Effects of microorganism on carbon, nitrogen and phosphorus of Dodonaea viscosa and the soils from different elevations in Yuanmou, Yunnan, China
WANGXue-Mei1,2, YANBang-Guo1,2,3, ZHAOGuang1,2, SHILiang-Tao3, LIUGang-Cai1, FANGHai-Dong3,*, 1Key Laboratory of Mountain Surface Processes and Ecological Regulation, Chinese Academy of Sciences, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences and Ministry of Water Conservancy, Chengdu 610041, China2University of Chinese Academy of Sciences, Beijing 100049, Chinaand 3Institute of Tropical Eco-Agriculture, Yunnan Academy of Agricultural Sciences, Yuanmou, Yunnan 651300, China 通讯作者:* 通信作者Author for correspondence (E-mail:rqsfhd@163.com) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41471232和31460127)
关键词:生态化学计量;土壤微生物;植物-土壤反馈;气候变化;车桑子 Abstract Aims Understanding the effects of soil microorganism at different elevations on plant C:N:P stoichiometry can help us to understand the plant-soil interactions in the context of climate change. Our aim was to quantify the independent and interactive effects of soil microbial communities and temperatures on the C, N, and P in the leaves of Dodonaea viscosa—a global widespread species. Methods Rhizosphere soils of D. viscosa were collected from two elevation zones in Yuanmou County, Yunnan Province. A 2 × 3 factorial experiment with six replications was conducted using climate chambers. The leaf C, N and P contents and the soil properties were measured after three months of the treatments. Important findings Compared with the autoclaved treatment, inoculated rhizosphere soils from both high and low elevations had higher nutrient absorption, especially P uptake. Temperature produced no significant effect on leaf C:N:P stoichiometry, but the interactive effect of temperature and microbial treatment appeared significant. For inoculated rhizosphere soils from high elevation, temperature had no significant effect on leaf C:N:P stoichiometry. For inoculated rhizosphere soils from low elevation, leaf N and P contents under low temperature were significantly lower than those with warmer soils. The promoting effect of soil microorganisms on nutrient uptake may be due to the direct effect of beneficial microorganisms (e.g., mycorrhizal fungi), but not through the alteration of nutrient cycling process. Because D. viscosa in the inoculated rhizosphere soils absorbed more N and P from the soil than those in autoclaved soil, the available N and P in inoculated rhizosphere soils were lower than those in autoclaved soils. As predicted future temperature will be lower in the studied region, the growth of D. viscosa may be negatively affected through plant-microbe feedbacks.
Keywords:ecological stoichiometry;soil microorganisms;plant-soil feedback;climate change;Dodonaea viscosa -->0 PDF (4853KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 王雪梅, 闫帮国, 赵广, 史亮涛, 刘刚才, 方海东. 云南元谋不同海拔土壤微生物对车桑子碳、氮、磷化学计量特征及土壤特性的影响. 植物生态学报, 2017, 41(3): 311-324 https://doi.org/10.17521/cjpe.2016.0267 WANGXue-Mei, YANBang-Guo, ZHAOGuang, SHILiang-Tao, LIUGang-Cai, FANGHai-Dong. Effects of microorganism on carbon, nitrogen and phosphorus of Dodonaea viscosa and the soils from different elevations in Yuanmou, Yunnan, China. Chinese Journal of Plant Ecology, 2017, 41(3): 311-324 https://doi.org/10.17521/cjpe.2016.0267 植物生长需要多种生命元素, 碳(C)、氮(N)、磷(P)是构成有机体的主要元素, 其含量及比例特征是生态化学计量学研究的主要内容。C是组成生物结构的最基本元素, 构成了约50%的植物干质量, 而N和P是多数植物生长过程中主要的限制性元素(Ågren, 2008; Vitousek et al., 2010)。植物C、N、P化学计量特征调节着植物生长(Fan et al., 2015), 其中, C:N和C:P在一定程度上反映了植物对N、P营养的利用效率, C:P和N:P可反映植物器官的生长速率, 并且N:P还是判断植物养分限制的重要指标(Elser et al., 2000; Tessier & Raynal, 2003; Güsewell, 2004; Zhang et al., 2015)。 植物的生态化学计量动态受到自身和外界环境因素的共同控制。一方面, 其生态化学计量特征具有内稳性(曾德慧和陈广生, 2005; 牛得草等, 2011)和物种特异性(石贤萌等, 2015); 另一方面, 植物的生态化学计量特征又随环境条件(如纬度、温度、降水、土壤肥力、群落演替等)的改变而发生改变(Reich & Oleksyn, 2004; Han et al., 2005; Sardans et al., 2012; 王振南和杨惠敏, 2013)。