Responses of soil respiration with biocrust cover to water and temperature in the southeastern edge of Tengger Desert, Northwest China
GUANChao1,2, ZHANGPeng1,*,, LIXin-Rong1 1Shapotou Desert Research and Experimental Station, Northwest Institute of Eco-environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, Chinaand 2University of Chinese Academy of Sciences, Beijing 100049, China 通讯作者:* 通信作者Author for correspondence (E-mail:zhangp1419@163.com) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41530746和41621001)和中国科学院“西部之光”博士项目
关键词:呼吸作用;生物结皮;降水格局;模拟增温 Abstract Aims Soil respiration of the lands covered by biocrusts is an important component in the carbon cycle of arid, semi-arid and dry-subhumid ecosystems (drylands hereafter), and one of the key processes in the carbon cycle of drylands. However, the responses of the rate of soil respiration with biocrusts to water and temperature are uncertain in the investigations of the effects of experimental warming and precipitation patterns on CO2 fluxes in biocrust dominated ecosystems. The objectives of this study were to investigate the relationships of carbon release from the biocrust-soil systems with water and temperature in drylands. Methods Intact soil columns with two types of biocrusts, including moss and algae-lichen crusts, were collected in a natural vegetation area in the southeastern fringe of the Tengger Desert. Open top chambers were used to simulate climate warming, and the soil respiration rate was measured under warming and non-warming treatments using an automated soil respiration system (LI-8150). Important findings Over the whole observational period (from April 2016 to July 2016), soil respiration rates varied from -0.16 to 4.69 μmol·m-2·s-1 for the moss crust-covered soils and from -0.21 to 5.72 μmol·m-2·s-1 for the algae-lichen crust-covered soils, respectively, under different rainfall events (the precipitations between 0.3-30.0 mm). The mean soil respiration rate of the moss crust-covered soils is 1.09 μmol·m-2·s-1, which is higher than that of the algae-lichen crust-covered soils of 0.94 μmol·m-2·s-1. The soil respiration rate of the two types of biocrust-covered soils showed different dynamics and spatial heterogeneities with rainfall events, and were positively correlated with precipitation. The mean soil respiration rate of the biocrust-covered soils without warming was 1.24 μmol·m-2·s-1, significantly higher than that with warming treatments of 0.79 μmol·m-2·s-1 (p < 0.05). By increasing the evaporation of soil moisture, the simulated warming impeded soil respiration. In most cases, soil temperature and soil respiration rate displayed a similar single-peak curve during the diel cycle. Our results show an approximately two hours’ lag between soil temperature at 5 cm depth and the soil respiration rate of the biocrust-covered soils during the diel cycle.
Keywords:respiration;biocrust;precipitation patterns;stimulated warming -->0 PDF (3479KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 管超, 张鹏, 李新荣. 腾格里沙漠东南缘生物结皮土壤呼吸对水热因子变化的响应. 植物生态学报, 2017, 41(3): 301-310 https://doi.org/10.17521/cjpe.2016.0326 GUANChao, ZHANGPeng, LIXin-Rong. Responses of soil respiration with biocrust cover to water and temperature in the southeastern edge of Tengger Desert, Northwest China. Chinese Journal of Plant Ecology, 2017, 41(3): 301-310 https://doi.org/10.17521/cjpe.2016.0326 荒漠生态系统是全球最大的陆地生态系统, 约占陆地总面积的41%, 为全球37%的人口提供生活支撑(Reynolds et al., 2007)。大多数荒漠生态系统受水分等非生物因子的调控和限制, 地表不可能连续分布大面积相对均匀的维管植物, 植物群落的斑块状分布, 为生物结皮的定居和拓殖提供了适宜的生态位, 使其覆盖度达到干旱区地表总面积的40% (李新荣等, 2009), 在一些受干扰较少的极端环境群落中, 生物结皮盖度甚至达到70% (Belnap, 1994)。生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009)。众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016)。 联合国政府间气候变化专门委员会(IPCC)第五次报告指出, 1880-2012年全球温度平均升高了 0.85 ℃, 北半球高纬度地区升温更明显, 预计到21世纪末将升温0.3-4.8 ℃ (IPCC, 2013)。气候变暖将导致极端天气和气候事件发生频率增加(IPCC, 2013), 同时将导致包括降水量、降水频率、降水强度和降水分布等在内的降水格局变化(Swemmer et al., 2007)。降水变化将会对土壤、植被、水文和土地荒漠化等多方面产生影响(朱雅娟等, 2012)。但这种降水格局的变化在不同区域范围内存在一定的差异, 我国极端干旱和干旱地区的降水量总体上呈增加趋势, 而半干旱和半湿润地区的降水量总体上呈减少趋势(朱雅娟等, 2012)。沙坡头地区的降水量在近30年内呈减少趋势(黄磊等, 2010)。对中国北方干旱区而言, 少量的降水增加不能弥补温度升高所带来的蒸发量的增加, 因此, 未来的气候变化将向暖干化趋势发展(施雅风, 1996)。土壤呼吸是一个复杂的过程, 受土壤温度、土壤湿度、降水、凋落物、土壤碳(C)、氮(N)含量等非生物因子, 以及植被类型、叶面积指数、根系生物量等生物因子的综合影响(Lundegardh, 1927), 是生物圈碳循环的重要组成部分, 占生态系统总呼吸的3/4 (Law et al., 2001)。干旱、半干旱生态系统对未来气候变化引起的温度和降水模式的变化高度敏感(Lane et al., 2013), 因而, 对于该生态系统相对有限的土壤有机碳库而言, 土壤呼吸速率的微小变化对整个生态系统碳储量的影响深远(West et al., 1994)。 有关温带荒漠生物结皮土壤呼吸已有大量研究, 如不同结皮类型、演替阶段生物结皮的呼吸动态特征(高艳红等, 2012; Su et al., 2013), 生物结皮土壤呼吸与水分(Zhao et al., 2014; Zhang et al., 2015)、温度(Zhang et al., 2015)、土壤理化性质(Zhang et al., 2013)等环境因子的关系等, 但仍然缺少生物结皮土壤呼吸对水热因子交互效应响应动态的研究, 尤其缺乏来自野外条件下的连续监测数据。因此, 本研究以腾格里沙漠东南缘天然植被区发育良好的两类典型生物结皮(藻-地衣和藓类为优势种)为研究对象, 野外采集原状结皮土壤样品, 利用开顶式生长室(OTC)模拟增温, 野外条件下连续监测两类生物结皮土壤呼吸动态, 研究降水变化和温度对其呼吸动态的影响, 以期为了解温带荒漠生态系统对全球变化的响应和适应机制提供数据支撑。
