Effluxes of nitrous oxide, methane and carbon dioxide and their responses to increasing nitrogen deposition in the Gurbantünggüt Desert of Xinjiang, China
ZHOUXiao-Bing1, ZHANGYuan-Ming1,*,, TAOYe1,2, WULin1,3 1Xinjiang Institute of Ecology and Geography, Key Laboratory of Biogeography and Bioresource in Arid Land, Chinese Academy of Sciences, Ürümqi 830011, China2Anqing Normal University, Anqing, Anhui 246000, Chinaand 3Hubei University for Nationalities, Enshi, Hubei 445000, China 通讯作者:* 通信作者Author for correspondence (E-mail:zhangym@ms.xjb.ac.cn) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:新疆维吾尔自治区自然科学基金面上项目(2015211A052)
关键词:氮沉降;N2O;CH4;CO2;生物量;结构方程模型 Abstract Aims Desert soils play an important role in the exchange of major greenhouse gas (GHG) between atmosphere and soil. However, many uncertainties existed in understanding of desert soil role, especially in efflux evaluation under a changing environment. Methods We conducted plot-based field study in center of the Gurbantünggüt Desert, Xinjiang, and applied six rates of simulated nitrogen (N) deposition on the plots, i.e. 0 (N0), 0.5 (N0.5), 1.0 (N1), 3.0 (N3), 6.0 (N6) and 24.0 (N24) g·m-2·a-1. The exchange rates of N2O, CH4 and CO2 during two growing seasons were measured for two years after N applications. Important findings The average efflux of two growing seasons from control plots (N0) were 4.8 μg·m-2·h-1, -30.5 μg·m-2·h-1 and 46.7 mg·m-2·h-1 for N2O, CH4 and CO2, respectively. The effluxes varied significantly among seasons. N0, N0.5 and N1 showed similar exchange of N2O in spring and summer, which was relatively higher than in autumn, while the rates of N2O in N6 and N24 were controled by time points of N applications. The uptake of CH4 was relatively higher in both spring and summer, and lower in autumn. Emission of CO2 changed minor from spring to summer, and greatly decreased in autumn in the first measured year. In the second year, the emission patterns were changed by rates of N added. N additions generally stimulated the emission of N2O, while the effects varied in different seasons and years. In addition, no obvious trends were found in the emission factor of N2O. The uptake of CH4 was not significantly affected by N additions. N additions did not change CO2 emissions in the first year, while high N significantly reduced the CO2 emissions in spring and summer of the second year, without affected in autumn. Structure equation model analysis on the factors suggested that N2O, CH4 and CO2 were dominantly affected by the N application rates, soil temperature or moisture and plant density, respectively. Over the growing seasons, both the net efflux and the global warming potential caused by N additions were small.
2010年和2011年的大气月平均温度呈现一致的变化趋势, 6-8月大气温度达到最高值(图1)。两年中大气降水有所差异, 尤其是7-8月的大气降水差异较大, 其他月份差异较小。土壤的温度和含水量呈现相反的变化趋势(图2), 各施N处理之间的变化也较一致。第一年, 从4月开始温度一直呈增加的趋势, 8月达到峰值, 随后开始缓慢降低; 第二年8月采样时的温度低于7月的温度。土壤含水量在4月和10月较高, 其他季节都较低, 大部分在0.2%- 0.4%之间。 显示原图|下载原图ZIP|生成PPT 图12010和2011年研究区域降水和气温变化。 -->Fig. 1Changes in air temperature and precipitation of the study area for 2010 and 2011. -->
显示原图|下载原图ZIP|生成PPT 图22010和2011年气体采集时各施氮(N)处理土壤温度和土壤含水量变化。N0、N0.5、N1、N3、N6和N24分别是6个样方, 以0、0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 6个不同模拟N沉降浓度进行N处理。 -->Fig. 2Changes in soil temperature and moisture at different nitrogen (N) treatments on the gas collected day in 2010 and 2011. N0, N0.5, N1, N3, N6 and N24 indicate 0, 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2 ·a-1 of simulated N deposition on the studied plots, respectively. -->
2.2 温室气体的季节变化动态
用对照样地的两年通量平均值反映荒漠未施N条件下温室气体排放动态。对通量的分析表明, N2O在春、夏、秋三季的平均值分别为5.0、6.3和 3.1 μg·m-2·h-1, 三季平均为4.8 μg·m-2·h-1。CH4在春、夏、秋三季的平均值分别为-32.9、-34.6和-23.9 μg·m-2·h-1, 三季平均为-30.5 μg·m-2·h-1。CO2在春、夏、秋三季的平均值分别为62.1、61.1和 17.1 mg·m-2·h-1, 三季平均为46.7 mg·m-2·h-1。 N2O的变化规律与N添加浓度和季节相关(表1;图3A)。2010年, 春季施N后, 高N (N24)处理样方的N2O在春季具有较高的释放速率, 在夏季降低, 秋季由于8月底的施N而再次升高。其他施N浓度的样方春季和夏季变化较为平缓, 秋季具有降低的趋势。2011年, 春季N施加后, N3和N24处理样方N2O的排放具有快速增加的趋势, 其他各处理N2O的排放在夏季达最大, 春季和秋季相对较低。 Table 1 表1 表1季节、氮(N)及其交互作用影响3种温室气体通量的重复测量的方差分析 Table 1Results of repeated measures ANOVAS for the gas efflux on the effects of seasons, nitrogen (N) treatments and their interactions
自由度 Degree of freedom
N2O
CH4
CO2
季节 Season
9
13.41**
12.40**
63.09**
N
5
53.19**
1.84
1.55
季节× N Season × N
45
5.07**
0.55
2.42**
**, p < 0.01. 新窗口打开 显示原图|下载原图ZIP|生成PPT 图32010和2011年N2O (A)、CH4 (B)、CO2 (C)通量的季节变化。N0、N0.5、N1、N3、N6和N24分别是6个样方, 以0、0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 6个不同模拟氮(N)沉降浓度进行N处理。 -->Fig. 3Seasonal changes in N2O (A), CH4 (B) and CO2 (C) effluxes in 2010 and 2011. N0, N0.5, N1, N3, N6 and N24 indicate 0, 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2·a-1 of simulated nitrogen (N) deposition on the studied plots, respectively. -->
N2O排放随着N浓度的增加具有增加的趋势(表1; 图4)。5月, 2010年各N处理之间差异不显著, 2011年N3、N6和N24处理显著高于N0、N0.5和N1处理(图4A)。8月, 2010年N6和N24处理显著高于N0、N0.5和N1处理, 2011年各N处理之间差异不显著(图4B)。10月, 2010年N24处理显著高于其他各处理, 2011年N排放无明显规律, 但以N24处理最高(图4C)。N2O的排放因子在各处理间无明显的规律(表2)。 CH4吸收受N影响较小(表1; 图5A、5B、5C), 2010和2011年两年中5月和10月, 各N处理之间差异不显著。仅2011年8月各处理之间具有显著差异。 显示原图|下载原图ZIP|生成PPT 图4不同N浓度添加下N2O通量(平均值±标准误差)。A, 5月。B, 8月。C, 10月。*表示两年间差异显著。不同小写字母表示同一年不同处理间差异显著。N0、N0.5、N1、N3、N6和N24分别是6个样方, 以0、0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 6个不同模拟氮(N)沉降浓度进行N处理。 -->Fig. 4N2O effluxes under different concentration nitrogen (N) treatments (mean ± SE). A, May. B, August. C, October. * indicates significant differences between two years. Different lowercase letters indicate significant differences among treatments in the same year. N0, N0.5, N1, N3, N6 and N24 indicate 0, 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2 ·a-1 of simulated N deposition on the studied plots, respectively. -->
显示原图|下载原图ZIP|生成PPT 图5不同N浓度添加下CH4吸收速率(平均值±标准误差)。A, 5月。B, 8月。C, 10月。*表示两年间差异显著。不同小写字母表示同一年不同处理间差异显著。N0、N0.5、N1、N3、N6和N24分别是6个样方, 以0、0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 6个不同模拟氮(N)沉降浓度进行N处理。 -->Fig. 5CH4 effluxes under different nitrogen (N) treatments (mean± SE). A, May. B, August. C, October. * indicates significant differences between two years. Different lowercase letters indicate significant differences among treatments in the same year. N0, N0.5, N1, N3, N6 and N24 indicate 0, 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2·a-1 of simulated N deposition on the plots, respectively. -->
