关键词:生态化学计量特征;荒漠草原;甘草;磷添加 Abstract Aims The increase in atmospheric nitrogen (N) deposition has accelerated N cycling of ecosystems, probably resulting in increases in phosphorus (P) demand of ecosystems. Studies on the effects of artificial N:P treatment on the growth and carbon (C), N, P ecological stoichiometry of desert steppe species could provide not only a new insight into the forecasting of how the interaction between soils and plants responses to long-term atmospheric N deposition increase, but also a scientific guidance for sustainable management of grassland in northern China under global climate change. Methods Based on a pot-cultured experiment conducted for Glycyrrhiza uralensis (an N-fixing species) during 2013 to 2014, we studied the effects of different N:P supply ratios (all pots were treated with the same amount of N but with different amounts of P) on aboveground biomass, root biomass, root/shoot ratio, and C:N:P ecological stoichiometry both in G. uralensis (leaves and roots) and in soils. Additionally, through the correlation analyses between biomass and C:N:P ecological stoichiometry in leaves, roots, and soils, we compared the differences among the C:N:P ecological stoichiometry of the three pools, and discussed the indication of C:N:P ecological stoichiometry in soils for the growth and nutrient uptake of G. uralensis. Important findings The results showed that, reducing N:P decreased C:P and N:P ratios both in G. uralensis (leaves and roots) and in soils but increased aboveground biomass and root biomass of G. uralensis, indicating that low to moderate P addition increased P availability of soils and P uptake of G. uralensis. However, excessive low N:P (high P addition) led to great decreases in soil C:P and N:P ratios, thus hindering N uptake and the growth of G. uralensis. C:N:P ratios in the two pools of G. uralensis (especially in leaves) had close correlations with soil C:N:P ratio, indicating that the change in soil C:N:P ratio would have a direct influence on plants. Our results suggest that, through regulating C:N:P ratio in leaves and soils, appropriate amounts of P addition could balance soil P supply and plant P demand and compensate the opposite influences of long-term atmospheric N deposition increase on the structure of desert steppe.
Keywords:ecological stoichiometry;desert steppe;Glycyrrhiza uralensis;phosphorus addition -->0 PDF (3552KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 黄菊莹, 余海龙, 王丽丽, 马凯博, 康扬眉, 杜雅仙. 不同氮磷比处理对甘草生长与生态化学计量特征的影响. 植物生态学报, 2017, 41(3): 325-336 https://doi.org/10.17521/cjpe.2016.0230 HUANGJu-Ying, YUHai-Long, WANGLi-Li, MAKai-Bo, KANGYang-Mei, DUYa-Xian. Effects of different nitrogen:phosphorus levels on the growth and ecological stoichiometry of Glycyrrhiza uralensis. Chinese Journal of Plant Ecology, 2017, 41(3): 325-336 https://doi.org/10.17521/cjpe.2016.0230 大气氮(N)沉降增加, 是人类面临的一个全球性气候变化问题(IPCC, 2013), 其生态学效应受到越来越多科研工作者的关注。一般而言, 在受N限制的生态系统, 短期或少量N沉降增加可以刺激N矿化(Jones & Power, 2012)、促进微生物-酶系统对土壤有机质和养分的分解(Liu et al., 2010), 从而提高土壤N和磷(P)的有效性。随着长期或过多N沉降, 不仅生态系统N输入增加, 而且N:P失衡、系统N饱和甚至P压力增加(Bobbink et al., 2010; Wardle et al., 2013), 以及植物生长P受限性增强(Yu et al., 2012)。在石灰性草原生态系统, 研究人员发现N添加显著提高了土壤磷酸单酯酶活性, 而土壤磷酸单酯酶活性与土壤有效P含量显著负相关, 因此认为长期N沉降增加导致该生态系统P受限性增加; 随着过多N输入, 植物根系和微生物通过分泌较多的磷酸酶提高植株对土壤P元素的吸收效率, 从而保证植物的正常生长(Phoenix et al., 2004)。在其他一些生态系统, 如热带森林生态系统(Zhu et al., 2013)、温带森林生态系统(Naples & Fisk, 2010)和沼泽生态系统(Phuyal et al., 2008), 研究人员也报道了类似的结果。由于P不仅是细胞间能量传递和核苷酸结构组成的重要元素, 而且是许多自然生态系统植物生长的限制因素, 因此P受限性的增加可能会影响到植物P策略和生态系统P循环, 从而间接地改变生态系统结构和功能(Bobbink et al., 2010)。 荒漠草原是最旱生的一类草原生态系统, 具有群落生产力低和稳定性差的特点, 对自然和人类活动的干扰比较敏感。受气候和地理位置的影响, 宁夏回族自治区境内分布有大面积荒漠草原, 其植被生长不仅受N的限制, 而且年际差异大, 导致生态系统的不稳定性和脆弱性。尽管在荒漠草原监测到的大气N沉降量低于其他生态系统(Liu et al., 2013), 但是该生态系统本身N沉降临界负荷低(段雷等, 2002), 而且即使是低剂量的N添加, 其长期的影响也是巨大的(Clark & Tilman, 2008), 因此有必要针对该生态系统开展大气N沉降效应的模拟研究。2011-2013年, 在国家自然科学基金青年项目的资助下, 本项目组在宁夏盐池荒漠草原设置了一个N沉降增加的野外模拟试验, 探讨了N添加对植物和土壤碳(C)、N、P化学计量学特征的影响。研究结果表明, 生长季内物种相对生长速率、群落生物量和物种数基本在10.0 g·m-2·a-1的N添加下达到最大, 超过10.0 g·m-2·a-1的N添加后, 各指标逐渐降低; 少量N添加对土壤C、N、P及其化学计量比的影响较小, 但高N添加提高了土壤速效N含量、降低了速效P含量(Huang et al., 2016; 黄菊莹和余海龙, 2016), 这意味着高N添加可能提高了荒漠草原P受限性。那么, 在高N添加的基础上施P将如何影响荒漠草原植物生长和化学计量学特征?P添加是否可以缓解N添加引起的P受限性增加?土壤养分受限性转变又将如何作用于植物?这些问题都值得我们进行深入研究。
