Responses of growth and litterfall production to nitrogen addition treatments from common shrublands in Mt. Dongling, Beijing, China
ZHANGJian-Hua1,2,*,, TANGZhi-Yao3, SHENHai-Hua2, FANGJing-Yun2,3 1Xinzhou Normal University, Xinzhou, Shanxi 034000, China2State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, Chinaand 3College of Urban and Environmental Sciences, and Key Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China 通讯作者:* E-mail: 1042584932@qq.com; wj123-2007@163.com* E-mail: 1042584932@qq.com; wj123-2007@163.com 责任编辑:ZHANGJian-HuaTANGZhi-YaoSHENHai-HuaFANGJing-Yun 收稿日期:2016-03-11 接受日期:2016-09-21 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略先导性科技专项(XDA05050300)和全球变化国家重大科学研究计划(2010CB950600和2014CB954004)
展开
摘要 我国北方灌丛土壤瘠薄, 近几十年来的氮沉降显著提高了北方灌丛土壤的可利用氮水平。灌木生长是灌丛碳吸存的重要组成部分, 凋落物在土壤和植物间充当着至关重要的纽带作用, 是陆地生态系统养分与能量循环的关键, 灌丛生长和凋落物生产受氮添加的影响很大。然而, 大气氮沉降对灌丛碳吸存和凋落物生产的影响人们知之甚少。该研究以荆条(Vitex negundo var. heterophylla)和绣线菊(Spiraea salicifolia)灌丛为例, 通过0 (N0)、20 (N1)、50 (N2)、100 (N3) kg N·hm-2·a-1施氮实验, 研究了短期(2012-2013年)氮添加对东灵山地区典型灌丛生长及凋落物生成的影响。研究结果显示: 在4种氮添加处理中, 荆条灌丛灌木基径年增长率分别为1.69%、2.78%、2.51%和1.80%, 相应处理中, 绣线菊灌丛灌木基径年增长率分别为1.38%、1.37%、1.59%和2.05%; 与之对应的株高年增长率分别为8.36%、8.48%、9.49%和9.83% (荆条灌丛)和2.12%、2.86、2.36%、2.52% (绣线菊灌丛)。虽然处理之间的差异没有达到显著性水平, 但N沉降在一定程度上促进了灌木的生长。不同处理间, 荆条地上生物量增加了0.19、0.23、0.14、0.15 t C·hm-2·a-1, 绣线菊灌丛地上生物量增加了0.027、0.025、0.032、0.041 t C·hm-2·a-1。在自然条件下, 荆条和绣线菊灌丛2013年凋落物的年产量分别为135.7和129.6 g·m-2。短期氮沉降对凋落物及组分的年产量有一定的促进作用, 但处理之间的差异没有达到显著性水平。研究结果表明施肥时间短、土壤含水量低等因素导致土壤可利用氮的利用效率很低, 从而使灌丛对施肥的响应比较缓慢。
关键词:氮沉降;碳循环;温带灌丛;生长率;凋落物;季节动态 Abstract Aims The shrublands of northern China have poor soil and nitrogen (N) deposition has greatly increased the local soil available N for decades. Shrub growth is one of important components of C sequestration in shrublands and litterfall acts as a vital link between plants and soil. Both are key factors in nutrient and energy cycling of terrestrial ecosystems, which greatly affected by nitrogen (N) addition (adding N fertilizer to the surface soil directly). However, the effects and significance of N addition on C sequestration and litterfall in shrublands remain unclear. Thus, a study was designed to investigate how N deposition and related treatments affected shrublands growth related to C sequestration and litterfall production of Vitex negundo var. heterophylla and Spiraea salicifolia in Mt. Dongling region of China. Methods A N enrichment experiment has been conducted for V. negundo var. heterophylla and S. salicifolia shrublands in Mt. Dongling, Beijing, including four N addition treatment levels (control (N0, 0 kg N·hm-2·a-1), low N (N1, 20 kg N·hm-2·a-1), medium N (N2, 50 kg N·hm-2·a-1) and high N (N3, 100 kg N·hm-2·a-1)). Basal diameter and plant height of shrub