-induced changes in biosynthesis of Rubisco and associated compensation to stress in foliage of hybrid poplar. 1 1998
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Ozone and forest trees. 1 1998
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
The role of ascorbate in drought-treated Cochlearia atlantica Pobed. and Armeria maritima (Mill.) Willd. 1 1991
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Ozone variations through vehicle emissions reductions based on air quality monitoring data in Taipei City, Taiwan, from 1994 to 2003. 1 2006
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Increasing tolerance to ozone by elevating foliar ascorbic acid confers greater protection against ozone than increasing avoidance. 2 2005
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Europe-wide reduction in primary productivity caused by the heat and drought in 2003. 2005
Weather highlights around the world. 1 1998
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Ascorbic acid, a familiar small molecule intertwined in the response of plants to ozone, pathogens, and the onset of senescence. 1 2004
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Stomatal behaviour and environment. 1 1978
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
1
... 抗坏血酸-谷胱甘肽(AsA-GSH)循环通过调节氧化还原反应防御ROS的伤害, 是植物细胞中主要的抗氧化途径.植物会通过抗氧化物质的结合消除ROS的毒害, 例如存在于叶绿体、细胞质、线粒体和过氧化物酶体中的一些抗氧化物质, 如抗坏血酸盐、谷胱甘肽、维生素a、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)以及AsA-GSH循环途径中抗坏血酸过氧化物酶(APX)、单脱水抗坏血酸还原酶(MDAR)、脱氢抗坏血酸还原酶(DHAR)和谷胱甘肽还原酶(GR)等构成细胞器中重要的抗氧化防御体系, 防止过氧化氢(H2O2)的产生(Potters et al., 2002).Iyer等(2013)证实由于还原型AsA和GSH的增加, O3和干旱复合胁迫下ROS的水平与无胁迫对照相近.干旱增加了Fagus crenata叶片的GSH含量, 提高了抗性机制, 减轻了植物受到的O3伤害(Watanabe et al., 2005).O3暴露下植物的APX、CAT、SOD等抗氧化酶类的活性和(或)丰度是增加的(Alonso et al., 2001), 且干旱条件下植物的GR和SOD也是增加的(Alonso et al., 2001; Huseynova et al., 2014), 而与O3和干旱的单独胁迫相比, 两者交互作用下抗氧化酶类反而是降低的, 从而推断两种胁迫的累积作用破坏了植物的防御体系(Wellburn et al., 1996; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
Drought under global warming: A review. 1 2011
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Chlorophyll a fluorescence analysis along a vertical gradient of the crown in a poplar (Oxford clone) subjected to ozone and water stress. 2012
Other factors than apoplastic ascorbate contribute to the differential ozone tolerance of two clones of Trifolium repens L. 1 2005
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Effects of ozone on the carbon metabolism of forest trees. 1 2001
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Metabolic-dependent changes in plant cell redox power after ozone exposure. 1 2009
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
Effects of ozone on stomatal responses to environmental parameters (blue light, red light, CO2 and vapour pressure deficit) in three Populus deltoides × Populus nigra genotypes. 1 2013
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Fungal community composition and function after long-term exposure of northern forests to elevated atmospheric CO2 and tropospheric O3. 1 2011
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Enhanced tolerance to ozone and drought stresses in transgenic tobacco overexpressing dehydroascorbate reductase in cytosol. 1 2006
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
1
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Photosynthesis, photorespiration and proline accumulation in water-stressed pea leaves. 1 1996
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
Effects of ozone and/or water stress on the growth of Abies veitchii seedlings. 2005
Evidence of widespread ozone-induced visible injury on plants in Beijing, China. 1 2014
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Assessing the impacts of current and future concentrations of surface ozone on crop yield with meta-analysis. 1 2009
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. 3 2002
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
... ), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
Regulation of phosphoenolpyruvate carboxylase in Pinus halepensis needles submitted to ozone and water stress. 1 2003
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
Oxygen processing in photosynthesis: Regulation and signalling. 1 2000
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. 1 2001
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Carbon metabolism enzyme activities and carbon partitioning in Pinus halepensis Mill. exposed to mild drought and ozone. 1 1996
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
a). Canopy transpiration of Jeffrey pine in mesic and xeric microsites: O3 uptake and injury response. 1 2003
... O3和干旱对不同植物的器官、组织和细胞具有不同程度的氧化伤害, 当胁迫达到一定程度时, 首先伤害敏感植物的叶片, 进而限制整株植物的生长.两种胁迫下叶片的表观伤害症状则各有特征.O3引起叶片坏疽斑块和不规则的褪绿、萎黄斑点, 而干旱引起叶片同质和渐进的变色, 同时伴有叶片的发育不良和卷曲下垂(Bohler et al., 2013).O3和干旱交互作用下的植物则并不表现为一致的加重伤害.与单独胁迫下的植物相比, 双重胁迫可能减弱(Showman, 1991; Iyer et al., 2013)或者加重(Bohler et al., 2013)叶片受伤害的表型特征.因此, 叶片的不同伤害症状表现可能是由于病斑的产生与胁迫诱导ROS大量生成并引发植物自身调节抗氧化系统有关, 不一定是气孔吸收O3所致(Grulke et al., 2003a).这也表明O3和干旱的相互作用对植物的影响在表观上和形态学上可能是相似的或累积的.但即使O3和(或)干旱胁迫下植物未表现出明显可见的叶片伤害症状, 其生长最后也可能受到限制(Shimizu & Feng, 2007). ...