降低施入的N:P (P添加量增加)提高了甘草地上生物量、地下生物量和总生物量, 并在8 g·m-2·a-1的P添加下3个指标达到最大值1.5、2.3和 3.8 g·plant-1, 表明当N:P处于中等水平时, 甘草具有较高的生物量, 过高和过低的N:P都不利于甘草生物量积累(图1A、1B、1C)。与地上生物量、地下生物量和总生物量不同, 8.0 g·m-2·a-1的P处理使根冠比与其他处理相比降低了21.7%-33.1% (图1D)。 显示原图|下载原图ZIP|生成PPT 图1不同氮(N)磷(P)比处理对甘草生物量和根冠比的影响(平均值±标准误差, n = 5)。N10P1、N10P2、N10P4、N10P8、N10P16和N10P32代表在统一施用10.0 g·m-2·a-1 N的基础上,分别施入1.0、2.0、4.0、8.0、16.0和32.0 g·m-2·a-1的P。不同小写字母表示N:P处理间指标差异显著(p < 0.05), 相同字母表示差异不显著(p > 0.05)。 -->Fig. 1Effects of nitrogen (N):phosphorus (P) level on root and aboveground biomass and root/shoot ratio of Glycyrrhiza uralensis (mean ± SE, n = 5). N10P1, N10P2, N10P4, N10P8, N10P16, and N10P32 represent all pots treated with 10.0 g·m-2·a-1 amount of N but with different amounts of P: 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 g·m-2·a-1, respectively. Different lowercase letters indicate significant difference (p < 0.05) between indices within N:P levels. The same lowercase letters indicate insignificant differences (p > 0.05). -->
2.3 不同N:P处理对甘草C、N、P含量及其化学计量比的影响
降低施入的N:P提高了甘草叶片P浓度, 降低了N浓度、C:P和N:P, 对C浓度和C:N的影响无显著的规律性(图2A、2D)。平均叶片C浓度、N浓度、P浓度、C:N、C:P和N:P分别为376.6 mg·g-1、33.1 mg·g-1、2.8 mg·g-1、12.2、143.5和12.5。 降低施入的N:P提高了甘草根系的C浓度、N浓度、P浓度和C:N, 降低了N:P (图3F), 对C:P的影响无明显的规律性。平均根系C浓度、N浓度、P浓度、C:N、C:P和N:P分别为426.5 mg·g-1、21.3 mg·g-1、1.9 mg·g-1、20.2、228.7和11.4。 显示原图|下载原图ZIP|生成PPT 图2不同氮(N)磷(P)比处理对甘草叶片碳(C)、N、P及其化学计量比的影响(平均值±标准误差, n = 5)。N10P1、N10P2、N10P4、N10P8、N10P16和N10P32代表在统一施用10.0 g·m-2·a-1 N的基础上, 分别施入1.0、2.0、4.0、8.0、16.0和32.0 g·m-2·a-1的P。不同小写字母表示N:P处理间指标差异显著(p < 0.05), 相同字母表示差异不显著(p > 0.05)。 -->Fig. 2Effects of nitrogen (N):phosphorus (P) level on leaf carbon (C), N, P and their stoichiometry ratios of Glycyrrhiza uralensis (mean ± SE, n = 5). N10P1, N10P2, N10P4, N10P8, N10P16, and N10P32 represent all pots treated with 10.0 g·m-2·a-1 amount of N but with different amounts of P: 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 g·m-2·a-1, respectively. Different lowercase letters indicate significant difference (p < 0.05) between indices within N:P levels. The same lowercase letters indicate insignificant difference (p > 0.05). -->
显示原图|下载原图ZIP|生成PPT 图3不同氮(N)磷(P)比处理对甘草根系碳(C)、N、P及其化学计量比的影响(平均值±标准误差, n = 5)。N10P1、N10P2、N10P4、N10P8、N10P16和N10P32代表在统一施用10.0 g·m-2·a-1 N的基础上, 分别施入1.0、2.0、4.0、8.0、16.0和32.0 g·m-2·a-1的P。不同小写字母表示N:P处理间指标差异显著(p < 0.05), 相同字母表示差异不显著(p > 0.05)。 -->Fig. 3Effects of nitrogen (N): phosphorus (P) level on root carbon (C), N, P and their stoichiometry ratios of Glycyrrhiza uralensis (mean ± SE, n = 5). N10P1, N10P2, N10P4, N10P8, N10P16, and N10P32 represent all pots were treated with 10.0 g·m-2·a-1 amount of N but with differing amounts of P: 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 g·m-2·a-1, respectively. Different lowercase letters indicate significant differences (p < 0.05) between indices within N:P levels. The same lowercase letters indicate insignificant differences (p > 0.05). -->
2.4 不同N:P处理对8月份土壤C、N、P含量及其化学计量比的影响
降低施入的N:P提高了土壤全P和速效P含量(图4C、4F), 降低了C:P和N:P (图5B、5C), 对其他指标的影响无一致的规律性。平均土壤有机C含量、全N含量、全P含量、NH4+-N含量、NO3--N含量、速效P含量、C:N、C:P和N:P分别为2.0 g·kg-1、0.3 g·kg-1、0.7 g·kg-1、1.6 mg·g-1、22.7 mg·g-1、103.9 mg·g-1、7.1、3.7和0.5。 显示原图|下载原图ZIP|生成PPT 图4不同氮(N)磷(P)比处理对土壤碳(C)、N、P含量的影响(平均值±标准误差, n = 5)。N10P1、N10P2、N10P4、N10P8、N10P16和N10P32代表在统一施用10.0 g·m-2·a-1 N的基础上, 分别施入1.0、2.0、4.0、8.0、16.0和32.0 g·m-2·a-1的P。不同小写字母表示N:P处理间指标差异显著(p < 0.05), 相同字母表示差异不显著(p > 0.05)。 -->Fig. 4Effects of nitrogen (N):phosphorus (P) level on soil carbon (C), N, and P content (mean ± SE, n = 5). N10P1, N10P2, N10P4, N10P8, N10P16, and N10P32 represent all pots treated with 10.0 g·m-2·a-1 amount of N but with different amounts of P: 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 g·m-2·a-1, respectively. Different lowercase letters indicate significant difference (p < 0.05) between indices within N:P levels. The same lowercase letters indicate insignificant difference (p > 0.05). -->
