韩玲, 赵成章
* , 徐婷, 冯威, 段贝贝
西北师范大学地理与环境科学学院, 甘肃省湿地资源保护与产业发展工程研究中心, 兰州 730070
Relationships between leaf thickness and vein traits of Achnatherum splendens under different soil moisture conditions in a flood plain wetland, Heihe River, China HANLing, ZHAOCheng-Zhang
* , XUTing, FENGWei, DUANBei-Bei
College of Geography and Environmental Science, Northwest Normal University, Research Center of Wetland Resources Protection and Industrial Development Engineering of Gansu Province, Lanzhou 730070, China 版权声明: 2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
展开
摘要
叶片厚度和叶脉性状的关联性影响着叶片水分的动态平衡, 对揭示植物叶片水分运输与生长速率耦合的生理生态学机制具有重要的意义。该文选择位于张掖市黑河干流边缘的洪泛平原湿地作为实验地, 以河边为起点, 沿垂直河岸线的方向依次设置I (50.07%)、II (38.77%)、III (31.5%)、IV (20.4%) 4个土壤含水量梯度样地, 采用标准化主轴估计(SMA)方法, 以叶脉密度和叶脉直径分别表示叶脉性状, 研究了不同土壤水分条件下芨芨草(Achnatherum splendens )种群的叶片厚度与叶脉性状的生长关系。结果表明: 随着湿地群落土壤含水量的逐渐降低, 湿地群落的植被密度和高度逐渐降低, 土壤电导率逐渐增大; 芨芨草的叶脉密度、叶片厚度、水分利用效率和光合有效辐射呈逐渐增加的趋势, 叶脉直径、蒸腾速率和分枝数呈逐渐减小的趋势, 净光合速率和株高呈先增大后减小的趋势; 随着湿地群落土壤含水量的逐渐降低, 芨芨草叶片厚度与叶脉密度、叶脉直径的相关性在不同样地间存在差异; 芨芨草叶片厚度与叶脉密度呈显著的正相关关系, SMA的斜率呈逐渐减小的趋势, 且在样地I和样地IV与1.0存在显著差异; 叶片厚度与叶脉直径呈显著的负相关关系, SMA的斜率呈逐渐增大的趋势, 且在样地I和样地IV与-1.0存在显著差异。芨芨草在土壤水分较充足的湿地群落倾向于少量粗脉的薄叶片构建方式, 在受土壤水分胁迫时选择大量细脉的厚叶片的生长模式, 体现了湿地植物在资源异质性分布的生境中根据其功能需求在自身性状之间进行的资源优化配置。 关键词: 叶片厚度 ;
叶脉密度 ;
叶脉直径 ;
异速生长 ;
芨芨草 ;
洪泛平原湿地 Abstract
Aims The correlations between leaf thickness and vein traits influenced the leaf hydraulic dynamic balance, and there were important meanings to reveal ecophysiological mechanisms of plant leaves water transport and growth rate. Our objective was to study the changes in the relationship between leaf thickness and vein traits (vein diameter, vein density) of Achnatherum splendens populations by using standardized major axis estimation (SMA) method under different soil moisture conditions located in flood plain wetland of Zhangye.Methods The study site was located at flood plains wetland of Zhangye, Gansu Province, China. Selecting a starting point along the vertical direction of the river, in turn, along the soil moisture gradient, four plots were set up at intervals of 40 m, plot I (50.07%), plot II (38.77%), plot III (31.5%) and plot IV (20.4%). From each of the four sample plots, seven samples were collected, resulting in (5 m × 5 m) a total of 28 samples. Community traits (height, density) and soil physical and chemical properties were investigated. Six individual samplings of A. splendens from each plot were used to measure the leaf thickness, vein density and vein diameter in laboratory. In addition, the photosynthetically active radiation (PAR ), leaf net photosynthetic rate (P n ) and transpiration rate (T r ) of A. splendens were measured in natural environment. The 28 plots were categorized into groups I, II, III and IV, and SMA estimation method was then used to examine the allometric relationship among leaf thickness, vein density and vein diameter. Important findings With a decreased soil moisture, the plant density and height displayed a pattern of steadily declining, while the soil electrical conductivity increased, In addition, the vein density, leaf thickness, water use efficiency (WUE ), PAR and twig number of A. splendens displayed a pattern of initial decrease, whereas the vein diameter and T r increase gradually, P n and plant high displayed changing trends of increasing-decreasing. The leaf thickness was negatively associated with the vein density, vein diameter, and the relationship varied with the soil moisture conditions (p < 0.05). There was a significant positive relationship (p < 0.05) between the leaf thickness and vein density. The SMA slope of the regression equation gradually decreased and was significantly different from 1.0 (p < 0.05) on plot I and IV. In addition, along decreased soil moisture, the standardized major axis slope of regression equation in the scaling relationships between the leaf thickness and vein diameter gradually increased and was significantly different from -1.0 (p < 0.05) on plot I and IV.Keywords: leaf thickness ;
vein density ;
vein diameter ;
allometry ;
Achnatherum splendens ;
-->
0
PDF (980KB) 元数据 多维度评价 相关文章 收藏文章 本文引用格式导出
EndNote Ris Bibtex 收藏本文 -->
韩玲 ,
赵成章 ,
徐婷 ,
冯威 ,
段贝贝 . 不同土壤水分条件下洪泛平原湿地芨芨草叶片厚度与叶脉性状的关系.
植物生态学报 , 2017, 41(5): 529-538
https://doi.org/10.17521/cjpe.2016.0123 HAN Ling ,
ZHAO Cheng-Zhang ,
XU Ting ,
FENG Wei ,
DUAN Bei-Bei . Relationships between leaf thickness and vein traits of
Achnatherum splendens under different soil moisture conditions in a flood plain wetland, Heihe River, China.
Chinese Journal of Plant Ecology , 2017, 41(5): 529-538
https://doi.org/10.17521/cjpe.2016.0123 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 )。叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 )。面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 )。干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 )。土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 )。叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 )。因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制。
芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 )。近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰。鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性。
1 材料和方法 1.1 研究区域和样地概况 研究区位于甘肃省张掖市甘州区西城驿黑河干流河床边缘缓坡地带的洪泛平原湿地, 地理位置为38.56° N, 100.26° E, 海拔1β482.7 m, 属温带大陆性气候, 雨热同期, 年平均气温7.8 ℃, ≥0 ℃积温为2 734 ℃, 年降水量132.6 mm, 主要集中在6-9月, 年蒸发量1β986.5 mm, 年日照时数3β077 h, 土壤以砾石土、灰棕荒漠土和草甸土为主, 湿地内土壤类型的多样化决定了旱生、盐生、湿生等植物群落的形成, 植被分布具有明显的水平分异特征, 以湿生植物、盐生植物与陆生乔灌林木为主。主要植物有芨芨草、芦苇(
Phragmites australis )、冰草(
Agropyron cristatum )、节节草(
Equisetum ramosissimum )、苦豆子(
Sophora alopecuroides )、宽苞水柏枝(
Myricaria bracteata )、多枝柽柳(
Tamarix ramosissima )、旱柳(
Salix matsudana )和沙枣(
Elaeagnus angustifolia )等。该研究区属于张掖黑河国家湿地保护区核心区, 不存在家畜放牧和人类活动等干扰因素, 能够真实地反映土壤水分等自然属性对植物生物学特性和繁殖等功能的影响(
韩玲等, 2016 )。
显示原图 |
下载原图ZIP |
生成PPT 图1研究区概况和样地设置图。 -->
Fig. 1Study area and locations of measured plots. -->
1.2 实验设置 黑河干流从莺落峡进入河西走廊, 河流两岸形成广阔的倾斜平原。 受季节性洪水和地下潜水的影响, 在河岸边缘发育了大片由芨芨草、芦苇、苦豆子等草本植物和多枝柽柳、沙枣、旱柳等乔木为主要植被的洪泛平原湿地。受地下水埋深的影响, 植被呈现明显的带状分布特征。在实地考察观测的基础上, 选择一处从河岸边到湿地与荒漠交界处长约200 m且以芨芨草为优势种的湿地群落进行实验。首先, 进行地下水埋深的测量(在每条样带上从近河岸开始每间隔10 m用铁铲挖至地下水流出, 待水位稳定后测量地下水埋深), 然后根据地下水埋深变化规律, 从黑河沿岸以垂直于河岸线的方向间隔40 m依次设置4条长20 m、宽150 m的平行样带I、II、III和IV, 分别为: 样地I, 地下水埋深0.5-1.0 m, 该样地受季节性和间歇性洪水的影响, 地表被水面半永久性覆盖, 其中11-4月份覆盖, 5-10月份不覆盖, 地下水位处在地表或附近, 主要植物有芨芨草、芦苇、宽苞水柏枝和多枝柽柳等; 样地II, 地下水埋深1.0-1.5 m, 该地段一般只是间歇性淹水, 4-11月份地表偶尔被覆盖, 地表水在一年中出现时间较短, 主要植物有芨芨草、苦豆子、旱柳等; 样地III, 地下水埋深1.5-2.0 m, 该地段一般不会淹水, 植被主要依靠地下潜水生存, 主要植物有芨芨草、节节草、沙枣等; 样地IV, 地下水埋深2.0-2.5 m, 土壤受河流水位影响极小, 土壤水分不能满足大多数植物的生长, 主要植物有芨芨草、冰草等。
