Differential uptakes of different forms of soil nitrogen among major tree species in subalpine coniferous forests of western Sichuan, China
ZOUTing-Ting1,2, ZHANGZi-Liang1,2, LINa1, YUANYuan-Shuang1,2, ZHENGDong-Hui1,2, LIUQin1, YINHua-Jun1,3,*, 1Chengdu Institute of Biology, Chinese Academy of Sciences, Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization of Chinese Academy of Sciences, and Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu 610041, China2University of Chinese Academy of Sciences, Beijing 100049, China3College of Environmental Science and Engineering, China West Normal University, Nanchong, Sichuan 637000, China 通讯作者:* 通信作者Author for correspondence (E-mail: yinhj@cib.ac.cn) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家重点研发计划项目(2017YFC0505200)、中国科学院前沿科学重点研究项目(QYZDB-SSW-SMC023)、国家自然科学基金(31670449)、四川省重点研发项目(2017SZ0038)和四川省青年科技基金(2016JQ0037)
关键词:有机氮;无机氮;氨基酸;稳定同位素标记;亚高山针叶林 Abstract Aims Although acquisition of soil organic nitrogen (N)(mainly amino acids) by plants is a widespread ecological phenomenon in many terrestrial ecosystems, the rate of organic N uptake and their contributions to plant nutrient supply are poorly understood. Our objective was to determine the relative contributions of inorganic N (NO3–-N and NH4+-N) and organic N (amino acids) to plant N uptake in a high-frigid forest ecosystem.Methods The differences in the uptake rate of three different forms of N (NO3–-N, NH4+-N and glycine) were quantified by exposing seedlings of two dominant tree species (Picea asperata and Betula albo-sinensis) in subalpine coniferous forests of western Sichuan, China, to trace quantities of K15NO3,15NH4Cl and (U-13C2/15N) glycine.Important findings Both 13C and 15N were significantly enriched in fine roots 2 h after tracer application, indicating the occurrence of glycine uptake in P. asperata and B. albo-sinensis seedlings. The seedlings of two tree species had a significant preference for NO3–-N compared with glycine and NH4+-N, and the uptake rate of NO3–-N was 5 to 10 times greater than that of glycine and NH4+-N. The roots of seedlings in the two species took up glycine more rapidly than NH4+-N, implying that soil organic N (i.e., amino acids) could be an important N source for the two species in subalpine coniferous forests. The results of this study are of great theoretical significance for understanding N utilization strategies and nutrient regulation processes in plants of the high-frigid forest ecosystems.
Keywords:organic nitrogen;inorganic nitrogen;amino acid;stable isotope labeling;subalpine coniferous forest -->0 PDF (531KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 邹婷婷, 张子良, 李娜, 袁远爽, 郑东辉, 刘庆, 尹华军. 川西亚高山针叶林主要树种对土壤中不同形态氮素的吸收差异. 植物生态学报, 2017, 41(10): 1051-1059 https://doi.org/10.17521/cjpe.2017.0165 ZOUTing-Ting, ZHANGZi-Liang, LINa, YUANYuan-Shuang, ZHENGDong-Hui, LIUQin, YINHua-Jun. Differential uptakes of different forms of soil nitrogen among major tree species in subalpine coniferous forests of western Sichuan, China. Chinese Journal of Plant Ecology, 2017, 41(10): 1051-1059 https://doi.org/10.17521/cjpe.2017.0165 氮(N)是植物需求量最大的一种矿质营养元素, 是植物体内核酸、蛋白质、酶等物质的主要组成成分, 同时也是陆地生态系统植物生长发育的主要限制因子(Jones et al., 2005), 对陆地生态系统净初级生产力和功能具有极其重要的作用(Elser et al., 2007)。土壤中的N有多种赋存形态和性质, 主要可分为无机N (以NH4+-N和NO3–-N为主)和有机N (如氨基酸、酰胺、尿素等)两大类。土壤无机N是土壤N的主要组成部分, 占土壤总N的90%以上(李合生, 2002)。传统的矿质营养学说通常认为植物只能吸收无机N (如NH4+-N和NO3–-N), 有机N必须被分解为无机N后才能被植物吸收。然而, 随着同位素技术的广泛运用和不断发展, 越来越多的研究表明: 许多植物(包括菌根植物和一些无菌根植物)能够直接吸收利用土壤中的多种氨基酸、简单蛋白质等小分子和大分子有机N化合物(Bennett & Prescott, 2004; Miller et al., 2007; Rothstein, 2014)。在一些常年低温、N矿化受到强烈限制的地区, 植物对有机N的吸收速率甚至超过无机N, 此现象引发了人们对有机N在陆地生态系统营养流动与N循环模型的重新思考和评价(Gallet-Budynek et al., 2009)。 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009)。比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001)。