Table 2 表2 表2N2O在各个处理的排放因子 Table 2Emission factor of N2O in different nitrogen (N)-added treatments
年 Year
N0.5
N1
N3
N6
N24
2010
0.05
0.11
0.09
0.16
0.11
2011
-0.33
0
0.03
0.04
0.03
N0.5, N1, N3, N6 and N24 indicate 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2·a-1 of simulated N deposition on the plots, respectively.N0.5、N1、N3、N6和N24分别是5个样方, 以0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 5个不同模拟氮(N)沉降浓度进行N处理。 新窗口打开 CO2的排放受N的影响也较小(表1; 图6)。5月, 2010年各处理之间差异不显著, 2011年随着N浓度的增加, CO2的通量在N3达到最大, 随后降低(图6A)。8月, 2010年CO2排放差异不显著, 2011年N0.5和N1处理CO2排放最高, 随后降低(图6B)。10月, 两年中各处理之间CO2排放差异不显著(图6C)。 显示原图|下载原图ZIP|生成PPT 图6不同N浓度添加下CO2排放速率(平均值±标准误差)。A, 5月。B, 8月。C, 10月。* 表示两年间差异显著。不同小写字母表示同一年不同处理间差异显著。N0、N0.5、N1、N3、N6和N24分别是6个样方, 以0、0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 6个不同模拟氮(N)沉降浓度进行N处理。 -->Fig. 6CO2 effluxes under different nitrogen (N) treatments (mean ± SE). A, May. B, August. C, October. * indicates significant differences between two years. Different lowercase letters indicate significant differences among treatments in the same year. N0, N0.5, N1, N3, N6 and N24 indicate 0, 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2·a-1 of simulated N deposition on the plots, respectively. -->
2.4 N增加的净效应及增温潜力
整个生长季N增加样方温室气体与对照相比, 仅N2O的比值随着N增加呈增加趋势, CH4与CO2的排放比值均在1左右(表3)。由温室气体排放差异所带来的全球增温潜力较小。 Table 3 表3 表3生长季各氮(N)处理的温室气体通量和增温潜力与对照(N0)之间的比值 Table 3The ratios of greenhouse gas effluxes and global warming potentials of nitrogen (N) added plots (N0.5, N1, N3, N6, N24) to controls (N0) during growing seasons
氮处理 N treatment
2010
2011
N2O
CH4
CO2
增温潜力 GWP
N2O
CH4
CO2
增温潜力 GWP
N0.5
1.02
0.92
0.98
0.99
0.89
0.89
0.91
0.91
N1
1.08
0.98
1.04
1.04
1.00
1.06
1.03
1.03
N3
1.19
0.88
1.14
1.15
1.07
0.86
0.91
0.94
N6
1.67
1.03
1.27
1.29
1.15
1.13
0.72
1.06
N24
2.81
0.90
1.06
1.12
1.48
1.00
0.55
0.92
N0.5, N1, N3, N6 and N24 indicate 0.5, 1.0, 3.0, 6.0 and 24.0 g·m-2·a-1 of simulated N deposition on the plots, respectively. GWP, global warming potential.N0.5、N1、N3、N6和N24分别是5个样方, 以0.5、1.0、3.0、6.0和24.0 g·m-2·a-1 5个不同模拟氮(N)沉降浓度进行N处理。 新窗口打开
2.5 温室气体与各因子之间的关系
SEM模型分析表明, 模型对N2O、CH4和CO2的解释量分别达到了56%、43%和70% (图7)。在N2O的模型中, N2O的变化受N施加浓度的影响最大, 其载重达到了0.77, 其次是植物密度(0.21)。地上或地下生物量对N2O影响不显著(图7A)。土壤温度或土壤含水量通过显著影响植株密度而间接影响N2O。在CH4模型中, 影响最大的是土壤温度或土壤含水量, 其次是生物量。N浓度和土壤温度或土壤含水量通过显著影响生物量而影响CH4的通量(图7B)。此外, 植株密度也能对CH4产生较大的影响。在CO2模型中, 植株密度和土壤温度是影响CO2动态的主要因子, 生物量对CO2的影响未达到显著水平(图7C)。 显示原图|下载原图ZIP|生成PPT 图7结构方程模型模拟生物和环境因子对温室气体的影响。A, N2O (χ2 = 0.00, p = 0.988; RMSEA = 0.000, p = 0.989)。B, CH4 (χ2 = 0.00, p = 0.988; RMSEA = 0.000, p = 0.989)。C, CO2 (χ2 = 5.12, p = 0.402; RMSEA = 0.020, p = 0.487)。其中, 实线指效应为正, 虚线指效应为负。箭头不同粗细与箭头旁载重系数成正比。RMSEA, 近似误差均方根。*, p < 0.05; **, p < 0.01。 -->Fig. 7Final fitted structural equation models depicting relative effects of biotic and abiotic variables on the effulxes of greenhouse gas. A, N2O (χ2 = 0.00, p = 0.988; RMSEA = 0.000, p = 0.989). B, CH4 (χ2 = 0.00, p = 0.988; RMSEA = 0.000, p = 0.989). C, CO2 (χ2 = 5.12, p = 0.402; RMSEA = 0.020, p = 0.487). Continuous and dashed arrows indicate positve and negative relationships, respectively. Arrow width is scaled to be proportional to path coefficients which appear adjacent to arrows. RMSEA, root mean square error of approximation. *, p < 0.05; **, p < 0.01. -->
3 讨论
3.1 N2O排放及其对N沉降的响应
古尔班通古特沙漠对照样地N2O的排放为该荒漠土壤的本底排放, 其通量(4.8 μg·m-2·h-1)与腾格里沙漠生物土壤结皮覆盖地的通量(3.2 μg·m-2·h-1)差异不大, 但后者季节变化不明显(徐冰鑫等, 2014)。内蒙古荒漠草原的研究也表明, N2O的排放在较冷的冬季(4.1 μg·m-2·h-1)小于其他季节的平均值(6.3 μg·m-2·h-1), 且不受施N (10 g N·m-2·a-1)的影响(Wang et al., 2011)。在青藏高原高山荒漠区, N2O的排放春季为0.7-1.1 μg·m-2·h-1, 夏季在1.2-1.9 μg·m-2·h-1之间, 秋季非常低, 春夏秋三季的排放值均远低于古尔班通古特沙漠等低海拔区域(Li et al., 2015b)。在土壤极端低温、低水分和低N的北极, 土壤也有可能作为N2O的库, 在一定的光照条件下吸收部分N2O, 但是不同植被覆盖条件下具有差异(Stewart et al., 2012)。因此, 荒漠N2O的排放也与其荒漠本身的立地条件密切相关。 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致。一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低。高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低。这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中。N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化。本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点。N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012)。土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生。一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015)。然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性。因此推断, 在该研究中, N2O的主要来源为硝化作用。在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加。然而, N2O的排放同时也受其他因子的影响。例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010)。通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用。同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性。不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征。沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态。因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著。
3.2 CH4吸收及其对N沉降的响应
在植被越稀少的区域, 土壤年CH4吸收量越高, 一般为荒漠>灌丛地>草地>森林(Zhuang et al., 2013)。荒漠土壤是陆地吸收CH4的重要组成部分, 荒漠土壤吸收CH4的测量值多数在10-38 μg·m-2·h-1之间(Striegl et al., 1992)。然而, CH4吸收随生态系统和区域变化非常大(Zhuang et al., 2013; Wang et al., 2014)。古尔班通古特沙漠对照样地吸收量在23-35 μg·m-2·h-1之间, 与全球大多数研究结果相近。 在腾格里沙漠生物土壤结皮覆盖区, 大部分情况下土壤为CH4汇, 平均值约为0.035 μg·m-2·h-1 (徐冰鑫等, 2014), 远低于古尔班通古特沙漠。内蒙古荒漠草原的平均吸收速率为46.4 μg·m-2·h-1, 夏季具有最高的CH4吸收速率, 其吸收不受N的影响(Wang et al., 2011)。在高山荒漠区域, 春季和秋季可能为CH4的释放季, 仅夏季表现为CH4的吸收, 且吸收值在4-8 μg·m-2·h-1之间(Li et al., 2015b)。北极荒漠区整个生长季CH4的吸收速率可达57.1 μg·m-2·h-1 (Emmerton et al., 2014), 高于其他荒漠类型。 在大部分季节, CH4的吸收不受N增加的影响, 这与亚热带种植园土壤以及内蒙古退化草地N添加后的CH4变化相一致(Chen et al., 2013; Li et al., 2015a)。然而, N增加通常抑制CH4的氧化, N对CH4单加氧酶具有竞争抑制作用, N转化过程生成的NH2OH和NO3-, 或者高浓度NH4+或NO3-带来的渗透调节压力也具有毒害抑制作用(Castro et al., 1995; Bodelier, 2011)。在全球尺度上, N沉降能在一定程度上降低土壤作为CH4库的能力(Zhuang et al., 2013)。在本研究中, 有大量的NH4+和NO3-输入, 然而与对照相比, 并没有大的差异, 因此该研究中N的增加并不是一个直接的驱动因子。N对CH4通量无显著影响的原因尚不明确, 可能是由于与CH4相关的微生物活动受其他环境因子的影响更大, 而掩盖了N效应造成。模型结果也表明, 能够对CH4产生最大影响的是非生物因子土壤温度或土壤含水量。例如, 秋季CH4吸收的减少, 可能是由秋季土壤含水量的增加所导致。以前的研究表明, 土壤含水量能够通过影响O2的浓度而影响CH4氧化细菌活性(Konda et al., 2010)。土壤温度可能影响CH4氧化细菌活性, 使得CH4氧化细菌对CH4的变化具有显著作用。另外, 土壤温度还可通过植物而间接影响CH4的吸收。
3.3 CO2排放及对N沉降的响应