大多数研究表明, 植物N、P养分特征与土壤养分具有很强的相关性, 其化学计量特征受土壤肥力的影响很大(阎凯等, 2011; 宾振钧等, 2015; Fan et al., 2015)。但这些研究基本上都是基于野外采样的数据, 植物养分是在长时间不同条件下形成的, 短期的控制试验能否造成物种水平上化学计量特征的改变还不得而知。但室内条件下化学计量特征对环境变化的响应更有利于研究单个物种的化学计量规律, 为野外研究不同物种、群落或生态系统的化学计量特征提供参考(严正兵等, 2013; 刘玉芳等, 2015)。 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011)。一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015)。土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011)。土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面。由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道。 车桑子(Dodonaea viscosa)是无患子科分布最广的一种植物, 除南极洲以外的所有大陆都能找到它的踪迹(Harrington & Gadek, 2009)。在我国, 车桑子主要分布于南方, 是干热河谷区最为典型的灌木。为了明确不同气候背景下土壤微生物群落对植物C、N、P化学计量特征可能造成的影响, 我们以车桑子为研究对象, 从两个不同海拔(分别属于南亚热带和北亚热带)的车桑子生长区采集根际土壤微生物, 并设定相应的两个温度梯度, 通过双因素交互试验, 研究了温度、土壤微生物及二者的交互作用对车桑子C、N、P计量特征和土壤特性的影响。我们假设: (1)接种微生物有利于土壤养分转化、增加土壤有效养分的含量, 从而提高车桑子的C、N、P含量, 并改变其C、N、P生态化学计量特征; (2)车桑子的N、P养分特征和土壤养分特征有一定的正相关关系, 但由于微生物对不同气候条件的长期适应, 温度会改变不同海拔区土壤微生物对车桑子和土壤的影响, 不同温度条件下车桑子和土壤的关系有所不同。我们希望通过本研究明确不同海拔地区土壤微生物在温度变化条件下对车桑子C、N、P化学计量特征的影响, 并揭示车桑子N、P养分特征与土壤特性的关系, 从而为未来气候变化下植物与土壤的关系提供理论参考。
1 材料和方法
1.1 研究区概况
元谋县位于云贵高原北缘的金沙江一级支流——龙川江下游的河谷地带(101.58°-102.08° E, 25.42°-26.12° N), 海拔898-2 836 m, 该地区相对高差约2 000 m, 生态环境垂直分异明显。从河谷到山顶依次为河谷南亚热带、山地中亚热带、山地北亚热带及山地温带-中温带4个垂直气候带(李坤, 1993)。通过野外考察, 选择元谋县高海拔区凉山乡和低海拔区苴林乡两个地点进行土壤样品采集。高海拔区的地理位置为101.94° E, 25.74° N, 海拔1 835 m, 年平均气温为14.6-15.9 ℃, 年降水量为 829 mm, 无霜期305-314天, 属北亚热带气候类型, 土壤类型为紫色土, 主要植被有车桑子、栎树(Quercus)、杜鹃(Rhododendron simsii)等乔灌植物; 低海拔区的地理位置为101.83° E, 25.85° N, 海拔 1 124 m, 年平均气温为21.9 ℃, 年降水量为621 mm, 无霜期363天, 属南亚热带气候类型(李坤, 1993), 土壤类型为燥红土, 主要植被为车桑子和黄茅(Heteropogon contortus)。
1.2 土壤采集与处理
参照Bell等(2014)的文献, 轻轻抖动植株后依然保留在根部的土壤即为根际土。分别在低海拔区苴林乡和高海拔区凉山乡采用抖土法采集车桑子根际土壤样品, 每个地点挖取5-10株车桑子植株, 并将同一地点的根际土混合为一个样品。将两地根际土分别过筛(2 mm)、混匀, 用冰盒运回实验室后保存于 4 ℃的冰箱中, 用作接种微生物用土, 测定其基本理化性质, 并用磷脂脂肪酸法初步测定两种土壤微生物群落结构的差异。两种根际土样的磷脂脂肪酸含量和理化性质分别见表1和表2。高海拔区和低海拔区根际土样磷脂脂肪酸种类分别为32种和21种, 总含量分别为11.995和7.667 nmol·g-1 dry soil。 此外, 采用多点混合法分别在上述高海拔紫色土区和低海拔燥红土区采集车桑子植株附近的表层土壤。将两地土样运回实验室过筛后, 用高压灭菌锅灭菌, 具体操作方法如下: 121 ℃, 103.5 kPa条件下, 用高压灭菌锅湿热灭菌1 h, 反复灭菌3次, 灭菌时间间隔为24 h, 以完全消除土壤微生物孢子。高压灭菌是土壤灭菌的有效方法, 并且对土壤理化性质的影响较小(Alphei & Scheu, 1993; Berns et al., 2008)。为了使所有处理的土壤理化性质具有一致性, 取等体积的灭菌紫色土和灭菌燥红土, 装入灭菌的盆钵中(121.1 ℃, 20 min), 用作盆栽基土, 每盆(用量筒)装基土1 400 mL (700 mL灭菌燥红土+ 700 mL灭菌紫色土), 充分混匀后, 取样测定基土的土壤理化性质(表2)。从表2来看, 接种的根际土P含量高于基土, 这可能是根际效应的缘故。本文所用基土为低海拔和高海拔车桑子生长区土壤的混合土, 能够代表车桑子生长环境的土壤性质。 Table 1 表1 表1高海拔区和低海拔区车桑子根际土样的磷脂脂肪酸含量(nmol·g-1) Table 1Abundant of main microbial communities varieties phospholipid fatty acid (PLFA) in the rhizosphere soils of Dodonaea viscosa at two elevation zones (nmol·g-1)
序号 No.
微生物标记 Sign of microorganism
高海拔区 High elevation
低海拔区 Low elevation
序号 No.
微生物标记 Sign of microorganism
高海拔区 High elevation
低海拔区 Low elevation
1
13:0
0
0.144 3
20
16:0 iso
0.725 0
0.601 9
2
14:0
0.176 6
0.193 1
21
16:1 2OH
0.237 6
0.247 6
3
16:0
1.990 0
1.078 4
22
16:1 iso G
0.079 4
0
4
17:0
0.133 7
0
23
16:1 w5c
0.349 2
0.099 7
5
18:0
0.506 1
0.276 7
24
16:1 w9c
0.074 1
0
6
19:0
1.250 0
1.250 0
25
17:0 10-methyl
0.235 5
0.249 9
7
20:0
0.065 0
0
26
17:0 anteiso
0.353 3
0.281 5
8
10:0 2OH
0.098 0
0
27
17:0 cyclo
0.299 3
0.127 2
9
11:0 iso
0.101 3
0
28
17:0 iso
0.299 1
0.230 2
10
11:0 iso 3OH
0
0.508 3
29
17:1 w8c
0.141 5
0
11
12:0 iso 3OH
0
0.148 5
30
17:1 w9c
0
0.115 3
12
13:0 iso
0.139 4
0
31
18:0 10-methyl, TBSA
0.470 5
0.302 5
13
14:0 iso
0.038 8
0
32
18:0 iso
0
0.118 3
14
14:1 w5c
0.085 5
0
33
18:1 w5c
0.289 0
0
15
15:0 3OH
0.114 8
0
34
18:1 w7c 11-methyl
0.143 1
0
16
15:0 anteiso
0.482 6
0.250 1
35
18:1 w9c
1.157 0
0.648 0
17
15:0 iso
1.107 4
0.531 1
36
18:3 w6c (6,9,12)
0.096 0
0
18
15:1 iso G
0.060 3
0
37
19:0 cyclo w8c
0.636 1
0.264 8
19
16:0 2OH
0.060 2
0
