关键词:被子植物;野外;分子标本;DNA;烘干;硅胶 Abstract Aims This study aims to identify a more convenient drying method for obtaining molecular specimen of angiosperms in the field than the conventional silica gel drying method. Methods The leaves of Prunus serrulata var. lannesiana and Liriope spicata were dried under temperatures of 150 °C, 80 °C, 40 °C as well as under natural conditions, and by the silica gel drying method, respectively. The DNA extracts of various specimens were then analyzed using techniques of spectrophotometer detection, electrophoresis and PCR to evaluate the impacts of different drying treatments to the genomic DNA of testing plants. Important findings The concentrations of total DNA were higher for the specimens dried at 40 °C treatment and by the silica gel drying method than other treatments when assessed by the techniques of spectrophotometer detection and electrophoresis. The concentration of PCR products was highest in the specimens dried at 40 °C. Based on the results, the 40 °C drying can be recommended for obtaining molecular specimens of angiosperms because of its minimum degree of degradation, for convenience of operation and avoiding carrying large amounts of silica gel in field investigations.
Keywords:angiosperms;field;molecular specimen;DNA;drying;silica gel -->0 PDF (2236KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 沈风娇, 任倩倩, 董琦, 朱丽, 张建芳, 杨婧, 张冉, 梁红柱, 赵建成, 石硕. 一种适合野外使用的被子植物分子标本干燥方式. 植物生态学报, 2017, 41(7): 787-794 https://doi.org/10.17521/cjpe.2016.0322 SHENFeng-Jiao, RENQian-Qian, DONGQi, ZHULi, ZHANGJian-Fang, YANGJing, ZHANGRan, LIANGHong-Zhu, ZHAOJian-Cheng, SHIShuo. A new angiosperms molecular specimen treatment method for field use. Chinese Journal of Plant Ecology, 2017, 41(7): 787-794 https://doi.org/10.17521/cjpe.2016.0322 提取高品质的DNA是植物材料用于分子生物学研究的先决条件, 对用于DNA提取的植物材料进行处理的理想方式是使用低温或液氮速冻处理, 但这一方法在野外不方便实施(Liston et al., 1990; 何天明等, 2004)。前人对野外如何处理分子标本(专门用于提取DNA的植物组织标本)这一问题进行了探索, 发现干燥处理后的分子标本可用于DNA提取(Doyle & Dickson, 1987), 而且分子标本经干燥处理后便于储存和携带, 此类处理方式特别适合野外使用。 ****们提出了不同干燥分子标本的处理方法, 如酒精法(Taggart et al., 1991)、硅胶干燥法(Chase & Hills, 1991)、硅藻土法、沙埋法(Cliquet & Jackson, 1997)等。研究发现, 分子标本干燥过程中DNA会发生不同程度的降解(Staats et al., 2011), 特别是标本的干燥方式严重影响分子标本中DNA的品质与总量(Särkinen et al., 2012), 而硅胶干燥法的效果较好, 优于上述其他分子标本干燥方法(Cliquet & Jackson, 1997; 蔡秀珍等, 2008)。野外采集标本时, 使用变色硅胶可将新鲜叶片较快干燥, 减少DNA降解, 使用也较为方便, 于是硅胶干燥法在处理分子标本时被广泛使用(Wang et al., 1996; Xie et al., 1998; Alexander et al., 2007; Xu et al., 2015)。