古尔班通古特沙漠生长季CO2排放的平均值约为46.7 mg·m-2·h-1, 春夏季排放速率约是秋季的3.5倍。腾格里沙漠生物土壤结皮覆盖地CO2的年平均值约46.7 mg·m-2·h-1, 与古尔班通古特沙漠生长季平均值一致, 且季节变化明显(徐冰鑫等, 2014)。同样, 对内蒙古荒漠草原的研究表明, 其CO2排放的平均值在夏季(109 mg·m-2·h-1)远高于其他季节 (32 mg·m-2·h-1), 且不受加N的影响(Wang et al., 2011)。青藏高原高山荒漠的春、夏、秋三季CO2排放量约为14、30和18 mg·m-2·h-1 (Li et al., 2015b), 春、夏季排放量远低于古尔班通古特沙漠和腾格里沙漠。这些差异可能是由研究区域温度和植被覆盖等因素造成。 CO2的排放主要包括土壤呼吸和植物呼吸, N对CO2的排放影响也随着季节不同具有差异性。2010年, 在生长旺季的5月和8月N对CO2的排放影响不显著可能是土壤和植物综合响应的结果。2011年, 植物和土壤呼吸通量的平衡被打破, N浓度变化带来的效应开始出现, 尤其是高N浓度对CO2的释放通量显著抑制。已有研究表明, 高N处理下土壤呼吸被抑制(Zhou & Zhang, 2014), 加上植物多样性降低, 造成CO2的排放较低。在秋季, 大部分植株死亡, 土壤温度也较低, 使得土壤呼吸变化不明显, 导致系统CO2的排放各处理差异不显著。模型结果表明, 显著影响CO2排放的主要有植株密度、土壤温度和土壤有机碳。因此, 生长季植物数量的变化对CO2的排放影响最大。 根据整个生长季(4月末至10月末)通量估算, 古尔班通古特沙漠(面积4.88 × 1010 m2) 3种温室气体交换的估算值分别是: N2O排放约为1.12 × 103 t, CH4吸收约为6.49 × 103 t, CO2排放约为1.03 × 107 t。其中CH4的吸收量与撒哈拉大沙漠对CH4的吸收量(1.36 × 106 t)相距甚远, 占中国年CH4估计吸收量(2.78 × 106 t)的0.23% (Zhuang et al., 2013)。该沙漠生长季N2O的排放量约占全国年排放量(2.15 × 106 t)的0.52% (Zhou et al., 2014)。然而, 这些数据的粗略估算需要荒漠更多点位的数据加以验证。与对照相比, N带来的3种通量的净变化除N2O外均较小, 增温潜力的比值也在1左右, 说明在荒漠土壤, 由N效应带来的净增温效应有限。同样, 对高山草甸的研究也表明, 短期的N增加不能影响净增温潜力(Jiang et al., 2010)。也有研究表明, 荒漠系统中增温对土壤系统温室气体的交换影响不大, 对气候变暖的贡献较小(徐冰鑫等, 2014)。并且, 对荒漠的碳通量研究表明, 荒漠系统本身是一个碳库, 能够储存碳(Wohlfahrt et al., 2008)。因此, 荒漠土壤在全球变化条件下可能不是全球变暖的一个重要贡献者。 The authors have declared that no competing interests exist.
AdamsMB (2003). Ecological issues related to N deposition to natural ecosystems: Research needs. Environment International, 29, 189-199. [本文引用: 1]
[2]
BaezS, FargioneJ, MooreDI, CollinsSL, GoszJR (2007). Atmospheric nitrogen deposition in the northern Chihu¬ahuan desert: Temporal trends and potential consequences. Journal of Arid Environments, 68, 640-651. [本文引用: 1]
[3]
BaiYF, WuJG, ClarkCM, NaeemS, PanQM, HuangJH, ZhangLX, HanXG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia Grasslands. Global Change Biology, 16, 358-372. [本文引用: 1]
[4]
Bellingrath-KimuraSD, Kishimoto-MoAW, OuraN, SekikawaS, YonemuraS, SudoS, HayakawaA, MinamikawaK, TakataY, HaraH (2015). Differences in the spatial variability among CO2, CH4, and N2O gas fluxes from an urban forest soil in Japan. Ambio, 44, 55-66. [本文引用: 1]
[5]
BodelierPLE (2011). Interactions between nitrogenous fertil¬izers and methane cycling in wetland and upland soils. Current Opinion in Environmental Sustainability, 3, 379-388. [本文引用: 1]
[6]
CanadellJG, KirschbaumMUF, KurzWA, SanzMJ, SchlamadingerB, YamagataY (2007). Factoring out natural and indirect human effects on terrestrial carbon sources and sinks. Environmental Science & Policy, 10, 370-384. [本文引用: 1]
ChenW, ZhengX, ChenQ, WolfB, Butterbach-BahlK, BrueggemannN, LinS (2013). Effects of increasing precipitation and nitrogen deposition on CH4 and N2O fluxes and ecosystem respiration in a degraded steppe in Inner Mongolia, China. Geoderma, 192, 335-340. [本文引用: 1]
[9]
ClarkCM, TilmanD (2008). Loss of plant species after chronic low level nitrogen deposition to prairie grasslands. Nature, 451, 712-715. [本文引用: 1]
[10]
DenmanKL, BrasseurGP, ChidthaisongA, CiaisP, CoxPM, DickinsonRE, HauglustaineDA, HeinzeC, HollandEA, JacobDJ, LohmannU, RamachandranS, Leite da Silva Dias P, Wofsy SC, Zhang X, Steffen W (2007). Couplings between changes in the climate system and biogeoche¬mistry. In: Solomon S, Qin D, Manning M, Chen Z, Mar¬quis M eds. Climate Change 2007: The Physical Science Basis Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. 501-564. [本文引用: 1]
[11]
EmmertonCA, St LouisVL, LehnherrI, HumphreysER, RydzE, KosolofskiHR (2014). The net exchange of methane with high Arctic landscapes during the summer growing season. Biogeosciences, 11, 3095-3106. [本文引用: 1]
[12]
ForsterP, RamaswamyV, ArtaxoP, BerntsenT, BettsR, FaheyDW, HaywoodJ, LeanJ, LoweDC, MyhreG, NgangaJ, PrinnR, RagaG, Schulz M, van Dorland R (2007). Cha¬nges in Atmospheric Constituents and in Radiative For¬cing Chapter 2. Cambridge University Press, Cambridge, UK. [本文引用: 1]
[13]
GalballyIE, KirstineWV, MeyerCP, WangYP (2008). Soil-atmosphere trace gas exchange in semiarid and arid zones. Journal of Environmental Quality, 37, 599-607. [本文引用: 1]
[14]
GraceJB (2006). Structural Equation Modeling and Natural Systems. Cambridge University Press, Cambridge, UK. 10. [本文引用: 1]
[15]
HeCE, LiuXJ, FangmeierA, ZhangFS (2007). Quantifying the total airborne nitrogen input into agroecosystems in the North China Plain. Agriculture, Ecosystems and Environment, 121, 395-400. [本文引用: 1]
[16]
HorváthL, GroszB, MachonA, TubaZ, NagyZ, CzóbelSZ, BaloghJ, PéliE, FótiSZ, WeidingerT, PintérK, FührerE (2010). Estimation of nitrous oxide emission from Hungarian semi-arid sandy and loess grasslands: Effect of soil parameters, grazing, irrigation and use of fertilizer. Agriculture, Ecosystems & Environment, 139, 255-263. [本文引用: 1]
[17]
InclánR, UribeC, SánchezL, SánchezDM, ClaveroÁ, FernándezAM, MoranteR, BlancoA, JandlR (2012). N2O and CH4 fluxes in undisturbed and burned holm oak, scots pine and pyrenean oak forests in central Spain. Biogeochemistry, 107, 19-41. [本文引用: 1]
[18]
IPCC (Intergovernmental Panel on Climate Change) (1996). Contribution of working group I to the second assessment report of the intergovernmental panel on climate change. In: Houghton JJ, Meiro Filho LG, Callander BA, Harris N, Kattenberg A, Maskell K eds. Climate Change 1995: The Science of Climate Change. Cambridge University Press, New York. [本文引用: 2]
[19]
JassalRS, BlackTA, RoyR, EthierG (2011). Effect of nitrogen fertilization on soil CH4 and N2O fluxes, and soil and bole respiration. Geoderma, 162, 182-186. [本文引用: 2]
[20]
JiangCM, YuGR, FangHJ, CaoGM, LiYN (2010). Short-term effect of increasing nitrogen deposition on CO2, CH4 and N2O fluxes in an alpine meadow on the Qinghai Tibetan Plateau, China. Atmospheric Environment, 44, 2920-2926. [本文引用: 1]
[21]
KondaR, OhtaS, IshizukaS, HeriyantoJ, WicaksonoA (2010). Seasonal changes in the spatial structures of N2O, CO2, and CH4 fluxes from Acacia mangium plantation soils in Indonesia. Soil Biology & Biochemistry, 42, 1512-1522. [本文引用: 1]
[22]
LiKH, GongYM, SongW, HeGX, HuYK, TianCY, LiuXJ (2012). Responses of CH4, CO2 and N2O fluxes to increasing nitrogen deposition in alpine grassland of the Tianshan Mountains. Chemosphere, 88, 140-143.