were measured from 2012-2013 within all treatments, and allometric models for different species of shrub’s live branch, leaf and root biomass were developed based on independent variables of basal diameter and plant height, which will be used to calculate biomass increment of shrub layer. Litterfall (litterfall sometimes is named litter, referring to the collective name for all organic matter produced by the aboveground part of plants and returned to the surface, and mainly includes leaves, bark, dead twigs, flowers and fruits.) also was investigated from 2012-2013 within all treatments. Important findings The results showed 1) mean basal diameter of shrubs in the V. negundo var. heterophylla and S. salicifolia shrublands were increased by 1.69%, 2.78%, 2.51%, 1.80% and 1.38%, 1.37%, 1.59%, 2.05% every year; 2) The height growth rate (the shrub height relative growth rate is defined with the percentage increase of plant height) of shrubs in the V. negundo var. heterophylla and S. salicifolia shrublands were 8.36%, 8.48%, 9.49%, 9.83% and 2.12%, 2.86%, 2.36%, 2.52% every year, respectively. Thee results indicated that N deposition stimulated growth of shrub layer both in V. negundo var. heterophylla and S. salicifolia shrublands, but did not reach statistical significance among all nitrogen treatments. The above-ground biomass increment of shrub layer in the V. negundo var. heterophylla and S. salicifolia shrublands were 0.19, 0.23, 0.14, 0.15 and 0.027, 0.025, 0.032, 0.041 t C·hm-2·a-1 respectively, which demonstrated that short-term N addition had no significant effects on the accumulation of C storage of the two shrublands. The litter production of the V. negundo var. heterophylla and S. salicifolia communities in 2013 were 135.7 and 129.6 g·m-2 under natural conditions, respectively. Nitrogen addition promoted annual production of total litterfall and different components of litterfall to a certain extent, but did not reach statistical significance among all nitrogen treatments. Above results indicated that short-term fertilization, together with extremely low soil moisture content and other related factors, lead to inefficient use of soil available nitrogen and slow response of shrublands to N addition treatments.
Keywords:nitrogen deposition;carbon cycle;temperate shrublands;growth rate;litterfall;seasonal dynamics -->0 PDF (655KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 张建华, 唐志尧, 沈海花, 方精云. 北京东灵山地区常见灌丛生长及凋落物生产对氮添加的响应. 植物生态学报, 2017, 41(1): 71-80 https://doi.org/10.17521/cjpe.2016.0093 ZHANGJian-Hua, TANGZhi-Yao, SHENHai-Hua, FANGJing-Yun. Responses of growth and litterfall production to nitrogen addition treatments from common shrublands in Mt. Dongling, Beijing, China. Chinese Journal of Plant Ecology, 2017, 41(1): 71-80 https://doi.org/10.17521/cjpe.2016.0093 过去几十年来, 人类活动所引起的大气氮(N)沉降增加显著改变了全球N循环(Vitousek et al., 1997; Mo et al., 2004), 这种情况在未来数十年将进一步加剧(Galloway et al., 2004)。