14:0 iso, 15:0 iso, 15:0 anteiso, 16:0 iso, 17:0 iso, 17:0 anteiso and 18:0 iso in the table represent Gram-positive bacteria; 16:1w9c and 17:0 cyclo represent Gram-negative bacteria; 18:0 indicates hydrogenobacter; 16:1 w5c indicates arbuscular mycorrhizal fungi (Qu et al., 2015).表中14:0 iso, 15:0 iso, 15:0 anteiso, 16:0 iso, 17:0 iso, 17:0 anteiso和18:0 iso代表革兰氏阳性菌; 16:1w9c和17:0 cyclo代表革兰氏阴性菌; 18:0表示嗜热解氢杆菌; 16:1 w5c表示丛枝菌根真菌(Qu et al., 2015)。 新窗口打开 Table 2 表2 表2高海拔区和低海拔区车桑子根际土以及盆栽基土的基本理化性质 Table 2Physical and chemical properties of the two inoculated rhizosphere soils and the potting medium of Dodonaea viscosa from high elevation and low elevation
2.1.1 不同处理下车桑子的C、N、P化学计量特征 土壤微生物对车桑子的C、N、P含量及其化学计量比有显著影响。除有机碳外, 温度对车桑子的N、P含量及其化学计量比均无显著影响, 但温度与微生物的交互作用对C、N、P含量及化学计量比有显著或极显著影响(表3)。 Table 3 表3 表3温度和土壤微生物处理对车桑子叶片碳(C)、氮(N)、磷(P)含量及化学计量比的双因素方差分析 Table 3Statistical results of a two-way analysis of variance (ANOVA) with temperature and soil microbial treatments on carbon (C), nitrogen (N), phosphorus (P) contents
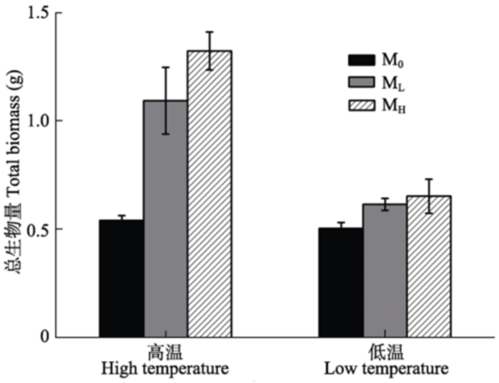
M0, autoclaved treatment; ML, inoculated rhizosphere soil from low elevation; MH, inoculated rhizosphere soil from high elevation. Bold values indicate the significant differences in the microbial treatment compared with the other two treatments under the same temperature (p < 0.05, LSD test).M0, 土壤灭菌处理; ML, 接种低海拔根际土处理; MH, 接种高海拔根际土处理。加粗数值表示同一温度条件下, 该微生物处理与其他两个微生物处理差异显著(p < 0.05, LSD检验)。 新窗口打开 2.1.2 不同处理下车桑子的生物量特征 双因素方差分析表明, 微生物(p < 0.001)、温度(p < 0.001)、温度×微生物(p < 0.01)对车桑子的生物 量都有显著影响。高温条件下, ML和MH处理的车桑子生物量显著高于M0处理, ML和MH处理的车桑子生物量分别比M0处理高出1.02倍和1.45倍, 但ML和MH处理的生物量没有显著差异(p > 0.05, LSD检验)。而在低温条件下, 各微生物处理间车桑子生物量均没有显著差异(图1)。 显示原图|下载原图ZIP|生成PPT 图1不同温度和微生物处理下车桑子总生物量(平均值±标准误差, n = 6)。M0, 土壤灭菌处理; ML, 接种低海拔根际土处理; MH, 接种高海拔根际土处理。 -->Fig. 1Total biomass of Dodonaea viscosa under different treatments (mean ± SE, n = 6). M0, autoclaved treatment; ML, inoculated rhizosphere soil from low elevation; MH, inoculated rhizosphere soil from high elevation. -->
2.2 不同处理下的土壤特性
2.2.1 土壤pH值 温度对土壤pH值无显著影响(p > 0.05), 微生物(p < 0.001)及温度与微生物的交互作用对土壤pH值影响显著(p < 0.05) (表5)。在两个温度条件下, 不同微生物处理的土壤pH值均为MH < ML < M0 (p < 0.001), 但无论何种微生物条件, 温度对土壤pH值均无显著影响(p > 0.05) (图2)。 Table 5 表5 表5温度和土壤微生物接种处理对土壤特性双因素方差分析 Table 5Statistical results of a two-way analysis of variance (ANOVA) with temperature and soil microbial treatments on soil properties
变量 Variables
微生物 Microorganism
温度 Temperature
微生物×温度 Microorganism × temperature
F
p
F
p
F
p
pH值 pH value
103.493
<0.001
0.688
0.413
4.071
0.027
微生物生物量C Microbial biomass C
68.654
<0.001
2.326
0.138
15.937
<0.001
微生物生物量N Microbial biomass N
449.038
<0.001
0.323
0.574
31.891
<0.001
铵态氮 Ammonium N
213.066
<0.001
29.048
<0.001
30.128
<0.001
硝态氮 Nitrate N
106.857
<0.001
0.182
0.673
6.240
0.005
有效磷 Available P
170.607
<0.001
5.110
0.031
57.256
0.001
有效钾 Available K
26.448
<0.001
68.391
<0.001
6.154
0.006
新窗口打开 显示原图|下载原图ZIP|生成PPT 图2不同温度和微生物处理下土壤pH值特征(平均值±标准误差, n = 6)。M0, 土壤灭菌处理; ML, 接种低海拔根际土处理; MH, 接种高海拔根际土处理。 -->Fig. 2Changes in soil pH value in different treatments (mean ± SE, n = 6). M0, autoclaved treatment; ML, inoculated rhizosphere soil from low elevation; MH, inoculated rhizosphere soil from high elevation. -->
2.2.2 土壤微生物生物量C、N 双因素方差分析表明, 温度对土壤微生物生物量C、N无显著影响(p > 0.05), 微生物及温度与微生物的交互作用对土壤微生物生物量C、N作用显著(表5)。除高温条件下MH处理的土壤微生物生物量C与M0处理的土壤微生物生物量C没有显著差异外, 其余条件下, 接种微生物处理(ML和MH)的土壤微生物生物量C、N都极显著低于灭菌处理(M0) (p < 0.001), 不论何种微生物条件, 温度对土壤微生物生物量C、N均没有显著影响(p > 0.05) (图3A、3B)。 显示原图|下载原图ZIP|生成PPT 图3不同温度和微生物处理下土壤微生物生物量(平均值±标准误差, n = 6)。A, 微生物生物量碳。B, 微生物生物量氮。M0, 土壤灭菌处理; ML, 接种低海拔根际土处理; MH, 接种高海拔根际土处理。 -->Fig. 3Characteristics of soil microbial biomass by treatments (mean ± SE, n = 6). A, Microbial biomass carbon. B, Microbial biomass nitrogen. M0, autoclaved treatment; ML, inoculated rhizosphere soil from low elevation; MH, inoculated rhizosphere soil from high elevation. -->