[23]
LiXY, ChengSL, FangHJ, YuGR, DangXS, XuMJ, WangL, SiGY, GengJ, HeS (2015a). The contrasting effects of deposited NH4+ and NO3- on soil CO2, CH4 and N2O fluxes in a subtropical plantation, southern China. Ecological Eng¬ineering, 85, 317-327. [本文引用: 1]
[24]
LiYY, DongSK, LiuSL, ZhouHK, GaoQZ, CaoGM, WangXX, SuXK, ZhangY, TangL, ZhaoHD, WuXY (2015b). Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the Qinghai Tibetan Plateau of China. Soil Biology & Biochemistry, 80, 306-314. [本文引用: 4]
[25]
LiuL, GreaverTL (2009). A review of nitrogen enrichment effects on three biogenic GHGs: The CO2 sink may be largely offset by stimulated N2O and CH4 emission. Ecology Letters, 12, 1103-1117. [本文引用: 1]
[26]
MarisSC, Teira EsmatgesMR, ArbonesA, RufatJ (2015). Effect of irrigation, nitrogen application, and a nitrification inhibitor on nitrous oxide, carbon dioxide and methane emissions from an olive (Olea europaea L.) orchard. Science of the Total Environment, 538, 966-978.
[27]
MoJM, FangYT, LinED, LiYE (2006). Soil N2O emission and its response to simulated N deposition in the main forests of Dinghushan in subtropical China. Journal of Plant Ecology (Chinese Version), 30, 901-910. (in Chinese with English abstract)[莫江明, 方运霆, 林而达, 李玉娥 (2006). 鼎湖山主要森林土壤N2O排放及其对模拟N沉降的响应 . 植物生态学报, 30, 901-910.] [本文引用: 1]
[28]
SchaefferSM, EvansRD (2005). Pulse additions of soil carbon and nitrogen affect soil nitrogen dynamics in an arid Colorado Plateau shrubland. Oecologia, 145, 425-433. [本文引用: 1]
[29]
SchoenbachP, WolfB, DickhoeferU, WiesmeierM, ChenWW, WanHW, GierusM, Butterbach BahlK, Koegel KnabnerI, SusenbethA, ZhengXH, TaubeF (2012). Grazing effects on the greenhouse gas balance of a temperate steppe ecosystem. Nutrient Cycling in Agroecosystems, 93, 357-371. [本文引用: 1]
[30]
StewartKJ, BrummellME, FarrellRE, SicilianoSD (2012). N2O flux from plant soil systems in polar deserts switch between sources and sinks under different light conditions. Soil Biology & Biochemistry, 48, 69-77. [本文引用: 2]
[31]
StrieglRG, McconnaugheyTA, ThorstensonDC, WeeksEP, WoodwardJC (1992). Consumption of atmospheric methane by desert soils. Nature, 357, 145-147. [本文引用: 1]
[32]
VourlitisGL, PasquiniS, ZorbaG (2007). Plant and soil N response of southern californian semi arid shrublands after 1 year of experimental N deposition. Ecosystems, 10, 263-279. [本文引用: 1]
WangZW, HaoXY, ShanD, HanGD, ZhaoML, WillmsWD, WangZ, HanX (2011). Influence of increasing temperature and nitrogen input on greenhouse gas emissions from a desert steppe soil in Inner Mongolia. Soil Science and Plant Nutrition, 57, 508-518. [本文引用: 3]
[35]
WohlfahrtG, FenstermakerLF, Arnone IiiJA (2008). Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem. Global Change Biology, 14, 1475-1487. [本文引用: 1]
[36]
XuBX, HuYG, ZhangZS, ChenYL, ZhangP, LiG (2014). Effects of experimental warming on CO2, CH4 and N2O fluxes of biological soil crust and soil system in a desert region. Chinese Journal of Plant Ecology, 38, 809-820. (in Chinese with English abstract)[徐冰鑫, 胡宜刚, 张志山, 陈永乐, 张鹏, 李刚 (2014). 模拟增温对荒漠生物土壤结皮-土壤系统CO2、CH4和N2O通量的影响 . 植物生态学报, 38, 809-820.]
[37]
XuR, WangM, WangY (2003a). Using a modified DNDC model to estimate N2O fluxes from semi arid grassland in China. Soil Biology & Biochemistry, 35, 615-620. [本文引用: 6]
[38]
XuR, WangYS, ZhengXH, JiBM, WangMX (2003b). A comparison between measured and modeled N2O emissions from Inner Mongolian semi-arid grassland. Plant and Soil, 255, 513-528. [本文引用: 1]
[39]
YanJH, ZhangW, WangKY, QinF, WangWT, DaiHT, LiPX (2014). Responses of CO2, N2O and CH4 fluxes between atmosphere and forest soil to changes in multiple environmental conditions. Global Change Biology, 20, 300-312.
[40]
ZaadyE, GroffmanPM, StandingD, ShachakM (2013). High N2O emissions in dry ecosystems. European Journal of Soil Biology, 59, 1-7. [本文引用: 1]
[41]
ZhangC, NiuD, HallSJ, WenH, LiX, FuH, WanC, ElserJJ (2014). Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biology & Biochemistry, 75, 113-123. [本文引用: 1]
[42]
ZhangJB, CaiZC, ChengY, ZhuTB (2009). Denitrification and total nitrogen gas production from forest soils of Eastern China. Soil Biology & Biochemistry, 41, 2551-2557. [本文引用: 2]
[43]
ZhangW, LiuXJ, HuYK, LiKH, ShenJL, LuoXS, SongW (2011). Analysis on input of atmospheric nitrogen dry deposition in Ürümqi. Arid Zone Research, 28, 771-716. (in Chinese with English abstract)[张伟, 刘学军, 胡玉昆, 李凯辉, 沈健林, 骆晓声, 宋韦 (2011). 乌鲁木齐市区大气N素干沉降的输入性分析 . 干旱区研究, 28, 771-716.] [本文引用: 1]
[44]
ZhangY, ZhengLX, LiuXJ, JickellsT, CapeJN, GouldingK, FangmeierA, ZhangFS (2008). Evidence for organic N deposition and its anthropogenic sources in China. Atmospheric Environment, 42, 1035-1041.
