Effects of light intensity variation on nitrogen and phosphorus contents, allocation and limitation in five shade-enduring plants
XIEHong-Tao, YUMu-Kui, CHENGXiang-Rong*, National Field Research Station of Eastern China Coastal Forest Ecosystem, Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Hangzhou 311400, China 通讯作者:* 通信作者Author for correspondence (E-mail: chxr@caf.ac.cn) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 浙江省与中国林业科学研究院省院合作林业科技项目(14204005和16204002)、国家自然科学基金(31300519)和中央级公益性科研院所基本科研业务费专项资金(RISF6930)
关键词:光强;耐阴植物;氮含量;磷含量;分配;适应策略 Abstract Aims To enhance the understanding on nitrogen (N) and phosphorus (P) physiological responses to different light environments in shade-enduring plants and provide references to improve the stand structure and ecosystem functions of plantation forests.Methods We selected seedlings of five shade-enduring species with high ecological and economic value in subtropical area of China to study the effects of light intensity on leaf N and P contents, allocation and nutrient limitation in shade-enduring plants. A light intensity gradient of five different levels was set to simulate the varying understory light environment.Important findings With decreasing light intensity, the total biomass and total N and P accumulation of five shade-enduring plants all showed a decreasing trend, but N, P contents in different organs increased. Among them, Gardenia jasminoides (GJ) had the highest while Illicium henryi (IH) had the lowest N content; The P contents of Quercus phillyraeoides (QP) and GJ were significantly higher than Elaeocarpus sylvestris (ES), Ardisia crenata (AC) and IH. QP and GJ had the highest N, P contents under extremely low light intensity (6% natural light intensity) condition (LIC), while AC and IH had the highest N and P contents in low (15% natural light intensity) and moderate (33% and 52% natural light intensity) LIC. ES demanded differently for LIC on N and P, which were 52% and 6% natural light intensity, respectively. N and P allocation of ES, AC and IH followed leaf > root > stem, but for QP and GJ were root > leaf > stem. Decreasing LIC significantly affected N and P allocation. N content variations shown good consistency among different organs under higher LIC (100% natural light intensity) while distinct variability under lower LIC (15% and 6% natural light intensity) in all five species. Phosphorus contents exhibited good consistency in IH, QP and GJ but varied in ES and AC. Decreasing LIC significantly affected organ N/P ratios of shade-enduring