2.2.3 土壤有效养分特征 不同处理下, 土壤有效氮与有效磷的变化规律相似, 即在两种温度处理下, 接种微生物均极显著降低了土壤铵态N、硝态N和有效P的含量(p < 0.001) (图4A-4C), 但极显著增加了土壤有效K的含量(p < 0.001), 尤以高温条件增加效果明显(图4D)。在各 种微生物条件下, 温度对土壤养分指标基本无显著影响。 显示原图|下载原图ZIP|生成PPT 图4不同温度和微生物处理下土壤有效养分含量(平均值±标准误差, n = 6)。A, 铵态氮。B, 硝态氮。C, 有效磷。D, 有效钾。M0, 土壤灭菌处理; ML, 接种低海拔根际土处理; MH, 接种高海拔根际土处理。 -->Fig. 4Amount of available nutrient in soils under different treatments (mean ± SE, n = 6). A, Ammonium nitrogen. B, Nitrate nitrogen. C, Available phosphorus. D, Available potassium. M0, autoclaved treatment; ML, inoculated rhizosphere soil from low elevation; MH, inoculated rhizosphere soil from high elevation. -->
2.3 车桑子叶片N、P与土壤特性的相关分析
不考虑温度因素, 车桑子N、P含量和土壤有效N、P、K养分之间均无显著相关性(p > 0.05)。但叶片N和土壤pH值有一定负相关关系(r = -0.32, p = 0.057), 叶片P与土壤pH值也显著负相关(r = -0.406, p =0.014), 并且叶片P和土壤K有一定的正相关关系 (r = 0.317, p = 0.059)。 但是, 我们发现温度会改变这种相关性。高温条件下, 叶片P和土壤pH值、微生物生物量C、微生物生物量N、铵态N、硝态N、有效P呈负相关关系, 而与土壤有效K呈极显著正相关关系; 叶片N与土壤各指标均无显著相关关系。而在低温条件下, 叶片N与土壤pH值显著负相关, 与土壤养分无显著相关性; 叶片P与土壤各指标均无显著相关关系(表6)。 Table 6 表6 表6不同温度条件下车桑子叶片氮(N)、磷(P)与土壤特性之间的相关系数(n = 18) Table 6Correlation coefficients between leaf nitrogen (N), phosphorus (P) concentration and soil properties under two temperatures (n = 18)
ÅgrenGI (2008). Stoichiometry and nutrition of plant growth in natural communities. Annual Review of Ecology Evolution and Systematics, 39, 153-170. [本文引用: 1]
[2]
AlpheiJ, ScheuS (1993). Effects of biocidal treatments on biological and nutritional properties of a mull-structured woodland soil. Geoderma, 56, 435-448. [本文引用: 1]
BardgettRD, ManningP, MorriënE, VriesFTD (2013). Hierarchical responses of plant-soil interactions to climate change: Consequences for the global carbon cycle. Journal of Ecology, 101, 334-343. [本文引用: 1]
[5]
BardgettRD, StreeterTC, ColeL, HartleyIR (2002). Linkages between soil biota, nitrogen availability, and plant nitrogen uptake in a mountain ecosystem in the Scottish Highlands. Applied Soil Ecology, 19, 121-134. [本文引用: 1]
[6]
BellC, CarrilloY, BootCM, RoccaJD, PendallE, WallensteinMD (2014). Rhizosphere stoichiometry: Are C: N: P ratios of plants, soils, and enzymes conserved at the plant species-level? New Phytologist, 201, 505-517. [本文引用: 1]
[7]
BernsAE, PhilippH, NarresHD, BurauelP, VereeckenH, TappeW (2008). Effect of gamma-sterilization and autoclaving on soil organic matter structure as studied by solid state NMR, UV and fluorescence spectroscopy. European Journal of Soil Science, 59, 540-550. [本文引用: 1]
[8]
BiYL, ChenSL, KongWP, FengYB (2014). Effects of microorganism inoculation on growth of soybean and its rhizosphere soil. Ecological Science, 33, 121-126. (in Chinese with English abstract)[毕银丽, 陈书琳, 孔维平, 冯颜博 (2014). 接种微生物对大豆生长及其根际土壤的影响 . 生态科学, 33, 121-126.]
[9]
BinZJ, ZhangRY, ZhangWP, XuDH (2015). Effects of nitrogen, phosphorus and silicon addition on leaf carbon, nitrogen, and phosphorus concentration of Elymus nutans of alpine meadow on Qinghai-Tibetan Plateau, China. Acta Ecologica Sinica, 35, 4699-4706. (in Chinese with English abstract)[宾振钧, 张仁懿, 张文鹏, 徐当会 (2015). 氮磷硅添加对青藏高原高寒草甸垂穗披碱草叶片碳氮磷的影响 . 生态学报, 35, 4699-4706.]
[10]
BowmanWD, DammM (2015). Alpine landscape variation in foliar nitrogen and phosphorus concentrations and the relation to soil nitrogen and phosphorus availability. Arctic, Antarctic, and Alpine Research, 35, 144-149.
BrownME (2003). Seed and root bacterization. Annual Review of Phytopathology, 12, 181-197. [本文引用: 1]
[13]
ChapmanSK, LangleyJA, HartSC, KochGW (2006). Plants actively control nitrogen cycling: Uncorking the microbial bottleneck. New Phytologist, 169, 27-34. [本文引用: 1]
[14]
DefossezE, CourbaudB, MarcaisB, ThuillerW, GrandaE, KunstlerG (2011). Do interactions between plant and soil biota change with elevation? A study on Fagus sylvatica. Biology Letters, 7, 699-701. [本文引用: 1]
[15]
DingWR, LüXX, MingQZ (2011). Hydrothermal analysis of environment of Yuanmou dry-hot river valley in Jinsha River, upper Yangtze River, China. China Rural Water and Hydropower, (2), 39-42. (in Chinese with English abstract)[丁文荣, 吕喜玺, 明庆忠 (2011). 变化环境下的元谋干热河谷区水热响应分析 . 中国农村水利水电, (2), 39-42.]