然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006)。造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014)。因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制。 川西亚高山针叶林是青藏高原东部高寒林区的重要组成部分, 其表现为低温、短生长季、高有机质、高有机N、低矿化速率和低养分有效性等一系列特征(尹华军等, 2008; Zhang et al., 2017), 这使得亚高山森林成为研究生态系统N养分来源与N素吸收利用的典型类型和重要区域。因此, 本研究选取川西亚高山针叶林两个代表性树种——云杉(Picea asperata)幼苗和红桦(Betula albo-sinensis)幼苗为研究对象, 采用稳定同位素标记法, 研究两个树种对无机N (NH4+-N和NO3–-N)和有机N (氨基酸)的吸收速率与差异, 旨在: (1)弄清川西亚高山针叶林生态系统植物能否直接吸收利用有机N作为植物N源? (2)在此基础上, 进一步比较该区植物对土壤中3种N素 (NH4+-N、NO3–-N和氨基酸)的吸收差异, 以揭示不同N源对该区植物N养分吸收的相对贡献。本研究结果可为更好地认识川西亚高山针叶林土壤N循环与养分获取策略提供一定的理论基础。
1 材料和方法
1.1 研究区概况
本研究地位于四川省理县境内杂谷脑镇胆扎木沟(31.41° N, 103.21° E, 海拔约2 300 m), 该区位于青藏高原东缘, 四川省西北部, 阿坝藏族羌族自治州东南缘, 地质结构属于龙门山断裂带中段, 山峦起伏, 地形复杂。该区气候类型属于山地季风气候, 夏季温凉多雨, 冬季寒冷干燥, 1月平均气温-8 ℃, 7月平均气温12.6 ℃, 年平均气温4.8-6.9 ℃, 无霜期115-150天, >10 ℃年有效积温1 400-2 500 ℃, 年降水量700-900 mm, 年干燥度0.59-1.18。生长季长度大概为6个月, 从4月下旬到10月下旬。冬季大概有4个月为冰雪覆盖期, 最大雪被厚度约为30 cm。土壤类型主要为山地棕壤(Nan et al., 2013)。
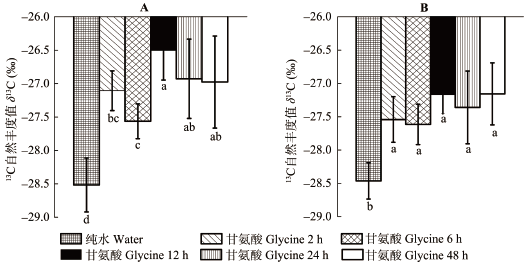
(U-13C2/15N)甘氨酸标记2 h后, 云杉和红桦幼苗细根δ13C表现出明显的富集现象, 在各取样时间点均显著高于对照(p < 0.05), 但两树种表现出不同的时间动态变化规律。(U-13C2/15N)甘氨酸标记12 h后, 云杉幼苗细根δ13C达到最大值, 并显著高于对照、2 h和6 h时细根中δ13C (p < 0.05), 而红桦幼苗细根δ13C在各取样时间点无显著差别(图2)。 与对照处理相比, 添加3种15N标记的N化合物后, 两树种幼苗细根表现出明显的15N富集现象, 各取样时间点δ15N值均显著高于对照(p < 0.05)。云杉幼苗细根中15NO3–-N、(U-13C2/15N)甘氨酸和 15NH4+-N 3种处理下δ15N值随时间增加, 总体表现出逐渐升高的趋势, 分别在处理48 h、24 h和48 h时达到最大值。类似地, 红桦幼苗细根中δ15N值在 15NO3–-N、(U-13C2/15N)甘氨酸和15NH4+-N 3种处理下也随时间增加逐渐升高, 分别在处理48 h、24 h和 12 h时达到最大值(图3)。 显示原图|下载原图ZIP|生成PPT 图2不同采样时间云杉(A)和红桦(B)幼苗细根中13C自然丰度值(δ13C)的动态变化(平均值±标准偏差, n = 6)。柱状图中不同小写字母表示同一树种δ13C在不同取样时间差异显著(p < 0.05)。 -->Fig. 2Dynamics of natural 13C abundance (δ13C) in fine roots of Picea asperata (A) and Betula albo-sinensis seedlings (B) at different sampling times (mean ± SD, n = 6). Different lowercase letters indicate significant differences in the δ13C value among sampling times within species (p < 0.05). --> 显示原图|下载原图ZIP|生成PPT 图3不同采样时间云杉和红桦幼苗细根中15N自然丰度值(δ15N)动态变化(平均值±标准偏差, n = 6)。A, 15NO3–-N处理下的δ15N。B, (U-13C2/15N)甘氨酸(Glycine)处理下的δ15N。C, 15NH4+-N处理下的δ15N。柱状图中不同小写字母表示同一树种δ15N在不同取样时间差异显著(p < 0.05)。 -->Fig. 3Dynamics of natural 15N abundance (δ15N) in fine roots of Picea asperata and Betula albo-sinensis seedlings at different sampling times (mean ± SD, n = 6). A, δ15N under 15NO3–-N labeling treatment. B, δ15N under glycine labeling treatment. C, δ15N for 15NH4+-N labeling treatment. Different lowercase letters indicate significant differences in the δ15N value among sampling times within species (p < 0.05). -->
2.2 云杉和红桦幼苗对3种N素吸收速率的差异
云杉幼苗对NO3–-N的吸收速率显著高于对甘氨酸和NH4+-N的吸收速率(p < 0.05)(图4)。云杉幼苗对NO3–-N、甘氨酸和NH4+-N的平均吸收速率分别为0.447、0.055和0.035 μg·g-1·h-1, 云杉幼苗对NO3–-N的平均吸收速率分别是甘氨酸和NH4+-N平均吸收速率的8.08和12.77倍; 类似地, 红桦幼苗对NO3–-N的吸收速率显著高于对甘氨酸和NH4+-N的吸收速率(p < 0.05)。红桦幼苗对NO3–-N的平均吸收速率(0.485 μg·g-1·h-1)分别是对甘氨酸(0.085 μg·g-1·h-1)和NH4+-N (0.070 μg·g-1·h-1)平均吸收速率的5.74和6.95倍。 显示原图|下载原图ZIP|生成PPT 图4不同采样时间点云杉(A)和红桦(B)幼苗对3种不同形态N素(NO3–-N、甘氨酸和NH4+-N)的吸收速率差异(平均值±标准偏差, n = 6)。柱状图中不同小写字母表示同一时间不同形态N吸收速率差异显著(p < 0.05)。 -->Fig. 4Differences in the mass-specific uptake rate for NO3–-N, glycine and NH4+-N in Picea asperata (A) and Betula albo-sinensis seedlings (B) at different sampling times (mean ± SD, n = 6). Different lowercase letters indicate significant differences in the uptake rate of different N forms at given sampling time (p < 0.05). -->
N是森林生态系统植物生产力和群落稳定性的最主要限制因素(Burton et al., 2007), 有关植物对土壤N的吸收和利用策略研究一直备受关注。目前, 人们对植物N吸收和利用的认识主要依赖于无菌栽培技术、15N示踪技术和13C/14C示踪技术的应用。在本研究中, 我们主要采用13C和15N双标记氨基酸技术来原位示踪植物对有机N的吸收, 通过此方法, 若能在植物组织中同时检测到13C和15N的富集, 且表明两者有相关性, 说明氨基酸以完整的形态被直接吸收(N?sholm et al., 1998)。通过研究发现, 在添加双标记物2 h后, 云杉和红桦幼苗根系中13C和15N均表现出明显的富集现象(图2, 图3), 强有力地证明了两种植物均能够直接吸收土壤中的氨基酸。此外, 该方法有效应用的另外一个重要前提是加入同位素N后不会对土壤产生施肥效应。在试验前我们基于样地3种N源含量, 所添加的3种同位素N含量均未超过土壤中各形态N背景浓度的10%。同时在试验结束时, 通过土壤3种N素分析也表明, 两种幼苗样地在标记试验后土壤中的各种形态N含量与试验前均无显著差异。因此, 该方法的有效性在本研究中得到有效保证, 即本研究中两种幼苗根系对3种同位素标记物(15NH4+-N、15NO3–-N和(U-13C2/15N)甘氨酸)吸收速率的差异能真实地反映其根系对土壤中不同形态N的吸收偏好。 显示原图|下载原图ZIP|生成PPT 图5云杉(A)和红桦(B)幼苗根系中13C和15N的单位摩尔原子百分超比值的差异(平均值±标准偏差, 6个重复)。虚线代表注射的双标记甘氨酸C:N原始值为2.0。 -->Fig. 5Differences in the molar ratio of excess 13C to excess 15N derived from double-labelled (U-13C2/15N) glycine in fine roots of Picea asperata (A) and Betula albo-sinensis seedlings (B). Horizontal line represents the 2:1 injection ratio of C:N administered with the doubly labeled glycine treatment. -->