[45]
ZhouF, ShangZY, CiaisP, TaoS, PiaoSL, RaymondP, HeCF, LiBG, WangRH, WangXH, PengSS, ZengZZ, ChenH, YingN, HouXK, XuP (2014). A new high-resolution N2O emission inventory for China in 2008. Environmental Science & Technology, 48, 8538-8547. [本文引用: 1]
[46]
ZhouXB, ZhangYM (2014). Seasonal pattern of soil respiration and gradual changing effects of nitrogen addition in a soil of the Gurbantünggüt Desert, northwestern China. Atmospheric Environment, 85, 187-194. [本文引用: 1]
[47]
ZhouXB, ZhangYM, DowningA (2012). Non linear response of microbial activity across a gradient of nitrogen addition to a soil from the Gurbantünggüt Desert, northwestern China. Soil Biology & Biochemistry, 47, 67-77. [本文引用: 1]
[48]
ZhuangQL, ChenM, XuK, TangJY, SaikawaE, LuYY, MelilloJM, PrinnRG, McGuireAD (2013). Response of global soil consumption of atmospheric methane to changes in atmospheric climate and nitrogen deposition. Global Biogeochemical Cycles, 27, 650-663. [本文引用: 2]
Ecological issues related to N deposition to natural ecosystems: Research needs. 1 2003
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
Atmospheric nitrogen deposition in the northern Chihu¬ahuan desert: Temporal trends and potential consequences. 1 2007
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia Grasslands. 1 2010
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
Differences in the spatial variability among CO2, CH4, and N2O gas fluxes from an urban forest soil in Japan. 1 2015
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
Interactions between nitrogenous fertil¬izers and methane cycling in wetland and upland soils. 1 2011
... 在大部分季节, CH4的吸收不受N增加的影响, 这与亚热带种植园土壤以及内蒙古退化草地N添加后的CH4变化相一致(Chen et al., 2013; Li et al., 2015a).然而, N增加通常抑制CH4的氧化, N对CH4单加氧酶具有竞争抑制作用, N转化过程生成的NH2OH和NO3-, 或者高浓度NH4+或NO3-带来的渗透调节压力也具有毒害抑制作用(Castro et al., 1995; Bodelier, 2011).在全球尺度上, N沉降能在一定程度上降低土壤作为CH4库的能力(Zhuang et al., 2013).在本研究中, 有大量的NH4+和NO3-输入, 然而与对照相比, 并没有大的差异, 因此该研究中N的增加并不是一个直接的驱动因子.N对CH4通量无显著影响的原因尚不明确, 可能是由于与CH4相关的微生物活动受其他环境因子的影响更大, 而掩盖了N效应造成.模型结果也表明, 能够对CH4产生最大影响的是非生物因子土壤温度或土壤含水量.例如, 秋季CH4吸收的减少, 可能是由秋季土壤含水量的增加所导致.以前的研究表明, 土壤含水量能够通过影响O2的浓度而影响CH4氧化细菌活性(Konda et al., 2010).土壤温度可能影响CH4氧化细菌活性, 使得CH4氧化细菌对CH4的变化具有显著作用.另外, 土壤温度还可通过植物而间接影响CH4的吸收. ...
Factoring out natural and indirect human effects on terrestrial carbon sources and sinks. 1 2007
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
... 在大部分季节, CH4的吸收不受N增加的影响, 这与亚热带种植园土壤以及内蒙古退化草地N添加后的CH4变化相一致(Chen et al., 2013; Li et al., 2015a).然而, N增加通常抑制CH4的氧化, N对CH4单加氧酶具有竞争抑制作用, N转化过程生成的NH2OH和NO3-, 或者高浓度NH4+或NO3-带来的渗透调节压力也具有毒害抑制作用(Castro et al., 1995; Bodelier, 2011).在全球尺度上, N沉降能在一定程度上降低土壤作为CH4库的能力(Zhuang et al., 2013).在本研究中, 有大量的NH4+和NO3-输入, 然而与对照相比, 并没有大的差异, 因此该研究中N的增加并不是一个直接的驱动因子.N对CH4通量无显著影响的原因尚不明确, 可能是由于与CH4相关的微生物活动受其他环境因子的影响更大, 而掩盖了N效应造成.模型结果也表明, 能够对CH4产生最大影响的是非生物因子土壤温度或土壤含水量.例如, 秋季CH4吸收的减少, 可能是由秋季土壤含水量的增加所导致.以前的研究表明, 土壤含水量能够通过影响O2的浓度而影响CH4氧化细菌活性(Konda et al., 2010).土壤温度可能影响CH4氧化细菌活性, 使得CH4氧化细菌对CH4的变化具有显著作用.另外, 土壤温度还可通过植物而间接影响CH4的吸收. ...
Effects of increasing precipitation and nitrogen deposition on CH4 and N2O fluxes and ecosystem respiration in a degraded steppe in Inner Mongolia, China. 1 2013
... 在大部分季节, CH4的吸收不受N增加的影响, 这与亚热带种植园土壤以及内蒙古退化草地N添加后的CH4变化相一致(Chen et al., 2013; Li et al., 2015a).然而, N增加通常抑制CH4的氧化, N对CH4单加氧酶具有竞争抑制作用, N转化过程生成的NH2OH和NO3-, 或者高浓度NH4+或NO3-带来的渗透调节压力也具有毒害抑制作用(Castro et al., 1995; Bodelier, 2011).在全球尺度上, N沉降能在一定程度上降低土壤作为CH4库的能力(Zhuang et al., 2013).在本研究中, 有大量的NH4+和NO3-输入, 然而与对照相比, 并没有大的差异, 因此该研究中N的增加并不是一个直接的驱动因子.N对CH4通量无显著影响的原因尚不明确, 可能是由于与CH4相关的微生物活动受其他环境因子的影响更大, 而掩盖了N效应造成.模型结果也表明, 能够对CH4产生最大影响的是非生物因子土壤温度或土壤含水量.例如, 秋季CH4吸收的减少, 可能是由秋季土壤含水量的增加所导致.以前的研究表明, 土壤含水量能够通过影响O2的浓度而影响CH4氧化细菌活性(Konda et al., 2010).土壤温度可能影响CH4氧化细菌活性, 使得CH4氧化细菌对CH4的变化具有显著作用.另外, 土壤温度还可通过植物而间接影响CH4的吸收. ...
Loss of plant species after chronic low level nitrogen deposition to prairie grasslands. 1 2008
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
1
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
The net exchange of methane with high Arctic landscapes during the summer growing season. 1 2014
... 在腾格里沙漠生物土壤结皮覆盖区, 大部分情况下土壤为CH4汇, 平均值约为0.035 μg·m-2·h-1 (徐冰鑫等, 2014), 远低于古尔班通古特沙漠.内蒙古荒漠草原的平均吸收速率为46.4 μg·m-2·h-1, 夏季具有最高的CH4吸收速率, 其吸收不受N的影响(Wang et al., 2011).在高山荒漠区域, 春季和秋季可能为CH4的释放季, 仅夏季表现为CH4的吸收, 且吸收值在4-8 μg·m-2·h-1之间(Li et al., 2015b).北极荒漠区整个生长季CH4的吸收速率可达57.1 μg·m-2·h-1 (Emmerton et al., 2014), 高于其他荒漠类型. ...
