C, N and P stoichiometry in different organs of Vitex rotundifolia in a Poyang Lake desertification hill
ZHOUHong-Yan1, WUQin1, CHENMing-Yue1, KUANGWei1, CHANGLing-Ling1, HUQi-Wu1,2,*, 1School of Geography and Environment, Jiangxi Normal University, Nanchang 330022, Chinaand 2Key Laboratory of Poyang Lake Wetland and Watershed Research, Ministry of Education (Jiangxi Normal University), Nanchang 330022, China 通讯作者:* 通信作者Author for correspondence (E-mail: huqiwu1979@gmail.com) 收稿日期:2016-07-5 接受日期:2017-02-14 网络出版日期:2017-04-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:江西省教育厅科技落地计划项目“鄱阳湖沙化土地与水土流失治理技术推广与示范”国家自然科学基金(31460129和31600371)
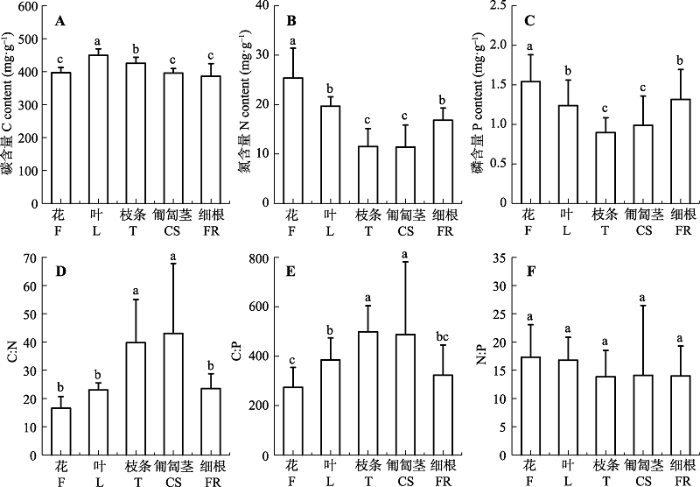
关键词:鄱阳湖;沙山;植物器官;养分;化学计量比 Abstract Aims The objectives were to clarify the responses of C, N and P stoichiometry of Vitex rotundifolia to desertification, and determine the C, N and P stoichiometric relationships among the organs. Methods In this study, different organs (e.g. flowers, leaves, twigs, creeping stems, fine roots) of V. rotundifolia were sampled along a desertification gradient in a typical Poyang Lak sandy hill. Subsequently, C, N and P contents of various organs were measured. Important findings The results showed nutrient contents in different organs ranged from 386.28 to 449.47 mg·g-1 for carbon, 11.40 to 25.37 mg·g-1 for nitrogen and 0.89 to 1.54 mg·g-1 for phosphorus, respectively. C, N and P contents differed significantly among the five organs. The maximum N and P content were found in flowers, whereas the minimums were observed in twigs and creping stems. Moreover, desertification intensity only significantly affected C, N and C:P. C:N and N:P ratios maintained relatively stable. Except N:P, the other nutrient elements and associated stoichiometry significantly differed among the organs. Hence, organs, rather than desertification intensity mainly controlled the C, N and P content and their stoichiometry variability. Although there was a positive correlation between mass-based N content (Nmass) and P content (Pmass) across the three desertification zones, the Nmass-Pmass relationship in V. rotundifolia did not shift. Irrespective desertification intensity and organs, N:P stoichiometry of V. rotundifolia was well constrained. In addition, significant correlations of C, N and P contents among organs were mainly found in the above-ground parts, especially between twigs and creeping stems.
