Effects of glyphosate on interspecific competition between Solidago canadensis and Imperata cylindrica
GUChun-Feng1,2, YEXiao-Qi2, WUMing2,*,, SHAOXue-Xin2, JIAOSheng-Wu2 1College of Life and Environment Sciences, Hangzhou Normal University, Hangzhou 310036, Chinaand 2Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Wetland Ecosystem Research Station of Hangzhou Bay, State Forestry Administration, Fuyang, Zhejiang 311400, China 通讯作者:* 通信作者Author for correspondence (E-mail: hangzhoubay@126.com) 责任编辑:GUChun-FengYEXiao-QiWUMingSHAOXue-XinJIAOSheng-Wu 收稿日期:2016-10-27 接受日期:2017-02-28 网络出版日期:2017-04-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家林业公益性行业科研专项(201404210)、国家自然科学基金(31400378)、浙江省公益技术研究农业项目(2015C32011)、浙江省-中国林业科学院科技合作重大项目(2015SY01)
关键词:除草剂;入侵植物;本地植物;生长;种间关系 Abstract Aims The extensive use of herbicide to control invasive plants would change the relationship between alien and neighboring plants. In order to provide data for rational use of herbicide and a theoretical reference for further studies on the ecological effects of glyphosate, we explored the variation of the relationship between an invasive plant Solidago canadensis and a native plant Imperata cylindrica when they were sprayed glyphosate. Methods A replacement series experiment was conducted from June to August 2016 in Wetland Ecosystem Research Station of Hangzhou Bay, State Forestry Administration, to examine the effects of glyphosate at seven concentration levels (0, 0.3, 0.6, 0.9, 1.2, 1.5, 1.8 mL·L-1) on the growth and interspecific competition of S. canadensis and I. cylindrica. Important findings (1) Glyphosate treatment significantly inhibited the growth of S. canadensis and I. cylindrica (p < 0.05). During the test, cumulative growth of height and leaf number of S. canadensis were apparently reduced with the increase of glyphosate concentration, but the leaf number of S. canadensis treated with 0.3- 1.5 mL·L-1 glyphosate was re-growing with time, while the one treated with 1.8 mL·L-1 was mostly dead. The withering rate of tiller and green leaf of I. cylindrica also significantly increased with the increase of glyphosate concentration, and the growth indices of this plant treated with 0.3-0.6 mL·L-1 were also re-growing with time. (2) Glyphosate treatment significantly affected interspecific competition (p < 0.05), which diminished as the glyphosate concentration increased. (3) Interspecific competition has significant influence on the biomass allocation of S. canadensis (p < 0.05). When facing competition, S. canadensis would allocate more organic matter to root and thus increase the ratio of root to shoot. Competition only inhibited the tiller number and total biomass of I. cylindrica, but insignificantly affected its ratio of root to shoot. (4) The interaction between glyphosate treatment and S. canadensis-I. cylindrica interspecific relationship also significantly influenced the biomass of S. canadensis and I. cylindrica (p < 0.05), but insignificantly affected the root/shoot ratio of two plants. Different plants have different tolerance to glyphosate stress. Compared with native plant I. cylindrica, S. canadensis has stronger tolerance to glyphosate. Low-concentration glyphosate could decrease the competitive intensity between S. canadensis and I. cylindrica, which may disturb the structure and dynamics of plant communities.
Keywords:herbicide;invasive plant;native plant;growth;interspecific relationship -->0 PDF (1365KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 古春凤, 叶小齐, 吴明, 邵学新, 焦盛武. 草甘膦对加拿大一枝黄花和伴生植物白茅种间竞争关系的影响. 植物生态学报, 2017, 41(4): 439-449 https://doi.org/10.17521/cjpe.2016.0331 GUChun-Feng, YEXiao-Qi, WUMing, SHAOXue-Xin, JIAOSheng-Wu. Effects of glyphosate on interspecific competition between Solidago canadensis and Imperata cylindrica. Chinese Journal of Plant Ecology, 2017, 41(4): 439-449 https://doi.org/10.17521/cjpe.2016.0331 随着全球变化加剧以及我国对外经济的发展, 外来植物的入侵频率和危害程度正在迅速增加, 现已成为我国重大环境问题(鞠瑞亭等, 2012; Pan et al., 2015)。入侵植物严重威胁了入侵地的生物多样性, 干扰了原生态系统的结构, 甚至引起生态系统服务功能退化, 造成我国每年超过70亿美元的经济损失(黄乔乔等, 2012)。防治外来植物普遍使用化学防除、人工拔除等方法(强胜, 2009)。有的****提议利用替代植被抵御外来植物入侵或者引进天敌以恢复稳定的生态平衡(徐正浩和王一平, 2004; Funk et al., 2008; 张瑞海, 2010), 这类生物防治法具有控制效果持久、对环境安全等优点, 但仍需要长期评估其生态风险性, 因此并未被大面积采用(强胜等, 2010); 由于人工拔除法存在成本过高的缺点, 目前我国防治外来植物主要依赖于经济、高效的化学防除法。自然条件下, 植物种内或种间普遍存在相互作用(Maestre et al., 2009), 正、负相互作用的动态平衡对植物群落结构和生态系统功能稳定有重要影响(Callaway, 2002; Brooker, 2008)。当受到环境胁迫或者生物因素影响时, 植物邻体之间的相互作用将会发生变化(Maestre et al., 2009; He et al., 2013; Zhang et al., 2013)。大量使用化学除草剂防除外来植物、抑制植物生长发育的同时, 必然直接或间接改变植物物种之间的相互联系(Brooker, 1998; Weidenhamer, 2010), 由此可能干扰植物群落的自然演替, 破坏当地生态系统的动态平衡。 