2.5 甘草叶、根系和土壤C、N、P及其化学计量比的相关性
甘草生物量与土壤C:N:P化学计量学特征的相关性较弱(表2), 只有地上生物量与土壤速效P含量、地下生物量与土壤C:N存在显著的相关性。 如表2所示, 叶片全C与土壤C:N:P化学计量学特征无显著的相关性; 叶片全N与土壤C:N和C:P正相关, 与全N负相关; 叶片全P与土壤全P正相关, 与C:P和N:P负相关; 叶片C:N与土壤C:N负相关; 叶片C:P与土壤C:P和N:P正相关, 与全P负相关; 叶片N:P与土壤C:N、C:P和N:P正相关, 与全P和速效P负相关。 根系全C与土壤速效P正相关, 与N:P负相关; 根系全N与土壤全P负相关; 根系全P和C:P均与土壤C:N:P化学计量学特征无显著的相关性; 根系C:N与土壤全P正相关, 与C:P和N:P负相关; 根系N:P与土壤C:P和N:P正相关, 与全P负相关。 Table 2 表2 表2甘草与土壤碳(C)、氮(N)、磷(P)含量及其化学计量比间的相关系数 Table 2Correlation coefficients between carbon (C), nitrogen (N), phosphorus (P ) and their stoichiometry ratios in Glycyrrhiza uralensis and in soils
指标 Index
土壤有机C Soil organic C (g·kg-1)
土壤全N Soil total N (g·kg-1)
土壤全P Soil total P (g·kg-1)
土壤速效P Soil available P (mg·kg-1)
土壤C:N Csoil:Nsoil
土壤C:P Csoil:Psoil
土壤N:P Nsoil:Psoil
地上生物量 Aboveground biomass (g·plant-1)
ns
ns
ns
0.51*
ns
ns
0.05
地下生物量 Belowground biomass (g·plant-1)
ns
ns
ns
ns
0.50*
ns
ns
叶片全C Leaf total C (mg·g-1)
ns
ns
ns
ns
ns
ns
ns
叶片全N Leaf total N (mg·g-1)
ns
-0.56*
ns
ns
0.69**
0.50*
ns
叶片全P Leaf total P (mg·g-1)
ns
ns
0.77**
ns
ns
-0.70**
-0.75**
叶片C:N Cleaf:Nleaf
ns
ns
ns
ns
-0.56*
ns
ns
叶片C:P Cleaf:Pleaf
ns
ns
-0.63**
ns
ns
0.58*
0.64**
叶片N:P Nleaf:Pleaf
ns
ns
-0.80**
-0.49*
0.57*
0.85**
0.79**
根系全C Root total C (mg·g-1)
ns
ns
ns
0.62**
ns
ns
-0.53*
根系全N Root total N (mg·g-1)
ns
ns
-0.50*
ns
ns
ns
ns
根系全P Root total P (mg·g-1)
ns
ns
ns
ns
ns
ns
ns
根系C:N Croot:Nroot
ns
ns
0.66**
ns
ns
-0.58*
-0.70**
根系C:P Croot:Proot
ns
ns
ns
ns
ns
ns
ns
根系N:P Nroot:Proot
ns
ns
-0.57*
ns
ns
0.65**
0.67**
ns indicates that correlation is insignificant at 0.05 level. * and ** indicate that correlations are significant at the 0.05 and 0.01 levels, respectively.ns代表显著性水平大于0.05。*和**分别代表显著性水平小于0.05和0.01。 新窗口打开
2.6 甘草生长与C:N:P化学计量学特征的关系
总体而言, 甘草生长与C:N:P化学计量学特征的相关性较弱(表3), 如: 地上生物量仅与土壤速效P含量以及根系全N浓度正相关, 而与其他指标无显著的相关性; 地下生物量与土壤C:N正相关, 与其他指标无显著的相关性; 总生物量和根冠比均与C、N、P及其化学计量比不存在显著的相关性。 Table 3 表3 表3碳(C):氮(N):磷(P)化学计量学特征与甘草生物量和根冠比的相关系数 Table 3Correlation coefficients between carbon (C):nitrogen (N):phosphorus (P) ecological stoichiometry and the biomass or root/shoot ratio of Glycyrrhiza uralensis
指标 Index
地上生物量 Aboveground biomass (g·plant-1)
地下生物量 Belowground biomass (g·plant-1)
总生物量 Total biomass (g·plant-1)
根冠比 Root/shoot ratio
土壤有机C Soil organic C (g·kg-1)
0.18
0.35
0.32
-0.37
土壤全N Soil total N (g·kg-1)
-0.01
-0.33
-0.23
-0.21
土壤全P Soil total P (g·kg-1)
-0.27
-0.39
-0.39
0.01
土壤速效P Soil available P (mg·kg-1)
0.51*
0.18
0.35
-0.05
土壤C:N Csoil:Nsoil
0.12
0.50*
0.40
0.04
土壤C:P Csoil:Psoil
-0.00
0.19
0.13
0.18
土壤N:P Nsoil:Psoil
0.05
0.07
0.07
0.13
叶片全C Leaf total C (mg·g-1)
0.07
-0.08
-0.03
-0.33
叶片全N Leaf total N (mg·g-1)
0.25
0.35
0.35
-0.27
叶片全P Leaf total P (mg·g-1)
-0.07
-0.31
-0.25
-0.25
叶片C:N Cleaf:Nleaf
-0.18
-0.21
-0.22
0.27
叶片C:P Cleaf:Pleaf
0.02
0.22
0.16
0.41
叶片N:P Nleaf:Pleaf
0.07
0.29
0.23
0.15
根系全C Root total C (mg·g-1)
0.39
0.39
0.44
-0.68
根系全N Root total N (mg·g-1)
0.49*
0.21
0.36
-0.58
根系全P Root total P (mg·g-1)
0.43
0.19
0.32
-0.55
根系C:N Croot:Nroot
-0.22
-0.03
-0.12
0.10
根系C:P Croot:Proot
-0.29
-0.05
-0.17
0.39
根系N:P Nroot:Proot
-0.07
-0.04
-0.06
0.18
*, p < 0.05。 新窗口打开 显示原图|下载原图ZIP|生成PPT 图5不同氮(N)磷(P)比处理对土壤碳(C):N:P化学计量比的影响(平均值±标准误差, n = 5)。N10P1、N10P2、N10P4、N10P8、N10P16和N10P32代表在统一施用10.0 g·m-2·a-1 N的基础上, 分别施入1.0、2.0、4.0、8.0、16.0和32.0 g·m-2·a-1的P。不同小写字母表示N:P处理间指标差异显著(p < 0.05), 相同字母表示差异不显著(p > 0.05)。 -->Fig. 5Effects of nitrogen (N): phosphorus (P) level on soil carbon (C) :N:P stoichiometry ratio (mean ± SE, n = 5). N10P1, N10P2, N10P4, N10P8, N10P16, and N10P32 represent all pots treated with 10.0 g·m-2·a-1 amount of N but with different amounts of P: 1.0, 2.0, 4.0, 8.0, 16.0, and 32.0 g·m-2·a-1, respectively. Different lowercase letters indicate significant difference (p < 0.05) between indices within N:P levels. The same lowercase letters indicate insignificant difference (p > 0.05). -->
BaoSD (2000). Soil and Agricultural Chemistry Analysis. 3rd ed. China Agriculture Press, Beijing. (in Chinese).[鲍士旦 (2000). 土壤农化分析. 第三版. 中国农业出版社, 北京.]