虽然使用硅胶干燥法处理后的标本可以得到较高品质的DNA, 但是使用硅胶干燥法也存在许多不便: 1)硅胶密度较大, 野外远距离采样时不便随身大量携带; 2)硅胶吸水后更换不及时会导致标本干燥不彻底, 影响分子标本品质, 而及时更换硅胶会大大增加工作量; 3)使用后的硅胶再次使用前, 需对其进行干燥处理, 回收利用的硅胶容易使样品之间相互污染。因此, 在野外标本采集时使用硅胶干燥法处理分子标本具有较大的局限性。那么是否有更适于野外使用的分子标本干燥方式呢? 我们在长期的野外采集实践中, 发现烘干法是一个可用的选择。前人做过尝试性的研究, 发现烘干处理后的标本也可提取到较高品质的DNA (靖相密等, 2008)。在本研究中, 我们对包括硅胶干燥法在内的几种干燥方式进行研究, 比较不同干燥方式对分子标本的影响; 并结合在野外可供使用的设备设置实验的具体方案, 试图得到一种在野外采集过程中可以替代硅胶干燥法的更为便捷的被子植物分子标本干燥方法, 供相关人员在野外标本采集过程中参考使用。
2.1.1 分光光度计检测总DNA 用分光光度计检测总DNA品质, 如表1所示: 日本晚樱OD260/280值均在1.70左右, DNA浓度在200-500 ng·μL-1之间; 山麦冬的OD260/280值在1.90左右, DNA浓度在40-250 ng·μL-1之间。 Table 1 表1 表1日本晚樱和山麦冬分子标本经不同干燥处理后DNA纯度、浓度及PCR产物浓度的比较(平均值±标准偏差) Table 1Comparisons of DNA purity, concentration and the concentration of PCR products in the specimens of Prunus serrulata var. lannesiana and Liriope spicata obtained with different drying methods (mean ± SD)
干燥方式 Drying method
日本晚樱 Prunus serrulata var. lannesiana
山麦冬 Liriope spicata
OD260/280
DNA (ng·μL-1)
PCR (ng·μL-1)
OD260/280
DNA (ng·μL-1)
PCR (ng·μL-1)
150
1.83 ± 0.17a
451.24 ± 150.91a
13.94 ± 3.97c
2.03 ± 0.04a
211.43 ± 64.31a
0.74 ± 2.30d
80
1.80 ± 0.13a
376.13 ± 121.04a
34.48 ± 9.42b
1.83 ± 0.16b
80.64 ± 48.16c
16.51 ± 17.49c
40
1.74 ± 0.16ab
470.44 ± 228.50a
49.07 ± 4.83a
1.90 ± 0.08ab
241.41 ± 88.07a
57.64 ± 6.51a
Y
1.73 ± 0.09ab
291.64 ± 90.77b
38.38 ± 4.75b
1.93 ± 0.14ab
160.01 ± 22.95b
46.98 ± 8.22b
S
1.64 ± 0.10b
432.59 ± 167.67a
39.95 ± 5.82b
1.90 ± 0.24ab
252.33 ± 61.74a
43.19 ± 8.72b
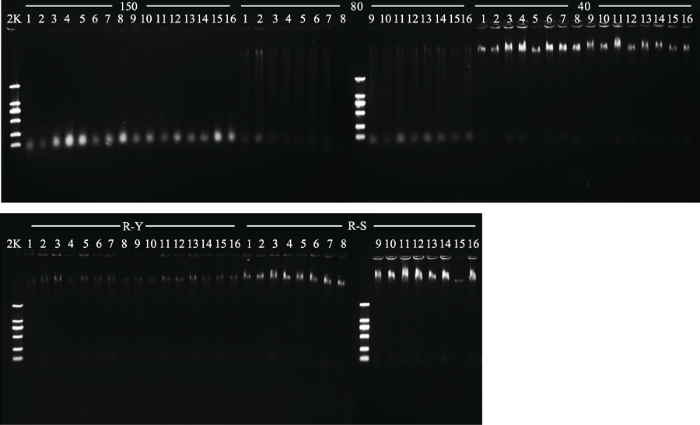
150, drying at 150 °C; 80, drying at 80 °C; 40, drying at 40 °C; Y, natural drying; S, silica gel drying. DNA, DNA concentration; PCR, concentration of PCR products. Values designated by different lowercase letters are significant different within the same columns (p < 0.05).150, 150 ℃烘干处理; 80, 80 ℃烘干处理; 40, 40 ℃烘干处理; Y, 吸水纸压制干燥; S, 硅胶干燥。DNA, DNA浓度; PCR, PCR产物浓度。同列不同小写字母表示差异显著(p < 0.05)。 新窗口打开 将5种不同干燥方式处理后所得到的数据进行统计学分析(表1), 比较日本晚樱及山麦冬的DNA浓度, 结果显示日本晚樱经吸水纸压制干燥后DNA浓度显著低于其余4种干燥方式(p < 0.05); 说明日本晚樱经吸水纸压制干燥后提取的DNA浓度较低, 经其他4种干燥方式处理后提取的DNA浓度较高。山麦冬经吸水纸压制干燥后DNA浓度显著低于硅胶干燥, 40和150 ℃烘干干燥, 差异显著(p < 0.05); 80 ℃烘干的DNA浓度显著低于吸水纸压制干燥, 说明山麦冬经硅胶干燥, 40和150 ℃烘干提取的DNA浓度较高, 吸水纸压制法提取的DNA浓度稍低, 80 ℃烘干提取的DNA浓度最低。 2.1.2 基因组DNA的电泳检测 从琼脂糖凝胶电泳检测结果(图1, 图2)可以看出, 经150和80 ℃烘干处理的样品DNA在胶孔内残留少, 说明这两种干燥方式提取的DNA较纯。 显示原图|下载原图ZIP|生成PPT 图15种不同干燥方式处理的日本晚樱(R)总DNA电泳图。