加拿大一枝黄花(Solidago canadensis)为菊科一枝黄花属(Solidago)的多年生草本植物, 原产于北美, 20世纪初作为观赏花卉引进我国上海、南京等地, 随后迅速向外扩散蔓延, 广泛入侵华东沿海湿地生态系统, 现已成为我国十大恶性杂草之一(董梅等, 2006; 陆彦等, 2006)。白茅(Imperata cylindrica)是多年生根茎禾草, 是常见的生长于河岸滩涂湿地的优势草本植物, 其植被的存在能够修复和改善脆弱的土壤环境, 有利于维持生态系统的稳定(白世红等, 2010)。长期观察发现, 在国家林业局杭州湾湿地生态系统定位观测研究站(以下简称观测站)邻近区域, 加拿大一枝黄花自然入侵到土著白茅群落, 并有不断向外蔓延的趋势。加拿大一枝黄花繁殖和抗逆境能力强, 入侵初期先与土著植物伴生, 随后抢占大量生存资源, 不断排挤土著植物而成为优势植物种(杨如意等, 2011; Gusev, 2015), 严重损害了入侵地的植物多样性、威胁原生态系统的稳定, 因此有必要采取措施防止加拿大一枝黄花的进一步入侵扩散。 草甘膦因理化性质稳定, 具有高效、低毒等优点, 是目前被大量使用的除草剂(Duke & Powles, 2008; 周垂帆等, 2013)。但目前对草甘膦影响入侵植物的研究主要集中在草甘膦的防除效果(陆彦等, 2006; Duke & Powles, 2008; 杨柳青等, 2011)、草甘膦对目标植物生理的影响(Lydon & Duke, 1988; Guo et al., 2009; 李光义等, 2010), 以及抗性或非抗性植物对草甘膦的敏感性(陈景超等, 2011; 刘文娟等, 2012)等方面, 草甘膦处理是否会影响植物之间相互作用及其机理还有待深入研究。为此, 本研究以国家林业局杭州湾湿地公园内自然入侵的加拿大一枝黄花和常见伴生植物白茅为研究对象, 设计了盆栽控制试验, 研究喷施草甘膦后加拿大一枝黄花与其伴生植物种间相互作用的变化规律, 为进一步研究草甘膦的生态效应提供理论参考, 为优化农药使用提供数据支持。
从表1可以看出, 当草甘膦作用60天后, G0.6的单种加拿大一枝黄花、白茅和混种白茅开始出现死亡植株; G1.2处理下, 白茅的死亡植株比例急剧升高, 混种白茅的植株死亡率达50%, 单种白茅的植株死亡率则达到71%; 在G1.8处理下, 加拿大一枝黄花也出现大量植株死亡的现象, 单种加拿大一枝黄花的死亡率已到46%, 混种加拿大一枝黄花的死亡率则达到64%。直到试验结束时, 最高浓度处理 (G1.8)组加拿大一枝黄花、白茅植株并未全部死亡, 故仍然收集所有处理下的植株进行相关统计分析。 Table 1 表1 表1不同浓度草甘膦处理下加拿大一枝黄花和白茅植株的累计死亡率(平均值±标准误差) Table 1Plant mortality rate of Solidago canadensis and Imperata cylindrica treated with different concentrations of glyphosate (mean ± SE)
草甘膦处理 Glyphosate treatment
植株死亡率 Plant mortality rate (%)
单种加拿大一枝黄花 S. canadensis grown in monoculture
混种加拿大一枝黄花 S. canadensis grown in mixture
单种白茅 I. cylindrica grown in monoculture
混种白茅 I. cylindrica grown in mixture
G0
0.00 ± 0.00aA
0.00 ± 0.00aA
0.00 ± 0.00aA
0.00 ± 0.00aA
G0.3
0.00 ± 0.00aA
0.00 ± 0.00aA
0.00 ± 0.00aA
0.00 ± 0.00aA
G0.6
14.28 ± 10.71aA
0.00 ± 0.00aB
14.29 ± 9.22bA
14.29 ± 9.22aA
G0.9
7.14 ± 4.61aA
7.14 ± 7.14aA
39.29 ± 12.02bA
7.14 ± 7.14aA
G1.2
25.00 ± 14.43abA
0.00 ± 0.00aB
71.43 ± 12.71cA
50.00 ± 15.43bA
G1.5
21.43 ± 8.50abA
50.00 ± 10.91bA
89.29 ± 7.43cA
57.14 ±14.86bcB
G1.8
46.43 ± 14.86bA
64.29 ± 14.28bA
92.86 ± 7.14cA
85.71 ± 9.2bA
G0, G0.3, G0.6, G0.9, G1.2, G1.5 and G1.8 represent 0, 0.3, 0.6, 0.9, 1.2, 1.5 and 1.8 mL·L-1 glyphosate, respectively. The different lowercase letters in each column indicate that there were significant differences among different concentrations (p < 0.05); the different uppercase letters in each row indicate that there were significant differences between different plant modes.G0、G0.3、G0.6、G0.9、G1.2、G1.5和G1.8分别表示草甘膦浓度为0 mL·L-1、0.3 mL·L-1、0.6 mL·L-1、0.9 mL·L-1、1.2 mL·L-1、1.5 mL·L-1、1.8 mL·L-1。同列不同小写字母表示不同处理浓度之间差异显著(p < 0.05); 同行不同大写字母表示不同种植方式之间差异显著。 新窗口打开 草甘膦处理显著抑制了加拿大一枝黄花和白茅的形态指标(p < 0.05, 表2)。7种浓度草甘膦处理后0-60天, 加拿大一枝黄花和白茅形态指标的变化见图1和图2。随着草甘膦处理浓度的升高, 加拿大一枝黄花的株高增长明显不断受到抑制; 随着处理时间的延长, 与对照相比, G0.3、G0.6、G0.9处理下单种、混种加拿大一枝黄花株高仍有增加, 但增加速率明显逐渐降低; 而G1.2、G1.5、G1.8处理下加拿大一枝黄花基本停止株高增长。加拿大一枝黄花能进行光合作用的绿叶片数同样随处理浓度递增而不断减少, 但随处理时间的延长, G0.3、G0.6、G0.9 (20-60天)和G1.2、G1.5 (40-60天)处理下, 草甘膦对植物的药效减小, 加拿大一枝黄花的绿叶数开始恢复生长。白茅分蘖的存活数和绿叶长度均随处理浓度递增而不断减少, 喷药后第20天, 白茅植株地上部分已大面积呈现发黄、枯萎等受害症状; 随着处理时间延长, G0.3 (20-60天)处理下,单种白茅的分蘖数和绿叶长度呈增加趋势; 在G0.3、G0.6 (20-60天)和G0.9 (40-60天)处理下混种白茅的分蘖数和绿叶长度也开始增加。 显示原图|下载原图ZIP|生成PPT 图1不同时期喷施草甘膦后加拿大一枝黄花的形态指标(平均值±标准误差)。A, B, 单种加拿大一枝黄花。C, D, 混种加拿大一枝黄花。G0、G0.3、G0.6、G0.9、G1.2、G1.5和G1.8同表1。 -->Fig. 1The growth indices of Solidago canadensis after spraying glyphosate at different growth stages (mean ± SE). A, B, S. canadensis grown in monoculture. C, D, S. canadensis grown in mixture. G0, G0.3, G0.6, G0.9, G1.2, G1.5 and G1.8 see Table 1. -->
显示原图|下载原图ZIP|生成PPT 图2不同时期喷施草甘膦后白茅的形态指标(平均值±标准误差)。A, B, 单种白茅。C, D, 混种白茅。G0、G0.3、G0.6、G0.9、G1.2、G1.5和G1.8同表1。 -->Fig. 2The growth indices of Imperata cylindrica after spraying glyphosate at different growth stages (mean ± SE). A, B, I. cylindrica grown in monoculture. C, D, I. cylindrica grown in mixture. G0, G0.3, G0.6, G0.9, G1.2, G1.5 and G1.8 see Table 1. -->
Table 2 表2 表2种植方式和草甘膦处理对加拿大一枝黄花和白茅生长指标的影响 Table 2Effects of planting method and glyphosate treatment on the growth of Solidago canadensis and Imperata cylindrica
生长指标 Growth index
种植方式 Planting method
草甘膦处理 Glyphosate treatment
交互作用 Interaction
F
p
F
p
F
p
加拿大一枝黄花 S. canadensis
总生物量 Total biomass
69.695
0.000**
20.433
0.000**
2.965
0.011*
地上生物量 Aboveground biomass
82.594
0.000**
20.185
0.000**
0.473
0.030*
地下生物量 Underground biomass
22.363
0.000**
11.761
0.000**
2.881
0.013*
株高 Plant height
18.520
0.000**
18.010
0.000**
3.000
0.010*
绿叶数 Green leaf number
0.604
0.439
11.192
0.000**
1.621
0.151
根冠比 Root /shoot ratio
6.773
0.011*
0.826
0.553
1.643
0.145
植株死亡率 Plant mortality rate
0.012
0.912
11.057
0.000**
2.336
0.039*
白茅 I. cylindrica
总生物量 Total biomass
37.776