3.2 亚高山针叶林树种对不同状态N吸收的偏好 分析
一般认为, 植物吸收NH4+-N的过程比吸收NO3–-N的过程节省更多能量(Wang & Macko, 2011), 土壤NH4+-N进入植物根系后直接被转化为谷氨酸盐从而被植物进一步利用, 而植物在吸收NO3–-N的过程中必须通过消耗较多能量的硝酸盐还原途径才能被植物吸收利用(Templer & Dawson, 2004)。因此, 相对于NO3–-N, 植物通常更偏好吸收土壤中的NH4+-N (Finzi & Berthrong, 2005; N?sholm et al., 2009)。然而, 本研究结果表明: 川西亚高山针叶林树种云杉和红桦更偏好吸收土壤NO3–-N。具体而言, 与氨基酸和NH4+-N处理相比, NO3–-N处理的植物细根表现出显著更高的δ15N值和更快的吸收速率(图2, 图3)。此外, 云杉和红桦幼苗根系对NO3–-N的平均吸收速率为NH4+-N和氨基酸平均吸收速率的5-10倍(图4)。该研究结果与大部分前人研究认为的植物偏好吸收和利用土壤NH4+-N的观点不一致。我们推测一个重要的原因可能与供试土壤有较高的NO3–-N含量有关。前期研究发现, 植物对土壤N的吸收具有可塑性, 当土壤中各种N的有效性发生变化时, 植物会调整自身对N的吸收偏好来维持甚至增加整体N的吸收(Ashton et al., 2010; Song et al., 2015)。Breteler和Siegerist (1984)的研究表明土壤中高浓度的NH4+-N抑制植物根系对NO3–-N的吸收, 而当NH4+- N浓度低于1 mmol·L-1时, 这种抑制作用将降至最小, 植物根系不再排斥NO3–-N, 可将其作为N源。在本研究中, 土壤NH4+-N浓度仅有-0.75 mmol·L-1, 因此其对植物吸收NO3–-N的抑制作用可忽略不计。Nordin等(2001)在研究北方森林植物N吸收过程中也发现植物对土壤中的不同形态N的偏好吸收与这些N素在土壤中的有效性呈正相关关系。因此, 我们认为本研究中植物对NO3–-N表现出显著更高的吸收速率(图3), 可能在很大程度上归因于土壤中相对较高的NO3–-N浓度。NO3–-N作为川西亚高山针叶林土壤中最主要的N形态(Xu et al., 2010), 其被云杉和红桦幼苗偏好吸收也是亚高山森林植物长期适应该区域土壤环境的结果。另外, NO3–-N在土壤中具有更高的可移动性也可导致植物对NO3–-N表现出明显的偏好吸收现象(Warren, 2009; Li et al., 2015)。比如, NO3–-N在土壤溶液中的扩散系数比NH4+-N高10-100倍(Gessler et al., 1998), 因此前者能够通过土壤溶液更快地扩散到根系从而为植物提供更大的吸收几率。尽管植物对NH4+-N的吸收消耗更少的能量, 但许多植物对NH4+-N具有较高的敏感性(Chen et al., 2013)。与NO3–-N相比, NH4+-N在植物体积累易产生毒性效应, 抑制K+、Ca2+、Mg2+等阳离子的吸收, 从而不利于植物的生长(Britto & Kronzucker, 2002; Coskun et al., 2013), 这可能也是植物偏好吸收NO3–-N的原因之一。然而, 关于不同生态系统、不同树种对NO3–-N和NH4+-N的偏好吸收与利用的影响因素与调控机制并不清楚, 需要进一步开展深入的研究。
AshtonIW, MillerAE, BowmanWD, SudingKN (2010). Niche complementarity due to plasticity in resource use: Plant partitioning of chemical N forms. Ecology, 91, 3252-3260.URL [本文引用: 1]
[2]
BennettJN, PrescottCE (2004). Organic and inorganic nitrogen nutrition of western red cedar, western hemlock and salal in mineral N-limited cedar-hemlock forests. Oecologia, 141, 468-476.https://doi.org/10.1007/s00442-004-1622-3URLPMID:15322895 [本文引用: 1]摘要 Western red cedar (Thuja plicata Donn.), western hemlock (Tsuga heterophylla Raf. Sarge) and salal (Gaultheria shallon Pursh) are the main species growing in cedar-hemlock forests on Vancouver Island, Canada. Based on the dominance of organic N in these systems, we tested the hypotheses that: (1) organic N can be utilized by the three plant species; and (2) salal, which is ericoid mycorrhizal and has high tannin concentration in its tissues, would absorb more N from the complex organic N compounds than the other two species. The abilities of cedar, hemlock and salal to take up$^{15} {\rm{N}},\,^{13} {\rm{C}}$-labelled glutamic acid were measured and the capacities of the three species to use nitrate$({\rm{NO}}_3 ^ - ),$ammonium$({\rm{NH}}_4 ^ +),$glutamic acid, protein and protein-tannin N were compared over a 20-day period. Based on$^{13}{\rm{C}}$enrichment, all three species absorbed at least a portion of glutamic acid intact. Cedar, hemlock and salal also showed similar patterns of N uptake from the${\rm{NO}}_3^ - ,$${\rm{NH}}_4 ^ +,$glutamic acid, protein and protein-tannin treatments. The largest proportions of applied N were taken up from the${\rm{NO}}_3^ - $and${\rm{NH}}_4^ + $treatments while smaller amounts of N were absorbed from the organic N compounds. Thus organic N was accessed to a modest degree by all three species, and salal did not have a greater capacity to utilize protein and protein-tannin-N.
