关键词:深色有隔内生真菌;群落组成;物种多样性;沙冬青;伴生植物 Abstract Aims Dark septate endophytes (DSE) extensively colonize in plant root tissues in different ecosystems. However, our understanding of the ecological significance of DSE in semiarid and arid lands is limited. The main objective of this study was to compare the community composition and species diversity of DSE in Ammopiptanthus mongolicus and its companion plants in desert habitats. Methods Healthy root samples of Ammopiptanthus mongolicus’s companion plants, Artemisia desertorum, Caragana rosea, Nitraria tangutorum and Caragana korshinskii, were collected separately from four main locations of Ammopiptanthus mongolicus plantation in Dengkou of Nei Mongol, Minqin of Gansu, Yinchuan and Shapotou of Ningxia in July 2013. Important findings A total of 103 strains belonging to seven species (i.e Alternaria, Curvularia, Embellisia, Phialocephala, Phoma, Podospora and Pleosporales) of DSE were isolated from the four types of companion plants by means of morphological identification and molecular identification. Among the seven genera, Alternaria, Embellisia, Curvularia and Podospora are found for the first time in desert ecosystems. Ammopiptanthus mongolicus and its companion plants at the same site had significantly different community composition of DSE. Ammopiptanthus mongolicus and its companion plants at different sites had significantly different DSE diversity index. The colonization and species richness of DSE, especially the species and the quantity of spores were higher in roots of Ammopiptanthus mongolicus than in its companion plants. This study supports the results that DSE can form a better symbiotic relationship with the roots of Ammopiptanthus mongolicus than with its companion plants, and can enhance the ecological adaptability of Ammopiptanthus mongolicus to the extreme desert environment.
Keywords:dark septate endophyte;community composition;species diversity;Ammopiptanthus mongolicus;companion plants -->0 PDF (2642KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 孙茜, 薛子可, 解琳琳, 贺学礼, 赵丽莉. 沙冬青及其伴生植物深色有隔内生真菌物种多样性. 植物生态学报, 2017, 41(7): 729-737 https://doi.org/10.17521/cjpe.2016.0270 SUNQian, XUEZi-Ke, XIELin-Lin, HEXue-Li, ZHAOLi-Li. Diversity of dark septate endophyte in the roots of Ammopiptanthus mongolicus and its companion plants. Chinese Journal of Plant Ecology, 2017, 41(7): 729-737 https://doi.org/10.17521/cjpe.2016.0270 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008)。已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用。与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006)。Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008)。因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007)。 荒漠生态系统中植物和微生物的生长和分布及其相互作用是荒漠生物学过程的重要部分, 揭示植物、微生物与环境之间的相互作用, 有助于我们深刻理解荒漠生态系统过程和植被演变规律; 而DSE作为荒漠化过程中非常重要的一类内生真菌, 能够连接地上生态系统和地下生态系统物质传输, 调节植物群落结构、植物多样性以及植物生态系统的生产力(Jumpponen, 2001; Porras-Alfaro et al., 2008; Li et al., 2015; 张娟等, 2015)。沙冬青(Ammopiptanthus mongolicus)是第三纪残遗种, 也是我国干旱荒漠区惟一超旱生常绿阔叶豆科灌木, 具有很强的抗逆性(杨期和等, 2004)。张淑容等(2013)和姜桥等(2014)研究发现, 荒漠植物根组织有较高的DSE定殖率。Li等(2015)研究了内蒙古地区沙冬青DSE和土壤因子的相关性, 并分离鉴定了Exophiala sp.、Phialophora sp.和Phialocephala sp. 3个DSE菌株; 陈严严等(2014)研究了甘肃和宁夏3个样地沙冬青DSE共生及分离培养特征, 共分离鉴定了10种DSE。在西北荒漠带, 同沙冬青相伴生长着一些生长年限较短的灌木和半灌木, 这些伴生植物与沙冬青既存在竞争抑制, 又协同进化, 与沙冬青共同发挥防风固沙的作用(孙茜等, 2015)。 本研究在前期对沙冬青及其伴生植物DSE定殖规律分析(孙茜等, 2015)的基础上, 分离培养定殖于根组织的DSE, 比较分析沙冬青及其伴生植物DSE群落结构和物种多样性的差异性, 为阐明沙冬青适应极端荒漠环境的机理提供依据。