[2]
BobbinkR, HicksK, GallowayJ, SprangerT, AlkemadeR, AshmoreM, BustamanteM, CinderbyS, DavidsonE, DentenerF, EmmettB, ErismanJW, FennM, GilliamF, NordinA, PardoL, de VriesW (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecological Applications, 20, 30-59.
[3]
ChenFS, HuXF, GeG (2007). Leaf N:P stoichiometry and nutrient resorption efficiency of Ophiopogon japonicus in Nanchang City. Acta Prataculturae Sinica, 16(4), 47-54. (in Chinese with English abstract)[陈伏生, 胡小飞, 葛刚 (2007). 城市地被植物麦冬叶片氮磷和养分再吸收效率 . 草业学报, 16(4), 47-54.] [本文引用: 2]
[4]
ClarkCM, TilmanD (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 451, 712-715.
[5]
ClevelandCC, LiptzinD (2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 85, 235-252. [本文引用: 1]
[6]
CrewsTE, FarringtonH, VitousekPM (2000). Changes in a symbiotic, heterotrophic nitrogen fixation on leaf litter of Metrosideros polymorpha with long-term ecosystem development in Hawaii. Ecosystems, 3, 386-395. [本文引用: 1]
[7]
CrossAF, SchlesingerWH (2001). Biological and geochemical controls on phosphorus fractions in semiarid soils. Biogeochemistry, 52, 155-172. [本文引用: 1]
[8]
CrousJW, MorrisAR, ScholesMC (2008). Growth and foliar nutrient response to recent applications of phosphorus (P) and potassium (K) and to residual P and K fertiliser applied to the previous rotation of Pinus patula at Usutu, Swaziland. Forest Ecology Management, 256, 712-721. [本文引用: 1]
DuanL, HaoJM, XieSD, ZhouZP (2002). Estimating critical loads of sulfur and nitrogen for Chinese soils by steady state method. Journal of Environmental Science, 23(2), 7-12. (in Chinese with English abstract).[段雷, 郝吉明, 谢绍东, 周中平 (2002). 用稳态法确定中国土壤的硫沉降和氮沉降临界负荷 . 环境科学, 23(2), 7-12.] [本文引用: 1]
[11]
FritzC, van DijkG, SmoldersAJP, PancottoVA, ElzengaTJTM, RoelofsJGM, GrootjansAP (2012). Nutrient additions in pristine Patagonian Sphagnum bog vegetation: Can phosphorus addition alleviate (the effects of) increased nitrogen loads. Plant Biology, 14, 491-499. [本文引用: 1]
[12]
GuoZW, ChenSL, YangQP, LiYC (2012). Responses of N and P stoichiometry on mulching management in the stand of Phyllostachys praecox. Acta Ecologica Sinica, 32, 6361-6368. (in Chinese with English abstract)[郭子武, 陈双林, 杨清平, 李迎春 (2012). 雷竹林土壤和叶片N、P化学计量特征对林地覆盖的响应 . 生态学报, 32, 6361-6368.]
[13]
GüsewellS (2005). Responses of wetland graminoids to the relative supply of nitrogen and phosphorus. Plant Ecology, 176, 35-55. [本文引用: 2]
[14]
HeLY, HuZM, GuoQ, LiSG, BaiWM, LiLH (2015). Influence of nitrogen and phosphorus addition on the aboveground biomass in Inner Mongolia temperate steppe, China. Chinese Journal of Applied Ecology, 26, 2291-2297. (in Chinese with English abstract)[何利元, 胡中民, 郭群, 李胜功, 白文明, 李凌浩 (2015). 氮磷添加对内蒙古温带草地地上生物量的影响 . 应用生态学报, 26, 2291-2297.] [本文引用: 1]
[15]
HuangJY, YuHL (2016). Responses of growth of four desert species to different N addition levels. Chinese Journal of Plant Ecology, 40, 165-176. (in Chinese with English abstract)[黄菊莹, 余海龙 (2016). 4个荒漠草原物种的生长对不同氮添加水平的响应 . 植物生态学报, 40, 165-176.]
[16]
HuangJY, YuHL, LinH, ZhangY, SearleEB, YuanZY (2016). Phosphorus amendment mitigates nitrogen addition-induced phosphorus limitation in two plant species in a desert steppe, China. Plant and Soil, 399, 221-232.
[17]
IPCC (Intergovernmental Panel on Climate Change) (2013). Climate Change 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK. [本文引用: 3]
[18]
JonesAG, PowerSA (2012). Field-scale evaluation of effects of nitrogen deposition on the functioning of heathland ecosystems. Journal of Ecology, 100, 331-342.
[19]
LiXB, ChenL, LIGQ, AnH (2013). Influence of enclosure on Glyeyrrhiza uralensis community and distribution pattern in arid and semi-arid areas. Acta Ecologica Sinica, 33, 3995-4001. (in Chinese with English abstract)[李学斌, 陈林, 李国旗, 安慧 (2013). 干旱半干旱地区围栏封育对甘草群落特征及其分布格局的影响 . 生态学报, 33, 3995-4001.] [本文引用: 1]
[20]
LiXB, ZhaoZF, ChenL, AnH, LiGQ, LiuBR (2012). The status and countermeasures of development on licorices industry of Ningxia. Ecological Economy, 12, 132-135. (in Chinese with English abstract)[李学斌, 赵志锋, 陈林, 安慧, 李国旗, 刘秉儒 (2012). 宁夏甘草产业发展现状及对策研究 . 生态经济, 12, 132-135.] [本文引用: 2]
[21]
LiuC, WangY, WangN, WangGX (2012). Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial ecosystems: A review. Chinese Journal of Plant Ecology, 36, 1205-1216. (in Chinese with English abstract)[刘超, 王洋, 王楠, 王根轩 (2012). 陆地生态系统植被氮磷化学计量研究进展 . 植物生态学报, 36, 1205-1216.] [本文引用: 1]
[22]
LiuP, HuangJH, SunOJX (2010). Litter decomposition and nutrient release as affected by soil nitrogen availability and litter quality in a semiarid grassland ecosystem. Oecologia, 162, 771-780.