Variation in morphological and biochemical O3 injury attributes of mature Jeffrey pine within canopies and between microsites. 2003
O3 uptake and drought stress effects on carbon acquisition of ponderosa pine in natural stands. 1 2002
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
From critical levels to critical loads for ozone: A discussion of a new experimental and modelling approach for establishing flux-response relationships for agricultural crops and native plant species. 1 2003
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
Effect of chronic ozone fumigation on the photosynthetic process of poplar clones showing different sensitivity. 1 2001
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Gene expression patterns of trembling aspen trees following long-term exposure to interacting elevated CO2 and tropospheric O3. 1 2005
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Effects of drought and canopy ozone exposure on antioxidants in fine roots of mature European beech (Fagus sylvatica). 2008
臭氧与干旱胁迫对海南蒲桃和盆架子幼苗光合生理的影响 2014
Light-induced changes in the content of some enzymes of the C4-dicarboxylic acid pathway of photosynthesis and its effect on other characteristics of photosynthesis. 1 1969
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
2 1994
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Modification of the biochemical pathways of plants induced by ozone: What are the varied routes to change? 1 2008
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
1997
1 1997
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Complex interactive effects of drought and ozone stress on the antioxidant defence systems of two wheat cultivars. 2002
Tree age dependence and within-canopy variation of leaf gas exchange and antioxidative defence in Fagus sylvatica under experimental free-air ozone exposure. 1 2005
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Determinants of stomatal sluggishness in ozone-exposed deciduous tree species. 3 2014
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... ), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... )及季节的变化有关(Hoshika et al., 2014). ...
Both ozone exposure and soil water stress are able to induce stomatal sluggishness. 2013
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Effects of ozone and mild drought stress on gas exchange, antioxidants and chloroplast pigments in current-year needles of young Norway spruce [Picea abies (L.) Karst]. 1998
Daily growth of European beech (Fagus sylvatica L.) on moist sites is affected by short-term drought rather than ozone uptake. 1 2015
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
Ozone induced microscopical changes and quantitative carbohydrate contents of hybrid poplar (Populus × euramericana). 2 1994
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
The effects of slightly elevated ozone concentrations and mild drought stress on the physiology and growth of Norway Spruce, Picea abies (L.) Karst. and beech, Fagus sylvatica L., in open-top chambers. 1 1994
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Ozone sensitivity analysis with the MM5-CMAQ modeling system for Shanghai. 2011
Chronic drought stress reduced but not protected Shantung maple (Acer truncatum Bunge) from adverse effects of ozone (O3) on growth and physiology in the suburb of Beijing, China. 2015
臭氧与干旱胁迫对华南地区3种绿化树种 2011
Extraordinary drought of 2003 overrules ozone impact on adult beech trees (Fagus sylvatica). 1 2006
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
Beech trees exposed to high CO2 and to simulated summer ozone levels: Effects on photosynthesis, chloroplast components and leaf enzyme activity. 1 2000
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Role of ascorbate in detoxifying ozone in the apoplast of spinach (Spinacia oleracea L.) leaves. 1 1993
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Image analysis of the stomatal cell walls of Picea abies (L.) Karst. in pure and ozone-enriched air. 1 1998
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
Dose- response relationships of ozone effects on foliar levels of antioxidants, soluble polyamines and peroxidase activity of Pinus taeda (L.): Assessment of the usefulness as early ozone indicators. 1991
Physiological response of Pinus halepensis needles under ozone and water stress conditions. 1 2001
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Ozone—A risk factor for trees and forests in Europe? 1999
3 2014
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Interactions between drought and O3 stress in forest trees. 1 2006
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Impact of ozone on trees: An ecophysiological perspective. 1 2003
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Forests under climate change and air pollution: Gaps in understanding and future directions for research. 2 2012
... 政府间气候变化专门委员会(IPCC, 2013)报告指出由气候变化导致的环境因素改变对生物体系有直接、间接以及交互的影响.在植物旺盛生长的时期, 充足的光照和高温往往导致O3浓度爆发性的升高, 并常常伴随着干旱的发生, 从而使得植物同时受到高浓度O3和干旱的双重胁迫(Matyssek et al., 2010). ...