新窗口打开 显示原图|下载原图ZIP|生成PPT 图3基于伴生植物深色有隔内生真菌ITS1-5.8S-ITS2序列构建的NJ树。DK-1、DK-0-10、DK-4是磴口样地伴生植物3种深色有隔内生真菌ITS1-5.8S-ITS2序列编号; MQ-1、MQ-4是民勤样地伴生植物两种深色有隔内生真菌ITS1-5.8S-ITS2序列编号; YC-1、YC-2-1、YC-3是银川样地伴生植物3种深色有隔内生真菌ITS1-5.8S-ITS2序列编号; SPT-1是沙坡头样地伴生植物深色有隔内生真菌ITS1-5.8S-ITS2序列编号。图中其他编号均为伴生植物深色有隔内生真菌在GenBank中Blast比对得到的ITS1-5.8S-ITS2序列的同源序列号。 -->Fig. 3Neighbor-Joining tree of dark septate endophyte in the roots of the companion plants of Ammopiptanthus mongolicus. DK-1, DK-0-10, DK-4 represent the sequence numbers of ITS1-5.8S-ITS2 form three kinds of dark septate endophyte in the roots of the companion plants of Ammopiptanthus mongolicus in Dengkou; MQ-1, MQ-4, represent the sequence numbers of ITS1-5.8S-ITS2 form two kinds of dark septate endophyte in the roots of the companion plants of Ammopiptanthus mongolicus in Minqin; YC-1, YC- 2-1, YC-3, represent the sequence numbers of ITS1-5.8S-ITS2 form three kinds of dark septate endophyte in the roots of the companion plants of Yinchuan; SPT-1 represents the sequence numbers of ITS1-5.8S-ITS2 form dark septate endophyte in the roots of the companion plants of Ammopiptanthus mongolicus in Shapotou. Other accession numbers in the figure are homologous sequence of dark septate endophyte in the roots of the companion plants of Ammopiptanthus mongolicus.obtained by Blast. -->
2.3 沙冬青与伴生植物深色有隔内生真菌多样性分析
2.3.1 深色有隔内生真菌分离频率 由表4可知, 在各样地伴生植物中, 磴口样地沙蒿的优势种为Alternaria alternata, 民勤样地红花锦鸡儿仅有Curvularia eragrostidis和Pleosporales sp., 银川样地白刺优势种为Embellisia属菌株, 沙坡头柠条锦鸡儿仅有Phoma medicaginis。不同样地DSE分离频率差异显著, 其中磴口最高为17.19%; 民勤最低为3.86%, 样地间表现为: 磴口>银川>沙坡头>民勤。各样地伴生植物均无共有种。 Table 4 表4 表4沙冬青及其伴生植物深色有隔内生真菌分离频率 Table 4Isolation frequency of dark septate endophyte in Ammopiptanthus mongolicus and its companion plants
样地 Site
植物 Plant
菌株编号 Number of strains
分类 Classification
菌株数量 Quantity of strains
分离频率 Isolation frequency (%)
磴口 Dengkou
沙冬青 Ammopiptanthus mongolicus
DK-S
Cladosporium sp.
16
5.61
DK-1
Podospora sp.
16
5.61
沙蒿 Artemisia desertorum
DK-0-10
Alternaria alternate
23
8.07
DK-4
Alternaria sp.
10
3.51
总菌株数 Total strains
65
22.74
民勤 Minqin
沙冬青 Ammopiptanthus mongolicus
0-10MQ1217-2-2
Paraphoma chrysanthemicola
30
10.53
2MQ0-10
Phoma sp.
23
8.07
MQ1125
Pleosporales sp.
12
4.21
0-10MQ1217-3
Cladosporium sphaerospermum
16
5.61
红花锦鸡儿 Caragana rosea
MQ-1
Pleosporales sp.
6
2.11
MQ-4
Curvularia eragrostidis
5
1.75
总菌株数 Total strains
92
32.29
银川 Yichuan
沙冬青 Ammopiptanthus mongolicus
8YC
Exophiala cancerae
8
2.81
4YC
Phoma sp.
9
3.16
白刺 Nitraria tangutorum
YC-1
Alternaria japonica
8
2.81
YC-2-1
Embellisia sp.
15
5.26
YC-3
Phialocephala fluminis
3
1.06
总菌株数 Total strains
43
15.08
沙坡头 Shapotou
沙冬青 Ammopiptanthus mongolicus
2SPT1218
Cladosporium cladosporioides
12
4.21
3ASPT1218
Exophiala sp.
28
9.83
6SPT
Phialophora mustea
13
4.56
SPT-1125
Exophiala salmonis
15
5.26
柠条锦鸡儿 Caragana korshinskii
SPT-1
Phoma medicaginis
17
5.96
总菌株数 Total strains
85
29.89
合计 Total
285
100
新窗口打开 在各样地沙冬青中, 磴口样地仅有Cladosporium属菌株, 民勤样地优势种为Paraphoma chrysanthemicola, 银川样地有Exophiala cancerae和Phoma sp., 沙坡头样地优势种为Exophiala属菌株。不同样地DSE分离频率差异显著, 其中民勤最高为28.42%; 磴口最低为5.61%, 样地间表现为: 民勤>沙坡头>银川>磴口。仅Pleosporales目是沙冬青与其伴生植物在民勤样地共有菌株。 2.3.2 深色有隔内生真菌属水平多样性测度 由表5可知, 样地间, 沙冬青及其伴生植物DSE种类组成存在差异, 从属水平看, 民勤的沙冬青DSE种类最多, 沙坡头次之, 而银川伴生植物白刺DSE种类比沙坡头柠条锦鸡儿多, Alternaria sp.、Embellisia sp.、Phialocephala sp.、Podospora sp.、Curvularia sp.仅存在于伴生种中, 而Paraphoma sp.、Exophiala sp.、Cladosporium sp.、Phialophora sp. 仅存在于沙冬青中, Pleosporales sp.和Phoma sp.在沙冬青和伴生植物中均有分布。 Table 5 表5 表5不同样地沙冬青及其伴生植物深色有隔内生真菌类群比较 Table 5The comparison between dark septate endophyte in roots of Ammopiptanthus mongolicus and that in A. mongolicus’s companion plants from different sampling sites
深色有隔内 生真菌种类 Species of dark septate endophyte
沙冬青 Ammopiptanthus mongolicus
沙蒿 Artemisia desertorum
红花锦鸡儿 Caragana rosea
白刺 Nitraria tangutorum
柠条锦鸡儿 Caragana korshinskii
磴口 Dengkou
民勤 Minqin
银川 Yinchuan
沙坡头 Shapotou
磴口 Dengkou
民勤 Minqin
银川 Yinchuan
沙坡头 Shapotou
Alternaria sp.