持续增加的N沉降导致了一系列生态问题, 如土壤酸化、土壤营养元素储存失衡和水体富营养化等(Howard & Richard, 2003; Galloway et al., 2004), N沉降及其生态后果已成为全球变化研究的热点内容。 灌丛是陆地生态系统的重要组成部分, 在群落演替过程中扮演着极其重要的角色, 在环境保护和替代能源方面都有非常重要的作用(胡会峰等, 2006)。灌丛生物量较低, 因而很少受到重视, 一般而言, 灌丛生态系统碳密度只占森林的10%-30% (Lieth & Whittaker, 1975; 贺金生等, 1997; 郑绍伟和唐敏, 2007)。同时, 灌丛处于比较贫瘠的土壤, 通常为N限制生态系统, 生态系统脆弱, 更易受环境变化的影响(Gorissen et al., 2004; Wessel et al., 2004)。在全球气候变化背景下, 尤其是大气N沉降显著增加的情况下, 灌丛碳循环一旦发生变化, 就会对全球气候变化产生深刻的影响。目前, 大气N沉降对灌丛生态系统影响的研究还十分缺乏, 灌丛生态系统对N沉降的响应方式还不清楚。 为此, 我们选择北京东灵山的典型灌丛——荆条(Vitex negundo var. heterophylla)和绣线菊(Spiraea salicifolia)灌丛为研究对象, 通过施氮的方式模拟N沉降增加情景, 探讨该灌丛类型生长及凋落物生产对N沉降的初期响应规律, 以期为揭示灌丛生态系统对全球变化响应机制研究积累数据。
1 材料和方法
1.1 研究地点
东灵山(39.80°-40.03° N, 115.40°-115.60° E)位于北京市门头沟区西部, 地理上属太行山系, 小五台山东部余脉, 主峰海拔2303 m。该区属暖温带半湿润性季风气候, 年平均气温6.3 ℃; 年降水量612 mm。植被类型以暖温带落叶阔叶林为主。从低山丘陵到山顶, 该山的植被垂直带依次划分为: 在400-1100 m的低山丘陵地区, 主要以荆条、绣线菊等灌丛为主; 在海拔1100-1700 m, 是由油松(Pinus tabuliformis)林、华北落叶松(Larix principis- rupprechtii)林、辽东栎(Quercus mongolica)林、核桃楸(Juglans mandshurica)林和落叶阔叶混交林组成的典型森林植被; 在海拔1700-2300 m, 主要是由蒿草(Kobresia spp.)、细叶薹草(Carex duriuscula subsp. stenophylloides)和多种亚高山成分的杂类草组成的亚高山草甸(刘贤赵等, 2009)。研究区背景N沉降为15 kg N·hm-2·a-1 (Du et al., 2013), 基本未受污染。实验样地分别选取低海拔的荆条和绣线菊灌丛, 两灌丛的样地情况见表1。 Table 1 表1 表1实验样地地形、土壤和植被特征(平均值±标准误差, n = 3) Table 1Topography, soil and vegetation characteristics of the experimental sites (mean ± SE, n = 3)
回归分析表明, 8种灌木的生物量模型均为幂函数方程(表2)。各灌木物种基于自变量D2H的生物量模型有着较大的R2, 因变量根、茎、叶、总生物量的生物量模型的R2值的分布范围是0.41 (胡枝子)至0.95 (小叶鼠李)、0.50 (小叶白蜡)至0.96 (小叶鼠李)、0.48 (河蒴荛花)至0.88 (小叶鼠李)、0.72 (小叶白蜡)至0.96 (小叶鼠李)(所有相关系数均极为显著, p < 0.001)(表2), 绝大部分模型可以解释因变量超过80%的方差, 表明8个灌木种的各组分的生物量回归模型能够满足估算的要求。 Table 2 表2 表2东灵山地区8种常见灌木物种生物量异速生长模型 Table 2Biomass allometric models for eight common shrub species in Mt. Dongling
物种 Species
器官 Organ
变量 Variable
R2
方程 Equation
山杏
根 Root
D2H
0.81
y = 1.26x0.77
Armeniaca sibirica
枝 Branch
D2H
0.94
y = 1.09x0.79
叶 Leaf
D2H
0.87
y = 1.61x0.56
总计 Total
D2H
0.95
y = 3.53x0.74
小叶白蜡
根 Root
D2H
0.88
y = 0.2x0.91
Fraxinus bungeana
枝 Branch
D2H
0.50
y = 5.47x0.51
叶 Leaf
D2H
0.77
y = 0.13x0.8
总计 Total
D2H
0.72
y = 4.47x0.62
小叶鼠李
根 Root
D2H
0.95
y = 0.7x0.85
Rhamnus
枝 Branch
D2H
0.96
y = 0.74x0.91
parvifolia
叶 Leaf
D2H
0.88
y = 0.89x0.58
总计 Total
D2H
0.96
y = 1.95x0.85
河朔荛花
根 Root
D2H
0.83
y = 0.81x0.61
Wikstroemia
枝 Branch
D2H
0.95
y = 0.92x0.78
chamedaphne
叶 Leaf
D2H
0.48
y = 1.32x0.35
总计 Total
D2H
0.92
y = 2.31x0.68
绣线菊
根 Root
D2H
0.77
y = 0.18x1.14
Spiraea salicifolia
枝 Branch
D2H
0.78
y = 0.23x1.18
叶 Leaf
D2H
0.62
y = 0.07x1.15
总计 Total
D2H
0.79
y = 0.48x1.16
荆条
根 Root
D2H
0.84
y = 0.2x1.09
Vitex negundo var.