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica)—Resume from the free-air fumigation study at Kranzberg Forest. 2010
Interactive effects of ozone and climate on tree growth and water use in a southern Appalachian forest in the USA. 2007
2 1999
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Evidence of widespread effects of ozone on crops and (semi-) natural vegetation in Europe (1990-2006) in relation to AOT40- and flux-based risk maps. 2011
中国水旱灾害公报 1 2006-2010
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. 2 1994
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
... ; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Protective role of kartolin-4 in wheat plants exposed to soil draught. 1 2002
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
How does elevated ozone impact soybean? A meta-analysis of photosynthesis, growth and yield. 1 2003
Ecophysiological and biochemical, strategies of response to ozone in Mediterranean evergreen broadleaf species. 2004
1 2007-2012
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
Belowground effects of enhanced tropospheric ozone and drought in a beech/spruce forest (Fagus sylvatica L./Picea abies (L.) Karst.). 1 2010
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Ascorbate and glutathione: Keeping active oxygen under control. 1 1998
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Plant water relations and control of cell elongation at low water potentials. 1 1998
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Responses of selected birch (Betula pendula Roth) clones to ozone change over time. 1 2003
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Induction of genes for the stress proteins PR-10 and PAL in relation to growth, visible injuries and stomatal conductance in birch (Betula pendula) clones exposed to ozone and/or drought. 1 1998
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
Hormonal regulation of photosynthetic enzymes in cotton under water stress. 2001
Ozone uptake, water loss and carbon exchange dynamics in annually drought-stressed Pinus ponderosa forests: Measured trends and parameters for uptake modeling. 3 2004
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Response of stomatal conductance to drought in ponderosa pine: Implications for carbon and ozone uptake. 2 2001
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
An evaluation of ozone exposure metrics for a seasonally drought-stressed ponderosa pine ecosystem. 1 2002
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Impact of ozone on Mediterranean forests: A review. 2 2006
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... ).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Ozone exposure and stomatal sluggishness in different plant physiognomic classes. 1 2010
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Rubisco activity: Effects of drought stress. 1 2002
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Effects of short-term ozone fumigation on tobacco plants: Response of the scavenging system and expression of the glutathione reductase. 1 2001
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Interacting effects of ozone and water stress on the stomatal resistance of beech (Fagus sylvatica L.). 2 1993
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
... )以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Response of radish to multiple stresses. 2. Influence of season and genotype on plant- response to ozone and soil-moisture deficit. 3 1993
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
... 的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
... 胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Changes in Rubisco and Rubisco activase gene expression and polypeptide content in Pinus halepensis M. subjected to ozone and drought. 1 2001
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
The function of ascorbate oxidase in tobacco. 2003
Steady state levels of free radicals in tomato fruit exposed to drought and ozone stress in a field experiment. 1 2003
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
Severe drought events increase the sensitivity to ozone on poplar clones. 2014
Intra-annual pattern of photosynthesis, growth and stable isotope partitioning in a poplar clone subjected to ozone and water stress. 2 2013
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
Growth and physiological responses to ozone and mild drought stress of tree species with different ecological requirements. 2 2010
... 抗坏血酸-谷胱甘肽(AsA-GSH)循环通过调节氧化还原反应防御ROS的伤害, 是植物细胞中主要的抗氧化途径.植物会通过抗氧化物质的结合消除ROS的毒害, 例如存在于叶绿体、细胞质、线粒体和过氧化物酶体中的一些抗氧化物质, 如抗坏血酸盐、谷胱甘肽、维生素a、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)以及AsA-GSH循环途径中抗坏血酸过氧化物酶(APX)、单脱水抗坏血酸还原酶(MDAR)、脱氢抗坏血酸还原酶(DHAR)和谷胱甘肽还原酶(GR)等构成细胞器中重要的抗氧化防御体系, 防止过氧化氢(H2O2)的产生(Potters et al., 2002).Iyer等(2013)证实由于还原型AsA和GSH的增加, O3和干旱复合胁迫下ROS的水平与无胁迫对照相近.干旱增加了Fagus crenata叶片的GSH含量, 提高了抗性机制, 减轻了植物受到的O3伤害(Watanabe et al., 2005).O3暴露下植物的APX、CAT、SOD等抗氧化酶类的活性和(或)丰度是增加的(Alonso et al., 2001), 且干旱条件下植物的GR和SOD也是增加的(Alonso et al., 2001; Huseynova et al., 2014), 而与O3和干旱的单独胁迫相比, 两者交互作用下抗氧化酶类反而是降低的, 从而推断两种胁迫的累积作用破坏了植物的防御体系(Wellburn et al., 1996; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Ascorbate and glutathione: Guardians of the cell cycle, partners in crime? 2002
The dependency of the size-growth relationship of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.) in forest stands on long-term site conditions, drought events, and ozone stress. 1 2011
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
1950~2010年中国干旱灾情频率-规模关系分析 1 2013
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Water-stress and free-radical mediated changes in sunflower seedlings. 1992
Method for automatic determination of soybean actual evapotranspiration under open top chambers (OTC) subjected to effects of water stress and air ozone concentration. 1 2012
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Ozone: A tool for probing programmed cell death in plants. 1 2000
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
The impact of atmospheric composition on plants: A case study of ozone and poplar. 2 2009
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... 和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
Use of a single-tree simulation model to predict effects of ozone and drought on growth of a white fir tree. 1 2000
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Contrasting effects of ozone under different water supplies in two Mediterranean tree species. 1 2005
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Soil-water thresholds for the responses of leaf expansion and gas exchange: A review. 1 1996
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
Ozone and plant health. 1 1996
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. 1 2003
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
On the cause of the 1930s Dust Bowl. 1 2004
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Effect of water stress on cell division and cell-division-cycle 2-like cell-cycle kinase activity in wheat leaves. 1998
One dry summer: A leaf proteome study on the response of oak to drought exposure. 1 2011
... 光合碳代谢中二磷酸核酮糖羧化酶(Rubisco)是C3植物碳固定的关键酶, 同时也是植物光呼吸的关键酶.O3通过降低Rubisco酶活性、酶亚基丰度(Brendley & Pell, 1998; Pelloux et al., 2001)影响植物的生理功能, 并导致净光合速率、最大羧化速率以及气孔导度的降低(Guidi et al., 2001; Morgan et al., 2003; Biswas & Jiang, 2011).在卡尔文循环中一些氧化还原调节酶类(如Rubisco活化酶、果糖-1,6-二磷酸酶、景天庚酮糖-1,7-二磷酸酶和核酮糖-5-磷酸激酶)在O3胁迫下受到抑制而导致其丰度降低(Bohler et al., 2007, 2010, 2013), 使卡尔文循环被激活.土壤因有效水分减少而降低光合速率也主要是因为Rubisco活性的降低(Bota et al., 2004), 进而导致植物的生长受限(Huang & Fu, 2000).Rubisco对胁迫的响应往往因植物品种不同和环境的改变而不同.如水分胁迫下烟草的Rubisco酶类受到抑制降低了Rubisco活性(Parry et al., 2002), 而夏栎(Guercus robur)的Rubisco活化酶增加导致Rubisco降解(Ser- geant et al., 2011), Pinus halepensis的Rubisco和Rubisco活化酶转录的丰度及蛋白含量则并未改变(Pelloux et al., 2001).在O3与干旱的交互作用下, 因两种胁迫各自对植物Rubisco的影响不同和植物本身生长阶段的不同, 表现出更加复杂多变的结果: 复合胁迫下杨树(Populus tremula × P. alba) Rubisco的降解是增加的, 但并不显著, 而且与干旱相比O3的影响更强(Bohler et al., 2013).O3对P. halepensis Rubisco的影响在植物生长后期也可能强于干旱, O3胁迫下Rubisco活性降低, 但是干旱无持续的显著作用, 使复合胁迫下Rubisco活性降低(Pelloux et al., 2001).但也有结果显示P. halepensis的Rubisco活性并未因O3而改变, 而是因干旱显著降低, 使两种胁迫下Rubisco活性显著降低(Gerant et al., 1996). ...
