Effects of enhanced UV-B radiation and nitrogen deposition on the growth of invasive plant Triadica sebifera
DENGBang-Liang, LIUQian, LIUXi-Shuai, ZHENGLi-Ya, JIANGLiang-Bo, GUOXiao-Min, LIUYuan-Qiu, ZHANGLing*, Jiangxi Key Laboratory of Silviculture/Collaborative Innovation Center of Jiangxi Typical Trees Cultivation and Utilization, College of Forestry, Jiangxi Agricultural University, Nanchang 330045, China 通讯作者:* 通信作者Author for correspondence (E-mail: lingzhang09@126.com) 责任编辑:DENGBang-LiangLIUQianLIUXi-ShuaiZHENGLi-YaJIANGLiang-BoGUOXiao-MinLIUYuan-QiuZHANGLing 收稿日期:2016-09-20 接受日期:2017-01-3 网络出版日期:2017-04-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41501317)
关键词:非生物胁迫;土壤微生物;资源可利用性;植物入侵 Abstract Aims Exotic plant invasions are important components of global change, threatening both the stability and function of invaded ecosystems. Shifts in competitive ability of invasive plants versus their native congeners have been documented. Enhanced UV-B radiation and nitrogen (N) deposition might interact with soil biota communities impacting the invasion process of exotic plant species. To understand the potential effects by UV-B and N with soil biota on plant growth would enhance our understanding of the mechanisms in plant invasions in the context of global change. Methods We conducted a full-factorial pot experiment in the native range (China) of Triadica sebifera invading US to investigate how UV-B radiation, N and soil biota together determined their seedling growth. Important findings The results showed that UV-B radiation, N and soil sterilization together impacted the growth of T. sebifera seedlings. UV-B radiation induced changes in biomass allocation with larger leaf biomass observed in response to UV-B radiation. In addition, N increased aboveground biomass and decreased root biomass simultaneously. Soil biota imposed positive effects on growth of T. sebifera, and the addition of N amplified these positive effects. The negative effects by UV-B radiation on growth of T. sebifera showed no response to N addition. Plant height, leaf biomass and total biomass of the invasive T. sebifera populations out- performed those of the native ones. In addition, invasive T. sebifera populations weakened the dependence of root/shoot ratio and root biomass on local soil microorganisms than native populations, but enhanced that of leaf area ratio.
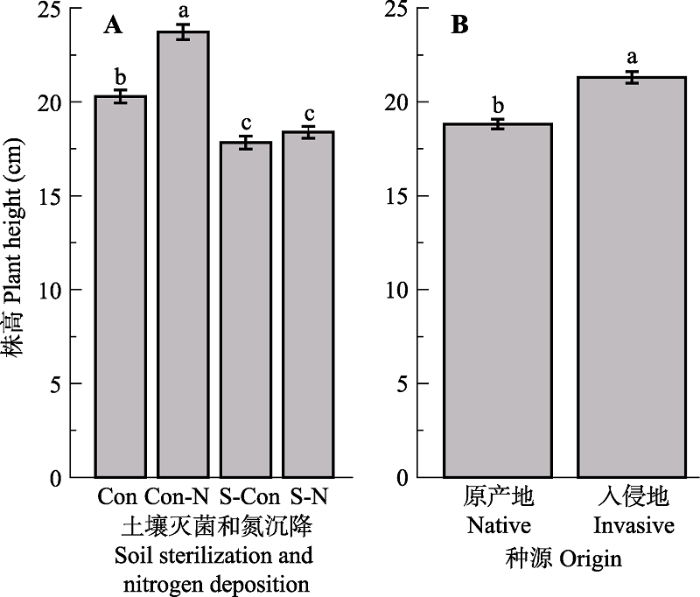
土壤灭菌、氮沉降以及土壤灭菌和氮沉降交互作用对株高具有极显著影响(p < 0.01)(表2)。氮沉降对株高具有显著正效应, 土壤灭菌以及土壤灭菌和氮沉降交互作用对株高具有显著负效应; 在氮沉降条件下, 土壤灭菌对株高具有显著负效应; 在土壤灭菌条件下, 氮沉降对株高没有影响(图1A)。入侵地乌桕较之原产地乌桕具有较高的株高(+13.17%)(图1B)。 显示原图|下载原图ZIP|生成PPT 图1不同处理对乌桕株高的影响(平均值±标准误差)。Con, 对照; N, 氮沉降; S, 土壤灭菌。相同小写字母表示没有显著差异(p > 0.05, LSD test)。 -->Fig. 1Effect of different treatments on plant height of Triadica sebifera (mean ± SE). Con, control; N, nitrogen deposition; S, soil sterilization. Same lowercase letters indicate no significant differences (p > 0.05, LSD test). -->
