Plant nutrient dynamics and stoichiometric homeostasis of invasive species Spartina alterniflora and native Cyperus malaccensis var. brevifolius in the Minjiang River estuarine wetlands
JIANGLi-Ling1, ZENGCong-Sheng2,3, SHAOJun-Jiong4, ZHOUXu-Hui1,*, 1Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, Institute of Biodiversity Science, Fudan University, Shanghai 200433, China2College of Geographical Science, Fujian Normal University, Fuzhou 350007, Chinaand 3Subtropical Wetland Research Center, Fujian Normal University, Fuzhou 350007, Chinaand 4School of Ecological and Environmental Sciences, East China Normal University, Shanghai 200062, China 通讯作者:* 通信作者Author for correspondence (E-mail: zxuhui14@163.com) 责任编辑:JIANGLi-LingZENGCong-ShengSHAOJun-JiongZHOUXu-Hui 收稿日期:2016-06-7 接受日期:2017-01-3 网络出版日期:2017-04-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家大学生创新项目(201210394011)、国家自然科学基金(31370489和31100352)、上海高校****(****)岗位计划和中共中央组织部青年****
关键词:生物入侵;养分动态;生态化学计量内稳性;生态系统功能;闽江河口湿地 Abstract Aims Stoichiometric homeostasis is an important mechanism in maintaining ecosystem structure, function, and stability. The invasion of exotic species, Spartina alterniflora, has largely threatened the structure and function of native ecosystems in the Minjiang River estuarine wetland. However, how S. alterniflora invasion affect plant stoichiometric homeostasis is largely unknown. This could enhance our understanding on wetland ecosystem stability and expand the applications of ecological stoichiometry theory. Methods Nitrogen (N) and phosphorus (P) contents of plant organs and soils in the S. alterniflora, Cyperus malaccensis var. brevifolius, and S. alterniflora-C. malaccensis var. brevifolius mixture were measured, and the homeostatic index (H) was calculated according to the stoichiometric homeostasis theory. Important findings Our results showed that the invasion of S. alterniflora significantly increased soil N:P ratio (p < 0.05), but did not affect soil N or P contents. The N and P contents of leaf and stem were the highest for S. alterniflora, and those of the stem were the highest for C. malaccensis var. brevifolius. At the ecosystem level, the average of homeostatic index (H) of N (HN, 25.31) was larger than those of P (HP, 10.33) and N:P (HN:P, 2.50). At the organ level, root HN was significantly larger than stem HN (p < 0.05) and sheath HN:P was greater than root HN:P (p < 0.05), while there was no significant difference for HP among root, stem, leaf, and sheath (p > 0.05). As for species, root HN of S. alterniflora was significantly larger than that of C. malaccensis var. brevifolius in the mixture community (p < 0.05). In the monoculture, stem HN:P of S. alterniflora was significantly higher than that of C. malaccensis var. brevifolius (p < 0.05). Furthermore, root HN, leaf HN and sheath HN of S. alterniflora in the mixed community was significantly larger than that of S. alterniflora in the monoculture (p < 0.05), suggesting that S. alterniflora