plants, but the fundamental growth restriction patterns remained. Light intensity variation and tree species co-regulated N, P utilization and allocation in shade-enduring plants, and then affected the total biomass and total N, P accumulation, which might result from the change of N and P utilization strategy. Therefore, light intensity preference and N, P nutrient balances in shade-enduring plants should be taken into account when constructing multiple layer and uneven-aged forests.
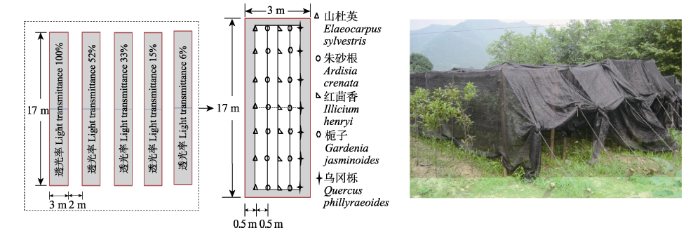
Keywords:light intensity;shade-enduring plant;N content;P content;allocation;adaptation strategy -->0 PDF (2181KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 颉洪涛, 虞木奎, 成向荣. 光照强度变化对5种耐阴植物氮磷养分含量、分配以及限制状况的影响. 植物生态学报, 2017, 41(5): 559-569 https://doi.org/10.17521/cjpe.2016.0248 XIEHong-Tao, YUMu-Kui, CHENGXiang-Rong. Effects of light intensity variation on nitrogen and phosphorus contents, allocation and limitation in five shade-enduring plants. Chinese Journal of Plant Ecology, 2017, 41(5): 559-569 https://doi.org/10.17521/cjpe.2016.0248 植物对养分的吸收、分配和利用是植物对不同环境适应策略的体现(Ingestad & Ågren, 1988; Suriyagoda et al., 2012)。氮(N)和磷(P)作为植物生长发育过程中的关键元素, 在蛋白质、核酸的合成以及能量传递等代谢过程起着至关重要的作用, 对植物的生长和发育有决定性影响(Vitousek et al., 2010)。在林下遮阴环境中, 植物对养分的吸收、分配和利用与林内小气候下的环境、土壤等多种因素有关(燕亚飞等, 2014)。作为最重要的一个影响因素, 林内光照可利用性与耐阴植物种类数量变化以及生长分布密切相关(尹华军等, 2008)。不同的光照条件(如光照强度、光质、光照时间等)不仅会影响植物光合能力和固碳相关酶的活性, 还会影响固氮相关酶(如硝酸还原酶)的活性, 进而影响植物N、P含量(王振南和杨惠敏, 2013)。在一定的光照强度范围内, 随着光照强度的增加, 植物的光合速率增加(关义新等, 2000), 而硝酸还原酶活性降低(刘国顺等, 2010), 这必然影响到依赖光合作用功能的N、P代谢过程。已有研究表明光照对耐阴植物的形态和结构特征有重要影响(管铭等, 2014), 那么光照可利用性的变化对林下耐阴植物的生理生化特征, 尤其是N、P养分在植物体各器官之间的吸收利用和分配格局会产生什么影响, 以及植物营养器官的化学计量会呈现怎样的环境分异规律和限制转变, 目前鲜有研究。此外, 就植物生态化学计量学来说, 已有大量有关非生物因素对植物N、P化学计量影响的研究(刘超等, 2012), 但对不同光环境下植物N、P生态化学计量变化特征的研究仍较少。 人工林是亚热带森林生态系统的重要组成部分, 具有多种生态和经济效益。但人工林普遍存在林分结构相对简单, 功能相对单一, 生态系统稳定性不强等突出问题。而构建完整的人工林空间结构体系有助于充分利用空间资源, 提升林分质量, 发挥更好的综合效益。构建人工林复层结构需要筛选对于特定光照环境生长和适应性良好的林下耐阴植物(许秀玉等, 2015), 而加强对耐阴植物生长和生活策略的研究有助于解决这一问题。目前我国亚热带人工林林下配置的具有较高经济价值的植物种有醉香含笑(Michelia macclurei)、木荷(Schima superba)和山杜英(Elaeocarpus sylvestris)等。本研究在对我国亚热带地区林下植被分布进行调查后, 筛选出5种分布比较广泛, 且具有较高综合价值的林下耐阴植物进行种苗培育, 然后进行光照处理实验, 以研究处于生长初期的5种耐阴植物在N、P调节和利用方面对光照强度变化的响应, 着重探讨以下3个问题: 1)不同植物N、P利用策略对光照强度变化的响应差异; 2)同种植物在不同光照强度下的N、P利用差异; 3)前二者的差异能否反映植物N、P限制格局的转变以及生物量积累的变化?