AllisonSD, TresederKK (2008). Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Global Change Biology, 14, 2898-2909. [本文引用: 2]
[2]
AustinAT, YahdjianL, StarkJM, BelnapJ, PorporatoA, NortonU, RavettaDA, SchaefferSM (2004). Water pluses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia, 141, 221-235. [本文引用: 1]
[3]
BelnapJ (1994). Cryptobiotic soil crusts: Basis for arid land restoration (Utah). Restoration and Management Notes, 12, 85-86. [本文引用: 1]
[4]
BelnapJ, PhillipsSL, MillerME (2004). Response of desert biological soil crusts to alterations in precipitation frequency. Oecologia, 141, 106-316. [本文引用: 1]
[5]
BowlingD, GroteE, BelnapJ (2011). Rain pulse response of soil CO2 exchange by biological soil crusts and grasslands of the semiarid Colorado Plateau, United States. Journal of Geophysical Research Atmospheres, 116, 2415-2422. [本文引用: 1]
[6]
ChenJ, LuoYQ, XiaJY, ShiZ, JiangLF, NiuSL, ZhouXH, CaoJJ (2016). Differential responses of ecosystem respiration components to experimental warming in a meadow grassland on the Tibetan Plateau. Agricultural and Forest Meteorology, 220, 21-29.
[7]
ConantRT, Dalla-BettaP, KlopatekCC, KlopatekJM (2004). Controls on soil respiration in semiarid soils. Soil Biology & Biochemistry, 36, 945-951. [本文引用: 1]
[8]
GaoYH, ZhangZS, LiuLC, JiaRL (2012). Soil respiration patterns during restoration of vegetation in the Shapotou Area, Northern China. Acta Ecologica Sinica, 32, 2474-2482. (in Chinese with English abstract)[高艳红, 张志山, 刘立超, 贾荣亮 (2012). 沙坡头人工植被演替过程的土壤呼吸特征 . 生态学报, 32, 2474-2482.]
[9]
GroismanPY, KnightRW, EasterlingDR, KarlTR, HegerlGC, RazuvaevVN (2005). Trends in intense precipitation in the climate record. Journal of Climate, 18, 1326-1350. [本文引用: 1]
[10]
HuYG, FengYL, ZhangZS, HuangL, ZhangP, XuBX (2014). Greenhouse gases fluxes of biological soil crusts and soil ecosystem in the artificial sand-fixing vegetation region in Shapotou area. Chinese Journal of Applied Ecology, 25, 61-68. (in Chinese with English abstract)[胡宜刚, 冯玉兰, 张志山, 黄磊, 张鹏, 徐冰鑫 (2014). 沙坡头人工植被固沙区生物结皮-土壤系统温室气体通量特征 . 应用生态学报, 25, 61-68.] [本文引用: 1]
[11]
HuangG, LiY, SuYG (2015). Effects of increasing precipitation on soil microbial community composition and soil respiration in a temperate desert, Northwestern China. Soil Biology & Biochemistry, 83, 52-56.
[12]
HuangL, ZhangZS, WuP (2010). Wavelet analysis of the precipitation time series in Shapotou desert area. Journal of Lanzhou University (Natural Sciences), 46(5), 63-66. (in Chinese with English abstract)[黄磊, 张志山, 吴攀 (2010). 沙坡头地区多年降水量时间序列的小波分析 . 兰州大学学报(自然科学版), 46(5), 63-66.]
[13]
IPCC (Intergovernmental Panel on Climate Change) (2013). Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. [本文引用: 3]
[14]
LaneRW, MenonM, McQuaidJB, AdamsDG, ThomasAD, HoonSR, DougillAJ (2013). Laboratory analysis of the effects of elevated atmospheric carbon dioxide on respira¬tion in biological soil crusts. Journal of Arid Environ¬ments, 98, 52-59.
[15]
LawBE, KelliherFM, BaldocchiDD, AnthoniPM, IrvineJ, MooreD, van TuylS (2001). Spatial and temporal variation in respiration in a young ponderosa pine forest during a summer drought. Agricultural and Forest Meteorology, 110, 27-43. [本文引用: 1]
[16]
LiXR, ZhangYM, ZhaoYG (2009). A study of biological soil crusts: Recent development, trend and prospect. Advances in Earth Science, 24, 11-14. (in Chinese with English abstract)[李新荣, 张元明, 赵允格 (2009). 生物土壤结皮研究: 进展、前沿与展望 . 地球科学进展, 24, 11-14.] [本文引用: 1]
[17]
LiXR, ZhouHY, WangXP, LiuLC, ZhangJG, ChenGX, ZhangZS, LiuYB, TanHJ, GaoYH (2016). Ecological restoration and recovery in arid desert regions of China: A review for 60-year research progresses of Shapotou Desert Research and Experiment Station, Chinese Academy of Sciences. Journal of Desert Research, 36, 247-264. (in Chinese with English abstract)[李新荣, 周海燕, 王新平, 刘立超, 张景光, 陈国雄, 张志山, 刘玉冰, 谭会娟, 高艳红 (2016). 中国干旱沙区的生态重建与恢复: 沙坡头站60年重要研究进展综述 . 中国沙漠, 36, 247-264.] [本文引用: 1]
[18]
LloydJ, TaylorJA (1994). On the temperature dependence of soil respiration. Functional Ecology, 8, 315-323.
