Growth-form regulates the altitudinal variation of interspecific seed mass of woody plants in Mt. Dalaoling, the Three Gorges Region, China
LIDao-Xin1, LIGuo2, SHENZe-Hao3,*,, XUShen-Dong1, HANQing-Yu1, WANGGong-Fang1, TIANFeng-Lei1 1Administration of the Dalaoling National Reserve in the Three Gorges, Yichang, Hubei 443000, China2Research Center of Biodiversity, Chinese Research Academy of Environmental Sciences, Beijing 100012, China3College of Urban and Environmental Sciences, Key Laboratory Ministry of Education for Earth Surface Processes, Peking University, Beijing 100871, China 通讯作者:* 通信作者Author for correspondence (E-mail: shzh@urban.pku.edu.cn) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 国家自然科学基金(31170449和41371190)
关键词:木本植物;种子质量;海拔;生长型;三峡大老岭 Abstract Aims Seed size is one of the most important characteristics of plant seeds, and has significant implications in plant ecological functions. Exploring the altitudinal pattern of seed size would help to detect environmental constraints on species distribution and understand the linkage between plant ecological function traits Methods The present study measured the quantitative features of seed size, including weight of 1 000 grain seeds, lengths of longer and shorter axes, and analyzed the relationships between seed size and altitude, as well as the influence of growth form. Seed samples were all collected from 201 locally common woody plants (belonging to 59 families and 87 genera) in the Dalaoling Natural Reserve in Yichang City, Hubei Province.Important findings Measured values of the seed mass, the longer axis, and the shorter axis of the 201 woody plant species all follow the lognormal distribution. Measurements of seed mass vary across five orders of magnitude. Significant correlation was found between seed mass, seed length of the longer and shorter axes (R2 = 0.755; 0.819; 0.630, p < 0.01). Moreover, seed mass of trees and small trees are significantly heavier than those of shrubs and woody vines. Seed mass values of evergreen broad leaved species are significantly heavier than those of deciduous broad leaved species and needle leaved species. Seed mass of all 201 species shows a slightly but statistically significant decreasing trend with the increase of altitude. In addition, altitudinal patterns of seed mass varied between species with different growth form. Our results indicated the variation of altitudinal trends of seed mass for different structural components of plant communities, implying the local community structure as a critical aspect of variation in macro-ecological patterns.
为了反映研究区域种子特征沿海拔梯度的分布, 根据对大老岭地区维管植物海拔分布的前期研究(沈泽昊等, 2001), 得到本研究中201种植物在本地区的海拔分布范围。在后续分析中, 以每种植物在大老岭的海拔分布范围中点作为其分布海拔高度值。根据《湖北植物志》(傅书遐, 2001-2002)查得不同木本植物的生长型, 分为乔木、小乔木、木质藤本和灌木4类, 并按叶性状分为常绿阔叶、落叶阔叶和常绿针叶3类。 频率直方图显示种子质量、种子长度和厚度3个指标的数值分布均强烈左偏(图1A、1B、1C), 表明数据可能服从泊松分布或对数正态分布。对每个数据进行对数变换, 并利用Kolmogorov-Smirnov检验比较种子3个指标的经验累积频率分布(ECFD)与参数值(即平均值和标准偏差)相同的正态分布累积密度函数(Norm CDF)的一致性(图1D、1E、1F)。结果表明, 对于样本量n = 201的种子质量、长度和厚度3个指标, Kolmogorov-Smirnov检验值D及其统计显著性检验p值分别为, 种子质量: D = 0.114, 0.05 < p < 0.10; 种子长度: D = 0.139, p < 0.05; 种子厚度: D = 0.121, p < 0.05, 均服从对数正态分布。