0.000**
37.701
0.000**
7.802
0.000**
地上生物量 Aboveground biomass
32.195
0.000**
29.126
0.000**
5.642
0.000**
地下生物量 Underground biomass
36.954
0.000**
36.851
0.000**
8.675
0.000**
分蘖数 Tiller number
13.195
0.000**
50.831
0.000**
5.75
0.000**
绿叶长度 Length of green leaf
1.379
0.244
52.523
0.000**
1.254
0.288
根冠比 Root/shoot ratio
0.615
0.435
0.599
0.730
2.357
0.058
植株死亡率 Plant mortality rate
5.242
0.025**
34.112
0.000**
0.971
0.450
* p < 0.05; ** p < 0.01。 新窗口打开 同样的, 草甘膦处理显著抑制了加拿大一枝黄花和白茅植株生物量的积累(p < 0.05), 但对两种植物根冠比的影响均不显著(p > 0.05, 表2)。随处理浓度增加, 加拿大一枝黄花、白茅的地上茎叶、地下部分积累的生物量均不断减少(图3B、3C, 图4B、4C)。 显示原图|下载原图ZIP|生成PPT 图3不同草甘膦浓度处理对加拿大一枝黄花植株总生物量(A)、地上生物量(B)、地下生物量(C)、根冠比(D)的影响(平均值±标准误差)。不同小写、大写字母分别表示单种、混种植物在不同处理浓度之间的差异显著(p < 0.05)。G0、G0.3、G0.6、G0.9、G1.2、G1.5和G1.8同表1。 -->Fig. 3Effects of different concentrations of glyphosate on total biomass (A), aboveground biomass (B), belowground biomass (C) and root/shoot ratio (D) of Solidago canadensis (mean ± SE). Different lowercase letters indicate that the indices are significantly different among different concentrations of glyphosate (p < 0.05) for plants grown in monocultures. Different uppercase letters indicate that the indices are significantly different among different concentrations of glyphosate (p < 0.05) for plants grown in mixtures. G0, G0.3, G0.6, G0.9, G1.2, G1.5 and G1.8 see Table 1. -->
显示原图|下载原图ZIP|生成PPT 图4不同草甘膦浓度处理对白茅植株总生物量(A)、地上生物量(B)、地下生物量(C)、根冠比(D)的影响(平均值±标准误差)。不同小写、大写字母分别表示单种、混种植物在不同处理浓度之间的差异显著(p < 0.05)。G0、G0.3、G0.6、G0.9、G1.2、G1.5和G1.8同表1。 -->Fig. 4Effects of different concentrations of glyphosate on total biomass (A), aboveground biomass (B), belowground biomass (C) and root/shoot ratio (D) of Imperata cylindrica (mean ± SE). Different lowercase letters indicate that the indices are significantly different among different concentrations of glyphosate (p < 0.05) for plants grown in monocultures. Different uppercase letters indicate that the indices are significantly different among different concentrations of glyphosate (p < 0.05) for plants grown in mixtures. G0, G0.3, G0.6, G0.9, G1.2, G1.5 and G1.8 see Table 1. -->
RY表明两种植物种内、种间竞争能力的强弱。如图5所示, 草甘膦处理显著影响了加拿大一枝黄花和白茅的RY (p < 0.05), 对照组加拿大一枝黄花的RY大于1, 表明在相同种植密度条件下, 加拿大一枝黄花的种内竞争效应大于种间竞争效应, 加拿大一枝黄花受到白茅的竞争影响较弱; 而白茅的RY小于1, 表明白茅的种间竞争强度大于种内竞争, 白茅受到加拿大一枝黄花的竞争影响强烈。喷施草甘膦后, 加拿大一枝黄花的RY呈现先升高后降低的趋势(除G1.2组外), 白茅的RY也显示出相似的变化趋势(除G1.8组外), 说明白茅对加拿大一枝黄花的竞争力越来越弱, 其受到加拿大一枝黄花竞争的影响也越来越弱。 显示原图|下载原图ZIP|生成PPT 图5不同浓度草甘膦处理对加拿大一枝黄花(A)和白茅(B)相对产量的影响(平均值±标准误差)。不同小写字母表示在不同处理浓度之间差异显著(p < 0.05)。G0、G0.3、G0.6、G0.9、G1.2、G1.5和G1.8同表1。 -->Fig. 5Effects of different concentrations of glyphosate on the relative yields of Solidago canadensis (A) and Imperata cylindrica (B) (mean ± SE). Different lowercase letters indicate that relative yields are significantly different among different concentrations of glyphosate (p < 0.05). G0, G0.3, G0.6, G0.9, G1.2, G1.5 and G1.8 see Table 1. -->
BaiSH, MaFY, HouD, WangD (2010).Change in population niche during vegetation community succession in the Yellow River Delta .Chinese Journal of Eco-agriculture, 18, 581-587. (in Chinese with English abstract)[白世红, 马风云, 侯栋, 王迪 (2010). 黄河三角洲植被演替过程种群生态位变化研究 . 中国生态农业学报, 18, 581-587.] [本文引用: 1]
[2]
BanZH, WangQ (2015).Responses of the competition between Alternanthera philoxeroides and Sambucus chinensis to simulated warming .Chinese Journal of Plant Ecology, 39, 43-51. (in Chinese with English abstract)[班芷桦, 王琼 (2015). 喜旱莲子草和接骨草竞争对模拟增温的响应 . 植物生态学报, 39, 43-51.] [本文引用: 1]
[3]
BrookerRW (2008). Facilitation in plant communities: The past, the present, and the future .Journal of Ecology, 95, 1-19. [本文引用: 1]
[4]
BrookerRW (1998). The balance between positive and negative plant interactions and its relationship to environmental gradients: A model .Oikos, 81, 196-207. [本文引用: 1]
[5]
CallawayRR (2002). Positive interactions among alpine plants increase with stress .Nature, 417, 844-848. [本文引用: 1]
[6]
ChenJC, ZhangCX, HuangHJ, WeiSH, ZhangM, GuoF (2011).Sensitivity to glyphosate and dynamics of shikimate accumulation in three weed species following glyphosate application .Chinese Journal of Pesticide Science, 13, 471-474. (in Chinese with English abstract)[陈景超, 张朝贤, 黄红娟, 魏守辉, 张猛, 郭峰 (2011). 三种杂草对草甘膦的敏感性及处理后植株体内莽草酸积累量差异 . 农药学学报, 13, 471-474.] [本文引用: 1]
[7]
ChenJF, XuHL, SunYB, HuangLL, ZhangPX, ZouCP, YuB, ZhuGF, ZhaoCY (2016). Interspecific differences in growth response and tolerance to the antibiotic sulfadiazine in ten clonal wetland plants in South China .Science of the Total Environment, 543, 197-205. [本文引用: 1]
[8]