-
-
-
-
+
-
+
-
Embellisia sp.
-
-
-
-
-
-
+
-
Phialocephala sp.
-
-
-
-
-
-
+
-
Pleosporales sp.
-
+
-
-
-
+
-
-
Podospora sp.
-
-
-
-
+
-
-
-
Curvularia sp.
-
-
-
-
-
+
-
-
Phoma sp.
-
+
+
-
-
-
-
+
Paraphoma sp.
-
+
-
-
-
-
-
-
Exophiala sp.
-
-
+
+
-
-
-
-
Cladosporium sp.
+
+
-
+
-
-
-
-
Phialophora sp.
-
-
-
+
-
-
-
-
-, no dark septate endophyte observed; +, presence of dark septate endophyte.-, 未出现深色有隔内生真菌; +, 出现深色有隔内生真菌。 新窗口打开 由表2和表6可知, 不同样地沙冬青与其伴生植物DSE多样性指数差异显著, 在一定程度上反映出群落DSE总数及分布数量的不同。其中, 多样性指数最大值出现在民勤沙冬青中, 为0.735 1, 最小值出现在红花锦鸡儿中, 为0.152 2; 均匀度指数、多样性指数和分离频率在样地间变化规律一致。而Simpson指数变化趋势与前三者相反, 以民勤沙冬青最高, 红花锦鸡儿最低。 Table 6 表6 表6沙冬青及其伴生植物深色有隔内生真菌物种多样性 Table 6Species diversity of dark septate endophyte in the roots of Ammopiptanthus mongolicus and its companion plants
样地 Sampling sites
植物 Plant
多样性指数 Diversity index
优势度 Dominance
均匀度 Uniformity
磴口 Dengkou
沙冬青 Ammopiptanthus mongolicus
0.161 6b
0.996 8a
-
沙蒿 Artemisia desertorum
0.482 3a
0.989 1a
0.439 0
民勤 Minqin
沙冬青 Ammopiptanthus mongolicus
0.735 1a
0.977 5a
0.530 3a
红花锦鸡儿 Caragana rosea
0.152 2b
0.999 2a
0.219 6b
银川 Yinchuan
沙冬青 Ammopiptanthus mongolicus
0.209 5b
0.998 2a
0.302 3a
白刺 Nitraria tangutorum
0.303 4a
0.996 3a
0.276 2b
沙坡头 Shapotou
沙冬青 Ammopiptanthus mongolicus
0.657 1a
0.983 7a
0.474 0
柠条锦鸡儿 Caragana korshinskii
0.168 1b
0.996 4a
-
Different letters indicate significant differences between diversity of dark septate endophyte in Ammopiptanthus mongolicus and that in Ammopiptanthus mongolicus’s companion plants in the same sampling site (p < 0.05).同列数据后不同小写字母表示同一样地沙冬青与伴生植物深色有隔内生真菌多样性差异显著(p < 0.05)。 新窗口打开
3 讨论
3.1 DSE群落组成和物种多样性
DSE广泛分布于各类生境, 尤其是广泛分布于胁迫环境和受干扰严重地区(Knapp et al., 2012; Postma et al., 2007; Zhao et al., 2015)。目前, 全世界已发现110科600余种植物根组织有DSE定殖(Mandyam & Jumpponen, 2005), 虽然DSE能在体外纯培养, 但大多数纯培养条件下, 无法还原DSE在植物体内的真实生境而不产孢, 加之DSE本身涵盖众多科属, 其组成的复杂性和不明确性, 使得仅通过形态特征难以确定DSE的分类地位, 分子测序技术极大地推动了DSE的分类研究(Bonfim et al., 2016; Yadav et al., 2016)。按照Sieber和Grünig (2013)对DSE的定义, 本研究分离的菌株均有DSE典型黑化结构, 不同样地的DSE菌株平均分离率为7.48%, 低于沙冬青DSE菌株10.0%的分离率(陈严严等, 2014)。 本研究每个样地都选取300条根段进行DSE分离培养, 能够反映该样地DSE群落组成情况(Bonfim et al., 2016), 在分离的DSE中, Alternaria、Embellisia、Curvularia和Podospora均为荒漠生态系统未见报道的DSE属, 其中Phialocephala、Phoma和Pleosporales 三类DSE在沙冬青根中也有分布(陈严严等, 2014; Li et al., 2015)。与沙冬青DSE多样性对比发现, 磴口样地沙冬青仅有Cladosporium sp., 与伴生植物沙蒿没有共有属种; 民勤样地沙冬青与伴生植物红花锦鸡儿共有菌株为Pleosporales, 但沙冬青DSE优势种为Paraphoma chrysanthemicola, 银川与沙坡头样地沙冬青与其伴生植物没有共有属种。同时, 沙冬青DSE分离率和多样性指数为民勤>沙坡头>银川, 而伴生植物为银川>沙坡头>民勤。说明DSE群落组成和物种多样性具有空间异质性, 并与植物种类和生长特性密切相关(Mandyam et al., 2012; Lugo et al., 2015)。
3.2 DSE与植物生态适应性的关系
许多研究表明, DSE与植物互惠共生, 赋予植物优良生长性状, 促进植物营养吸收, 提高植物抗病性及胁迫环境中的抗逆性(Usuki & Narisawa, 2007; Likar & Regvar, 2013; Mahmoud & Narisawa, 2013; Zhao et al., 2015)。与陈严严(2014)等的研究结果对比可知, 在民勤、银川和沙坡头样地, 沙冬青DSE平均总定殖率及种类数量显著高于该样地伴生植物。同时发现, 甘肃、宁夏地区沙冬青可培养DSE共6属10种, 4种产孢, 1种10 ℃诱导后产孢, 其余不产孢, 而伴生植物仅6属6种, 2种产孢, 1种诱导后产孢, 其余不产孢; 磴口样地, 沙冬青仅分离1属1种产孢DSE, 而伴生植物有2属3种, 1种在诱导后产孢(Li et al., 2015)。Mandyam和Jumpponen (2015)研究发现, 与植物有良好共生关系的内生真菌可以促进植物对生存环境的适应能力, 宿主植物进化与DSE丰富度协同一致。伴生植物与第三纪孑遗植物沙冬青相伴相生, 在漫长的进化过程中与DSE协同发展, 与伴生植物相比, DSE可能与沙冬青根系能够形成更好的共生关系, 增强了沙冬青对极端荒漠环境的生态适应性(Mandyam & Jumpponen, 2005; Wu & Guo, 2008; Lugo et al., 2015)。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
BanY, TangM, ChenH, XuZ, ZhangH, YangY (2012). The response of dark septate endophytes (DSE) to heavy metals in pure culture. PLOS ONE, 7, e47968. doi: 10.1371/journal.pone.0047968. [本文引用: 1]
[2]