150, 150 ℃烘干处理; 80, 80 ℃烘干处理; 40, 40 ℃烘干处理; Y, 吸水纸压制干燥; S, 硅胶干燥; 2K, DNA ladder中最长的DNA片段为2 000 bp。 -->Fig. 1Genomic DNA in specimens of Prunus serrulata var. lannesiana (R) obtained with five different drying methods. 150, drying at 150 °C; 80, drying at 80 °C; 40, drying at 40 °C; Y, natural drying; S, silica gel drying; 2K, 2 kb plus DNA ladder. -->
显示原图|下载原图ZIP|生成PPT 图25种不同干燥方式处理的山麦冬(M)总DNA电泳图。150, 150 ℃烘干处理; 80, 80 ℃烘干处理; 40, 40 ℃烘干处理; Y, 吸水纸压制干燥; S, 硅胶干燥; 2K, DNA ladder中最长的DNA片段为2 000 bp。 -->Fig. 2Genomic DNA in specimens of Liriope spicata (M) obtained with five different drying methods. 50, drying at 150 °C; 80, drying at 80 °C; 40, drying at 40 °C; Y, natural drying; S, silica gel drying; 2K, 2 kb plus DNA ladder. -->
5种不同干燥方式处理后DNA电泳检测均可检测到DNA的存在(图1、图2), 日本晚樱和山麦冬经40 ℃烘干、硅胶干燥后所得长片段条带(接近胶孔位置的条带, 胶孔在胶图上部)最亮; 吸水纸压制干燥法处理所得的长片段条带稍暗; 150 ℃烘干处理所得的长片段条带最暗。日本晚樱经80 ℃烘干后提取的长片段DNA条带较亮, 而山麦冬提取的长片段DNA较暗。这说明日本晚樱与山麦冬经40 ℃烘干与硅胶干燥处理所得的长片段DNA浓度最大, 吸水纸压制干燥法处理所得的长片段DNA浓度稍低, 经150 ℃烘干所得的长片段DNA浓度最低, 降解严重。日本晚樱经80 ℃烘干所得的长片段DNA浓度较大, 而山麦冬经80 ℃烘干所得的长片段DNA浓度较低。 显示原图|下载原图ZIP|生成PPT 图3日本晚樱(R)经5种不同干燥方式处理后PCR凝胶电泳检测结果。150, 150 ℃烘干处理; 80, 80 ℃烘干处理; 40, 40 ℃烘干处理; Y, 吸水纸压制干燥; S, 硅胶干燥; 2K, DNA ladder中最长的DNA片段为2 000 bp。 -->Fig. 3Agarose gel electrophoresis of PCR products in specimens of Prunus serrulata var. lannesiana (R) obtained with five different drying methods. 150, drying at 150 °C; 80, drying at 80 °C; 40, drying at 40 °C; Y, natural drying; S, silica gel drying; 2K, 2kb plus DNA ladder. -->
2.2 PCR体外扩增电泳检测
使用Image J软件将PCR扩增结果定量分析, 再将PCR产物浓度进行统计分析(表1), 可以看出, 日本晚樱40 ℃烘干处理后PCR产物的浓度显著高于硅胶干燥、吸水纸压制干燥、80 ℃烘干(p < 0.05); 硅胶干燥、吸水纸压制干燥、80 ℃烘干3种干燥方式处理后体外扩增的浓度无显著性差异(p > 0.05); 150 ℃烘干处理后PCR产物浓度显著低于硅胶干燥、吸水纸压制干燥、80 ℃烘干(p < 0.05)。山麦冬40 ℃烘干处理后PCR产物浓度显著高于吸水纸压制干燥、硅胶干燥(p < 0.05); 吸水纸压制干燥、硅胶干燥两种干燥方式处理后PCR产物浓度无显著性差异(p > 0.05); 80 ℃烘干处理后PCR产物浓度显著低于吸水纸压制干燥、硅胶干燥两种干燥方式; 150 ℃烘干处理后PCR产物浓度显著低于80 ℃烘干(p < 0.05)。这说明日本晚樱和山麦冬经40 ℃烘干处理后PCR产物浓度最高, 硅胶干燥和吸水纸压制干燥法处理后PCR产物浓度稍低, 150 ℃烘干处理PCR产物浓度最低。日本晚樱经80 ℃烘干处理后PCR产物浓度与硅胶干燥、吸水纸压制干燥浓度无显著区别, 而山麦冬经80 ℃烘干处理后PCR产物浓度与硅胶干燥、吸水纸压制干燥相比浓度较低。 显示原图|下载原图ZIP|生成PPT 图4山麦冬(M)经5种不同干燥方式处理后PCR凝胶电泳检测结果。150, 150 ℃烘干处理; 80, 80 ℃烘干处理; 40, 40 ℃烘干处理; Y, 吸水纸压制干燥; S, 硅胶干燥; 2K, DNA ladder中最长的DNA片段为2 000 bp。 -->Fig. 4Agarose gel electrophoresis of PCR products in specimens of Liriope spicata (M) obtained with five different drying methods. 150, drying at 150 °C; 80, drying at 80 °C; 40, drying at 40 °C; Y, natural drying; S, silica gel drying; 2K, 2 kb plus DNA ladder. -->