光强变化显著影响植物总生物量, 且种间差异明显。随着光强减弱, 山杜英和朱砂根的总生物量均先增加后减少, 而栀子和乌冈栎则持续减少, 红茴香总生物量先增加后减少, 但在极低光强(6R)下也表现出增加的趋势(图2)。朱砂根的生物量明显低于其他4种植物。山杜英总生物量随光照减弱骤降, 而红茴香总生物量变化幅度相对较小, 表明植物对光强变化响应的种间差异非常明显。 显示原图|下载原图ZIP|生成PPT 图2不同光强下5种耐阴植物的总N、P积累和总生物量(平均值±标准偏差)。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。不同小写字母表示同一物种不同光强下总N、P积累和总生物量差异显著(p < 0.05); 不同大写字母表示同一光强下不同物种间总N、P积累和总生物量差异显著(p < 0.05)。 -->Fig. 2Total N, P accumulations and biomass of five shade- enduring plants under varying light intensity condition (mean ± SD). AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively. Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels. -->
光照强度变化对植物总N和各器官N含量产生显著影响。在所有光照强度下, 栀子的总N含量均高于其他4种植物(图3); 而红茴香的总N含量最低且稳定性高, 随光照变化不明显, 说明它具有较强的N内稳性(图4)。随着光强的减弱, 山杜英、红茴香总N含量均表现为先增加, 后减少(p < 0.05), 且均 在光强为33R时总N含量最高(图3)。而朱砂根、乌冈栎和栀子总N含量随光强减弱波动变化, N含量最高时对应的光强分别为15R、15R和6R, 这表明5种植物在低光强环境中均有更好的N吸收能力。 显示原图|下载原图ZIP|生成PPT 图3不同光强下5种耐阴植物总N和各器官N含量(平均值±标准偏差)。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。不同小写字母表示同一物种不同光强下N含量差异显著(p < 0.05); 不同大写字母表示同一光强下不同物种间N含量差异显著(p < 0.05)。 -->Fig. 3Total N content and organ N contents of five shade-enduring plants under varying light intensity condition (mean ± SD). AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively. Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels. -->
显示原图|下载原图ZIP|生成PPT 图4不同光照强度下植物各器官N、P含量的变异系数。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。 -->Fig. 4Coefficients of variation of N and P contents in plant organs under varying light intensity condition. AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. -->
光强变化显著影响植物总P和各器官P含量(p < 0.05), 且物种差异明显。在所有光照强度下, 乌冈栎和栀子的总P含量均明显高于山杜英、朱砂根和红茴香(图5)。随着光强减弱, 朱砂根、栀子和乌冈栎的总P含量均显著增加, 山杜英先减少后增加, 呈现“U”形曲线, 红茴香则先增加后减少(p < 0.05), 表现出趋势各异的P吸收特征。山杜英、朱砂根、栀子和乌冈栎均在较低光强时有最高的P含量(15R和6R), 而红茴香则在中等光强(52R)时有最高的P含量, 这表明5种植物在低光强环境中同样具备良好的P吸收能力。 在不同光强下, 植物叶片、枝干和根系P含量有显著差异(p < 0.05)。各器官P含量随物种差异而不同, 山杜英、朱砂根和红茴香的P含量关系为: 叶片>根系>枝干; 栀子明显表现为: 根系>叶片>枝干; 乌冈栎则是根系>叶片≈枝干。随着光强减弱, 红茴香、栀子和乌冈栎P含量在器官之间的变化表现出一致性, 但山杜英和朱砂根有一定的分异性(图5)。这与N利用格局随光强的变化趋势不同, 并未明显体现光照对于器官间P含量变化的主导作用, 表明植物体内的P在器官间的分配利用可能更多地受到内部机制的调节。 显示原图|下载原图ZIP|生成PPT 图5不同光强下五种耐阴植物总P和各器官P含量(平均值±标准偏差)。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。不同小写字母表示同一物种不同光强下P含量差异显著(p < 0.05); 不同大写字母表示同一光强下不同物种间P含量差异显著(p < 0.05)。 -->Fig. 5Total P content and organ P contents of five shade-enduring plants under varying LIC (mean ± SD). AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively. Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels. -->
2.4 不同光强下植物各器官的N:P及限制状况