Table 2 表2 表2UV-B辐射(UV)、土壤灭菌(S)和氮沉降(N)处理对不同种源(O)乌桕形态学特征的影响方差分析 Table 2The dependence of morphological traits on Triadica sebifera origin (O), UV-B radiation (UV), soil sterilization (S) and nitrogen deposition (N) treatment, and their interactions in ANOVAs
处理 Treatment
df
F value
株高 Plant height
叶片数 Leaf number
叶面积 Leaf areas
比叶面积 Specific leaf area
叶面积比 Leaf area ratio
UV
1
0.01
4.96*
0.07
5.19*
0.00
S
1
144.85**
343.02**
820.08**
14.66**
78.98**
N
1
37.90**
3.60
40.55**
2.03
59.97**
O
1
58.13**
0.76
1.20
0.16
1.82
UV × S
1
0.52
0.26
0.02
0.05
0.58
UV × N
1
2.46
2.58
2.27
0.00
1.21
UV × O
1
0.05
0.22
0.20
1.01
0.05
S × N
1
19.70**
18.90**
73.34**
0.11
44.87**
S × O
1
0.22
1.86
2.09
2.59
8.57**
N × O
1
0.01
0.28
0.52
0.01
1.12
UV × S × N
1
0.01
2.49
0.06
0.23
0.10
UV × S × O
1
0.75
1.12
0.22
0.46
0.16
UV × N × O
1
0.07
0.08
0.24
0.12
0.40
S × N × O
1
1.17
2.18
2.76
1.52
2.81
UV × S × N × O
1
0.37
0.01
1.05
0.12
0.46
*, 0.01 < p < 0.05, **, p < 0.01, significant results (p < 0.05) are shown in bold.*, 0.01 < p < 0.05, **, p < 0.01, 显著性结果(p < 0.05)以粗体形式展示。 新窗口打开 土壤灭菌以及土壤灭菌和氮沉降交互作用对叶片数具有极显著影响(p < 0.01), UV-B辐射对叶片数具有显著影响(p < 0.05)(表2)。氮沉降对叶片数具有显著正效应, 土壤灭菌以及土壤灭菌和氮沉降的交互作用对叶片数具有显著负效应; 在氮沉降条件下, 土壤灭菌对叶片数具有显著负效应; 在土壤灭菌条件下, 氮沉降对叶片数没有影响(图2A)。UV-B辐射增加叶片数(+5.59%)(图2B)。 显示原图|下载原图ZIP|生成PPT 图2不同处理对乌桕叶形态学特征的影响(平均值±标准误差)。Con, 对照; N, 氮沉降; S, 土壤灭菌; UV, UV-B辐射; Native, 原产地; Invasive, 入侵地; SLA, 比叶面积; LAR, 叶面积比。相同小写字母表示没有显著差异(p > 0.05, LSD test)。 -->Fig. 2Effect of different treatments on leaf morphological traits of Triadica sebifera (mean ± SE). Con, control; N, nitrogen deposition; S, soil sterilization; UV, UV-B radiation; SLA, specific leaf area; LAR, leaf area ratio. Same lowercase letters indicate no significant differences (p > 0.05, LSD test). -->
BallaréCL (2014). Light regulation of plant defense .Annual Review of Plant Biology, 65, 335-363. [本文引用: 1]
[2]
BlosseyB, NotzoldR (1995). Evolution of increased competitive ability in invasive nonindigenous plants: A hypothesis .Journal of Ecology, 83, 887-889. [本文引用: 1]
[3]
BradleyBA, BlumenthalDM, WilcoveDS, ZiskaLH (2010). Predicting plant invasions in an era of global change .Trends in Ecology & Evolution, 25, 310-318. [本文引用: 1]
[4]
CallawayRM, BedmarEJ, ReinhartKO, SilvanCG, KlironomosJ (2011). Effects of soil biota from different ranges on Robinia invasion: Acquiring mutualists and escaping pathogens .Ecology, 92, 1027-1035. [本文引用: 1]
[5]