BonfimJA, VasconcellosRLF, BaldesinLF, SieberTN, CardosoEJBN (2016). Dark septate endophytic fungi of native plants along an altitudinal gradient in the Brazilian Atlantic forest. Fungal Ecology, 20, 202-210. [本文引用: 2]
[3]
ChenYY, HeXL, LiBK, ChengCQ, JiangQ, ChenWY (2014). The symbiotic and isolated culture characteristics of DSE in root of Ammopiptanthus mongolicus. Journal of Agricultural University of Hebei, 37, 18-24. (in Chinese with English abstract)[陈严严, 贺学礼, 李宝库, 程春泉, 姜桥, 陈伟燕 (2014). 沙冬青根系深色有隔内生真菌共生及分离培养特征研究 . 河北农业大学学报, 37, 18-24.] [本文引用: 3]
[4]
DengX, SongXS, YinDC, SongRQ (2015). Effects of salt stress on growth and nutritional metabolism of two dark septate endophyte. Journal of Central South University of Forestry & Technology, 35(5), 1-7. (in Chinese with English abstract)[邓勋, 宋小双, 尹大川, 宋瑞清 (2015). 盐胁迫对2株深色有隔内生真菌(DSE)生长及营养代谢的影响 . 中南林业科技大学学报, 35(5), 1-7.] [本文引用: 1]
[5]
GrünigCR, QuelozV, SieberTN, HoldenriederO (2008). Dark septate endophytes (DSE) of the Phialocephala fortinii-Acephala applanata species complex in tree roots: Classification, population biology, and ecology. Botany-Botanique, 86, 1355-1369. [本文引用: 1]
[6]
JiangQ, HeXL, ChenWY, ZhangYJ, RongXR, WangL (2014). Spatial distribution of AM and DSE fungi in the rhizosphere of Ammopiptanthus nanus. Acta Ecologica Sinica, 34, 2929-2937. (in Chinese with English abstract)[姜桥, 贺学礼, 陈伟燕, 张玉洁, 荣心瑞, 王雷 (2014).新疆沙冬青AM和DSE真菌的空间分布 . 生态学报, 34, 2929-2937.] [本文引用: 1]
[7]
JumpponenA (2001). Dark septate endophytes—Are they mycorrhizal? Mycorrhiza, 11, 207-211. [本文引用: 2]
[8]
KauppinenM, RavealaK, WäliPR, RuotsalainenAL (2014). Contrasting preferences of arbuscular mycorrhizal and dark septate fungi colonizing boreal and subarctic Avenella flexuosa. Mycorrhiza, 24, 171-177. [本文引用: 1]
[9]
KnappDG, PintyeA, KovacsGM (2012). The dark side is not fastidious-dark septate endophytic fungi of native and invasive plants of semiarid sandy areas. PLOS ONE, 7, e32570. doi: 10.1371/journal.pone.0032570. [本文引用: 1]
[10]
LiBK, HeXL, HeC, ChenYY, WangXQ (2015). Spatial dynamics of dark septate endophytes and soil factors in the rhizosphere of Ammopiptanthus mongolicus in Inner Mongolia, China .Symbiosis, 65, 75-84. [本文引用: 3]
[11]
LikarM, RegvarM (2013). Isolates of dark septate endophytes reduce metal uptake and improve physiology of Salix caprea L. Plant and Soil, 370, 593-604. [本文引用: 1]
[12]
LugoMA, ReinhartKO, MenoyoE, CrespoEM, UrcelayC (2015). Plant functional traits and phylogenetic relatedness explain variation in associations with root fungal endophytes in an extreme arid environment. Mycorrhiza, 25, 85-95. [本文引用: 2]
[13]
MahmoudRS, NarisawaK (2013). A new fungal endophyte, Scolecobasidium humicola, promotes tomato growth under organic nitrogen conditions. PLOS ONE, 8, e78746. doi: 10.1371/journal.pone.0078746. [本文引用: 1]
[14]
MandyamK, FoxC, JumpponenA (2012). Septate endophyte colonization and host responses of grasses and forbs native to a tallgrass prairie. Mycorrhiza, 22, 109-119. [本文引用: 1]
[15]
MandyamK, JumpponenA (2005). Seeking the elusive function of the root-colonising dark septate endophytic fungi. Studies in Mycology, 53, 173-189. [本文引用: 2]
[16]
MandyamK, JumpponenA (2008). Seasonal and temporal dynamics of arbuscular mycorrhizal and dark septate endophytic fungi in a tallgrass prairie ecosystem are minimally affected by nitrogen enrichment. Mycorrhiza, 18, 145-155. [本文引用: 1]
[17]
MandyamK, JumpponenA (2015). Mutualism-parasitism paradigm synthesized from results of root-endophyte models. Frontiers in Microbiology, 5, 1-13.