[3]
BinkleyD, AberJ, PastorJ, NadelhofferK (1986). Nitrogen availability in some Wisconsin forests: Comparisons of resin bags and on-site incubations. Biology Fertility of Soils, 2, 77-82.https://doi.org/10.1007/BF00257583URL [本文引用: 1]摘要 Estimates of ammonium and nitrate availability in conifer and hardwood forests using an ion exchange resin (IER) bag method and with on-site incubations of soil cores in buried bags were compared. Correlations between the two methods were generally high. Correlation coefficients ( r ) between IER nitrate and buried-bag mineralized nitrate ranged from 0.87 to 0.92. Both methods also correlated well with aboveground net primary production, litter fall N content, and fine root biomass. The major differences between the methods related to the relative importances of ammonium and nitrate forms of available N. The IER method indicated that both ammonium and nitrate were important on all sites, with nitrate predominating in most soils. The buried-bag results indicated that available N was primarily in the form of nitrate (all ammonium was oxidized), but that nitrate was insignificant on infertile sites.
[4]
BretelerH, SiegeristM (1984). Effect of ammonium on nitrate utilization by root of dwarf bean. Plant Physiology, 75, 1099-1103.https://doi.org/10.1016/S0031-9422(00)84871-2URLPMID:16663742摘要 Amino compounds (1 mM, pH 5) were given prior to, together with, or after the addition of nitrate to study their effect on nitrate uptake and in vivo nitrate reductase activity (NRA) in roots of Phaseolus vulgaris. The effect of amino compounds varied with the amino species, the nitrate status of the plant (induced vs uninduced) and the aspect of nitrate utilization. Cysteine inhibited the nitrate uptake rate and root NRA under all conditions tested. NRA in uninduced roots was stimulated by tryptophan, and arginine inhibited NRA under all conditions tested. Uptake was inhibited by aspartate and glutamate and stimulated by leucine when these amino compounds were given prior to or after completion of the apparent induction of nitrate uptake. In the presence of 尾-alanine and tryptophan, induction of uptake was accelerated.
[5]
BrittoDT, KronzuckerHJ (2002). NH4+ toxicity in higher plants: A critical review. Journal of Plant Physiology, 159, 567-584.https://doi.org/10.1078/01761610222260815URL [本文引用: 1]摘要 Ammonium (NH 4 +) toxicity is an issue of global ecological and economic importance. In this review, we discuss the major themes of NH 4 + toxicity, including the occurrence of NH 4 + in the biosphere, response differences to NH 4 + nutrition among wild and domesticated species, symptoms and proposed mechanisms underlying toxicity, and means by which it can be alleviated. Where possible, nitrate (NO 3 鈭) nutrition is used as point of comparison. Particular emphasis is placed on issues of cellular pH, ionic balance, relationships with carbon biochemistry, and bioenergetics of primary NH 4 + transport. Throughout, we attempt to identify areas that are controversial, and areas that are in need of further examination.
[6]
BurtonJ, ChenCR, XuZH, GhadiriH (2007). Soluble organic nitrogen pools in adjacent native and plantation forests of subtropical Australia. Soil Biology & Biochemistry, 39, 2723-2734.https://doi.org/10.1016/j.soilbio.2007.05.021URL [本文引用: 1]摘要 Soil soluble organic nitrogen (SON) can play an important role in soil nitrogen (N) cycling in forest ecosystems. This study examined the effect of land-use change from a native forest (NF) to a first rotation (1R) and subsequent second rotation (2R) hoop pine ( Araucaria cunninghamii ) plantation on soil SON pools. The impact of residue management on SON pools was also investigated in the 2R forest, where SON was measured in tree rows (2R-T) and windrows (2R-W). Various extraction techniques were used to measure SON pool size in the 0–10, 10–20 and 20–3002cm layers of soil. The results showed that land-use change had a significant impact on soil SON pools. In the 0–1002cm layer, 3.2–8.7, 14–23, 20–28, 60–160 and 127–34002mg02SON02kg 611 were extracted by water, 0.502M K 2 SO 4 , 202M KCl, hot water and hot 202M KCl, respectively. The size of the SON pools and the potential production of SON (PPSON) were generally highest in the NF soil and lowest in the 2R-T soil, and in all forest types decreased with soil depth. The larger SON pools in the NF soil coincided with lower soil, litter and root C:N ratios, suggesting that the difference in the size of SON pools between the NF and 1R soil may be related to differences in the quality of organic matter input under the different forest ecosystems. Differences in the size of SON pools between the 1R soil and the 2R soils and between the 2R-T soil and the 2R-W soil may be related to the quantity of organic matter input and time since disturbance. Significant relationships were found between the SON extracted by 0.502M K 2 SO 4 (SON ps ) and 202M KCl (SON KCl ), and also among the SON extracted by hot 202M KCl (SON hKCl ), hot water (SON hw ) and water (SON w ), suggesting that the organic N released by these groups of extracts may be at least partly from similar pools.