invasions increased their stoichiometric homeostasis. Meanwhile, the stoichiometric homeostasis of invasive and native plants were influenced by multiple factors, such as nutrients, organs, vegetation, and invasion. However, larger homeostasis was found in S. alterniflora than in C. malaccensis var. brevifolius in some particular organs either in mixture or monoculture communities. Therefore, the successful invasion of S. alterniflora may result from higher homeostatic index than the native species, C. malaccensis var. brevifolius.
Keywords:biological invasion;nutrient dynamics;ecological stoichiometric homeostasis;ecosystem function;Minjiang River estuary wetland -->0 PDF (1427KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 蒋利玲, 曾从盛, 邵钧炯, 周旭辉. 闽江河口入侵种互花米草和本地种短叶茳芏的养分动态及植物化学计量内稳性特征. 植物生态学报, 2017, 41(4): 450-460 https://doi.org/10.17521/cjpe.2016.0193 JIANGLi-Ling, ZENGCong-Sheng, SHAOJun-Jiong, ZHOUXu-Hui. Plant nutrient dynamics and stoichiometric homeostasis of invasive species Spartina alterniflora and native Cyperus malaccensis var. brevifolius in the Minjiang River estuarine wetlands. Chinese Journal of Plant Ecology, 2017, 41(4): 450-460 https://doi.org/10.17521/cjpe.2016.0193 生物入侵作为全球变化的一个重要方面, 严重威胁了土著生物群落的生物多样性和生态系统功能(Davis, 2003; Hooper et al., 2005)。作为我国中亚热带和南亚热带过渡区最具代表性的河口湿地之一的闽江河口湿地(刘剑秋等, 2006), 自2002年以来逐渐遭到互花米草(Spartina alterniflora)的大规模入侵。互花米草以其极强的适应力和繁殖力迅速占据空生态位, 侵占本地种短叶茳芏(Cyperus malaccensis var. brevifolius)的生境, 在潮间带形成了短叶茳芏单种群落、互花米草单种群落和互花米草-短叶茳芏混生群落共存的竞争格局。入侵的互花米草通过影响土著生态系统的结构, 进而影响生态系统的养分循环, 生态系统养分的可利用性改变反过来又影响植物的生长状况, 从而可能改变入侵种与土著种之间的竞争关系。养分元素氮(N)和磷(P)是湿地生态系统植物生长的主要限制资源, 其可利用性及生态化学计量关系是植物生长和生态系统物质循环的重要调节因子(熊汉锋和王运华, 2005)。植物能在外界养分元素组成改变的情况下, 一定程度上维持自身各元素之间的相对比例。这种由生物在长期进化过程中为适应外界环境变化而形成的维持自身化学组成相对稳定的能力就叫作生态化学计量内稳性(简称内稳性)(Kooijman, 1995)。 国外内稳性研究的对象由最初的真菌和细菌(Levi & Cowling, 1969; Goldman et al., 1987)发展到藻类、浮游动物和草本植物(Rhee, 1978; Andersen & Hessen, 1991; Güsewell et al., 2005)。由于高等植物体内元素的内稳性和化学计量特征复杂, 对高等植物的研究甚少(Cernusak et al., 2009; Matzek & Vitousek, 2009; Yu et al., 2010)。生态系统水平上对内稳性的研究多集中于海洋生态系统和陆地生态系统(Güsewell, 2004; 苏强, 2012), 对湿地生态系统的研究较少(Wang et al., 2014)。国内内稳性的研究还处于起步阶段。庾强(2009)以内蒙古草原为研究对象, 通过1 200 km的横断范围的样带试验和内蒙古草原生态系统定位研究站27年的监测数据发现, 内稳性高的物种具有较高的优势度和生物量稳定性, 内稳性高的生态系统则具有较高的生产力和稳定性。因此, 内稳性是衡量物种竞争力的一个重要指标。已有研究表明内稳性的高低受多种因子的影响, 其中包括植物器官(庾强, 2009)、养分供应(严正兵等, 2013)、光强以及施肥(Sterner & Elser, 2002)等, 影响因子的不确定性使得不同研究中内稳性对环境的响应不一致。目前有关互花米草入侵对本地生态系统的影响多集中在物质循环以及生态化学计量学, 而物种水平上内稳性的研究甚少(张仁懿等, 2015)。已有研究发现, 闽江河口湿地生长季阶段的入侵种互花米草和本地种短叶茳芏均具有一定的内稳性, 互花米草N和N:P的内稳性高于短叶茳芏N和N:P的内稳性, 而短叶茳芏P的内稳性高于互花米草P的内稳性(蒋利玲等, 2014)。 为了进一步研究湿地生态系统植物内稳性与生态系统功能的关系, 我们以互花米草入侵为切入点, 以短叶茳芏单种群落、互花米草单种群落以及互花米草-短叶茳芏混生群落为研究对象, 对其土壤和植物的养分动态以及植物的内稳性进行了研究, 旨在解决以下问题: 1)入侵过程如何影响外来种和本地种的限制性养分以及植物内稳性?2)元素类型、器官类型和物种类型等不同因子对物种植物内稳性有什么影响?
1 材料和方法
1.1 试验设计
本试验在福建省闽江河口鳝鱼滩湿地(26.01°- 26.06° N, 119.57°-119.68° E, 图1)进行。该湿地为闽江河口区面积最大的砂泥质洲滩天然湿地。该区域气候暖热湿润, 年平均气温为19.3 ℃, 年降水量为 1 346 mm。土壤以滨海盐土、潮土以及水稻土等人工土壤为主。潮汐为正规半日潮。湿地内有维管植物109科294属408种(含变种), 本地优势种主要有短叶茳芏、芦苇(Phragmites australis)和藨草(Scirpus triqueter)。自2002年互花米草入侵以来, 其入侵斑块由零星分布发展到超过200万m2的大面积分布, 已在潮间带和部分潮上带形成了明显的大面积入侵斑块(郑彩红等, 2006)。 显示原图|下载原图ZIP|生成PPT 图1采样点位置图。 -->Fig. 1Location of the sampling site. -->