Keywords:Poyang Lake;desertification hill;plant organ;nutrient;stoichiometry -->0 PDF (1334KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 周红艳, 吴琴, 陈明月, 匡伟, 常玲玲, 胡启武. 鄱阳湖沙山单叶蔓荆不同器官碳、氮、磷化学计量特征. 植物生态学报, 2017, 41(4): 461-470 https://doi.org/10.17521/cjpe.2016.0231 ZHOUHong-Yan, WUQin, CHENMing-Yue, KUANGWei, CHANGLing-Ling, HUQi-Wu. C, N and P stoichiometry in different organs of Vitex rotundifolia in a Poyang Lake desertification hill. Chinese Journal of Plant Ecology, 2017, 41(4): 461-470 https://doi.org/10.17521/cjpe.2016.0231 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用。C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005)。N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004)。植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011)。当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010)。近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究。然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010)。同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014)。 土地沙化长期以来一直是我国最为严重的生态环境问题(董光荣等, 1999), 不仅出现在我国北方干旱、半干旱地区, 亦出现于南方湖滨及河流下游冲积平原地区(朱震达和崔书红, 1996)。以鄱阳湖区为例, 沙化土地面积多达3.89万hm2 (左长青, 1986)。鄱阳湖沙化土地虽处于亚热带湿润区, 但水热不同期, 特别是8-10月的高温、少雨与地表强蒸发, 以及冬春季节固定的风向与强劲的风速是影响植被恢复的重要因素。近几十年来, 马鞭草科落叶小灌木单叶蔓荆(Vitex rotundifolia)被成功地应用于鄱阳湖沙化土地的植被恢复, 并且在不同沙化程度区形成稳定的群落。研究表明生物有机体具有极高的化学计量内稳性(Elser et al., 2010), 并且通过调节生物对环境变化的响应, 化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机制(Yu et al., 2010)。单叶蔓荆的养分及其化学计量关系在不同沙化程度区会发生怎样的变化?其N、P功能性状之间的关系是否因沙化梯度而发生策略位移?本研究通过测定鄱阳湖典型沙山重度、中度、轻度沙化区单叶蔓荆花、叶、枝条、匍匐茎、细根等部位的C、N、P含量, 分析各器官养分元素分配格局、化学计量比的差异与关联, 以及随沙化程度的变化, 试图探讨上述两个科学问题, 为进一步揭示单叶蔓荆的沙地适应机制提供科学依据, 为鄱阳湖沙化区植被恢复提供参考。
1 材料和方法
1.1 研究区概况
研究区设置在鄱阳湖沙化土地的典型分布区——江西省都昌县多宝乡鄱阳湖湖滨沙山, 地理位置29.36°-29.46° N, 116.05°-116.13° E。该区属于亚热带湿润性季风气候, 年降水量1β310 mm, 年蒸发量1β883 mm, 日照充足。多宝沙山主要是由冬季风吹蚀湖滨沙地堆积于近湖滨的一级阶地而形成的。依据野外实地调查, 参考沙丘分布形态、地表裸露程度、植被覆盖度、土壤有机质、氮磷养分等指标, 研究区沙山可以划分为3个沙化梯度, 其中湖滨 800 m范围内为重度沙化区, 800-2β000 m为中度沙化区, 2β000 m之外为轻度沙化区。重度沙化区主要由流动沙丘、半流动沙丘组成, 呈面状、条带状分布;中度沙化区地表裸露呈斑块状, 主要为半固定沙丘;轻度沙化区以固定沙丘为主, 地表裸露呈斑点状零散分布。3种不同沙化程度区土壤(0-10 cm)、植被基本特征见表1。 Table 1 表1 表1不同沙化程度区土壤植被特征(平均值±标准偏差) Table 1Characteristics of soil and vegetation in different desertification zones (mean ± SD)
Different lowercase letters indicate significant difference between sampling sites at p < 0.05 levels. Coverage values range from 0 to 1. SOC, TN and TP were abbreviation of soil organic carbon, total nitrogen and total phosphorus, respectively. HD, MD and LD indicate high, moderate and low desertification, respectively.表中不同小写字母表示采样区间差异显著(p < 0.05)。盖度范围为0-1。 新窗口打开