故在后续分析中3个种子指标均采用1og10转换值。 因此, 采用种子质量和大小测量值的对数变化值进行后续分析, 包括Pearson相关分析、多元线性回归模型和方差分析。多元线性回归模型只以海拔、叶片性状、木本生长型为自变量, 并考虑变量两两之间的交互作用项, 用调整R2和F检验值作为模型比较的参数。 各项分析均用统计软件R3.2.5(www.r-project.org)完成。 显示原图|下载原图ZIP|生成PPT 图1种子质量(A)、长度(B)和厚度(C)值的频率分布直方图及3个指标的对数转换值的经验累积频率分布(ECFD)与正态分布的累积密度函数(Normal CDF)拟合曲线的比较(D, E, F)。 -->Fig. 1The frequency distribution of seed mass (A), seed length (B) and thickness (C), and a comparison between the curves of empirical cumulative frequency distribution (ECFD) and cumulative density function (CDF) of normal distribution, for log10-transformed seed mass, seed length and seed thickness (D, E, F). -->
2 结果
2.1 种子的基本统计特征
本文测定的木本植物种子数据总计201种, 按生长型, 包括: 乔木85种, 小乔木42种, 灌木61种, 藤本植物13种。所测定的种子质量介于0.03-7 870.00 mg (图2), 平均值为178.4 mg; 最大值与最小值之间相差5个数量级。其中, 种子质量介于1.0- 500.0 mg的植物种数占到全部统计物种的64.4%。 显示原图|下载原图ZIP|生成PPT 图2对数转换的种子质量与种子长度、厚度(A)及种子长度与厚度(B)的相关性。 -->Fig. 2The correlation between seed mass (mg, log10-transformed) and the length and thickness (mm, both log10-transformed) (A), and the correlation between the length and thickness (B) of seeds. -->
全部测量种子的长度介于0.82-34.90 mm, 平均值和中位值分别为6.81 mm和5.46 mm, 种子长度的最大值与最小值之间相差2个数量级; 种子厚度介于0.12-27.97 mm; 平均值和中位值分别为4.08 mm和3.05 mm。种子厚度的最大值与最小值之间也相差2个数量级。201种木本植物种子的长度与厚度比 介于1.005-22.020, 其平均值和中位值分别为2.410和1.653。种子长度与厚度比的最大值与最小值之间相差1个数量级。 大老岭主要木本植物种子质量与种子长度、厚度(对数值)之间均存在极显著(p < 0.001)的线性相关关系, 相关系数分别为0.744和0.873 (图3A), 因此, 种子厚度比长度对种子质量的指示性更好。而种子长度与厚度之间也有显著的线性关系(r = 0.694; p < 0.001)(图3B)。
2.2 种子质量与生长型之间的关系
根据F检验结果, 乔木、小乔木、灌木、木质藤本4种生长型植物的种子质量对数值之间存在如下 关系: 乔木和小乔木的种子质量显著大于灌木和木质藤本(p < 0.05), 但乔木和小乔木之间、灌木和木质藤本之间的种子质量差异并不显著(图3A)。按照叶性状把这些物种分成常绿(阔叶)、落叶(阔叶)和针叶三类, 结果常绿阔叶种的种子质量显著大于落叶阔叶种, 但在针叶种与常绿阔叶种、落叶阔叶种之间, 种子质量的差异都不显著(图3B)。 显示原图|下载原图ZIP|生成PPT 图3不同生长型(A)和叶片类型(B)的种子质量对数值。a、b代表显著性阈值p < 0.05下的不同水平值。 -->Fig. 3Seed mass (log10-transformed) of different growth forms (A) and leaf types (B). a, and b indicate significantly different levels with p < 0.05. -->
2.3 种子质量的海拔格局
全部201种木本植物的种子质量的对数值与海拔有统计上显著但较弱的负相关关系(图4A)。分为常绿阔叶种、落叶阔叶种和针叶种(图4B、4C、4D)来看, 常绿阔叶种的种子质量对数值与海拔有统计上显著(n = 47, p = 0.018)的负相关关系, 落叶阔叶种与海拔没有显著的相关性(n = 146), 针叶种种子质量与海拔之间却显著正相关(n = 8, p = 0.019)。但从不同生长型来看, 乔木、灌木和木质藤本种(图4E、4G、4H)种子质量(对数值)与海拔都没有显著的相关关系, 而小乔木(图4F)种子质量对数值与海拔有显著的负相关关系。 显示原图|下载原图ZIP|生成PPT 图4不同生长型植物种组的种子质量对数值的海拔格局。A, 全部种。B, 常绿阔叶种。C,落叶阔叶种。D, 针叶种。E, 乔木。F, 小乔木。G, 灌木。H, 木质藤本。 -->Fig. 4Altitudinal patterns of seed mass (log10-transformed) of different growth-forms of plant species. A, All species. B, Evergreen-broadleaved species. C, Deciduous-broadleaved species. D, Coniferous species. E, Tree. F, Small tree. G, Shrub. H, Liana. -->
2.4 生长型和海拔对植物种子质量的影响
综合考虑不同物种的生长型、叶性状和分布的海拔高度三方面因素, 比较几个不同模型的结果可以发现, 3个因素合计可以解释20%左右的植物种子质量变化, 其中反映物种分布海拔与落叶性的交互作用的模型(M4)最理想(表1)。结合多元线性回归模型和方差分析, 表明生长型对种子质量的影响最大, 占总解释量的67.7%; 其次是物种分布的海拔中点値(代表其生态适宜海拔高度)占总解释量的9.3%; 叶属性效应接近显著, 占解释量的7.9%; 而落叶性与物种适宜海拔的交互作用对植物种子质量的影响显著, 占解释总量的14.9%。但是, 在考虑了交互效应之后, 模型M4中海拔本身的回归系数并不显著。 Table 1 表1 表1解释种子质量对数值的不同多元线性回归模型比较 Table 1Comparison of linear regression models for seed mass (log10-transformed)
模型 Model
自变量 Independent variables
自由度 d.f.