ChenT, LiuWL, ZhangCB, WangJ (2012). Effects of Solidago canadensis invasion on dynamics of native plant communities and their mechanisms .Chinese Journal of Plant Ecology, 36, 253-261. (in Chinese with English abstract)[陈彤, 刘文莉, 张崇邦, 王江 (2012). 加拿大一枝黄花入侵对本土植物群落动态的影响及其机制 . 植物生态学报, 36, 253-261.] [本文引用: 1]
[9]
DongM, LuJZ, ZhangWJ, ChenJK, LiB (2006).Canada goldenrod ( Solidago canadensis): An invasive alien weed rapidly spreading in China. Acta Phytotaxonomica Sinica, 44, 72-85. (in Chinese with English abstract)[董梅, 陆建忠, 张文驹, 陈家宽, 李博 (2006). 加拿大一枝黄花——一种正在迅速扩张的外来入侵植物 . 植物分类学报, 44, 72-85.] [本文引用: 1]
FunkJL, ClelandEE, SudingKN, ZavaletaES (2008). Restoration through reassembly: Plant traits and invasion resistance . Trends in Ecology and Evolution, 23, 695-703. [本文引用: 1]
[12]
GuoSL, JiangHW, FangF, ChenGQ (2009). Influences of herbicides, uprooting and use as cut flowers on sexual reproduction of Solidago canadensis . Weed Research, 49, 291-299. [本文引用: 1]
[13]
GusevAP (2015). The impact of invasive Canadian goldenrod (Solidago canadensis L.) on regenerative succession in old fields .Russian Journal of Biological Invasions, 6(2), 74-77. [本文引用: 1]
[14]
HeQ, BertnessMD, AltieriAH (2013). Global shifts towards positive species interactions with increasing environmental stress .Ecology Letters, 16, 695-706. [本文引用: 1]
[15]
HuangQQ, ShenYD, LiXX, ChengHT, SongX, FanZW (2012). Research progress on the distribution and invasiveness of alien invasive plants in China .Ecology and Environment Sciences, 21, 977-985. (in Chinese with English abstract )[黄乔乔, 沈奕德, 李晓霞, 程汉亭, 宋鑫, 范志伟 (2012). 外来入侵植物在中国的分布及入侵能力研究进展 . 生态环境学报, 21, 977-985.] [本文引用: 1]
[16]
HuangX, LiCJ, NanZB (2012).Competitive effects between Medicago sativa and Achnatherum inebrians .Acta Prataculturae Sinica, 21(1), 59-65. (in Chinese with English abstract)[黄玺, 李春杰, 南志标 (2012). 紫花苜蓿与醉马草的竞争效应 . 草业学报, 21(1), 59-65.] [本文引用: 2]
[17]
JuRT, LiH, ShihCJ, LiB (2012). Progress of biological invasions research in China over the last decade .Biodiversity Science, 20, 581-611. (in Chinese with English abstract)[鞠瑞亭, 李慧, 石正人, 李博 (2012). 近十年中国生物入侵研究进展 . 生物多样性, 20, 581-611.] [本文引用: 1]
[18]
KeddyP, NielsenK, WeiherE (2002). Relative competitive performance of 63 species of terrestrial herbaceous plants .Journal of Vegetation Science, 13, 5-16. [本文引用: 1]
[19]
LiGY, HouXW, DengX, WangZ, ZhangGH, LiQF (2010). The effect on allelopathy of Wedelia chinensis Merr. resulting from herbicide .Chinese Agricultural Science Bulletin, 26(1), 173-181. (in Chinese with English abstract)[李光义, 侯宪文, 邓晓, 王中, 张桂花, 李勤奋 (2010). 除草剂对蟛蜞菊化感作用的影响研究 . 中国农学通报, 26(1), 173-181.] [本文引用: 2]
[20]
LiuWJ, LiuY, HuangXQ, ZhouXQ, SongJ, YinQ, WangD, TaoL, ZhangFL, ChangLJ, ZhangL, LeiSR (2012). Impact of spraying glyphosate on growth and yield component of glyphosate-tolerant soybean at different growth stages .Scientia Agricultura Sinica, 45, 675-684. (in Chinese with English abstract)[刘文娟, 刘勇, 黄小琴, 周西全, 宋君, 尹全, 王东, 陶李, 张富丽, 常丽娟, 张蕾, 雷绍荣 (2012). 不同时期喷施草甘膦对抗草甘膦转基因大豆生长和产量构成的影响 . 中国农业科学, 45, 675-684.] [本文引用: 1]
[21]
LuY, SunGC, ZhuJY, FanMJ, MaoHP (2006). Comparison of control effects of glyphosate application in different time on Solidago canadensis L. Weed Science, (1), 51-52. (in Chinese with English abstract)[陆彦, 孙国才, 朱建亚, 范美娟, 毛慧萍 (2006). 草甘膦防除加拿大一枝黄花不同时间用药效果比较 . 杂草科学, (1), 51-52.] [本文引用: 2]
[22]
LydonJ, DukeSO (1988). Glyphosate induction of elevated levels of hydroxybenzoic acids in higher plants .Journal of Agricultural Food and Chemistry, 36, 813-818. [本文引用: 2]
[23]
MaestreFT, CallawayRM, FernandoV, LortieCJ (2009). Refining the stress-gradient hypothesis for competition and facilitation in plant communities .Journal of Ecology, 97, 199-205. [本文引用: 2]
[24]
MullanCC, KristyK, HalpernBS (2008). Interactive and cumulative effects of multiple human stressors in marine systems .Ecology Letters, 11, 1304-1315. [本文引用: 1]
[25]
PanXB, ZhangJQ, XuH, ZhangXL, ZhangW, SongHH, ZhuSF (2015). Spatial similarity in the distribution of invasive alien plants and animals in China .Natural Hazards, 77, 1751-1764. [本文引用: 1]
[26]
PengH, GuiFR, LiZY, LiJ, WanFH (2010). Competition effect of Imperata cylindrical to Ageratina adenophora. Chinese Journal of Ecology, 29, 1931-1936. (in Chinese with English abstract)[彭恒, 桂富荣, 李正跃, 李隽, 万方浩 (2010). 白茅对紫茎泽兰的竞争效应 . 生态学杂志, 29, 1931-1936.] [本文引用: 2]
[27]
PingXY, ZhouGS, SunJS (2010). Advances in the study of photosynthetic allocation and its controls .Chinese Journal of Plant Ecology, 34, 100-111. (in Chinese with English abstract)[平晓燕, 周广胜, 孙敬松 (2010). 植物光合产物分配及其影响因子研究进展 . 植物生态学报, 34, 100-111.] [本文引用: 1]
