 , 杨玉盛
, 杨玉盛福建师范大学地理科学学院, 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007
Interactive effects of soil warming and nitrogen addition on fine root production of Chinese fir seedlings
SHIShun-Zeng, XIONGDe-Cheng, DENGFei, FENGJian-Xin, XUChen-Sen, ZHONGBo-Yuan, CHENYun-Yu, CHENGuang-Shui, YANGYu-Sheng通讯作者:
收稿日期:2016-09-6
接受日期:2016-11-10
网络出版日期:2017-02-10
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Methods A mesocosm experiment, with a factorial design of soil warming (ambient, +5 °C) and nitrogen addition (ambient, ambient + 40 kg·hm-2·a-1, ambient + 80 kg·hm-2·a-1), was carried out in the Chenda State-owned Forest Farm in Sanming City, Fujian Province, China. Fine root production (indexed by the number of fine roots emerged per tube of one year) was measured biweekly using minirhizotrons from March of 2014 to February of 2015.
Important findings (1) The two-way ANOVA showed that soil warming had a significant effect on fine root production, while nitrogen addition and soil warming × nitrogen addition had no effect. (2) The three-way ANOVA (soil warming, nitrogen addition and diameter class) showed that soil warming, diameter class and soil warming × diameter class had significant effects on fine root production, especially for the number of fine roots in 0-1 mm diameter class that had been significantly increased by soil warming. Compared with the 1-2 mm roots, the 0-1 mm roots seemed more flexible. (3) Repeated measures of ANOVA (soil warming, nitrogen addition and season) showed that soil warming, season, soil warming × season, and soil warming × nitrogen addition × season had significant effects on fine root production. In spring, the number of fine roots was significantly increased both by soil warming and soil warming × season, while soil warming, nitrogen addition, soil warming × nitrogen addition significantly decreased fine root production in the summer. (4) Soil warming, soil layer, soil warming × soil layer had significant effects on fine root production. The number of in-growth fine roots was significantly increased by soil warming at the 20-30 cm depth only. It seemed that warming forced fine roots to grow deeper in the soil. In conclusion, soil warming significantly increased fine root production, but they had different responses and were dependent of different diameter classes, seasons and soil layers. Nitrogen addition had no effect on fine root production. Only in spring and summer, soil warming and nitrogen addition had significant interactive effects.
Keywords:
-->0
PDF (5656KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
据国际气候变化委员会(IPCC)第五次评估报告预测: 到21世纪末, 全球地表平均增温0.3-4.8 ℃ (IPCC, 2013)。大量研究表明, 温度升高可能直接或间接地影响森林生态系统的地上和地下生态过程(Way & Oren, 2010)。由于化石燃料的使用和农业施肥的增加(Davidson, 2009), 预计到21世纪末全球氮沉降速率将增加2-3倍(Lamarque et al., 2005)。我国中东部森林生态系统氮沉降已超过20 kg·hm-2·a-1, 成为全球三大氮沉降区之一, 并呈逐渐加重的趋势(Liu et al., 2011)。因而, 研究土壤增温和氮沉降对森林生态系统的影响对维持生态系统生产力和森林碳汇储量等具有重要的意义。
细根(直径≤2 mm)是水分和养分吸收的主要器官, 也是根系中活跃性和敏感度最高的部分(Hendricks et al., 1993; Nadelhoffer, 2000); 同时, 细根周转又是土壤碳输入的主要途径。目前关于增温对不同的陆地生态系统细根生物量和生产的影响有大量的研究, 但是并没有一致的结论。Wan等(2004)通过空气增温的方法发现增温促进了Acer rubrum和A. saccharum幼苗细根的生产。Bai等(2010)对中国北方半干旱温带草原研究发现, 增温后细根平均年生产量降低。Wan等(2005)对高秆草(tallgrass)的研究表明0-5、5-15和15-30 cm土层中增温与对照处理之间细根生物量均没有显著差异。由于森林生态系统类型、林龄和系统氮水平等因素不同, 各生态系统细根对氮沉降的响应存在很大的不确定性, 以往研究表明模拟氮沉降或施肥可能对森林细根生物量有促进、抑制作用或无影响(Li et al., 2015)。Liu和Greaver (2010)针对已发表的110篇文献进行了meta分析, 其中涵盖10种类型的生态系统, 添加氮肥的范围为10-650 kg·hm-2·a-1, 分析显示氮添加总体上使根的生物量增加, 但对细根生产量没有影响, 仅温带针叶林和苔原细根生产量在氮添加的情形下各自显著地增加了9%和74%。Leppälammi-Kujansuu等(2013)对挪威云杉(Picea abies, 又称欧洲云杉)的长期增温和施肥试验表明, 土壤中出现了大量的活根和死根, 表明细根的生产率(量)和死亡率(量)都增加, 增温、施肥及其交互作用将会显著地增加细根生物量, 导致地下凋落物生产量增加了3-4倍。与气候相关的环境因子都是协同作用的, 但目前有关增温和氮沉降的交互作用对生态系统(尤其是地下部分)的影响的研究仍比较缺乏(Leppälammi- Kujansuu et al., 2014)。