2015年8月初, 于基本无明显降水、土壤水盐状况相对稳定的期间进行取样。在选定的4个样地中各随机选取7个5 m × 5 m的样方, 共计28个样方。首先分别测定每个样方内湿地群落所有植物的高度和密度, 芨芨草以丛为单位; 然后在每个小样方随机挖掘 1 m × 1 m × 0.5 m的土壤剖面, 用环刀(200 cm
3 )分5层间隔10 cm采取土样, 重复3次, 现场编号、各土层均匀混合称鲜质量, 然后带回实验室在105 ℃的烘箱内烘干12 h, 取出称质量, 计算出各样地0-50 cm土层土壤质量含水量(
韩玲等, 2016 ); 最后在每一个样方选择6丛芨芨草(株高为大中小各2丛), 用卷尺测量从基部到最高分枝的高度, 定为株高, 并记录芨芨草株丛分枝数, 然后选取每个株丛最外层4个方位充分伸展且健康完整的2-3个叶片, 做好标记后进行以下步骤:
(1)植物光合参数测定。于8月15-18日(代表植物生长旺盛期), 选择晴朗天气的9:00-12:00进行气体交换参数的测定。光合测定使用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany), 测量过程中使用人工红蓝光源, 光合有效辐射(
PAR )为1β200 μmol·m
-2 ·s
-1 , CO
2 浓度约为340 μmol·mol
-1 , 相对湿度(
RH )保持在40%-50%, 流速设定为 750 μmol·s
-1 , 叶室面积为8 cm
2 , 对做好标记的每个叶片记录5组数据用于统计分析, 分别测定叶片的净光合速率(
P n )、蒸腾速率(
T r )等参数(
任青吉等, 2015 ), 对于不能充满叶室的叶片, 则保存于湿润的封口袋中, 带回实验室, 用便携式激光叶面积仪(CI-202, Walz, Camas, USA)测定叶面积, 进而计算
P n 和
T r , 水分利用效率(
WUE ) =
P n /
T r (
韩玲等, 2016 )。
(2)光合有效辐射测定。于8月15-18日用手持光量子计(3415F, Walz, Plainfield, USA)在9:00-11:00对4个土壤水分梯度内所有供试芨芨草株丛进行
PAR 测定, 分别选择植株上方、中层(株高1/2处)和地表处向外距灌层15 cm处测量(
韩玲等, 2016 )。
(3)叶片厚度和叶脉性状的测定。基于每个芨芨草被标记的2-3个叶片, 采集后保存于湿润的封口袋中, 带回实验室用福尔马林-乙酸溶液(37%甲醛溶液, 50%乙醇和13%冰醋酸溶液)固定(
徐婷等, 2016 )。①先进行叶片厚度(单位: mm)的测定, 使用游标卡尺测量(精确度0.01 mm), 测量时尽量选择叶片中心部位, 并避开叶脉, 每个叶片重复测量5次, 最后取平均值为叶片厚度; ②叶脉性状的测定, 先用5% NaOH的乙醇进行化学清理、番红-固绿染色(
Berlyn & Miksche, 1976 )、用水包埋呈透明薄膜状, 然后在体视显微镜(SMZ168-BL, Motic, 香港, 中国)下放大10倍后拍照, 每个叶片拍摄10个视野用于叶脉性状的观测, 最后用Motic Images Plus 2.0软件获得各个照片中的叶脉直径和叶脉的总长度, 所有照片中得出的平均值即为叶脉直径和叶脉密度; ③最后用便携式激光叶面积仪(CI-202, Walz, Camas, USA)对每个叶片进行扫描, 得到叶面积, 叶脉密度用单位叶面积(单位: mm
2 )的叶脉总长度(单位: mm)表示(
Sack et al ., 2012 )。
(4)土壤盐分测定。用电导法测定土壤含盐量, 在室温下称取过2 mm筛的风干土样10 g, 加50 mL去CO
2 蒸馏水(水土质量比为5:1), 取浸出液, 置振荡机上振荡5 min。将布氏漏斗与抽气系统相连后把悬浊的土浆缓缓倒入漏斗, 直至抽滤完毕, 滤液倒入三角瓶备用(
张雅琼等, 2010 )。用上海雷磁仪器厂生产的DDS-11C便携式电导仪测浸出液的电导率, 3次重复, 取平均值即为土壤电导率值(
韩玲等, 2016 )。
1.3 数据分析 对湿地群落中4个水分梯度中芨芨草叶片厚度、叶脉密度和叶脉直径的实验数据先进行以10为底的对数转换, 使之符合正态分布后再进行分析。用于各土壤水分梯度中任意两个叶性状的异速生长方程的参数估计, 主要采用标准化主轴估计(SMA)方法(
Warton et al ., 2006 ), 采用
y = axb , 线性转换成log(
y ) =
b log(
x ) + log(
a ), 式中
x 和
y 表示两个特征参数,
a 为性状关系的截距,
b 为斜率, 即异速生长参数或相对生长的指数, 当|
b | = 1时, 表示两者是等速变化关系; 当
|b| 显著偏离1时, 两者间为异速变化关系(
Harvey & Pagel, 1991 ), 由软件SMATR Version 2.0 (
Falster et al ., 2012 )计算完成, 每一个回归斜率的置信区间根据Pitman (1939)方法计算, 并采用Warton和Weber (2002)的方法对每个密度的斜率进行异质性测试。不同水分梯度间植物功能性状平均值的差异比较采用单因素方差分析(
a = 0.05)。实验所用数据采用SPSS 16.0软件进行处理分析, 用Excel软件绘图。
2 结果分析 2.1 不同土壤水分条件下湿地群落生物学特征和土壤理化性质 不同样地湿地群落的生物学特征、土壤理化性质和
PAR 的变化如
表1 所示。不同样地间的湿地群落的土壤含水量、土壤电导率、群落密度、高度和
PAR 存在显著差异(
p < 0.05), 从样地I到样地II、样地III和样地IV, 土壤含水量呈逐渐减小趋势, 样地IV比样地I减少了59.26%; 随着不同样地土壤含水量的逐渐降低, 湿地群落的平均高度和密度均呈现逐渐减小的趋势, 样地IV比样地I群落高度减少42.42%, 密度降低67.69%;
PAR 和土壤电导率随着土壤含水量的降低均呈逐渐增加的趋势, 样地IV比样地I
PAR 增加了1.17倍, 土壤电导率增加了2.68倍。
Table 1 表1 表1不同样地的湿地群落生物学特征和土壤理化性质(平均值±标准误差, n = 30) Table 1Wetland community biology characteristics and soil physicochemical properties in different plots (mean ± SE, n = 30) 样地 土壤含水量 土壤电导率 -1 ) 高度 密度-2 ) PAR (μmol·m-2 ·s-1 )I 50.07 ± 1.24a 3.01 ± 0.19d 171.20 ± 5.24a 19.5 ± 0.55a 645.30 ± 13.18d II 38.77 ± 1.03b 5.88 ± 0.23c 146.37 ± 4.62b 16.4 ± 0.34b 833.10 ± 22.48c III 31.50 ± 1.01c 7.79 ± 0.33b 127.33 ± 3.25c 10.3 ± 0.24c 921.40 ± 31.24b IV 20.40 ± 0.67d 11.90 ± 0.55a 98.57 ± 3.66d 6.3 ± 0.1d 1β397.10 ± 45.25a
PAR, photosynthetically active radiation. Different lowercase letters in the same column indicate significant differences among plots (p < 0.05).PAR, 光合有效辐射。同列不同小写字母表示样地间差异显著(p < 0.05)。新窗口打开 2.2 不同土壤水分条件下湿地植物芨芨草各功能性状变化 芨芨草各功能性状在不同样地的变化如
表2 所示。随着土壤含水量的逐渐降低, 芨芨草的株高和净光合速率(
P n )呈先增大后减小的趋势, 株高的变化趋势为样地III > II > I > IV,
P n 的变化趋势为样地III > II > IV > I, 二者均在样地III出现较大值, 样地IV较样地I,
P n 增加了5.07%, 株高减小了26.6%; 芨芨草分枝数、叶脉直径和蒸腾速率(
T r )均呈逐渐减小的趋势, 样地IV较样地I分别减少了21.15%、44.26%和7.03%; 叶片厚度、叶脉密度和水分利用效率(
WUE )均呈逐渐增加的趋势, 从样地I到样地IV分别增加了14.48%、23.69%和12.95%; 芨芨草各功能性状的变化在样地II和样地III之间均无显著差异(
p > 0.05)。表明样地II和样地III的湿地群落生境胁迫较小, 较适宜湿地群落芨芨草种群的生存。
Table 2 表2 表2不同样地芨芨草各功能性状(平均值±标准误差, n = 30) Table 2Functional characteristics of Achnatherum splendens in different plots (mean ± SE, n = 30) 各功能性状 Functional characteristic 样地 Plot I II III IV 株高 Plant height (cm) 130.34 ± 4.50b 149.67 ± 4.62a 152.00 ± 5.24a 95.67 ± 3.56c 分枝数 Twig number (No.·clump-1 ) 203.33 ± 8.83a 173.67 ± 6.34b 155.33 ± 5.90b 113.33 ± 4.29c 叶片厚度 Leaf thickness (mm) 0.145 ± 0.01c 0.158 ± 0.02b 0.162 ± 0.02b 0.166 ± 0.02a 叶脉直径 Vein diameter (mm) 0.208 ± 0.04a 0.195 ± 0.03b 0.184 ± 0.02b 0.164 ± 0.02c 叶脉密度 Vein destiny (mm·mm-1 ) 1.283 ± 0.05c 1.406 ± 0.04b 1.439 ± 0.03b 1.587 ± 0.03a P n (μmol CO2 ·m-2 ·s-1 )13.20 ± 0.12c 13.94 ± 0.17a 14.05 ± 0.18a 13.87 ± 0.13b T r (mmol H2 O·m-2 ·s-1 )6.83 ± 0.11a 6.67 ± 0.10b 6.63 ± 0.09b 6.35 ± 0.07c WUE (μmol CO2 ·mmol-1 H2 O)1.93 ± 0.01c 2.08 ± 0.02b 2.12 ± 0.02b 2.18 ± 0.03a
P n , net photosynthetic rate; T r , transpiration rate. WUE, water use efficiency. Different lowercase letters in the same line indicate significant differences among plots (p < 0.05).P n , 净光合速率; T r , 蒸腾速率; WUE, 水分利用效率。同行不同小写字母表示样地间差异显著(p < 0.05)。新窗口打开 2.3 不同土壤水分条件下湿地植物芨芨草叶片厚度和叶脉密度的关系 如
图2 所示, 不同土壤水分条件下芨芨草叶片厚度和叶脉密度呈显著的正相关生长关系(
p < 0.05)。通过SMA分析得到, 芨芨草在不同样地间的SMA斜率存在差异, 样地I为1.196β4 (95%的置信区间:
CI = 0.992β0-1.163β1)、样地II为0.974β2 (95%的置信区间:
CI = 0.802β2-0.994β3)、样地III为0.962β6 (95%的置信区间:
CI = 0.885β4-0.972β1)、样地IV为0.808β9 (95%的置信区间:
CI = 0.764β4-0.857β2)。从样地I到样地IV SMA斜率依次呈现逐渐减小的趋势, 样地I的SMA斜率显著大于1.0 (
p < 0.05), 样地II和III的SMA斜率均与1.0无显著差异(
p > 0.05), 样地IV的SMA斜率显著小于1.0 (
p < 0.05)。表明芨芨草叶片厚度随着叶脉密度的增加而呈显著增加的趋势, 在不同样地间呈现不同程度的异速生长关系(
p < 0.05), 在样地I芨芨草叶片厚度的增加速率大于叶脉密度的增加速率; 样地II和III二者呈近等速生长关系; 样地IV叶脉密度增加的速度大于叶片厚度增加的速度, 土壤水分的异质性分布影响了芨芨草叶片厚度和叶脉密度的资源权衡机制。
显示原图 |
下载原图ZIP |
生成PPT 图2不同土壤水分条件下芨芨草叶片厚度与叶脉密度的关系。I, 样地I (土壤含水量50.07%); II, 样地II (土壤含水量38.77%); III, 样地III (土壤含水量31.5%); IV, 样地IV (土壤含水量20.4%)。 -->
Fig. 2Relationship between leaf thickness and vein density of Achnatherum splendens among different soil moisture conditions. I, plot I (soil moisture content 50.07%); II, plot II (soil moisture content 38.77%); III, plot III (soil moisture content 31.5%); IV, plot IV (soil moisture content 20.4%). -->
2.4 不同土壤水分条件下湿地植物芨芨草叶片厚度和叶脉直径的关系 如
图3 所示, 不同土壤水分条件下芨芨草叶片厚度和叶脉直径呈显著的负相关关系(
p < 0.05)。通过SMA分析得到, 芨芨草在不同样地间的标准主轴化斜率存在差异, 样地I为-0.777β3 (95%的置信区间:
CI = -0.783 0, -0.741 6)、样地II为-0.956 6 (95%的置信区间:
CI = -0.101 2, -0.943 1)、样地III为-1.035 4 (95%的置信区间:
CI = -1.074 5, -0.981 2)、样地IV为-1.116 5 (95%的置信区间:
CI = -1.174 4, -1.082β7), 从样地I到样地IV, SMA斜率的绝对值呈现逐渐增大的趋势, 样地I的SMA斜率显著大于-1.0 (
p < 0.05), 样地II和III的SMA斜率均与-1.0无显著差异(
p > 0.05), 样地IV的SMA斜率显著小于-1.0 (
p < 0.05)。表明芨芨草随着叶片厚度的逐渐增大, 叶脉直径呈相反的变化趋势, 二者在不同样地呈显著的权衡生长关系(