ChenBM, PengSL, WuXP, WangPL, MaJX (2016). A bibliometric analysis of researches on topics related to the ecological damage caused by and risk assessments of exotic invasive species from 1995 to 2014 .Acta Ecologica Sinica, 36, 1-9. (in Chinese with English abstract)[陈宝明, 彭少麟, 吴秀平, 王鹏龙, 马建霞 (2016). 近20年外来生物入侵危害与风险评估研究的文献计量分析 . 生态学报, 36, 1-9.] [本文引用: 1]
[6]
ChenHZ, HanR (2015). Plants respond to UV-B radiation: A review .Chinese Bulletin of Botany, 50, 790-801. (in Chinese with English abstract)[陈慧泽, 韩榕 (2015). 植物响应UV-B辐射的研究进展 . 植物学报, 50, 790-801.] [本文引用: 2]
[7]
ChenT, LiuWL, ZhangCB, WangJ (2012). Effects of Solidago canadensis invasion on dynamics of native plant communities and their mechanisms .Chinese Journal of Plant Ecology, 36, 253-261. (in Chinese with English abstract)[陈彤, 刘文莉, 张崇邦, 王江 (2012). 加拿大一枝黄花入侵对本土植物群落动态的影响及其机制 . 植物生态学报, 36, 253-261.] [本文引用: 1]
[8]
DavisMA, GrimeJP, ThompsonK (2000). Fluctuating resources in plant communities: A general theory of invasibility .Journal of Ecology, 88, 528-534. [本文引用: 1]
[9]
FengYL, FuGL, ZhengYL (2008). Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners .Planta, 228, 383-390. [本文引用: 1]
[10]
HeneryML, BowmanG, MrázP, TreierUA, Gex-FabryE, SchaffnerU, Müller-SchärerH (2010). Evidence for a combination of pre-adapted traits and rapid adaptive change in the invasive plantCentaurea stoebe .Journal of Ecology, 98, 800-813. [本文引用: 1]
[11]
HockM, BeckmannM, HofmannRR, BruelheideH, ErfmeierA (2015). Effects of UV-B radiation on germination characteristics in invasive plants in New Zealand .NeoBiota, 26, 21-37. [本文引用: 1]
[12]
HuangJX, XuX, WangM, NieM, QiuSY, WangQ, QuanZX, XiaoM, LiB (2016). Responses of soil nitrogen fixation to Spartina alterniflora invasion and nitrogen addition in a Chinese salt marsh .Scientific Reports, 6, 20384. doi: 10.1038/srep20384. [本文引用: 1]
HuangW, WangY, DingJQ (2013). A review of adaptive evolution of defense strategies in an invasive plant species, Chinese tallow (Triadica sebifera) .Chinese Journal of Plant Ecology, 37, 889-900. (in Chinese with English abstract)[黄伟, 王毅, 丁建清 (2013). 入侵植物乌桕防御策略的适应性进化研究 . 植物生态学报, 37, 889-900.] [本文引用: 1]
[15]
JuRT, LiH, ShiZR, LiB (2012). Progress of biological invasions research in China over the last decade .Biodiversity Science, 20, 581-611. (in Chinese with English abstract)[鞠瑞亭, 李慧, 石正人, 李博 (2012). 近十年中国生物入侵研究进展 . 生物多样性, 20, 581-611.]
[16]
KanakidouM, MyriokefalitakisS, DaskalakisN, FanourgakisG, NenesA, BakerAR, TsigaridisK, MihalopoulosN (2016). Past, present, and future atmospheric nitrogen deposition .Journal of the Atmospheric Sciences, 73, 2039-2047. [本文引用: 2]
[17]
KeaneRM, CrawleyMJ (2002). Exotic plant invasions and the enemy release hypothesis .Trends in Ecology & Evolution, 17, 164-170.