LuoCY, XuGP, ChaoZG, WangSP, LinXW, HuYG, ZhangZH, DuanJC, ChangXF, SuAL, LiYN, ZhaoXQ, DuMY, TangYH, KimballB (2010). Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan Plateau. Global Change Biology, 16, 1606-1617.
[21]
LuoYQ, ZhouXH (2007). Soil Respiration and the Environment. Translated by Jiang LF, Qu LY, Zhou YM, Wen YX. Higher Education Press, Beijing. 18-21. (in Chinese)[骆亦其, 周旭辉 (2007). 土壤呼吸与环境. 姜丽芬, 曲来叶, 周玉梅, 温逸馨, 译. 高等教育出版社, 北京.] [本文引用: 2]
[22]
MaestreFT, EscolarC, de GuevaraML, QueroJL, LázaroR, Delgado-BaquerizoM, OchoaV, BerdugoM, GozaloB, GallardoA (2013). Changes in biocrust cover drive carbon cycle responses to climate change in drylands. Global Change Biology, 19, 3835-3847. [本文引用: 1]
NewM, ToddM, HulmeM, JonesP (2001). Precipitation measurements and trends in the twentieth century. International Journal of Climatology, 21, 1899-1922. [本文引用: 1]
[25]
Noy-MeirI (1973). Desert ecosystems environment and producers. Annual Review of Ecology and Systematics, 4, 25-51.
[26]
OberbauerSF, TweedieCE, WelkerJM, FahnestockJM, HenryGHR, WebberPJ, HollisterRD, WalkerMD, KuchyA, ElmoreE, StarrG (2007). Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients. Ecological Monographs, 77, 221-238.
[27]
ReynoldsJF, SmithDM, LambinEF, TurnerBL, MortimoreM, BatterburyS, DowningTE, DowlatabadiH, FernandezRJ, HerrickJE, Huber-SannwaldE, JiangH, LeemansR, LynamT, MaestreFT, AyarzaM, WalkerBG (2007). Global desertification: Building a science for dryland development. Science, 316, 847-851. [本文引用: 1]
[28]
Riveros-IreguiDA, EmanuelRE, MuthDJ, McGlynnBL, EpsteinHE, WelschDL, PacificVJ, WraithJM (2007). Diurnal hysteresis between soil CO2 and soil temperature is controlled by soil water content. Geophysical Research Letters, 34, L17404.
[29]
SalaOE, LaurenrothWK (1982). Small rainfall events: An ecological role in semiarid regions. Oecologia, 53, 301-304. [本文引用: 1]
[30]
SchwinningS, SalaOE (2004). Hierarchy of responses to re¬source pulses in arid and semi-arid ecosystems. Oecologia, 141, 211-220. [本文引用: 2]
[31]
SharkhuuA, PlanteAF, EnkhmandalO, GonneauC, CasperBB, BoldgivB, PetraitisPS (2016). Soil and ecosystem respiration responses to grazing, watering and experimen¬tal warming chamber treatments across topographical gra¬dients in northern Mongolia. Geoderma, 269, 91-98.
[32]
ShenZX, LiYL, FuG (2015). Response of soil respiration to short-term experimental warming and precipitation pulses over the growing season in an alpine meadow on the Northern Tibet. Applied Soil Ecology, 90, 35-40. [本文引用: 1]
[33]
ShiYF (1996). Features and tendency of global warming and its implications for China. Journal of Natural Disasters, 5(2), 1-10. (in Chinese with English abstract)[施雅风 (1996). 全球和中国变暖特征及未来趋势 . 自然灾害学报, 5(2), 1-10.] [本文引用: 2]
[34]
SuYG, WuL, ZhouZB, LiuYB, ZhangYM (2013). Carbon flux in deserts depends on soil cover type: A case study in the Gurbantunggute desert, North China. Soil Biology & Biochemistry, 58, 332-340. [本文引用: 1]
[35]
SubkeJA, ReichsteinM, TenhunenJD (2003). Explaining temporal variation in soil CO2 efflux in a mature spruce forest in Southern Germany. Soil Biology & Biochemistry, 35, 1467-1483. [本文引用: 1]
[36]
SwemmerAM, KnappAK, SnymanHA (2007). Intra-seasonal precipitation patterns and aboveground productivity in three perennial grasslands. Journal of Ecology, 95, 780-788. [本文引用: 1]
[37]
WangB, ZhaTS, JiaX, WuB, ZhangYQ, QinSG (2014). Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem. Biogeosciences, 11, 259-268. [本文引用: 3]
[38]
WangQ, HeNP, LiuY, LiML, XuL (2016). Strong pulse effects of precipitation events on soil microbial respiration in temperate forests. Geoderma, 275, 67-73.