p < 0.05), 且在样地I芨芨草叶片厚度减小的速度小于叶脉直径减小的速度; 样地II和III叶片厚度增加的速度接近叶脉直径减小的速度; 样地IV叶片厚度增加的速度大于叶脉直径减小的速度, 不同生境中土壤含水量的差异加剧了叶片厚度与叶脉直径的资源权衡分配机制。
显示原图 |
下载原图ZIP |
生成PPT 图3不同土壤水分条件下芨芨草叶片厚度与叶脉直径的关系。I, 样地I (土壤含水量50.07%); II, 样地II (土壤含水量38.77%); III, 样地III (土壤含水量31.5%); IV, 样地IV (土壤含水量20.4%)。 -->
Fig. 3Relationship between leaf thickness and vein diameter of Achnatherum splendens among different soil moisture conditions. I, plot I (soil moisture content 50.07%); II, plot II (soil moisture content 38.77%); III, plot III (soil moisture content 31.5%); IV, plot IV (soil moisture content 20.4%). -->
3 讨论 叶片水力阻力是植物应对环境水分和光照胁迫的主要限制因子之一(
Scoffoni et al ., 2011 ), 被视作植物水分传输过程中的一个水力瓶颈, 是应对灾难性水力失调的安全阀门(
Villagra et al ., 2013 )。分析叶片厚度与叶脉密度、叶脉直径的异速生长关系对认识叶片适应不同水分环境具有重要作用。为了缓解生理活动中的水分胁迫, 植物往往在叶片水分防御和输导的构造上进行资源权衡投资(
许洺山等, 2015 ), 这影响着植物叶片的形态构建及代谢活动中的碳分配(
Moles et al ., 2014 )。本研究发现, 在不同土壤水分条件下, 芨芨草叶片厚度与叶脉密度、叶脉直径间存在不同的生长关系(
p < 0.05), 在样地I和IV, SMA斜率的绝对值均与1.0存在显著差异(
p < 0.05), 在样地II和III, SMA斜率的绝对值与1.0无显著差异(
p > 0.05)(
图2 ,
图3 )。不同土壤水分样地芨芨草叶性状的异速生长关系及敏感性差异, 表明湿地植物在维持水力安全和碳利用效率间存在着潜在的权衡机制, 对于理解和预测不同环境下植物的存活、生长、繁衍和分布具有重要的意义。
3.1 高土壤含水量条件下芨芨草叶片厚度和叶脉性状之间的生长关系 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 )。研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 )。在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 )。主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新。
3.2 低土壤含水量条件下芨芨草叶片厚度和叶脉性状的生长关系 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 )。单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 )。湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力。
叶形态和生理特性在植物碳同化、水分关系和能量平衡等方面有重要作用, 决定了叶片功能属性在不同环境条件下的差异(
Wright et al ., 2004 ;
Poorter & Bongers, 2006 )。芨芨草喜生于地下水埋深1.5 m左右的盐碱滩沙质土壤上(
张明娟等, 2012 ), 在地下水埋深为1.0-2.0 m, 且土壤水分适中的样地II和样地III, 湿地群落环境中群落高度、密度和地表土壤水分、理化过程和强度均处于干旱到湿润的过渡区, 种间对空间、光和土壤水分、养分的竞争有所减弱(
表1 ), 比较适宜芨芨草的生长, 芨芨草
P n 和株高最大(
表2 ), 使其能利用较为充足的物质和能量资源进行高效的光合作用和物质运输, 芨芨草选择了折中水分输送和存储结构的稳步投资, 形成了叶片厚度与叶脉密度、叶脉直径均近等速生长的格局(
p < 0.05、
图2 -3), 也是叶片厚度由薄到厚、叶脉密度由小到大、叶脉直径由粗到细转变的区域, 即在该环境中保持较高的光合能力和竞争力, 保证了植株的顺利生长和种群的繁衍更新, 从而成为湿地群落中的优势种。
4 结论 生长在土壤含水量丰富的近河岸湿地群落样地中的芨芨草叶片厚度增长的速度快于叶脉密度增加的速度, 从而导致产生了较薄的叶片, 叶脉粗, 数量少, 倾向于采取高蒸腾速率的开拓性策略, 而远离河岸的芨芨草叶脉密度增加的速度大于叶片厚度增加的速度, 具有较厚的叶片, 叶脉细, 数量多, 选择了以降低水分消耗为主的保守性策略, 在不同水分环境下形成了湿地植物叶片水分传输通道表型可塑性机制。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
[1] Berlyn GP Miksche JP Botanical Microtechnique and Cytochemistry . Iowa State University Press, Ames. [本文引用: 1] [2] Blonder B Violle C Bentley LP Enquist BJ 2010 ). Venation networks and the origin of the leaf economics spectrum. Ecology Letters, 91 -100 . [本文引用: 1] [3] Brodribb TJ Jordan GJ 2011 ). Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees. New Phytologist, 437 -448 . [本文引用: 1] [4] Brodribb TJ Feild TS Jordan GJ 2007 ). Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiology, 1890 -1898 . [本文引用: 1] [5] Domínguez MT Aponte C Perez-Ramos IM 2012 ). Relationships between leaf morphological traits, nutrient concentrations and isotopic signatures for Mediterranean woody plant species and communities. Plant and Soil, 407 -424 . [本文引用: 1] [6] Falster DS Warton DI Wright IJ URL [本文引用: 2] [7] Gago J Douthe C Florez-Sarasa I Escalona JM Galmes J Fernie AR Flexas J Medrano H 2014 ). Opportunities for improving leaf water use efficiency under climate change conditions. Plant Science, 108 -119 . [本文引用: 2] [8] Givnish TJ 1987 ). Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints. The New Phytologist, 131 -160 . [本文引用: 1] [9] Gong R Gao Q 2015 ). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions. Chinese Journal of Plant Ecology, 300 -308 . (in English with Chinese abstract)[龚容 , 高琼 (2015 ). 叶片结构的水力学特性对植物生理功能影响的研究进展 植物生态学报 , 39, 300 -308 .] [10] Harvey PH Pagel MD [本文引用: 7] [11] Han L Zhao CZ Xu T Feng W Duan BB Zheng HL 2016 ). Trade-off between leaf size and vein density of Achnatherum splendens in Zhangye wetland. Chinese Journal of Plant Ecology, 788 -797 . (in Chinese with English abstract)[韩玲 , 赵成章 , 徐婷 , 冯威 , 段贝贝 , 郑慧玲 (2016 ). 张掖湿地芨芨草叶大小和叶脉密度的权衡关系 植物生态学报 , 40, 788 -797 .] [本文引用: 1] [12] Hou ZJ Zhao CZ Li Y Zhang Q Ma XL 2014 ). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland. Chinese Journal of Plant Ecology, 281 -288 . (in English with Chinese abstract)[侯兆疆 , 赵成章 , 李钰 , 张茜 , 马小丽 (2014 ). 不同坡向高寒退化草地狼毒株高和枝条数的权衡关系 植物生态学报 , 38, 281 -288 .] [本文引用: 1] [13] Hultine KR Marshall JD 2000 ). Altitude trends in conifer leaf morphology and stable carbon isotope composition. Oecologia, 32 -40 . [本文引用: 1] [14] Kang SZ Zhang JH 2004 ). Controlled alternate partial root- zone irrigation: Its physiological consequences and impact on water use efficiency. Journal of Experimental Botany, 2437 -2446 . [本文引用: 4] [15] Li R Jiang ZM Zhang SX Cai J 2015 ). A review of new research progress on the vulnerability of xylem embolism of woody plants. Chinese Journal of Plant Ecology, 838 -848 . (in English with Chinese abstract)[李荣 , 姜在民 , 张硕新 , 蔡靖 (2015 ). 木本植物木质部栓塞脆弱性研究新进展 植物生态学报 , 39, 838 -848 .] [本文引用: 1] [16] Matthew-Ogburn R Edwards EJ 2013 ). Repeated origin of three-dimensional leaf venation releases constraints on the evolution of succulence in plants. Current Biology, 722 -726 . [本文引用: 1] [17] McKown AD Cochard H Sack L 2010 ). Decoding leaf hydraulics with a spatially explicit model: Principles of venation architecture and implications for its evolution. The American Naturalist, 447 -460 . [本文引用: 2] [18] Moles AT Perkins SE Laffan SW 2014 ). Which is a better predictor of plant traits: Temperature or precipitation? Journal of Vegetation Science, 1167 -1180 . [本文引用: 1] [19] Nardini A Luglio J 2014 ). Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes. Functional Ecology, 810 -818 .[20] Nardini A Pedà G La Rocca N 2012 ). Trade-offs between leaf hydraulic capacity and drought vulnerability: Morpho- anatomical bases, carbon costs and ecological consequences. New Phytologist, 788 -798 . [本文引用: 1] [21] Nardini A Raimondo F LoGullo MA 2010 ). Leaf miners help us understand leaf hydraulic design. Plant, Cell & Environment, 1091 -1100 . [本文引用: 2] [22] Ogburn RM Edwards EJ 2012 ). Quantifying succulence: A rapid, physiologically meaningful metric of plant water storage. Plant, Cell & Environment, 1533 -1542 . [本文引用: 1] [23] Pausas JG Austin MP 2001 ). Patterns of plant species richness in relation to different environments: An appraisal. Journal of Vegetation Science, 153 -166 . [本文引用: 1] [24] Perez-Ramos IM Roumet C Cruz P 2012 ). Evidence for a plant community economics spectrum driven by nutrient and water limitations in a Mediterranean rangeland of southern France. Journal of Ecology, 1315 -1327 . [本文引用: 1] [25] Poorter L Bongers F 2006 ). Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology, 1733 -1743 . [本文引用: 1] [26] Quan XK Wang CK 2015 ). Comparison of foliar water use efficiency among 17 provenances of Larix gmelinii in the Mao’ershan area. Chinese Journal of Plant Ecology, 352 -361 . (in English with Chinese abstract)[全先奎 , 王传宽 (2015 ). 帽儿山17个种源落叶松针叶的水分利用效率比较 植物生态学报 , 39, 352 -361 .] [本文引用: 1] [27] Ren QJ Li HL Bu HY 2015 ). Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow. Chinese Journal of Plant Ecology, 593 -603 . (in English with Chinese abstract)[任青吉 , 李宏林 , 卜海燕 (2015 ). 玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较 植物生态学报 , 39, 593 -603 .] [本文引用: 2] [28] Reich PB Cornelissen H 2014 ). The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. Journal of Ecology, 275 -301 . [本文引用: 1] [29] Sack L Scoffoni C McKown AD 2012 ). Developmentally based scaling of leaf venation architecture explains global ecological patterns. Nature Communications, 837 . [本文引用: 2] [30] Sack L Scoffoni C 2013 ). Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 983 -1000 . [本文引用: 3] [31] Sack L Scoffoni C John GP Poorter H Mason CM Mendez-Alonzo R Donovan LA 2013 ). How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis. Journal of Experimental Botany, 4053 -4080 . [本文引用: 3] [32] Scoffoni C Rawls M McKown A Cochard H Sack L 2011 ). Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture. Plant Physiology, 832 -843 . [本文引用: 3] [33] Scoffoni C Kunkle J Pasquet-Kok J Vuong C Patel AJ Montgomery RA Givnish TJ Sack L 2015 ). Light induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse Hawaiian lobeliads. New Phytologist, 43 -58 . [本文引用: 1] [34] Silvertown J Charlesworth D 2001 ). Introduction to Plant Population Biology. BlackwellLondon. [本文引用: 1] [35] Song LL Fan JW Wu SH Zhong HP Wang N 2012 ). Response characteristics of leaf traits of common species along an altitudinal gradient in Hongchiba Grassland, Chongqing. Acta Ecologica Sinica, 2759 -2767 . (in English with Chinese abstract)[宋璐璐 , 樊江文 , 吴绍洪 , 钟华平 , 王宁 (2012 ). 红池坝草地常见物种叶片性状沿海拔梯度的响应特征 生态学报 , 32, 2759 -2767 .] [本文引用: 1] [36] Sperry JS 2000 ). Hydraulic constraints on plant gas exchange. Agricultural and Forest Meteorology, 13 -23 . [本文引用: 1] [37] Villagra M Campanello PI Bucci SJ Goldstein G 2013 ). Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species. Tree Physiology, 1308 -1318 . [本文引用: 1] [38] Wang RL Yu GR He NP Wang QF Zhao N Xu ZW 2015 ). Latitudinal patterns and influencing factors of leaf functional traits in Chinese forest ecosystems. Acta Geographica Sinica, 1735 -1746 . (in English with Chinese abstract)[王瑞丽 , 于贵瑞 , 何念鹏 , 王秋凤 , 赵宁 , 徐志伟 (2015 ). 中国森林叶片功能属性的纬度格局及其影响因素 地理学报 , 70, 1735 -1746 .] [39] Warton DI Weber NC 2002 ). Common slope tests for bivariate errors-in-variables models. Biometrical Journal, 161 -174 . [本文引用: 1] [40] Warton DI Wright IJ Falster DS Westoby M 2006 ). Bivariate line-fitting methods for allometry. Biological Reviews, 259 -291 . [本文引用: 1] [41] Wang J Cheng JM Wan HE Fang F 2004 ). Study on soil moisture characteristics and water use efficiency of Achnatherum splendens grassland in Loess Plateau. Arid Meteorology, 51 -55 . [本文引用: 1] [42] Wilson PJ Thompson K Hodgson JG 1999 ). Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 155 -162 . [本文引用: 1] [43] Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JH Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas ML Niinemets U Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The worldwide leaf economics spectrum. Nature, 821 -827 . [本文引用: 1] [44] Wu HW Li XY Jing ZY Li J Zheng XR Zhao DZ 2015 ). Variations in water use for Achnatherum splendens in Lake Qinghai watershed based on δ D and δ 18 O. Acta Ecologica Sinica, 8174 -8183 . (in English with Chinese abstract)[吴华武 , 李小雁 , 蒋志云 , 李静 , 郑肖然 , 赵殿智 (2015 ). 基于δ δ 18 O的青海湖流域芨芨草水分利用来源变化研究 生态学报 , 35, 8174 -8183 .] [本文引用: 3] [45] Xu MS Huang HX Shi QR Yang XD Zhou LL Zhao YT Zhang QQ Yan ER 2015 ). Responses of soil water content to change in plant functional traits in evergreen broadleaved forests in eastern Zhejiang Province. Chinese Journal of Plant Ecology, 857 -866 . (in English with Chinese abstract)[许洺山 , 黄海侠 , 史青茹 , 杨晓东 , 周刘丽 , 赵延涛 , 张晴晴 , 阎恩荣 (2015 ). 浙东常绿阔叶林植物功能性状对土壤含水量变化的响应 植物生态学报 , 39, 857 -866 .] [本文引用: 2] [46] Xu T Zhao CZ Duan BB Han L Zhen HL Feng W 2016 ). Slope-related variations of different levels of vein density and leaf size in Robinia pseudoacacia in the northern mountains of Lanzhou. Chinese Journal of Ecology, 41 -47 . (in English with Chinese abstract)[徐婷 , 赵成章 , 段贝贝 , 韩玲 , 郑慧玲 , 冯威 (2016 ). 兰州北山刺槐不同等级叶脉密度与叶大小关系的坡向差异性 生态学杂志 , 35, 41 -47 .] [本文引用: 1] [47] Yu HY Chen YT Xu ZZ Zhou GS 2014 ). Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol. Chinese Journal of Plant Ecology, 1029 -1040 . (in English with Chinese abstract)[于鸿莹 , 陈莹婷 , 许振柱 , 周广胜 (2014 ). 内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析 植物生态学报 , 38, 1029 -1040 .] [本文引用: 1] [48] Zhang SB Guan ZJ Sun M Zhang JJ Cao KF Hu H 2012 ). Evolutionary association of stomatal traits with leaf vein density in Paphiopedilum, Orchidaceae. PLOS ONE, e40080 . doi: 10.1371/journal.pone.0040080 . [本文引用: 1] [49] Zhang ZL Liu GD Zhang FC Zheng CX Kang YH 2014 ). Research progress of plant leaf hydraulic conductivity. Chinese Journal of Ecology, 1663 -1670 . (in English with Chinese abstract)[张志亮 , 刘国东 , 张富仓 , 郑彩霞 , 康银红 (2014 ). 植物叶片导水率的研究进展 生态学杂志 , 33, 1663 -1670 .] [本文引用: 3] [50] Zhang MJ Liu MS Xu C Chi T Hong C 2012 ). Spatial pattern responses of Achnatherum splendens to environmental stress in different density levels. Acta Ecologica Sinica, 595 -604 . (in English with Chinese abstract)[张明娟 , 刘茂松 , 徐驰 , 池婷 , 洪超 (2012 ). 不同密度条件下芨芨草空间格局对环境胁迫的响应 生态学报 , 32, 595 -604 .] [本文引用: 1] [51] Zhang YQ Liang CZ Wang W Wang LX Peng JT Yan JC Jia JC 2010 ). Soil salinity and Achnatherum splendens distribution. Chinese Journal of Ecology, 2438 -2443 . (in English with Chinese abstract)[张雅琼 , 梁存柱 , 王炜 , 王立新 , 彭江涛 , 闫建成 , 贾成朕 (2010 ). 芨芨草群落土壤盐分特征 生态学杂志 , 29, 2438 -2443 .] [本文引用: 1] [52] Zhou HH Li WH 2015 ). Responses and adaptation of xylem hydraulic conductivity to salt stress in Populus euphratica . Chinese Journal of Plant Ecology, 81 -91 . (in English with Chinese abstract)[周洪华 , 李卫红 (2015 ). 胡杨木质部水分传导对盐胁迫的响应与适应 植物生态学报 , 39, 81 -91 .]