单叶蔓荆各器官C含量随着沙化程度的变化发生较大的波动, 在5种器官中, 除了匍匐茎表现出显著的下降趋势外, 其他器官的波动没有表现出一致的规律(图2A)。多数器官N含量随着沙化程度的降低表现出增加趋势, 且最低值多出现于湖岸线边缘500 m范围内的重度沙化区, N含量的高值则多分布于离湖岸线2β000 m之外的轻度沙化区。匍匐茎N含量与其他器官相比表现出下降的趋势(图2B)。各器官中P含量随沙化程度的变化没有明显规律, 变异系数范围为7.4%-28.9% (图2C)。 显示原图|下载原图ZIP|生成PPT 图2单叶蔓荆各器官碳(C)、氮(N)、磷(P)含量随沙化程度变化(平均值±标准偏差)。HD、MD和LD同表1。 -->Fig. 2Changes of carbon(C), nitrogen(N) and phosphorus(P) contents in various organs of Vitex rotundifolia along desertification gradient (mean ± SD). HD, MD and LD see Table 1. -->
2.4 单叶蔓荆Nmass-Pmass关系沿沙化梯度的变化
Nmass-Pmass间呈极显著正相关关系(r = 0.49, p < 0.001, n = 160), 且重度、中度、轻度沙化区单叶蔓荆N、P含量之间均呈显著线性关系(p < 0.01)(图3)。Nmass-Pmass之间一元线性关系的斜率表现为轻度沙化区高于中度与重度沙化区, 截距则反之, 表现为重度沙化区高于中度与轻度沙化区。协方差分析表明3种沙化程度区Nmass-Pmass线性关系的斜率与截距均未达到显著性差异(表3)。 显示原图|下载原图ZIP|生成PPT 图3不同沙化程度区单叶蔓荆基于质量的氮含量(Nmass)和磷含量(Pmass)之间的关系。HD、MD和LD同表1。 -->Fig. 3Relationship between mass-based nitrogen content (Nmass) and mass-based phosphorus content (Pmass) of Vitex rotundifolia at different desertification zones. HD, MD and LD see Table 1. -->
Table 3 表3 表3不同沙化程度区单叶蔓荆基于质量的氮含量(Nmass)和磷含量(Pmass)之间线性关系的斜率与截距差异检验 Table 3A test of differences in slopes and intercepts of linear regressions of mass-based nitrogen content (Nmass) against mass-based phosphorus content (Pmass) of Vitex rotundifolia among different desertification zones using analysis of covariance
Nmass-Pmass (n = 160)
重度沙化区 HD zone
中度沙化区 MD zone
轻度沙化区 LD zone
F
p
斜率 Slope
5.2
8.3
10.1
1.13
0.33
截距 Intercept
9.3
7.5
5.8
2.66
0.073
HD, MD and LD represented high, moderate and low desertification zone, respectively. 新窗口打开
通过沙化程度和各器官对单叶蔓荆C、N、P含量及其化学计量关系影响的一般线性模型(GLM)分析发现: 沙化程度对单叶蔓荆的C、N含量影响显著, 对P含量的影响未达到显著性水平。相应的C:N和N:P受沙化程度的影响不显著, 但C:P受沙化程度影响显著。各器官这一因子除对N:P影响不显著外, 对其他元素及化学计量关系的影响都极为显著(p < 0.01)。沙化程度与各器官的交互作用对所有元素及其化学计量关系均有显著影响(表4)。 Table 4 表4 表4沙化程度和各器官对单叶蔓荆碳氮磷含量及其计量比影响的一般线性模型(GLM)分析 Table 4General linear model (GLM) analysis of effects of desertification intensity and organs on C, N and P contents of Vitex rotundifolia and their stoichiometry
自变量 Variable
因变量 Dependent variable
平方和 Sum of squares
自由度 d.f.
均方 Mean square
统计量 f
显著性 p
沙化程度 Desertification intensity
C
113.260
2
56.630
4.998
0.008*
N
3 454.735
2
1 727.368
3.717
0.027*
P
0.563
2
0.281
2.941
0.056
C:N
100.579
2
50.289
0.326
0.723
C:P
187 733.990
2
93 866.995
4.373
0.014*
N:P
240.789
2
120.394
2.773
0.066
器官 Organs
C
3 792.949
4
948.237
83.690
0.000**
N
78 810.635
4
19 702.659
42.392
0.000**
P
7.656
4
1.914
20.009
0.000**
C:N
15 152.136
4
3 788.034
24.534
0.000**
C:P
1 036 736.242
4
259 184.060
12.076
0.000**
N:P
333.015
4
83.254
1.918
0.111