FanH, WuJ, LiuW, YuanY, HuL, CaiQ (2015). Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil, 392, 127-138. [本文引用: 1]
GüsewellS (2004). N:P ratios in terrestrial plants: Variation and functional significance. New Phytologist, 164, 243-266. [本文引用: 2]
[21]
HanW, FangJ, GuoD, ZhangY (2005). Leaf N and P stoichiometry across 753 terrestrial plant species in China. New Phytologist, 168, 377-385. [本文引用: 3]
[22]
HarringtonMG, GadekPA (2009). A species well travelled — The Dodonaea viscosa (Sapindaceae) complex based on phylogenetic analyses of nuclear ribosomal ITS and ETSf sequences. Journal of Biogeography, 36, 2313-2323. [本文引用: 1]
[23]
HuangJY, SongCC, SongYY, LiuDY, WanZM, LiaoYJ (2008). Influence of freshwater marsh tillage on microbial biomass and dissolved organic carbon and nitrogen. Environmental Science, 29, 1380-1387. (in Chinese with English abstract)[黄靖宇, 宋长春, 宋艳宇, 刘德燕, 万忠梅, 廖玉静 (2008). 湿地垦殖对土壤微生物量及土壤溶解有机碳、氮的影响 . 环境科学, 29, 1380-1387.] [本文引用: 1]
[24]
JaegerCH, MonsonRK, FiskMC, SchmidtSK (1999). Seasonal partitioning of nitrogen by plants and soil microorganisms in an alpine ecosystem. Ecology, 80, 1883-1891. [本文引用: 1]
[25]
JiangJ, SongMH (2010). Review of the roles of plants and soil microorganisms in regulating ecosystem nutrient cycling. Chinese Journal of Plant Ecology, 34, 979-988. (in Chinese with English abstract)[蒋婧, 宋明华 (2010). 植物与土壤微生物在调控生态系统养分循环中的作用 . 植物生态学报, 34, 979-988.] [本文引用: 2]
[26]
JonassonS, MichelsenA, SchmidtIK, NielsenEV (1999). Responses in microbes and plants to changed temperature, nutrient, and light regimes in the Arctic. Ecology, 80, 1828-1843.
[27]
LauJA, LennonJT (2011). Evolutionary ecology of plant-microbe interactions: Soil microbial structure alters selection on plant traits. New Phytologist, 192, 215-224.
[28]
LauJA, LennonJT (2012). Rapid responses of soil microorganisms improve plant fitness in novel environments. Proceedings of the National Academy of Sciences of the United States of America, 109, 14058-14062. [本文引用: 1]
[29]
LenoirJ, GégoutJC, MarquetPA, de RuffrayP, BrisseH (2008). A significant upward shift in plant species optimum elevation during the 20th century. Science, 32, 1768-1771.
[30]
LiJP, LiT, ZhaoZW (2003). Diversity of arbuscular mycorrhizal fungi in the hot-dry valley of Jinsha River. Mycosystema, 22, 604-612. (in Chinese with English abstract)[李建平, 李涛, 赵之伟 (2003). 金沙江干热河谷 (元谋段)丛枝菌根真菌多样性研究 . 菌物学报, 22, 604-612.] [本文引用: 1]
[31]
LiK (1993). Yuanmou County Annals. The Peoples Press of Yunnan, Kunming.(in Chinese) [李坤 (1993). 元谋县志. 云南人民出版社, 昆明.] [本文引用: 3]
LiuYF, ChenSL, LiYC, ChenS, GuoZW, YangQP (2015). Effect of long-term flooded conditions on nutrient stoi¬chiometric characteristics of Phyllostachys rivalis rhizome roots. Acta Botanica Boreali-Occidentalia Sinica, 35, 350-355. (in Chinese with English abstract)[刘玉芳, 陈双林, 李迎春, 陈珊, 郭子武, 杨清平 (2015). 长期淹水对河竹鞭根养分化学计量特征的影响 . 西北植物学报, 35, 350-355.]
[34]
LuRK (2000). Analysis Methods of Soil Agricultural Chemistry. China Agricultural Science and Technology Press, Beijing, (in Chinese)[鲁如坤 (2000). 土壤农业化学分析方法. 中国农业科技出版社, 北京.] [本文引用: 1]
[35]
MiransariM (2011). Soil microbes and plant fertilization. Applied Microbiology and Biotechnology, 92, 875-885.
[36]
NiuDC, DongXY, FuH (2011). Seasonal dynamics of carbon, nitrogen and phosphorus stoichiometry in Stipa bungeana. Pratacultural Science, 28, 915-920. (in Chinese with English abstract)[牛得草, 董晓玉, 傅华 (2011). 长芒草不同季节碳氮磷生态化学计量特征 . 草业科学, 28, 915-920.] [本文引用: 1]
[37]
OlanderL, VitousekP (2000). Regulation of soil phosphatase and chitinase activity by N and P availability. Biog¬eo¬chemistry, 49, 175-191.
[38]
PengSL (2014). Responses of Soil Microbial Communities to Water and Temperature Changes in Yuanmou Dry-Hot Valley. PhD dissertation, University of Chinese Academy of Sciences, Beijing. 111-113. (in Chinese with English abstract)[彭思利 (2014). 元谋干热河谷土壤微生物群落对水热变化的响应 . 博士学位论文, 中国科学院大学, 北京. 111-113.] [本文引用: 1]
[39]
Pernilla BrinkmanE, van der PuttenWH, BakkerEJ, VerhoevenKJ (2010). Plant-soil feedback: Experimental approaches, statistical analyses and ecological interpretations. Journal of Ecology, 98, 1063-1073.
[40]
QiSH, WangJB (2007). The primary analysis of climate change in dry-hot valley of Yuanmou, China. Journal of Meteorological Research and Application, 28, 125-127. (in Chinese with English abstract)[起树华, 王建彬 (2007). 元谋干热河谷气候生态环境变化的初步分析 . 气象研究与应用, 28, 125-127.] [本文引用: 1]
[41]
QuL, HuangY, MaK, ZhangY, BierecA (2015). Effects of plant cover on properties of rhizosphere and inter-plant soil in a semiarid valley, SW China. Soil Biology & Biochemistry, 94, 1-9.
[42]
ReichPB, OleksynJ (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006.