1 2007
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
Soil-atmosphere trace gas exchange in semiarid and arid zones. 1 2008
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
Estimation of nitrous oxide emission from Hungarian semi-arid sandy and loess grasslands: Effect of soil parameters, grazing, irrigation and use of fertilizer. Agriculture, 1 2010
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
N2O and CH4 fluxes in undisturbed and burned holm oak, scots pine and pyrenean oak forests in central Spain. 1 2012
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
2
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
Effect of nitrogen fertilization on soil CH4 and N2O fluxes, and soil and bole respiration. 2 2011
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
Short-term effect of increasing nitrogen deposition on CO2, CH4 and N2O fluxes in an alpine meadow on the Qinghai Tibetan Plateau, China. 1 2010
... 在大部分季节, CH4的吸收不受N增加的影响, 这与亚热带种植园土壤以及内蒙古退化草地N添加后的CH4变化相一致(Chen et al., 2013; Li et al., 2015a).然而, N增加通常抑制CH4的氧化, N对CH4单加氧酶具有竞争抑制作用, N转化过程生成的NH2OH和NO3-, 或者高浓度NH4+或NO3-带来的渗透调节压力也具有毒害抑制作用(Castro et al., 1995; Bodelier, 2011).在全球尺度上, N沉降能在一定程度上降低土壤作为CH4库的能力(Zhuang et al., 2013).在本研究中, 有大量的NH4+和NO3-输入, 然而与对照相比, 并没有大的差异, 因此该研究中N的增加并不是一个直接的驱动因子.N对CH4通量无显著影响的原因尚不明确, 可能是由于与CH4相关的微生物活动受其他环境因子的影响更大, 而掩盖了N效应造成.模型结果也表明, 能够对CH4产生最大影响的是非生物因子土壤温度或土壤含水量.例如, 秋季CH4吸收的减少, 可能是由秋季土壤含水量的增加所导致.以前的研究表明, 土壤含水量能够通过影响O2的浓度而影响CH4氧化细菌活性(Konda et al., 2010).土壤温度可能影响CH4氧化细菌活性, 使得CH4氧化细菌对CH4的变化具有显著作用.另外, 土壤温度还可通过植物而间接影响CH4的吸收. ...
Seasonal changes in the spatial structures of N2O, CO2, and CH4 fluxes from Acacia mangium plantation soils in Indonesia. 1 2010
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
Responses of CH4, CO2 and N2O fluxes to increasing nitrogen deposition in alpine grassland of the Tianshan Mountains. 2012
a). The contrasting effects of deposited NH4+ and NO3- on soil CO2, CH4 and N2O fluxes in a subtropical plantation, southern China. 1 2015
... 在大部分季节, CH4的吸收不受N增加的影响, 这与亚热带种植园土壤以及内蒙古退化草地N添加后的CH4变化相一致(Chen et al., 2013; Li et al., 2015a).然而, N增加通常抑制CH4的氧化, N对CH4单加氧酶具有竞争抑制作用, N转化过程生成的NH2OH和NO3-, 或者高浓度NH4+或NO3-带来的渗透调节压力也具有毒害抑制作用(Castro et al., 1995; Bodelier, 2011).在全球尺度上, N沉降能在一定程度上降低土壤作为CH4库的能力(Zhuang et al., 2013).在本研究中, 有大量的NH4+和NO3-输入, 然而与对照相比, 并没有大的差异, 因此该研究中N的增加并不是一个直接的驱动因子.N对CH4通量无显著影响的原因尚不明确, 可能是由于与CH4相关的微生物活动受其他环境因子的影响更大, 而掩盖了N效应造成.模型结果也表明, 能够对CH4产生最大影响的是非生物因子土壤温度或土壤含水量.例如, 秋季CH4吸收的减少, 可能是由秋季土壤含水量的增加所导致.以前的研究表明, 土壤含水量能够通过影响O2的浓度而影响CH4氧化细菌活性(Konda et al., 2010).土壤温度可能影响CH4氧化细菌活性, 使得CH4氧化细菌对CH4的变化具有显著作用.另外, 土壤温度还可通过植物而间接影响CH4的吸收. ...
b). Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the Qinghai Tibetan Plateau of China. 4 2015
... 古尔班通古特沙漠对照样地N2O的排放为该荒漠土壤的本底排放, 其通量(4.8 μg·m-2·h-1)与腾格里沙漠生物土壤结皮覆盖地的通量(3.2 μg·m-2·h-1)差异不大, 但后者季节变化不明显(徐冰鑫等, 2014).内蒙古荒漠草原的研究也表明, N2O的排放在较冷的冬季(4.1 μg·m-2·h-1)小于其他季节的平均值(6.3 μg·m-2·h-1), 且不受施N (10 g N·m-2·a-1)的影响(Wang et al., 2011).在青藏高原高山荒漠区, N2O的排放春季为0.7-1.1 μg·m-2·h-1, 夏季在1.2-1.9 μg·m-2·h-1之间, 秋季非常低, 春夏秋三季的排放值均远低于古尔班通古特沙漠等低海拔区域(Li et al., 2015b).在土壤极端低温、低水分和低N的北极, 土壤也有可能作为N2O的库, 在一定的光照条件下吸收部分N2O, 但是不同植被覆盖条件下具有差异(Stewart et al., 2012).因此, 荒漠N2O的排放也与其荒漠本身的立地条件密切相关. ...
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
... 在腾格里沙漠生物土壤结皮覆盖区, 大部分情况下土壤为CH4汇, 平均值约为0.035 μg·m-2·h-1 (徐冰鑫等, 2014), 远低于古尔班通古特沙漠.内蒙古荒漠草原的平均吸收速率为46.4 μg·m-2·h-1, 夏季具有最高的CH4吸收速率, 其吸收不受N的影响(Wang et al., 2011).在高山荒漠区域, 春季和秋季可能为CH4的释放季, 仅夏季表现为CH4的吸收, 且吸收值在4-8 μg·m-2·h-1之间(Li et al., 2015b).北极荒漠区整个生长季CH4的吸收速率可达57.1 μg·m-2·h-1 (Emmerton et al., 2014), 高于其他荒漠类型. ...
... 古尔班通古特沙漠生长季CO2排放的平均值约为46.7 mg·m-2·h-1, 春夏季排放速率约是秋季的3.5倍.腾格里沙漠生物土壤结皮覆盖地CO2的年平均值约46.7 mg·m-2·h-1, 与古尔班通古特沙漠生长季平均值一致, 且季节变化明显(徐冰鑫等, 2014).同样, 对内蒙古荒漠草原的研究表明, 其CO2排放的平均值在夏季(109 mg·m-2·h-1)远高于其他季节 (32 mg·m-2·h-1), 且不受加N的影响(Wang et al., 2011).青藏高原高山荒漠的春、夏、秋三季CO2排放量约为14、30和18 mg·m-2·h-1 (Li et al., 2015b), 春、夏季排放量远低于古尔班通古特沙漠和腾格里沙漠.这些差异可能是由研究区域温度和植被覆盖等因素造成. ...
A review of nitrogen enrichment effects on three biogenic GHGs: The CO2 sink may be largely offset by stimulated N2O and CH4 emission. 1 2009
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
Effect of irrigation, nitrogen application, and a nitrification inhibitor on nitrous oxide, carbon dioxide and methane emissions from an olive (Olea europaea L.) orchard. 2015
鼎湖山主要森林土壤N2O排放及其对模拟N沉降的响应 1 2006
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
Pulse additions of soil carbon and nitrogen affect soil nitrogen dynamics in an arid Colorado Plateau shrubland. 1 2005
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
Grazing effects on the greenhouse gas balance of a temperate steppe ecosystem. 1 2012
... 氧化亚氮(N2O)、甲烷(CH4)和CO2是引起全球变暖的3种主要温室气体.近年来, 3种气体分别以每年0.3%、0.9%和0.5%的速率增加, 它们对全球变暖的贡献占到了87% (Forster et al., 2007).在100年的尺度上, N2O和CH4的增温潜力分别是CO2的298倍和25倍(IPCC, 1996; Denman et al., 2007).土壤能够产生和消耗温室气体, 是调节陆地生态系统中大气温室气体含量的主要贡献者(Canadell et al., 2007).然而, 土壤和大气界面的温室气体的交换变化非常大, 许多环境因子参与其中, 土壤温度、土壤含水量和pH值均有可能成为不同系统的主要驱动因子(Yan et al., 2014; Bellingrath-Kimura et al., 2015).有研究表明, 干旱半干旱区土壤-大气界面的CH4、CO和N2O等交换可以占到全球土壤-大气界面交换的5%-40% (Schoenbach et al., 2012; Zaady et al., 2013), 然而, 目前干旱半干旱区气体交换与环境变化关系的定量化研究还较少(Galbally et al., 2008). ...