中国亚热带被称为“回归带上的绿洲”, 是全球气候变化的敏感地带。杉木(Cunninghamia lanceolata)是中国南方重要的造林和用材树种, 被广泛种植, 面积达1239.1×104 hm2, 蓄积量为47357.33 × 104 m3, 分别占全国人工林面积和蓄积量的26.55%和46.89%, 在中国人工林中占有重要地位(中华人民共和国国家林业局, 2005)。为此, 本研究通过模拟增温和氮沉降试验, 研究土壤增温和氮添加对杉木幼苗细根生产力的影响, 以期为进一步揭示全球环境变化下杉木人工林生产力和碳吸存的变化提供基础数据。
1 试验区概况和研究方法
1.1 试验区概况
试验区位于福建三明森林生态系统与全球变化研究站陈大观测点(26.32° N, 117.60° E)。平均海拔300 m, 属中亚热带季风气候, 年平均气温19.1 ℃, 年降水量1749 mm (主要集中在3-8月), 年蒸发量1585 mm, 相对湿度81%。该研究区土壤以花岗岩发育的红壤和黄壤为主。1.2 试验设计
试验采用完全随机设计, 依据IPCC第五次评估报告对21世纪末全球地表平均增温的预测(IPCC, 2013)和该研究区氮沉降背景值36 kg·hm-2·a-1, 设置对照(CT, 不增温、不添加额外氮素的处理), 增温(W, +5 ℃, 不添加额外氮素), 低氮(LN, +40 kg·hm-2·a-1, 指自然氮沉降背景下, 再额外添加的氮素,下同), 高氮(HN, +80 kg·hm-2·a-1), 增温低氮(WLN, +5 ℃, +40 kg·hm-2·a-1), 增温高氮(WHN, +5 ℃, +80 kg·hm-2·a-1) 6个处理, 每个处理5个重复, 小区面积2 m × 2 m。试验小区四周采用4块PVC板(200 cm × 70 cm深)焊接而成, 与周围土壤隔开, 防止小区之间相互干扰。小区土壤取自附近的杉木林土壤, 按0-10 cm、10-20 cm、20-70 cm分层取回, 剔除根系、石块和其他杂物后, 分层混合均匀, 并按20-70 cm、10-20 cm和0-10 cm重填回2 m × 2 m试验小区内, 同时采用压实法调整土壤容重与原位土壤容重接近。于2013年10月安装加热电缆(所有小区都布设相同电缆), 平行布设, 深度为10 cm, 间距20 cm, 并在最外围环绕一圈, 保证样地增温的均匀性。同时在实验布设完成后在每个小区布设4个温度传感器(T109, Campbell Scientific, Logan, USA)和4个水分传感器(EC-5, Decagon, Pullman, USA), 其中2个温度传感器埋在10 cm处, 另外2个温度传感器分别埋在5 cm和20 cm深土壤中; 2个水分传感器测定土壤10 cm处的含水量,另外2个土壤水分传感器监测5 cm和20 cm深处的土壤湿度。温度和水分传感器均布设在两条电缆线中间位置。增温技术环节详细描述见参考文献(陈仕东等, 2013)。2013年11月, 每个2 m × 2 m小区均匀种植4株一年生二代半短侧枝杉木幼苗, 杉木位置均处于两条电缆线之间。在每个小区中的2株杉木幼苗正下方布设两根微根管用以监测杉木细根生长和死亡动态, 微根管长度为90 cm, 微根管与地面呈45°角插入土壤中, 换算成垂直深度约为40 cm。样地布设完成后, 于2014年3月开始通电增温。并于2014年3月开始对氮添加处理施氮肥(NH4NO3, 分析纯), 每月初以溶液的形式对小区喷洒, 全年共喷洒12次。按照处理水平要求, 将每个小区每次所需要喷洒的NH4NO3溶解在800 mL (相当年降水量增加约2 mm)去离子水中(LN和WLN处理喷洒溶液浓度约为1.67 g·L-1, HN和WHN处理喷洒溶液浓度约为3.33 g·L-1), 用手提式喷雾器在小区四周从幼苗林冠上方对小区均匀喷洒。其他小区喷洒等量的去离子水。同时定期清除小区内自然萌发的其他植物。1.3 根系影像采集和数据采集
从2014年3月至2015年2月对细根进行了为期1年的连续性观测。于每月上旬和下旬(间隔大约15天)使用微根管影像成像系统BTC-2 Minirhizotron Camera System (Bartz Technology, Santa Barbara, USA)对细根生长过程进行影像收集, 在1天内完成每次影像的收集工作。图片收集后带回实验室用Rootfly图像分析软件对其进行处理, 按照0-10 cm、10-20 cm、20-30 cm、30-40 cm进行分层处理, 并获得每张影像中细根直径、细根长度和细根数量等数据。同时鉴别活根和死根: 白色根、白色带红色斑点或红色根定义为活根; 暗黑色、皮层脱落或表皮褶皱的根定义为死根(Huang et al., 2016)。本研究中, 细根生产量用影像中每根管细根一年总出生数量作为指标。观测期间的土壤增温和对照小区的土壤温度和含水量变化见图1。土壤增温对不同深度土壤增温的影响见参考文献(章宪等, 2014)。2015年1月, 采用土钻法在每个小区随机取4个土样, 土钻直径3.5 cm, 测定0-20 cm土壤的铵态氮和硝态氮(图2)。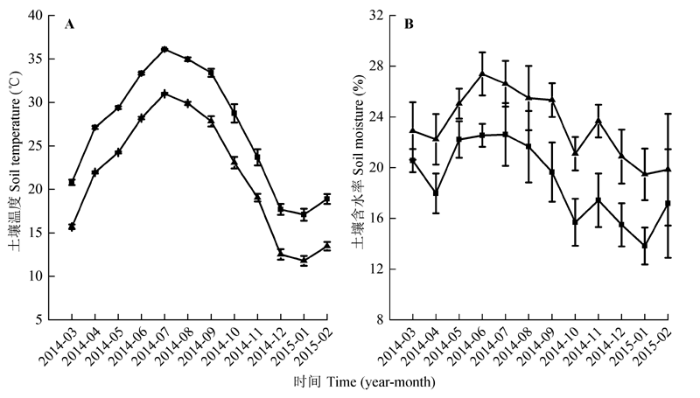 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1不同处理(以对照(不增温、不添加额外氮素的处理, ▲)和土壤增温处理(+5 ℃、不添加额外氮素的处理, ■)为例) 0-10 cm土壤温度(A)和含水率(B)的年变化(平均值±标准偏差)。
-->Fig. 1Annual changes of soil (0-10 cm) temperature (A) and moisture (B) under different treatments (mean ± SD). ▲, control treatment (ambient, ambient); ■, soil warming treatment (+5 °C, ambient).
-->
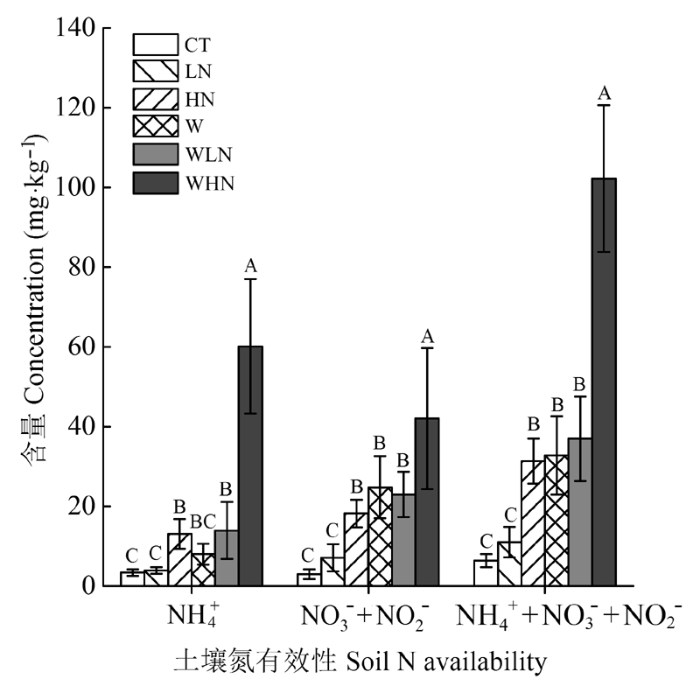 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2不同处理0-20 cm土壤氮有效性(平均值±标准偏差)。NH4+, 铵态氮。NO3- + NO2-, 硝态氮。CT, 对照(不增温、不添加额外氮素); HN, 高氮(不增温, +80 kg·hm-2·a-1, 指自然氮沉降背景下, 再额外添加的氮素量,下同); LN, 低氮(不增温, +40 kg·hm-2·a-1); W, 土壤增温(+5 ℃, 不添加额外氮素); WHN, 土壤增温+高氮(+5 ℃, +80 kg·hm-2·a-1); WLN, 土壤增温+低氮(+5 ℃, +40 kg·hm-2·a-1)。不同大写字母表示不同处理间差异显著(p < 0.05)。
-->Fig. 2Soil (0-20 cm) nitrogen availability under different treatments (mean ± SD). NH4+, ammonium nitrogen. NO3- + NO2-, nitrate nitrogen. CT, control treatment (ambient, ambient); HN, high nitrogen addition (ambient, ambient + 80 kg·hm-2·a-1); LN, low nitrogen addition (ambient, ambient + 40 kg·hm-2·a-1); W, soil warming (+5 °C, ambient); WHN, soil warming plus high nitrogen addition (+5 °C, ambient + 80 kg·hm-2·a-1); WLN, soil warming plus low nitrogen addition (+5 °C, ambient + 40 kg·hm-2·a-1). Different capital letters indicate significant differences among treatments (p < 0.05).