在不同光强下, 山杜英、朱砂根、红茴香、栀子和乌冈栎总N:P变化范围分别为9.01-16.48、11.57-13.57、8.16-11.51、9.86-12.47和4.39-5.41, 变异系数分别为23.08%、5.94%、12.04%、9.72%和9.25%, 表明光强变化强烈影响山杜英的总N:P, 而对朱砂根的影响最弱。总N:P存在器官间的差异, 山杜英、栀子和乌冈栎表现为叶片>枝干>根系, 而朱砂根为叶片>根系>枝干, 红茴香则是根系>叶片>枝干(表2)。 光强变化显著影响植物器官N:P (p < 0.05)。叶片N:P变化幅度相对较小, 而枝干和根系的N:P变化幅度较大。随着光强减弱, 山杜英各器官N:P先增加后减小, 呈现倒“U”形变化; 朱砂根枝干N:P一直减小, 根系N:P先减小后增大; 红茴香、栀子和乌冈栎各器官N:P变化趋势相似, 均为先减小后增加。各物种在器官水平上的N:P变化反映了它们在不同光环境下的生理生化自我调节, 体现了耐阴植物良好的N、P养分调节能力。 随着光强减弱, 植物叶片N:P出现波动, 但几乎未改变植物的生长受限情况(表2)。山杜英和朱砂根一直受P限制, 红茴香和乌冈栎一直受N限制, 而栀子则几乎一直受N、P共同限制。 Table 2 表2 表2不同光强下5种耐阴植物氮磷比(N:P) (平均值±标准偏差) Table 2Nitrogen/phosphorus ratio (N:P) of five shade-enduring plant species under varying light intensity condition (mean ± SD)
物种 Species
相对光强 Relative light intensity
100R
52R
33R
15R
6R
山杜英 Elaeocarpus sylvestris
总N:P Total N:P
9.01 ± 0.59Cd
15.95 ± 0.53Aa
16.48 ± 1.19Aa
14.50 ± 0.29Ab
10.87 ± 0.38ABc
叶片N:P Leaf N:P
15.90 ± 1.19Aab
18.96 ± 0.59Aab
19.11 ± 3.11Aa
17.31 ± 0.45Aab
15.67 ± 1.79Bb
枝干N:P Stem N:P
6.79 ± 0.60Dc
15.70 ± 0.02Aa
16.14 ± 0.40Aa
12.23 ± 0.37Bb
6.66 ± 0.10Cc
根系N:P Root N:P
5.17 ± 1.08Db
8.72 ± 0.59Ba
9.31 ± 0.83Aa
9.39 ± 1.34Aa
8.28 ± 1.06Ba
朱砂根 Ardisia crenata
总N:P Total N:P
13.57 ± 0.45Aa
12.62 ± 0.11Bb
12.26 ± 0.34Bbc
12.67 ± 0.41Bb
11.57 ± 0.47Ac
叶片N:P磷 Leaf N:P
16.15 ± 0.71Ab
16.59 ± 0.48Bb
17.63 ± 2.80Bab
16.61 ± 1.17Ab
19.41 ± 0.11Aa
枝干N:P Stem N:P
15.28 ± 1.18Aa
10.27 ± 0.01Bb
9.63 ± 0.25Cbc
8.75 ± 0.44Ccd
8.28 ± 0.85Bd
根系N:P Root N:P
11.61 ± 0.92Aa
10.26 ± 0.09Abc
9.64 ± 0.23Ac
9.58 ± 0.08Ac
10.58 ± 0.17Ab
红茴香 Illicium henryi
总N:P Total N:P
10.14 ± 0.73Ca
8.16 ± 0.28Da
11.11 ± 0.04Ca
11.51 ± 0.07BCa
10.37 ± 0.02Ba
叶片N:P Leaf N:P
10.73 ± 1.06Cb
9.91 ± 0.28Db
12.24 ± 0.68Da
12.09 ± 0.05Ca
11.87 ± 0.40Ca
枝干N:P Stem N:P
13.22 ± 1.00Bab
8.16 ± 0.44Cc
14.05 ± 0.34Bab
14.53 ± 1.07Aa
12.37 ± 1.39Ab
根系N:P Root N:P
7.97 ± 0.28Bb
6.33 ± 0.15Cd
8.24 ± 0.51Bb
8.92 ± 0.27Aa
7.19 ± 0.13Bc
栀子 Gardenia jasminoides
总N:P Total N:P
12.47 ± 0.24Ba
10.30 ± 0.15Cb
11.59 ± 0.91Ca
9.86 ± 0.42Cb
11.83 ± 0.29Aa
叶片N:P Leaf N:P
15.75 ± 0.51Aa
14.22 ± 0.34Cb
15.12 ± 0.91Cab
15.62 ± 1.26Bab
16.30 ± 0.48Ba
枝干N:P Stem N:P
10.24 ± 0.72Ca
6.41 ± 0.39Dc
8.15 ± 0.55Cb
5.84 ± 0.06Dcd
5.14 ± 0.09Dd
根系N:P Root N:P
7.05 ± 0.15Ca
5.29 ± 0.01Ccd
5.88 ± 0.89Cbc
4.75 ± 0.36Bd
6.48 ± 0.31Cab
乌冈栎 Quercus phillyraeoides
总N:P Total N:P
5.38 ± 0.01Da
5.41 ± 0.04Ea
4.39 ± 0.04Db
5.23 ± 0.25Da
4.54 ± 0.05Cb
叶片N:P Leaf N:P
11.64 ± 0.23Ba
9.84 ± 0.15Dbc
9.24 ± 0.85Ec
10.39 ± 0.13Db
9.92 ± 0.16Dbc
枝干N:P Stem N:P
3.65 ± 0.12Ea
3.32 ± 0.05Eb
2.98 ± 0.03Dc
3.57 ± 0.14Ea
3.59 ± 0.02Ea
根系N:P Root N:P
3.82 ± 0.12Ea
3.52 ± 0.02Da
2.79 ± 0.37Db
3.54 ± 0.35Ca
2.65 ± 0.02Db
Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively.同行不同小写字母表示同一植物各器官在不同光照水平下N:P差异显著(p < 0.05); 同列同项不同大写字母表示不同植物同一器官在相同光照条件下N:P差异显著(p < 0.05)。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。 新窗口打开