[18]
MuthukumarT, SenthilkumarM, RajangamM (2006). Arbuscular mycorrhizal morphology and dark septate fungal associations in medicinal and aromatic plants of Western Ghats, Southern India. Mycorrhiza, 17, 11-24. [本文引用: 1]
[19]
Porras-AlfaroA, HerreraJ, SinsabaughRL, OdenbachKJ, LowreyT, NatvigDO (2008). Novel root fungal consortium associated with a dominant desert grass. Applied and Environment Microbiology, 74, 2805-2813. [本文引用: 1]
[20]
PostmaJWM, OlssonPA, Kalkengren-GrerupU (2007). Root colonization by arbuscular mycorrhizal, fine endophytic and dark septate fungi across a pH gradient in acid beech forests. Soil Biology & Biochemistry, 39, 400-408. [本文引用: 1]
[21]
RudgersJA, ClayK (2007). Endophyte symbiosis with tall fescue: How strong are the impacts on communities and ecosystems? Fungal Biology Reviews, 21, 107-124. [本文引用: 1]
[22]
ScervinoJM, GottliebA, SilvaniVA (2009). Exudates of dark septate endophyte (DSE) modulate the development of the arbuscular mycorrhizal fungus (AMF) Gigaspora rosea. Soil Biology & Biochemistry, 41, 1753-1756.
[23]
SieberTN, GrünigCR (2013). Fungal root endophytes. In: Eshel A, Beeckman T eds. Plant Roots—The Hidden Half. 4th edn. CRC Press, Florida, USA. 38-49.
[24]
SunQ, HeC, HeXL, ZhaoLL (2015). Colonization of dark septate endophytes in roots of Ammopiptanthus mongolicus and its associated plants as influenced by soil properties. Chinese Journal of Plant Ecology, 39, 878-889. (in Chinese with English abstract)[孙茜, 贺超, 贺学礼, 赵丽莉 (2015). 沙冬青与伴生植物深色有隔内生真菌定殖规律及其与土壤因子的相关性 . 植物生态学报, 39, 878-889.] [本文引用: 2]
[25]
UsukiF, NarisawaK (2007). A mutualistic symbiosis between a dark septate endophytic fungus, Heteroconium chaetospira, and a nonmycorrhizal plant, Chinese cabbage. Mycorrhiza, 99, 175-184. [本文引用: 1]
[26]
WuL, GuoS (2008). Interaction between an isolate of dark- septate fungi and its host plant Saussurea involucrata. Mycorrhiza, 18, 79-85. [本文引用: 1]
[27]
YadavM, YadavA, KumarS, YadavJP (2016). Spatial and seasonal influences on culturable endophytic mycobiota associated with different tissues of Eugenia jambolana Lam. and their antibacterial activity against MDR strains. BMC Microbiology, 16, 1-12. [本文引用: 2]
[28]
YanJ, HeXL, ZhangYJ, XuW, ZhangJ, ZhaoLL (2014). Colonization of arbuscular mycorrhizal fungi and dark septate endophytes in roots of desert Salix psammophila. Chinese Journal of Plant Ecology, 38, 949-958. (in Chinese with English abstract)[闫姣, 贺学礼, 张亚娟, 许伟, 张娟, 赵丽莉 (2014). 荒漠北沙柳根系丛枝菌根真菌和黑隔内生真菌定殖状况 . 植物生态学报, 38, 949-958.] [本文引用: 1]
[29]
YangQH, GeXJ, YeWH, DengX, LiaoFL (2004). Characteristics of Ammopiptanthus nanus seed and factors affecting its germination. Acta Phytoecologica Sinica, 28, 651-656. (in Chinese with English abstract)[杨期和, 葛学军, 叶万辉, 邓雄, 廖富林 (2004). 矮沙冬青种子特性和萌发影响因素的研究 . 植物生态学报, 28, 651-656.] [本文引用: 1]
[30]
YuZS, ZhangBH, SunW, ZhangFL, LiZY (2013). Phylogenetically diverse endozoic fungi in the South China Sea sponges and their potential in synthesizing bioactive natural products suggested by PKS gene and cytotoxic activity analysis. Fungal Diversity, 58, 127-141. [本文引用: 1]
[31]
ZhanFD, HeYM, LiT, YangYY, GurpalST, ZhaoZW (2015). Tolerance and antioxidant response of a dark septate endophyte (DSE), Exophiala pisciphila, to cadmium stress. Bulletin Environmental Contamination Toxicology, 94, 96-102. [本文引用: 1]
[32]
ZhangJ, HeXL, ZhaoLL, XuW, YanJ (2015). Responses of desert soil factors and dark septate endophytes colonization to clonal plants invasion. Acta Ecologica Sinica, 35, 1095-1103. (in Chinese with English abstract)[张娟, 贺学礼, 赵丽莉, 许伟, 闫娇 (2015). 荒漠土壤因子和DSE定殖对克隆植物入侵的响应 . 生态学报, 35, 1095-1103.] [本文引用: 1]
[33]
ZhangSR, HeXL, XuHB, LiuCM, NiuK (2013). Correlation study of AM and DSE fungi and soil factors in the rhizosphere of Ammopiptanthus mongolicus. Acta Botanica Boreali-Occidentalia Sinica, 33, 1891-1897. (in Chinese with English abstract)[张淑容, 贺学礼, 徐浩博, 刘春卯, 牛凯 (2013). 蒙古沙冬青根围AM和DSE真菌与土壤因子的相关性研究 . 西北植物学报, 33, 1891-1897.]