-->
1.4 数据处理和分析
试验观测期从2014年3月至2015年2月, 依据亚热带气候条件, 将3-5月设为春季, 6-8月设为夏季, 9-11月设为秋季, 12月和翌年1、2月设为冬季。采用双因素方差分析检验土壤增温、氮添加对细根生产量的影响; 采用重复测量方差分析检验土壤增温、氮添加、季节对细根生产量的影响; 采用三因素方差分析检验土壤增温、氮添加、径级或土层对细根生产量的影响。采用单因素方差分析和LSD检验各处理间的细根生产量差异。所有的统计分析均在SPSS 20.0软件上进行, 显著性水平设定为p = 0.05。
2 结果
2.1 土壤微环境
增温对土壤温度和水分的影响均达到极显著水平(p < 0.01)(表1)。与CT相比, W处理土壤温度显著增加(p < 0.05) (图1A), 土壤含水率则显著下降(p < 0.05)(图1B)。增温和氮添加对铵态氮、硝态氮和土壤有效氮的影响均达到极显著水平(p < 0.01); 增温和氮添加的交互作用对铵态氮和土壤有效氮的影响达到极显著水平(p < 0.01), 对硝态氮没有影响(p > 0.05)(表1)。处理间比较可知, WHN的铵态氮显著高于其他处理(p < 0.01), HN和WLN处理的铵态氮均显著高于CT和LN处理(p < 0.01); WHN的硝态氮和土壤有效氮均显著高于其他处理(p < 0.01), 而HN、W和WLN处理的硝态氮和土壤有效氮显著高于CT和LN (p < 0.01)(图2)。Table 1
表1
表1土壤增温、氮添加及其交互作用对土壤温度、含水率和土壤有效氮影响的双因素方差分析p值
Table 1p-value of two-way ANOVA on the effects of soil warming, nitrogen addition and their interaction on soil temperature, soil moisture and soil nitrogen availability
| 因子 Factor | 土壤温度 Soil temperature (℃) | 土壤湿度 Soil moisture (%) | 铵态氮 Ammonium nitrogen | 硝态氮 Nitrate nitrogen | 土壤有效氮(铵态氮和硝态氮) Soil nitrogen availability (ammonium nitrogen and nitrate nitrogen) |
|---|---|---|---|---|---|
| W | <0.001 | 0.005 | <0.001 | <0.001 | <0.001 |
| N | <0.001 | <0.001 | <0.001 | ||
| W × N | <0.001 | 0.317 | <0.001 |
新窗口打开
2.2 细根生产量
土壤增温对细根生产量有显著的促进作用(p < 0.05), 而氮添加、土壤增温和氮添加的交互作用对细根生产量没有影响(p > 0.05) (表2)。W处理的细根生产量显著高于LN和HN处理(p < 0.05); WHN处理细根生产量显著高于HN处理(p < 0.05); 其他处理之间均无显著差异(p > 0.05)(图3A)。Table 2
表2
表2土壤增温、氮添加及其交互作用对每根管细根一年总出生数量影响的双因素方差分析p值
Table 2p-value of two-way ANOVA on the effects of soil warming, nitrogen addition and their interaction on total number of fine roots emerged per tube of one year
| 指标 Index | 因子 Factor | ||
|---|---|---|---|
| W | N | W × N | |
| 每根管细根一年总出生数量 Total number of fine roots emerged per tube of one year (No.·tube-1·a-1) | 0.005 | 0.616 | 0.483 |
新窗口打开
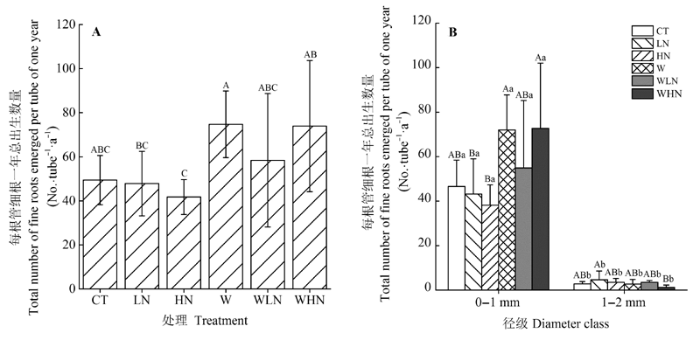 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3不同处理每根管细根一年总出生数量(A)和细根径级分布(B)(平均值±标准偏差)。不同大写字母表示不同处理差异显著(p < 0.05), 不同小写字母表示相同处理不同径级间差异显著(p < 0.05)。CT, 对照(不增温、不添加额外氮素); HN, 高氮(不增温, +80 kg·hm-2·a-1, 指自然氮沉降背景下, 再额外添加的氮素量,下同); LN, 低氮(不增温, +40 kg·hm-2·a-1); W, 土壤增温(+5 ℃, 不添加额外氮素); WHN, 土壤增温+高氮(+5 ℃, +80 kg·hm-2·a-1); WLN, 土壤增温+低氮(+5 ℃, +40 kg·hm-2·a-1)。
-->Fig. 3Total number of fine roots emerged of one year (A) under different treatments and number of different diameter class (B) (mean ± SD). Different capital letters indicate significant differences among treatments (p < 0.05). Different lowercase letters indicate significant differences among diameters (p < 0.05). CT, control treatment (ambient, ambient); HN, high nitrogen addition (ambient, ambient + 80 kg·hm-2·a-1); LN, low nitrogen addition (ambient, ambient + 40 kg·hm-2·a-1); W, soil warming (+5 °C, ambient); WHN, soil warming plus high nitrogen addition (+5 °C, ambient + 80 kg·hm-2·a-1); WLN, soil warming plus low nitrogen addition (+5 °C, ambient + 40 kg·hm-2·a-1).