光强变化对耐阴植物N、P养分含量、利用格局以及限制状况均有显著影响, 且种间差异明显。总体上, 光强的减弱降低了植物总生物量和总N、P积累, 但植物器官水平上的N、P含量均有所增加。光强变化会影响植物受限状况, 但基本未改变其受限状态。在异质性光强中的N、P养分吸收和分配特征反映了耐阴植物在不良光环境中的适应性策略和在理想光环境中的生长性策略。植物N含量变化受光强影响程度大于P, P可能更多地受到内部调节。综合来看, 可优先考虑将山杜英和红茴香作为亚热带地区人工林林下植被恢复的先锋乔灌物种。本研究的局限性在于缺少野外实验验证, 未来还需要在多尺度上研究光强对于耐阴植物氮、磷养分积累、营养器官含量以及限制状况的影响。 致谢 感谢千家村试验基地工作人员在样品采集、分析工作中给予的帮助。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
ChengXR, ZhouJH, ChenYH, WuKZ, YuMK (2016). Effects of light intensity on phenotypic plasticity of Gardenia jasminoides seedlings. Acta Agriculturae Universitatis Jiangxiensis, 38, 180-186. (in Chinese with English abstract)[成向荣, 周俊宏, 陈永辉, 武克壮, 虞木奎 (2016). 山栀子幼苗表型可塑性对不同光环境的响应 . 江西农业大学学报, 38, 180-186.] [本文引用: 2]
[3]
DonovanLA, MaheraliH, CarusoCM, HuberH, KroonHD (2011). The evolution of the worldwide leaf economics spectrum. Trends in Ecology & Evolution, 26, 88-95. [本文引用: 1]
[4]
GrootDC, BoogaardR, MarcelisLM, HarbinsonJ, LambersH (2003). Contrasting effects of N and P deprivation on the regulation of photosynthesis in tomato plants in relation to feedback limitation. Journal of Experimental Botany, 54, 1957-1967. [本文引用: 1]
[5]
GuanM, JinZX, WangQ, LiYL, ZuoW (2012). Response of photosynthesis traits of dominant plant species to different light regimes in the secondary forest in the area of Qiandao Lake, Zhejiang, China. Chinese Journal of Applied Ecology, 25, 1615-1622. (in Chinese with English abstract)[管铭, 金则新, 王强, 李月灵, 左威 (2012). 千岛湖次生林优势种植物光合特性对不同光环境的响应 . 应用生态学报, 25, 1615-1622.] [本文引用: 1]
[6]
GuanYX, LinB, LingBY (2000). Interaction effects of light density and nitrogen on maize leaf pigments, chlorophyll fluorescence and energy transition. Plant Nutrition and Fertilizer Science, 6, 152-158. (in Chinese with English abstract)[关义新, 林葆, 凌碧莹 (2000). 光氮互作对玉米叶片光合色素及其荧光特性与能量转换的影响 . 植物营养与肥料学报, 6, 152-158.] [本文引用: 1]
[7]
GüsewellS (2004). N:P ratios in terrestrial plants: Variation and functional significance. New Phytologist, 164, 243-266. [本文引用: 1]
[8]
IngestadT, ÅgrenGI (1988). Nutrient uptake and allocation at steady-state nutrition. Physiologia Plantarum, 72, 450-459. [本文引用: 1]
[9]
KangM, XieYM, XuY, XuYL, YanER (2013). Within- community variability of plant leaf N and P contents in Tiantong, Zhejiang Province. Journal of East China Normal University (Natural Science), (2), 20-29. (in Chinese with English abstract)[康蒙, 谢一鸣, 许月, 徐艺露, 阎恩荣 (2013). 浙江天童植物叶片氮磷含量的群落内变异 . 华东师范大学学报(自然科学版), (2), 20-29.] [本文引用: 1]
[10]
KoerselmanW, MeulemanAFM (1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 33, 1441-1450.