R2
F
p
M1
Elevation, Leaf, GF
194
0.186
8.543
<0.001
M2
Elevation, Leaf, GF, Leaf : GF
191
0.192
6.218
<0.001
M3
Elevation, Leaf, GF, Elevation : GF
191
0.198
6.414
<0.001
M4
Elevation, Leaf, GF, Elevation : Leaf
192
0.216
7.815
<0.001
Elevation, elevation of mid-point of altitudinal species range; GF, growth form, including tree, small tree, shrub and aliana; Leaf, leaf features, including evergreen broad-leaved, deciduous broad-leaved, and coniferous; d.f., degree of freedom.Elevation, 物种海拔分布范围的中点; GF, 生长型, 包括乔木、小乔木、灌木和木质藤本; Leaf, 叶性状, 包括常绿阔叶、落叶阔叶和针叶。 新窗口打开 常绿阔叶种的种子质量大于落叶阔叶种且都显著大于针叶树的种子质量。乔木和小乔木的种子质量显著大于藤本和灌木种的种子质量, 后两者之间差异不显著。常绿阔叶种和落叶阔叶种的种子质量与海拔的相关系数相对于针叶种而言都显著较小(表2)。 Table 2 表2 表2最优回归模型(M4)的参数估计 Table 2Parameter estimate for the optimal regression model (M4)
对三峡宜昌大老岭地区常见木本植物种子大小的海拔格局的初步研究表明, 所分析物种的种子质量和大小值均服从对数正态分布, 数值变异性的顺序是质量值>种子长度>种子厚度; 种子质量、长度厚度之间均存在极显著的相关关系; 乔木和小乔木种子质量显著大于灌木和藤本; 常绿阔叶种的种子质量大于落叶阔叶和针叶树种。大老岭地区主要木本植物种子的质量与海拔有一个弱而统计上显著的负相关关系, 但这种关系在不同生长型和叶性状之间并不一致。种子质量的海拔格局可能受到群落和环境多因素的影响, 需要一个多尺度的解释框架。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
BakerHG (1972). Seed weight in relation to environmental conditions in California. Ecology, 53, 997-1010. [本文引用: 1]
[3]
BolmgrenK, CowanPD (2008). Time-size tradeoffs: A phylogenetic comparative study of flowering time, plant height and seed mass in a north-temperate flora. Oikos, 117, 424-429. [本文引用: 2]
[4]
BuHY, CuXL, XuXL, LiuK, JiaP, DuGZ (2006). Seed mass and germination in an alpine meadow on the eastern Tsinghai-Tibet Plateau. Plant Ecology, 191, 127-149. [本文引用: 1]
[5]
Castro-DíeP, Montserrat-MartíG, CornelissenJHC (2003). Trade-offs between phenology, relative growth rate, life form and seed mass among 22 Mediterranean woody species. Plant Ecology, 166, 117-129. [本文引用: 1]
[6]
DaineseM, SitziaT (2013). Assessing the influence of environmental gradients on seed mass variation in mountain grasslands using a spatial phylogenetic filtering approach. Perspectives in Plant Ecology, Evolution and Systematics, 15, 12-19. [本文引用: 1]
[7]
DíazS, KattgeJ, CornelissenJHC, WrightIJ, LavorelS, DrayS, ReuB, KleyerM, WirthC, PrenticeIC, GarnierE, BönischG, WestobyM, PoorterH, ReichPB, MolesAT, DickieJ, GillisonAN, ZanneAE, ChaveJ, WrightSJ, Sheremet’evSN, JactelH, BaralotoC, CeraboliniB, PierceS, ShipleyB, KirkupD, CasanovesF, JoswigJS, GüntherA, FalczukV, RügerN, MahechaMD, GornéLD (2016). The global spectrum of plant form and function. Nature, 529, 167-171. [本文引用: 1]
[8]
DuY, HeHJ, ZhangZF, YangYJ, LiLY, YangXY (2014). Correlation of seed mass with elevation. Plant Diversity and Resources, 36, 109-115. (in Chinese with English abstract)[杜燕, 何华杰, 张志峰, 杨娅娟, 李涟漪, 杨湘云(2014). 种子重量与海拔的相关性分析 . 植物分类与资源学报, 36, 109-115.] [本文引用: 2]
[9]
FieldingCA, WhittakerJB, ButterfieldJEL, CoulsonJC (1999). Predicting responses to climate change: The effect of altitude and latitude on the phenology of the Spittlebug Neophilaenus lineatus. Functional Ecology, 13(Suppl. 1), 65-73. [本文引用: 1]
[10]
FuSX (2001-2002). Flora of Hubei. Vol. I, II, III. Hubei Press of Science and Technology., Wuhan (in Chinese)[傅书遐 (2001-2002). 湖北植物志. 第I, II, III卷. 湖北科学技术出版社.], 武汉
[11]
GarlandT, HarveyPH, IvesAR (1992). Procedures for the analysis of comparative data using phylogenetically independent contrasts. Systematic Biology, 41, 18-32. [本文引用: 1]
[12]