[18]
LeiYB, XiaoHF, FengYL (2010). Impacts of alien plant invasions on biodiversity and evolutionary responses of native species .Biodiversity Science, 18, 622-630. (in Chinese with English abstract)[类延宝, 肖海峰, 冯玉龙 (2010). 外来植物入侵对生物多样性的影响及本地生物的进化响应 . 生物多样性, 18, 622-630.] [本文引用: 1]
[19]
LiLB, TangTX, HaiMR, ChenJW, ZhouP (2015). Response and molecular mechanisms of plants to enhanced UV-B radiation .Chinese Agricultural Science Bulletin, 31(13), 159-163. (in Chinese with English abstract)[李良博, 唐天向, 海梅荣, 陈军文, 周平 (2015). 植物对UV-B辐射增强的响应及其分子机制 . 中国农学通报, 31(13), 159-163.] [本文引用: 1]
[20]
LiLB, ZhangLG, TangTX, HaiMR, ZhouP (2016). Influence of enhanced UV-B on physiological characteristics of Eupatorium adenophorum L. and Artemisia argyi Levl .Acta Botanica Boreali-Occidentalia Sinica, 36, 343-352. (in Chinese with English abstract)[李良博, 张连根, 唐天向, 海梅荣, 周平 (2016). UV-B辐射增强对紫茎泽兰和艾草抗性生理特性的影响 . 西北植物学报, 36, 343-352.] [本文引用: 3]
[21]
LiY, LiX, HeYM, XiaY, ZuYQ (2015). Effect and evaluation of enhanced UV-B radiation on interactions between plant and pathogen .Plant Physiology Journal, 51, 1557-1566. (in Chinese with English abstract)[李元, 李想, 何永美, 夏杨, 祖艳群 (2015). UV-B辐射增强对植物-病原菌互作体系的影响及评价 . 植物生理学报, 51, 1557-1566.] [本文引用: 1]
[22]
LiuJ, LiJM, YuH, HeWM, YuFH, SangWG, LiuGF, DongM (2010). The relationship between functional traits and invasiveness of alien plants .Biodiversity Science, 18, 569-576. (in Chinese with English abstract)[刘建, 李钧敏, 余华, 何维明, 于飞海, 桑卫国, 刘国方, 董鸣 (2010). 植物功能性状与外来植物入侵 . 生物多样性, 18, 569-576.] [本文引用: 2]
[23]
LuRK (2000). Methods of Soil Agricultural Chemical Analysis. China Agriculture Scientech Press, Beijing. (in Chinese)[鲁如坤 (2000). 土壤农业化学分析方法. 中国农业科技出版社, 北京.] [本文引用: 3]
[24]
NiY, SongC, LiJN (2015). Effect of enhanced ultraviolet-B radiation on epicuticular wax inArabidopsis thaliana .Acta Ecologica Sinica, 35, 1505-1512. (in Chinese with English abstract)[倪郁, 宋超, 李加纳 (2015). UV-B辐射增强对拟南芥表皮蜡质的影响 . 生态学报, 35, 1505-1512.]
[25]
NijjerS, RogersWE, SiemannE (2007). Negative plant-soil feedbacks may limit persistence of an invasive tree due to rapid accumulation of soil pathogens .Proceedings of the Royal Society B: Biological Sciences, 274, 2621-2627. [本文引用: 1]
[26]
SucharVA, RobberechtR (2016). Integration and scaling of UV-B radiation effects on plants: From molecular interactions to whole plant responses . Ecology and Evolution, 1-19. [本文引用: 1]
[27]
van der PuttenWH, KlironomosJN, WardleDA (2007). Microbial ecology of biological invasions .The ISME Journal, 1, 28-37. [本文引用: 2]
WangH, MaXC, ZhangL, ZouJW, SiemannE (2016). UV-B has larger negative impacts on invasive populations of Triadica sebifera but ozone impacts do not vary .Journal of Plant Ecology, 9, 61-68. [本文引用: 2]
[30]
WangHX, LiuWZ (2011). Effects of enhanced UV-B radiation on leaf morphology and structure and pigment contents of Camptotheca acuminata .Chinese Agricultural Science Bulletin, 27(5), 209-213. (in Chinese with English abstract)[王海霞, 刘文哲 (2011). UV-B辐射增强对喜树叶片色素含量和形态结构的影响 . 中国农学通报, 27(5), 209-213.]