Photosynthetic responses of groundnut to moisture stress. 2 1993
... 光合碳代谢的磷酸烯醇丙酮酸羧化酶(PEPc)对固定CO2有重要作用(Hatch et al., 1969), 而且PEPc活性增强可激发三羧酸循环保持氮代谢运行的补偿机制(Tietz & Wild, 1991).PEPc活性的变化不仅取决于胁迫时间, 而且因品种变化而不同.例如有实验证实干旱条件下豌豆(Pisum sativum)和Arachis pintoi中的PEPc活性是增强的(Sharma & Nisha, 1993; Fedina & Popova, 1996), 棉花(Gossypium hirsutum)中PEPc活性表现出降低的趋势(Pandey et al., 2001), 而P. halepensis中PEPc的活性、数量和转录都无显著改变(Fontaine et al., 2003).在高浓度O3下, 植物叶片的PEPc活性是增加的(Landolt et al., 1994; Lutz et al., 2000; Inclan et al., 2005), 但也可能随时间改变而发生变化.Fontaine等(2003)就指出三年生P. halepensis的PEPc活性只有在O3暴露后期才是增加的, 同时O3和干旱交互作用下生长后期的PEPc活性也是增加的, 但是比单独O3的影响低.也有结果显示O3和干旱复合胁迫对二年生P. halepensis的PEPc活性无影响(Inclan et al., 2005).因此, PEPc的活性因植物品种, 胁迫发生时间、持续时间以及两种胁迫的作用强度不同而不同. ...
... O3和干旱对不同植物的器官、组织和细胞具有不同程度的氧化伤害, 当胁迫达到一定程度时, 首先伤害敏感植物的叶片, 进而限制整株植物的生长.两种胁迫下叶片的表观伤害症状则各有特征.O3引起叶片坏疽斑块和不规则的褪绿、萎黄斑点, 而干旱引起叶片同质和渐进的变色, 同时伴有叶片的发育不良和卷曲下垂(Bohler et al., 2013).O3和干旱交互作用下的植物则并不表现为一致的加重伤害.与单独胁迫下的植物相比, 双重胁迫可能减弱(Showman, 1991; Iyer et al., 2013)或者加重(Bohler et al., 2013)叶片受伤害的表型特征.因此, 叶片的不同伤害症状表现可能是由于病斑的产生与胁迫诱导ROS大量生成并引发植物自身调节抗氧化系统有关, 不一定是气孔吸收O3所致(Grulke et al., 2003a).这也表明O3和干旱的相互作用对植物的影响在表观上和形态学上可能是相似的或累积的.但即使O3和(或)干旱胁迫下植物未表现出明显可见的叶片伤害症状, 其生长最后也可能受到限制(Shimizu & Feng, 2007). ...
Ozone and/or water stresses could have influenced the Betula ermanii Cham. forest decline observed at Oku-Nikko, Japan. 1 2007
... O3和干旱对不同植物的器官、组织和细胞具有不同程度的氧化伤害, 当胁迫达到一定程度时, 首先伤害敏感植物的叶片, 进而限制整株植物的生长.两种胁迫下叶片的表观伤害症状则各有特征.O3引起叶片坏疽斑块和不规则的褪绿、萎黄斑点, 而干旱引起叶片同质和渐进的变色, 同时伴有叶片的发育不良和卷曲下垂(Bohler et al., 2013).O3和干旱交互作用下的植物则并不表现为一致的加重伤害.与单独胁迫下的植物相比, 双重胁迫可能减弱(Showman, 1991; Iyer et al., 2013)或者加重(Bohler et al., 2013)叶片受伤害的表型特征.因此, 叶片的不同伤害症状表现可能是由于病斑的产生与胁迫诱导ROS大量生成并引发植物自身调节抗氧化系统有关, 不一定是气孔吸收O3所致(Grulke et al., 2003a).这也表明O3和干旱的相互作用对植物的影响在表观上和形态学上可能是相似的或累积的.但即使O3和(或)干旱胁迫下植物未表现出明显可见的叶片伤害症状, 其生长最后也可能受到限制(Shimizu & Feng, 2007). ...