-->
2.3 细根生产量的径级分布
土壤增温、径级、土壤增温和径级的交互作用对细根生产量有显著影响(p < 0.05), 但氮添加、土壤增温和氮添加的交互作用、氮添加和径级的交互作用, 以及三者的交互作用对细根生产量没有影响(p > 0.05)(表3)。在0-1 mm径级中, 土壤增温对细根生产量有显著的促进作用(p < 0.05)(表4); 各处理间比较发现, W和WHN处理细根生产量显著高于LN和HN处理(p < 0.05)(图3B); 而在1-2 mm径级中, 土壤增温、氮添加及其交互作用对细根生产量均没有显著的影响(p > 0.05)(表4); 只有WHN处理细根生产量显著低于LN处理(p < 0.05)(图3B)。在所有处理中, 0-1 mm径级细根生产量均极显著高于1-2 mm径级(p < 0.01)(图3B)。Table 3
表3
表3土壤增温、氮添加和径级对每根管细根一年总出生数量影响的方差分析p值
Table 3p-value of ANOVA on the effects of soil warming, nitrogen addition and diameter class on total number of fine roots emerged per tube of one year
| 指标 Index | 因子 Factor | ||||||
|---|---|---|---|---|---|---|---|
| W | N | D | W × N | W × D | N × D | W × N × D | |
| 每根管细根一年总出生数量 Total number of fine roots emerged per tube of one year (No.·tube-1·a-1) | 0.004 | 0.624 | <0.001 | 0.491 | 0.002 | 0.44 | 0.431 |
新窗口打开
Table 4
表4
表4不同径级、不同季节、不同土层中的土壤增温、氮添加及其交互作用对每根管细根一年总出生数量影响的双因素方差分析p值
Table 4p-value of two-way ANOVA on the effects of soil warming, nitrogen addition and their interaction on total number of fine roots emerged per tube of one year in different diameter classes, seasons and soil layers
| 指标 Index | 因子 Factor | W | N | W × N | |
|---|---|---|---|---|---|
| 每根管细根一年总出生数量 Total mumber of fine roots emerged per tube of one year (No.·tube-1·a-1) | 径级 Diameter class | 0-1 mm | 0.004 | 0.535 | 0.465 |
| 1-2 mm | 0.137 | 0.182 | 0.505 | ||
| 季节 Season | 春季 Spring | <0.001 | 0.529 | 0.010 | |
| 夏季 Summer | 0.003 | 0.001 | 0.041 | ||
| 秋季 Autumn | 0.226 | 0.555 | 0.971 | ||
| 冬季 Winter | 0.702 | 0.175 | 0.313 | ||
| 土层 Soil layer | 0-10 cm | 0.547 | 0.488 | 0.423 | |
| 10-20 cm | 0.158 | 0.114 | 0.052 | ||
| 20-30 cm | 0.005 | 0.424 | 0.892 | ||
| 30-40 cm | 0.124 | 0.379 | 0.892 |
新窗口打开
2.4 细根生产量的季节动态
土壤增温、季节、土壤增温和季节的交互作用, 以及土壤增温、氮添加和季节三者的交互作用对细根生产量的影响均达到显著水平(p < 0.05)(表5)。在春季, 土壤增温、土壤增温和氮添加的交互作用对细根生产量有显著的促进作用(p < 0.05), 但氮添加对细根生产量则无显著的影响(p > 0.05)(表4); 处理间比较发现: W和WHN处理细根生产量显著高于CT和HN处理(p < 0.05)(图4); 在夏季, 土壤增温、氮添加、土壤增温和氮添加的交互作用对细根生产量有显著的抑制作用(p < 0.05)(表4); CT处理细根生产量显著高于其他处理(p < 0.05), W、LN、WLN、HN和WLN处理间均无显著差异(p > 0.05); 在秋季, 土壤增温、氮添加、土壤增温和氮添加的交互作用对细根生产量均无显著影响(p > 0.05); 不同处理间也没有显著差异(p > 0.05)(表4); 在冬季, 土壤增温、氮添加、土壤增温和氮添加的交互作用对细根生产量均无显著影响(p > 0.05)(表4); 只有WLN处理细根生产量显著高于W处理。不同处理细根出生的季节模式不同, CT处理夏季细根生产量显著高于春、秋、冬季(p < 0.05); W和LN处理春季细根生产量显著高于其他季节(p < 0.05); WLN和WHN处理春秋季细根生产量高于夏季和冬季; HN处理四个季节之间细根生产量没有显著差异(p > 0.05), 但在秋季最高(图4)。Table 5
表5
表5土壤增温、氮添加和季节对每根管细根一年总出生数量影响的重复测量方差分析的p值
Table 5p-value of repeated measures ANOVA on the effects of soil warming, nitrogen addition and season on total number of fine roots emerged per tube of one year
| 指标 Index | 因子 Factor | ||||||
|---|---|---|---|---|---|---|---|
| W | N | S | W × N | W × S | N × S | W × N × S | |
| 每根管细根一年总出生数量 Total number of fine roots emerged per tube of one year (No.·tube-1·a-1) | 0.005 | 0.616 | <0.001 | 0.483 | <0.001 | 0.193 | 0.025 |
新窗口打开
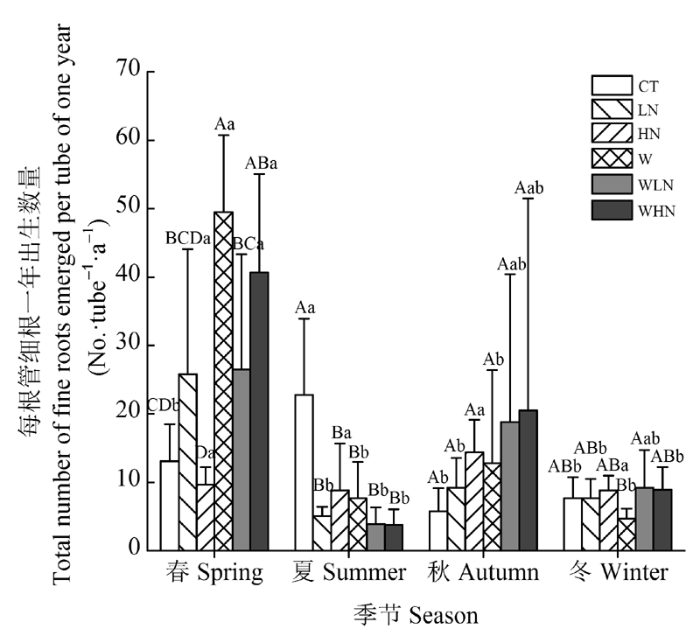 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4不同季节每根管细根一年总出生数量(平均值±标准偏差)。不同大写字母表示相同季节不同处理间差异显著(p < 0.05), 不同小写字母表示相同处理不同季节间差异显著(p < 0.05)。CT, 对照 (不增温、不添加额外氮素); HN, 高氮(不增温, +80 kg·hm-2·a-1, 指自然氮沉降背景下, 再额外添加的氮素量,下同); LN, 低氮(不增温, +40 kg·hm-2·a-1); W, 土壤增温(+5 ℃, 不添加额外氮素); WHN, 土壤增温+高氮(+5 ℃, +80 kg·hm-2·a-1); WLN, 土壤增温+低氮(+5 ℃, +40 kg·hm-2·a-1)。
-->Fig. 4Total number of fine roots emerged per tube of one year under different seasons (mean ± SD). Different capital letters indicate significant differences among treatments in the same season (p < 0.05). Different lowercase letters indicate significant differences among seasons in the same treatment (p < 0.05). CT, control treatment (ambient, ambient); HN, high nitrogen addition (ambient, ambient + 80 kg·hm-2·a-1); LN, low nitrogen addition (ambient, ambient + 40 kg·hm-2·a-1); W, soil warming (+5 °C, ambient); WHN, soil warming plus high nitrogen addition (+5 °C, ambient + 80 kg·hm-2·a-1); WLN, soil warming plus low nitrogen addition (+5 °C, ambient + 40 kg·hm-2·a-1).