[31]
WilliamsonCE, ZeppRG, LucasRM, MadronichS, AustinAT, BallaréCL, NorvalM, SulzbergerB, BaisAF, McKenzieRL, RobinsonSA, HäderDP, PaulND, BornmanJF (2014). Solar ultraviolet radiation in a changing climate .Nature Climate Change, 4, 434-441. [本文引用: 1]
YangQ, LiB, SiemannE (2015). The effects of fertilization on plant-soil interactions and salinity tolerance of invasive Triadica sebifera .Plant and Soil, 394, 99-107. [本文引用: 2]
[34]
ZhangL, ZhangY, WangH, ZouJW, SiemannE (2013). Chinese tallow trees (Triadica sebifera) from the invasive range outperform those from the native range with an active soil community or phosphorus fertilization .PLOS ONE, 8, e74233. doi: 10.1371/journal.pone.0074233. [本文引用: 3]
[35]
ZouJ, RogerWE, SiemannE (2007). Differences in morphological and physiological traits between native and invasive populations of Sapium sebiferum .Functional Ecology, 21, 721-730. [本文引用: 6]
[36]
ZouJW, RogerWE, DeWaltSJ, SiemannE (2006). The effect of Chinese tallow tree (Sapium sebiferum) ecotype on soil-plant system carbon and nitrogen processes .Oecologia, 150, 272-281. [本文引用: 1]
... 氮作为植物生长所需的大量元素, 其水平的高低直接影响植物的新陈代谢和生长发育.工业革命以来, 人类活动导致大气氮沉降加剧(Kanakidou et al., 2016), 直接提高了土壤氮素可利用水平.“资源可利用性增强”假说认为: 某一群落内资源可利用水平增加会导致外来植物入侵该群落的可能性增加(Davis et al., 2000).由于部分入侵植物较之本地植物表现出更强的氮吸收速率(Zou et al., 2006), 因而氮沉降加剧可能对植物入侵具有促进作用(Bradley et al., 2010; 陈彤等, 2012). ...
Fluctuating resources in plant communities: A general theory of invasibility 1 2000
... 氮作为植物生长所需的大量元素, 其水平的高低直接影响植物的新陈代谢和生长发育.工业革命以来, 人类活动导致大气氮沉降加剧(Kanakidou et al., 2016), 直接提高了土壤氮素可利用水平.“资源可利用性增强”假说认为: 某一群落内资源可利用水平增加会导致外来植物入侵该群落的可能性增加(Davis et al., 2000).由于部分入侵植物较之本地植物表现出更强的氮吸收速率(Zou et al., 2006), 因而氮沉降加剧可能对植物入侵具有促进作用(Bradley et al., 2010; 陈彤等, 2012). ...
Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners 1 2008
Effects of UV-B radiation on germination characteristics in invasive plants in New Zealand 1 2015
... 由于氟氯烃等大气污染物的大量排放, 致使O3层空洞日趋严重, 最终导致地表UV-B辐射强度增加(Williamson et al., 2014).UV-B辐射增强会造成植物DNA损伤(如嘧啶二聚体)并诱发一系列形态学和生理生化方面的防御反应, 甚至通过干扰植物-土壤反馈过程而进一步影响土壤养分吸收和微生物活性等过程(陈慧泽和韩榕, 2015; 李良博等, 2015; Suchar & Robberecht, 2016).由于适应策略以及响应机制的不同, 使得入侵植物与本地植物对UV-B辐射损伤的修复和抵御能力不同(Hock et al., 2015; 李良博等, 2015, 2016; Wang et al., 2015), 因此, UV-B作为一种非生物胁迫, 可能在植物入侵过程中扮演着至关重要的角色. ...
Responses of soil nitrogen fixation to Spartina alterniflora invasion and nitrogen addition in a Chinese salt marsh 1 2016
... “天敌逃逸”假说认为, 外来植物能够成功入侵可能与入侵地无专食性天敌危害有关(Keane & Crawley, 2002; Huang et al., 2012; 黄伟等, 2013), 随着研究的深入, 土壤有害微生物(如病原菌)也被认为具有类似于地上天敌的作用(van der Putten et al., 2007; Callaway et al., 2011); 此外, 土壤有益微生物共生体(如根瘤菌、菌根真菌等)通常能够协助入侵植物高效地利用土壤氮(Huang et al., 2016)、磷(Zhang et al., 2013)等养分, 因此, 土壤有害和有益微生物的综合效应可能促进或者阻碍植物入侵. ...