沙化程度×器官 Desertification intensity × Organs
C
699.213
8
87.402
7.714
0.000**
N
9 851.142
8
1 231.393
2.649
0.010*
P
2.181
8
0.273
2.850
0.006*
C:N
5 414.247
8
676.781
4.393
0.000**
C:P
653 464.339
8
81 683.042
3.806
0.000**
N:P
1 302.409
8
162.801
3.750
0.001*
*, p < 0.05; **, p < 0.01。 新窗口打开 植物体营养元素含量及化学计量关系不仅受植物的进化史、生活史、遗传、不同器官的生理功能等因素影响, 同时还受到土壤营养元素的供给及其在植物体内的运输、储藏、利用等过程的影响。本研究中除了匍匐茎N含量与土壤0-10 cm全N含量呈显著负相关之外, 未发现单叶蔓荆其他器官与土壤表层0-10 cm、10-30 cm之间相应养分元素之间的相关关系(表5)。 Table 5 表5 表5单叶蔓荆各器官N、P含量与土壤相应元素的相关关系 Table 5The correlations of N, P contents between soil and various organs of Vitex rotundifolia
土层 Soil layer
养分元素 Nutrient element
花 Flower
叶 Leaf
枝条 Twig
匍匐茎 Creeping stem
细根 Fine root
0-10 cm
N
0.336
0.405
0.211
-0.628*
0.303
P
-0.337
0.370
0.184
0.091
0.493
10-30 cm
N
0.212
0.390
0.180
-0.587
0.253
P
-0.297
0.351
0.102
0.123
-0.409
*, p < 0.05; **, p < 0.01。 新窗口打开 上述GLM分析结果表明: 相对于沙化程度的变化所引起的植物生境条件, 如土壤水分、养分供给的变化, 不同器官的生理功能差异是影响单叶蔓荆C、N、P含量及化学计量比的主导因子。化学计量内稳性(stoichiometric homoeostasis)是生态化学计量学存在的基础(Sterner & Elser, 2002)。Elser等(2010)认为尽管C、N、P化学计量比具有较大的时空变异, 但生物有机体仍保持极高的化学计量内稳性。化学计量内稳性通过调节生物对环境因子的响应, 成为生态系统结构、功能和稳定性维持的重要机制(Yu et al., 2010)。本研究中3种元素的化学计量关系中C:N、N:P在不同沙化程度区均未表现出显著差异。这一结果表明单叶蔓荆能够在沙山不同沙化区形成稳定的群落并保持较高的优势度, 化学计量内稳性很可能是其中非常重要的维持机制, 但这一机制仍然有待于进一步进行植物养分与土壤养分, 特别是速效养分之间关系的动态监测来加以验证。以往研究表明植物叶片与细根具有相似的N:P化学计量比(Yuan et al., 2011), 同样, 本研究也发现N、P养分元素虽然在单叶蔓荆不同器官中的含量具有显著差异, 但是具有相似的N:P化学计量比, 保持了相对稳定性。
3.3 沙化程度对单叶蔓荆Nmass-Pmass关系的影响
由于降水、土壤养分等环境梯度而引起的植物功能性状之间关系的策略位移现象不仅出现在种间水平上(Wright et al., 2001; 李永华等, 2005; Wei et al., 2011), 也出现在种内水平上(Wei et al., 2011)。但这一现象是否具有普遍性目前仍不清楚, 例如, 胡梦瑶等(2012)并没有发现紫花针茅(Stipa purpurea)在不同降水梯度下具有明显的策略位移现象。此外, 上述功能性状的位移现象多出现于较大尺度下的环境梯度中, 局地尺度上是否也存在类似的现象仍缺乏研究。本研究区尽管重度、中度与轻度沙化区土壤有机质、全氮、全磷等养分, 以及地表植被覆盖度等存在显著差异(表1), 但未发现单叶蔓荆Nmass-Pmass线性关系的斜率与截距在3种沙化程度区发生显著变化(表3), 表明单叶蔓荆这一对功能性状关系在不同沙化区未发生明显的位移现象。成对植物功能性状关系发生策略位移主要是由于环境梯度对其中一个功能性状影响更大, 例如, Wei等(2011)发现低降水区的油蒿(Artemisia ordosica)种群Nmass高于较高降水区, 但比叶面积差异不大。本研究中Nmass-Pmass在不同沙化程度区未出现明显的策略位移现象, 表明土壤沙化所引起的水、肥等限制因素可能共同作用于这两个性状, 使得植物Nmass与Pmass发生同步变化, 同时这也进一步佐证了N:P化学计量关系在不同沙化区的相对稳定。此外, 本研究结果也表明局地尺度种内水平上很可能因为环境梯度过小而不具备发生Nmass-Pmass关系的位移现象。
4 结论
单叶蔓荆不同器官间C、N、P含量存在显著差异, 其中花的N、P含量显著高于其他器官。各器官中以枝条与匍匐茎之间的关系最为密切, 二者之间的C、N、P含量及C:N、C:P化学计量比均表现为显著性相关。沙化程度仅对C、N含量及C:P影响显著, 不同器官是影响单叶蔓荆C、N、P含量及化学计量比的主导因子。单叶蔓荆N:P化学计量比具有保守性特征, 没有随沙化程度与器官的改变而发生显著变化, Nmass-Pmass关系在不同沙化程度区未发生位移现象。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
CraineJM, LeeWG (2003). Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand .Oecologia, 134, 471-478. [本文引用: 1]
[4]