1
... (3)叶片厚度和叶脉性状的测定.基于每个芨芨草被标记的2-3个叶片, 采集后保存于湿润的封口袋中, 带回实验室用福尔马林-乙酸溶液(37%甲醛溶液, 50%乙醇和13%冰醋酸溶液)固定(
徐婷等, 2016 ).①先进行叶片厚度(单位: mm)的测定, 使用游标卡尺测量(精确度0.01 mm), 测量时尽量选择叶片中心部位, 并避开叶脉, 每个叶片重复测量5次, 最后取平均值为叶片厚度; ②叶脉性状的测定, 先用5% NaOH的乙醇进行化学清理、番红-固绿染色(
Berlyn & Miksche, 1976 )、用水包埋呈透明薄膜状, 然后在体视显微镜(SMZ168-BL, Motic, 香港, 中国)下放大10倍后拍照, 每个叶片拍摄10个视野用于叶脉性状的观测, 最后用Motic Images Plus 2.0软件获得各个照片中的叶脉直径和叶脉的总长度, 所有照片中得出的平均值即为叶脉直径和叶脉密度; ③最后用便携式激光叶面积仪(CI-202, Walz, Camas, USA)对每个叶片进行扫描, 得到叶面积, 叶脉密度用单位叶面积(单位: mm
2 )的叶脉总长度(单位: mm)表示(
Sack et al ., 2012 ). ...
Venation networks and the origin of the leaf economics spectrum.
1
2010
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
Water supply and demand remain balanced during leaf acclimation of
1
2011
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Leaf maximum photosynthetic rate and venation are linked by hydraulics.
1
2007
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Relationships between leaf morphological traits, nutrient concentrations and isotopic signatures for Mediterranean woody plant species and communities.
1
2012
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
2
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 对湿地群落中4个水分梯度中芨芨草叶片厚度、叶脉密度和叶脉直径的实验数据先进行以10为底的对数转换, 使之符合正态分布后再进行分析.用于各土壤水分梯度中任意两个叶性状的异速生长方程的参数估计, 主要采用标准化主轴估计(SMA)方法(
Warton et al ., 2006 ), 采用
y = axb , 线性转换成log(
y ) =
b log(
x ) + log(
a ), 式中
x 和
y 表示两个特征参数,
a 为性状关系的截距,
b 为斜率, 即异速生长参数或相对生长的指数, 当|
b | = 1时, 表示两者是等速变化关系; 当
|b| 显著偏离1时, 两者间为异速变化关系(
Harvey & Pagel, 1991 ), 由软件SMATR Version 2.0 (
Falster et al ., 2012 )计算完成, 每一个回归斜率的置信区间根据Pitman (1939)方法计算, 并采用Warton和Weber (2002)的方法对每个密度的斜率进行异质性测试.不同水分梯度间植物功能性状平均值的差异比较采用单因素方差分析(
a = 0.05).实验所用数据采用SPSS 16.0软件进行处理分析, 用Excel软件绘图. ...
Opportunities for improving leaf water use efficiency under climate change conditions.
2
2014
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints.
1
1987
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
叶片结构的水力学特性对植物生理功能影响的研究进展
2015
7
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
... 研究区位于甘肃省张掖市甘州区西城驿黑河干流河床边缘缓坡地带的洪泛平原湿地, 地理位置为38.56° N, 100.26° E, 海拔1β482.7 m, 属温带大陆性气候, 雨热同期, 年平均气温7.8 ℃, ≥0 ℃积温为2 734 ℃, 年降水量132.6 mm, 主要集中在6-9月, 年蒸发量1β986.5 mm, 年日照时数3β077 h, 土壤以砾石土、灰棕荒漠土和草甸土为主, 湿地内土壤类型的多样化决定了旱生、盐生、湿生等植物群落的形成, 植被分布具有明显的水平分异特征, 以湿生植物、盐生植物与陆生乔灌林木为主.主要植物有芨芨草、芦苇(
Phragmites australis )、冰草(
Agropyron cristatum )、节节草(
Equisetum ramosissimum )、苦豆子(
Sophora alopecuroides )、宽苞水柏枝(
Myricaria bracteata )、多枝柽柳(
Tamarix ramosissima )、旱柳(
Salix matsudana )和沙枣(
Elaeagnus angustifolia )等.该研究区属于张掖黑河国家湿地保护区核心区, 不存在家畜放牧和人类活动等干扰因素, 能够真实地反映土壤水分等自然属性对植物生物学特性和繁殖等功能的影响(
韩玲等, 2016 ). ...
... 2015年8月初, 于基本无明显降水、土壤水盐状况相对稳定的期间进行取样.在选定的4个样地中各随机选取7个5 m × 5 m的样方, 共计28个样方.首先分别测定每个样方内湿地群落所有植物的高度和密度, 芨芨草以丛为单位; 然后在每个小样方随机挖掘 1 m × 1 m × 0.5 m的土壤剖面, 用环刀(200 cm
3 )分5层间隔10 cm采取土样, 重复3次, 现场编号、各土层均匀混合称鲜质量, 然后带回实验室在105 ℃的烘箱内烘干12 h, 取出称质量, 计算出各样地0-50 cm土层土壤质量含水量(
韩玲等, 2016 ); 最后在每一个样方选择6丛芨芨草(株高为大中小各2丛), 用卷尺测量从基部到最高分枝的高度, 定为株高, 并记录芨芨草株丛分枝数, 然后选取每个株丛最外层4个方位充分伸展且健康完整的2-3个叶片, 做好标记后进行以下步骤: ...
... (1)植物光合参数测定.于8月15-18日(代表植物生长旺盛期), 选择晴朗天气的9:00-12:00进行气体交换参数的测定.光合测定使用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany), 测量过程中使用人工红蓝光源, 光合有效辐射(
PAR )为1β200 μmol·m
-2 ·s
-1 , CO
2 浓度约为340 μmol·mol
-1 , 相对湿度(
RH )保持在40%-50%, 流速设定为 750 μmol·s
-1 , 叶室面积为8 cm
2 , 对做好标记的每个叶片记录5组数据用于统计分析, 分别测定叶片的净光合速率(
P n )、蒸腾速率(
T r )等参数(
任青吉等, 2015 ), 对于不能充满叶室的叶片, 则保存于湿润的封口袋中, 带回实验室, 用便携式激光叶面积仪(CI-202, Walz, Camas, USA)测定叶面积, 进而计算
P n 和
T r , 水分利用效率(
WUE ) =
P n /
T r (
韩玲等, 2016 ). ...
... (2)光合有效辐射测定.于8月15-18日用手持光量子计(3415F, Walz, Plainfield, USA)在9:00-11:00对4个土壤水分梯度内所有供试芨芨草株丛进行
PAR 测定, 分别选择植株上方、中层(株高1/2处)和地表处向外距灌层15 cm处测量(
韩玲等, 2016 ). ...
... (4)土壤盐分测定.用电导法测定土壤含盐量, 在室温下称取过2 mm筛的风干土样10 g, 加50 mL去CO
2 蒸馏水(水土质量比为5:1), 取浸出液, 置振荡机上振荡5 min.将布氏漏斗与抽气系统相连后把悬浊的土浆缓缓倒入漏斗, 直至抽滤完毕, 滤液倒入三角瓶备用(
张雅琼等, 2010 ).用上海雷磁仪器厂生产的DDS-11C便携式电导仪测浸出液的电导率, 3次重复, 取平均值即为土壤电导率值(
韩玲等, 2016 ). ...