Interactive effects of herbivory and competition intensity determine invasive plant performance 2 2012
... “天敌逃逸”假说认为, 外来植物能够成功入侵可能与入侵地无专食性天敌危害有关(Keane & Crawley, 2002; Huang et al., 2012; 黄伟等, 2013), 随着研究的深入, 土壤有害微生物(如病原菌)也被认为具有类似于地上天敌的作用(van der Putten et al., 2007; Callaway et al., 2011); 此外, 土壤有益微生物共生体(如根瘤菌、菌根真菌等)通常能够协助入侵植物高效地利用土壤氮(Huang et al., 2016)、磷(Zhang et al., 2013)等养分, 因此, 土壤有害和有益微生物的综合效应可能促进或者阻碍植物入侵. ...
... 乌桕(Triadica sebifera)原产于中国, 18世纪末作为油料和观赏树种引入美国, 如今在美国东南部出现严重入侵现象, 形成茂密的单一优势种, 严重破坏入侵地生态系统(Huang et al., 2012; Zhang et al., 2013; Yang et al., 2015).乌桕入侵种群通过“地上优先”的分配策略以及高耐性、低抗性的进化选择, 使其比原产地种群具有更高的相对生长速率、氮吸收速率和比叶面积等特征(Zou et al., 2006, 2007; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
入侵植物乌桕防御策略的适应性进化研究 1 2013
... “天敌逃逸”假说认为, 外来植物能够成功入侵可能与入侵地无专食性天敌危害有关(Keane & Crawley, 2002; Huang et al., 2012; 黄伟等, 2013), 随着研究的深入, 土壤有害微生物(如病原菌)也被认为具有类似于地上天敌的作用(van der Putten et al., 2007; Callaway et al., 2011); 此外, 土壤有益微生物共生体(如根瘤菌、菌根真菌等)通常能够协助入侵植物高效地利用土壤氮(Huang et al., 2016)、磷(Zhang et al., 2013)等养分, 因此, 土壤有害和有益微生物的综合效应可能促进或者阻碍植物入侵. ...
近十年中国生物入侵研究进展 2012
Past, present, and future atmospheric nitrogen deposition 2 2016
... 乌桕(Triadica sebifera)原产于中国, 18世纪末作为油料和观赏树种引入美国, 如今在美国东南部出现严重入侵现象, 形成茂密的单一优势种, 严重破坏入侵地生态系统(Huang et al., 2012; Zhang et al., 2013; Yang et al., 2015).乌桕入侵种群通过“地上优先”的分配策略以及高耐性、低抗性的进化选择, 使其比原产地种群具有更高的相对生长速率、氮吸收速率和比叶面积等特征(Zou et al., 2006, 2007; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
UV-B辐射增强对喜树叶片色素含量和形态结构的影响 2011
Solar ultraviolet radiation in a changing climate 1 2014
... 由于氟氯烃等大气污染物的大量排放, 致使O3层空洞日趋严重, 最终导致地表UV-B辐射强度增加(Williamson et al., 2014).UV-B辐射增强会造成植物DNA损伤(如嘧啶二聚体)并诱发一系列形态学和生理生化方面的防御反应, 甚至通过干扰植物-土壤反馈过程而进一步影响土壤养分吸收和微生物活性等过程(陈慧泽和韩榕, 2015; 李良博等, 2015; Suchar & Robberecht, 2016).由于适应策略以及响应机制的不同, 使得入侵植物与本地植物对UV-B辐射损伤的修复和抵御能力不同(Hock et al., 2015; 李良博等, 2015, 2016; Wang et al., 2015), 因此, UV-B作为一种非生物胁迫, 可能在植物入侵过程中扮演着至关重要的角色. ...
The effects of fertilization on plant-soil interactions and salinity tolerance of invasive Triadica sebifera 2 2015
... 由于氟氯烃等大气污染物的大量排放, 致使O3层空洞日趋严重, 最终导致地表UV-B辐射强度增加(Williamson et al., 2014).UV-B辐射增强会造成植物DNA损伤(如嘧啶二聚体)并诱发一系列形态学和生理生化方面的防御反应, 甚至通过干扰植物-土壤反馈过程而进一步影响土壤养分吸收和微生物活性等过程(陈慧泽和韩榕, 2015; 李良博等, 2015; Suchar & Robberecht, 2016).由于适应策略以及响应机制的不同, 使得入侵植物与本地植物对UV-B辐射损伤的修复和抵御能力不同(Hock et al., 2015; 李良博等, 2015, 2016; Wang et al., 2015), 因此, UV-B作为一种非生物胁迫, 可能在植物入侵过程中扮演着至关重要的角色. ...