CuiGY, CaoY, ChenYM (2015). Characteristics of nitrogen and phosphorus stoichiometry across components of forest ecosystem in Shaanxi Province .Chinese Journal of Plant Ecology, 39, 1146-1155. (in Chinese with English abstract)[崔高阳, 曹扬, 陈云明 (2015). 陕西省森林各生态系统组分氮磷化学计量特征 . 植物生态学报, 39, 1146-1155.] [本文引用: 1]
[5]
DongGR, WuB, CiLJ, ZhouHS, LuQ, LuoB (1999). Present situation, causes and control way of desertification in China .Journal of Desert Research, 19(4), 22-36. (in Chinese with English abstract)[董光荣, 吴波, 慈龙骏, 周欢水, 卢琦, 罗斌 (1999). 我国荒漠化现状、成因与防治对策 . 中国沙漠, 19(4), 22-36.] [本文引用: 1]
ElserJJ, FaganWF, KerkhoffAJ, SwensonNG, EnquisBJ (2010). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change .New Phytologist, 186, 593-608. [本文引用: 3]
[8]
EvinerVT, Chapin FSIII (2003). Functional matrix: A conceptual framework for predicting multiple plant effects on ecosystem processes .Ecology, Evolution and Systematics, 34, 455-485. [本文引用: 1]
[9]
GengY, WangL, JinDM, LiuHY, HeJS (2014). Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits .Oecologia, 175, 445-455. [本文引用: 1]
[10]
GuoDL, LiH, MitchellRJ, HanWX, HendricksJJ, FaheyTJ, HendrickRL (2008). Heterogeneity by root branch order: Exploring the discrepancy in root longevity and turnover estimates between minirhizotron and C isotope methods .New Phytologist, 177, 443-456. [本文引用: 1]
[11]
GüsewellS (2004). N:P ratios in terrestrial plants: Variation and functional significance .New Phytologist, 164, 243-266. [本文引用: 1]
[12]
HeJS, HanXG (2010). Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems .Chinese Journal of Plant Ecology, 34, 2-6. (in Chinese with English abstract)[贺金生, 韩兴国 (2010). 生态化学计量学: 探索从个体到生态系统的统一化理论 . 植物生态学报, 34, 2-6.] [本文引用: 1]
[13]
HuMY, ZhangL, LuoTX, ShenW (2012). Variations in leaf functional traits of Stipa purpurea along a rainfall gradient in Xizang, China .Chinese Journal of Plant Ecology, 36, 136-143. (in Chinese with English abstract)[胡梦瑶, 张林, 罗天祥, 沈维 (2012). 西藏紫花针茅叶功能性状沿降水梯度的变化 . 植物生态学报, 36, 136-143.]
[14]
HuangJY, YuHL (2016). Responses of growth of four desert species to different N addition levels .Chinese Journal of Plant Ecology, 40, 165-176. (in Chinese with English abstract)[黄菊莹, 余海龙 (2016). 四种荒漠草原植物的生长对不同氮添加水平的响应 . 植物生态学报, 40, 165-176.] [本文引用: 1]
KerkhoffAJ, FaganWF, ElserJJ, EnquistBJ (2006). Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants .The American Naturalist, 168, 103-122. [本文引用: 3]
[17]
KillingbeckKT, WhitfordWG (1996). High foliar nitrogen in desert shrubs: An important ecosystem trait or defective desert doctrine .Ecology, 77, 1728-1737.