-->
2.5 细根生产量的垂直分布
土壤增温、土层以及土壤增温和土层的交互作用对细根生产量有显著影响(p < 0.01)(表6)。在0-10 cm土层, 土壤增温、氮添加、土壤增温和氮添加的交互作用对细根生产量均无显著影响(p > 0.05)(表4); 不同处理间细根生产量也没有差异(p > 0.05)(图5)。在10-20 cm土层, 土壤增温、氮添加、土壤增温和氮添加的交互作用对细根生产量均无显著影响 (p > 0.05)(表4); 处理间比较, W和WHN处理细根生产量显著高于WLN和HN处理(p < 0.05)(图5)。在20-30 cm土层, 土壤增温对细根生产量有显著的促进作用(p < 0.05), 但氮添加、土壤增温和氮添加的交互作用对细根生产量无显著影响(p > 0.05)(表4); W和WHN处理细根生产量显著高于LN和HN处理(p < 0.05)(图5); 在30-40 cm土层, 土壤增温、氮添加、土壤增温和氮添加的交互作用对细根生产量均无显著影响(p > 0.05)(表4); 不同处理间细根生产量没有差异(p > 0.05)(图5)。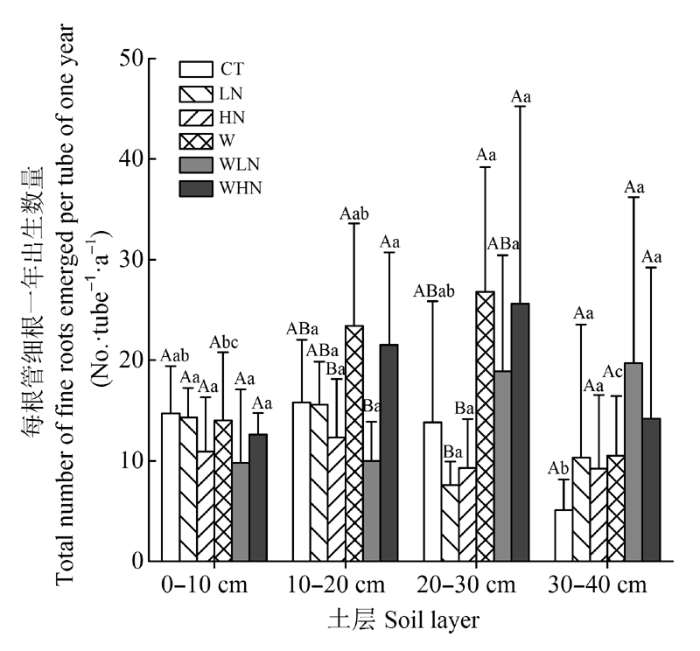 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5不同土层每根管细根一年总出生数量(平均值±标准偏差)。不同大写字母表示相同土层不同处理间差异显著(p < 0.05); 不同小写字母表示相同处理不同土层间差异显著(p < 0.05)。CT, 对照(不增温、不添加额外氮素); HN, 高氮(不增温, +80 kg·hm-2·a-1, 指自然氮沉降背景下, 再额外添加的氮素量,下同); LN, 低氮(不增温, +40 kg·hm-2·a-1); W, 土壤增温(+5 ℃, 不添加额外氮素); WHN, 土壤增温+高氮(+5 ℃, + 80 kg·hm-2·a-1); WLN, 土壤增温+低氮(+5 ℃, + 40 kg·hm-2·a-1)。
-->Fig. 5Total number of fine roots emerged per tube of one year under different soil layer (mean ± SD). Different capital letters indicate significant differences among treatments in the same soil layer (p < 0.05). Different lowercase letters indicate significant differences among soil layers in the same treatment (p < 0.05). CT, control treatment (ambient, ambient); HN, high nitrogen addition (ambient, ambient + 80 kg·hm-2·a-1); LN, low nitrogen addition (ambient, ambient + 40 kg·hm-2·a-1); W, soil warming (+5 °C, ambient); WHN, soil warming plus high nitrogen addition (+5 °C, ambient + 80 kg·hm-2·a-1); WLN, soil warming plus low nitrogen addition (+5 °C, ambient + 40 kg·hm-2·a-1).