选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样。各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复。在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤。土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009), 土壤有效磷的测定采用NaHCO3浸提-钼锑抗比色法①(① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005)。
互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地土壤N、P含量均在4月份达到峰值, 表现出先增后减的变化趋势, 且N、P含量之间具有显著的相关性(p < 0.05或p < 0.01); 此外, 3种群落土壤N:P值随季节变化的规律不明显(图2)。互花米草单种群落、短叶茳芏单种群落及其混生群落土壤N含量平均值大小分别为33.91、33.21和31.54 mg·kg-1 (F2,61 = 0.064, p > 0.05), 土壤P含量平均值大小分别为2.78、3.15和2.39 mg·kg-1 (F2,62 = 1.131, p > 0.05), 土壤N:P值平均值大小分别为7.68、7.00和9.48, 且混生群落N:P值显著高于短叶茳芏单种群落N:P值(F2,61 = 2.302, p < 0.05)。 显示原图|下载原图ZIP|生成PPT 图23种群落类型植物器官及土壤的N、P含量和N:P值动态变化(平均值±标准误差, n = 4)。CM, 短叶茳芏单种群落; CMMC, 混生群落中的短叶茳芏; MC, 互花米草-短叶茳芏混生群落; SA, 互花米草单种群落; SAMC, 混生群落中的互花米草。 -->Fig. 2Dynamic of N, P and N:P in plant organs and soils across the three community types (mean ± SE, n = 4). CM, Cyperus malaccensis var. brevifolius community; CMMC, C. malaccensis var. brevifolius in mixed community; MC, S. alterniflora-C. malaccensis var. brevifolius mixture community; SA, Spartina alterniflora community; SAMC, S. alterniflora in mixed community. -->
互花米草和短叶茳芏植物器官中的N、P含量和N:P值对土壤的响应模式不同(图2)。N元素方面, 单种群落和混生群落中的互花米草叶N含量均显著高于对应单种群落和混生群落中的互花米草根、茎和鞘N含量(F3,80 = 20.207, p < 0.05), 单种群落中的短叶茳芏茎N含量显著高于对应单种群落中的短叶茳芏根N含量(F3,78 = 5.062, p < 0.05); 在混生群落中, 短叶茳芏根N含量显著高于对应混生群落中的互花米草根N含量(p < 0.05); 在单种群落中, 短叶茳芏根N含量显著高于对应单种群落中的互花米草根N含量(F3,78 = 12.168, p < 0.05)。P元素方面, 单种群落和混生群落中的互花米草叶P含量均显著高于对应单种群落和混生群落中的互花米草根、茎和鞘P含量(F3,81 = 8.867, p < 0.05), 混生群落中的短叶茳芏茎P含量显著高于对应混生群落中的短叶茳芏根P含量(F3,69 = 3.169, p < 0.05); 在混生群落中, 短叶茳芏茎P含量显著高于对应混生群落中的互花米草茎P含量(p < 0.05); 在单种群落中, 短叶茳芏茎P含量显著高于对应单种群落中的互花米草茎P含量(F3,74 = 3.265, p < 0.05)。植物器官间的N:P值无显著差异(F11,227 = 0.593, p > 0.05)。混生群落中的互花米草和短叶茳芏N:P值范围分别为7.54-30.71和6.26- 19.54, 平均值分别为16.17和12.87; 单种群落中的互花米草和短叶茳芏N:P值范围分别为2.31-33.75和5.45-19.86, 平均值分别为14.64和12.19; 所有植物的根、茎、叶和鞘N:P值范围分别为4.84-25.15、2.31-33.75、9.45-30.71和6.49-28.09, 平均值分别为13.43、13.49、17.22和15.76。混生群落中的互花米草叶和鞘N:P值均显著高于单种群落中的互花米草根N:P值和单种群落中的短叶茳芏根N:P值, 显著 高于单种群落和混生群落中的短叶茳芏茎N:P值 (F11,227 = 2.358, p < 0.05)。
2.2 土壤与植物养分相关性分析
对土壤和植物器官之间养分相关性进行分析的结果(表1)表明: 单种群落和混生群落中的短叶茳芏茎N含量均与对应的短叶茳芏单种群落和短叶茳芏混生群落的土壤N含量显著正相关(p < 0.05); 单种群落中的互花米草茎N含量与对应互花米草单种群落土壤N含量极显著正相关(p < 0.01), 单种群落中的互花米草鞘N含量与对应互花米草单种群落土壤N含量显著正相关 (p < 0.05); 单种群落中的互花米草根和叶P含量与对应互花米草单种群落土壤P含量显著负相关(p < 0.05); 混生群落中的互花米草茎N和茎P含量均与对应混生群落土壤N和土壤P含量显著正相关(p < 0.05)。 Table 1 表1 表13种群落类型植物器官与土壤氮(N)、磷(P)含量的相关系数 Table 1The correlation coefficients between soil and plant organs in nitrogen (N) and phosphorus (P) concentration of three community types
群落类型中的植物 Plant in community type
土壤N含量 Soil N concentration
土壤P含量 Soil P concentration
根N Root N
茎N Stem N
叶N Leaf N
鞘N Sheath N
根P Root P
茎P Stem P
叶P Leaf P
鞘P Sheath P
SAMC
-0.177