[39]
WestNE, StarkJM, JohnsonDW, AbramsMM, WightJR, HeggemD, PeckS (1994). Effects of climatic-change on the edaphic features of arid and semiarid lands of western north America. Arid Soil Research Rehabilitation, 8, 307-351. [本文引用: 1]
[40]
XuBX, HuYG, ZhangZS, ChenYL, ZhangP, LiG (2014). Effects of experimental warming on CO2, CH4 and N2O fluxes of biological soil crust and soil system in a desert region. Chinese Journal of Plant Ecology, 38, 809-820. (in Chinese with English abstract)[徐冰鑫, 胡宜刚, 张志山, 陈永乐, 张鹏, 李刚 (2014). 模拟增温对荒漠生物土壤结皮-土壤系统CO2、CH4和N2O通量的影响 . 植物生态学报, 38, 809-820.] [本文引用: 1]
[41]
XuWF, LiXL, LiuW, LiLH, HouLY, ShiHQ, XiaJZ, LiuD, ZhangHC, ChenY, CaiWW, FuY, YuanWP (2016). Spatial patterns of soil and ecosystem respiration regulated by biological and environmental variables along a precipitation gradient in semi-arid grasslands in China. Ecological Research, 31, 505-513. [本文引用: 1]
[42]
ZhangZS, DongXJ, LiuYB, LiXR, JiaRL, HuYG, HeMZ, HuangL (2012). Soil oxidases recovered faster than hydrolases in a 50-year chronosequence of desert revegetation. The Plant Soil, 358, 275-287. [本文引用: 1]
[43]
ZhangZS, DongXJ, XuBX, ChenYL, ZhaoY, GaoYH, HuYG, HuangL (2015). Soil respiration sensitivities to water and temperature in a revegetated desert. Journal of Geophysical Research: Biogeosciences, 120, 773-787. [本文引用: 2]
[44]
ZhangZS, LiXR, NowakRS, WuP, GaoYH, ZhaoY, HuangL, HuYG, JiaRL (2013). Effect of sand-stabilizing shrubs on soil respiration in a temperate desert. The Plant Soil, 367, 449-463. [本文引用: 3]
[45]
ZhaoY, LiXR, ZhangZS (2014). Biological soil crusts influence carbon release response following rainfall in a temperate desert, northern China. Ecological Research, 29, 889-896. [本文引用: 1]
[46]
ZhuYJ, WuB, LuQ (2012). Progress in the study on response of arid zones to precipitation change. Forest Research, 25, 100-106. (in Chinese with English abstract)[朱雅娟, 吴波, 卢琦 (2012). 干旱区对降水变化响应的研究进展 . 林业科学研究, 25, 100-106.]
Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. 2 2008
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
... ), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Water pluses and biogeochemical cycles in arid and semiarid ecosystems. 1 2004
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Cryptobiotic soil crusts: Basis for arid land restoration (Utah). 1 1994
... 荒漠生态系统是全球最大的陆地生态系统, 约占陆地总面积的41%, 为全球37%的人口提供生活支撑(Reynolds et al., 2007).大多数荒漠生态系统受水分等非生物因子的调控和限制, 地表不可能连续分布大面积相对均匀的维管植物, 植物群落的斑块状分布, 为生物结皮的定居和拓殖提供了适宜的生态位, 使其覆盖度达到干旱区地表总面积的40% (李新荣等, 2009), 在一些受干扰较少的极端环境群落中, 生物结皮盖度甚至达到70% (Belnap, 1994).生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
Response of desert biological soil crusts to alterations in precipitation frequency. 1 2004
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Rain pulse response of soil CO2 exchange by biological soil crusts and grasslands of the semiarid Colorado Plateau, United States. 1 2011
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Differential responses of ecosystem respiration components to experimental warming in a meadow grassland on the Tibetan Plateau. 2016
Controls on soil respiration in semiarid soils. 1 2004
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
沙坡头人工植被演替过程的土壤呼吸特征 2012
Trends in intense precipitation in the climate record. 1 2005
... 有关温带荒漠生物结皮土壤呼吸已有大量研究, 如不同结皮类型、演替阶段生物结皮的呼吸动态特征(高艳红等, 2012; Su et al., 2013), 生物结皮土壤呼吸与水分(Zhao et al., 2014; Zhang et al., 2015)、温度(Zhang et al., 2015)、土壤理化性质(Zhang et al., 2013)等环境因子的关系等, 但仍然缺少生物结皮土壤呼吸对水热因子交互效应响应动态的研究, 尤其缺乏来自野外条件下的连续监测数据.因此, 本研究以腾格里沙漠东南缘天然植被区发育良好的两类典型生物结皮(藻-地衣和藓类为优势种)为研究对象, 野外采集原状结皮土壤样品, 利用开顶式生长室(OTC)模拟增温, 野外条件下连续监测两类生物结皮土壤呼吸动态, 研究降水变化和温度对其呼吸动态的影响, 以期为了解温带荒漠生态系统对全球变化的响应和适应机制提供数据支撑. ...