-->
Table 6
表6
表6土壤增温、氮添加和土层对每根管细根一年总出生数量影响的方差分析p值
Table 6p-value of ANOVA on the effects of soil warming, nitrogen addition and soil layer on total number of fine roots emerged per tube of one year
| 指标 Index | 因子 Factor | ||||||
|---|---|---|---|---|---|---|---|
| W | N | L | W × N | W × L | N × L | W × N × L | |
| 每根管细根一年总出生数量 Total number of fine roots emerged per tube of one year (No.·tube-1·a-1) | 0.001 | 0.563 | 0.06 | 0.419 | 0.025 | 0.256 | 0.765 |
新窗口打开
3 讨论
3.1 土壤增温和氮添加对杉木幼苗细根生产的影响机制
增温能促进土壤温度升高, 提高土壤氮有效性, 同时也会降低土壤水分有效性, 进而促进细根生产。氮添加虽然会提高土壤氮有效性, 但对细根生产没有显著影响。从总体上看, 土壤增温和氮添加对细根生产并没有显著的交互作用(图6)。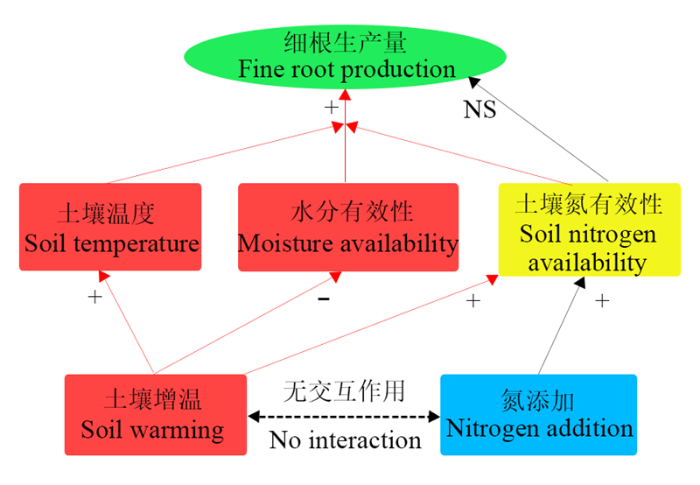 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6土壤增温和氮添加对杉木幼苗细根生产的影响机制。“+”表示促进; “-”表示抑制; “NS”表示无显著影响。红色箭头表示受土壤增温的影响; 黑色箭头表示受氮添加的影响。
-->Fig. 6Proposed mechanism on the effects of soil warming and nitrogen addition on fine root production. “+” means increase; “-” means decrease; “NS” means have no significant effect. The red arrows mean effects by soil warming; while the black arrows mean effects by nitrogen addition.
-->
3.2 细根生产量
本研究中, 土壤增温对细根生产量有显著的促进作用, 尤其是W处理细根生产量显著高于LN和HN处理, WHN处理细根生产量显著高于HN处理。Majdj和Öhrvik (2004)对瑞典北部的挪威云杉进行增温和施肥交互试验发现, 增温样地和增温+施肥样地的细根生产量显著高于对照样地和施肥样地, 主要原因是土壤增温促进了土壤矿化速率, 提高了养分有效性, 从而使细根生产量增加(Fitter et al., 1999; Wan et al., 2004)。此外, 增温对细根生产量有显著促进作用, 也有可能要归因于增温会延长细根的生长季节(Majdi & Öhrvik, 2004)。Leppälammi- Kujansuu等(2013)对挪威云杉的长期增温和施肥试验结果表明, 土壤增温后生长季节延长且土壤中氮有效性增加, 从而促进了细根的生产量增加。另有研究表明: 在降水量或者水分有效性丰富的地带, 增温会导致细根生产量和死亡量提高(Fitter et al., 1999; Wan et al., 2004)。而在水分受限制的生态系统, 如温带半干旱草原, 增温会显著降低细根生产量(Bai et al., 2010)。该研究区地处亚热带, 降水丰富, 即使在土壤增温后水分有效性也相对较高(特别是在雨季), 因而土壤增温后细根生产量较高。由于森林生态系统类型、林龄和系统背景值等因素不同, 各生态系统细根对氮沉降的响应并不明确(Li et al., 2015)。Ostertag (2001)采用土钻法对夏威夷山地以Metrosideros polymorpha为主的林分研究表明, 当土壤氮添加量增加时, <2 mm细根生产量下降。Liu和Greaver (2010)针对已发表的110篇文献进行了一个meta分析, 其中涵盖10种类型的生态系统, 添加氮肥的范围为10-650 kg·hm-2·a-1, 分析显示氮添加在总体上对细根生产量没有影响, 从不同生态系统类型来看, 温带针叶林和苔原细根生产量在氮添加后分别增加了9%和74%。本研究中氮添加对细根生产量没有显著影响可能有两个原因: (1)该研究区本底土壤氮有效性较高, 且氮添加对土壤氮有效性的影响比增温小, 因而氮添加对细根生产没有显著影响; (2)细根生产量受不同的碳分配机制的共同调控(Chen et al., 2013)。在氮受限制的生态系统(如温带和苔原生态系统), 氮有效性增加会促进林木生长, 从而促进细根生产, 以异速生长限制为主; 而在生长稳定且氮有效性较高的生态系统, 氮有效性增加可能会抑制细根生产, 以木质生产和细根生产权衡为主(Chen et al., 2013)。本研究中杉木幼苗处于个体快速生长之中, 可能受到异速生长限制、木质生产和细根生产权衡的共同限制, 导致细根生产对氮添加的响应不显著。
3.3 细根生产量的径级分布
0-1 mm细根是吸收水分和养分的主要器官, 相对于1-2 mm细根更为活跃和敏感, 各处理的0-1 mm细根生产量占细根总生产量的比重均超过90%。黄锦学等(2012)对细柄阿丁枫(Altingia grlilipes)和米槠(Castanopsis carlesii)天然林细根分布的研究也发现0-1 mm细根占0-2 mm细根总生产力的比重达到90%以上, 表明0-1 mm细根对细根生产的贡献更为重要。本研究中, 土壤增温和径级的交互作用对细根生产量有显著影响, 土壤增温只对0-1 mm径级的细根生产量有显著的促进作用, 这表明小径级的吸收根对于增温的响应更具有可塑性, 也体现了增温增强了杉木幼苗的养分获取能力。而氮添加对两个径级中的细根生产量均没有影响, 这可能是因为该研究区土壤氮有效性较高, 氮添加对细根生产的影响较小。3.4 细根生产量的季节动态
本研究发现土壤增温、氮添加及两者的交互作用对杉木幼苗细根生产的影响因季节而异。在春季, 土壤增温后幼苗细根可能会较早地进入生长季; 春季亦是细根生产的高峰, 需要大量的土壤养分, 而增温后养分矿化速率增加; 同时春季的土壤含水量高, 增温后细根生产不大可能受土壤水分限制, 这些因素综合作用使增温以及增温和氮添加的交互作用对春季细根生产有显著的促进作用(Majdi & Öhrvik, 2004)。在夏季, 土壤温度较高, 而增温后土壤温度更高, 土壤含水率亦降低, 细根生长容易受到高温和干旱限制, 因而增温对细根生产有显著的抑制作用。Bai等(2010)也指出在水分受限制的生态系统如温带半干旱草原, 细根生长对水分有效性非常敏感, 增温会显著降低细根生产量。而氮添加可能进一步加剧了杉木幼苗的生理干旱, 从而导致土壤增温和氮添加对细根生产的影响存在显著的交互作用。在秋季和冬季, 土壤增温、氮添加以及两者的交互作用对细根生产均没有显著影响, 这可能与这些季节幼苗对土壤养分、水分需求量较低, 细根生产量较低, 因此细根生产对土壤环境变化的敏感性较低有关。3.5 细根生产量的垂直分布