[7]
Chapin FSⅢ, MoilanenL, KiellandK (1993). Preferential use of organic nitrogen for growth by a non-mycorrhizal arctic sedge. Nature, 361, 150-153.URL [本文引用: 1]
[8]
ChenG, GuoSW, KronzuckerHJ, ShiWM (2013). Nitrogen use efficiency ( NUE) in rice links to NH4+ toxicity and futile NH4+ cycling in roots. Plant and Soil, 369, 351-363. [本文引用: 1]
[9]
CoskunD, BrittoDT, LiMY, BeckerA, KronzuckerHJ (2013). Rapid ammonia gas transport accounts for futile transmembrane cycling under NH3/NH4+ toxicity in plant roots. Plant Physiology, 163, 1859-1867.URL [本文引用: 1]
[10]
CuiXY (2007). Organic nitrogen use by plants and its significance in some natural ecosystems. Acta Ecologica Sinica, 27, 3500-3512. (in Chinese with English abstract)[崔晓阳 (2007). 植物对有机氮源的利用及其在自然生态系统中的意义 . 生态学报, 27, 3500-3512.]https://doi.org/10.3321/j.issn:1000-0933.2007.08.049URL [本文引用: 2]摘要 近来大量实验研究表明,许多植物能够在不经矿化的情况下直接吸收、利用环境介质中的生物有机氮,尤其氨基酸类。而且,有些植物利用氨基酸的效率可以与矿质氮源(NH4+、NO3)相当或更高。自然界植物赖以生存的土壤生境中同时存在多种有机氮和矿质氮养分,这是导致植物(至少部分植物)进化产生利用各种不同氮源能力的环境驱动力。土壤中的游离氨基酸尽管含量不高,但其周转快、通量大,理论上可远大于植物的氮需求。尽管植物在与土壤微生物的有机氮源竞争中处于根本性劣势,但土壤中氨基酸的巨大潜在通量和植物相对于微生物的生命周期仍可使植物在长期竞争中获取数量可观的氮。基于植物根对氨基酸的吸收能力、土壤中游离氨基酸库的大小和通量、植物与土壤微生物对氨基酸氮源的竞争以及有关的原位实验结果,近来许多研究者都认为植物有机氮营养在多种生态系统中是重要或潜在重要的。尤其是在一些极地、高山、亚高山、北方针叶林或泰加林生态系统中,由于低温等因素限制有机氮矿化,土壤氨基酸浓度常超过矿质氮(NH4+、NO3-)浓度,氨基酸可能代表着植物的一个主要氮源。认识到现实生态系统中植物对有机氮源利用的重要性意味着传统的矿质营养观念的更新,这将在很大程度上改变人们对某些重要生态过程的理解,并导致对若干生态学中心问题的再认识。研究以森林生态系统为例,阐述了我国开展该领域研究的科学意义和基本框架。
[11]
ElserJJ, BrackenMES, ClelandEE, GrunerDS, HarpoleWS (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 10, 1135-1142.URL [本文引用: 1]
[12]
FinziAC, BerthrongST (2005). The uptake of amino acids bymicrobes and trees in three cold-temperate forests. Ecology, 86, 3345-3353.https://doi.org/10.1890/04-1460URL [本文引用: 2]摘要 Amino acids are emerging as a critical component of the terrestrial N cycle, yet there is little understanding of amino acid cycling in temperate forests. This research studied the uptake and turnover of amino acid N by soil microbes and the capacity of forest trees to take up the amino acid glycine in comparison to NH4+and NO3-. This research was conducted in three temperate forests located in northwest Connecticut, USA. The three forests differed in soil parent material and canopy tree species composition. At all three sites, amino acids were released from soil organic matter through the activity of proteolytic enzymes resulting in a pool of free amino acids in soil. Free amino acids were rapidly immobilized by soil microbes. A15N-enriched-glycine-addition experiment also showed that a significant fraction of the amino acid N taken up by soil microbes was mineralized to NH4+with substantial nitrification at one site. Tree species from all three sites had the physiological capacity to absorb the amino acid glycine but took up amino acid N, NH4+, and NO3-in proportion to their availability in the soil. At the site with the highest gross fluxes of N, nearly all the N in amino acids was mineralized, and fine roots assimilated inorganic N much more rapidly than amino acid N. At the two sites with slower rates of gross amino acid production, the pool of free amino acids was larger, and fine roots assimilated amino acid N almost as fast as inorganic N. This study demonstrates that amino acids are an important component of the N cycle in temperate forests.
[13]
Gallet-BudynekA, BrzostekE, RodgersVL, TalbotJM, HyzyS, FinziAC (2009). Intact amino acid uptake by northern hardwood and conifer trees. Ecosystem Ecology Original Paper, 160, 129-138.https://doi.org/10.1007/s00442-009-1284-2URLPMID:19238450 [本文引用: 2]摘要 Empirical and modeling studies of the N cycle in temperate forests of eastern North America have focused on the mechanisms regulating the production of inorganic N, and assumed that only inorganic forms of N are available for plant growth. Recent isotope studies in field conditions suggest that amino acid capture is a widespread ecological phenomenon, although northern temperate forests have yet to be studied. We quantified fine root biomass and applied tracer-level quantities of U-0106C60-0167 N-glycine, 0167NH6262 and 0167NO6163 in two stands, one dominated by sugar maple and white ash, the other dominated by red oak, beech, and hemlock, to assess the importance of amino acids to the N nutrition of northeastern US forests. Significant enrichment of 0106C in fine roots 2 and 5 h following tracer application indicated intact glycine uptake in both stands. Glycine accounted for up to 77% of total N uptake in the oak-beech-hemlock stand, a stand that produces recalcitrant litter, cycles N slowly and has a thick, amino acid-rich organic horizon. By contrast, glycine accounted for only 20% of total N uptake in the sugar maple and white ash stand, a stand characterized by labile litter and rapid rates of amino acid production and turnover resulting in high rates of mineralization and nitrification. This study shows that amino acid uptake is an important process occurring in two widespread, northeastern US temperate forest types with widely differing rates of N cycling.