A comparison of ozone injury to vegetation during moist and drought years. 1 1991
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Biochemical responses in leaves of two apple tree cultivars subjected to progressing drought. 1 2005
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
Indirect radiative forcing of climate change through ozone effects on the land-carbon sink. 1 2007
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Impacts of ozone on forests: A European perspective. 1 1998
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Influence of water deficit and rewatering on the components of the ascorbate-glutathione cycle in four interspecific Prunus hybrids. 2005
... 叶绿体电子传递链中, O3使光合系统和三磷腺苷(ATP)酶的一些亚基显著降低(Bohler et al., 2007, 2011), 而植物对O3伤害的抵御与O3通量有关(Dizengremel et al., 2008).干旱加强植物气孔关闭, 限制了O3的吸收, 使气体交换受限, 但同时也影响了ATP酶(Tezara et al., 1999; Flexas et al., 2002), 限制了光合电子传递速率(Vitale et al., 2008).O3诱导铁氧还蛋白-NADP+-氧化还原酶在胁迫初期快速增加(Bohler et al., 2013), 这与氧化胁迫下植物的响应一致(Dizengremel et al., 2008, 2009; Heath, 2008).但在长期的O3胁迫下铁氧还蛋白-NADP+-氧化还原酶会由于防御体系超负荷而降低, 一直降低的光合系统亚基也可能是由叶绿体中ATP和NADPH累积引起, 导致卡尔文循环活力降低, 而这些都未在干旱胁迫中发现.O3和干旱交互作用下引起卡尔文循环中的蛋白质含量降低, 这与O3单独胁迫的结果相似, 但影响程度较低, 并未产生显著的改变(Bohler et al., 2013).可见, O3和干旱对植物光合碳代谢的影响与品种和胁迫强度及实际的环境状况有关, 目前尚缺乏复合胁迫下相关蛋白质组调控机理研究. ...
Physiological responses of Quercus ilex leaves to water stress and acute ozone exposure under controlled conditions. 1 2008
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Reduced soil water availability did not protect two competing grassland species from the negative effects of increasing background ozone. 2012
Impact of four years exposure to different levels of ozone, phosphorus and drought on chlorophyll, mineral nutrients, and stem volume of Norway spruce, Picea abies. 2002
利用植物作为大气污染“监测警报器” 1974
Effects of ozone and soil water stress, singly and in combination, on leaf antioxidative systems of Fagus crenata seedlings. 2 2005
... 抗坏血酸-谷胱甘肽(AsA-GSH)循环通过调节氧化还原反应防御ROS的伤害, 是植物细胞中主要的抗氧化途径.植物会通过抗氧化物质的结合消除ROS的毒害, 例如存在于叶绿体、细胞质、线粒体和过氧化物酶体中的一些抗氧化物质, 如抗坏血酸盐、谷胱甘肽、维生素a、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)以及AsA-GSH循环途径中抗坏血酸过氧化物酶(APX)、单脱水抗坏血酸还原酶(MDAR)、脱氢抗坏血酸还原酶(DHAR)和谷胱甘肽还原酶(GR)等构成细胞器中重要的抗氧化防御体系, 防止过氧化氢(H2O2)的产生(Potters et al., 2002).Iyer等(2013)证实由于还原型AsA和GSH的增加, O3和干旱复合胁迫下ROS的水平与无胁迫对照相近.干旱增加了Fagus crenata叶片的GSH含量, 提高了抗性机制, 减轻了植物受到的O3伤害(Watanabe et al., 2005).O3暴露下植物的APX、CAT、SOD等抗氧化酶类的活性和(或)丰度是增加的(Alonso et al., 2001), 且干旱条件下植物的GR和SOD也是增加的(Alonso et al., 2001; Huseynova et al., 2014), 而与O3和干旱的单独胁迫相比, 两者交互作用下抗氧化酶类反而是降低的, 从而推断两种胁迫的累积作用破坏了植物的防御体系(Wellburn et al., 1996; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
... 和干旱的单独胁迫相比, 两者交互作用下抗氧化酶类反而是降低的, 从而推断两种胁迫的累积作用破坏了植物的防御体系(Wellburn et al., 1996; Alonso et al., 2001).因此, 植物对O3与干旱的去氧化响应取决于两种胁迫诱导产生的抗氧化分子和酶类的多少.但目前仍缺乏转录水平与AsA-GSH循环中基因调控的蛋白酶活性的相关性研究(Creissen & Mullineaux, 2002; Iyer et al., 2013). ...
Drought and air pollution affect nitrogen cycling and free radical scavenging in Pinus halepensis (Mill). 1996
O3和干旱胁迫对元宝枫叶片气孔特征的复合影响 2 2014
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Ozone suppresses soil drying- and abscisic acid (ABA)-induced stomatal closure via an ethylene-dependent mechanism. 1 2009
... O3和干旱交互作用下, 干旱诱导的气孔关闭被认为是保护植物免受O3伤害的防御措施.干旱条件下种植园的西黄松(Pinus ponderosa)也显示O3伤害的短期保护效应(Panek & Goldstein, 2001).然而, 这一现象尚未得到普遍证实.尽管干旱诱导的气孔关闭降低了植物对O3的吸收, 但干旱胁迫同时也增加了叶片的自由基(Buckland et al., 1991; Biehler & Fock, 1996), 使植物受到伤害, 因而干旱和O3的复合胁迫对植物可能是有害的(Heber et al., 1995).自然环境中O3与干旱对气孔的交互作用取决于品种(Pell et al., 1993; Ribas et al., 2005; Biswas & Jiang, 2011; Wagg et al., 2012)、胁迫出现次序(Bohler et al., 2013)、胁迫程度(Le Thiec et al., 1994)、一天内胁迫的时段(Le Thiec et al., 1994)、植物的生长期 (Alonso et al., 2001; Skärby et al., 1998)以及季节的变化(Pell et al., 1993).对O3敏感的冬小麦品种, 在O3和干旱的交互作用下显示出对O3的抗性; 然而另一抗性品种(Triticum turgidum ssp. durum)则丧失了对O3的抗性(Biswas & Jiang, 2011).两种胁迫发生的次序对交互作用也至关重要(Le Thiec et al., 1994; Bohler et al., 2013).如果干旱发生在O3胁迫之前, 干旱首先促发气孔导度关闭, 进而保护了随后发生的O3对植物的伤害.有实验证实O3暴露下的Leontodon hispidus施用外源脱落酸(ABA)后, 渐进的干旱胁迫会降低气孔导度(Wilkinson & Davies, 2009), 通过干旱影响气孔对ABA的敏感性和离子泵, 降低了细胞的膜损伤(Torsethaugen et al., 1999), 减轻了植物的伤害程度.而当干旱发生在O3胁迫之后, 因ABA诱导的气孔关闭被O3诱导植物释放的乙烯引起的气孔缓慢响应所抑制(Tanaka et al., 2005), 导致的气孔滞后效应加重了干旱对植物的伤害(Wilkinson & Davies, 2009).但也有科学家指出干旱诱导的ABA对气孔在O3胁迫下的敏感程度的影响仍未明确(Wilkinson & Davies, 2010). ...