沙坡头人工植被固沙区生物结皮-土壤系统温室气体通量特征 1 2014
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Effects of increasing precipitation on soil microbial community composition and soil respiration in a temperate desert, Northwestern China. 2015
On the temperature dependence of soil respiration. 1994
Carbon dioxide evolution and crop growth. 3 1927
... 荒漠生态系统是全球最大的陆地生态系统, 约占陆地总面积的41%, 为全球37%的人口提供生活支撑(Reynolds et al., 2007).大多数荒漠生态系统受水分等非生物因子的调控和限制, 地表不可能连续分布大面积相对均匀的维管植物, 植物群落的斑块状分布, 为生物结皮的定居和拓殖提供了适宜的生态位, 使其覆盖度达到干旱区地表总面积的40% (李新荣等, 2009), 在一些受干扰较少的极端环境群落中, 生物结皮盖度甚至达到70% (Belnap, 1994).生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
... ).生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
... ), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan Plateau. 2010
2
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Changes in biocrust cover drive carbon cycle responses to climate change in drylands. 1 2013
... 另外, 本研究发现, 5 cm土壤温度日变化曲线峰值出现的时间通常滞后于生物结皮土壤呼吸2 h (图6, 图7).类似的, 来自宁夏盐池半干旱生态系统的研究发现, 10 cm土壤温度日变化曲线出现峰值的时间滞后于土壤呼吸, 并且滞后时间与土壤含水量呈显著的线性负相关关系(Wang et al., 2014).来自山地森林生态系统的研究发现, 20 cm土壤温度日变化曲线出现峰值的时间滞后于土壤呼吸, 并且滞后时间与土壤含水量显著正相关(Riveros-Iregui et al., 2007).然而, 目前对造成这种滞后性的原因尚不明确.现有解释认为或许是因为土壤呼吸与所测量的土壤温度的深度不匹配, 或许是因为光合作用的日变化对根际呼吸作用的影响(Subke et al., 2003).土壤呼吸包括自养呼吸和异养呼吸两部分, 自养呼吸主要受空气温度影响, 而异养呼吸主要受土壤温度影响(Lloyd & Taylor, 1994), 自养呼吸和异养呼吸对环境因子响应的差异也可能是造成这种滞后性的一个可能原因(Riveros-Iregui et al., 2007).因此, 未来需进一步通过长期控制实验来研究造成这种滞后性的原因. ...
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Precipitation measurements and trends in the twentieth century. 1 2001
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Desert ecosystems environment and producers. 1973
Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients. 2007
Global desertification: Building a science for dryland development. 1 2007
... 荒漠生态系统是全球最大的陆地生态系统, 约占陆地总面积的41%, 为全球37%的人口提供生活支撑(Reynolds et al., 2007).大多数荒漠生态系统受水分等非生物因子的调控和限制, 地表不可能连续分布大面积相对均匀的维管植物, 植物群落的斑块状分布, 为生物结皮的定居和拓殖提供了适宜的生态位, 使其覆盖度达到干旱区地表总面积的40% (李新荣等, 2009), 在一些受干扰较少的极端环境群落中, 生物结皮盖度甚至达到70% (Belnap, 1994).生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
Diurnal hysteresis between soil CO2 and soil temperature is controlled by soil water content. 2007
Small rainfall events: An ecological role in semiarid regions. 1 1982
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Hierarchy of responses to re¬source pulses in arid and semi-arid ecosystems. 2 2004
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Soil and ecosystem respiration responses to grazing, watering and experimen¬tal warming chamber treatments across topographical gra¬dients in northern Mongolia. 2016
Response of soil respiration to short-term experimental warming and precipitation pulses over the growing season in an alpine meadow on the Northern Tibet. 1 2015
... 荒漠生态系统是全球最大的陆地生态系统, 约占陆地总面积的41%, 为全球37%的人口提供生活支撑(Reynolds et al., 2007).大多数荒漠生态系统受水分等非生物因子的调控和限制, 地表不可能连续分布大面积相对均匀的维管植物, 植物群落的斑块状分布, 为生物结皮的定居和拓殖提供了适宜的生态位, 使其覆盖度达到干旱区地表总面积的40% (李新荣等, 2009), 在一些受干扰较少的极端环境群落中, 生物结皮盖度甚至达到70% (Belnap, 1994).生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
全球和中国变暖特征及未来趋势 2 1996
... 另外, 本研究发现, 5 cm土壤温度日变化曲线峰值出现的时间通常滞后于生物结皮土壤呼吸2 h (图6, 图7).类似的, 来自宁夏盐池半干旱生态系统的研究发现, 10 cm土壤温度日变化曲线出现峰值的时间滞后于土壤呼吸, 并且滞后时间与土壤含水量呈显著的线性负相关关系(Wang et al., 2014).来自山地森林生态系统的研究发现, 20 cm土壤温度日变化曲线出现峰值的时间滞后于土壤呼吸, 并且滞后时间与土壤含水量显著正相关(Riveros-Iregui et al., 2007).然而, 目前对造成这种滞后性的原因尚不明确.现有解释认为或许是因为土壤呼吸与所测量的土壤温度的深度不匹配, 或许是因为光合作用的日变化对根际呼吸作用的影响(Subke et al., 2003).土壤呼吸包括自养呼吸和异养呼吸两部分, 自养呼吸主要受空气温度影响, 而异养呼吸主要受土壤温度影响(Lloyd & Taylor, 1994), 自养呼吸和异养呼吸对环境因子响应的差异也可能是造成这种滞后性的一个可能原因(Riveros-Iregui et al., 2007).因此, 未来需进一步通过长期控制实验来研究造成这种滞后性的原因. ...
... ), 自养呼吸和异养呼吸对环境因子响应的差异也可能是造成这种滞后性的一个可能原因(Riveros-Iregui et al., 2007).因此, 未来需进一步通过长期控制实验来研究造成这种滞后性的原因. ...