AlexanderPJ, RajanikanthG, BaconCD, DonovanbaileyC (2007). Recovery of plant DNA using a reciprocating saw and silica-based columns. Molecular Ecology Notes, 7, 5-9. [本文引用: 1]
[2]
CaiXZ, LiuKM, LongCL (2008). DNA extraction from dried leaves and PCR amplification of Colocasia. Chinese Wild Plant Resources, 27(1), 51-57. (in Chinese with English abstract)[蔡秀珍, 刘克明, 龙春林 (2008). 野生芋属植物干叶片DNA的提取及PCR扩增 . 中国野生植物资源, 27(1), 51-57.] [本文引用: 1]
[3]
ChaseMW, HillsHH (1991). Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon, 40, 215-220. [本文引用: 1]
[4]
CliquetS, JacksonMA (1997). Comparison of air-drying methods for evaluating the desiccation tolerance of liquid culture-produced blastospores of Paecilomyces fumosoroseus. World Journal of Microbiology and Biotechnology, 13, 299-303. [本文引用: 2]
[5]
DoyleJJ, DicksonEE (1987). Preservation of plant samples for DNA restriction endonuclease analysis. Taxon, 36, 715-722. [本文引用: 1]
[6]
HeTM, ChenXS, WuY (2004). Preparation of DNA from silica gel dried leaves of Rosaceae. Journal of Shihezi University ( Natural Science), 22, 316-319. (in Chinese with English abstract)[何天明, 陈学森, 吴燕 (2004). 从蔷薇科果树硅胶干燥叶片中制备DNA . 石河子大学学报(自然科学版), 22, 316-319.] [本文引用: 1]
[7]
Heenan PB, GoekeDF, HoulistonGJ, LysakMA (2012). Phylogenetic analyses of ITS and rbcL DNA sequences for sixteen genera of Australian and New Zealand Brassicaceae result in the expansion of the tribe Microlepidieae. Taxon, 61, 970-979. [本文引用: 1]
[8]
JingXM, ChuYX, TangGG, ZhangYC, LiuZ, ZhangH (2008). Effects of different sample preserving methods on genomic DNA extraction of Chimonanthus praecox and their ISSR-PCR validation. Molecular Plant Breeding, 6, 387-392. (in Chinese with English abstract)[靖相密, 褚云霞, 汤庚国, 张永春, 刘忠, 张慧 (2008). 不同保存方法对蜡梅总DNA提取效果的影响及ISSR-PCR验证 . 分子植物育种, 6, 387-392.] [本文引用: 1]
[9]
LiJL, WangS, YuJ, WangL, ZhouSL (2013). A modified CTAB protocol for plant DNA extraction. Chinese Bulletin of Botany, 48, 72-78. (in Chinese with English abstract)[李金璐, 王硕, 于婧, 王玲, 周世良 (2013). 一种改良的植物DNA提取方法 . 植物学报, 48, 72-78.] [本文引用: 1]
[10]
ListonA, RiesebergLH, AdamsRP, DoN, ZhuGL (1990). A method for collecting dried plant specimens for DNA and isozyme analyses, and the results of a field test in Xinjiang, China. Annals of the Missouri Botanical Garden, 77, 859-863. [本文引用: 1]