GentiliR, ArmiraglioS, SgorbatiS, BaroniC (2013). Geomorphological disturbance affects ecological driving forces and plant turnover along an altitudinal stress gradient on alpine slopes. Plant Ecology, 214, 571-586. [本文引用: 1]
[13]
GuoH, MazerSJ, DuG (2010). Geographic variation in seed mass within and among nine species of Pedicularis(Orobanchaceae): Effects of elevation, plant size and seed number per fruit. Journal of Ecology, 98, 1232-1242. [本文引用: 2]
[14]
GuoSQ (2007). Relationship Between Seed Size and Climate Along Elevational Gradient on the Eastern Qinghai-Tibetan Plateau. Master degree dissertation, Lanzhou University, Lanzhou. (in Chinese with English abstract)[郭淑青 (2007). 青藏高原东部植物种子大小与海拔及气候关系研究 . 硕士学位论文, 兰州大学, 兰州.] [本文引用: 1]
[15]
HammondDS, BrownVK (1995). Seed size of woody plants in relation to disturbance, dispersal, soil type in wet neotropical forests. Ecology, 76, 2544-2561. [本文引用: 1]
[16]
HarperJL, LovellPH, MooreKG (1970). The shapes and sizes of seeds. Annual Review of Ecology and Systematics, 1, 327-356. [本文引用: 2]
[17]
HoweHF, SmallwoodJ (1982). Ecology of seed dispersal. Annual Review of Ecology and Systematics, 13, 201-228. [本文引用: 2]
[18]
HuangZL (2002). Biodiversity conservation for the Three Gorges Project. Biodiversity Science, 9, 472-481. (in Chinese with English abstract)[黄真理 (2002). 三峡工程中的生物多样性保护 . 生物多样性, 9, 472-481.] [本文引用: 1]
[19]
JiangZ, MaK (2015). Environmental filtering drives herb community composition and functional trait changes across an elevational gradient. Plant Ecology and Evolution, 148, 301-310. [本文引用: 1]
[20]
JinYX, ZhengZ, ChenZL, WangYM, PengPS, ZhaoZE (1987). Studies on the strategies for the impacts of the Three Gorges Project on the valuable and rare plant species in the reservoir region. In: The Leading Group of the Chinese Academy of Sciences of the Project for the Ecology and Environment in the Three Gorges Project eds. Studies on the Strategies for the Impacts of the Three Gorges Project of the Yangtze River on the Ecology and Environment. Science Press, Beijing. 114-122. (in Chinese)[金义兴, 郑重, 陈卓良, 王映明, 彭蒲松, 赵子恩 (1987). 三峡工程对库区珍稀植物的影响及对策的研究. 见: 中国科学院三峡工程生态与环境项目领导小组编, 长江三峡工程对生态与环境影响及其对策研究. 科学出版社, 北京. 114-122.] [本文引用: 1]
[21]
KitayamaK, AibaS (2002). Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. Journal of Ecology, 90, 37-51. [本文引用: 1]
[22]
KolbA, BarschF, DiekmannM (2006). Determinants of local abundance and range size in forest vascular plants. Global Ecology and Biogeography, 15, 237-247. [本文引用: 1]
[23]
Ko?rnerC (2000). Why are there global gradients in species richness? Mountains might hold the answer. Trends in Ecology and Evolution, 15, 513-514. [本文引用: 1]
[24]
Ko?rnerC (2007). The use of “altitude” in ecological research. Trends in Ecology and Evolution, 22, 569-574. [本文引用: 1]
[25]
LeishmanMR (2001). Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. Oikos, 93, 294-302. [本文引用: 1]
[26]
LeishmanMR, MurrayBR (2001). The relationship between seed size and abundance in plant communities: Model predictions and observed patterns. Oikos, 94, 151-161. [本文引用: 1]
[27]
LuoL, ShenGZ, XieZQ, YuJ (2011). Leaf functional traits of four typical forests along the altitudinal gradient in Mt. Shennongjia. Acta Ecologica Sinica, 31, 6420-6428. (in Chinese with English abstract)[罗璐, 申国珍, 谢宗强, 喻杰 (2011). 神农架海拔梯度上4种典型森林的乔木叶片功能性状特征 . 生态学报, 31, 6420-6428.] [本文引用: 1]
[28]
MolesAT, WartonDI, StevensRD, WestobyM (2004). Does a latitudinal gradient in seedling survival favor larger seeds in the tropics? Ecology Letter, 7, 911-914.