0.503*
0.023
0.297
-0.099
0.443*
0.087
0.156
SA
0.290
0.701**
-0.032
0.537*
-0.598**
0.182
-0.544*
-0.287
CMMC
0.195
0.653**
-
-
0.051
0.332
-
-
CM
0.433
0.538*
-
-
0.073
-0.079
-
-
*, p < 0.05; **, p < 0.01。The abbreviations are the same as in Fig. 2.*表示在p < 0.05水平下显著相关; **表示在p < 0.01水平下显著相关。缩写见图2。 新窗口打开 单种群落中的互花米草茎N含量与其土壤N含量的显著相关程度大于对应单种群落中的短叶茳芏茎N含量与其土壤N含量的相关程度; 混生群落中的短叶茳芏茎N含量与其土壤N含量的显著相关程度大于对应混生群落中的互花米草茎N含量与其土壤N含量的相关程度。互花米草入侵后, 单种群落中的互花米草鞘N含量与其土壤N含量间的相关性由显著正相关转变为混生群落中的不显著正相关。同时, 单种群落中的互花米草根P含量与其土壤P含量的相关性由显著负相关转变为混生群落中的不显著正相关; 单种群落中的互花米草茎P含量与其土壤P含量的相关性由不显著正相关转变为混生群落中的显著正相关; 单种群落中的互花米草叶P含量与其土壤P含量的相关性由显著负相关转变为混生群落中的不显著正相关。相比互花米草, 单种群落和混生群落中的短叶茳芏茎N含量与其土壤N含量的相关性均显著, 且混生群落中的相关程度较大;单种群落中的短叶茳芏茎P含量与其土壤P含量的相关性由负相关转变为混生群落中的正相关, 且两者的相关关系均不显著。
2.3 植物化学计量内稳性特征
不同元素的内稳性不同。该湿地生态系统N、P和N:P值的内稳性指数(HN、HP和HN:P)平均值分别为25.31、2.5和10.33, 其中HN显著高于HP (F2,35 = 2.632, p < 0.05)(图3A)。根据内稳性大小的界定, HP为弱稳态型, HN和HN:P为稳态型。对互花米草而言, 混生群落中的互花米草根和叶HN显著高于对应混生群落中的互花米草根和叶HP以及根和叶HN:P, 混生群落中的互花米草鞘HN和鞘HN:P显著高于对应混生群落中的互花米草鞘HP (p < 0.05); 单种群落中的互花米草茎HN:P显著高于对应单种群落中的互花米草茎HN和茎HP, 单种群落中的互花米草鞘HN显著高于对应单种群落中的互花米草鞘HP (p < 0.05)(图4)。 对短叶茳芏而言, 无论单种群落还是混生群落, 短叶茳芏HN、HP和HN:P之间均无显著差异(p > 0.05) (图4)。 显示原图|下载原图ZIP|生成PPT 图3不同元素、器官以及植物类型的内稳性指数(H)(平均值±标准误差, n = 4)。不同小写字母表示不同因子之间差异显著。缩写见图2。 -->Fig. 3The homeostatic index (H) in different element, organ and plant types (mean ± SE, n = 4). Different lowercase letters denote significant differences among different factors. The abbreviations are the same as in Fig. 2. -->
显示原图|下载原图ZIP|生成PPT 图43种群落类型植物器官的N、P和N:P值的内稳性指数(H)(平均值±标准误差, n = 4)。不同小写字母表示不同因子之间差异显著。缩写见图2。 -->Fig. 4The homeostatic index (H) of N, P and N:P in plant organs across the three community types (mean ± SE, n = 4). Different lowercase letters denote significant differences among different factors. The abbreviations are the same as in Fig. 2. -->
植物内稳性的大小在不同器官中存在差异。生态系统根、茎、叶和鞘的内稳性指数平均值分别为13.37、5.55、15.75和22.7 (F3,35 = 0.614, p > 0.05) (图3B)。对互花米草而言, 对比混生群落中的互花米草器官间的HN大小发现, 根HN > 叶HN > 鞘HN > 茎HN, 各器官间差异显著(p < 0.05); 混生群落中的互花米草鞘HN:P显著高于对应混生群落中的互花米草根、茎和叶HN:P; 单种群落中的互花米草鞘HN显著高于对应单种群落中的互花米草根、茎和叶HN, 单种群落中的互花米草茎HN:P显著高于对应单种群落中的互花米草根HN:P和叶HN:P (图4)。对短叶茳芏而言, 无论单种群落还是混生群落, 植物器官间的内稳性均无显著差异(p > 0.05)(图4)。所有植物根、茎、叶和鞘HN分别为35.22、3.24、42.38和32.56, 且根HN显著高于茎HN (F3,11 = 2.566, p < 0.05); 器官间HP差异不显著(F3,11 = 1.577, p > 0.05), 鞘HN:P显著高于根HN:P (F3,11 = 2.256, p < 0.05)(图5A)。 显示原图|下载原图ZIP|生成PPT 图5不同器官和植物类型N、P和N:P值的内稳性指数(H)(平均值±标准误差, n = 4)。不同小写字母表示不同因子之间差异显著。缩写见图2。 -->Fig. 5The homeostatic index (H) for different organs and plants in N, P and N:P (mean ± SE, n = 4). Different lowercase letters denote significant differences among different factors. The abbreviations are the same as in Fig. 2. -->
AndersenT, HessenDO (1991). Carbon, nitrogen, and phosphorus content of freshwater zooplankton .Limnology and Oceanography, 36, 807-814. [本文引用: 1]
[2]
BagwellCE, LovellCR (2000). Microdiversity of culturable diazotrophs from the rhizoplanes of the salt marsh grasses Spartina alterniflora and Juncus roemerianus . Microbial Ecology, 39, 128-136. [本文引用: 1]