QiangS, ChenGJ, LiBP, MengL (2010). Invasive alien species in Chinese agricultural ecosystems and their management .Biodiversity Science, 18, 647-659. (in Chinese with English abstract)[强胜, 陈国奇, 李保平, 孟玲 (2010). 中国农业生态系统外来种入侵及其管理现状 . 生物多样性, 18, 647-659.] [本文引用: 1]
[30]
ThorpeAS, AschehougET, AtwaterDZ, CallawayRM (2011). Interactions among plants and evolution .Journal of Ecology, 99, 729-740. [本文引用: 1]
[31]
WangCY, XiaoHG, ZhaoLL, LiuJ, WangL, ZhangF, ShiYC, DuDL (2016). The allelopathic effects of invasive plant Solidago canadensis on seed germination and growth of Lactuca sativa enhanced by different types of acid deposition .Ecotoxicology, 25, 555-562. [本文引用: 1]
[32]
WangXJ, LangZH, ShanAS, HuangDF (2008). Advances in mechanism of herbicide in inhibiting amino acid biosynthesis and herbicide-tolerant transgenic plants .China Biotechnology, 28, 110-116. (in Chinese with English abstract)[王秀君, 郎志宏, 单安山, 黄大昉 (2008). 氨基酸生物合成抑制剂类除草剂作用机理及耐除草剂转基因植物研究进展 . 中国生物工程杂志, 28, 110-116.] [本文引用: 1]
[33]
WeidenhamerJR (2010). Direct and indirect effects of invasive plants on soil chemistry and ecosystem function .Journal of Chemical Ecology, 36, 59-69. [本文引用: 2]
[34]
WilliamsAC, McCarthyBC (2001). A new index of interspecific competition for replacement and additive designs .Ecological Research, 16, 29-40. [本文引用: 1]
[35]
XuZH, WangYP (2004). Disastrous mechanisms and control strategies of alien invasive plants .Chinese Journal of Ecology, 23, 124-127. (in Chinese with English abstract)[徐正浩, 王一平 (2004). 外来入侵植物成灾的机制及防除对策 . 生态学杂志, 23, 124-127.] [本文引用: 1]
[36]
YangLQ, LiaoFY, ZhaoK, XuQ, FengJS, WuHQ (2011). The effects of different herbicide on the growth of Solidago canadensis .Journal of Central South University of Forestry & Technology, 31(4), 109-113. (in Chinese with English abstract)[杨柳青, 廖飞勇, 赵坤, 许潜, 冯家生, 吴红强 (2011). 不同除草剂对加拿大一枝黄花生长的影响 . 中南林业科技大学学报, 31(4), 109-113.] [本文引用: 2]
[37]
YangRY, ZanST, TangJJ, ChenX (2011).Invasion mechanisms of Solidago canadensis L.: A review .Acta Ecologica Sinica, 31, 1185-1194. (in Chinese with English abstract)[杨如意, 昝树婷, 唐建军, 陈欣 (2011). 加拿大一枝黄花的入侵机理研究进展 . 生态学报, 31, 1185-1194.] [本文引用: 3]
[38]
ZhangRH (2010). Research on Screening of Replacement Plants for Flaveria bidentis and Competition Between Replacement Plants with F. bidentis. Master degree dissertation, Fujian Agriculture and Forestry University, Fuzhou. 12-37. (in Chinese with English abstract)[张瑞海 (2010). 黄顶菊替代植物的筛选及其与黄顶菊竞争效应的研究 . 硕士学位论文, 福建农业大学, 福州. 12-37.] [本文引用: 1]
[39]
ZhangSS, JinYL, TangJJ, ChenX (2009). The invasive plant Solidago canadensis L. suppresses local soil pathogens through allelopathy .Applied Soil Ecology, 41, 215-222. [本文引用: 1]
[40]
ZhangWP, JiaX, DamagaardC, BaiyY, PanS, WangGX (2013). The interplay between above- and below-ground plant-plant interactions along an environmental gradient: Insights from two-layer zone-of-influence models .Oikos, 122, 1147-1156. [本文引用: 1]
[41]
ZhouCF, LiY, ZhangXY, YuYC (2013). Research advance in eco-toxicity of glyphosate .Ecology and Environment Sciences, 22, 1737-1743. (in Chinese with English abstract)[周垂帆, 李莹, 张晓勇, 俞元春 (2013). 草甘膦毒性研究进展 . 生态环境学报, 22, 1737-1743.] [本文引用: 2]
[42]
ZhouJ, LiHL, LuoFL, HuangWJ, ZhangMX, YuFH (2015). Effects of nitrogen addition on interspecific competition between Alternanthera philoxeroides and Alternanthera sessilis .Acta Ecologica Sinica, 35, 8258-8267. (in Chinese with English abstract)[周建, 李红丽, 罗芳丽, 黄文军, 张明祥, 于飞海 (2015). 施氮对空心莲子草(Alternanthera philoxeroides)和莲子草(Alternanthera sessilis)种间关系的影响 . 生态学报, 35, 8258-8267.] [本文引用: 2]
[43]
ZhuQG, JinAW, WangYK, QiuYH, LiXT, ZhangSH (2013). Biomass allocation of branches and leaves in Phyllostachys heterocycla ‘Pubescens’ under different management modes: Allometric scaling analysis .Chinese Journal of Plant Ecology, 37, 811-819. (in Chinese with English abstract)[朱强根, 金爱武, 王意锟, 邱永华, 李雪涛, 张四海 (2013). 不同营林模式下毛竹枝叶的生物量分配: 异速生长分析 . 植物生态学报, 37, 811-819.] [本文引用: 1]
... 随着全球变化加剧以及我国对外经济的发展, 外来植物的入侵频率和危害程度正在迅速增加, 现已成为我国重大环境问题(鞠瑞亭等, 2012; Pan et al., 2015).入侵植物严重威胁了入侵地的生物多样性, 干扰了原生态系统的结构, 甚至引起生态系统服务功能退化, 造成我国每年超过70亿美元的经济损失(黄乔乔等, 2012).防治外来植物普遍使用化学防除、人工拔除等方法(强胜, 2009).有的****提议利用替代植被抵御外来植物入侵或者引进天敌以恢复稳定的生态平衡(徐正浩和王一平, 2004; Funk et al., 2008; 张瑞海, 2010), 这类生物防治法具有控制效果持久、对环境安全等优点, 但仍需要长期评估其生态风险性, 因此并未被大面积采用(强胜等, 2010); 由于人工拔除法存在成本过高的缺点, 目前我国防治外来植物主要依赖于经济、高效的化学防除法.自然条件下, 植物种内或种间普遍存在相互作用(Maestre et al., 2009), 正、负相互作用的动态平衡对植物群落结构和生态系统功能稳定有重要影响(Callaway, 2002; Brooker, 2008).当受到环境胁迫或者生物因素影响时, 植物邻体之间的相互作用将会发生变化(Maestre et al., 2009; He et al., 2013; Zhang et al., 2013).大量使用化学除草剂防除外来植物、抑制植物生长发育的同时, 必然直接或间接改变植物物种之间的相互联系(Brooker, 1998; Weidenhamer, 2010), 由此可能干扰植物群落的自然演替, 破坏当地生态系统的动态平衡. ...