N2O flux from plant soil systems in polar deserts switch between sources and sinks under different light conditions. 2 2012
... 古尔班通古特沙漠对照样地N2O的排放为该荒漠土壤的本底排放, 其通量(4.8 μg·m-2·h-1)与腾格里沙漠生物土壤结皮覆盖地的通量(3.2 μg·m-2·h-1)差异不大, 但后者季节变化不明显(徐冰鑫等, 2014).内蒙古荒漠草原的研究也表明, N2O的排放在较冷的冬季(4.1 μg·m-2·h-1)小于其他季节的平均值(6.3 μg·m-2·h-1), 且不受施N (10 g N·m-2·a-1)的影响(Wang et al., 2011).在青藏高原高山荒漠区, N2O的排放春季为0.7-1.1 μg·m-2·h-1, 夏季在1.2-1.9 μg·m-2·h-1之间, 秋季非常低, 春夏秋三季的排放值均远低于古尔班通古特沙漠等低海拔区域(Li et al., 2015b).在土壤极端低温、低水分和低N的北极, 土壤也有可能作为N2O的库, 在一定的光照条件下吸收部分N2O, 但是不同植被覆盖条件下具有差异(Stewart et al., 2012).因此, 荒漠N2O的排放也与其荒漠本身的立地条件密切相关. ...
... 本研究发现, N的增加能够促进N2O的排放, 该研究结果与大部分的研究结果(Liu & Greaver, 2009; Jassal et al., 2011; Li et al., 2012)一致.一般来说, N0、 N0.5和N1等低浓度的N处理N2O的排放在春季和夏季较高, 秋季较低.高浓度的N处理受N施加季节的影响, N添加季N2O高, 随后降低.这些结果表明, 在荒漠地区, N的脉冲带来的N, 部分能够经过土壤过程以N2O的形式重新回到大气中.N2O季节动态在低浓度的N脉冲条件下不随脉冲发生时间点而变化.本研究中春夏生长季N2O排放较高, 也与Li等(2015b)的结果相一致, 认为高山草原N2O呈现生长期比生长早期和非生长期要高的特点.N2O的排放通常受微生物参与的两个过程——硝化和反硝化作用影响, 这些过程都与温度和水分相关(Inclán et al., 2012).土壤的高含水量不仅能够刺激土壤微生物活性, 而且能够降低土壤中O2的流动, 导致反硝化作用产生.一些研究认为, 在森林系统中, 土壤反硝化作用主要控制N2O的产生和排放(Zhang et al., 2009; Maris et al., 2015).然而, 在本研究中, 整个生长季土壤水分处于较低含量水平, 并未达到田间含水量(约16%), 并且沙质土壤具有较好的O2流动性.因此推断, 在该研究中, N2O的主要来源为硝化作用.在施N后, NH4+的输入也为硝化作用快速增加提供了大量的基质, 导致增N样方的N2O在一定时期内增加.然而, N2O的排放同时也受其他因子的影响.例如, 植物能够吸收N2O (Stewart et al., 2012),一些研究也表明土壤温度对N2O排放也具有重要作用(莫江明等, 2006; Horváth et al., 2010).通过模型分析可以看出, N添加在N2O的动态变化中具有最重要的作用.同时, 植物多样性的变化也能影响N2O, 从而增加了N2O排放的不确定性.不同季节植物密度相差很大, 因此造成了N2O排放具有明显的季节特征.沙漠土壤的非生物因子土壤温度和水分可以通过影响植株密度而影响N2O动态.因此, 年际间各N处理间N排放的差异性可能不同, 如在2010年5月和2011年7月差异不显著. ...
Consumption of atmospheric methane by desert soils. 1 1992
... 在植被越稀少的区域, 土壤年CH4吸收量越高, 一般为荒漠>灌丛地>草地>森林(Zhuang et al., 2013).荒漠土壤是陆地吸收CH4的重要组成部分, 荒漠土壤吸收CH4的测量值多数在10-38 μg·m-2·h-1之间(Striegl et al., 1992).然而, CH4吸收随生态系统和区域变化非常大(Zhuang et al., 2013; Wang et al., 2014).古尔班通古特沙漠对照样地吸收量在23-35 μg·m-2·h-1之间, 与全球大多数研究结果相近. ...
Plant and soil N response of southern californian semi arid shrublands after 1 year of experimental N deposition. 1 2007
... 随着全球变化的加剧, 氮(N)沉降的增加日益成为影响干旱区生态系统结构和功能的重要因子(Baez et al., 2007; Bai et al., 2010).干旱半干旱区占全球陆地面积的1/3, 通常受N限制, 但是该区域消耗N的能力有限(Vourlitis et al., 2007; Clark & Tilman, 2008).干旱区少量的N增加能够带来比其他类型生态系统更大的影响(Adams, 2003).干旱区的一些生物土壤化学过程, 如营养转化、微生物活动和气体交换等对N增加较敏感(Schaeffer & Evans, 2005; Zhou et al., 2012; Zhang et al., 2014).目前, 在干旱区大部分研究集中于CO2如何响应N增加(Zhang et al., 2014; Zhou & Zhang, 2014), 而针对N增加条件下N2O和CH4的通量研究较为缺乏.N沉降的增加通常能刺激N2O的排放, 部分抵消和平衡土壤界面N总量的增加(Xu et al., 2003a, 2003b; Jiang et al., 2010).荒漠土壤一般表现为吸收CH4, 因为荒漠面积大, 因此其吸收总量不可小视.由于控制CH4通量的土壤过程对N的响应不同, N对CH4的影响也具有差异性.在目前全球努力控制温室气体排放的背景下, 研究温室气体的动态以及温室气体如何响应N的增加显得十分重要.由于冬季气温低, 微生物活性低, 通量通常极低(Xu et al., 2003a), 大部分研究集中于生长季.生长季是温室气体活动的旺季, 这时非生物因子适宜生物活动.本研究选择古尔班通古特荒漠, 研究人工模拟N增加条件下主要温室气体N2O、CH4和CO2的交换速率, 并分析影响其动态的主要因子, 为该区域温室气体的评估和生态系统管理提供数据支持. ...
Soil methane uptake by grasslands and forests in China. 1 2014
... 在植被越稀少的区域, 土壤年CH4吸收量越高, 一般为荒漠>灌丛地>草地>森林(Zhuang et al., 2013).荒漠土壤是陆地吸收CH4的重要组成部分, 荒漠土壤吸收CH4的测量值多数在10-38 μg·m-2·h-1之间(Striegl et al., 1992).然而, CH4吸收随生态系统和区域变化非常大(Zhuang et al., 2013; Wang et al., 2014).古尔班通古特沙漠对照样地吸收量在23-35 μg·m-2·h-1之间, 与全球大多数研究结果相近. ...
Influence of increasing temperature and nitrogen input on greenhouse gas emissions from a desert steppe soil in Inner Mongolia. 3 2011
... 古尔班通古特沙漠对照样地N2O的排放为该荒漠土壤的本底排放, 其通量(4.8 μg·m-2·h-1)与腾格里沙漠生物土壤结皮覆盖地的通量(3.2 μg·m-2·h-1)差异不大, 但后者季节变化不明显(徐冰鑫等, 2014).内蒙古荒漠草原的研究也表明, N2O的排放在较冷的冬季(4.1 μg·m-2·h-1)小于其他季节的平均值(6.3 μg·m-2·h-1), 且不受施N (10 g N·m-2·a-1)的影响(Wang et al., 2011).在青藏高原高山荒漠区, N2O的排放春季为0.7-1.1 μg·m-2·h-1, 夏季在1.2-1.9 μg·m-2·h-1之间, 秋季非常低, 春夏秋三季的排放值均远低于古尔班通古特沙漠等低海拔区域(Li et al., 2015b).在土壤极端低温、低水分和低N的北极, 土壤也有可能作为N2O的库, 在一定的光照条件下吸收部分N2O, 但是不同植被覆盖条件下具有差异(Stewart et al., 2012).因此, 荒漠N2O的排放也与其荒漠本身的立地条件密切相关. ...
... 在腾格里沙漠生物土壤结皮覆盖区, 大部分情况下土壤为CH4汇, 平均值约为0.035 μg·m-2·h-1 (徐冰鑫等, 2014), 远低于古尔班通古特沙漠.内蒙古荒漠草原的平均吸收速率为46.4 μg·m-2·h-1, 夏季具有最高的CH4吸收速率, 其吸收不受N的影响(Wang et al., 2011).在高山荒漠区域, 春季和秋季可能为CH4的释放季, 仅夏季表现为CH4的吸收, 且吸收值在4-8 μg·m-2·h-1之间(Li et al., 2015b).北极荒漠区整个生长季CH4的吸收速率可达57.1 μg·m-2·h-1 (Emmerton et al., 2014), 高于其他荒漠类型. ...