枝 Branch
D2H
0.82
y = 0.54x0.95
heterophylla
叶 Leaf
D2H
0.66
y = 0.86x0.59
总计 Total
D2H
0.90
y = 1.07x0.95
蚂蚱腿子
根 Root
D2H
0.95
y = 0.07x1.16
Myripnois dioica
枝 Branch
D2H
0.91
y = 0.42x1.00
叶 Leaf
D2H
0.79
y = 0.31x0.71
总计 Total
D2H
0.95
y = 0.64x1.00
胡枝子
根 Root
D2H
0.41
y = 0.19x1.06
Lespedeza bicolor
枝 Branch
D2H
0.87
y = 0.15x1.5
叶 Leaf
D2H
0.84
y = 0.29x1.25
总计 Total
D2H
0.84
y = 0.61x1.29
D2H is regarded as independent variables and organ biomass as dependent variables.D2H为自变量, 器官生物量为因变量。 新窗口打开
2.2 氮添加对灌木生长的影响
实验前, 各样地间的灌木的D和H无显著差异(表3), 可以用来比较不同N处理对灌木生长的影响。施肥2年后, N0、N1、N2和N3处理的荆条灌丛灌木层D增长率分别为: 1.69%、2.78%、2.51%、1.80%, 相应处理中绣线菊灌丛灌木层D增长率为1.38%、1.37%、1.59%、2.05% (图1A); 不同处理中, 荆条灌丛H增长率分别为8.36%、8.48%、9.49%、9.83%, 相应处理中绣线菊灌丛H增长率分别为2.12%、2.86%、2.36%、2.52% (图1B)。氮添加对荆条和绣线菊样地灌木的D和H增长率均无显著的影响。 Table 3 表3 表3不同氮添加处理样地灌木的基径和株高(平均值±标准偏差) Table 3Shrub diameter (D, cm) and height (H, cm) of shrubland under different nitrogen addition treatments (mean ± SD)
物种 Species
变量 Variable
对照 Control (N0)
低氮 Low-N (N1)
中氮 Medium-N (N2)
高氮 High-N (N3)
荆条灌丛 Vitex negundo var. heterophylla shrubland
D
0.70 ± 0.10
0.90 ± 0.20
0.70 ± 0.00
0.70 ± 0.10
H
78.30 ± 7.80
84.20 ± 10.10
74.60 ± 3.00
75.40 ± 9.00
绣线菊灌丛 Spiraea salicifolia shrubland
D
0.53 ± 0.00
0.57 ± 0.00
0.57 ± 0.00
0.54 ± 0.00
H
76.40 ± 0.60
78.80 ± 4.00
81.40 ± 7.10
82.80 ± 4.70
N0, 0 kg N·hm-2·a-1; N1, 20 kg N·hm-2·a-1; N2, 50 kg N·hm-2·a-1; N3, 100 kg N·hm-2·a-1. 新窗口打开 显示原图|下载原图ZIP|生成PPT 图1氮添加对灌木基径(D)和株高(H)相对生长速率的影响(平均值±标准偏差)。相同字母a和b分别表示各处理间无显著差异(p > 0.05)。N0、N1、N2、N3氮添加量分别为0、20、50、100 kg N·hm-2·a-1。 -->Fig. 1Influence of nitrogen addition treatments on the relative growth rates of shrub diameter (D) and height (H) (mean ± SD). The same letters indicates no significant difference (p > 0.05) among treatments. N0, N1, N2 and N3 denote 0, 20, 50 and 100 kg N·hm-2·a-1 nitrogen addition, respectively. -->
2.3 氮添加对灌木层生物量的影响
施肥2年后, 各处理中荆条灌丛灌木地上生物量(AGB)增量分别为0.19、0.23、0.14、0.15 t C·hm-2·a-1, 相应处理中绣线菊灌丛灌木地上生物量增量分别为0.027、0.025、0.032、0.041 t C·hm-2·a-1 (图2A); 荆条和绣线菊灌丛各处理样地总生物量(TB)的增量分别为0.29、0.36、0.22、0.22 t C·hm-2·a-1和0.043、0.046、0.048、0.047 t C·hm-2·a-1 (图2B)。N添加对荆条和绣线菊灌丛灌木层的生物量无明显的影响。 显示原图|下载原图ZIP|生成PPT 图2氮添加对灌木层地上生物量(AGB)和总生物量(TB)的净增量的影响(平均值±标准偏差)。相同字母a和b分别表示各处理间无显著差异(p > 0.05)。N0、N1、N2、N3同图1。 -->Fig. 2Influence of nitrogen addition treatments on above ground biomass (AGB) and total biomass (TB) (mean ± SD). The same letter indicates no significant difference (p > 0.05) among treatments. N0, N1, N2, N3 see Fig. 1. -->
2.4 凋落物产量对N添加的初期响应