[29]
MolesAT, WestobyM (2003). Latitude, seed predation and seed mass. Journal of Biogeography, 30, 105-128. [本文引用: 1]
[30]
MolesAT, WestobyM (2006). Seed size and plant strategy across the whole life cycle. Oikos, 113, 91-105. [本文引用: 2]
[31]
Montaño-CentellasFA, Garitano-ZavalaA (2015). Andean bird responses to human disturbances along an elevational gradient. Acta Oecologica, 65-66, 51-60. [本文引用: 1]
[32]
PieterDF, GraaeBJ, AnnetteK, AnnaS, LerB (2011). An intraspecific application of the leaf-height—Seed ecology strategy scheme to forest herbs along a latitudinal gradient. Ecography, 34, 132-140. [本文引用: 1]
[33]
PluessAR, SchützW, StöcklinJ (2005). Seed weight increases with altitude in the Swiss Alps between related species but not among populations of individual species. Oecologia, 144, 55-61. [本文引用: 1]
[34]
QiW, BuH, CornelissenJHC, ZhangC, GuoS, WangJ, ZhouX, LiW, DuG (2015). Untangling interacting mechanisms of seed mass variation with elevation: Insights from the comparison of inter-specific and intra-specific studies on eastern Tibetan angiosperm species. Plant Ecology, 216, 283-292. [本文引用: 2]
[35]
RahbekC (1997). The relationship among area, elevation, and regional species richness in Neotropical birds. The American Naturalist, 149, 875-902. [本文引用: 1]
[36]
ReesM, VenableDL (2007). Why do big plants make big seeds? Journal of Ecology, 95, 926-936. [本文引用: 1]
[37]
RockwoodLL (1985). Seed weight as a function of life form, elevation and life zone in Neotropical forests. Biotropica, 17, 32-39. [本文引用: 1]
[38]
SalisburyE (1974). Seed size and mass in relation to environment. Proceedings of the Royal Society of London. Series B, Biological Sciences, 186, 83-88. [本文引用: 1]
[39]
ShenZH, TangYY, LüN, ZhaoJ, LiDX, WangGF (2007). Community dynamics of seed rain in mixed evergreen broad-leaved and deciduous forests in a subtropical mountain of central China. Journal of Integrative Plant Biology, 49, 1294-1303. [本文引用: 1]
[40]
ShenZH, JinYX, WuJQ, ZhaoZE, HuangHD (2000). A quantitative classification of the forest communities at Dalaoling Mountain in the Three Gorges. Journal of Wuhan Botanical Research, 18, 99-107. (in Chinese with English abstract)[沈泽昊, 金义兴, 吴金清, 赵子恩, 黄汉东 (2000). 三峡大老岭地区森林群落的数量分类 . 武汉植物学研究, 18, 99-107.] [本文引用: 3]
[41]
ShenZH, WangGF, LiDX (2002). Gap related disturbance in mixed mountain forests at Mt. Dalaoling in the Three Gorges II. Topographic patterns. Acta Phytoecologica Sinica, 26, 149-156. (in Chinese with English abstract)[沈泽昊, 王功芳, 李道兴 (2002). 三峡大老岭山地常绿落叶阔叶混交林林隙干扰研究II. 林隙干扰的地形格局 . 植物生态学报, 26, 149-156.] [本文引用: 1]
[42]
ShenZH, ZhangXS, JinYX (2001). A vertical gradient analysis of the flora of Dalaoling Mountain in the Three Gorges region, China. Acta Phytotaxonomica Sinica, 39, 260-268. (in Chinese with English abstract)[沈泽昊, 张新时, 金义兴 (2001). 三峡大老岭植物区系的垂直梯度分析 . 植物分类学报, 39, 260-268.] [本文引用: 2]
[43]
SidorCG, PopaI, VladR, CherubiniP (2015). Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees, 29, 985-997. [本文引用: 1]
[44]
SinghSP, AdhikariBS, ZobelDB, SinghSP, ZobelDB (1994). Biomass, productivity, leaf longevity and forest structure in the Central Himalaya. Ecological Monographs, 64, 401-421. [本文引用: 1]
[45]
StevensonPR, PinedaM, SamperT (2005). Influence of seed size on dispersal patterns of woolly monkeys ( Lagothrix lagothricha) at Tinigua Park, Colombia. Oikos, 110, 435-440. [本文引用: 1]
[46]
SuzukiS (1998). Leaf phenology, seasonal changes in leaf quality and herbivory pattern of Sanguisorba tenuifolia at different altitudes. Oecologia, 117(1-2), 69-176. [本文引用: 1]
[47]
WalkerJJ, BeursKM, WynneRH (2014). Dryland vegetation phenology across an elevation gradient in Arizona, USA, investigated with fused MODIS and Landsat data. Remote Sensing of Environment, 144, 85-97. [本文引用: 1]
[48]
WangT, RenH, MaK (2005). Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, Northwestern China. Trees, 19, 735-741. [本文引用: 1]
[49]
WangYF, YueYC (2014). Effects of resource allocation and floral traits on the number and mass of Saussurea undulate seeds from different elevations in eastern Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology, 38, 366-374. (in Chinese with English abstract)[王一峰, 岳永成 (2014). 青藏高原东缘不同海拔波缘风毛菊资源分配及花部特征对种子数目和质量的影响 . 植物生态学报, 38, 366-374.]