中国农业生态系统外来种入侵及其管理现状 1 2010
... 随着全球变化加剧以及我国对外经济的发展, 外来植物的入侵频率和危害程度正在迅速增加, 现已成为我国重大环境问题(鞠瑞亭等, 2012; Pan et al., 2015).入侵植物严重威胁了入侵地的生物多样性, 干扰了原生态系统的结构, 甚至引起生态系统服务功能退化, 造成我国每年超过70亿美元的经济损失(黄乔乔等, 2012).防治外来植物普遍使用化学防除、人工拔除等方法(强胜, 2009).有的****提议利用替代植被抵御外来植物入侵或者引进天敌以恢复稳定的生态平衡(徐正浩和王一平, 2004; Funk et al., 2008; 张瑞海, 2010), 这类生物防治法具有控制效果持久、对环境安全等优点, 但仍需要长期评估其生态风险性, 因此并未被大面积采用(强胜等, 2010); 由于人工拔除法存在成本过高的缺点, 目前我国防治外来植物主要依赖于经济、高效的化学防除法.自然条件下, 植物种内或种间普遍存在相互作用(Maestre et al., 2009), 正、负相互作用的动态平衡对植物群落结构和生态系统功能稳定有重要影响(Callaway, 2002; Brooker, 2008).当受到环境胁迫或者生物因素影响时, 植物邻体之间的相互作用将会发生变化(Maestre et al., 2009; He et al., 2013; Zhang et al., 2013).大量使用化学除草剂防除外来植物、抑制植物生长发育的同时, 必然直接或间接改变植物物种之间的相互联系(Brooker, 1998; Weidenhamer, 2010), 由此可能干扰植物群落的自然演替, 破坏当地生态系统的动态平衡. ...
Interactions among plants and evolution 1 2011
... 植物之间最基本的相互作用是资源竞争和化感作用(Thorpe et al., 2011), 植物只要共享某种生存资源, 其种内个体或种间个体之间就会发生程度不一的竞争.加拿大一枝黄花获取和利用资源的能力要强于白茅, 在自然群落中, 白茅对加拿大一枝黄花的竞争作用小于加拿大一枝黄花种内个体间的相互竞争, 使得与白茅混种时, 加拿大一枝黄花植株积累了更多的有机物质, 而白茅积累了较少的物质.低浓度草甘膦影响了植物的种间竞争强度(图5), 混种加拿大一枝黄花和白茅地上、地下生物量积累出现高于单种植株的趋势, 表明种内抑制作用逐渐大于种间抑制.加拿大一枝黄花种内对地下有限资源的竞争更为激烈, 因而其根冠比低于混种加拿大一枝黄花的根冠比; 而白茅对草甘膦处理较敏感, 单种、混种时白茅整株均受到较大损伤.当受到环境胁迫时, 种间相互作用的类型和强度可能会随之发生改变(Mullan et al., 2008).草甘膦处理抑制加拿大一枝黄花、白茅茎叶和根正常的生长发育, 降低了两种植物地上部分竞争光照资源的能力, 以及地下部分从环境吸收养分的能力, 由此减弱了两种植物的种间竞争效应(图5).作为一种环境胁迫因子, 草甘膦进入植物体内将会影响其分泌的次生代谢物质, 从而改变植物的化感作用强度(Lydon & Duke, 1988; 李光义等, 2010; Weidenhamer, 2010).加拿大一枝黄花是广为人知的具有强化感作用的入侵植物(杨如意等, 2011; Wang et al., 2016), 在入侵过程中会向环境中释放化感物质以抑制邻近植物的生长发育, 还能促进有利于自身生长的根系微生物群落的增长(Zhang et al., 2009; 杨柳青等, 2011), 由此增加自身竞争能力.进入加拿大一枝黄花体内的草甘膦很可能影响其根系分泌的具有化感效力的次生代谢产物, 减弱了其对伴生植物白茅的化感抑制作用, 使得喷施草甘膦后混种白茅的生物量略比单种种 群高. ...
The allelopathic effects of invasive plant Solidago canadensis on seed germination and growth of Lactuca sativa enhanced by different types of acid deposition 1 2016
... 植物之间最基本的相互作用是资源竞争和化感作用(Thorpe et al., 2011), 植物只要共享某种生存资源, 其种内个体或种间个体之间就会发生程度不一的竞争.加拿大一枝黄花获取和利用资源的能力要强于白茅, 在自然群落中, 白茅对加拿大一枝黄花的竞争作用小于加拿大一枝黄花种内个体间的相互竞争, 使得与白茅混种时, 加拿大一枝黄花植株积累了更多的有机物质, 而白茅积累了较少的物质.低浓度草甘膦影响了植物的种间竞争强度(图5), 混种加拿大一枝黄花和白茅地上、地下生物量积累出现高于单种植株的趋势, 表明种内抑制作用逐渐大于种间抑制.加拿大一枝黄花种内对地下有限资源的竞争更为激烈, 因而其根冠比低于混种加拿大一枝黄花的根冠比; 而白茅对草甘膦处理较敏感, 单种、混种时白茅整株均受到较大损伤.当受到环境胁迫时, 种间相互作用的类型和强度可能会随之发生改变(Mullan et al., 2008).草甘膦处理抑制加拿大一枝黄花、白茅茎叶和根正常的生长发育, 降低了两种植物地上部分竞争光照资源的能力, 以及地下部分从环境吸收养分的能力, 由此减弱了两种植物的种间竞争效应(图5).作为一种环境胁迫因子, 草甘膦进入植物体内将会影响其分泌的次生代谢物质, 从而改变植物的化感作用强度(Lydon & Duke, 1988; 李光义等, 2010; Weidenhamer, 2010).加拿大一枝黄花是广为人知的具有强化感作用的入侵植物(杨如意等, 2011; Wang et al., 2016), 在入侵过程中会向环境中释放化感物质以抑制邻近植物的生长发育, 还能促进有利于自身生长的根系微生物群落的增长(Zhang et al., 2009; 杨柳青等, 2011), 由此增加自身竞争能力.进入加拿大一枝黄花体内的草甘膦很可能影响其根系分泌的具有化感效力的次生代谢产物, 减弱了其对伴生植物白茅的化感抑制作用, 使得喷施草甘膦后混种白茅的生物量略比单种种 群高. ...