Drought, ozone, ABA and ethylene: New insights from cell to plant to community. 2 2010
... 气孔是植物体与外界环境进行气体交换的门户, 通过控制CO2和水分对蒸腾、光合、呼吸等重要生理过程进行调节, 也是诸如O3等气体污染物进入植物的主要通道.气孔调节的首要功能是平衡与碳收支相关的水分消耗(Cowan, 1978).植物应对O3和干旱胁迫的最初防御门户就是气孔.O3在通过气孔进入植物的同时伴随水汽的蒸发, 气孔关闭的响应机制理论上会保护植物抵抗这两种胁迫, 形成物理防御抵制O3吸收和水汽的散失(图1).渐进式(pro- gressive)的土壤干旱阻碍了植物对O3的吸收(Panek et al., 2002; Matyssek et al., 2006).Ciais等(2005)的结果显示, 即使O3浓度在增加了40%的严重胁迫下, 由于初夏的干旱迫使气孔关闭, 植物对O3的吸收仍低于潮湿年份的平均吸收值.据此一些田间限制性灌溉的保护措施也在实际中得到应用.例如古巴农民在预防夏季O3浓度升高对作物的毒害方面, 通过提前1-2天控制对叶类作物(莴苣(Lactuca sativa)和烟草(Nicotiana tabacum))的灌溉, 成功地降低了O3带来的伤害(ICP Vegetation, 2011).然而, 此种方法仅针对一些在干旱胁迫下表现出诸如气孔关闭和叶表面积减少的品种才是有效的(ICP Vegetation, 2011).在大气和土壤都潮湿的环境中, 即使暴露在低浓度O3下, 通过气孔进入到植物的O3通量也是较高的; 而在水分缺乏的环境中, 即使在高浓度O3条件下, 干旱诱导的气孔关闭仍极大地限制了植物对O3的吸收(Retzlaff et al., 2000; Paoletti, 2006).而且干旱对气孔导度和O3吸收的限制有记忆效应(memory-effects) (Karlsson et al., 2000).可见气孔内在行为机制的改变可能是为了降低O3胁迫引起的植物细胞壁木质化和膜损伤(Heath & Taylor, 1997; Maier-Maercker, 1998).但干旱条件下的气孔关闭虽然降低了植物对O3的吸收, 同时也限制了植物对CO2的吸收(Panek & Goldstein, 2001), 进而影响植物生长.气孔的响应(关闭)也并不是始终如一的(Wittig et al., 2007), 如O3诱发的气孔反应滞后(sluggish)(Paoletti & Grulke, 2010; Hoshika et al., 2012, 2014; Dumont et al., 2013)所导致的气孔关闭缓慢(Pearson & Mansfield, 1993; Karlsson et al., 1995)或气孔导度增大(Oksanen, 2003), 降低了气孔在干旱条件下对蒸腾的抑制, 从而扰乱了植物对水分亏缺的响应(Hoshika et al., 2014), 导致O3和干旱交互作用下植物的伤害加重(Retzlaff et al., 2000; Pollastrini et al., 2010).这种气孔在时间上的滞后效应与植物物种品种(Paoletti & Grulke, 2010; Hoshika et al., 2012)、胁迫的程度(Hoshika et al., 2012)及季节的变化有关(Hoshika et al., 2014). ...
... O3和干旱都会引起光合速率的降低并由此影响净CO2的固定(Flexas et al., 2002; Wittig et al., 2007; Biswas & Jiang, 2011), 而在O3胁迫之后发生的干旱加重了对植物净同化率和光合效率的伤害(Grulke et al., 2002), 导致生物量降低.生物量的降低包括种子质量和数量的降低(Flexas et al., 2002; Biswas & Jiang, 2011), 且O3和干旱同时发生对植物生物量降低是累积效应(Biswas & Jiang, 2011).不同的植物对胁迫的响应是不同的, 且干旱对植物的影响可能更大.水分胁迫对夏栎的伤害影响并不明显, 对欧洲山毛榉甚至没有影响, 但降低了杨树(Pop- ulus maximowiczii × P. berolinensis)的生长, 包括茎粗和生物量, 而且O3和干旱的交互作用加剧了对杨树叶绿素荧光参数和生长的伤害(Pollastrini et al., 2010).即使高浓度O3促进土壤呼吸, 增加了细根的生长, 但是在干旱胁迫下欧洲云杉根系的生长仍然受到限制, 而山毛榉却未有此现象, 可能对云杉而言干旱更占主导地位(Nikolova et al., 2010).在德国克兰茨贝格的森林实验点也证实干旱的影响可能更大.O3对山毛榉树的影响也因水分亏缺而大大降低, 对茎粗和整个茎杆体积增量的有害影响很大程度上是因干旱造成的(Matyssek et al., 2010). ...