Carbon flux in deserts depends on soil cover type: A case study in the Gurbantunggute desert, North China. 1 2013
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Explaining temporal variation in soil CO2 efflux in a mature spruce forest in Southern Germany. 1 2003
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
Intra-seasonal precipitation patterns and aboveground productivity in three perennial grasslands. 1 2007
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem. 3 2014
... 荒漠生态系统是全球最大的陆地生态系统, 约占陆地总面积的41%, 为全球37%的人口提供生活支撑(Reynolds et al., 2007).大多数荒漠生态系统受水分等非生物因子的调控和限制, 地表不可能连续分布大面积相对均匀的维管植物, 植物群落的斑块状分布, 为生物结皮的定居和拓殖提供了适宜的生态位, 使其覆盖度达到干旱区地表总面积的40% (李新荣等, 2009), 在一些受干扰较少的极端环境群落中, 生物结皮盖度甚至达到70% (Belnap, 1994).生物结皮是由蓝细菌, 藻类、地衣、苔藓等隐花植物, 及其他相关非维管植物利用菌丝体、假根和分泌物等与浅层土壤颗粒“胶结”而成的复杂团聚结构(李新荣等, 2009), 是干旱、半干旱荒漠生态系统地表景观的重要组成部分, 在促进维管植物定居生长、稳定土壤表面、影响水分再分配等方面具有重要的作用, 同时也是该系统重要的碳源, 对该系统的碳循环起着直接和间接的作用(李新荣等, 2009).众多研究表明, 生物结皮土壤呼吸对温度和湿度等非生物因子变化敏感(Maestre et al., 2013; Shen et al., 2015; Wang et al., 2016). ...
... 水分有效性是限制干旱、半干旱地区一切生命活动的关键因子(Noy-Meir, 1973).降水格局的变化会影响包括土壤呼吸在内的生态系统碳循环(Bowling et al., 2011).干旱、半干旱地区通常以小降水事件(降水量<5 mm)为主, 降水直接关系到土壤水分的有效性(Sala & Laurenroth, 1982).本研究中, 小降水事件约占全部降水事件的68.8% (图1), 与沙坡头地区近30年的小降水事件所占比例(68.3%)相近.这些小的降水虽然只能湿润地表0-5 cm土壤, 但足以激发植被和土壤中的生命活动(Schwinning & Sala, 2004).而生物结皮的存在延长了水分在浅层土壤中的保存时间, 特别是当干旱胁迫发生时, 生物结皮增加浅层土壤水分有效性的功能显得尤为重要(李新荣等, 2016).生物结皮属变水(poikilohydric)生物, 自身对水分的调节能力极差, 无法利用深层土壤水分(Nash, 1996).而生物结皮仅在湿润时才能进行新陈代谢活动, 因此其生物活性主要与降水量、降水频度和降水的持续时间密切相关(Belnap et al., 2004).本研究发现, 不同降水事件下, 生物结皮土壤呼吸呈现出不同的变化动态, 具有明显的时空异质性(图2, 图3), 这是由每次降水的大小、频度、持续时间及降水前所经历干旱时间的差异造成的.土壤微生物呼吸对降水响应迅速且强烈(Wang et al., 2016), 降水的增加会通过协同增加细菌和真菌的丰富度而增加微生物的生物量, 并进一步增加荒漠生态系统土壤碳释放量(Huang et al., 2015).因此, 通过土壤微生物呼吸速率的变化可以确定全球变化背景下降水模式变化对土壤碳含量的影响(Wang et al., 2016).另外, 研究表明, 降水加快了土壤呼吸速率, 且降水对土壤呼吸的影响取决于降水的大小、前期情况及土壤水分状况等因素(Shen et al., 2015).也就是说, 当土壤处于湿润状态时, 土壤呼吸对降水的响应减弱, 而当土壤处于干旱状态时, 土壤呼吸对降水的响应加强.然而, 干旱半干旱地区降水稀少, 蒸发强烈, 土壤主要处于干旱状态, 因此降水对土壤呼吸的影响主要取决于降水的大小.本研究发现, 生物结皮土壤呼吸速率与降水量有显著的线性正相关关系(图5).类似的, Huang等(2015)的研究表明, 土壤呼吸随着降水量的增加而线性增加.Xu等(2016)来自内蒙古草原的研究表明, 土壤呼吸和生态系统呼吸均与降水量呈显著的线性正相关关系.胡宜刚等(2014)和徐冰鑫等(2014)来自沙坡头地区的研究表明, 生物结皮土壤系统CO2通量与土壤湿度呈显著的线性正相关关系.此外, Zhao等(2014)来自沙坡头地区的研究表明, 降水模式的变化, 特别是较大降水事件对生物结皮土壤碳释放的影响占主导地位.然而, 在全球、地区及小尺度上的极端降水事件呈现出增加趋势(New et al., 2001), 且极端降水事件出现的频率还会继续增加(Groisman et al., 2005).因此, 关于不同降水事件下生物结皮土壤呼吸的变化, 有待未来长期深入的 研究. ...
... 除了水分之外, 温度也是影响生物结皮土壤呼吸的重要环境因子(Noy-Meir, 1973).Oberbauer等(2007)的研究表明, 增温显著增加干旱苔原生态系统的呼吸作用.Maestre等(2013)研究发现, 在半干旱地区增温显著提高了生物结皮的呼吸速率.来自沙坡头天然植被区的研究表明, 增温降低了藓类结皮和藻类结皮土壤系统的年平均CO2通量, 但其降低程度的差异不显著(徐冰鑫等, 2014).而本研究中, 增温降低了两类结皮土壤呼吸速率(图4), 这与上述的研究结果不一致.主要原因是: 尽管增温能够通过刺激土壤酶的活性而加速土壤有机质的分解(Luo et al., 2010), 进而促进土壤微生物的呼吸, 但干旱区受水分制约明显, 水分几乎是干旱区所有生态过程的第一驱动力(Austin et al., 2004), 土壤有机质分解、微生物、酶活性受到抑制(Allison & Treseder, 2008), 进而影响土壤呼吸.增温主要是通过降低土壤水分有效性而抑制土壤微生物活性, 进而抑制土壤呼吸(Allison & Treseder, 2008; Shen et al., 2015), 土壤呼吸速率受土壤含水量的影响大于受土壤温度的影响(Conant et al., 2004; Sharkhuu et al., 2016).另外, 荒漠生态系统有机质贫乏(Zhang et al., 2012), 缺少根系呼吸(骆亦其和周旭辉, 2007)等自养呼吸, 而Chen等(2016)研究发现, 增温显著降低了异养呼吸, 这也是增温降低生物结皮土壤呼吸的重要原因.此外, 采用OTC模拟增温并不能完全代表野外的真实情况, 未来将通过野外原位监测试验来进一步完善相关研究结果. ...