[11]
LiDF, YuSL, WangGX, FangWW (2015). Environmental heterogeneity and mechanism of stoichiometry properties of vegetative organs in dominant shrub communities across the Loess Plateau. Chinese Journal of Plant Ecology, 39, 453-465. (in Chinese with English abstract)[李单凤, 于顺利, 王国勋, 方伟伟 (2015). 黄土高原优势灌丛营养器官化学计量特征的环境分异和机制 . 植物生态学报, 39, 453-465.] [本文引用: 1]
[12]
LiYL, MaoW, ZhaoXY, ZhangTH (2010). Leaf nitrogen and phosphorus stoichiometry in typical desert and desertified regions, North China. Environmental Science, 31, 1716-1725. (in Chinese with English abstract)[李玉霖, 毛伟, 赵学勇, 张铜会 (2010). 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究 . 环境科学, 31, 1716-1725.] [本文引用: 1]
[13]
LiZA, WangBS, KongGH, ZhangZP, WengH (1999). The element content of plant in Cryptocarya concinna community in Dinghushan evergreen monsoon broad-leaf forest. Acta Phytoecologica Sinica, 23, 411-417. (in Chinese with English abstract)[李志安, 王伯荪, 孔国辉, 张祝平, 翁轰 (1999). 鼎湖山季风常绿阔叶林黄果厚壳桂群落植物元素含量特征 . 植物生态学报, 23, 411-417.] [本文引用: 1]
[14]
LiuC, WangY, WangN, WangGX (2012). Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial ecosystems: A review. Chinese Journal of Plant Ecology, 36, 1205-1216. (in Chinese with English abstract)[刘超, 王洋, 王楠, 王根轩 (2012). 陆地生态系统植被氮磷化学计量研究进展 . 植物生态学报, 36, 1205-1216.] [本文引用: 1]
[15]
LiuGS, YunF, ShiHZ, WangK, ZhangCH, SongJ (2010). Effects of cooperation of light and nitrogen on the content of nitric compound, antioxidant system and quality of flue- cured tobacco. Scientia Agricultura Sinica, 43, 3732-3741. (in Chinese with English abstract)[刘国顺, 云菲, 史宏志, 王可, 张春华, 宋晶 (2010). 光、氮及其互作对烤烟含氮化合物含量、抗氧化系统及品质的影响 . 中国农业科学, 43, 3732-3741.] [本文引用: 1]
[16]
LiuWF, FanHB (2008). Characteristics of nitrogen cycles in the ecosystem of mixed stands of Masson pine and broadleaved forests. Journal of Beijing Forestry University, 30(6), 52-58. (in Chinese with English abstract)[刘文飞, 樊后保 (2008). 马尾松阔叶树混交林生态系统的氮素循环特征 . 北京林业大学学报, 30(6), 52-58.] [本文引用: 1]
[17]
LiuXZ, KangSZ (2002). Effects of shading on photosynthesis, dry matter partitioning and N, P, K concentrations in leaves of tomato plants at different growth stages. Acta Ecologica Sinica, 22, 2264-2271. (in Chinese with English abstract)[刘贤赵, 康绍忠 (2002). 不同生长阶段遮荫对番茄光合作用、干物质分配与叶N、P、K的影响 . 生态学报, 22, 2264-2271.]
[18]
MatzekV, VitousekPM (2009). N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis. Ecology Letters, 12, 765-771. [本文引用: 1]
[19]
QiaoXR (2007). Effects of Light Intensity on Growth Photosynthetic Characteristics and Quality of Flue-cured Tobacco. Master degree dissertation, Henan Agricultural University, Zhengzhou. (in Chinese with English abstract)[乔新荣 (2007). 光照强度对烤烟生长发育、光合特性及品质的影响 . 硕士学位论文, 河南农业大学, 郑州.]
[20]
ShiJG, ZhuKL, CaoHY, DongST, LiuP, ZhaoB, ZhangJW (2015). Effect of light from flowering to maturity stage on dry matter accumulation and nutrient absorption of summer maize. Chinese Journal of Applied Ecology, 26, 46-52.(in Chinese with English abstract). [史建国, 朱昆仑, 曹慧英, 董树亭, 刘鹏, 赵斌, 张吉旺 (2015). 花粒期光照对夏玉米干物质积累和养分吸收的影响. 应用生态学报, 26, 46-52.] [本文引用: 1]
[21]
ShiJZ, WangTD (1994). A mechanistic model describing the photosynthate partitioning during vegetative phase. Acta Botanica Sinica, 36, 181-189. (in Chinese with English abstract)[施建忠, 王天铎 (1994). 植物营养生长期同化物分配的机理模型 . 植物学报, 36, 181-189.]