LiuY, ZhangJ, ChenYM, ChenL, LiuQ (2013). Effect of nitrogen and phosphorus fertilization on biomass allocation and C:N:P stoichiometric characteristics of Eucalyptus grandis seedlings. Chinese Journal of Plant Ecology, 37, 933-941. (in Chinese with English abstract)[刘洋, 张健, 陈亚梅, 陈磊, 刘强 (2013). 氮磷添加对巨桉幼苗生物量分配和C:N:P化学计量特征的影响 . 植物生态学报, 37, 933-941.]
[25]
MayorJR, WrightSJ, TurnerBL (2014). Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest. Journal of Ecology, 102, 36-44. [本文引用: 1]
[26]
NaplesBK, FiskMC (2010). Belowground insights into nutrient limitation in northern hardwood forests. Biogeochemistry, 97, 109-121.
[27]
PhoenixGK, BoothRE, LeakeJR, ReadDJ, GrimeJP, LeeJA (2004). Simulated pollutant nitrogen deposition increases P demand and enhances root-surface phosphatase activities of three plant functional types in a calcareous grassland. New Phytologist, 161, 279-289. [本文引用: 1]
[28]
PhuyalM, ArtzRRE, SheppardL, LeithID, JohnsonD (2008). Long-term nitrogen deposition increases phosphorus limitation of bryophytes in an Ombrotrophic Bog. Plant Ecology, 196, 111-121.
[29]
ShiXM, QiJH, SongL, LiuWY, HuangJB, LiS, LuHZ, ChenX (2015). C, N and P stoichiometry of two dominant seedlings and their responses to nitrogen additions in the montane moist evergreen broad-leaved forest in Ailao Mountains, Yunnan. Chinese Journal of Plant Ecology, 39, 962-970. (in Chinese with English abstract)[石贤萌, 杞金华, 宋亮, 刘文耀, 黄俊彪, 李苏, 卢华正, 陈曦 (2015). 哀牢山中山湿性常绿阔叶林两种优势幼苗C、N、P化学计量特征及其对N沉降增加的响应 . 植物生态学报, 39, 962-970.] [本文引用: 1]
[30]
TianHQ, ChenGS, ZhangC, MelilloJM, HallCAS (2010). Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry, 98, 139-151. [本文引用: 1]
[31]
WardleDA, GundaleMJ, JaderlundA, NilssonMC (2013). Decoupled long-term effects of nutrient enrichment on aboveground and belowground properties in subalpine tundra. Ecology, 94, 904-919. [本文引用: 2]
[32]
YangXX, RenF, ZhouHK, HeJS (2014). Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology, 38, 159-166. (in Chinese with English abstract)[杨晓霞, 任飞, 周华坤, 贺金生 (2014). 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应 . 植物生态学报, 38, 159-166.]
[33]
YangYH, FangJY, JiCJ, DattaA, LiP, MaWH, MohammatA, ShenHH, HuHF, KnappBO, SmithP (2014). Stoichiometric shifts in surface soils over broad geographical scales: Evidence from China’s grasslands. Global Ecology and Biogeography, 23, 947-955. [本文引用: 1]
[34]
YuQ, WuHH, HeNP, LüXT, WangZP, ElserJJ, WuJG, HanXG (2012). Testing the growth-rate hypothesis in vascular plants with above- and below-ground biomass. PLOS ONE, 7, e32162. doi: 10.1371/journal.pone.0032162. [本文引用: 1]
[35]
YuanZY, ChenHYH (2015). Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nature Climate Change, 5, 465-469. [本文引用: 1]
[36]
ZhaoYF, XuFL, WangWL, WangLL, WangGX, SunPY, BaiXF (2014). Seasonal variation in contents of C, N and P and stoichiometry characteristics in fine roots, stems and needles of Larix principis-rupprechtii. Chinese Bulletin of Botany, 49, 560-568. (in Chinese with English abstract)[赵亚芳, 徐福利, 王渭玲, 王玲玲, 王国兴, 孙鹏跃, 白小芳 (2014). 华北落叶松根茎叶碳氮磷含量及其化学计量学特征的季节变化 . 植物学报, 49 , 560-568.] [本文引用: 1]
[37]
ZhuFF, YohM, GilliamFS, LuXK, MoJM (2013). Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions. PLOS ONE, 8, e82661. doi:10.1371/journal.pone.0082661. [本文引用: 2]
Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. 2010
城市地被植物麦冬叶片氮磷和养分再吸收效率 2 2007
... 大气氮(N)沉降增加, 是人类面临的一个全球性气候变化问题(IPCC, 2013), 其生态学效应受到越来越多科研工作者的关注.一般而言, 在受N限制的生态系统, 短期或少量N沉降增加可以刺激N矿化(Jones & Power, 2012)、促进微生物-酶系统对土壤有机质和养分的分解(Liu et al., 2010), 从而提高土壤N和磷(P)的有效性.随着长期或过多N沉降, 不仅生态系统N输入增加, 而且N:P失衡、系统N饱和甚至P压力增加(Bobbink et al., 2010; Wardle et al., 2013), 以及植物生长P受限性增强(Yu et al., 2012).在石灰性草原生态系统, 研究人员发现N添加显著提高了土壤磷酸单酯酶活性, 而土壤磷酸单酯酶活性与土壤有效P含量显著负相关, 因此认为长期N沉降增加导致该生态系统P受限性增加; 随着过多N输入, 植物根系和微生物通过分泌较多的磷酸酶提高植株对土壤P元素的吸收效率, 从而保证植物的正常生长(Phoenix et al., 2004).在其他一些生态系统, 如热带森林生态系统(Zhu et al., 2013)、温带森林生态系统(Naples & Fisk, 2010)和沼泽生态系统(Phuyal et al., 2008), 研究人员也报道了类似的结果.由于P不仅是细胞间能量传递和核苷酸结构组成的重要元素, 而且是许多自然生态系统植物生长的限制因素, 因此P受限性的增加可能会影响到植物P策略和生态系统P循环, 从而间接地改变生态系统结构和功能(Bobbink et al., 2010). ...