[11]
QiuNW, LiuQ, LiuH (2015). Multiple comparison data in statistical analysis. Journal of Biomathematics, 30, 535-541. (in Chinese with English abstract)[邱念伟, 刘倩, 刘慧 (2015). 生物学实验数据统计分析中的多重比较法 . 生物数学学报, 30, 535-541.] [本文引用: 1]
[12]
SärkinenT, StaatsM, RichardsonJE, CowanRS, BakkerFT (2012). How to open the treasure chest? Optimising DNA extraction from herbarium specimens. PLOS ONE, 7, e43808. doi: 10.1371/journal.pone.0043808. [本文引用: 1]
TaggartJB, HynesRA, ProdöuhlPA, FergusonA (1991). A simplified protocol for routine total DNA isolation from salmonid fishes. Journal of Fish Biology, 40, 963-965. [本文引用: 1]
[15]
WangXD, WangZP, ZouYP (1996). An improved procedure for the isolation of nuclear DNA from leaves of wild grapevine dried with silica gel. Plant Molecular Biology Reporter, 14, 369-373. [本文引用: 1]
[16]
WeiCX, XiePS, ZhouWD, ChenYF, ZhangJ, HuaiHY (2008). Observation on morphological structure of leaf epidermis of Ophiopogon japonicus, Liriope spicata and L. platyphylla. Journal of Plant Resources and Environment, 17(4), 9-15. (in Chinese with English abstract)[韦存虚, 谢佩松, 周卫东, 陈义芳, 张军, 淮虎银 (2008). 麦冬、土麦冬和阔叶土麦冬叶表皮形态结构的观察 . 植物资源与环境学报, 17(4), 9-15.] [本文引用: 1]
[17]
XieZ, GeS, HongD (1998). Preparation of DNA from silica gel dried mini-amount of leaves of Oryza rufipogon for RAPD study and total DNA bank construction. Acta Botanica Sinica, 41, 807-812. [本文引用: 1]
[18]
XuC, DongW, ShiS, ChengT, LiCH, LiuYL, WuP, WuHK, GaoP (2015). Accelerating plant DNA barcode reference library construction using herbarium specimens: Improved experimental techniques. Molecular Ecology Resources, 15, 1366-1374. [本文引用: 1]
Recovery of plant DNA using a reciprocating saw and silica-based columns. 1 2007
... ****们提出了不同干燥分子标本的处理方法, 如酒精法(Taggart et al., 1991)、硅胶干燥法(Chase & Hills, 1991)、硅藻土法、沙埋法(Cliquet & Jackson, 1997)等.研究发现, 分子标本干燥过程中DNA会发生不同程度的降解(Staats et al., 2011), 特别是标本的干燥方式严重影响分子标本中DNA的品质与总量(Särkinen et al., 2012), 而硅胶干燥法的效果较好, 优于上述其他分子标本干燥方法(Cliquet & Jackson, 1997; 蔡秀珍等, 2008).野外采集标本时, 使用变色硅胶可将新鲜叶片较快干燥, 减少DNA降解, 使用也较为方便, 于是硅胶干燥法在处理分子标本时被广泛使用(Wang et al., 1996; Xie et al., 1998; Alexander et al., 2007; Xu et al., 2015).虽然使用硅胶干燥法处理后的标本可以得到较高品质的DNA, 但是使用硅胶干燥法也存在许多不便: 1)硅胶密度较大, 野外远距离采样时不便随身大量携带; 2)硅胶吸水后更换不及时会导致标本干燥不彻底, 影响分子标本品质, 而及时更换硅胶会大大增加工作量; 3)使用后的硅胶再次使用前, 需对其进行干燥处理, 回收利用的硅胶容易使样品之间相互污染.因此, 在野外标本采集时使用硅胶干燥法处理分子标本具有较大的局限性.那么是否有更适于野外使用的分子标本干燥方式呢? ...