... 植物之间最基本的相互作用是资源竞争和化感作用(Thorpe et al., 2011), 植物只要共享某种生存资源, 其种内个体或种间个体之间就会发生程度不一的竞争.加拿大一枝黄花获取和利用资源的能力要强于白茅, 在自然群落中, 白茅对加拿大一枝黄花的竞争作用小于加拿大一枝黄花种内个体间的相互竞争, 使得与白茅混种时, 加拿大一枝黄花植株积累了更多的有机物质, 而白茅积累了较少的物质.低浓度草甘膦影响了植物的种间竞争强度(图5), 混种加拿大一枝黄花和白茅地上、地下生物量积累出现高于单种植株的趋势, 表明种内抑制作用逐渐大于种间抑制.加拿大一枝黄花种内对地下有限资源的竞争更为激烈, 因而其根冠比低于混种加拿大一枝黄花的根冠比; 而白茅对草甘膦处理较敏感, 单种、混种时白茅整株均受到较大损伤.当受到环境胁迫时, 种间相互作用的类型和强度可能会随之发生改变(Mullan et al., 2008).草甘膦处理抑制加拿大一枝黄花、白茅茎叶和根正常的生长发育, 降低了两种植物地上部分竞争光照资源的能力, 以及地下部分从环境吸收养分的能力, 由此减弱了两种植物的种间竞争效应(图5).作为一种环境胁迫因子, 草甘膦进入植物体内将会影响其分泌的次生代谢物质, 从而改变植物的化感作用强度(Lydon & Duke, 1988; 李光义等, 2010; Weidenhamer, 2010).加拿大一枝黄花是广为人知的具有强化感作用的入侵植物(杨如意等, 2011; Wang et al., 2016), 在入侵过程中会向环境中释放化感物质以抑制邻近植物的生长发育, 还能促进有利于自身生长的根系微生物群落的增长(Zhang et al., 2009; 杨柳青等, 2011), 由此增加自身竞争能力.进入加拿大一枝黄花体内的草甘膦很可能影响其根系分泌的具有化感效力的次生代谢产物, 减弱了其对伴生植物白茅的化感抑制作用, 使得喷施草甘膦后混种白茅的生物量略比单种种 群高. ...
黄顶菊替代植物的筛选及其与黄顶菊竞争效应的研究 1 2010
... 随着全球变化加剧以及我国对外经济的发展, 外来植物的入侵频率和危害程度正在迅速增加, 现已成为我国重大环境问题(鞠瑞亭等, 2012; Pan et al., 2015).入侵植物严重威胁了入侵地的生物多样性, 干扰了原生态系统的结构, 甚至引起生态系统服务功能退化, 造成我国每年超过70亿美元的经济损失(黄乔乔等, 2012).防治外来植物普遍使用化学防除、人工拔除等方法(强胜, 2009).有的****提议利用替代植被抵御外来植物入侵或者引进天敌以恢复稳定的生态平衡(徐正浩和王一平, 2004; Funk et al., 2008; 张瑞海, 2010), 这类生物防治法具有控制效果持久、对环境安全等优点, 但仍需要长期评估其生态风险性, 因此并未被大面积采用(强胜等, 2010); 由于人工拔除法存在成本过高的缺点, 目前我国防治外来植物主要依赖于经济、高效的化学防除法.自然条件下, 植物种内或种间普遍存在相互作用(Maestre et al., 2009), 正、负相互作用的动态平衡对植物群落结构和生态系统功能稳定有重要影响(Callaway, 2002; Brooker, 2008).当受到环境胁迫或者生物因素影响时, 植物邻体之间的相互作用将会发生变化(Maestre et al., 2009; He et al., 2013; Zhang et al., 2013).大量使用化学除草剂防除外来植物、抑制植物生长发育的同时, 必然直接或间接改变植物物种之间的相互联系(Brooker, 1998; Weidenhamer, 2010), 由此可能干扰植物群落的自然演替, 破坏当地生态系统的动态平衡. ...
The invasive plant Solidago canadensis L. suppresses local soil pathogens through allelopathy 1 2009
... 植物之间最基本的相互作用是资源竞争和化感作用(Thorpe et al., 2011), 植物只要共享某种生存资源, 其种内个体或种间个体之间就会发生程度不一的竞争.加拿大一枝黄花获取和利用资源的能力要强于白茅, 在自然群落中, 白茅对加拿大一枝黄花的竞争作用小于加拿大一枝黄花种内个体间的相互竞争, 使得与白茅混种时, 加拿大一枝黄花植株积累了更多的有机物质, 而白茅积累了较少的物质.低浓度草甘膦影响了植物的种间竞争强度(图5), 混种加拿大一枝黄花和白茅地上、地下生物量积累出现高于单种植株的趋势, 表明种内抑制作用逐渐大于种间抑制.加拿大一枝黄花种内对地下有限资源的竞争更为激烈, 因而其根冠比低于混种加拿大一枝黄花的根冠比; 而白茅对草甘膦处理较敏感, 单种、混种时白茅整株均受到较大损伤.当受到环境胁迫时, 种间相互作用的类型和强度可能会随之发生改变(Mullan et al., 2008).草甘膦处理抑制加拿大一枝黄花、白茅茎叶和根正常的生长发育, 降低了两种植物地上部分竞争光照资源的能力, 以及地下部分从环境吸收养分的能力, 由此减弱了两种植物的种间竞争效应(图5).作为一种环境胁迫因子, 草甘膦进入植物体内将会影响其分泌的次生代谢物质, 从而改变植物的化感作用强度(Lydon & Duke, 1988; 李光义等, 2010; Weidenhamer, 2010).加拿大一枝黄花是广为人知的具有强化感作用的入侵植物(杨如意等, 2011; Wang et al., 2016), 在入侵过程中会向环境中释放化感物质以抑制邻近植物的生长发育, 还能促进有利于自身生长的根系微生物群落的增长(Zhang et al., 2009; 杨柳青等, 2011), 由此增加自身竞争能力.进入加拿大一枝黄花体内的草甘膦很可能影响其根系分泌的具有化感效力的次生代谢产物, 减弱了其对伴生植物白茅的化感抑制作用, 使得喷施草甘膦后混种白茅的生物量略比单种种 群高. ...