... ), 研究人员也报道了类似的结果.由于P不仅是细胞间能量传递和核苷酸结构组成的重要元素, 而且是许多自然生态系统植物生长的限制因素, 因此P受限性的增加可能会影响到植物P策略和生态系统P循环, 从而间接地改变生态系统结构和功能(Bobbink et al., 2010). ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. 2008
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? 1 2007
... 著降低(图5B、图5C).受土壤P供给水平变化特点的影响, 叶片P浓度表现出逐渐增加的趋势, 并在低、中N:P水平下表现显著, 表明中、高P添加使叶片P摄取增强, 与以往研究结果(Crous et al., 2008; Mayor et al., 2014)相似.由于供试土壤属于轻度碱化土壤, 土壤中的P易与游离态CaCO3结合形成难溶性的磷酸钙盐(Cross & Schlesinger, 2001), 同时, 10.0 g·m-2·a-1的N添加加剧了供试土壤P受限性, 少量P添加可能不足以消除叶片P限制(Fritz et al., 2012), 因而此时叶片P浓度的变化幅度较小(图2C).高N:P施入水平下叶片N浓度的变化幅度也很小, N10P32的N:P施入水平使叶片N浓度下降至 26.4 mg·g-1, 表明低N:P施入水平限制了甘草N摄取, 导致甘草叶片N保持能力增强.关于植物根系C:N:P化学计量学特征, 有研究表明少量N和P共施提高了麦冬(Ophiopogon japonicus)根部N浓度、P浓度和N:P (陈伏生等, 2007).本研究发现较高水平的N:P施入也提高了甘草根系N和P浓度, 但降低了C:P和N:P (图3E、图3F), 说明少量P添加促进了根系对N和P的共同吸收, 但由于根系对P吸收的速度快于对N的吸收, 从而导致根系C:P和N:P下降. ...
Changes in a symbiotic, heterotrophic nitrogen fixation on leaf litter of Metrosideros polymorpha with long-term ecosystem development in Hawaii. 1 2000
Growth and foliar nutrient response to recent applications of phosphorus (P) and potassium (K) and to residual P and K fertiliser applied to the previous rotation of Pinus patula at Usutu, Swaziland. 1 2008
Belowground insights into nutrient limitation in northern hardwood forests. 2010
Simulated pollutant nitrogen deposition increases P demand and enhances root-surface phosphatase activities of three plant functional types in a calcareous grassland. 1 2004
Testing the growth-rate hypothesis in vascular plants with above- and below-ground biomass. 1 2012
... 著降低(图5B、图5C).受土壤P供给水平变化特点的影响, 叶片P浓度表现出逐渐增加的趋势, 并在低、中N:P水平下表现显著, 表明中、高P添加使叶片P摄取增强, 与以往研究结果(Crous et al., 2008; Mayor et al., 2014)相似.由于供试土壤属于轻度碱化土壤, 土壤中的P易与游离态CaCO3结合形成难溶性的磷酸钙盐(Cross & Schlesinger, 2001), 同时, 10.0 g·m-2·a-1的N添加加剧了供试土壤P受限性, 少量P添加可能不足以消除叶片P限制(Fritz et al., 2012), 因而此时叶片P浓度的变化幅度较小(图2C).高N:P施入水平下叶片N浓度的变化幅度也很小, N10P32的N:P施入水平使叶片N浓度下降至 26.4 mg·g-1, 表明低N:P施入水平限制了甘草N摄取, 导致甘草叶片N保持能力增强.关于植物根系C:N:P化学计量学特征, 有研究表明少量N和P共施提高了麦冬(Ophiopogon japonicus)根部N浓度、P浓度和N:P (陈伏生等, 2007).本研究发现较高水平的N:P施入也提高了甘草根系N和P浓度, 但降低了C:P和N:P (图3E、图3F), 说明少量P添加促进了根系对N和P的共同吸收, 但由于根系对P吸收的速度快于对N的吸收, 从而导致根系C:P和N:P下降. ...
Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. 1 2015
... 大气氮(N)沉降增加, 是人类面临的一个全球性气候变化问题(IPCC, 2013), 其生态学效应受到越来越多科研工作者的关注.一般而言, 在受N限制的生态系统, 短期或少量N沉降增加可以刺激N矿化(Jones & Power, 2012)、促进微生物-酶系统对土壤有机质和养分的分解(Liu et al., 2010), 从而提高土壤N和磷(P)的有效性.随着长期或过多N沉降, 不仅生态系统N输入增加, 而且N:P失衡、系统N饱和甚至P压力增加(Bobbink et al., 2010; Wardle et al., 2013), 以及植物生长P受限性增强(Yu et al., 2012).在石灰性草原生态系统, 研究人员发现N添加显著提高了土壤磷酸单酯酶活性, 而土壤磷酸单酯酶活性与土壤有效P含量显著负相关, 因此认为长期N沉降增加导致该生态系统P受限性增加; 随着过多N输入, 植物根系和微生物通过分泌较多的磷酸酶提高植株对土壤P元素的吸收效率, 从而保证植物的正常生长(Phoenix et al., 2004).在其他一些生态系统, 如热带森林生态系统(Zhu et al., 2013)、温带森林生态系统(Naples & Fisk, 2010)和沼泽生态系统(Phuyal et al., 2008), 研究人员也报道了类似的结果.由于P不仅是细胞间能量传递和核苷酸结构组成的重要元素, 而且是许多自然生态系统植物生长的限制因素, 因此P受限性的增加可能会影响到植物P策略和生态系统P循环, 从而间接地改变生态系统结构和功能(Bobbink et al., 2010). ...
华北落叶松根茎叶碳氮磷含量及其化学计量学特征的季节变化 1 2014
... 大气氮(N)沉降增加, 是人类面临的一个全球性气候变化问题(IPCC, 2013), 其生态学效应受到越来越多科研工作者的关注.一般而言, 在受N限制的生态系统, 短期或少量N沉降增加可以刺激N矿化(Jones & Power, 2012)、促进微生物-酶系统对土壤有机质和养分的分解(Liu et al., 2010), 从而提高土壤N和磷(P)的有效性.随着长期或过多N沉降, 不仅生态系统N输入增加, 而且N:P失衡、系统N饱和甚至P压力增加(Bobbink et al., 2010; Wardle et al., 2013), 以及植物生长P受限性增强(Yu et al., 2012).在石灰性草原生态系统, 研究人员发现N添加显著提高了土壤磷酸单酯酶活性, 而土壤磷酸单酯酶活性与土壤有效P含量显著负相关, 因此认为长期N沉降增加导致该生态系统P受限性增加; 随着过多N输入, 植物根系和微生物通过分泌较多的磷酸酶提高植株对土壤P元素的吸收效率, 从而保证植物的正常生长(Phoenix et al., 2004).在其他一些生态系统, 如热带森林生态系统(Zhu et al., 2013)、温带森林生态系统(Naples & Fisk, 2010)和沼泽生态系统(Phuyal et al., 2008), 研究人员也报道了类似的结果.由于P不仅是细胞间能量传递和核苷酸结构组成的重要元素, 而且是许多自然生态系统植物生长的限制因素, 因此P受限性的增加可能会影响到植物P策略和生态系统P循环, 从而间接地改变生态系统结构和功能(Bobbink et al., 2010). ...
Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions. 2 2013
... 大气氮(N)沉降增加, 是人类面临的一个全球性气候变化问题(IPCC, 2013), 其生态学效应受到越来越多科研工作者的关注.一般而言, 在受N限制的生态系统, 短期或少量N沉降增加可以刺激N矿化(Jones & Power, 2012)、促进微生物-酶系统对土壤有机质和养分的分解(Liu et al., 2010), 从而提高土壤N和磷(P)的有效性.随着长期或过多N沉降, 不仅生态系统N输入增加, 而且N:P失衡、系统N饱和甚至P压力增加(Bobbink et al., 2010; Wardle et al., 2013), 以及植物生长P受限性增强(Yu et al., 2012).在石灰性草原生态系统, 研究人员发现N添加显著提高了土壤磷酸单酯酶活性, 而土壤磷酸单酯酶活性与土壤有效P含量显著负相关, 因此认为长期N沉降增加导致该生态系统P受限性增加; 随着过多N输入, 植物根系和微生物通过分泌较多的磷酸酶提高植株对土壤P元素的吸收效率, 从而保证植物的正常生长(Phoenix et al., 2004).在其他一些生态系统, 如热带森林生态系统(Zhu et al., 2013)、温带森林生态系统(Naples & Fisk, 2010)和沼泽生态系统(Phuyal et al., 2008), 研究人员也报道了类似的结果.由于P不仅是细胞间能量传递和核苷酸结构组成的重要元素, 而且是许多自然生态系统植物生长的限制因素, 因此P受限性的增加可能会影响到植物P策略和生态系统P循环, 从而间接地改变生态系统结构和功能(Bobbink et al., 2010). ...