[22]
SimsL, PastorJ, LeeT, DeweyB (2012). Nitrogen, phosphorus and light effects on growth and allocation of biomass and nutrients in wild rice. Oecologia, 170, 65-76. [本文引用: 1]
[23]
SuriyagodaLB, LambersH, RentonM, RentonM, RyanMH (2012). Growth, carboxylate exudates and nutrient dynamics in three herbaceous perennial plant species under low, moderate and high phosphorus supply. Plant and Soil, 358, 105-117. [本文引用: 1]
[24]
ThompsonK, ParkinsonJA, BandSR, SpencerRE (1997). A comparative study of leaf nutrient concentrations in a regional herbaceous flora. New Phytologist, 136, 679-689. [本文引用: 1]
[25]
ValladaresF, MartinezFE, BalaguerL, CoronaEP, ManriqueE (2000). Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource- use strategy? New Phytologist, 148, 79-91. [本文引用: 1]
WangH, CaiZQ, CaiCT, ZhangSX (2008). A comparative study of two tropical pioneer species with different life span under different light and nutrient conditions. Journal of Wuhan Botanical Research, 26, 134-141. (in Chinese with English abstract)[王辉, 蔡志全, 蔡传涛, 张硕新 (2008). 不同光照和营养条件下两种不同寿命热带先锋种的对比研究 . 武汉植物学研究, 26, 134-141.] [本文引用: 1]
[28]
WangSQ, YuGR (2008). Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus element. Acta Ecologica Sinica, 28, 3937-3947. (in Chinese with English abstract)[王绍强, 于贵瑞 (2008). 生态系统碳氮磷元素的生态化学计量学特征 . 生态学报, 28, 3937-3947.] [本文引用: 1]
[29]
WangZN, YangHM (2013). Response of ecological stoichiometry of carbon, nitrogen and phosphorus in plants to abiotic environmental factors. Pratacultural Science, 30, 927-934. (in Chinese with English abstract)[王振南, 杨惠敏 (2013). 植物碳氮磷生态化学计量对非生物因子的响应 . 草业科学, 30, 927-934.] [本文引用: 1]
[30]
XuQX (2013). Effect of Thinning on Soil Physicochemical Property and Parbon Ptorage of the Patural Larix gmelinii Forest in Great Xing’an Mountains. Master degree dissertation, Northeast Forestry University, Harbin. (in Chinese with English abstract)[徐庆祥 (2013). 抚育间伐对大兴安岭兴安落叶松天然林碳储量的影响 . 硕士学位论文, 东北林业大学, 哈尔滨.] [本文引用: 1]
[31]
XuXY, XiaoL, WangMH, ZhangHX (2015). A comprehensive evaluation system for anti-typhoon performance of trees in coastal areas. Journal of Zhejiang A & F University, 32, 516-522. (in Chinese with English abstract)[许秀玉, 肖莉, 王明怀, 张华新 (2015). 沿海抗台风树种评价体系构建与选择 . 浙江农林大学学报, 32, 516-522.] [本文引用: 1]
[32]
YanYF, FangSZ, TianY, SongH, TunXJ (2014). The response of understory plant diversity and nutrient accumulation to stand structure of poplar plantation. Chinese Journal of Ecology, 33, 1170-1177. (in Chinese with English abstract)[燕亚飞, 方升佐, 田野, 宋浩, 囤兴建 (2014). 林下植物多样性及养分积累量对杨树林分结构的响应 . 生态学杂志, 33, 1170-1177.] [本文引用: 1]
[33]
YangX, TangZY, JiC, LiuHY, MaWH, MohhamotA, ShiZY, SunW, WangT, WangXP, WuX, YuSL, YueM, ZhengCY (2014). Scaling of nitrogen and phosphorus across plant organs in shrub land biomes across Northern China. Scientific Reports, 4, 5448. doi: 10.1038/srep05448.