[18]
KoerselmanW, MeulemanAFM (1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation .Journal of Applied Ecology, 33, 1441-1450. [本文引用: 1]
[19]
LiYH, LuoTX, LuQ, TianXY, WuB, YangHH (2005). Comparisons of leaf traits among 17 major plant species in Shazhuyu Sand Control Experimental Station of Qinghai Province .Acta Ecology Sinica, 25, 994-999. (in Chinese with English abstract)[李永华, 罗天祥, 卢琦, 田晓娅, 吴波, 杨恒华 (2005). 青海省沙珠玉治沙站17种主要植物叶性因子的比较 . 生态学报, 25, 994-999.] [本文引用: 2]
[20]
LiYL, MaoW, ZhaoXY, ZhangTH (2010). Leaf nitrogen and phosphorus stoichiometry in typical desert and desertified regions, North China .Environment Science, 31, 1716-1725. (in Chinese with English abstract)[李玉霖, 毛伟, 赵学勇, 张铜会 (2010). 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究 . 环境科学, 31, 1716-1725.] [本文引用: 3]
[21]
LiuWD, SuJR, LiSF, ZhangZJ, LiZG (2010). Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province .Acta Ecologica Sinica, 30, 6581-6590. (in Chinese with English abstract)[刘万德, 苏建荣, 李帅锋, 张志钧, 李忠文 (2010). 云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征 . 生态学报, 30, 6581-6590.] [本文引用: 1]
[22]
LuoMW, MaoL, LiQQ, ZhaoX, XiaoY, JiaTT, GuoZG (2015). C, N, and P stoichiometry of plant and soil in the restorable plant communities distributed on the land used for Qinghai-Tibet Highway Construction in the Qinghai-Tibetan Plateau, China .Acta Ecologica Sinica, 35, 7832-7841. (in Chinese with English abstract)[雒明伟, 毛亮, 李倩倩, 赵旭, 肖玉, 贾婷婷, 郭正刚 (2015). 青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征 . 生态学报, 35, 7832-7841.] [本文引用: 1]
[23]
MaYZ, ZhongQL, JinBJ, LuHD, GuoBQ, ZhengY, LiM, ChengDL (2015). Spatial changes and influencing factors of fine root carbon, nitrogen and phosphorus stoichiometry of plants in China .Chinese Journal of Plant Ecology, 39, 159-166. (in Chinese with English abstract)[马玉珠, 钟全林, 靳冰洁, 卢宏典, 郭炳桥, 郑媛, 李曼, 程栋梁 (2015). 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子 . 植物生态学报, 39, 159-166.]
[24]
MarschnerH (1995). Mineral Nutrition of Higher Plants. Academic Press, London. [本文引用: 1]
[25]
NieLQ, WuQ, YaoB, FuS, HuQW (2016). Leaf litter and soil carbon, nitrogen, and phosphorus stoichiometry of dominant plant species in the Poyang Lake wetland .Acta Eclogica Sinica, 36, 1898-1906. (in Chinese with English abstract)[聂兰琴, 吴琴, 尧波, 付姗, 胡启武 (2016). 鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征 . 生态学报, 36, 1898-1906.] [本文引用: 1]
[26]
NiklasKJ, OwensT, ReichPB, CobbED (2005). Nitrogen phosphorus leaf stoichiometry and the scaling of plant growth .Ecology Letters, 8, 636-642. [本文引用: 1]
[27]
PregitzerKS, DeforestJL, BurtonAJ, AllenME, RuessRW, HendrickRL (2002). Fine root architecture of nine North American trees . Ecological Monographs, 72, 293-309. [本文引用: 1]
[28]
ReichPB (2014). The world-wide “fast-slow” plant economics spectrum: A traits manifesto .Journal of Ecology, 102, 275-301. [本文引用: 1]
[29]
SternerRW, ElserJJ (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton, USA. [本文引用: 1]
[30]
TjoelkerMG, CraineJM, WedinD, ReichPB, TilmanD (2005). Linking leaf and root trait syndromes among 39 grassland and savannah species .New Phytologist, 167, 493-508. [本文引用: 1]
[31]
WangWQ, WangC, SardansJ, ZengCS, TongC, PenuelasJ (2015). Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil-plant system in the Minjiang River tidal estuarine wetlands of China .Wetlands Ecology and Management, 23, 865-880. [本文引用: 1]
WeiH, WuB, YangW, LuoT (2011). Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China .Plant Biology, 13, 85-92. [本文引用: 2]
[34]
WrightIJ, ReichPB, WestobyM (2001). Strategy shifts in leaf physiology, structure and nutrient content between species of high -and low-rainfall and high -and low-nutrient habitats .Functional Ecology, 15, 423-434. [本文引用: 4]
[35]
XuB, ChengYX, GanHJ, ZhouWJ, HeJS (2010). Correlations between leaf and fine root traits among and within species of typical temperate grassland in Xilin River Basin, Inner Mongolia, China .Chinese Journal of Plant Ecology, 34, 29-38. (in Chinese with English abstract)[徐冰, 程雨曦, 甘慧洁, 周文嘉, 贺金生 (2010). 内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联 . 植物生态学报, 34, 29-38.] [本文引用: 1]
YuanZY, ChenHYH, ReichPB (2011). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus .Nature Communications, 2, 1-6. [本文引用: 2]
[38]
ZhuZD, CuiSH (1996). The problem of land desertification in South China .Journal of Desert Research, 16, 331-337. (in Chinese with English abstract)[朱震达, 崔书红. 中国南方的土地荒漠化问题 (1996). 中国沙漠 , 16, 331-337.] [本文引用: 1]
[39]
ZuoCQ (1986). Analysis for the causes, utilization and governance in sand hill, Poyang Lake .Soil and Water Conservation, (4), 2-5. (in Chinese)[左长青 (1986). 鄱阳湖沙山成因及治理利用浅析 . 中国水土保持, (4), 2-5.] [本文引用: 1]
The C:N:P stoichiometry of autotrophs-theory and observations 1 2004
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010).近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014). ...