在2012年, N2和N3处理增加了荆条灌丛凋落量; 各N处理均提高了绣线菊灌丛凋落量。在2013年, 各N处理对两灌丛凋落量均有促进作用, 但均未达到统计学上的显著差异(图1)。N0、N1、N2和N3处理的荆条和绣线菊灌丛总凋落量在2012年分别为89.1、87.3、119.2、99.7和104.9、133.7、117.1、138.7 g·m-2; 2013年的产量分别为135.7、148.4、155.9、153.4和129.6、191.5、156.4、182.2 g·m-2。不同N处理下的两灌丛凋落物年产量均表现为2013年的产量高于2012年。在凋落物的组成中, 叶占大部分, 约80%以上, 其次为花果和枝, 两者约占20% (图3)。 显示原图|下载原图ZIP|生成PPT 图3实验期间2012 (左)和2013 (右)年氮添加对荆条灌丛(V.n)和绣线菊灌丛(S.t)凋落物总量及各组分量的影响(平均值±标准偏差)。N0、N1、N2、N3同图1。 -->Fig. 3Influence of nitrogen addition treatments on gross litterfall and different components of litterfall of Vitex negundo var. heterophylla (V.n) and Spiraea salicifolia (S.t) shrublands in 2012 (left) and 2013 (right) (mean ± SD). N0, N1, N2, N3 see Fig. 1. -->
由图4可知, 在荆条样地, 落叶量的月动态变化特征与凋落物总量的变化趋势相吻合。总凋落物和落叶在5至10月表现为逐渐增加的趋势, 在10月达到峰值, 年际间存在波动。花果凋落量的峰值在2012年和2013年分别出现在8月和7月; 凋落枝的变化则相对复杂, 2012年波动规律不明显, 在2013年5月出现一个峰值(图4A-4D)。在绣线菊灌丛样地, 总凋落物和落叶季节动态曲线相似, 高峰期均出现在10月。花果凋落量和落枝量在2012年变化规律不明显。2013年两者的最大值分别在7月和5月(图4E-4H)。N0、N1、N2和N3处理的花果凋落量在7月的产量分别为: 5.61、9.06、6.26和10.07 g·m-2; 相应处理的凋落枝在同年5月的产量分别为: 4.16、6.18、2.99和2.95 g·m-2。重复测量方差分析表明, N3处理显著增加了荆条灌丛叶凋落量和枝凋落量, N2处理显著增加了荆条灌丛花果凋落量。N3处理增加了绣线菊灌丛叶凋落量(图4)。 显示原图|下载原图ZIP|生成PPT 图4氮添加对凋落物总量及各组分的季节变化的影响(平均值±标准偏差)。左侧为荆条, 右侧为绣线菊。带星号的N2和N3处理与对照N0差异达到显著水平。N0、N1、N2、N3同图1。 -->Fig. 4Influence of nitrogen addition treatments on seasonal variation of gross litterfall and its component (mean ± SD). Left for Vitex negundo var. heterophylla and right for Spiraea salicifolia shrublands, respectively. N2 and N3 marked with an asterisk have a very significant difference (p < 0.05) compared to the control. N0, N1, N2, N3 see Fig. 1. -->
DrayJR, GorhamE (1964). Litter production in forests of the world .Advance in Ecological Research, 2, 101-157. [本文引用: 1]
[2]
DuEZ, ZhouZ, LiP, HuXY, MaYC, WangW, ZhengCY, ZhuJX, HeJS, FangJY (2013). NEECF: A project of nutrient enrichment experiments in China’s forests .Journal of Plant Ecology, 6, 428-435. [本文引用: 1]
[3]
FanHB, LiuWF, QiuXQ, XuL, WangQ, ChenQF (2007). Responses of litterfall production in Chinese fir plantation to increased nitrogen deposition .Chinese Journal of Ecology, 26, 1335-1338. (in Chinese with English abstract)[樊后保, 刘文飞, 裘秀群, 徐雷, 王强, 陈秋凤 (2007). 杉木人工林凋落物量对氮沉降增加的初期响应 . 生态学杂志, 26, 1335-1338.] [本文引用: 2]
[4]
FangJY, LiuGH, ZhuB, WangXK, LiuSH (2006). Carbon budgets of three temperate forest ecosystems in Dongling Mt., Beijing, China .Science in China (Series D: Earth Sciences), 36, 533-543. (in Chinese with English abstract)[方精云, 刘国华, 朱彪, 王效科, 刘绍辉 (2006). 北京东灵山三种温带森林生态系统的碳循环 . 中国科学D辑: 地球科学, 36, 533-543.] [本文引用: 1]