[34]
ZhaoD, TaoL, ShenM, WangJ, ZhaoZ (2015). Diverse strategies conferring extreme cadmium (Cd) tolerance in the dark septate endophyte (DSE), Exophiala pisciphila: Evidence from RNA-seq data. Microbiological Research, 170, 27-35. [本文引用: 2]
The response of dark septate endophytes (DSE) to heavy metals in pure culture. 1 2012
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Dark septate endophytic fungi of native plants along an altitudinal gradient in the Brazilian Atlantic forest. 2 2016
... DSE广泛分布于各类生境, 尤其是广泛分布于胁迫环境和受干扰严重地区(Knapp et al., 2012; Postma et al., 2007; Zhao et al., 2015).目前, 全世界已发现110科600余种植物根组织有DSE定殖(Mandyam & Jumpponen, 2005), 虽然DSE能在体外纯培养, 但大多数纯培养条件下, 无法还原DSE在植物体内的真实生境而不产孢, 加之DSE本身涵盖众多科属, 其组成的复杂性和不明确性, 使得仅通过形态特征难以确定DSE的分类地位, 分子测序技术极大地推动了DSE的分类研究(Bonfim et al., 2016; Yadav et al., 2016).按照Sieber和Grünig (2013)对DSE的定义, 本研究分离的菌株均有DSE典型黑化结构, 不同样地的DSE菌株平均分离率为7.48%, 低于沙冬青DSE菌株10.0%的分离率(陈严严等, 2014). ...
... 本研究每个样地都选取300条根段进行DSE分离培养, 能够反映该样地DSE群落组成情况(Bonfim et al., 2016), 在分离的DSE中, Alternaria、Embellisia、Curvularia和Podospora均为荒漠生态系统未见报道的DSE属, 其中Phialocephala、Phoma和Pleosporales 三类DSE在沙冬青根中也有分布(陈严严等, 2014; Li et al., 2015).与沙冬青DSE多样性对比发现, 磴口样地沙冬青仅有Cladosporium sp., 与伴生植物沙蒿没有共有属种; 民勤样地沙冬青与伴生植物红花锦鸡儿共有菌株为Pleosporales, 但沙冬青DSE优势种为Paraphoma chrysanthemicola, 银川与沙坡头样地沙冬青与其伴生植物没有共有属种.同时, 沙冬青DSE分离率和多样性指数为民勤>沙坡头>银川, 而伴生植物为银川>沙坡头>民勤.说明DSE群落组成和物种多样性具有空间异质性, 并与植物种类和生长特性密切相关(Mandyam et al., 2012; Lugo et al., 2015). ...
... DSE广泛分布于各类生境, 尤其是广泛分布于胁迫环境和受干扰严重地区(Knapp et al., 2012; Postma et al., 2007; Zhao et al., 2015).目前, 全世界已发现110科600余种植物根组织有DSE定殖(Mandyam & Jumpponen, 2005), 虽然DSE能在体外纯培养, 但大多数纯培养条件下, 无法还原DSE在植物体内的真实生境而不产孢, 加之DSE本身涵盖众多科属, 其组成的复杂性和不明确性, 使得仅通过形态特征难以确定DSE的分类地位, 分子测序技术极大地推动了DSE的分类研究(Bonfim et al., 2016; Yadav et al., 2016).按照Sieber和Grünig (2013)对DSE的定义, 本研究分离的菌株均有DSE典型黑化结构, 不同样地的DSE菌株平均分离率为7.48%, 低于沙冬青DSE菌株10.0%的分离率(陈严严等, 2014). ...
... 本研究每个样地都选取300条根段进行DSE分离培养, 能够反映该样地DSE群落组成情况(Bonfim et al., 2016), 在分离的DSE中, Alternaria、Embellisia、Curvularia和Podospora均为荒漠生态系统未见报道的DSE属, 其中Phialocephala、Phoma和Pleosporales 三类DSE在沙冬青根中也有分布(陈严严等, 2014; Li et al., 2015).与沙冬青DSE多样性对比发现, 磴口样地沙冬青仅有Cladosporium sp., 与伴生植物沙蒿没有共有属种; 民勤样地沙冬青与伴生植物红花锦鸡儿共有菌株为Pleosporales, 但沙冬青DSE优势种为Paraphoma chrysanthemicola, 银川与沙坡头样地沙冬青与其伴生植物没有共有属种.同时, 沙冬青DSE分离率和多样性指数为民勤>沙坡头>银川, 而伴生植物为银川>沙坡头>民勤.说明DSE群落组成和物种多样性具有空间异质性, 并与植物种类和生长特性密切相关(Mandyam et al., 2012; Lugo et al., 2015). ...