... 乌桕(Triadica sebifera)原产于中国, 18世纪末作为油料和观赏树种引入美国, 如今在美国东南部出现严重入侵现象, 形成茂密的单一优势种, 严重破坏入侵地生态系统(Huang et al., 2012; Zhang et al., 2013; Yang et al., 2015).乌桕入侵种群通过“地上优先”的分配策略以及高耐性、低抗性的进化选择, 使其比原产地种群具有更高的相对生长速率、氮吸收速率和比叶面积等特征(Zou et al., 2006, 2007; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
Chinese tallow trees (Triadica sebifera) from the invasive range outperform those from the native range with an active soil community or phosphorus fertilization 3 2013
... 乌桕(Triadica sebifera)原产于中国, 18世纪末作为油料和观赏树种引入美国, 如今在美国东南部出现严重入侵现象, 形成茂密的单一优势种, 严重破坏入侵地生态系统(Huang et al., 2012; Zhang et al., 2013; Yang et al., 2015).乌桕入侵种群通过“地上优先”的分配策略以及高耐性、低抗性的进化选择, 使其比原产地种群具有更高的相对生长速率、氮吸收速率和比叶面积等特征(Zou et al., 2006, 2007; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
... ; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
Differences in morphological and physiological traits between native and invasive populations of Sapium sebiferum 6 2007
... “天敌逃逸”假说认为, 外来植物能够成功入侵可能与入侵地无专食性天敌危害有关(Keane & Crawley, 2002; Huang et al., 2012; 黄伟等, 2013), 随着研究的深入, 土壤有害微生物(如病原菌)也被认为具有类似于地上天敌的作用(van der Putten et al., 2007; Callaway et al., 2011); 此外, 土壤有益微生物共生体(如根瘤菌、菌根真菌等)通常能够协助入侵植物高效地利用土壤氮(Huang et al., 2016)、磷(Zhang et al., 2013)等养分, 因此, 土壤有害和有益微生物的综合效应可能促进或者阻碍植物入侵. ...