GaoXM, DuXJ, WangZL (2003). Comparison of seedling recruitment and establishment of Quercus wutaishanica in two habitats in Dongling Mountainous area, Beijing . Acta Phytoecologica Sinica, 27, 404-411. (in Chinese with English abstract)[高贤明, 杜晓军, 王中磊 (2003). 北京东灵山区两种生境条件下辽东栎幼苗补充与建立的比较 . 植物生态学报, 27, 404-411.] [本文引用: 1]
[7]
GorissenA, TietemaA, JoostenNN, EstiarteM, PeñuelasJ, SowerbyA, EmmettBA, BeierC (2004). Climate change affects carbon allocation to the soil in shrublands .Ecosystems, 7, 650-661. [本文引用: 2]
[8]
GuanLL, ZhouGY, ZhangDQ, LiuJX, ZhangQM (2004). Twenty years of litterfall dynamics in subtropical evergreen broad-leaved forests at the Dinghushan forest ecosystem research station . Acta Phytoecologica Sinica, 28, 445-456. (in Chinese with English abstract)[官丽莉, 周国逸, 张德强, 刘菊秀, 张倩媚 (2004). 鼎湖山南亚热带常绿阔叶林凋落物量20年动态研究 . 植物生态学报, 28, 445-456.] [本文引用: 1]
[9]
HeJS, WangQB, HuD (1997). Studies on the biomass of typical shrubland and their regeneration capacity after cutting . Acta Phytoecologica Sinica, 21, 512-520. (in Chinese with English abstract)[贺金生, 王其兵, 胡东 (1997). 长江三峡地区典型灌丛的生物量及其再生能力 . 植物生态学报, 21, 512-520.] [本文引用: 1]
[10]
HowardD, RichardB (2003). Nitrogen: The essential public enemy .Journal of Applied Ecology, 40, 771-781. [本文引用: 1]
[11]
HuHF, WangZH, LiuGH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Ecology (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 1]
[12]
KikuzawaK, AsaiT, FukuchiM (1977). Leaf litter production in a plantation of Alnus inokumae . Journal of Ecology, 72, 993-999. [本文引用: 1]
[13]
KiraT, ShideiT (1967). Primary production and turnover of organic matter in different forest ecosystems of the western Pacific .Japanese Journal of Ecology, 17, 70-87. [本文引用: 1]
[14]
KozovitsAR, BustamanteMMC, GarofaloCR, BucciS, GoldsteinG (2007). Nutrient resorption and patterns of litter production and decomposition in a Neotropical Savanna . Functional Ecology, 21, 1034-1043. [本文引用: 1]
[15]
LiethH, WhittakerRH (1975). Primary Productivity of the Biosphere . Springer-Verlag, New York. [本文引用: 1]
[16]
LiuL, GreaverTL (2010). A global perspective on below- ground carbon dynamics under nitrogen enrichment . Ecology Letters, 13, 819-828. [本文引用: 1]
[17]
LiuXZ, WangGA, LiJZ, WangQ (2009). Nitrogen isotope composition characteristics of modern plants and their variations along an altitudinal gradient in Dongling Mountain in Beijing .Science in China (Series D: Earth Sciences), 39, 1347-1359. (in Chinese with English abstract)[刘贤赵, 王国安, 李嘉竹, 王庆 (2009). 北京东灵山地区现代植物氮同位素组成及其对海拔梯度的响应 . 中国科学D辑: 地球科学, 39, 1347-1359.] [本文引用: 1]
[18]