The interplay between above- and below-ground plant-plant interactions along an environmental gradient: Insights from two-layer zone-of-influence models 1 2013
... 随着全球变化加剧以及我国对外经济的发展, 外来植物的入侵频率和危害程度正在迅速增加, 现已成为我国重大环境问题(鞠瑞亭等, 2012; Pan et al., 2015).入侵植物严重威胁了入侵地的生物多样性, 干扰了原生态系统的结构, 甚至引起生态系统服务功能退化, 造成我国每年超过70亿美元的经济损失(黄乔乔等, 2012).防治外来植物普遍使用化学防除、人工拔除等方法(强胜, 2009).有的****提议利用替代植被抵御外来植物入侵或者引进天敌以恢复稳定的生态平衡(徐正浩和王一平, 2004; Funk et al., 2008; 张瑞海, 2010), 这类生物防治法具有控制效果持久、对环境安全等优点, 但仍需要长期评估其生态风险性, 因此并未被大面积采用(强胜等, 2010); 由于人工拔除法存在成本过高的缺点, 目前我国防治外来植物主要依赖于经济、高效的化学防除法.自然条件下, 植物种内或种间普遍存在相互作用(Maestre et al., 2009), 正、负相互作用的动态平衡对植物群落结构和生态系统功能稳定有重要影响(Callaway, 2002; Brooker, 2008).当受到环境胁迫或者生物因素影响时, 植物邻体之间的相互作用将会发生变化(Maestre et al., 2009; He et al., 2013; Zhang et al., 2013).大量使用化学除草剂防除外来植物、抑制植物生长发育的同时, 必然直接或间接改变植物物种之间的相互联系(Brooker, 1998; Weidenhamer, 2010), 由此可能干扰植物群落的自然演替, 破坏当地生态系统的动态平衡. ...
 , 邵学新
, 邵学新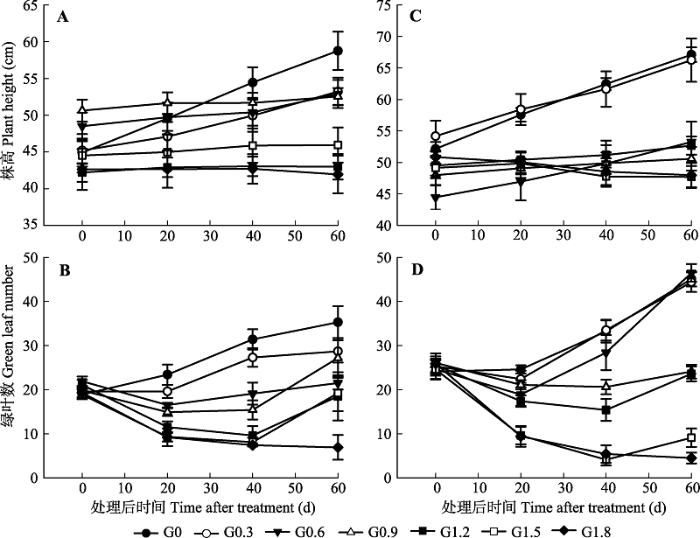 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT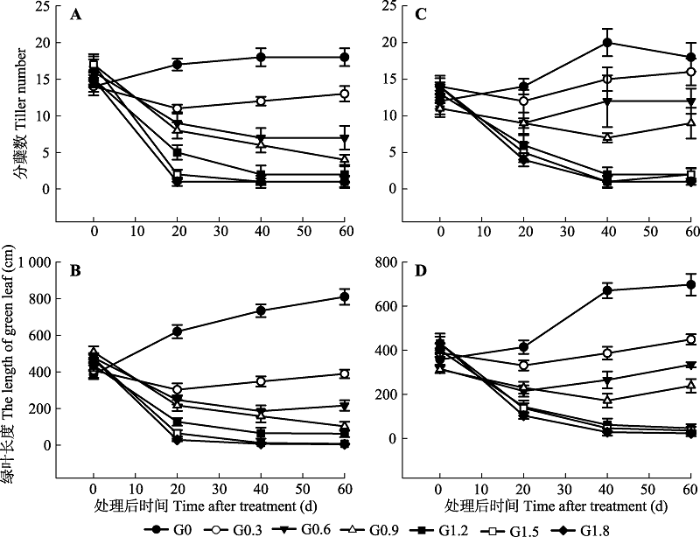 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT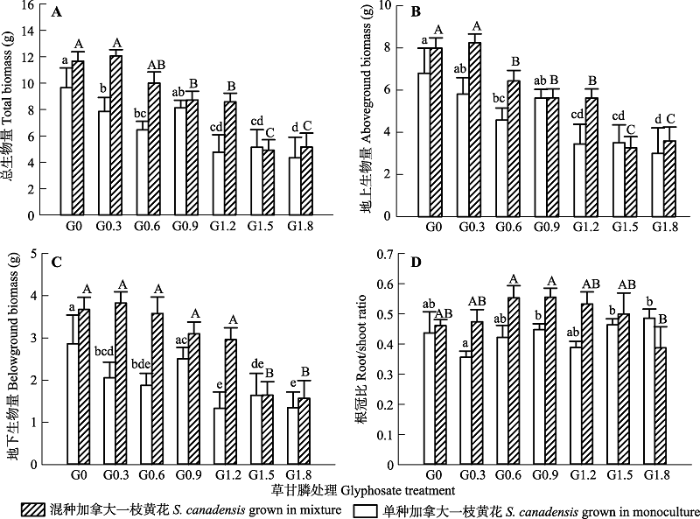 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT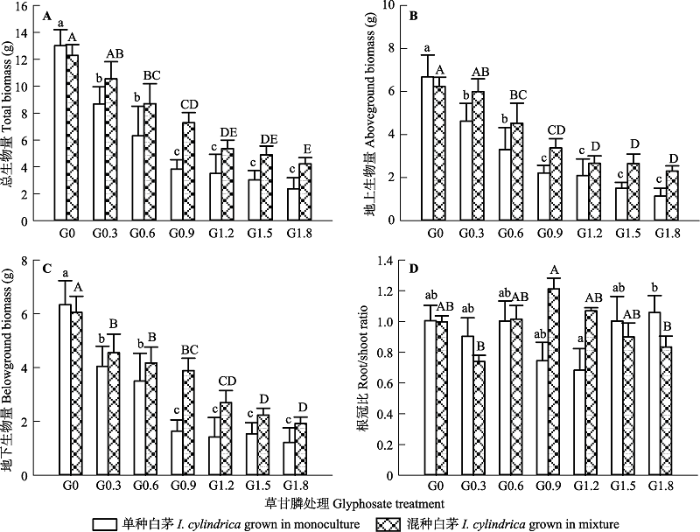 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT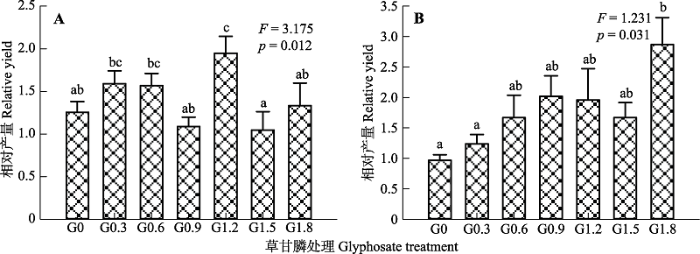 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}