... 古尔班通古特沙漠生长季CO2排放的平均值约为46.7 mg·m-2·h-1, 春夏季排放速率约是秋季的3.5倍.腾格里沙漠生物土壤结皮覆盖地CO2的年平均值约46.7 mg·m-2·h-1, 与古尔班通古特沙漠生长季平均值一致, 且季节变化明显(徐冰鑫等, 2014).同样, 对内蒙古荒漠草原的研究表明, 其CO2排放的平均值在夏季(109 mg·m-2·h-1)远高于其他季节 (32 mg·m-2·h-1), 且不受加N的影响(Wang et al., 2011).青藏高原高山荒漠的春、夏、秋三季CO2排放量约为14、30和18 mg·m-2·h-1 (Li et al., 2015b), 春、夏季排放量远低于古尔班通古特沙漠和腾格里沙漠.这些差异可能是由研究区域温度和植被覆盖等因素造成. ...
Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem. 1 2008
Seasonal pattern of soil respiration and gradual changing effects of nitrogen addition in a soil of the Gurbantünggüt Desert, northwestern China. 1 2014
 , 陶冶
, 陶冶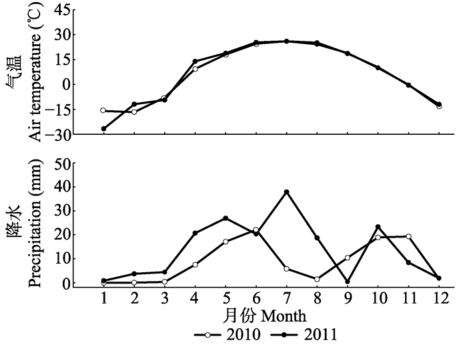 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT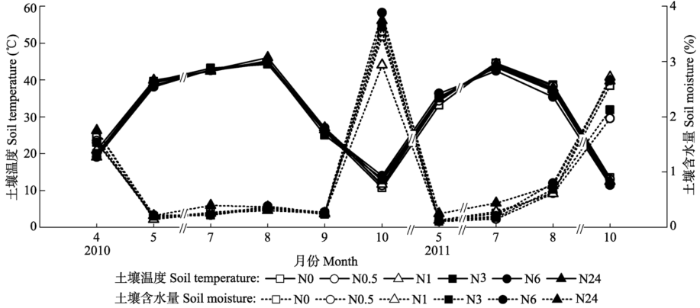 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT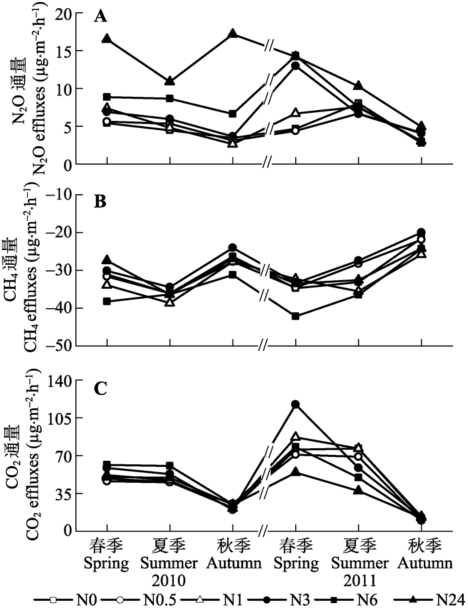 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT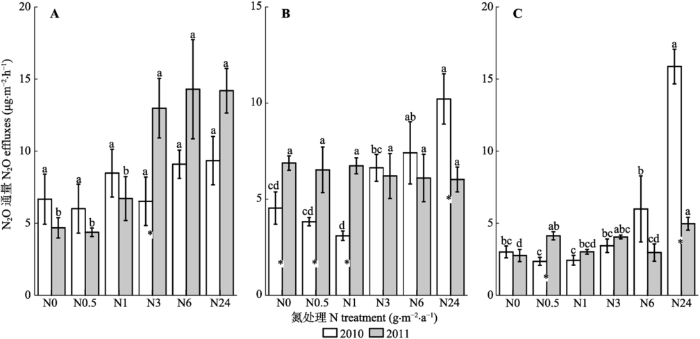 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT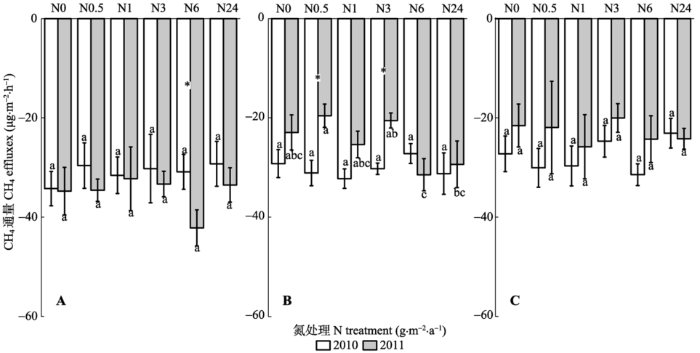 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT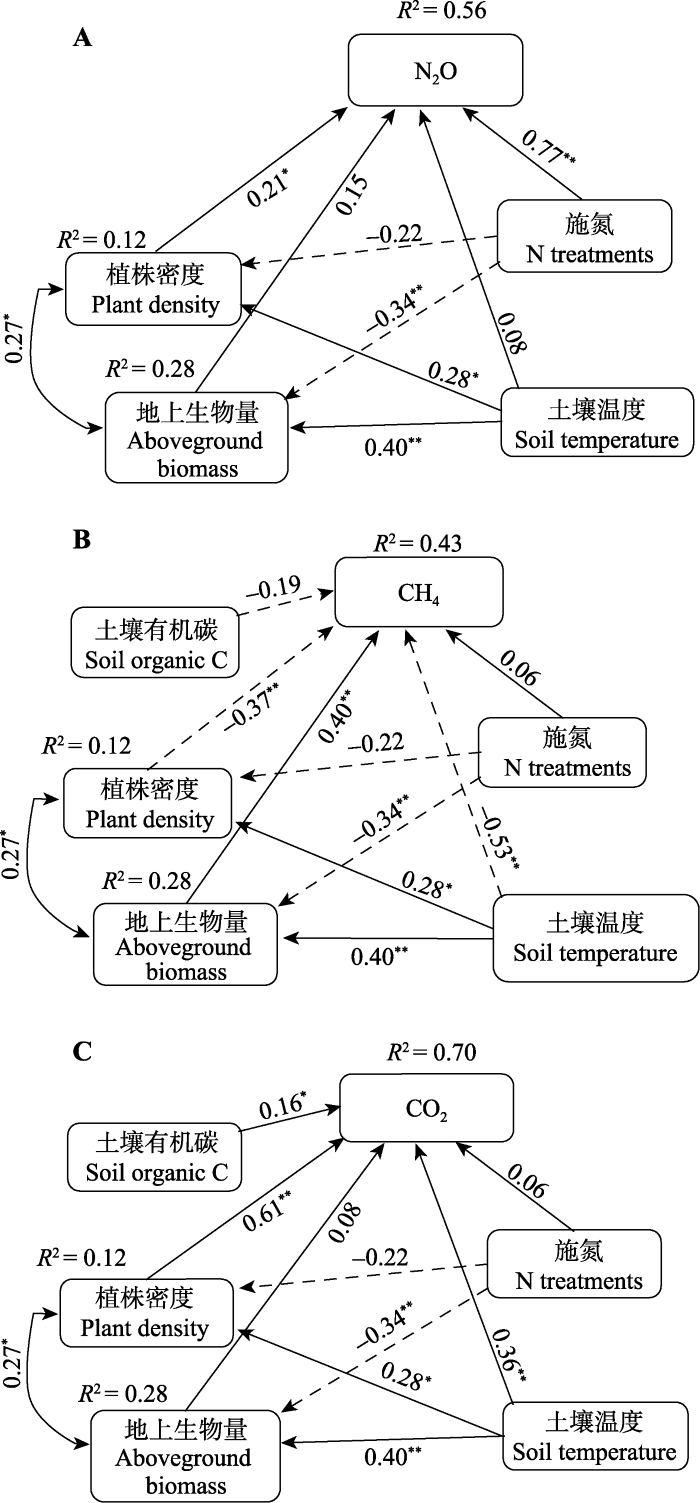 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}