BottT, MeyerGA, YoungEB (2008). Nutrient limitation and morphological plasticity of the carnivorous pitcher plant Sarracenia purpurea in contrasting wetland environments .New Phytologist, 180, 631-641. [本文引用: 1]
[5]
CernusakLA, WinterK, TurnerBL (2009). Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physiological and environmental controls .New Phytologist, 185, 770-779. [本文引用: 1]
[6]
DavisMA (2003). Biotic globalization: Does competition from introduced species threaten biodiversity? Bioscience, 53, 481-489. [本文引用: 1]
[7]
ElserJJ, FaganWF, KerkhoffAJ, SwensonNG, EnquistBJ (2010). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change .New Phytologist, 186, 593-608. [本文引用: 1]
[8]
FanXY (2012). Spatial Variation in Nutrient of Dominant Plant and Ecological Stoichiometry from Laohu Gou of Qilian Mountain . Master degree dissertation, Lanzhou University, Lanzhou. (in Chinese with English abstract)[樊晓勇 (2012). 祁连山老虎沟优势植物的养分空间变化与生态化学计量学研究 . 硕士学位论文, 兰州大学, 兰州.] [本文引用: 1]
[9]
FridCLJ, ChandrasekaraWU, DaveyP (1999). The restoration of mud flats invaded by common cord-grass (Spartina anglica, CE Hubbard) using mechanical disturbance and its effects on the macrobenthic fauna . Aquatic Conservation Marine & Freshwater Ecosystems, 9, 47-61. [本文引用: 1]
[10]
GarnierE (1998). Interspecific Variation in Plasticity of Grasses in Response to Nitrogen Supply . Cambridge University Press, Cambridge, UK. 155-181. [本文引用: 1]
[11]
GoldmanJC, CaronDA, DennettMR (1987). Regulations of gross growth efficiency and ammonium regeneration in bacteria by substrate C:N ratio .Limnology and Oceanography, 32, 1239-1252. [本文引用: 1]
[12]
GrevstadFS, StrongDR, Garcia-RossiD, SwitzerRW, WeckerMS (2003). Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: Agent specificity and early results . Biological Control, 27, 32-42. [本文引用: 1]
[13]
GüsewellS (2004). N:P ratios in terrestrial plants: Variation and functional significance .New Phytologist, 164, 243-266. [本文引用: 2]
[14]
GüsewellS, JewellPL, EdwardsPJ (2005). Effects of heterogeneous habitat use by cattle on nutrient availability and litter decomposition in soils of an Alpine pasture .Plant and Soil, 268, 135-149. [本文引用: 1]
[15]
GüsewellS, KoerselmanW (2002). Variation in nitrogen and phosphorus concentrations of wetland plants .Perspectives in Plant Ecology Evolution and Systematics, 5, 37-61. [本文引用: 3]
[16]
HooperDU, ChapinFS, EwelJJ, HectorA, InchaustiP, LavorelS, LawtonJH, LodgeDM, LoreauM, NaeemS, SchmidB, SetäläH, SymstadAJ, VandermeerJ, WardleDA (2005). Effects of biodiversity on ecosystem functioning: A consensus of current knowledge .Ecological Monographs, 75, 3-35. [本文引用: 1]
[17]
HuWF, ZhangWL, ZhangLH, ChenXY, LinW, ZengCS, TongC (2014). Stoichiometric characteristics of nitrogen and phosphorus in major wetland vegetation of China . Chinese Journal of Plant Ecology, 38, 1041-1052. (in Chinese with English abstract)[胡伟芳, 章文龙, 张林海, 陈晓艳, 林伟, 曾从盛, 仝川 (2014). 中国主要湿地植被氮和磷生态化学计量学特征 . 植物生态学报, 38, 1041-1052.]