盐胁迫对2株深色有隔内生真菌(DSE)生长及营养代谢的影响 1 2015
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Dark septate endophytes (DSE) of the Phialocephala fortinii-Acephala applanata species complex in tree roots: Classification, population biology, and ecology. 1 2008
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
新疆沙冬青AM和DSE真菌的空间分布 1 2014
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Dark septate endophytes—Are they mycorrhizal? 2 2001
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Plant functional traits and phylogenetic relatedness explain variation in associations with root fungal endophytes in an extreme arid environment. 2 2015
... 本研究每个样地都选取300条根段进行DSE分离培养, 能够反映该样地DSE群落组成情况(Bonfim et al., 2016), 在分离的DSE中, Alternaria、Embellisia、Curvularia和Podospora均为荒漠生态系统未见报道的DSE属, 其中Phialocephala、Phoma和Pleosporales 三类DSE在沙冬青根中也有分布(陈严严等, 2014; Li et al., 2015).与沙冬青DSE多样性对比发现, 磴口样地沙冬青仅有Cladosporium sp., 与伴生植物沙蒿没有共有属种; 民勤样地沙冬青与伴生植物红花锦鸡儿共有菌株为Pleosporales, 但沙冬青DSE优势种为Paraphoma chrysanthemicola, 银川与沙坡头样地沙冬青与其伴生植物没有共有属种.同时, 沙冬青DSE分离率和多样性指数为民勤>沙坡头>银川, 而伴生植物为银川>沙坡头>民勤.说明DSE群落组成和物种多样性具有空间异质性, 并与植物种类和生长特性密切相关(Mandyam et al., 2012; Lugo et al., 2015). ...
Seasonal and temporal dynamics of arbuscular mycorrhizal and dark septate endophytic fungi in a tallgrass prairie ecosystem are minimally affected by nitrogen enrichment. 1 2008
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Mutualism-parasitism paradigm synthesized from results of root-endophyte models. 2015
Arbuscular mycorrhizal morphology and dark septate fungal associations in medicinal and aromatic plants of Western Ghats, Southern India. 1 2006
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Novel root fungal consortium associated with a dominant desert grass. 1 2008
Spatial and seasonal influences on culturable endophytic mycobiota associated with different tissues of Eugenia jambolana Lam. and their antibacterial activity against MDR strains. 2 2016
... 选用核糖体内转录间隔区(ITS)基因序列对不产孢菌株进行分子鉴定(Yadav et al., 2016).每个菌种选择2-5株菌株, 在PDA平板上27 ℃培养14天后, 刮取新鲜菌丝10 mg, 置于Eppendorf管中, 杵棒研磨充分后, 采用真菌基因组DNA提取试剂盒(北京索莱宝科技有限公司提供)提取菌丝DNA, 将所得PCR原液进行测序.将本研究所测序列和参考序列用DNASTAR软件包的Megign程序进行排序, 用 Mega 4软件构建NJ树.结合菌株形态特征确定菌株分类地位(Yu et al., 2013). ...
... DSE广泛分布于各类生境, 尤其是广泛分布于胁迫环境和受干扰严重地区(Knapp et al., 2012; Postma et al., 2007; Zhao et al., 2015).目前, 全世界已发现110科600余种植物根组织有DSE定殖(Mandyam & Jumpponen, 2005), 虽然DSE能在体外纯培养, 但大多数纯培养条件下, 无法还原DSE在植物体内的真实生境而不产孢, 加之DSE本身涵盖众多科属, 其组成的复杂性和不明确性, 使得仅通过形态特征难以确定DSE的分类地位, 分子测序技术极大地推动了DSE的分类研究(Bonfim et al., 2016; Yadav et al., 2016).按照Sieber和Grünig (2013)对DSE的定义, 本研究分离的菌株均有DSE典型黑化结构, 不同样地的DSE菌株平均分离率为7.48%, 低于沙冬青DSE菌株10.0%的分离率(陈严严等, 2014). ...
荒漠北沙柳根系丛枝菌根真菌和黑隔内生真菌定殖状况 1 2014
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Phylogenetically diverse endozoic fungi in the South China Sea sponges and their potential in synthesizing bioactive natural products suggested by PKS gene and cytotoxic activity analysis. 1 2013
... 选用核糖体内转录间隔区(ITS)基因序列对不产孢菌株进行分子鉴定(Yadav et al., 2016).每个菌种选择2-5株菌株, 在PDA平板上27 ℃培养14天后, 刮取新鲜菌丝10 mg, 置于Eppendorf管中, 杵棒研磨充分后, 采用真菌基因组DNA提取试剂盒(北京索莱宝科技有限公司提供)提取菌丝DNA, 将所得PCR原液进行测序.将本研究所测序列和参考序列用DNASTAR软件包的Megign程序进行排序, 用 Mega 4软件构建NJ树.结合菌株形态特征确定菌株分类地位(Yu et al., 2013). ...