[14]
GesslerA, SchneiderS, von SengbuschD, WeberP, HanemannU, HuberC, RotheA, KreutzerK, RennenbergH (1998). Field and laboratory experiments on net uptake of nitrate and ammonium by the roots of spruce ( Picea abies) and beech(Fagus sylvatica) trees. New Phytologist, 138, 275-285. [本文引用: 1]
[15]
JonesDL, HealeyJR, WillettVB, FarrarJF, HodgeA (2005). Dissolved organic nitrogen uptake by plants—An important N uptake pathway. Soil Biology & Biochemistry, 37, 413-423.https://doi.org/10.1016/j.soilbio.2004.08.008URL [本文引用: 2]摘要 The direct uptake of dissolved organic nitrogen (DON) by plants has the potential to be a primary Factor in ecosystem functioning and vegetation succession particularly in N-limiting environments. Clear experimental evidence to support this view, however, is still lacking. Further, many of the experimental approaches used to assess whether DON is important may be compromised due to the use of inappropriate methods for comparing and quantifying plant available inorganic and organic soil N pools. In addition, experiments aimed at quantifying plant DON capture using dual-labelled ( 15N, 13C) organic N tracers often do not consider important aspects such as isotope pool dilution, differences in organic and inorganic N pool turnover times, bi-directional DON flows at the soil 搑oot interface, and the differential fate of the 15N and 13C in the tracer compounds. Based upon experimental evidence, we hypothesize that DON uptake from the soil may not contribute largely to N acquisition by plants but may instead be primarily involved in the recapture of DON previously lost during root exudation. We conclude that while root uptake of amino acids in intact form has been shown, evidence demonstrating this as a major plant N acquisition pathway is still lacking.
[16]
LajthaK (1994). Nutrient uptake in eastern deciduous tree seedlings. Plant and Soil, 160, 193-199.https://doi.org/10.1007/BF00010145URL [本文引用: 1]摘要 Tree seedlings that colonize large treefall gaps are generally shade-intolerant species with high potential relative growth rates. Nutrient availability may be significantly elevated in disturbance-induced gaps, however, little is known about the role of differences in nutrient uptake capacities of different species in structuring the community response to gap openings in eastern North American deciduous forests. Seven tree species were grown from seed under both a high and a low nutrient regime, and uptake kinetics of phosphate, ammonium, and nitrate were studied. Yellow birch, a species with intermediate shade tolerance and relative growth rate, had the highest maximum rates of uptake of all ions, while tulip tree, a gap-colonizing species with high relative growth rate, had the lowest rate of phosphate uptake and intermediate rates of ammonium and nitrate uptake. Beech and hickory, which have low relative growth rates and are not gap-colonizing species, had intermediate levels of nutrient uptake. There was no evidence that species with the highest maximum uptake rates measured at high supply concentrations had relatively low uptake at low nutrient supply concentrations. Although birch increased phosphate absorption capacity when grown under a low nutrient regime, this pattern did not hold for nitrate or ammonium uptake, and other species showed no change in nutrient uptake capacity according to nutrient growth regime. Clearly, factors other than nutrient absorption capacity, such as nutrient use efficiency or allocation to root vs. shoot biomass, underlie differences in species' capacities to colonize and maintain a high relative growth rate in canopy gaps.
[17]
LiHS (2002). Modern Plant Physiology. Higher Education Press, Beijing. (in Chinese)[李合生 (2002). 现代植物生理学. 高等教育出版社, 北京.] [本文引用: 1]
[18]
LiYJ, SunDD, LiDD, XuZF, ZhaoCZ, LinHH, LiuQ (2015). Effects of warming on ectomycorrhizal colonization and nitrogen nutrition of Picea asperata seedlings grown in two contrasting forest ecosystems. Scientific Report, 5, 17546. doi: 10.1038/srep17546. [本文引用: 1]
[19]
LipsonD, NasholmT (2001). The unexpected versatility of plants: Organic nitrogen use and availability in terrestrial ecosystems. Oecologia, 128, 305-316. [本文引用: 1]
[20]
LuJL (2003). Plant Nutriology. China Agricultural University Press, Beijing. (in Chinese)[陆景陵 (2003).植物营养学. 中国农业大学出版社, 北京.] [本文引用: 1]
[21]
McKaneRB, JohnsonLC, ShaverGR, NadelhofferKJ, RastetterEB, FryB, GiblinAE, KiellandK, KwiatkowskiBL, LaundreJA, MurrayG (2002). Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. Nature, 415, 68-71. [本文引用: 1]
[22]
MillerAE, BowmanWD, SudingKN (2007). Plant uptake of inorganic and organic nitrogen: Neighbor identity matters. Ecology, 88, 1832-1840. [本文引用: 1]
[23]
NanHW, LiuQ, ChenJS, ChengXY, YinHJ, YinCY, ZhaoCZ (2013). Effects of nutrient heterogeneity and competition on root architecture of spruce seedlings: implications for an essential feature of root foraging. PLOS ONE, 8, e65650. doi: 10.1371/journal.pone.0065650. [本文引用: 1]
[24]
N?sholmT, EkbladA, NordinA, GieslerR, HogbergM, HogbergP (1998). Boreal forest plants take up organic nitrogen. Nature, 392, 914-916. [本文引用: 2]
[25]
N?sholmT, KiellandK, GanetegU (2009). Uptake of organic nitrogen by plants. New Phytologist, 182, 31-48. [本文引用: 3]
[26]
NordinA, HogbergP, N?sholmT (2001). Soil nitrogen form and plant nitrogen uptake along a boreal forest productivity gradient. Oecologia, 129, 125-132.