... 在森林和酸性泥炭沼泽等N:P较高的生态系统, 研究人员已证实长期N沉降增加会导致系统P限制增加(Phuyal et al., 2008; Zhu et al., 2013).对宁夏荒漠草原而言, 尽管已监测到的N沉降量低于其他生态系统(Liu et al., 2013), 但是其本身N沉降临界负荷低, 因此可能对长期低剂量N沉降增加比较敏感(段雷等, 2002).本研究结果表明, 随着N:P施入水平降低, 土壤C:P和N:P呈降低趋势, 从而缓解了系统P限制、增强了叶片P摄取、提高了叶片和根系C:P和N:P, 促进了甘草生长.因此, 适当的人为P添加可通过调节土壤和植物叶片C:N:P化学计量学特征, 缓解土壤和植物间P的供需压力, 并削弱长期大气N沉降增加对群落结构的负面影响, 研究结果可为全球变化背景下荒漠草原生态系统的适应性管理提供科学依据.考虑到甘草自身的固N特性, 今后进行田间试验验证时, 有必要通过长期的野外模拟试验在不同功能群物种间进行比较和探讨. ...
 , 余海龙
, 余海龙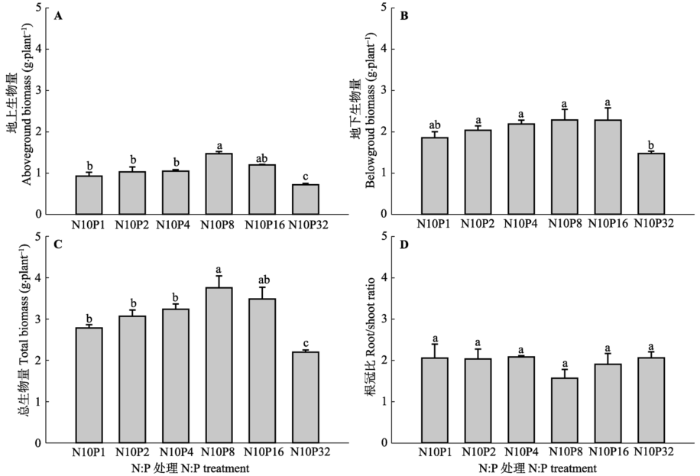 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT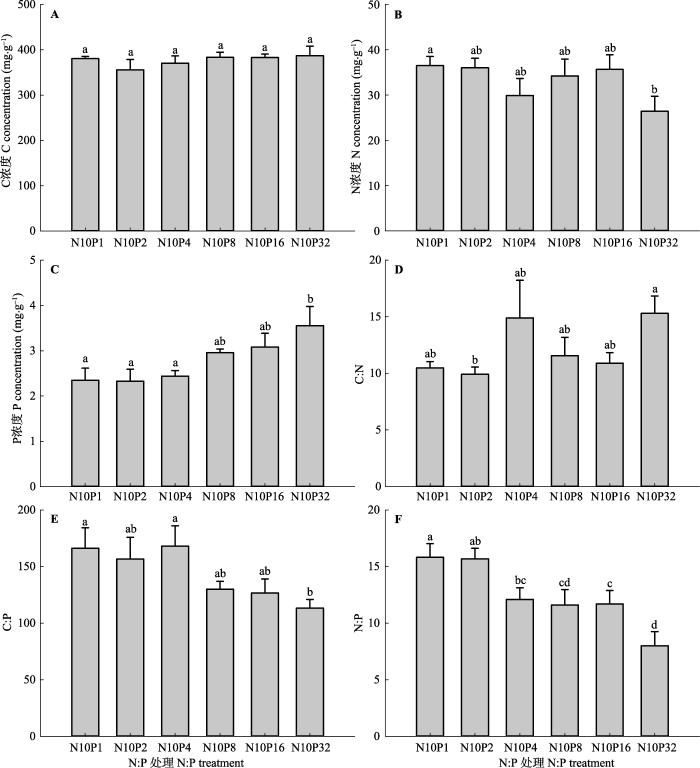 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT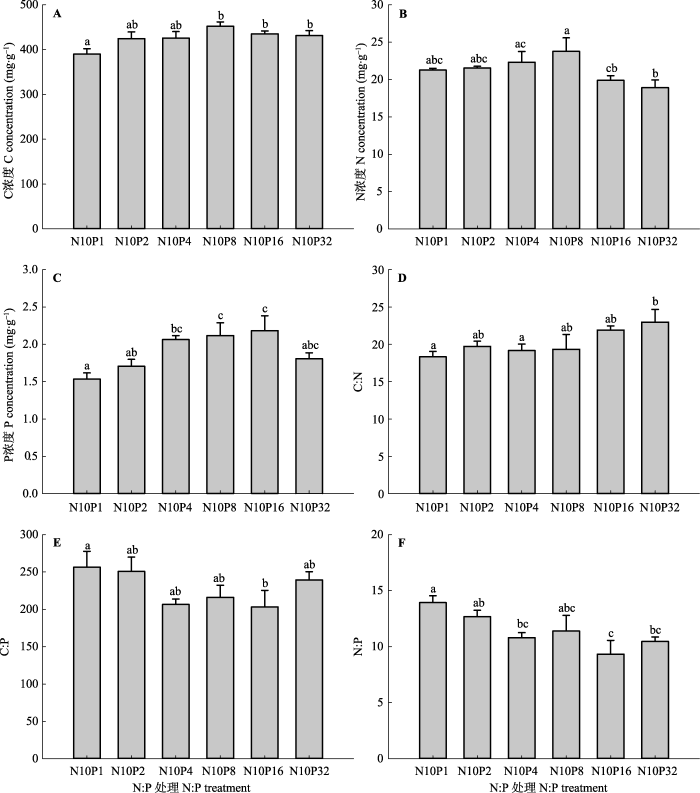 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT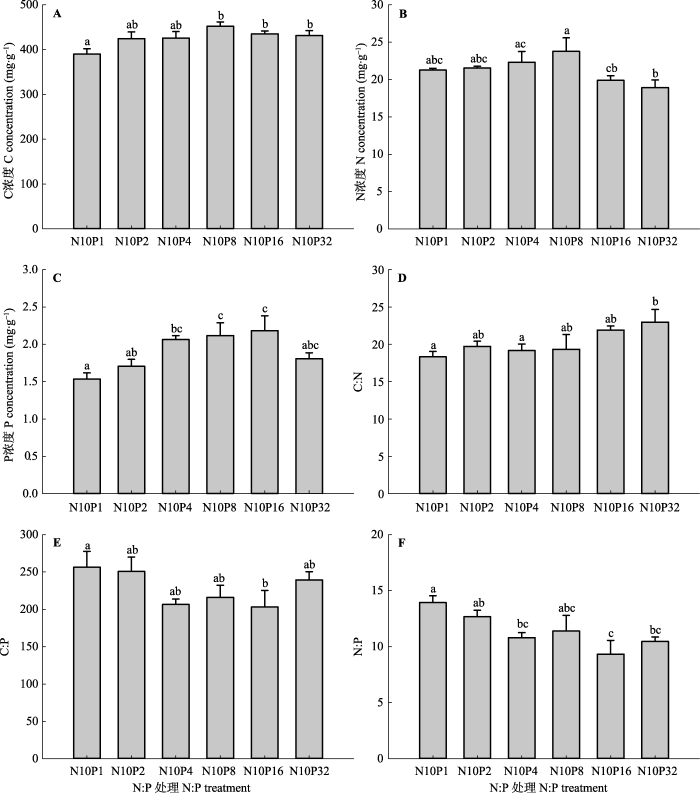 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT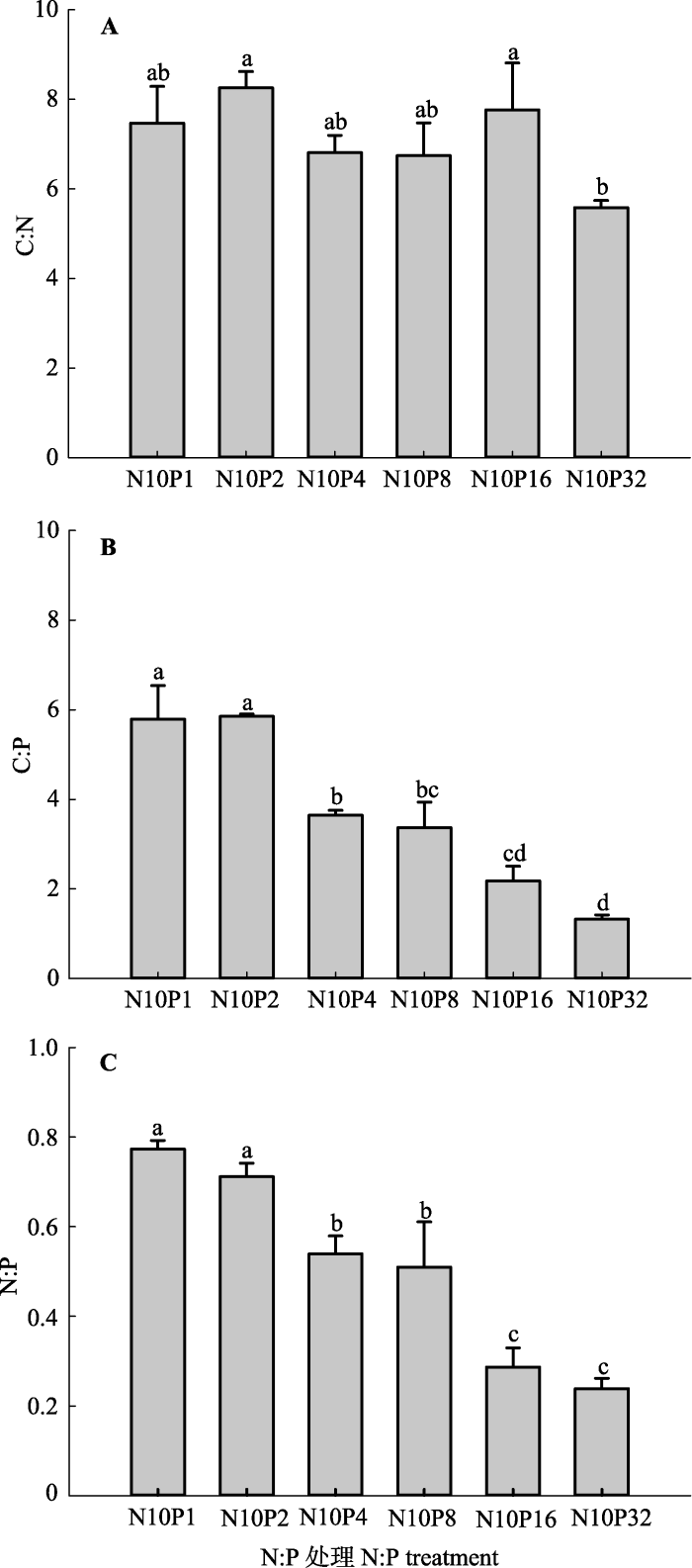 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}