LuM, ZhouXH, LuoYQ, YangYH, FangCM, ChenJK, LiB (2011). Minor stimulation of soil carbon storage by nitro- gen addition: A meta-analysis .Agriculture Ecosystems and Environment, 140, 234-244. [本文引用: 1]
[19]
LüY, ZhengZM, MeilibanM, LuJ, WangXH (2013). Effects of nitrogen and phosphorus fertilization on litterfall production and nutrient dynamics in a Schima superba forest in Zhejiang Province of East China . Chinese Journal of Applied Ecology, 24, 3027-3034. (in Chinese with English abstract)[吕妍, 郑泽梅, 美丽班·马木提, 陆江, 王希华 (2013). 增施氮磷肥对木荷林凋落物生产量及其养分的影响 . 应用生态学报, 24, 3027-3034.] [本文引用: 3]
[20]
MoJM, XueH, FangYT (2004). Litter decomposition and its responses to simulated N deposition for the major plants of Dinghushan forest in subtropical China .Acta Ecologica Sinica, 24, 1413-1420. [本文引用: 1]
[21]
MurtyD, McMurtrieRE, RyanMG (1996). Declining forest productivity in aging forest stands: A modeling analysis of alternative hypotheses . Tree Physiology, 16, 187-200. [本文引用: 1]
[22]
SmaillSJ, ClintonPW, GreenfieldLG (2008). Nitrogen fertilizer effects on litter fall, FH layer and mineral soil characteristics in New Zealand Pinus radiate plantations . Forest Ecology and Management, 256, 564-569.
[23]
SpieckerH (1999). Overview of recent growth trends in Euro- pean forests .Water Air and Soil Pollution, 116, 33-46. [本文引用: 1]
[24]
TammCO (1991). Nitrogen in Terrestrial Ecosystems: Questions of Productivity, Vegetational Changes, and Ecosystem Stability . Springer-Verlag, Berlin. 50-97. [本文引用: 2]
[25]
VitousekPM, AberJD, HowarthRW, LikensGE, MatsonPA, SchindlerDW, SchlesingerWH, TilmanDG (1997). Human alteration of the global nitrogen cycle: Sources and consequences .Ecological Applications, 7, 737-750. [本文引用: 1]
[26]
WangFY (1989). Review on the study of forest litterfall .Advances in Ecology, 6, 82-89. (in Chinese with English abstract)[王凤友 (1989). 森林凋落量研究综述 . 生态学进展, 6, 82-89.] [本文引用: 1]
[27]
WesselWW, TietemaA, BeierC, EmmettBA, PeñuelasJ, Riis-NielsonT (2004). A qualitative ecosystem assessment for different shrublands in western Europe under impact of climate change .Ecosystems, 7, 662-671. [本文引用: 2]
[28]
XuXN, HirataE, ShibataH (2004). Effect of typhoon disturbance on fine litterfall and related nutrient input in a sub-tropical forest on Okinawa Island, Japan .Basic and Applied Ecology, 5, 271-282. [本文引用: 1]
[29]
ZhangC, ZhangL, LiP, ShiWT, XuXN (2014). Response of litter production and its seasonality to increased nitrogen deposition in a subtropical evergreen broad-leaved forest . Chinese Journal of Ecology, 33, 1205-1210. (in Chinese with English abstract)[张驰, 张林, 李鹏, 施文涛, 徐小牛 (2014). 亚热带常绿阔叶林凋落物生产及季节动态对模拟氮沉降增加的响应 . 生态学杂志, 33, 1205-1210.]