[27]
OhlundJ, N?sholmT (2001). Growth of conifer seedlings on organic and inorganic nitrogen sources. Tree Physiology, 18, 1319-1326. [本文引用: 1]
[28]
PastorJ, AberJ, McClaughertyC, MelilloJ (1984). Aboveground production and N and P cycling along a nitrogen mineralization gradient on Blackhawk Island, Wisconsin. Ecology, 65, 256-268. [本文引用: 1]
[29]
PerssonJ, Gardestr?mP, N?sholmT (2006). Uptake, metabolism and distribution of organic and inorganic nitrogen sources by Pinus sylvestris. Journal of Experimental Botany, 57, 2651-2659. [本文引用: 1]
[30]
RothsteinDE (2014). In-situ root uptake and soil transformations of glycine, glutamine and ammonium in two temperate deciduous forests of contrasting N availability. Soil Biology & Biochemistry, 75, 233-236. [本文引用: 1]
[31]
SongFQ, TianXJ, YangCL, HeXB, ChenBin, ZhuJ, HaoJJ (2006). Ectomycorrhizal infection intensity of subalpine forest ecosystems in western Sichuan, China. Acta Ecologica Sinica, 26, 4171-4178. (in Chinese with English abstract)[宋福强, 田兴军, 杨昌林, 何兴兵, 陈彬, 朱静, 郝杰杰 (2006). 川西亚高山带森林生态系统外生菌根的形成 . 生态学报, 26, 4171-4178.] [本文引用: 1]
[32]
SongMH, ZhengLL, SudingKN, YinTF, YuFH (2015). Plasticity in nitrogen form uptake and preference in response to long-term nitrogen fertilization. Plant and Soil, 394, 215-224. [本文引用: 1]
[33]
StribleyDP, ReadDJ (1980). The biology of mycorrhizae in the Ericaceae VII. The relationship between mycorrhizal infection and the capacity to utilize simple and complex organic nitrogen sources. New Phytologist, 86, 365-371. [本文引用: 1]
[34]
TemplerP, DawsonT (2004). Nitrogen uptake by four tree species of the Catskill Mountains, New York: Implications for forest N dynamics. Plant and Soil, 262, 251-261. [本文引用: 1]
[35]
WangLX, MackoSA (2011). Constrained preferences in nitrogen uptake across plant species and environments. Plant, Cell & Environment, 34, 525-534. [本文引用: 1]
[36]
WarrenCR (2009). Does nitrogen concentration affect relative uptake rates of nitrate, ammonium, and glycine? Journal of Plant Nutrition and Soil Science, 172, 224-229. [本文引用: 1]
[37]
WarrenCR, TarantoMT (2010). Temporal variation in pools of amino acids, inorganic and microbial N in a temperate grassland soil. Soil Biology & Biochemistry, 42, 353-359. [本文引用: 1]
[38]
WeiLL, ChenCR, YuS (2014). Uptake of organic nitrogen and preference for inorganic nitrogen by two Australian native Araucariaceae species. Plant Ecology & Diversity, 8, 259-264. [本文引用: 1]
[39]
XuZF, HuR, XiongP, WanCA, CaoG, LiuQ (2010). Initial soil responses to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China: Nutrient availabilities, microbial properties and enzyme activities. Applied Soil Ecology, 46, 291-299. [本文引用: 1]
[40]
YinHJ, LaiT, ChenXY, JiangXM, LiuQ (2008). Warming effects on growth and physiology of seedlings of Betula albo-sinensis and Abies faxoniana under two contrasting light conditions in subalpine coniferous forest of western Sichuan, China. Journal of Plant Ecology (Chinese Version), 32, 1072-1083. (in Chinese with English abstract)[尹华军, 赖挺, 程新颖, 蒋先敏, 刘庆 (2008). 增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响 . 植物生态学报, 32, 1072-1083.] [本文引用: 1]
[41]
ZhangZL, YuanYS, ZhaoWQ, HeHL, LiDD, HeW, LiuQ, YinHJ (2017). Seasonal variations in the soil amino acid pool and flux following the conversion of a natural forest to a pine plantation on the eastern Tibetan Plateau, China. Soil Biology & Biochemistry, 105, 1-11. [本文引用: 2]
Niche complementarity due to plasticity in resource use: Plant partitioning of chemical N forms. 1 2010