... 乌桕(Triadica sebifera)原产于中国, 18世纪末作为油料和观赏树种引入美国, 如今在美国东南部出现严重入侵现象, 形成茂密的单一优势种, 严重破坏入侵地生态系统(Huang et al., 2012; Zhang et al., 2013; Yang et al., 2015).乌桕入侵种群通过“地上优先”的分配策略以及高耐性、低抗性的进化选择, 使其比原产地种群具有更高的相对生长速率、氮吸收速率和比叶面积等特征(Zou et al., 2006, 2007; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
... ; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
... ), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...
The effect of Chinese tallow tree (Sapium sebiferum) ecotype on soil-plant system carbon and nitrogen processes 1 2006
... 乌桕(Triadica sebifera)原产于中国, 18世纪末作为油料和观赏树种引入美国, 如今在美国东南部出现严重入侵现象, 形成茂密的单一优势种, 严重破坏入侵地生态系统(Huang et al., 2012; Zhang et al., 2013; Yang et al., 2015).乌桕入侵种群通过“地上优先”的分配策略以及高耐性、低抗性的进化选择, 使其比原产地种群具有更高的相对生长速率、氮吸收速率和比叶面积等特征(Zou et al., 2006, 2007; Zhang et al., 2013; 吴昊和丁建清, 2014; Yang et al., 2015), 在生长方面已经进化出明显的优势.鉴于乌桕成功入侵与非生物和生物因素的复杂性, 国内外****相继开展了原产地和入侵地乌桕种群对UV-B辐射增强(Wang et al., 2015), 氮沉降和土壤微生物交互作用(Zhang et al., 2013), 施肥、盐度和土壤微生物交互作用(Yang et al., 2015)等相关研究, 通过了解不同非生物和生物条件下入侵地和原产地乌桕种群在形态学和生理特征方面的差异, 逐步揭示了乌桕成功入侵的关键因素, 然而, 土壤肥力是如何直接或间接调控微生物介导的植物生长来应对非生物胁迫尚不明确. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT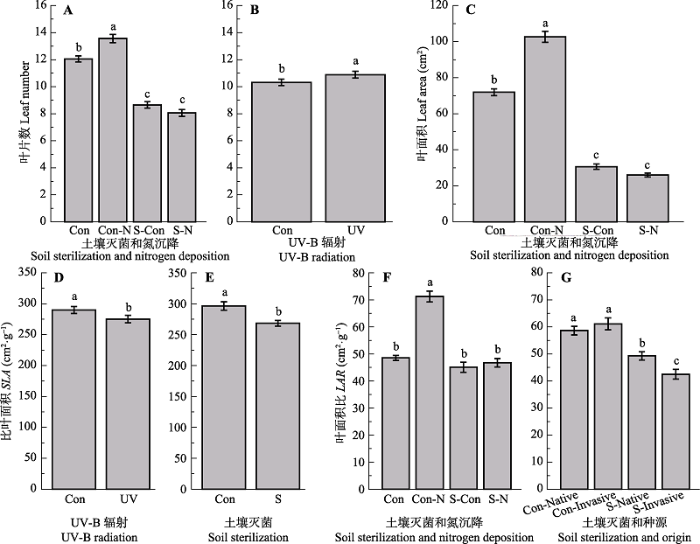 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT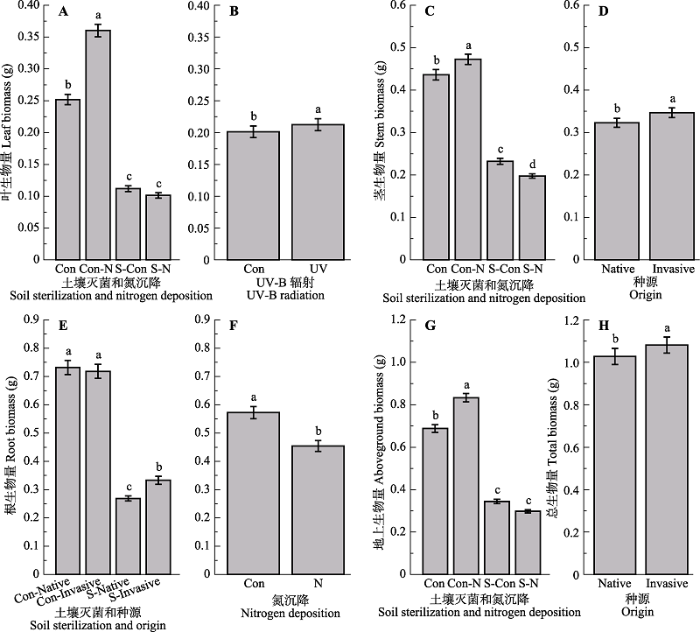 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT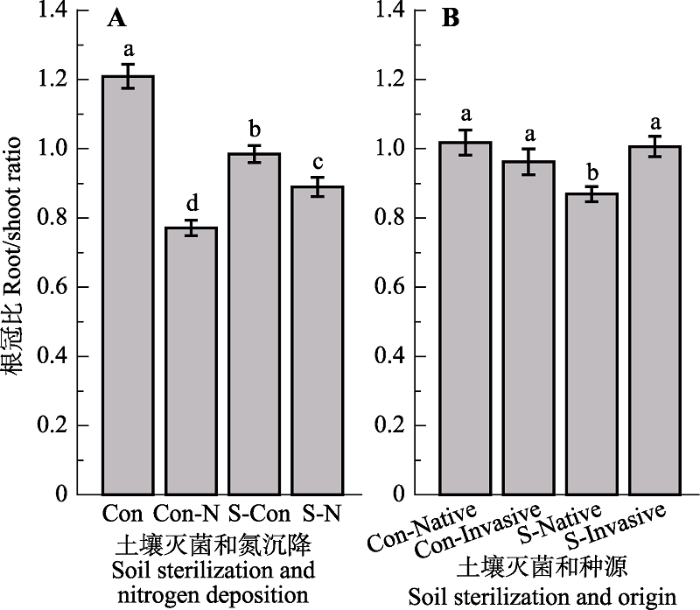 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}