[50]
WestobyM, JuradoE, LeishmanM (1992). Comparative evolutionary ecology of seed size. Trends in Ecology and Evolution, 7, 368-372. [本文引用: 2]
[51]
YingJS, ZhangZS (1984). Endemism in the flora of China: Studies on the endemic genera. Acta Phytotaxonomica Sinica, 22, 259-268. (in Chinese with English abstract)[应俊生, 张志松 (1984). 中国植物区系中的特有现象——特有属的研究 . 植物分类学报, 22, 259-268.] [本文引用: 1]
[52]
ZhangST, DuGZ, ChenJK (2003). The present situation and prospect of studies on evolutionary ecology of seed size variation. Acta Ecologica Sinica, 23, 353-364. (in Chinese with English abstract)[张世挺, 杜国祯, 陈家宽 (2003). 种子大小变异的进化生态学研究现状与展望 . 生态学报, 23, 353-364.] [本文引用: 1]
Why don’t bigger plants have proportionately bigger seeds? 1 2005
... 植物的功能性状与其生活史策略之间的关系是植物生态与进化研究的基本问题(Howe & Smallwood, 1982; Westoby et al., 1992; Moles & Westoby, 2006).种子大小(size)或质量(mass)是植物的重要生态功能性状, 对物种扩散、种群更新以及群落动态具有关键作用(Salisbury, 1974; Leishman & Murray, 2001; Stevenson et al., 2005), 并对植物群落构建的环境筛选过程与结果具有不可替代的影响(Arssen, 2005; Díaz et al., 2016). ...
Seed weight in relation to environmental conditions in California. 1 1972
... 尽管存在反例(Pluess et al., 2005), 但多数研究发现植物种子质量随海拔上升而减小(Baker, 1972; Bu et al., 2006; 郭淑青, 2007; Guo et al., 2010; Dainese & Sitzia, 2013), 并认为能量是海拔梯度上种间种子质量变化的主要限制因子(Qi et al., 2015).不过值得注意的是, 这些研究基本针对草本植物群落的物种以及同一分类群内部的物种.在仅有涉及多个生长型大量植物种的种子海拔分布研究中, 分析物种也以草本植物占绝大多数, 并且没有分生长型来分析(杜燕等, 2014; Qi et al., 2015).因此对木本植物而言, 种子质量的海拔格局及生物与非生物环境因子的相对贡献尚有待进一步研究.同时, 由于草本植物群落往往不足以覆盖研究区域的整体海拔梯度, 因而采样尺度限制有可能对结果显示的海拔格局产生影响. ...
Time-size tradeoffs: A phylogenetic comparative study of flowering time, plant height and seed mass in a north-temperate flora. 2 2008
Assessing the influence of environmental gradients on seed mass variation in mountain grasslands using a spatial phylogenetic filtering approach. 1 2013
... 尽管存在反例(Pluess et al., 2005), 但多数研究发现植物种子质量随海拔上升而减小(Baker, 1972; Bu et al., 2006; 郭淑青, 2007; Guo et al., 2010; Dainese & Sitzia, 2013), 并认为能量是海拔梯度上种间种子质量变化的主要限制因子(Qi et al., 2015).不过值得注意的是, 这些研究基本针对草本植物群落的物种以及同一分类群内部的物种.在仅有涉及多个生长型大量植物种的种子海拔分布研究中, 分析物种也以草本植物占绝大多数, 并且没有分生长型来分析(杜燕等, 2014; Qi et al., 2015).因此对木本植物而言, 种子质量的海拔格局及生物与非生物环境因子的相对贡献尚有待进一步研究.同时, 由于草本植物群落往往不足以覆盖研究区域的整体海拔梯度, 因而采样尺度限制有可能对结果显示的海拔格局产生影响. ...
The global spectrum of plant form and function. 1 2016
... 植物的功能性状与其生活史策略之间的关系是植物生态与进化研究的基本问题(Howe & Smallwood, 1982; Westoby et al., 1992; Moles & Westoby, 2006).种子大小(size)或质量(mass)是植物的重要生态功能性状, 对物种扩散、种群更新以及群落动态具有关键作用(Salisbury, 1974; Leishman & Murray, 2001; Stevenson et al., 2005), 并对植物群落构建的环境筛选过程与结果具有不可替代的影响(Arssen, 2005; Díaz et al., 2016). ...
Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. 1 2002
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
Determinants of local abundance and range size in forest vascular plants. 1 2006
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
Why are there global gradients in species richness? Mountains might hold the answer. 1 2000
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
The use of “altitude” in ecological research. 1 2007
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. 1 2001
... 种子质量大小是影响植物扩散能力和扩散方式的关键性状, 又与物种萌发率和幼苗补给(recruitment)成功率密切相关, 也是决定物种竞争能力的关键因素(Howe & Smallwood, 1982; Westoby et al., 1992).因此种子质量大小受到多种生态与进化驱动力的影响, 包括物种竞争与繁殖能力的权衡(Harper et al., 1970; Leishman, 2001; 张世挺等, 2003).这些影响因素的作用具有不同的时空尺度, 导致即使在局部群落中, 种子质量大小的种间差异也往往达到几个数量级(Hammond & Brown, 1995; Shen et al., 2007). ...