[43]
SardansJ, PeñuelasJ, EstiarteM (2008). Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Applied Soil Ecology, 39, 223-235. [本文引用: 1]
[44]
SardansJ, Rivas-UbachA, PeñuelasJ (2012). Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). Forest Ecology and Management, 262, 2024-2034.
[45]
ShawLJ, BeatonbY, GloverLA, KillhamK, MehargAA (1999). Re-inoculation of autoclaved soil as a non-sterile treatment for xenobiotic sorption and biodegradation studies. Applied Soil Ecology, 11, 217-226. [本文引用: 1]
[46]
ShenRF, ZhaoXQ (2015). Role of soil microbes in the acquisition of nutrients by plants. Acta Ecologica Sinica, 35, 6584-6591. (in Chinese with English abstract)[沈仁芳, 赵学强 (2015). 土壤微生物在植物获得养分中的作用 . 生态学报, 35, 6584-6591.] [本文引用: 1]
[47]
ShiXM, QiJH, SongL, LiuWY, HuangJB, LiS, LuHZ, ChenX (2015). C, N and P stoichiometry of two dominant seedlings and their responses to nitrogen additions in the montane moist evergreen broad-leaved forest in Ailao Mountains, Yunnan. Chinese Journal of Plant Ecology, 39, 962-970. (in Chinese with English abstract)[石贤萌, 杞金华, 宋亮, 刘文耀, 黄俊彪, 李苏, 卢华正, 陈曦 (2015). 哀牢山中山湿性常绿阔叶林两种优势幼苗C、N、P化学计量特征及其对N沉降增加的响应 . 植物生态学报, 39, 962-970.]
[48]
SongYT, ZhouDW, LiQ, WangP, HuangYX (2012). Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in Northeast China. Chinese Journal of Plant Ecology, 36, 222-230. (in Chinese with English abstract)[宋彦涛, 周道玮, 李强, 王平, 黄迎新 (2012). 松嫩草地80种草本植物叶片氮磷化学计量特征 . 植物生态学报, 36, 222-230.] [本文引用: 1]
[49]
SteinauerK, TilmanD, WraggPD, CesarzS, CowlesJM, PritschK, ReichPB, WeisserWW, EisenhauerN (2015). Plant diversity effects on soil microbial functions and enzymes are stronger than warming in a grassland experiment. Ecology, 96, 99-112. [本文引用: 2]
[50]
TessierJT, RaynalDJ (2003). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. Journal of Applied Ecology, 40, 523-534.
[51]
TresederKK, VitousekPM (2001). Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests. Ecology, 82, 946-954. [本文引用: 1]
[52]
Van der PuttenWH, BardgettRD, BeverJD, BezemerTM, CasperBB, FukamiT, KardolP, KlironomosJN, KulmatiskiA, SchweitzerJA, SudingKN, Van de VoordeTFJ, WardleDA (2013). Plant-soil feedbacks: The past, the present and future challenges. Journal of Ecology, 101, 265-276. [本文引用: 4]
[53]
van GrunsvenRHA, van der PuttenWH, BezemerTM, BerendseF, VeenendaalEM (2010). Plant-soil interactions in the expansion and native range of a poleward shifting plant species. Global Change Biology, 16, 380-385. [本文引用: 1]
WangGB, RuanHH, TangYF, HeR (2009). A review on the dynamics of soil microbial biomass in forest ecosystems. Journal of Anhui Agricultural University, 36, 100-104. (in Chinese with English abstract)[王国兵, 阮宏华, 唐燕飞, 何容 (2009). 森林土壤微生物生物量动态变化研究进展 . 安徽农业大学学报, 36, 100-104.] [本文引用: 1]
[56]
WangXM, YanBG, LiuGC, FangHD (2016). Spatial variation in seed dormancy and germination of Dodonaea viscose in Yuanmou dry-hot valley, China. Journal of Tropical and Subtropical Botany, 24, 375-380. (in Chinese with English abstract)[王雪梅, 闫帮国, 刘刚才, 方海东 (2016). 元谋干热河谷车桑子种子休眠与萌芽的空间变异特征研究 . 热带亚热带植物学报, 24, 375-380.] [本文引用: 1]
[57]
WangZN, YangHM (2013). Responses of ecological stoi¬chiometry of carbon, nitrogen and phosphorus in plants to abiotic environmental factors. Pratacultural Science, 30, 927-934. (in Chinese with English abstract)[王振南, 杨惠敏 (2013). 植物碳氮磷生态化学计量对非生物因子的响应 . 草业科学, 30, 927-934.]
[58]
WuTG, WuM, LiuL, XiaoJH (2010). Seasonal variations of leaf nitrogen and phosphorus stoichiometry of three herbaceous species in Hangzhou Bay coastal wetlands, China. Chinese Journal of Plant Ecology, 34, 23-28. (in Chinese with English abstract)[吴统贵, 吴明, 刘丽, 萧江华 (2010). 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化 . 植物生态学报, 34, 23-28.] [本文引用: 2]
[59]
WuY, YuX, WangH, DingN, XuJ (2010). Does history matter? Temperature effects on soil microbial biomass and community structure based on the phospholipid fatty acid (PLFA) analysis. Journal of Soils and Sediments, 10, 223-230.
[60]
XuX, OuyangH, RichterA, WanekW, CaoG, KuzyakovY (2011). Spatio-temporal variations determine plant-micr¬obe competition for inorganic nitrogen in an alpine meadow. Journal of Ecology, 99, 563-571. [本文引用: 1]
[61]
YanK, FuDG, HeF, DuanCQ (2011). Leaf nutrient stoichiometry of plants in the phosphorus-enriched soils of the Lake Dianchi watershed, southwestern China. Chinese Journal of Plant Ecology, 35, 353-361. (in Chinese with English abstract)[阎凯, 付登高, 何峰, 段昌群 (2011). 滇池流域富磷区不同土壤磷水平下植物叶片的养分化学计量特征 . 植物生态学报, 35, 353-361.]
[62]
YanZB, KimNY, HanTS, FangJY, HanWX (2013). Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of Arabidopsis thaliana. Chinese Journal of Plant Ecology, 37, 551-557. (in Chinese with English abstract)[严正兵, 金南瑛, 韩廷申, 方精云, 韩文轩 (2013). 氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响 . 植物生态学报, 37, 551-557.] [本文引用: 1]
[63]
ZengDH, ChenGS (2005). Ecological stoichiometry: A science to explore the complexity of living systems. Acta Phytoecologica Sinica, 29, 1007-1019. (in Chinese with English abstract)[曾德慧, 陈广生 (2005). 生态化学计量学: 复杂生命系统奥秘的探索 . 植物生态学报, 29, 1007-1019.]