... 对湿地群落中4个水分梯度中芨芨草叶片厚度、叶脉密度和叶脉直径的实验数据先进行以10为底的对数转换, 使之符合正态分布后再进行分析.用于各土壤水分梯度中任意两个叶性状的异速生长方程的参数估计, 主要采用标准化主轴估计(SMA)方法(
Warton et al ., 2006 ), 采用
y = axb , 线性转换成log(
y ) =
b log(
x ) + log(
a ), 式中
x 和
y 表示两个特征参数,
a 为性状关系的截距,
b 为斜率, 即异速生长参数或相对生长的指数, 当|
b | = 1时, 表示两者是等速变化关系; 当
|b| 显著偏离1时, 两者间为异速变化关系(
Harvey & Pagel, 1991 ), 由软件SMATR Version 2.0 (
Falster et al ., 2012 )计算完成, 每一个回归斜率的置信区间根据Pitman (1939)方法计算, 并采用Warton和Weber (2002)的方法对每个密度的斜率进行异质性测试.不同水分梯度间植物功能性状平均值的差异比较采用单因素方差分析(
a = 0.05).实验所用数据采用SPSS 16.0软件进行处理分析, 用Excel软件绘图. ...
张掖湿地芨芨草叶大小和叶脉密度的权衡关系
1
2016
... 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 ).研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 ).在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 ).主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新. ...
不同坡向高寒退化草地狼毒株高和枝条数的权衡关系
1
2014
... 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 ).研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 ).在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 ).主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新. ...
Altitude trends in conifer leaf morphology and stable carbon isotope composition.
1
2000
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Controlled alternate partial root- zone irrigation: Its physiological consequences and impact on water use efficiency.
4
2004
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
... 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 ).研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 ).在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 ).主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
木本植物木质部栓塞脆弱性研究新进展
1
2015
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Repeated origin of three-dimensional leaf venation releases constraints on the evolution of succulence in plants.
1
2013
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Decoding leaf hydraulics with a spatially explicit model: Principles of venation architecture and implications for its evolution.
2
2010
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 叶片水力阻力是植物应对环境水分和光照胁迫的主要限制因子之一(
Scoffoni et al ., 2011 ), 被视作植物水分传输过程中的一个水力瓶颈, 是应对灾难性水力失调的安全阀门(
Villagra et al ., 2013 ).分析叶片厚度与叶脉密度、叶脉直径的异速生长关系对认识叶片适应不同水分环境具有重要作用.为了缓解生理活动中的水分胁迫, 植物往往在叶片水分防御和输导的构造上进行资源权衡投资(
许洺山等, 2015 ), 这影响着植物叶片的形态构建及代谢活动中的碳分配(
Moles et al ., 2014 ).本研究发现, 在不同土壤水分条件下, 芨芨草叶片厚度与叶脉密度、叶脉直径间存在不同的生长关系(
p < 0.05), 在样地I和IV, SMA斜率的绝对值均与1.0存在显著差异(
p < 0.05), 在样地II和III, SMA斜率的绝对值与1.0无显著差异(
p > 0.05)(
图2 ,
图3 ).不同土壤水分样地芨芨草叶性状的异速生长关系及敏感性差异, 表明湿地植物在维持水力安全和碳利用效率间存在着潜在的权衡机制, 对于理解和预测不同环境下植物的存活、生长、繁衍和分布具有重要的意义. ...
Which is a better predictor of plant traits: Temperature or precipitation?
1
2014
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes.
2014
Trade-offs between leaf hydraulic capacity and drought vulnerability: Morpho- anatomical bases, carbon costs and ecological consequences.
1
2012
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Leaf miners help us understand leaf hydraulic design.
2
2010
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Quantifying succulence: A rapid, physiologically meaningful metric of plant water storage.
1
2012
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Patterns of plant species richness in relation to different environments: An appraisal.
1
2001
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
Evidence for a plant community economics spectrum driven by nutrient and water limitations in a Mediterranean rangeland of southern France.
1
2012
... 叶形态和生理特性在植物碳同化、水分关系和能量平衡等方面有重要作用, 决定了叶片功能属性在不同环境条件下的差异(
Wright et al ., 2004 ;
Poorter & Bongers, 2006 ).芨芨草喜生于地下水埋深1.5 m左右的盐碱滩沙质土壤上(
张明娟等, 2012 ), 在地下水埋深为1.0-2.0 m, 且土壤水分适中的样地II和样地III, 湿地群落环境中群落高度、密度和地表土壤水分、理化过程和强度均处于干旱到湿润的过渡区, 种间对空间、光和土壤水分、养分的竞争有所减弱(
表1 ), 比较适宜芨芨草的生长, 芨芨草
P n 和株高最大(
表2 ), 使其能利用较为充足的物质和能量资源进行高效的光合作用和物质运输, 芨芨草选择了折中水分输送和存储结构的稳步投资, 形成了叶片厚度与叶脉密度、叶脉直径均近等速生长的格局(
p < 0.05、
图2 -3), 也是叶片厚度由薄到厚、叶脉密度由小到大、叶脉直径由粗到细转变的区域, 即在该环境中保持较高的光合能力和竞争力, 保证了植株的顺利生长和种群的繁衍更新, 从而成为湿地群落中的优势种. ...
Leaf traits are good predictors of plant performance across 53 rain forest species.
1
2006
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
帽儿山17个种源落叶松针叶的水分利用效率比较
1
2015
... (1)植物光合参数测定.于8月15-18日(代表植物生长旺盛期), 选择晴朗天气的9:00-12:00进行气体交换参数的测定.光合测定使用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany), 测量过程中使用人工红蓝光源, 光合有效辐射(
PAR )为1β200 μmol·m
-2 ·s
-1 , CO
2 浓度约为340 μmol·mol
-1 , 相对湿度(
RH )保持在40%-50%, 流速设定为 750 μmol·s
-1 , 叶室面积为8 cm
2 , 对做好标记的每个叶片记录5组数据用于统计分析, 分别测定叶片的净光合速率(
P n )、蒸腾速率(
T r )等参数(
任青吉等, 2015 ), 对于不能充满叶室的叶片, 则保存于湿润的封口袋中, 带回实验室, 用便携式激光叶面积仪(CI-202, Walz, Camas, USA)测定叶面积, 进而计算
P n 和
T r , 水分利用效率(
WUE ) =
P n /
T r (
韩玲等, 2016 ). ...
玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较
2
2015
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto.
1
2014
... (3)叶片厚度和叶脉性状的测定.基于每个芨芨草被标记的2-3个叶片, 采集后保存于湿润的封口袋中, 带回实验室用福尔马林-乙酸溶液(37%甲醛溶液, 50%乙醇和13%冰醋酸溶液)固定(
徐婷等, 2016 ).①先进行叶片厚度(单位: mm)的测定, 使用游标卡尺测量(精确度0.01 mm), 测量时尽量选择叶片中心部位, 并避开叶脉, 每个叶片重复测量5次, 最后取平均值为叶片厚度; ②叶脉性状的测定, 先用5% NaOH的乙醇进行化学清理、番红-固绿染色(
Berlyn & Miksche, 1976 )、用水包埋呈透明薄膜状, 然后在体视显微镜(SMZ168-BL, Motic, 香港, 中国)下放大10倍后拍照, 每个叶片拍摄10个视野用于叶脉性状的观测, 最后用Motic Images Plus 2.0软件获得各个照片中的叶脉直径和叶脉的总长度, 所有照片中得出的平均值即为叶脉直径和叶脉密度; ③最后用便携式激光叶面积仪(CI-202, Walz, Camas, USA)对每个叶片进行扫描, 得到叶面积, 叶脉密度用单位叶面积(单位: mm
2 )的叶脉总长度(单位: mm)表示(
Sack et al ., 2012 ). ...
Developmentally based scaling of leaf venation architecture explains global ecological patterns.
2
2012
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future.
3
2013
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis.
3
2013
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
... 叶片水力阻力是植物应对环境水分和光照胁迫的主要限制因子之一(
Scoffoni et al ., 2011 ), 被视作植物水分传输过程中的一个水力瓶颈, 是应对灾难性水力失调的安全阀门(
Villagra et al ., 2013 ).分析叶片厚度与叶脉密度、叶脉直径的异速生长关系对认识叶片适应不同水分环境具有重要作用.为了缓解生理活动中的水分胁迫, 植物往往在叶片水分防御和输导的构造上进行资源权衡投资(
许洺山等, 2015 ), 这影响着植物叶片的形态构建及代谢活动中的碳分配(
Moles et al ., 2014 ).本研究发现, 在不同土壤水分条件下, 芨芨草叶片厚度与叶脉密度、叶脉直径间存在不同的生长关系(
p < 0.05), 在样地I和IV, SMA斜率的绝对值均与1.0存在显著差异(
p < 0.05), 在样地II和III, SMA斜率的绝对值与1.0无显著差异(
p > 0.05)(
图2 ,
图3 ).不同土壤水分样地芨芨草叶性状的异速生长关系及敏感性差异, 表明湿地植物在维持水力安全和碳利用效率间存在着潜在的权衡机制, 对于理解和预测不同环境下植物的存活、生长、繁衍和分布具有重要的意义. ...
Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture.
3
2011
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Light induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse Hawaiian lobeliads.
1
2015
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Introduction to Plant Population Biology. Blackwell
1
2001
... 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 ).研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 ).在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 ).主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新. ...
红池坝草地常见物种叶片性状沿海拔梯度的响应特征
1
2012
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
Hydraulic constraints on plant gas exchange.
1
2000
... 叶片水力阻力是植物应对环境水分和光照胁迫的主要限制因子之一(
Scoffoni et al ., 2011 ), 被视作植物水分传输过程中的一个水力瓶颈, 是应对灾难性水力失调的安全阀门(
Villagra et al ., 2013 ).分析叶片厚度与叶脉密度、叶脉直径的异速生长关系对认识叶片适应不同水分环境具有重要作用.为了缓解生理活动中的水分胁迫, 植物往往在叶片水分防御和输导的构造上进行资源权衡投资(
许洺山等, 2015 ), 这影响着植物叶片的形态构建及代谢活动中的碳分配(
Moles et al ., 2014 ).本研究发现, 在不同土壤水分条件下, 芨芨草叶片厚度与叶脉密度、叶脉直径间存在不同的生长关系(
p < 0.05), 在样地I和IV, SMA斜率的绝对值均与1.0存在显著差异(
p < 0.05), 在样地II和III, SMA斜率的绝对值与1.0无显著差异(
p > 0.05)(
图2 ,
图3 ).不同土壤水分样地芨芨草叶性状的异速生长关系及敏感性差异, 表明湿地植物在维持水力安全和碳利用效率间存在着潜在的权衡机制, 对于理解和预测不同环境下植物的存活、生长、繁衍和分布具有重要的意义. ...
Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species.
1
2013
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
中国森林叶片功能属性的纬度格局及其影响因素
2015
Common slope tests for bivariate errors-in-variables models.
1
2002
... 对湿地群落中4个水分梯度中芨芨草叶片厚度、叶脉密度和叶脉直径的实验数据先进行以10为底的对数转换, 使之符合正态分布后再进行分析.用于各土壤水分梯度中任意两个叶性状的异速生长方程的参数估计, 主要采用标准化主轴估计(SMA)方法(
Warton et al ., 2006 ), 采用
y = axb , 线性转换成log(
y ) =
b log(
x ) + log(
a ), 式中
x 和
y 表示两个特征参数,
a 为性状关系的截距,
b 为斜率, 即异速生长参数或相对生长的指数, 当|
b | = 1时, 表示两者是等速变化关系; 当
|b| 显著偏离1时, 两者间为异速变化关系(
Harvey & Pagel, 1991 ), 由软件SMATR Version 2.0 (
Falster et al ., 2012 )计算完成, 每一个回归斜率的置信区间根据Pitman (1939)方法计算, 并采用Warton和Weber (2002)的方法对每个密度的斜率进行异质性测试.不同水分梯度间植物功能性状平均值的差异比较采用单因素方差分析(
a = 0.05).实验所用数据采用SPSS 16.0软件进行处理分析, 用Excel软件绘图. ...
Bivariate line-fitting methods for allometry.
1
2006
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
Study on soil moisture characteristics and water use efficiency of
1
2004
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
Specific leaf area and leaf dry matter content as alternative predictors of plant strategies.
1
1999
... 叶形态和生理特性在植物碳同化、水分关系和能量平衡等方面有重要作用, 决定了叶片功能属性在不同环境条件下的差异(
Wright et al ., 2004 ;
Poorter & Bongers, 2006 ).芨芨草喜生于地下水埋深1.5 m左右的盐碱滩沙质土壤上(
张明娟等, 2012 ), 在地下水埋深为1.0-2.0 m, 且土壤水分适中的样地II和样地III, 湿地群落环境中群落高度、密度和地表土壤水分、理化过程和强度均处于干旱到湿润的过渡区, 种间对空间、光和土壤水分、养分的竞争有所减弱(
表1 ), 比较适宜芨芨草的生长, 芨芨草
P n 和株高最大(
表2 ), 使其能利用较为充足的物质和能量资源进行高效的光合作用和物质运输, 芨芨草选择了折中水分输送和存储结构的稳步投资, 形成了叶片厚度与叶脉密度、叶脉直径均近等速生长的格局(
p < 0.05、
图2 -3), 也是叶片厚度由薄到厚、叶脉密度由小到大、叶脉直径由粗到细转变的区域, 即在该环境中保持较高的光合能力和竞争力, 保证了植株的顺利生长和种群的繁衍更新, 从而成为湿地群落中的优势种. ...
The worldwide leaf economics spectrum.
1
2004
... 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 ).研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 ).在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 ).主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新. ...
基于
δ D、
δ 18 O的青海湖流域芨芨草水分利用来源变化研究
3
2015
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
... 叶片水力阻力是植物应对环境水分和光照胁迫的主要限制因子之一(
Scoffoni et al ., 2011 ), 被视作植物水分传输过程中的一个水力瓶颈, 是应对灾难性水力失调的安全阀门(
Villagra et al ., 2013 ).分析叶片厚度与叶脉密度、叶脉直径的异速生长关系对认识叶片适应不同水分环境具有重要作用.为了缓解生理活动中的水分胁迫, 植物往往在叶片水分防御和输导的构造上进行资源权衡投资(
许洺山等, 2015 ), 这影响着植物叶片的形态构建及代谢活动中的碳分配(
Moles et al ., 2014 ).本研究发现, 在不同土壤水分条件下, 芨芨草叶片厚度与叶脉密度、叶脉直径间存在不同的生长关系(
p < 0.05), 在样地I和IV, SMA斜率的绝对值均与1.0存在显著差异(
p < 0.05), 在样地II和III, SMA斜率的绝对值与1.0无显著差异(
p > 0.05)(
图2 ,
图3 ).不同土壤水分样地芨芨草叶性状的异速生长关系及敏感性差异, 表明湿地植物在维持水力安全和碳利用效率间存在着潜在的权衡机制, 对于理解和预测不同环境下植物的存活、生长、繁衍和分布具有重要的意义. ...
浙东常绿阔叶林植物功能性状对土壤含水量变化的响应
2
2015
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
... (3)叶片厚度和叶脉性状的测定.基于每个芨芨草被标记的2-3个叶片, 采集后保存于湿润的封口袋中, 带回实验室用福尔马林-乙酸溶液(37%甲醛溶液, 50%乙醇和13%冰醋酸溶液)固定(
徐婷等, 2016 ).①先进行叶片厚度(单位: mm)的测定, 使用游标卡尺测量(精确度0.01 mm), 测量时尽量选择叶片中心部位, 并避开叶脉, 每个叶片重复测量5次, 最后取平均值为叶片厚度; ②叶脉性状的测定, 先用5% NaOH的乙醇进行化学清理、番红-固绿染色(
Berlyn & Miksche, 1976 )、用水包埋呈透明薄膜状, 然后在体视显微镜(SMZ168-BL, Motic, 香港, 中国)下放大10倍后拍照, 每个叶片拍摄10个视野用于叶脉性状的观测, 最后用Motic Images Plus 2.0软件获得各个照片中的叶脉直径和叶脉的总长度, 所有照片中得出的平均值即为叶脉直径和叶脉密度; ③最后用便携式激光叶面积仪(CI-202, Walz, Camas, USA)对每个叶片进行扫描, 得到叶面积, 叶脉密度用单位叶面积(单位: mm
2 )的叶脉总长度(单位: mm)表示(
Sack et al ., 2012 ). ...
兰州北山刺槐不同等级叶脉密度与叶大小关系的坡向差异性
1
2016
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析
1
2014
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
Evolutionary association of stomatal traits with leaf vein density in
1
2012
... 植物叶片的水分输导及动态平衡是SPAC系统中水分迁移的一个重要环节, 不仅受植物本身的调节和控制, 而且还受外界环境条件的影响(
张志亮等, 2014 ), 水流通过直径递减的一系列叶脉从维管束木质部末端进入表皮细胞和气孔下腔附近的蒸发部位, 叶片中的水分运输路径及效率对整个植物体的叶片几何构型构建、水力特性及资源的获取与权衡利用效率等具有较大的影响(
Sack & Scoffoni, 2013 ).叶片功能性状作为联系植物生理适应性和环境梯度间的桥梁, 叶片厚度影响着叶片的水分供应、存储以及光合作用中物质和能量的交换过程(
Scoffoni et al ., 2011 ;
Ogburn & Edwards, 2012 ), 叶脉密度和叶脉直径可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力(
Brodribb & Jordan, 2011 ;
Sack & Scoffoni, 2013 ).面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾, 叶片各功能性状的自我调整与适应显得至关重要(
许洺山等, 2015 ), 叶片厚度的资源投入有利于减少水分的蒸腾散失并提高固碳能力(
Gago et al ., 2014 ;
全先奎和王传宽, 2015 ); 较粗直径导管的高水分流动速率提高了叶脉水分输导的“有效性”, 却降低了水分运输中的“安全性” (
Givnish, 1987 ;
Matthew-Ogburn & Edwards, 2013 ), 而增加叶脉密度能够增加叶片水分供应的面积和路径, 使得水分运输的安全性得到保障(
Nardini et al ., 2010 ), 三者的资源经济权衡机制与协同进化策略共同对叶片水分供应与散失的平衡、植物生理活动的能量供给和生存适应方面具有不可替代的调控作用, 体现了叶脉和叶性状间存在着蒸腾成本和光合收益的匹配和权衡关系(
Sack et al ., 2013 ).干旱区土壤水分格局作为影响植物水分来源及利用策略的关键因子, 与光照和生存空间等多重环境变化及选择压力的交互作用对植物性状的资源获取方式和生理活性的表达具有直接的限制作用(
Pausas & Austin, 2001 ).土壤水分亏缺会限制植物固碳功能的发挥及引起植物体木质部空穴化和栓塞现象的产生, 进而影响叶片的气体交换和植物的正常生长发育(
李荣等, 2015 ;
许洺山等, 2015 ).叶片厚度和叶脉结构等功能性状能够敏锐地感应叶片的输水阻力对生理功能的阻碍(
Scoffoni et al ., 2015 ), 为有效地加强植物体内的水分运输与利用效率和进一步增强现实竞争力, 植物在叶片的叶肉组织和叶脉网络系统的构造上进行权衡投资(
Wilson et al ., 1999 ;
Reich & Cornelissen, 2014 ), 促使植物调整叶脉结构和叶片厚度的生物量分配和叶片构型构建, 从而提高植物对土壤水分的有效利用(
Moles et al ., 2014 ;
王瑞丽等, 2015 ), 实现叶片生理特性和环境空间异质性的精细协调, 体现了植物最经济的生境适应性和表型可塑性机制(
Silvertown & Charlesworth, 2001 ), 即植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(
Nardini & Luglio, 2014 ).因此, 研究湿地群落芨芨草叶片厚度和叶脉性状对不同土壤水分梯度的响应规律, 可以探索湿地植物碳水代谢关系和高效用水的生理生态学机制. ...