[18]
JiangLL, HeS, WuLF, YanYF, WengSF, LiuJ, WangWQ, ZengCS (2014). Characteristics of stoichiometric homeostasis of three plant species in wetlands in Minjiang estuary .Wetland Science, 3, 293-298. (in Chinese with English abstract)[蒋利玲, 何诗, 吴丽凤, 颜远烽, 翁少峰, 刘静, 王维奇, 曾从盛 (2014). 闽江河口湿地3种植物化学计量内稳性特征 . 湿地科学, 3, 293-298.] [本文引用: 2]
[19]
JiangW, LiXQ, JiangQ, HuangDK, ChengHG (2006). Kjeldahl method and the elemental analyzer method in measurement of total nitrogen in sediments: Comparison and its significance .Geochimica, 35, 221-226. (in Chinese with English abstract)[江伟, 李心清, 蒋倩, 黄代宽, 程红光 (2006). 凯氏蒸馏法和元素分析仪法测定沉积物中全氮含量的异同及其意义 . 地球化学, 35, 221-226.] [本文引用: 1]
[20]
JonasP, PatrickF, AkiraG, JamesMH, JayneJ, SatoshiK (2010). To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs .Oikos, 119, 741-751. [本文引用: 1]
[21]
KoerselmanW, MeulemanAMF (1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation .Journal of Applied Ecology, 33, 1441-1450. [本文引用: 1]
[22]
KooijmanSALM (1995). The stoichiometry of animal energetics .Journal of Theoretical Biology, 177, 139-149. [本文引用: 1]
[23]
LeviMP, CowlingEB (1969). Role of nitrogen in wood deterioration. VII. Physiological adaptation of wood-destroying and other fungi to substrates deficient in nitrogen .Phytopathology, 59, 460-468. [本文引用: 1]
[24]
LiHP, ZhangLQ (2007). Experimental study on physical controls of an exotic plant Spartina alterniflora in Shanghai .Journal of East China Normal University (Natural Science), 6, 44-55. (in Chinese with English abstract)[李贺鹏, 张利权 (2007). 外来植物互花米草的物理控制实验研究 . 华东师范大学学报(自然科学版), 6, 44-55.] [本文引用: 1]
[25]
LiYY, SunJH, YuCB, ChengX, ZhangFS, LiL (2009). Effects of nitrogen fertilization application and faba bean/maize intercropping on the spatial and temporal distribution of soil inorganic nitrogen .Plant Nutrition and Fertilizer Science, 15, 815-823. (in Chinese with English abstract)[李玉英, 孙建好, 余常兵, 程序, 张福锁, 李隆 (2009). 施氮量和蚕豆/玉米间作对土壤无机氮时空分布的影响 . 植物营养与肥料学报, 15, 815-823.] [本文引用: 1]
[26]
LiuJQ, ZengCS, ChenN (2006). Study on the Wetlands of Minjiang River Estuary. Science Press, Beijing. 21-47. (in Chinese)[刘剑秋, 曾从盛, 陈宁 (2006). 闽江河口湿地研究. 科学出版社, 北京. 21-47.] [本文引用: 1]
[27]
MatzekV, VitousekPM (2009). N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis .Ecology Letters, 12, 765-771. [本文引用: 1]
[28]
MoisanderPH, PiehlerMF, PaerlHW (2005). Diversity and activity of epiphytic nitrogen fixers on standing dead stems of the salt marsh grass Spartina alterniflora .Aquatic Microbial Ecology, 39, 271-279. [本文引用: 1]
[29]
PattenK (2002). Smooth cordgrass (Spartina alterniflora) control with Imazapyr .Weed Technology, 16, 826-832. [本文引用: 1]
[30]
RheeGY (1978). Effects of N:P atomic ratios and nitrate limitation on algal growth, cell composition and nitrate uptake .Limnology and Oceanography, 23, 10-25. [本文引用: 1]
[31]
SternerRW, ElserJJ (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press, New Jersey. 226-226. [本文引用: 2]
[32]
SuQ (2012). The framework of stoichiometry homeostasis in zooplankton elemental composition .Acta Ecologica Sinica, 32, 7213-7219. (in Chinese with English abstract)[苏强 (2012). 浮游动物化学计量学稳态性特征研究进展 . 生态学报, 32, 7213-7219.] [本文引用: 1]
[33]
TessierJT, RaynalDJ (2003). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation .Journal of Applied Ecology, 40, 523-534. [本文引用: 1]
[34]
TilmanD, ReichPB, KnopsJMH (2006). Biodiversity and ecosystem stability in a decade-long grassland experiment .Nature, 441, 629-632. [本文引用: 1]
[35]
WangH, LiHY, ZhangZJ, MuehlbauerJD, HeQ, XuXH, YueCL, JiangDQ (2014). Linking stoichiometric homeostasis of microorganisms with soil phosphorus dynamics in wetlands subjected to microcosm warming .PLOS ONE, 9, e85575-e85575. doi: 10.1371/journal.pone.0085575. [本文引用: 1]
[36]
WuMY, HackerS, AyresD, StrongDR (1999). Potential of Prokelisia spp. as biological control agents of english cordgrass, Spartina anglica .Biological Control, 16, 267-273. [本文引用: 1]
[37]
XiongHF, WangYH (2005). Advances in researches on biogeochemical circulation of C, N and P in wetlands .Chinese Journal of Soil Science, 36, 240-243. (in Chinese with English abstract)[熊汉锋, 王运华 (2005). 湿地碳氮磷的生物地球化学循环研究进展 . 土壤通报, 36, 240-243.] [本文引用: 1]
[38]
YanZB, KimNY, HanTS, FangJY, HanWX (2013). Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of Arabidopsis thaliana .Chinese Journal of Plant Ecology, 37, 551-557. (in Chinese with English abstract)[严正兵, 金南瑛, 韩廷申, 方精云, 韩文轩 (2013). 氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响 . 植物生态学报, 37, 551-557.] [本文引用: 1]
[39]
YuQ (2009). Ecological Stoichiometric Study on Vascular Plants in the Inner Mongolia Steppe . PhD dissertation, Institute of Botany,Chinese Academy of Science, Beijing. 56-67. (in Chinese with English abstract)[庾强 (2009). 内蒙古草原植物化学计量生态学研究 . 博士学位论文, 中国科学院植物研究所, 北京. 56-67.] [本文引用: 1]
YuQ, WilcoxK, PierreKL, KnappAK, HanXG, SmithMD (2015). Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change .Ecology, 96, 2328-2335.