Strong pulse effects of precipitation events on soil microbial respiration in temperate forests. 2016
Effects of climatic-change on the edaphic features of arid and semiarid lands of western north America. 1 1994
... 有关温带荒漠生物结皮土壤呼吸已有大量研究, 如不同结皮类型、演替阶段生物结皮的呼吸动态特征(高艳红等, 2012; Su et al., 2013), 生物结皮土壤呼吸与水分(Zhao et al., 2014; Zhang et al., 2015)、温度(Zhang et al., 2015)、土壤理化性质(Zhang et al., 2013)等环境因子的关系等, 但仍然缺少生物结皮土壤呼吸对水热因子交互效应响应动态的研究, 尤其缺乏来自野外条件下的连续监测数据.因此, 本研究以腾格里沙漠东南缘天然植被区发育良好的两类典型生物结皮(藻-地衣和藓类为优势种)为研究对象, 野外采集原状结皮土壤样品, 利用开顶式生长室(OTC)模拟增温, 野外条件下连续监测两类生物结皮土壤呼吸动态, 研究降水变化和温度对其呼吸动态的影响, 以期为了解温带荒漠生态系统对全球变化的响应和适应机制提供数据支撑. ...
Spatial patterns of soil and ecosystem respiration regulated by biological and environmental variables along a precipitation gradient in semi-arid grasslands in China. 1 2016
... 另外, 本研究发现, 5 cm土壤温度日变化曲线峰值出现的时间通常滞后于生物结皮土壤呼吸2 h (图6, 图7).类似的, 来自宁夏盐池半干旱生态系统的研究发现, 10 cm土壤温度日变化曲线出现峰值的时间滞后于土壤呼吸, 并且滞后时间与土壤含水量呈显著的线性负相关关系(Wang et al., 2014).来自山地森林生态系统的研究发现, 20 cm土壤温度日变化曲线出现峰值的时间滞后于土壤呼吸, 并且滞后时间与土壤含水量显著正相关(Riveros-Iregui et al., 2007).