[30]
ZhengSW, TangM (2007). Summary of research on shrub biomass in China .Journal of Chengdu University (Natural Science Edition), 26, 189-192. (in Chinese with English abstract)[郑绍伟, 唐敏 (2007). 灌木群落及生物量研究综述 . 成都大学学报(自然科学版), 26, 189-192.] [本文引用: 1]
... 过去几十年来, 人类活动所引起的大气氮(N)沉降增加显著改变了全球N循环(Vitousek et al., 1997; Mo et al., 2004), 这种情况在未来数十年将进一步加剧(Galloway et al., 2004).持续增加的N沉降导致了一系列生态问题, 如土壤酸化、土壤营养元素储存失衡和水体富营养化等(Howard & Richard, 2003; Galloway et al., 2004), N沉降及其生态后果已成为全球变化研究的热点内容. ...
... ; Galloway et al., 2004), N沉降及其生态后果已成为全球变化研究的热点内容. ...
... 过去几十年来, 人类活动所引起的大气氮(N)沉降增加显著改变了全球N循环(Vitousek et al., 1997; Mo et al., 2004), 这种情况在未来数十年将进一步加剧(Galloway et al., 2004).持续增加的N沉降导致了一系列生态问题, 如土壤酸化、土壤营养元素储存失衡和水体富营养化等(Howard & Richard, 2003; Galloway et al., 2004), N沉降及其生态后果已成为全球变化研究的热点内容. ...
Litter decomposition and its responses to simulated N deposition for the major plants of Dinghushan forest in subtropical China 1 2004
... 过去几十年来, 人类活动所引起的大气氮(N)沉降增加显著改变了全球N循环(Vitousek et al., 1997; Mo et al., 2004), 这种情况在未来数十年将进一步加剧(Galloway et al., 2004).持续增加的N沉降导致了一系列生态问题, 如土壤酸化、土壤营养元素储存失衡和水体富营养化等(Howard & Richard, 2003; Galloway et al., 2004), N沉降及其生态后果已成为全球变化研究的热点内容. ...
Declining forest productivity in aging forest stands: A modeling analysis of alternative hypotheses 1 1996
Human alteration of the global nitrogen cycle: Sources and consequences 1 1997
... 过去几十年来, 人类活动所引起的大气氮(N)沉降增加显著改变了全球N循环(Vitousek et al., 1997; Mo et al., 2004), 这种情况在未来数十年将进一步加剧(Galloway et al., 2004).持续增加的N沉降导致了一系列生态问题, 如土壤酸化、土壤营养元素储存失衡和水体富营养化等(Howard & Richard, 2003; Galloway et al., 2004), N沉降及其生态后果已成为全球变化研究的热点内容. ...
 , 唐志尧
, 唐志尧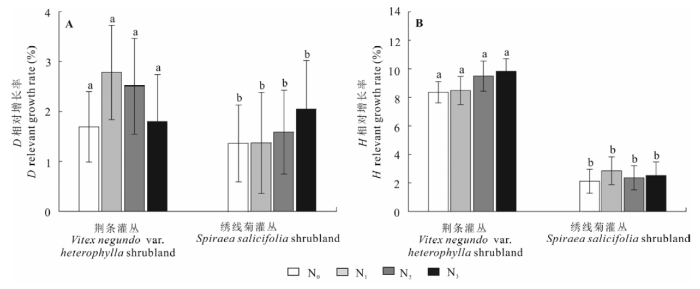 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT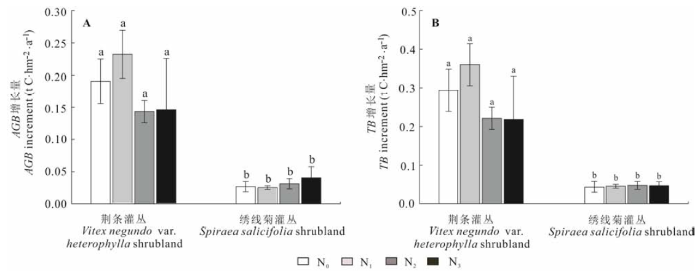 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}