[42]
ZengY, TianGH, ChenLY, LiJ, AnD, LeiZS, TangH, PengSL (2011). Influence ofSpartina alterniflora invasion on soil ecosystem: A review .Chinese Journal of Ecology, 30, 2080-2087. (in Chinese with English abstract)[曾艳, 田广红, 陈蕾伊, 李静, 安东, 雷振胜, 唐虹, 彭少麟 (2011). 互花米草入侵对土壤生态系统的影响 . 生态学杂志, 30, 2080-2087.] [本文引用: 1]
[43]
ZhangLX, BaiYF, HanXG (2004). Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol .Acta Botanica Sinica, 46, 259-270. [本文引用: 1]
[44]
ZhangRY, ShiXM, LiWJ, GuoR, WangG (2015). Response of species homeostasis and biomass on a sub-alpine grassland .Pratacultural Science, 32, 1539-1547. (in Chinese with English abstract)[张仁懿, 史小明, 李文金, 郭睿, 王刚 (2015). 亚高寒草甸物种内稳性与生物量变化模式 . 草业科学, 32, 1539-1547.] [本文引用: 1]
[45]
ZhengCH, ZengCS, ChenZQ, LinMC (2006). A study on the changes of landscape pattern of estuary wetlands of the Minjiang River .Wetland Science, 4, 29-35. (in Chinese with English abstract)[郑彩红, 曾从盛, 陈志强, 林茂昌 (2006). 闽江河口区湿地景观格局演变研究 . 湿地科学, 4, 29-35.] [本文引用: 1]
Carbon, nitrogen, and phosphorus content of freshwater zooplankton 1 1991
... 选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样.各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复.在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤.土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009), 土壤有效磷的测定采用NaHCO3浸提-钼锑抗比色法①(① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005). ...
Nutrient limitation and morphological plasticity of the carnivorous pitcher plant Sarracenia purpurea in contrasting wetland environments 1 2008
The restoration of mud flats invaded by common cord-grass (Spartina anglica, CE Hubbard) using mechanical disturbance and its effects on the macrobenthic fauna 1 1999
... 目前, 互花米草的防治措施主要有物理控制(Frid et al., 1999)、化学控制(Patten, 2002)和生物控制(Wu et al., 1999, Grevstad et al., 2003)这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
Interspecific Variation in Plasticity of Grasses in Response to Nitrogen Supply 1 1998
Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: Agent specificity and early results 1 2003
... 目前, 互花米草的防治措施主要有物理控制(Frid et al., 1999)、化学控制(Patten, 2002)和生物控制(Wu et al., 1999, Grevstad et al., 2003)这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
N:P ratios in terrestrial plants: Variation and functional significance 2 2004
... 目前, 互花米草的防治措施主要有物理控制(Frid et al., 1999)、化学控制(Patten, 2002)和生物控制(Wu et al., 1999, Grevstad et al., 2003)这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
凯氏蒸馏法和元素分析仪法测定沉积物中全氮含量的异同及其意义 1 2006
... 选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样.各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复.在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤.土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009), 土壤有效磷的测定采用NaHCO3浸提-钼锑抗比色法①(① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005). ...
To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs 1 2010
... 其中, x为土壤无机N或有效P含量(mg·kg-1)或N:P值, y为植物体器官对应的全氮(N)或全磷(P)含量(mg·kg-1)或N:P值,c为常数, H为植物内稳性指数.H使用年数据进行拟合计算得到, 其中植物器官的内稳性为各器官在N、P以及N:P值水平上的拟合, 物种的内稳性为所有器官内稳性的平均值, 生态系统N、P以及N:P值的内稳性为该生态系统所有植物整体在相应N、P和N:P值水平上的拟合.H的类型可根据H > 4为稳态型、2 < H < 4为弱稳态型、4/3 < H < 2为弱敏感型, 以及H < 4/3为敏感型来界定(Jonas et al., 2010). ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation 1 1996
... 目前, 互花米草的防治措施主要有物理控制(Frid et al., 1999)、化学控制(Patten, 2002)和生物控制(Wu et al., 1999, Grevstad et al., 2003)这3种方法, 其中以收割、碎根水淹等物理方法居多.研究表明, 翻耕和碎根处理方法仅在当年有一定的控制效果, 不具有长期有效性, 且割除碎根也需要在最佳季节进行(李贺鹏和张利权, 2007).已有的研究认为: 闽江河口湿地生态系统在7月份时互花米草内稳性最低, 是治理互花米草的最佳时期(蒋利玲等, 2014).充分考虑植物内稳性特征的影响, 建议采用物理-化学综合方法进行互花米草的控制, 在7月份对互花米草进行碎根, 再通过施加N缓解短叶茳芏根和茎的养分限制, 从而增大短叶茳芏群落在湿地生态系统中的内稳性. ...