然而, 目前对造成这种滞后性的原因尚不明确.现有解释认为或许是因为土壤呼吸与所测量的土壤温度的深度不匹配, 或许是因为光合作用的日变化对根际呼吸作用的影响(Subke et al., 2003).土壤呼吸包括自养呼吸和异养呼吸两部分, 自养呼吸主要受空气温度影响, 而异养呼吸主要受土壤温度影响(Lloyd & Taylor, 1994), 自养呼吸和异养呼吸对环境因子响应的差异也可能是造成这种滞后性的一个可能原因(Riveros-Iregui et al., 2007).因此, 未来需进一步通过长期控制实验来研究造成这种滞后性的原因. ...
Soil oxidases recovered faster than hydrolases in a 50-year chronosequence of desert revegetation. 1 2012
 , 李新荣
, 李新荣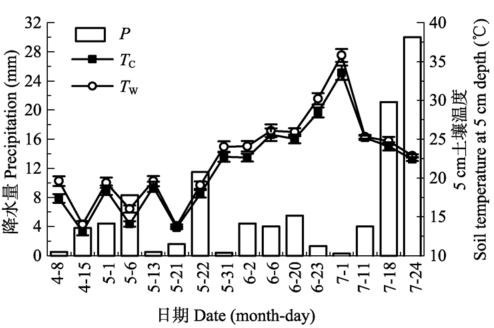 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT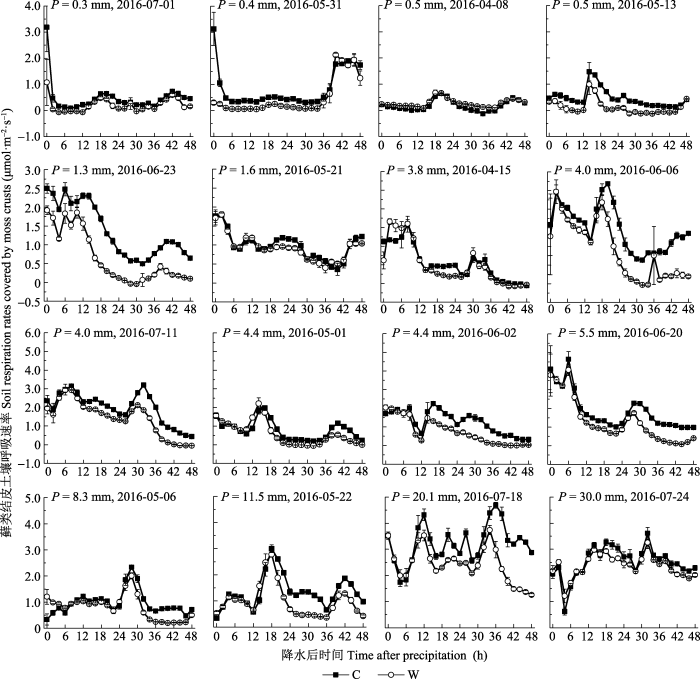 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT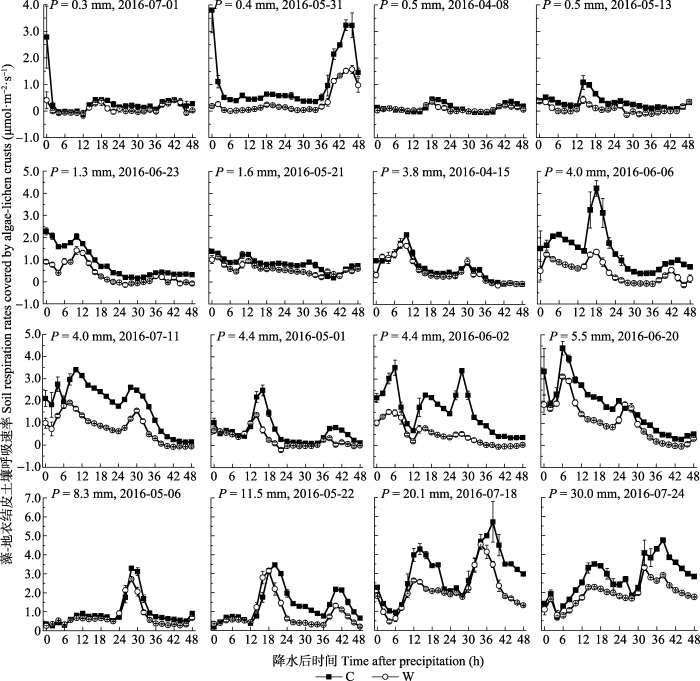 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT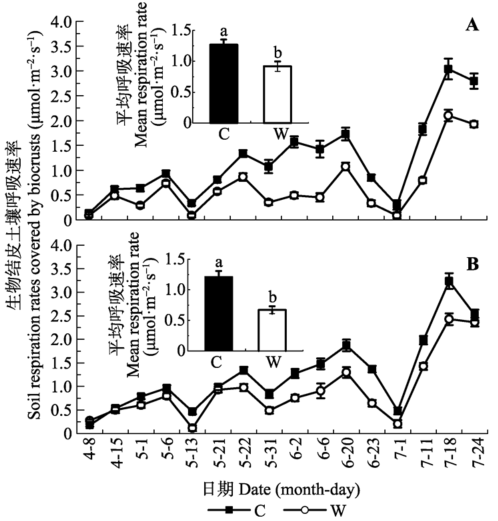 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT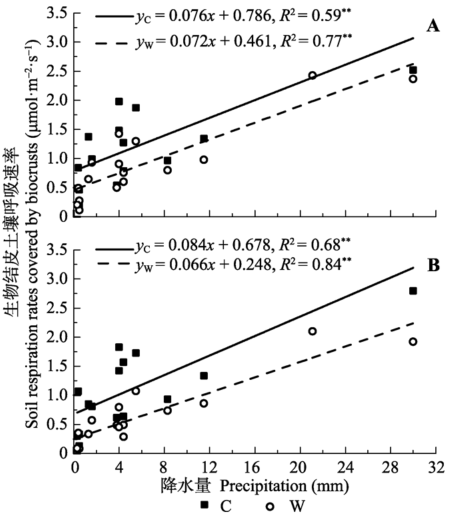 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT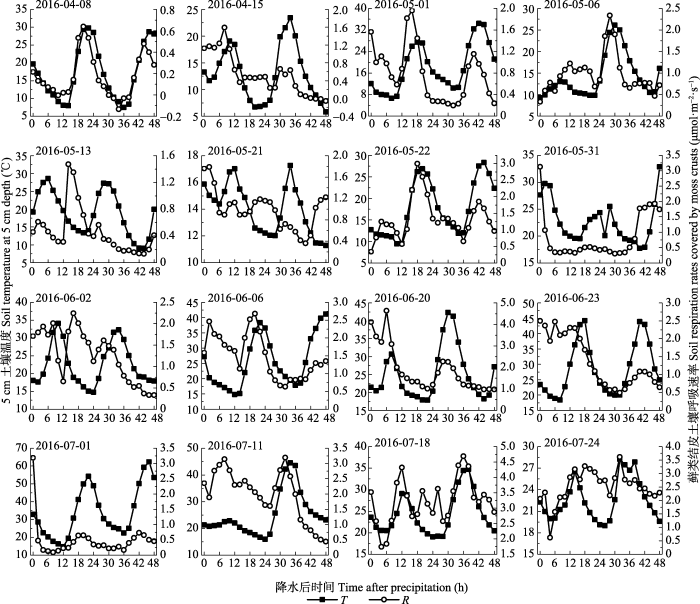 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT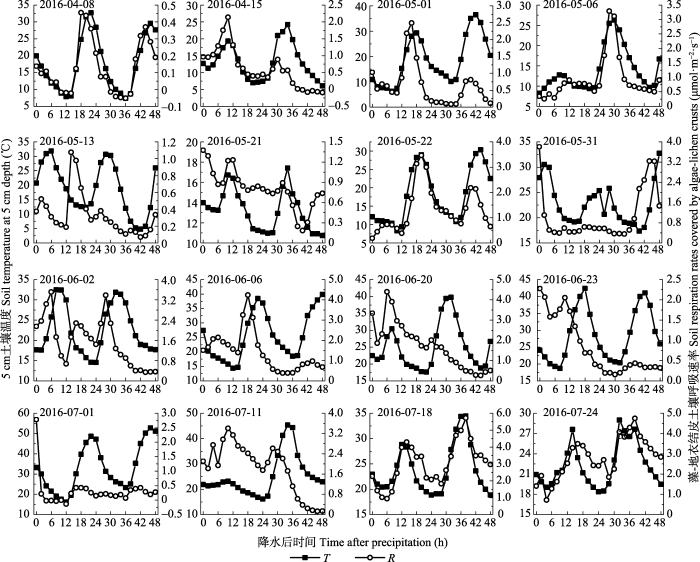 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}