野生芋属植物干叶片DNA的提取及PCR扩增 1 2008
... ****们提出了不同干燥分子标本的处理方法, 如酒精法(Taggart et al., 1991)、硅胶干燥法(Chase & Hills, 1991)、硅藻土法、沙埋法(Cliquet & Jackson, 1997)等.研究发现, 分子标本干燥过程中DNA会发生不同程度的降解(Staats et al., 2011), 特别是标本的干燥方式严重影响分子标本中DNA的品质与总量(Särkinen et al., 2012), 而硅胶干燥法的效果较好, 优于上述其他分子标本干燥方法(Cliquet & Jackson, 1997; 蔡秀珍等, 2008).野外采集标本时, 使用变色硅胶可将新鲜叶片较快干燥, 减少DNA降解, 使用也较为方便, 于是硅胶干燥法在处理分子标本时被广泛使用(Wang et al., 1996; Xie et al., 1998; Alexander et al., 2007; Xu et al., 2015).虽然使用硅胶干燥法处理后的标本可以得到较高品质的DNA, 但是使用硅胶干燥法也存在许多不便: 1)硅胶密度较大, 野外远距离采样时不便随身大量携带; 2)硅胶吸水后更换不及时会导致标本干燥不彻底, 影响分子标本品质, 而及时更换硅胶会大大增加工作量; 3)使用后的硅胶再次使用前, 需对其进行干燥处理, 回收利用的硅胶容易使样品之间相互污染.因此, 在野外标本采集时使用硅胶干燥法处理分子标本具有较大的局限性.那么是否有更适于野外使用的分子标本干燥方式呢? ...
Silica gel: An ideal material for field preservation of leaf samples for DNA studies. 1 1991
... ****们提出了不同干燥分子标本的处理方法, 如酒精法(Taggart et al., 1991)、硅胶干燥法(Chase & Hills, 1991)、硅藻土法、沙埋法(Cliquet & Jackson, 1997)等.研究发现, 分子标本干燥过程中DNA会发生不同程度的降解(Staats et al., 2011), 特别是标本的干燥方式严重影响分子标本中DNA的品质与总量(Särkinen et al., 2012), 而硅胶干燥法的效果较好, 优于上述其他分子标本干燥方法(Cliquet & Jackson, 1997; 蔡秀珍等, 2008).野外采集标本时, 使用变色硅胶可将新鲜叶片较快干燥, 减少DNA降解, 使用也较为方便, 于是硅胶干燥法在处理分子标本时被广泛使用(Wang et al., 1996; Xie et al., 1998; Alexander et al., 2007; Xu et al., 2015).虽然使用硅胶干燥法处理后的标本可以得到较高品质的DNA, 但是使用硅胶干燥法也存在许多不便: 1)硅胶密度较大, 野外远距离采样时不便随身大量携带; 2)硅胶吸水后更换不及时会导致标本干燥不彻底, 影响分子标本品质, 而及时更换硅胶会大大增加工作量; 3)使用后的硅胶再次使用前, 需对其进行干燥处理, 回收利用的硅胶容易使样品之间相互污染.因此, 在野外标本采集时使用硅胶干燥法处理分子标本具有较大的局限性.那么是否有更适于野外使用的分子标本干燥方式呢? ...