土壤增温、土层以及土壤增温和土层的交互作用对细根生产有显著影响, 表明土壤增温对细根的垂直分布有显著的作用, 而氮添加对细根的垂直分布没有影响。不同处理的细根峰值有不同的分布模式: 不增温处理细根生产峰值均出现在10-20 cm 土层; 而增温处理中, W和WHN处理细根生产峰值出现在20-30 cm土层, WLN处理细根生产峰值出现在30-40 cm土层, 表明增温对较深层土壤中的细根生产有促进作用。Leppälammi-Kujansuu等 (2013)对挪威云杉的试验也表明, 增温使较深层土壤中的细根生物量增加; Johnson等(2006)对花旗松(Pse- udotsuga menziesii)的研究表明增温会使细根向更深层土壤分布。这与Wu等(2014)发现增温会使高山草甸的细根有向0-10 cm表土层分布的趋势不同。进一步分析发现, 仅在20-30 cm土层, 土壤增温对细根生产量有显著的促进作用。其可能原因是: 土壤增温电缆布设在土壤10 cm处, 土壤表层的增温幅度最大, 土壤水分蒸发较快, 土壤含水量降低最多, 因而增温对表层土壤(0-10 cm, 10-20 cm)细根生产没有促进作用; 相反, 较深土层(20-30 cm)土壤含水量较高, 细根生产受水分限制小, 增温促进细根生产。但在更深土层(30-40 cm)没有发现增温对细根生产的促进作用, 这可能与该土层土壤增温幅度较低有关(章宪等, 2014)。
4 结论
(1)土壤增温对细根生产量有显著的促进作用, 土壤增温可能通过提高养分有效性使细根生产量增加。(2)土壤增温和径级的交互作用对细根生产量有显著影响, 土壤增温显著增加了0-1 mm径级的细根生产量。(3)土壤增温、季节、土壤增温和季节的交互作用, 以及土壤增温、氮添加和季节三者的交互作用对细根生产量均有显著影响。特别是春季增温后养分矿化速率增加, 且春季的土壤含水量高, 这些因素的综合作用使增温以及增温和氮添加的交互作用对春季细根生产有显著的促进作用; 在夏季, 由于土壤增温与环境高温的叠加作用以及氮添加可能进一步加剧杉木幼苗的生理干旱, 导致土壤增温、氮添加以及两者的交互作用对细根生产量有显著的抑制作用。(4)土壤增温仅对20-30 cm土层的细根生产有显著的促进作用, 表明土壤增温促使细根向更深层土壤分布。研究表明, 土壤增温促进了杉木幼苗细根生产, 但其影响因径级、季节和土层而异; 氮添加对细根生产没有影响; 土壤增温和氮添加仅在春季和夏季才存在显著的交互作用。但是本文仅以细根出生数量作为生产量的指标, 而没有考虑细根长度、体积等形态指标, 也没有对细根生产量进行转换, 对估计细根的生产量有一定的难度。致谢 感谢福建三明森林生态系统与全球变化研究站刘小飞、陈仕东、林伟盛、胥超老师在野外工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | . |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | . |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | . |
| [22] | |
| [23] | |
| [24] | |
| [25] | . |
1
2010
... 本研究中, 土壤增温对细根生产量有显著的促进作用, 尤其是W处理细根生产量显著高于LN和HN处理, WHN处理细根生产量显著高于HN处理.Majdj和Öhrvik (2004)对瑞典北部的挪威云杉进行增温和施肥交互试验发现, 增温样地和增温+施肥样地的细根生产量显著高于对照样地和施肥样地, 主要原因是土壤增温促进了土壤矿化速率, 提高了养分有效性, 从而使细根生产量增加(
Allocation of gross primary production in forest ecosystems: Allometric constraints and environmental responses.
2
2013
... 由于森林生态系统类型、林龄和系统背景值等因素不同, 各生态系统细根对氮沉降的响应并不明确(
... ).在氮受限制的生态系统(如温带和苔原生态系统), 氮有效性增加会促进林木生长, 从而促进细根生产, 以异速生长限制为主; 而在生长稳定且氮有效性较高的生态系统, 氮有效性增加可能会抑制细根生产, 以木质生产和细根生产权衡为主(
持续性主动增温对中亚热带森林土壤呼吸影响研究初报
1
2013
... 试验采用完全随机设计, 依据IPCC第五次评估报告对21世纪末全球地表平均增温的预测(
The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860.
1
2009
... 据国际气候变化委员会(IPCC)第五次评估报告预测: 到21世纪末, 全球地表平均增温0.3-4.8 ℃ (
Root production and turnover in an upland grassland subjected to artificial soil warming respond to radiation flux and nutrients, not temperature.
2
1999
... 本研究中, 土壤增温对细根生产量有显著的促进作用, 尤其是W处理细根生产量显著高于LN和HN处理, WHN处理细根生产量显著高于HN处理.Majdj和Öhrvik (2004)对瑞典北部的挪威云杉进行增温和施肥交互试验发现, 增温样地和增温+施肥样地的细根生产量显著高于对照样地和施肥样地, 主要原因是土壤增温促进了土壤矿化速率, 提高了养分有效性, 从而使细根生产量增加(
... ).Leppälammi- Kujansuu等(2013)对挪威云杉的长期增温和施肥试验结果表明, 土壤增温后生长季节延长且土壤中氮有效性增加, 从而促进了细根的生产量增加.另有研究表明: 在降水量或者水分有效性丰富的地带, 增温会导致细根生产量和死亡量提高(
Assessing the role of fine roots in carbon and nutrient cycling.
1
1993
... 细根(直径≤2 mm)是水分和养分吸收的主要器官, 也是根系中活跃性和敏感度最高的部分(
Understory fine roots are more ephemeral than those of trees in subtropical Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) stands.