[64]
ZhanXH, JiangYH (1999). Advances in researches on mechanism of microbial inoculants on promoting plant growth. Plant Nutrition and Fertilizer Science, 5, 97-105. (in Chinese with English abstract)[占新华, 蒋延惠 (1999). 微生物制剂促进植物生长机理的研究进展 . 植物营养与肥料学报, 5, 97-105.] [本文引用: 1]
[65]
ZhangAM, LiNK, ZhaoGY, ZhangSF (2015). Research progress on the phosphate-solubilizing and potassium-solubilizing microorganisms. Journal of Hebei University (Natural Science Edition), 35, 442-448. (in Chinese with English abstract)[张爱民, 李乃康, 赵钢勇, 张双凤 (2015). 土壤中解磷、解钾微生物研究进展 . 河北大学学报(自然科学版), 35, 442-448.] [本文引用: 1]
[66]
ZhangQY, SunHL, LiSC, HuX, PangL (2013). Different environmental conditions on germination characteristics of Dodonaea viscosa (L.) Jacq. Seed, 32, 12-14. (in Chinese with English abstract)[张琼瑛, 孙海龙, 李绍才, 胡兴, 庞亮 (2013). 不同环境条件对车桑子萌发的影响 . 种子, 32, 12-14.]
[67]
ZhangW, ZhaoJ, PanF, LiD, ChenH, WangK (2015). Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant and Soil, 391, 77-91.
[68]
ZhaoZW, RenLC, LiT, LiJP (2003). Arbuscular mycorrhizas in the dry-hot valley of Jinsha River. Acta Botanica Yunnanica, 25, 199-204. (in Chinese with English abstract)[赵之伟, 任立成, 李涛, 李建平 (2003). 金沙江干热河谷(元谋段)的丛枝菌根 . 云南植物研究, 25, 199-204.] [本文引用: 1]
Stoichiometry and nutrition of plant growth in natural communities. 1 2008
... 植物生长需要多种生命元素, 碳(C)、氮(N)、磷(P)是构成有机体的主要元素, 其含量及比例特征是生态化学计量学研究的主要内容.C是组成生物结构的最基本元素, 构成了约50%的植物干质量, 而N和P是多数植物生长过程中主要的限制性元素(Ågren, 2008; Vitousek et al., 2010).植物C、N、P化学计量特征调节着植物生长(Fan et al., 2015), 其中, C:N和C:P在一定程度上反映了植物对N、P营养的利用效率, C:P和N:P可反映植物器官的生长速率, 并且N:P还是判断植物养分限制的重要指标(Elser et al., 2000; Tessier & Raynal, 2003; Güsewell, 2004; Zhang et al., 2015). ...
Effects of biocidal treatments on biological and nutritional properties of a mull-structured woodland soil. 1 1993
Alpine landscape variation in foliar nitrogen and phosphorus concentrations and the relation to soil nitrogen and phosphorus availability. Arctic, Antarctic, 2015
Plants actively control nitrogen cycling: Uncorking the microbial bottleneck. 1 2006
... 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011).一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015).土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011).土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面.由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道. ...
Do interactions between plant and soil biota change with elevation? A study on Fagus sylvatica. 1 2011
Growth rate-stoichiometry couplings in diverse biota. 2 2003
... 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011).一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015).土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011).土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面.由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道. ...
A species well travelled — The Dodonaea viscosa (Sapindaceae) complex based on phylogenetic analyses of nuclear ribosomal ITS and ETSf sequences. 1 2009
... 元谋县位于云贵高原北缘的金沙江一级支流——龙川江下游的河谷地带(101.58°-102.08° E, 25.42°-26.12° N), 海拔898-2 836 m, 该地区相对高差约2 000 m, 生态环境垂直分异明显.从河谷到山顶依次为河谷南亚热带、山地中亚热带、山地北亚热带及山地温带-中温带4个垂直气候带(李坤, 1993).通过野外考察, 选择元谋县高海拔区凉山乡和低海拔区苴林乡两个地点进行土壤样品采集.高海拔区的地理位置为101.94° E, 25.74° N, 海拔1 835 m, 年平均气温为14.6-15.9 ℃, 年降水量为 829 mm, 无霜期305-314天, 属北亚热带气候类型, 土壤类型为紫色土, 主要植被有车桑子、栎树(Quercus)、杜鹃(Rhododendron simsii)等乔灌植物; 低海拔区的地理位置为101.83° E, 25.85° N, 海拔 1 124 m, 年平均气温为21.9 ℃, 年降水量为621 mm, 无霜期363天, 属南亚热带气候类型(李坤, 1993), 土壤类型为燥红土, 主要植被为车桑子和黄茅(Heteropogon contortus). ...
... )等乔灌植物; 低海拔区的地理位置为101.83° E, 25.85° N, 海拔 1 124 m, 年平均气温为21.9 ℃, 年降水量为621 mm, 无霜期363天, 属南亚热带气候类型(李坤, 1993), 土壤类型为燥红土, 主要植被为车桑子和黄茅(Heteropogon contortus). ...
Regulation of soil phosphatase and chitinase activity by N and P availability. 2000
元谋干热河谷土壤微生物群落对水热变化的响应 1 2014
... 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011).一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015).土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011).土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面.由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道. ...
Plant-soil feedback: Experimental approaches, statistical analyses and ecological interpretations. 2010
元谋干热河谷气候生态环境变化的初步分析 1 2007
... 植物的生态化学计量动态受到自身和外界环境因素的共同控制.一方面, 其生态化学计量特征具有内稳性(曾德慧和陈广生, 2005; 牛得草等, 2011)和物种特异性(石贤萌等, 2015); 另一方面, 植物的生态化学计量特征又随环境条件(如纬度、温度、降水、土壤肥力、群落演替等)的改变而发生改变(Reich & Oleksyn, 2004; Han et al., 2005; Sardans et al., 2012; 王振南和杨惠敏, 2013).大多数研究表明, 植物N、P养分特征与土壤养分具有很强的相关性, 其化学计量特征受土壤肥力的影响很大(阎凯等, 2011; 宾振钧等, 2015; Fan et al., 2015).但这些研究基本上都是基于野外采样的数据, 植物养分是在长时间不同条件下形成的, 短期的控制试验能否造成物种水平上化学计量特征的改变还不得而知.但室内条件下化学计量特征对环境变化的响应更有利于研究单个物种的化学计量规律, 为野外研究不同物种、群落或生态系统的化学计量特征提供参考(严正兵等, 2013; 刘玉芳等, 2015). ...