The relationship between seed size and abundance in plant communities: Model predictions and observed patterns. 1 2001
... 植物的功能性状与其生活史策略之间的关系是植物生态与进化研究的基本问题(Howe & Smallwood, 1982; Westoby et al., 1992; Moles & Westoby, 2006).种子大小(size)或质量(mass)是植物的重要生态功能性状, 对物种扩散、种群更新以及群落动态具有关键作用(Salisbury, 1974; Leishman & Murray, 2001; Stevenson et al., 2005), 并对植物群落构建的环境筛选过程与结果具有不可替代的影响(Arssen, 2005; Díaz et al., 2016). ...
神农架海拔梯度上4种典型森林的乔木叶片功能性状特征 1 2011
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
Does a latitudinal gradient in seedling survival favor larger seeds in the tropics? 2004
Latitude, seed predation and seed mass. 1 2003
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
Seed size and plant strategy across the whole life cycle. 2 2006
... 植物的功能性状与其生活史策略之间的关系是植物生态与进化研究的基本问题(Howe & Smallwood, 1982; Westoby et al., 1992; Moles & Westoby, 2006).种子大小(size)或质量(mass)是植物的重要生态功能性状, 对物种扩散、种群更新以及群落动态具有关键作用(Salisbury, 1974; Leishman & Murray, 2001; Stevenson et al., 2005), 并对植物群落构建的环境筛选过程与结果具有不可替代的影响(Arssen, 2005; Díaz et al., 2016). ...
Seed weight increases with altitude in the Swiss Alps between related species but not among populations of individual species. 1 2005
... 尽管存在反例(Pluess et al., 2005), 但多数研究发现植物种子质量随海拔上升而减小(Baker, 1972; Bu et al., 2006; 郭淑青, 2007; Guo et al., 2010; Dainese & Sitzia, 2013), 并认为能量是海拔梯度上种间种子质量变化的主要限制因子(Qi et al., 2015).不过值得注意的是, 这些研究基本针对草本植物群落的物种以及同一分类群内部的物种.在仅有涉及多个生长型大量植物种的种子海拔分布研究中, 分析物种也以草本植物占绝大多数, 并且没有分生长型来分析(杜燕等, 2014; Qi et al., 2015).因此对木本植物而言, 种子质量的海拔格局及生物与非生物环境因子的相对贡献尚有待进一步研究.同时, 由于草本植物群落往往不足以覆盖研究区域的整体海拔梯度, 因而采样尺度限制有可能对结果显示的海拔格局产生影响. ...
Untangling interacting mechanisms of seed mass variation with elevation: Insights from the comparison of inter-specific and intra-specific studies on eastern Tibetan angiosperm species. 2 2015
... 尽管存在反例(Pluess et al., 2005), 但多数研究发现植物种子质量随海拔上升而减小(Baker, 1972; Bu et al., 2006; 郭淑青, 2007; Guo et al., 2010; Dainese & Sitzia, 2013), 并认为能量是海拔梯度上种间种子质量变化的主要限制因子(Qi et al., 2015).不过值得注意的是, 这些研究基本针对草本植物群落的物种以及同一分类群内部的物种.在仅有涉及多个生长型大量植物种的种子海拔分布研究中, 分析物种也以草本植物占绝大多数, 并且没有分生长型来分析(杜燕等, 2014; Qi et al., 2015).因此对木本植物而言, 种子质量的海拔格局及生物与非生物环境因子的相对贡献尚有待进一步研究.同时, 由于草本植物群落往往不足以覆盖研究区域的整体海拔梯度, 因而采样尺度限制有可能对结果显示的海拔格局产生影响. ...
... ; Qi et al., 2015).因此对木本植物而言, 种子质量的海拔格局及生物与非生物环境因子的相对贡献尚有待进一步研究.同时, 由于草本植物群落往往不足以覆盖研究区域的整体海拔梯度, 因而采样尺度限制有可能对结果显示的海拔格局产生影响. ...
The relationship among area, elevation, and regional species richness in Neotropical birds. 1 1997
... 海拔是山地植被分异的主导环境梯度(Ko?rner, 2007), 也是生物多样性宏观分布格局的重要调控因素(Rahbek, 1997; Ko?rner, 2000).海拔对植物物种的分布具有多重影响, 能量和降水都具有显著的海拔变化, 因而生态系统的复杂性、功能强度(如植被生产力)、种间相互作用, 也随着植被的海拔带谱显示了梯度变化(Suzuki, 1998; Kitayama & Aiba, 2002); 另一方面, 在山地的海拔梯度上, 各种人类活动、动物觅食和地貌、水文过程对植被产生显著的干扰(沈泽昊等, 2002; Gentili et al., 2013; Montaño-Centellas & Garitano-Zavala, 2015).物种功能性状沿海拔梯度的变化及其限制因子是理解物种海拔分布的关键.研究表明, 植物的功能性状, 如物候(Fielding et al., 1999; Walker et al., 2014)、年轮宽度(Wang et al., 2005; Sidor et al., 2015)、叶片性状(Singh et al., 1994; 罗璐等, 2011; Jiang & Ma, 2015)等, 都受到海拔梯度的环境变化影响, 并成为物种海拔分布范围阈值形成的非生物(如低温限制)和生物(如种间竞争)机制.众所周知, 种子的生产和萌发更新是植物生命史中对环境变化最为敏感的环节, 往往构成植物分布边界的限制因子.因此, 种子性状, 如大小、数量沿气候梯度的变化应与植物种分布范围存在密切的联系(Moles & Westoby, 2003; Kolb et al., 2006). ...