施氮量和蚕豆/玉米间作对土壤无机氮时空分布的影响 1 2009
... 选择潮间带生境的互花米草单种群落、互花米草-短叶茳芏混生群落和短叶茳芏单种群落沼泽湿地为研究对象, 对沼泽湿地的土壤和植物进行取样.各群落斑块面积约为30 m × 30 m, 在每个斑块的样方内随机选取4个面积大小为25 cm × 25 cm的小样方作为重复.在2012年4月至2013年2月, 每2个月采样一次, 采用齐地面收割方式获得植物的地上部分, 采用挖掘法获取0-30 cm深度的地下所有根系和土壤.土壤无机氮的测定采用KCl浸提-连续流动分析仪法(李玉英等, 2009), 土壤有效磷的测定采用NaHCO3浸提-钼锑抗比色法①(① Olsen SR, Cole CV, Watanabe FS, Dean LAC (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. US Department of Agriculture, Washington, DC, USA. 939.); 植物全氮的测定采用元素分析仪法(江伟等, 2006), 植物全磷的测定采用浓硫酸-高氯酸消煮-连续流动分析仪法(鲍士旦, 2005). ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT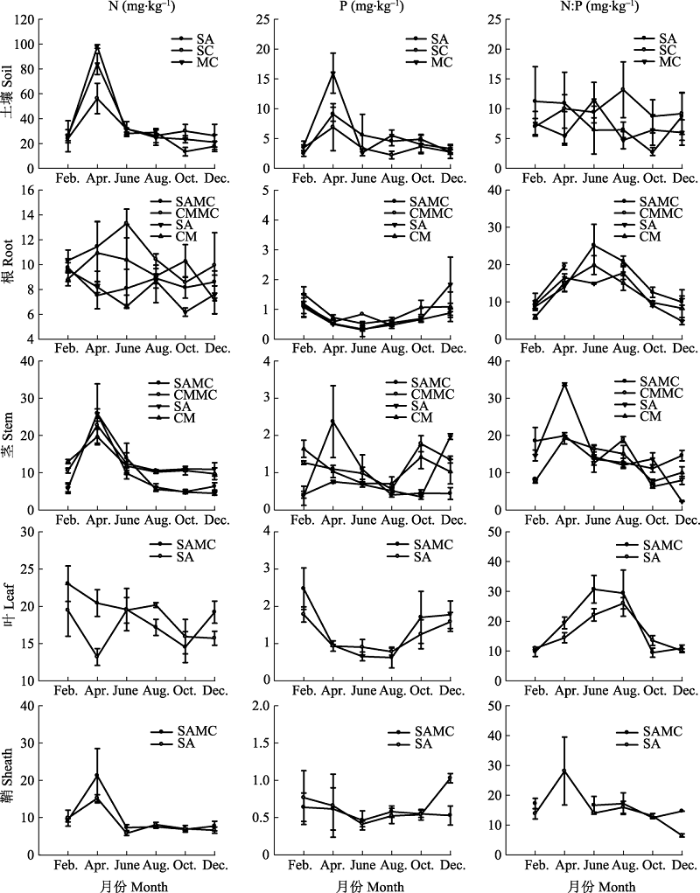 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT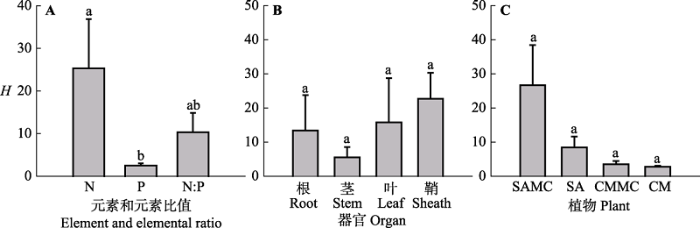 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT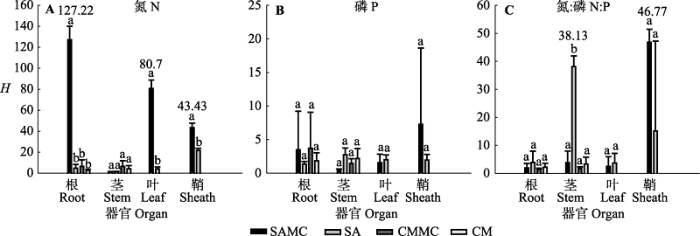 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT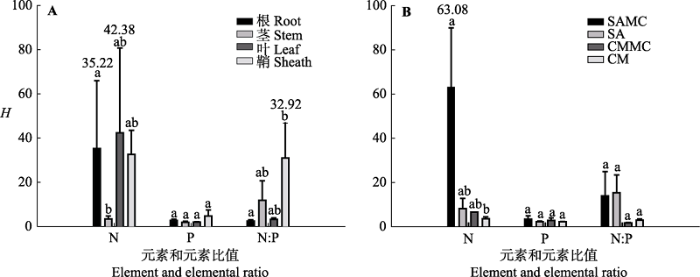 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}