Effects of plant cover on properties of rhizosphere and inter-plant soil in a semiarid valley, SW China. 2015
Global patterns of plant leaf N and P in relation to temperature and latitude. 2004
Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. 1 2008
... 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011).一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015).土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011).土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面.由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道. ...
Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). 2012
Re-inoculation of autoclaved soil as a non-sterile treatment for xenobiotic sorption and biodegradation studies. 1 1999
... 植物的生态化学计量动态受到自身和外界环境因素的共同控制.一方面, 其生态化学计量特征具有内稳性(曾德慧和陈广生, 2005; 牛得草等, 2011)和物种特异性(石贤萌等, 2015); 另一方面, 植物的生态化学计量特征又随环境条件(如纬度、温度、降水、土壤肥力、群落演替等)的改变而发生改变(Reich & Oleksyn, 2004; Han et al., 2005; Sardans et al., 2012; 王振南和杨惠敏, 2013).大多数研究表明, 植物N、P养分特征与土壤养分具有很强的相关性, 其化学计量特征受土壤肥力的影响很大(阎凯等, 2011; 宾振钧等, 2015; Fan et al., 2015).但这些研究基本上都是基于野外采样的数据, 植物养分是在长时间不同条件下形成的, 短期的控制试验能否造成物种水平上化学计量特征的改变还不得而知.但室内条件下化学计量特征对环境变化的响应更有利于研究单个物种的化学计量规律, 为野外研究不同物种、群落或生态系统的化学计量特征提供参考(严正兵等, 2013; 刘玉芳等, 2015). ...
Plant-soil feedbacks: The past, the present and future challenges. 4 2013
... 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011).一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015).土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011).土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面.由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道. ...
... 14:0 iso, 15:0 iso, 15:0 anteiso, 16:0 iso, 17:0 iso, 17:0 anteiso and 18:0 iso in the table represent Gram-positive bacteria; 16:1w9c and 17:0 cyclo represent Gram-negative bacteria; 18:0 indicates hydrogenobacter; 16:1 w5c indicates arbuscular mycorrhizal fungi (Qu et al., 2015). ...
... 土壤微生物是土壤活有机体的重要组成部分, 与植物养分密切相关(Miransari, 2011).一方面, 微生物(如共生固氮菌、丛枝菌根真菌、植物根际促生菌)对植物养分获得具有直接的促进作用; 另一方面, 微生物通过土壤养分循环特别是N、P循环间接地影响植物营养(沈仁芳和赵学强, 2015).土壤微生物对植物存在着反馈效应(Brinkman et al., 2010; van der Putten et al., 2013), 但在不同气候条件下土壤微生物的群落结构和生物多样性是不同的(Liang et al., 2015), 土壤微生物在面对不同环境变化尤其是温度变化时会有不同的响应(Wu et al., 2010), 并且, 土壤微生物对植物的反馈效应存在着海拔差异(Defossez et al., 2011).土壤微生物对植物的反馈效应研究目前主要集中于植物生物量、植物性状等方面.由于植物营养与土壤微生物间的紧密关系, 不同气候下的土壤微生物也会影响植物化学计量特征, 但关于土壤微生物对植物生态化学计量特征影响的研究还少见报道. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT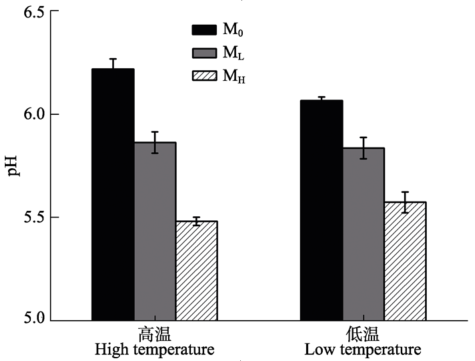 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT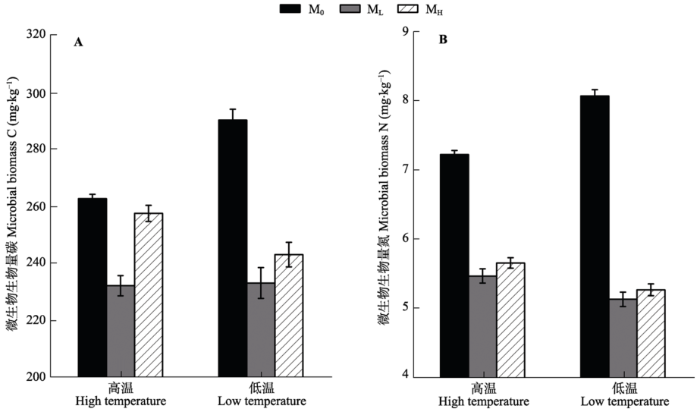 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT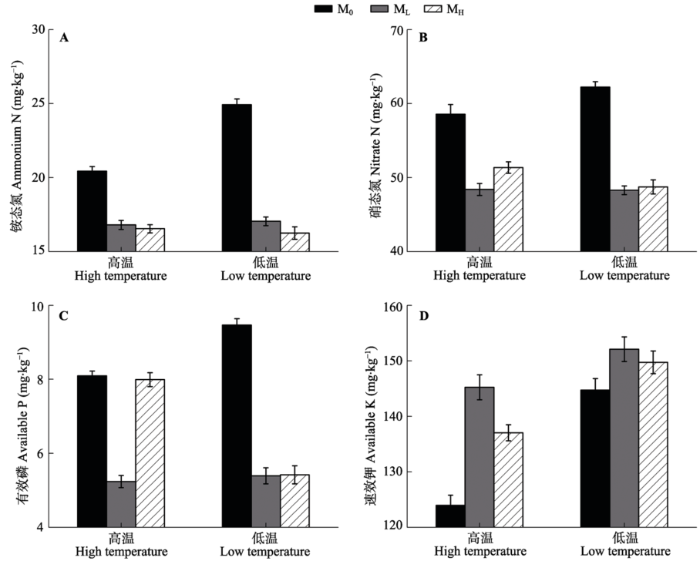 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}