1
2016
... 从2014年3月至2015年2月对细根进行了为期1年的连续性观测.于每月上旬和下旬(间隔大约15天)使用微根管影像成像系统BTC-2 Minirhizotron Camera System (Bartz Technology, Santa Barbara, USA)对细根生长过程进行影像收集, 在1天内完成每次影像的收集工作.图片收集后带回实验室用Rootfly图像分析软件对其进行处理, 按照0-10 cm、10-20 cm、20-30 cm、30-40 cm进行分层处理, 并获得每张影像中细根直径、细根长度和细根数量等数据.同时鉴别活根和死根: 白色根、白色带红色斑点或红色根定义为活根; 暗黑色、皮层脱落或表皮褶皱的根定义为死根(
中亚热带细柄阿丁枫和米槠群落细根的生产和死亡动态
2012
3
2013
... 据国际气候变化委员会(IPCC)第五次评估报告预测: 到21世纪末, 全球地表平均增温0.3-4.8 ℃ (
... 试验采用完全随机设计, 依据IPCC第五次评估报告对21世纪末全球地表平均增温的预测(
... 土壤增温、土层以及土壤增温和土层的交互作用对细根生产有显著影响, 表明土壤增温对细根的垂直分布有显著的作用, 而氮添加对细根的垂直分布没有影响.不同处理的细根峰值有不同的分布模式: 不增温处理细根生产峰值均出现在10-20 cm 土层; 而增温处理中, W和WHN处理细根生产峰值出现在20-30 cm土层, WLN处理细根生产峰值出现在30-40 cm土层, 表明增温对较深层土壤中的细根生产有促进作用.Leppälammi-Kujansuu等 (2013)对挪威云杉的试验也表明, 增温使较深层土壤中的细根生物量增加;
Elevated CO2 and elevated temperature have no effect on Douglas-fir fine-root dynamics in nitrogen-poor soil.
1
2006
... 据国际气候变化委员会(IPCC)第五次评估报告预测: 到21世纪末, 全球地表平均增温0.3-4.8 ℃ (
Assessing future nitrogen deposition and carbon cycle feedback using a multimodel approach: Analysis of nitrogen deposition.
2005
Effects of long-term temperature and nutrient manipulation on Norway spruce fine roots and mycelia production.
1
2013
... 细根(直径≤2 mm)是水分和养分吸收的主要器官, 也是根系中活跃性和敏感度最高的部分(
Fine root turnover and litter production of Norway spruce in a long-term temperature and nutrient manipulation experiment.
2
2014
... 细根(直径≤2 mm)是水分和养分吸收的主要器官, 也是根系中活跃性和敏感度最高的部分(
... 由于森林生态系统类型、林龄和系统背景值等因素不同, 各生态系统细根对氮沉降的响应并不明确(
The effects of simulated nitrogen deposition on plant root traits: A meta-analysis.
2015
A global perspective on belowground carbon dynamics under nitrogen enrichment.
1
2010
... 据国际气候变化委员会(IPCC)第五次评估报告预测: 到21世纪末, 全球地表平均增温0.3-4.8 ℃ (
Nitrogen deposition and its ecological impact in China: An overview.
2
2011
... 本研究中, 土壤增温对细根生产量有显著的促进作用, 尤其是W处理细根生产量显著高于LN和HN处理, WHN处理细根生产量显著高于HN处理.Majdj和Öhrvik (2004)对瑞典北部的挪威云杉进行增温和施肥交互试验发现, 增温样地和增温+施肥样地的细根生产量显著高于对照样地和施肥样地, 主要原因是土壤增温促进了土壤矿化速率, 提高了养分有效性, 从而使细根生产量增加(
... 本研究发现土壤增温、氮添加及两者的交互作用对杉木幼苗细根生产的影响因季节而异.在春季, 土壤增温后幼苗细根可能会较早地进入生长季; 春季亦是细根生产的高峰, 需要大量的土壤养分, 而增温后养分矿化速率增加; 同时春季的土壤含水量高, 增温后细根生产不大可能受土壤水分限制, 这些因素综合作用使增温以及增温和氮添加的交互作用对春季细根生产有显著的促进作用(
Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden.
1
2004
... 细根(直径≤2 mm)是水分和养分吸收的主要器官, 也是根系中活跃性和敏感度最高的部分(
The potential effects of nitrogen deposition on fine-root production in forest ecosystems.
2000
Effects of nitrogen and phosphorus availability on fine-root dynamics in Hawaiian montane forests.
2001
1
2005
... 中国亚热带被称为“回归带上的绿洲”, 是全球气候变化的敏感地带.杉木(Cunninghamia lanceolata)是中国南方重要的造林和用材树种, 被广泛种植, 面积达1239.1×104 hm2, 蓄积量为47357.33 × 104 m3, 分别占全国人工林面积和蓄积量的26.55%和46.89%, 在中国人工林中占有重要地位(
Direct and indi- rect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie
2
2005
... 本研究中, 土壤增温对细根生产量有显著的促进作用, 尤其是W处理细根生产量显著高于LN和HN处理, WHN处理细根生产量显著高于HN处理.Majdj和Öhrvik (2004)对瑞典北部的挪威云杉进行增温和施肥交互试验发现, 增温样地和增温+施肥样地的细根生产量显著高于对照样地和施肥样地, 主要原因是土壤增温促进了土壤矿化速率, 提高了养分有效性, 从而使细根生产量增加(
... ;
CO2 enrichment and warming of the atmosphere enhance both productivity and mortality of maple tree fine roots.
1
2004
... 据国际气候变化委员会(IPCC)第五次评估报告预测: 到21世纪末, 全球地表平均增温0.3-4.8 ℃ (
Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data.
2010
Effects of warming on root diameter, distribution, and longevity in an alpine meadow.
2
2014
... 从2014年3月至2015年2月对细根进行了为期1年的连续性观测.于每月上旬和下旬(间隔大约15天)使用微根管影像成像系统BTC-2 Minirhizotron Camera System (Bartz Technology, Santa Barbara, USA)对细根生长过程进行影像收集, 在1天内完成每次影像的收集工作.图片收集后带回实验室用Rootfly图像分析软件对其进行处理, 按照0-10 cm、10-20 cm、20-30 cm、30-40 cm进行分层处理, 并获得每张影像中细根直径、细根长度和细根数量等数据.同时鉴别活根和死根: 白色根、白色带红色斑点或红色根定义为活根; 暗黑色、皮层脱落或表皮褶皱的根定义为死根(
... 进一步分析发现, 仅在20-30 cm土层, 土壤增温对细根生产量有显著的促进作用.其可能原因是: 土壤增温电缆布设在土壤10 cm处, 土壤表层的增温幅度最大, 土壤水分蒸发较快, 土壤含水量降低最多, 因而增温对表层土壤(0-10 cm, 10-20 cm)细根生产没有促进作用; 相反, 较深土层(20-30 cm)土壤含水量较高, 细根生产受水分限制小, 增温促进细根生产.但在更深土层(30-40 cm)没有发现增温对细根生产的促进作用, 这可能与该土层土壤增温幅度较低有关(
土壤增温对不同深度土壤温度的影响
2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}