 , 徐慎东
, 徐慎东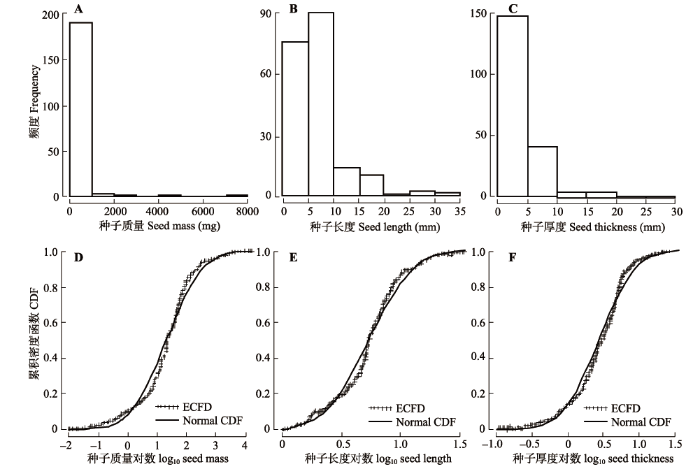 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT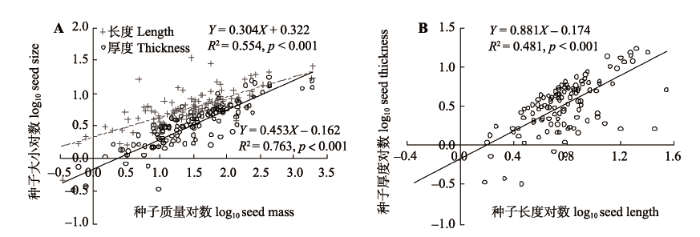 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT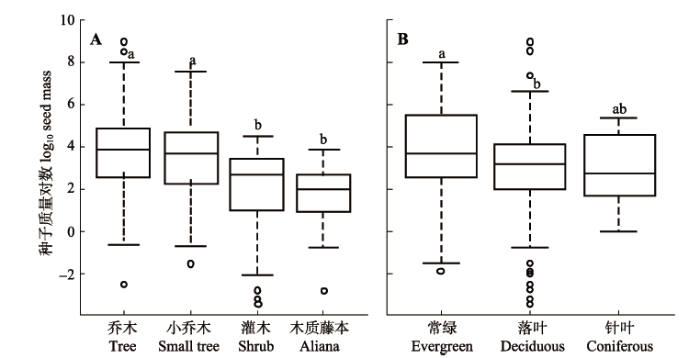 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT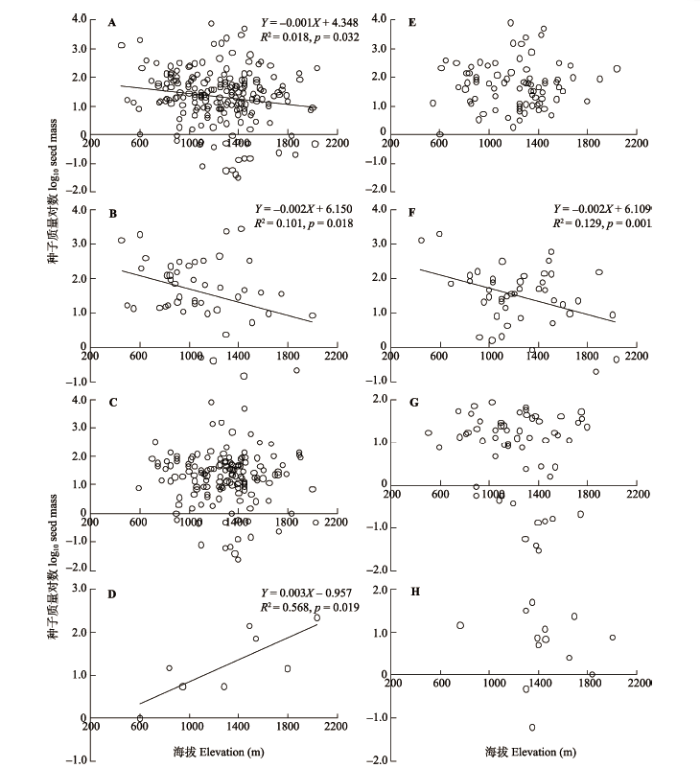 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}