Comparison of air-drying methods for evaluating the desiccation tolerance of liquid culture-produced blastospores of Paecilomyces fumosoroseus. 2 1997
... ****们提出了不同干燥分子标本的处理方法, 如酒精法(Taggart et al., 1991)、硅胶干燥法(Chase & Hills, 1991)、硅藻土法、沙埋法(Cliquet & Jackson, 1997)等.研究发现, 分子标本干燥过程中DNA会发生不同程度的降解(Staats et al., 2011), 特别是标本的干燥方式严重影响分子标本中DNA的品质与总量(Särkinen et al., 2012), 而硅胶干燥法的效果较好, 优于上述其他分子标本干燥方法(Cliquet & Jackson, 1997; 蔡秀珍等, 2008).野外采集标本时, 使用变色硅胶可将新鲜叶片较快干燥, 减少DNA降解, 使用也较为方便, 于是硅胶干燥法在处理分子标本时被广泛使用(Wang et al., 1996; Xie et al., 1998; Alexander et al., 2007; Xu et al., 2015).虽然使用硅胶干燥法处理后的标本可以得到较高品质的DNA, 但是使用硅胶干燥法也存在许多不便: 1)硅胶密度较大, 野外远距离采样时不便随身大量携带; 2)硅胶吸水后更换不及时会导致标本干燥不彻底, 影响分子标本品质, 而及时更换硅胶会大大增加工作量; 3)使用后的硅胶再次使用前, 需对其进行干燥处理, 回收利用的硅胶容易使样品之间相互污染.因此, 在野外标本采集时使用硅胶干燥法处理分子标本具有较大的局限性.那么是否有更适于野外使用的分子标本干燥方式呢? ...
... ), 而硅胶干燥法的效果较好, 优于上述其他分子标本干燥方法(Cliquet & Jackson, 1997; 蔡秀珍等, 2008).野外采集标本时, 使用变色硅胶可将新鲜叶片较快干燥, 减少DNA降解, 使用也较为方便, 于是硅胶干燥法在处理分子标本时被广泛使用(Wang et al., 1996; Xie et al., 1998; Alexander et al., 2007; Xu et al., 2015).虽然使用硅胶干燥法处理后的标本可以得到较高品质的DNA, 但是使用硅胶干燥法也存在许多不便: 1)硅胶密度较大, 野外远距离采样时不便随身大量携带; 2)硅胶吸水后更换不及时会导致标本干燥不彻底, 影响分子标本品质, 而及时更换硅胶会大大增加工作量; 3)使用后的硅胶再次使用前, 需对其进行干燥处理, 回收利用的硅胶容易使样品之间相互污染.因此, 在野外标本采集时使用硅胶干燥法处理分子标本具有较大的局限性.那么是否有更适于野外使用的分子标本干燥方式呢? ...
Preservation of plant samples for DNA restriction endonuclease analysis. 1 1987
Phylogenetic analyses of ITS and rbcL DNA sequences for sixteen genera of Australian and New Zealand Brassicaceae result in the expansion of the tribe Microlepidieae. 1 2012

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT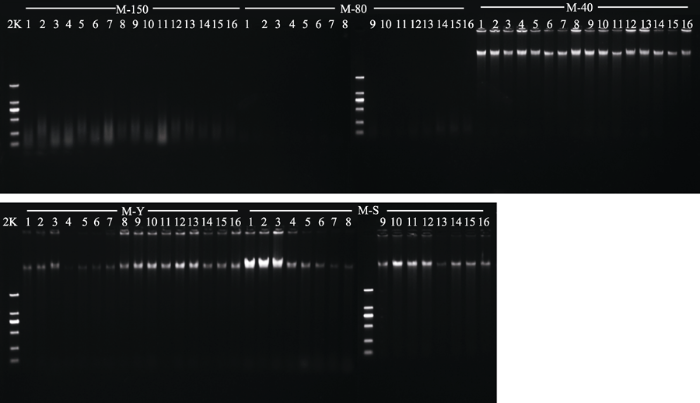 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT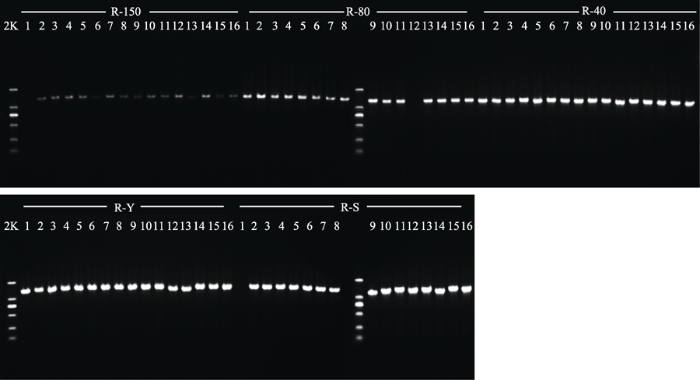 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}