植物叶片导水率的研究进展
3
2014
... 芨芨草(
Achnatherum splendens )是我国西北地区常见的多年生密丛性草本植物, 草秆坚硬、直立丛生、根系发达、须根较多, 叶片拥有较强的叶脉机械支撑能力和水分输导组织, 芨芨草并非喜盐植物, 与典型盐生植物相比, 仅是生态幅较宽的耐盐旱生物种, 通常发育于轻度盐化的河湖边缘以及低洼地等隐域性生境, 是一种优良的饲用植物、纤维植物和水土保持植物(
张明娟等, 2012 ;
韩玲等, 2016 ).近年来, 叶脉功能性状与叶片厚度(
Scoffoni et al ., 2011 )、叶大小(
Scoffoni et al ., 2015 ;
徐婷等, 2016 )的异速生长关系、叶脉网络性状与叶片水力特性(
Blonder et al ., 2010 ;
Zhang et al ., 2012 )、叶片碳构建(
Reich & Cornelissen, 2014 )的关联性, 以及植物输水结构与耐旱性的关系(
李荣等, 2015 )、叶片功能属性沿水分和光照梯度的变化趋势(
Domínguez et al ., 2012 ;
Perez-Ramos et al ., 2012 )等方面已进行了大量的研究, 解释了植物适应特定环境的生理生态学机制; 芨芨草的水分利用策略(
Wang et al ., 2004 )等也受到了关注, 但芨芨草叶片形态和叶脉性状的异速生长规律研究不深入, 尤其是湿地群落不同土壤水分生境下叶片厚度与叶脉密度、叶脉直径的生长关系对植物生理功能和异质性资源分布的响应机理尚不清晰.鉴于此, 本文以张掖湿地芨芨草种群为研究对象, 探讨张掖湿地群落不同土壤水分梯度下芨芨草叶片厚度-叶脉密度, 叶片厚度-叶脉直径的相互关系及变化规律, 旨在理解不同环境下植物叶片的资源权衡分配和物理构建方式, 有助于揭示湿地植物为获得最大光合生产力所形成的水资源利用策略和生理生态适应性. ...
... 植物叶脉系统水分运输的有效性与抗栓塞的安全性有一定的动态权衡, 这种权衡性可能是水分有效运输的最佳值(
Scoffoni et al ., 2015 ).单位叶面积上木质部通道较多的植物, 其叶片的流体传导性和叶水力导度更强(
Sack & Scoffoni, 2013 ), 有利于输送水分、养分和光合产物(
Brodribb et al ., 2007 ;
McKown et al ., 2010 ).湿地群落土壤含水量最小的样地IV, 群落高度和密度显著减小(
表1 ), 芨芨草接收到的
PAR 最大(
表1 ), 随着土壤水分潜在蒸散的增大, 伴随着土壤电导率最大(
表1 ), 盐胁迫引起土壤水势降低, 诱导植物发生水分胁迫(
Kang & Zhang, 2004 ;
李荣等, 2015 ), 与典型盐生植物相比, 芨芨草并非喜盐植物(
张明娟等, 2012 ), 在水盐胁迫为主的条件下, 往往易造成植物木质部导管栓塞(
Sperry, 2000 ;
龚容和高琼, 2015 ), 芨芨草水分输导的有效性和安全性显著下降, 促使芨芨草进行叶性状间的资源权衡分配, 并构建与其相适应的叶片几何构型以缓解植物体水分胁迫, 芨芨草趋向于叶片厚度的增加速率小于叶脉密度增加速率、大于叶脉直径减小速率的异速生长模式(
p < 0.05,
图2 ,
图3 ), 主要原因有: (1)面对该生境中较高的
PAR , 芨芨草选择增加叶片厚度的资源投资, 既能通过增大水蒸气进出表皮阻力降低叶片蒸腾速率(
表2 ), 也能减少植物内部的水分散失(
Ogburn & Edwards, 2012 ), 且厚叶片较长的栅栏细胞和更多的细胞层数能减少光辐射的穿透量, 又避免了强光对叶片的伤害(
于鸿莹等, 2014 ); (2)该生境中土壤含水量较样地I显著减少59.26%, 为了降低水分胁迫下叶片水力功能紊乱和脱水的风险, 芨芨草增加叶脉密度使得水分运输的路径增多, 叶片可以绕过气蚀化的木质部把水分运输到蒸腾部位的细胞(
Sack et al ., 2013 ), 且植物小径叶脉的导管壁厚(
Givnish, 1987 ), 芨芨草减小叶脉直径的资源分配, 降低了导管坍塌和破坏的几率, 大大提高了水分运输的安全性和水分利用效率(
表2 ), 与Nardini (2012)等关于“直径小且数量多的叶脉水分运输效率高”一致, 体现了芨芨草通过环境因子对植物水力结构的影响和驯化, 减少了营养的“流失”, 并提高应对水分胁迫、维持光合生产和自我保护的调控能力. ...
... 叶形态和生理特性在植物碳同化、水分关系和能量平衡等方面有重要作用, 决定了叶片功能属性在不同环境条件下的差异(
Wright et al ., 2004 ;
Poorter & Bongers, 2006 ).芨芨草喜生于地下水埋深1.5 m左右的盐碱滩沙质土壤上(
张明娟等, 2012 ), 在地下水埋深为1.0-2.0 m, 且土壤水分适中的样地II和样地III, 湿地群落环境中群落高度、密度和地表土壤水分、理化过程和强度均处于干旱到湿润的过渡区, 种间对空间、光和土壤水分、养分的竞争有所减弱(
表1 ), 比较适宜芨芨草的生长, 芨芨草
P n 和株高最大(
表2 ), 使其能利用较为充足的物质和能量资源进行高效的光合作用和物质运输, 芨芨草选择了折中水分输送和存储结构的稳步投资, 形成了叶片厚度与叶脉密度、叶脉直径均近等速生长的格局(
p < 0.05、
图2 -3), 也是叶片厚度由薄到厚、叶脉密度由小到大、叶脉直径由粗到细转变的区域, 即在该环境中保持较高的光合能力和竞争力, 保证了植株的顺利生长和种群的繁衍更新, 从而成为湿地群落中的优势种. ...
不同密度条件下芨芨草空间格局对环境胁迫的响应
1
2012
... (4)土壤盐分测定.用电导法测定土壤含盐量, 在室温下称取过2 mm筛的风干土样10 g, 加50 mL去CO
2 蒸馏水(水土质量比为5:1), 取浸出液, 置振荡机上振荡5 min.将布氏漏斗与抽气系统相连后把悬浊的土浆缓缓倒入漏斗, 直至抽滤完毕, 滤液倒入三角瓶备用(
张雅琼等, 2010 ).用上海雷磁仪器厂生产的DDS-11C便携式电导仪测浸出液的电导率, 3次重复, 取平均值即为土壤电导率值(
韩玲等, 2016 ). ...
芨芨草群落土壤盐分特征
1
2010
... 植物的水分适应能力是长期适应环境的一种复杂的遗传特性, 与植物形态结构、内部生理生化活动和外界条件等密切相关(
李荣等, 2015 ), 植物会通过调整光合构件生长率、权衡资源分配等长期的表型可塑性来增加异质性资源的觅食效率(
侯兆疆等, 2014 ).研究区属于河西走廊温带干旱区的天然湿地生态系统, 土壤水和地下水可能是芨芨草的主要利用水源(
吴华武等, 2015 ).在土壤水分较高的样地I, 湿地群落密度和高度最大(
表1 ), 光竞争成为植被生长中面对的首要问题, 群落冠层内的芨芨草获得的
PAR 更少(
表1 ,
表2 ), 为了保证自身的生理活性和体内水分的动态平衡, 芨芨草增加分枝数以获取更多的光资源(
表2 ), 并进行叶内生物量的权衡分配和叶片构型塑造的调整, 形成了增加叶脉直径, 减小叶片厚度和叶脉密度的资源分配机制, 形成了叶片厚度生长速度显著大于叶脉密度的生长速度,而小于叶脉直径减小速率的异速生长格局(
p < 0.05,
图2 ,
图3 ).主要原因有: (1)该样地中芨芨草株高较小(
表2 ), 为了保证有限光环境中最大的碳同化速率和光合能力, 需加强叶脉直径的营养物质投资, 以较强的机械支撑强度支持更大的展叶面积, 伴随着叶片厚度的减小(
表2 ), 薄叶片能够使CO
2 和水分等从气孔到达叶绿体的路径缩短, 有利于光的透过和光能的吸收(
Hultine & Marshall, 2000 ;
宋璐璐等, 2012 ), 进而提高了植物的生存适合度和群落竞争力; (2)样地中土壤含水量较高时植物木质部导水速率较高(
周洪华等和李卫红, 2015 ), 叶片水分充足, 较小的叶脉密度也能满足较高蒸腾速率(
T r )的水分需求(
表2 ), 芨芨草以有限的生物量增加叶脉直径来提高水分运输的量与速率, 消除了叶脉密度变小的弊端, 同时通过减小叶片厚度减少了水分输送阻力, 以确保植物保持自身碳、矿物质以及水分的收支平衡, 从而保证了芨芨草种群的顺利繁衍和更新. ...
胡杨木质部水分传导对盐胁迫的响应与适应
2015
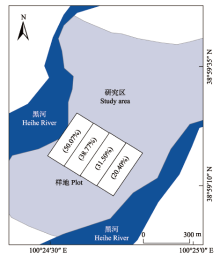 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT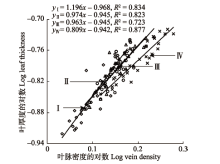 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT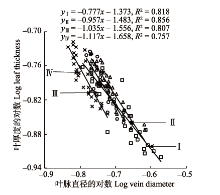 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}