[34]
YinHJ, LaiT, ChengXY, JiangXM, LiuQ (2008). Warming effects on growth and physiology of seedlings of Betula albo-sinensis and Abies faxoniana under two contrasting light conditions in subalpine coniferous forest of western Sichuan, China. Journal of Plant Ecology (Chinese Version), 32, 1072-1083. (in Chinese with English abstract)[尹华军, 赖挺, 程新颖, 蒋先敏, 刘庆 (2008). 增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响 . 植物生态学报, 32, 1072-1083.] [本文引用: 1]
[35]
ZhaoP, KriebitzsehW, ZhangZQ (1999). Gas exchange, chlorophyll and nitrogen contents in leaves of three common trees in middle Europe under two contrasting light regime. Journal of Tropical and Subtropical Botany, 7, 133-139. (in Chinese with English abstract)[赵平, KriebitzsehW, 张志权 (1999). 欧洲3种常见乔木幼苗在两种光环境下叶片的气体交换、叶绿素含量和氮素含量 . 热带亚热带植物学报, 7, 133-139.] [本文引用: 1]
[36]
ZhengSX, ShangguanZP (2007). Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees, 21, 357-370. [本文引用: 1]

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT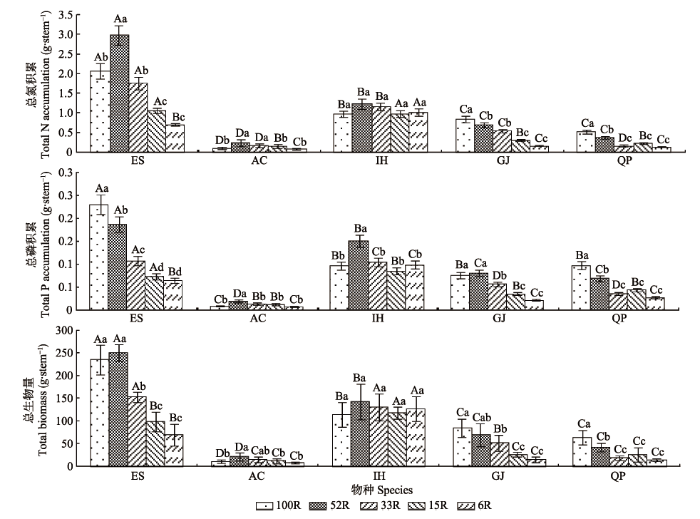 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT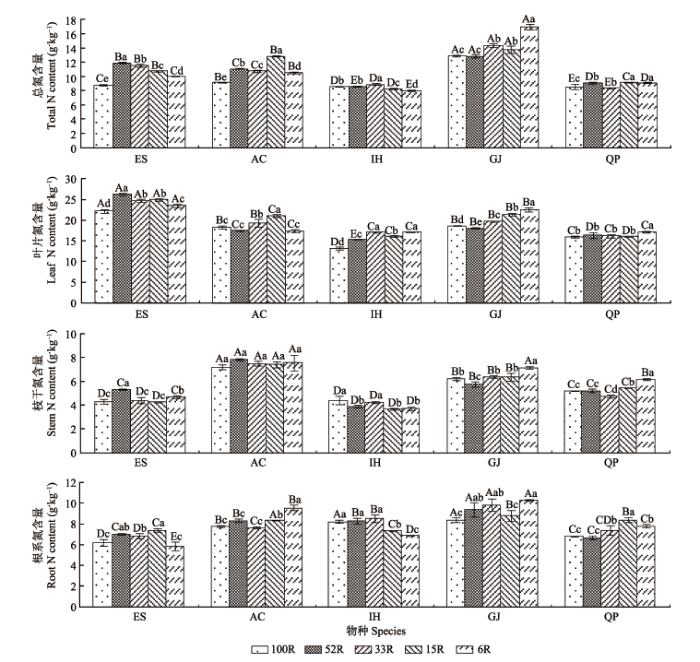 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT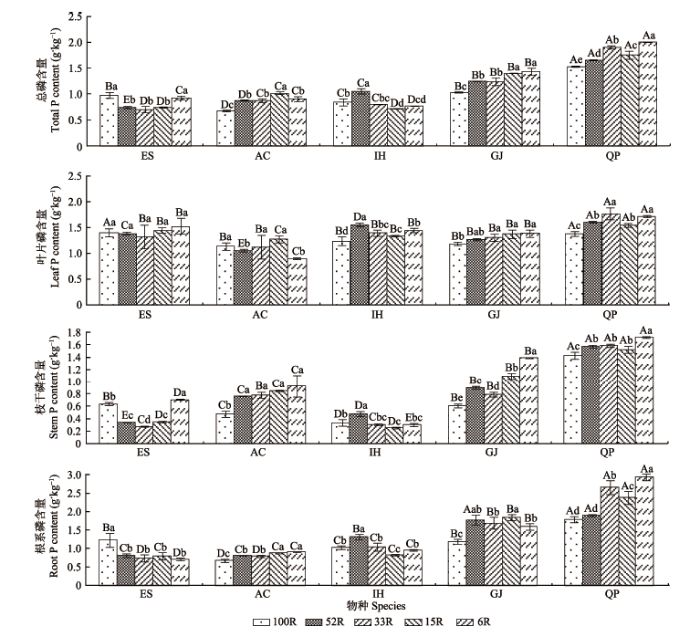 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}