To what extent do current and projected increases in surface ozone affect photosynthesis and stomatal conductance of trees? A meta- analytic review of the last 3 decades of experiments. 2 2007
... 21世纪快速的都市化、工业化和交通运输使地表臭氧(O3)浓度逐渐增加, 世界各地出现不同程度的O3污染.其中, 英国、法国、德国、瑞士和意大利等多个欧洲国家持续10天的O3浓度都超过了90 nmol·mol-1 (ICP Vegetation, 2011), 亚洲和拉丁美洲等快速发展中国家的O3平均浓度也超过40 nmol·mol-1 (Matyssek et al., 2014).在中国人口密集的特大城市(如北京、上海、广州等) O3浓度超过100 nmol·mol-1 (Chang & Lee, 2006; Li et al., 2011), 预计到2100年, 全球大部分地区的年平均O3浓度将达到40 nmol·mol-1以上, 夏季北半球地区甚至将超过70 nmol·mol-1 (Sitch et al., 2007).地表O3作为具有植物毒性的空气污染物, 对自然植物种群、作物和森林的危害被广泛关注(Matyssek & Innes, 1999; Ashmore, 2005; Matyssek et al., 2012).长时间暴露于O3下的敏感植物会出现表观可见的叶片伤害症状(Mills et al., 2011; Feng et al., 2014), 并伴随光合速率下降(Dizengremel, 2001; Renaut et al., 2009; Wittig et al., 2009), 生长缓慢及早衰(Matyssek & Sandermann, 2003), 进而削弱植物的抗逆性、竞争力和适应力, 最终降低碳汇潜力(Miller & McBride, 1999; Karnosky et al., 2007; Edwards & Zak, 2011), 对生态系统的结构、功能和碳循环产生长远影响.现有的O3污染可导致森林树种生物量损失达10%, 农作物产量的损失为5.3%-17.5% (Broadmeadow, 1998; Feng & Kobayashi, 2009).至2030年, 全球农作物因O3的影响年经济损失将高达170-350亿美元(Avnery et al., 2011). ...
... 目前, O3和干旱对植物的影响存在如下有争议的结论: 一方面, O3引起的气孔滞后效应加重了植物在干旱胁迫下的水分蒸散损失, 从而对植物产生协同伤害(Grulke et al., 2003b; Karnosky et al., 2005); 另一方面, 干旱诱导气孔关闭, 限制进入叶片的O3剂量, 起到保护植物的拮抗作用, 降低了O3伤害(Panek & Goldstein, 2001; Panek et al., 2002; Grünhage & Jager, 2003; Dizengremel et al., 2013); 也有一些研究显示二者胁迫下植物并不受交互作用的影响(Le Thiec et al., 1994; Wittig et al., 2009).在O3暴露下, 干旱对植物的保护作用仅限于叶片水平, 但对整株植物而言, 干旱的伤害大于O3的伤害(Alonson et al., 2014; Pollastrini et al., 2014).此外, 由于O3刺激糖基酶类基因表达促进了葡聚糖代谢, 而干旱则是抑制其代谢, 导致两种胁迫下上述代谢过程并没有受到影响(Iyer et al., 2013), 减轻了植物在交互作用下所受到的伤害.因此, O3和干旱对植物的伤害不仅取决于O3进入细胞的量(Heath, 1994), 而且与植物通过酶类和非酶类反应去除氧自由基的能力(Manderscheid et al., 1991)及自身防御和修复过程的活跃度有关(Sandermann, 1996). ...
Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: A quantitative meta-analysis. 2009
Photosynthesis and yield responses of ozone- polluted winter wheat to drought. 1 2007
... 植物受到胁迫时, 细胞内部需要复杂的调控体系, 包括活性氧(ROS)的信号传导、产生及清除来控制其受到的毒害作用.因此, 植物进行氧化应激平衡ROS的活性和抗氧化系统的能力至关重要.O3和干旱胁迫对植物的影响最主要是诱导氧化应激的不同, O3会进入叶片内部并迅速转换为ROS, 导致氧化胁迫(Rao et al., 2000; Ernst et al., 2012), 虽然严重的干旱也可导致氧化胁迫(Foyer & Noctor, 2000), 但干旱胁迫下的植物应激更多是利用ROS作为内部产生信号的分子(Yao et al., 2013).因此, O3胁迫下ROS会不断累积, 而在干旱胁迫下ROS只是作为逆境胁迫的响应因子存在于叶绿体中.双重胁迫诱导大量ROS产生, 使许多细胞成分(如细胞液、蛋白质、碳水化合物、核酸等)受到氧化伤害, 进而激发了与防御有关的蛋白进行的解毒作用(Pääkkönen et al., 1998), 其中高活性的清除氧化伤害的酶类保护了植物免受胁迫伤害(Asada, 1997; Pasqualini et al., 2001). ...