Plant responses to multiple environmental factors 1 1987
Functional matrix: A conceptual framework for predicting multiple plant effects on ecosystem processes 1 2003
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003)、植物生长过程中对资源的利用和分配(Kerkhoff et al., 2006), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003; Tjoelker et al., 2005)、P含量(Kerkhoff et al., 2006; Geng et al., 2014)均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al., 2002; 王政权和郭大立, 2008), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al., 2002; Guo et al., 2008), 从而影响了与叶片之间相应元素的相关关系. ...
Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits 1 2014
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003)、植物生长过程中对资源的利用和分配(Kerkhoff et al., 2006), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003; Tjoelker et al., 2005)、P含量(Kerkhoff et al., 2006; Geng et al., 2014)均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al., 2002; 王政权和郭大立, 2008), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al., 2002; Guo et al., 2008), 从而影响了与叶片之间相应元素的相关关系. ...
Heterogeneity by root branch order: Exploring the discrepancy in root longevity and turnover estimates between minirhizotron and C isotope methods 1 2008
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003)、植物生长过程中对资源的利用和分配(Kerkhoff et al., 2006), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003; Tjoelker et al., 2005)、P含量(Kerkhoff et al., 2006; Geng et al., 2014)均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al., 2002; 王政权和郭大立, 2008), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al., 2002; Guo et al., 2008), 从而影响了与叶片之间相应元素的相关关系. ...
N:P ratios in terrestrial plants: Variation and functional significance 1 2004
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010).近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014). ...
生态化学计量学: 探索从个体到生态系统的统一化理论 1 2010
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010).近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014). ...
西藏紫花针茅叶功能性状沿降水梯度的变化 2012
四种荒漠草原植物的生长对不同氮添加水平的响应 1 2016
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010).近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014). ...
TRY—A global database of plant traits 2011
Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants 3 2006
Linking leaf and root trait syndromes among 39 grassland and savannah species 1 2005
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003)、植物生长过程中对资源的利用和分配(Kerkhoff et al., 2006), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003; Tjoelker et al., 2005)、P含量(Kerkhoff et al., 2006; Geng et al., 2014)均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al., 2002; 王政权和郭大立, 2008), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al., 2002; Guo et al., 2008), 从而影响了与叶片之间相应元素的相关关系. ...
Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil-plant system in the Minjiang River tidal estuarine wetlands of China 1 2015
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010).近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014). ...
根系生态学 1 2008
... 近年来, 越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制(Eviner & Chapin, 2003)、植物生长过程中对资源的利用和分配(Kerkhoff et al., 2006), 还对进一步预测植物对环境变化的响应具有重要意义.本研究发现单叶蔓荆各器官的养分元素及化学计量关系的相关性主要集中在地上部分, 即花、叶、枝条、匍匐茎之间, 其中尤以枝条与匍匐茎之间表现明显, 二者除了N:P没有达到显著相关之外, 其他的养分元素及其化学计量关系均达到了显著性相关(表2), 显示了二者之间在养分存储与运输之间的密切关系.在植物的所有器官中, 叶片与细根之间的性状关联最受关注.一些大尺度多物种间的研究表明叶片和细根的N含量(Craine & Lee, 2003; Tjoelker et al., 2005)、P含量(Kerkhoff et al., 2006; Geng et al., 2014)均呈显著正相关.本研究未发现叶片与细根之间的3种元素及其化学计量关系显著相关, 一个可能的原因是叶片与细根性状的关联主要发生在种间, 小尺度上植物生境条件相对一致, 种内个体间功能性状因变异幅度较小而关联不明显(徐冰等, 2010).另一种可能的原因是本研究中的细根样品的采集是以直径<2 mm为标准的.由于根系在发育过程中形成明显的分枝结构, 具有高度的形态异质性(Pregitzer et al., 2002; 王政权和郭大立, 2008), 直径<2 mm的细根并不是一个均质的单一组分, 而是由形态、寿命、生理差异巨大的不同个体根组成的高度异质的混合体(Pregitzer et al., 2002; Guo et al., 2008), 从而影响了与叶片之间相应元素的相关关系. ...
Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China 2 2011
Strategy shifts in leaf physiology, structure and nutrient content between species of high -and low-rainfall and high -and low-nutrient habitats 4 2001
... 碳(C)、氮(N)和磷(P)是植物基本营养元素, 在植物生长和各种生理调节机能中发挥着重要作用.C是构成植物体内干物质的最主要元素, 而N和P则分别与植物的光合作用和细胞分裂等重要生理活动有关(Niklas et al., 2005).N:C和P:C与植物的相对生长率关系密切(Ågren, 2004), N:P则可以反映植物生长受N或P的限制情况(Koerselman & Meuleman, 1996; Elser et al., 2000; Güsewell, 2004).植物不同组织器官营养元素水平与变化是调整它们的生长速率以适应环境变化的一种策略(Marschner, 1995), 如干旱区植物相对于湿润区植物通常拥有更高的单位面积(或质量)叶N、P含量(Wright et al., 2001; 李玉霖等, 2010), 植物地下根系的N、P含量低于地上部分的叶片, 但二者具有相似的N:P化学计量比(Yuan et al., 2011).当前, 区域C、N、P化学计量格局及其驱动因素仍然是生态化学计量学的一个重要研究领域, 其结果不仅有助于揭示物种间化学计量的趋同与分异, 对于预测生态系统功能、过程对全球变化的响应亦具有重要意义(贺金生和韩兴国, 2010; Elser et al., 2010).近年来, 国内****在不同时空尺度, 针对森林(刘万德等, 2010; 崔高阳等, 2015)、草地(Yu et al., 2010; 雒明伟等, 2015)、荒漠(李玉霖等, 2010; 黄菊莹和余海龙, 2016)、湿地(Wang et al., 2015; 聂兰琴等, 2016)等生态系统类型开展了大量的研究.然而, 集中于叶片的化学计量信息亟需拓展至植物其他器官(Elser et al., 2010).同时, 从丰富资源经济谱(economics spectrum)理论的角度看, 加强植物不同器官养分化学计量关系, 以及各器官之间的关联研究亦十分必要(Reich, 2014). ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT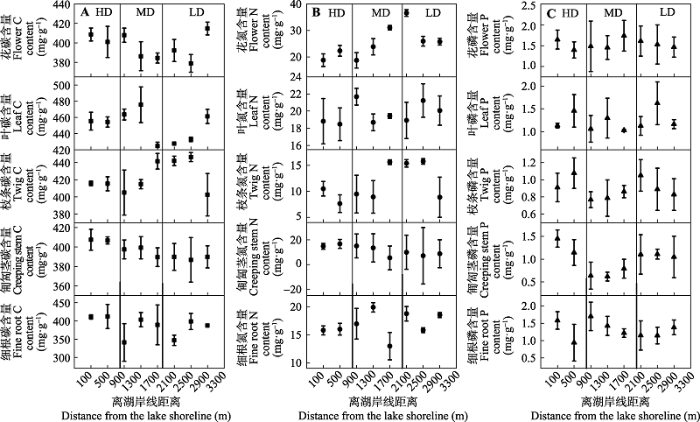 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT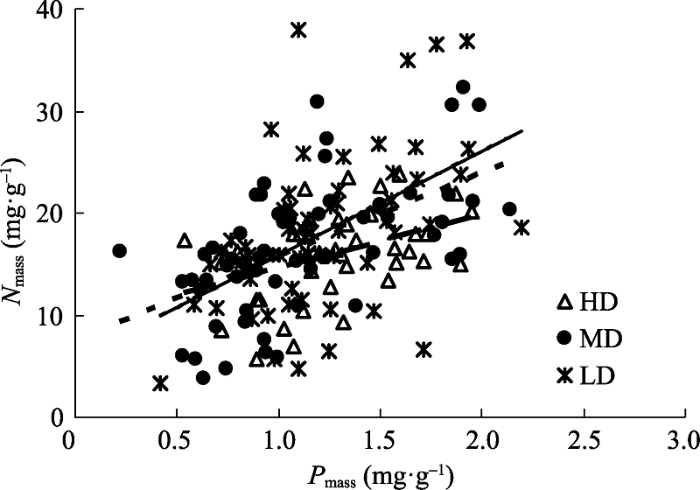 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}