Tolerance and antioxidant response of a dark septate endophyte (DSE), Exophiala pisciphila, to cadmium stress. 1 2015
... 深色有隔内生真菌(DSE)泛指一群定居在植物根组织细胞内或细胞间隙的子囊菌和无性繁殖真菌, 其典型特征是细胞壁有黑色素沉淀, 能够形成深色有隔菌丝(Jumpponen, 2001; Grünig et al., 2008).已有研究表明, DSE具有广泛的宿主和生态分布特性, 广泛存在于沙漠(姜桥等, 2014)、干旱草原(闫姣等, 2014)和亚北极高山地区(Kauppinen et al., 2014)等不同生态环境中, 并在重金属矿区(Ban et al., 2012; Zhan et al., 2015)、盐胁迫地区(邓勋等, 2015)发挥着重要的生态修复作用.与丛枝菌根(AM)真菌仅能侵染宿主植物根皮层组织不同, DSE既可侵染宿主植物根皮层, 又能以薄壁菌丝侵染根维管组织(Muthukumar et al., 2006).Scervino等(2009)的研究表明, DSE能够改变植物菌根状况, 调节根际不同共生关系, 并在一定程度上补偿自然条件下AM真菌定殖率低或共生前期对宿主植物营养物质消耗的问题(Mandyam & Jumpponen, 2008).因此, 研究荒漠环境中DSE多样性和生态分布规律, 有助于充分理解土壤真菌共生体在荒漠生态系统中的功能和意义(Rudgers & Clay, 2007). ...
Diverse strategies conferring extreme cadmium (Cd) tolerance in the dark septate endophyte (DSE), Exophiala pisciphila: Evidence from RNA-seq data. 2 2015
... DSE广泛分布于各类生境, 尤其是广泛分布于胁迫环境和受干扰严重地区(Knapp et al., 2012; Postma et al., 2007; Zhao et al., 2015).目前, 全世界已发现110科600余种植物根组织有DSE定殖(Mandyam & Jumpponen, 2005), 虽然DSE能在体外纯培养, 但大多数纯培养条件下, 无法还原DSE在植物体内的真实生境而不产孢, 加之DSE本身涵盖众多科属, 其组成的复杂性和不明确性, 使得仅通过形态特征难以确定DSE的分类地位, 分子测序技术极大地推动了DSE的分类研究(Bonfim et al., 2016; Yadav et al., 2016).按照Sieber和Grünig (2013)对DSE的定义, 本研究分离的菌株均有DSE典型黑化结构, 不同样地的DSE菌株平均分离率为7.48%, 低于沙冬青DSE菌株10.0%的分离率(陈严严等, 2014). ...
 , 赵丽莉
, 赵丽莉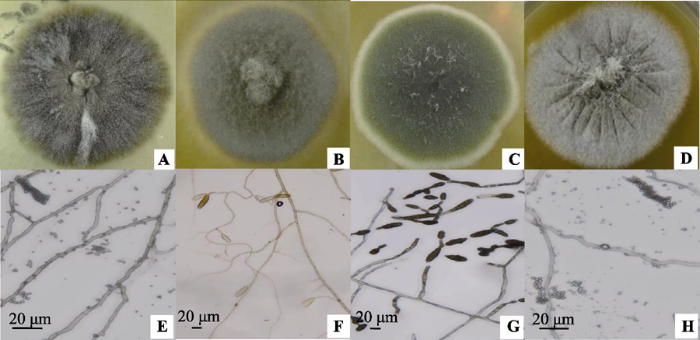 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT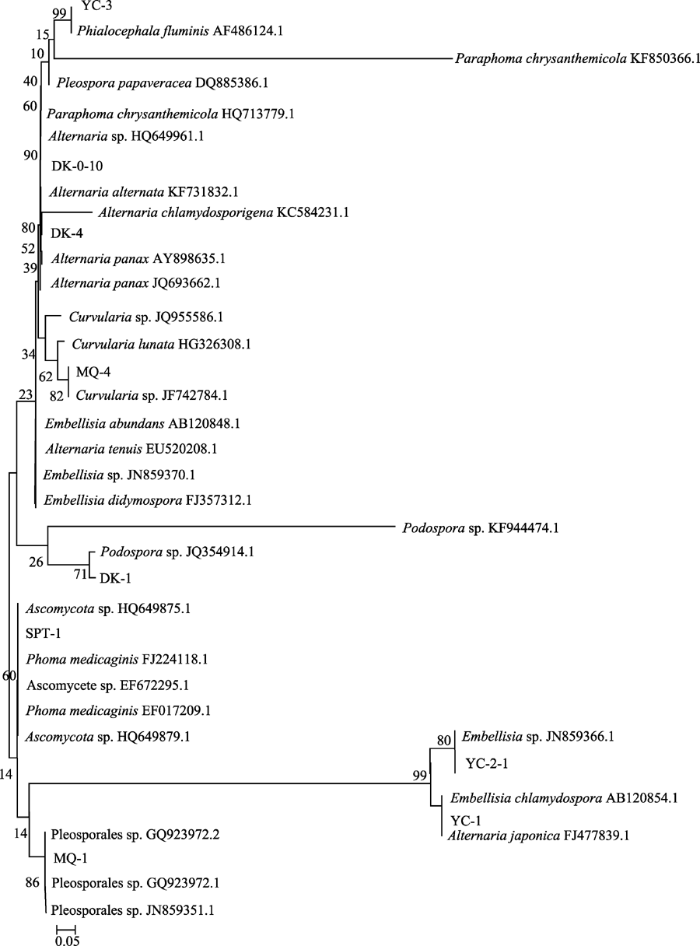 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}