Drought-induced H2O2 accumulation in subsidiary cells is involved in regulatory signaling of stomatal closure in maize leaves. 2013
Effects of ozone and drought on biomass allocation of four seedlings in South China. 2014
b). The influences of ozone and soil water stress, singly and in combination, on leaf gas exchange rates, leaf ultrastructural characteristics and annual ring width of Fagus crenata seedlings. 2001
Effects of ozone and/or soil water stress on growth and photosynthesis of Fagus crenata seedlings. 2001
Carry-over effects of ozone and water stress on leaf phenological characteristics and bud frost hardiness of Fagus crenata seedlings. 1 2004
... 抗氧化系统在植物对O3和干旱胁迫的抗性中发挥重要的作用, O3增加了抗氧化物的浓度和抗氧化酶的活性(Alonso et al., 2001; Herbinger et al., 2005)且增强了抗氧化系统相关的基因表达(Gupta et al., 2005).虽然AsA含量与植物的抗性密切相关, 如烟草中DHAR过量表达也证实了增加的AsA含量增强了其对O3和干旱的抗性(Chen & Gallie, 2005; Eltayeb et al., 2006), 但也有研究指出两种白车轴草(Trifolium repens)质外体中的AsA含量差异无法说明O3的抗性差异(D’Haese et al., 2005).干旱胁迫也增加了植物叶片抗氧化物质的浓度或改变其氧化还原态(Šircelj et al., 2005; Talbi et al., 2015).然而当干旱达到一定程度时, 抗氧化物的活性却降低, 从而使植物受到伤害(Quartacci & Navari-Izzo, 1992; Zhang & Kirkham 1994; Alonso et al., 2001).因此, O3和干旱胁迫下植物的抗氧化响应是多变的, 因植物品种、胁迫时间及胁迫强度各异, 其内在的作用机制也需进一步明确. ...
Drought-stress-induced chang- es in activities of superoxide-dismutase, catalase, and peroxidase in wheat species. 1 1994
... AsA是AsA-GSH循环中一种主要的抗氧化剂, 直接与羟基自由基、超氧化物和单线态氧反应, 在光合作用调节中也起重要作用(Noctor & Foyer, 1998), 并作为电子供体将H2O2还原为水, 而AsA的生成需要NADPH的还原力, 同时Rubisco活性降低, PEPC活性升高.质外体系统(质外体AsA和APX)是对氧化伤害的第一层防御体系(Luwe et al., 1993; Sanmartin et al., 2003).植物对O3的敏感性也与叶片组织AsA的氧化还原态有关(Conklin & Barth, 2004; Chen & Gallie, 2005).另外, AsA也是植物细胞质、叶绿体、液泡、线粒体和质外体中主要的氧化还原缓冲剂(Potters et al., 2002; Pignocchi et al., 2003).总AsA水平降低了由ROS对细胞造成的伤害程度, 与植物的耐旱性有重要关系, 而DHAR的活性增加与干旱胁迫的程度也有关(Sofo et al., 2005).干旱条件下, 细胞质和叶绿体中的AsA发挥重要的解毒作用, 且主要取决于植物的种类(Mittler & Zilinskas, 1994; Zhang & Kirkham, 1996).欧洲云杉(Picea abies)针叶中总AsA含量因O3胁迫而增加, 同时干旱胁迫使其质外体AsA也显著增加, 且在共同胁迫下两者都是显著增加的(Kronfuß et al., 1998).因此, O3和干旱复合胁迫可能增加了还原力, 同时促进了氧化胁迫下植物的防护作用. ...
Enzymatic responses of the ascorbate-glutathione cycle to drought in sorghum and sunflower plants. 1 1996
... 随着人口增长以及农业、能源和工业部门的扩增, 对水的需求也迅猛增加, 近年来不断发生的大规模密集干旱事件在欧洲、非洲、亚洲、澳大利亚、南美洲、中美洲和北美洲产生了广泛影响(Comte, 1998; Schubert et al., 2004; Dai, 2011).1950-2010年中国平均每年的受灾面积达到了21.6 × 106 hm2 (邱海军等, 2013).干旱对植物的影响表现为: 降低气孔导度、影响水分蒸散(Sadras & Milroy, 1996; Bréda et al., 2006), 改变叶片光系统组分含量(Anjum et al., 2003), 损害光合反应器官(Fu & Huang, 2001), 降低卡尔文循环的酶活性(Monakhova & Chernyadèv, 2002), 使植物的细胞分裂和增长受损(Nonami, 1998), 限制叶片的数量, 缩减叶片的伸张, 降低比叶面积(Abrams et al., 1994), 从而限制植物生长, 最终导致产量降低(Schuppler et al., 1998; Monakhova & Chernyadèv, 2002; Kaya et al., 2006).中国在2006-2011年间, 由干旱造成的直接经济损失达到1150亿元(中国水利部, 2006-2010; 国家统计局, 2007-2012).气候变化导致的干旱使全球森林在2000-2009年间减少了5.5亿T净初级生产力(Zhao & Running, 2010). ...
Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. 2010

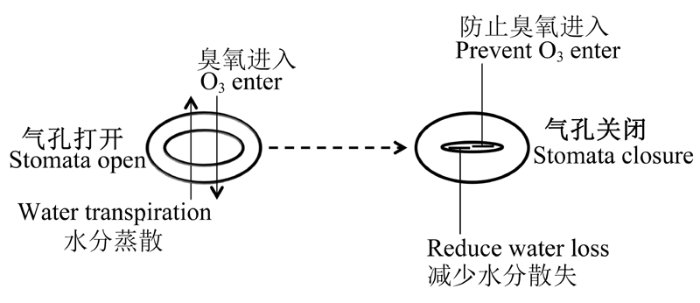 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}