... 一般认为, 植物吸收NH4+-N的过程比吸收NO3–-N的过程节省更多能量(Wang & Macko, 2011), 土壤NH4+-N进入植物根系后直接被转化为谷氨酸盐从而被植物进一步利用, 而植物在吸收NO3–-N的过程中必须通过消耗较多能量的硝酸盐还原途径才能被植物吸收利用(Templer & Dawson, 2004).因此, 相对于NO3–-N, 植物通常更偏好吸收土壤中的NH4+-N (Finzi & Berthrong, 2005; N?sholm et al., 2009).然而, 本研究结果表明: 川西亚高山针叶林树种云杉和红桦更偏好吸收土壤NO3–-N.具体而言, 与氨基酸和NH4+-N处理相比, NO3–-N处理的植物细根表现出显著更高的δ15N值和更快的吸收速率(图2, 图3).此外, 云杉和红桦幼苗根系对NO3–-N的平均吸收速率为NH4+-N和氨基酸平均吸收速率的5-10倍(图4).该研究结果与大部分前人研究认为的植物偏好吸收和利用土壤NH4+-N的观点不一致.我们推测一个重要的原因可能与供试土壤有较高的NO3–-N含量有关.前期研究发现, 植物对土壤N的吸收具有可塑性, 当土壤中各种N的有效性发生变化时, 植物会调整自身对N的吸收偏好来维持甚至增加整体N的吸收(Ashton et al., 2010; Song et al., 2015).Breteler和Siegerist (1984)的研究表明土壤中高浓度的NH4+-N抑制植物根系对NO3–-N的吸收, 而当NH4+- N浓度低于1 mmol·L-1时, 这种抑制作用将降至最小, 植物根系不再排斥NO3–-N, 可将其作为N源.在本研究中, 土壤NH4+-N浓度仅有-0.75 mmol·L-1, 因此其对植物吸收NO3–-N的抑制作用可忽略不计.Nordin等(2001)在研究北方森林植物N吸收过程中也发现植物对土壤中的不同形态N的偏好吸收与这些N素在土壤中的有效性呈正相关关系.因此, 我们认为本研究中植物对NO3–-N表现出显著更高的吸收速率(图3), 可能在很大程度上归因于土壤中相对较高的NO3–-N浓度.NO3–-N作为川西亚高山针叶林土壤中最主要的N形态(Xu et al., 2010), 其被云杉和红桦幼苗偏好吸收也是亚高山森林植物长期适应该区域土壤环境的结果.另外, NO3–-N在土壤中具有更高的可移动性也可导致植物对NO3–-N表现出明显的偏好吸收现象(Warren, 2009; Li et al., 2015).比如, NO3–-N在土壤溶液中的扩散系数比NH4+-N高10-100倍(Gessler et al., 1998), 因此前者能够通过土壤溶液更快地扩散到根系从而为植物提供更大的吸收几率.尽管植物对NH4+-N的吸收消耗更少的能量, 但许多植物对NH4+-N具有较高的敏感性(Chen et al., 2013).与NO3–-N相比, NH4+-N在植物体积累易产生毒性效应, 抑制K+、Ca2+、Mg2+等阳离子的吸收, 从而不利于植物的生长(Britto & Kronzucker, 2002; Coskun et al., 2013), 这可能也是植物偏好吸收NO3–-N的原因之一.然而, 关于不同生态系统、不同树种对NO3–-N和NH4+-N的偏好吸收与利用的影响因素与调控机制并不清楚, 需要进一步开展深入的研究. ...
Organic and inorganic nitrogen nutrition of western red cedar, western hemlock and salal in mineral N-limited cedar-hemlock forests. 1 2004
... 氮(N)是植物需求量最大的一种矿质营养元素, 是植物体内核酸、蛋白质、酶等物质的主要组成成分, 同时也是陆地生态系统植物生长发育的主要限制因子(Jones et al., 2005), 对陆地生态系统净初级生产力和功能具有极其重要的作用(Elser et al., 2007).土壤中的N有多种赋存形态和性质, 主要可分为无机N (以NH4+-N和NO3–-N为主)和有机N (如氨基酸、酰胺、尿素等)两大类.土壤无机N是土壤N的主要组成部分, 占土壤总N的90%以上(李合生, 2002).传统的矿质营养学说通常认为植物只能吸收无机N (如NH4+-N和NO3–-N), 有机N必须被分解为无机N后才能被植物吸收.然而, 随着同位素技术的广泛运用和不断发展, 越来越多的研究表明: 许多植物(包括菌根植物和一些无菌根植物)能够直接吸收利用土壤中的多种氨基酸、简单蛋白质等小分子和大分子有机N化合物(Bennett & Prescott, 2004; Miller et al., 2007; Rothstein, 2014).在一些常年低温、N矿化受到强烈限制的地区, 植物对有机N的吸收速率甚至超过无机N, 此现象引发了人们对有机N在陆地生态系统营养流动与N循环模型的重新思考和评价(Gallet-Budynek et al., 2009). ...
Nitrogen availability in some Wisconsin forests: Comparisons of resin bags and on-site incubations. 1 1986
... 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009).比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001).然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006).造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014).因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制. ...
Effect of ammonium on nitrate utilization by root of dwarf bean. 1984
NH4+ toxicity in higher plants: A critical review. 1 2002
... 一般认为, 植物吸收NH4+-N的过程比吸收NO3–-N的过程节省更多能量(Wang & Macko, 2011), 土壤NH4+-N进入植物根系后直接被转化为谷氨酸盐从而被植物进一步利用, 而植物在吸收NO3–-N的过程中必须通过消耗较多能量的硝酸盐还原途径才能被植物吸收利用(Templer & Dawson, 2004).因此, 相对于NO3–-N, 植物通常更偏好吸收土壤中的NH4+-N (Finzi & Berthrong, 2005; N?sholm et al., 2009).然而, 本研究结果表明: 川西亚高山针叶林树种云杉和红桦更偏好吸收土壤NO3–-N.具体而言, 与氨基酸和NH4+-N处理相比, NO3–-N处理的植物细根表现出显著更高的δ15N值和更快的吸收速率(图2, 图3).此外, 云杉和红桦幼苗根系对NO3–-N的平均吸收速率为NH4+-N和氨基酸平均吸收速率的5-10倍(图4).该研究结果与大部分前人研究认为的植物偏好吸收和利用土壤NH4+-N的观点不一致.我们推测一个重要的原因可能与供试土壤有较高的NO3–-N含量有关.前期研究发现, 植物对土壤N的吸收具有可塑性, 当土壤中各种N的有效性发生变化时, 植物会调整自身对N的吸收偏好来维持甚至增加整体N的吸收(Ashton et al., 2010; Song et al., 2015).Breteler和Siegerist (1984)的研究表明土壤中高浓度的NH4+-N抑制植物根系对NO3–-N的吸收, 而当NH4+- N浓度低于1 mmol·L-1时, 这种抑制作用将降至最小, 植物根系不再排斥NO3–-N, 可将其作为N源.在本研究中, 土壤NH4+-N浓度仅有-0.75 mmol·L-1, 因此其对植物吸收NO3–-N的抑制作用可忽略不计.Nordin等(2001)在研究北方森林植物N吸收过程中也发现植物对土壤中的不同形态N的偏好吸收与这些N素在土壤中的有效性呈正相关关系.因此, 我们认为本研究中植物对NO3–-N表现出显著更高的吸收速率(图3), 可能在很大程度上归因于土壤中相对较高的NO3–-N浓度.NO3–-N作为川西亚高山针叶林土壤中最主要的N形态(Xu et al., 2010), 其被云杉和红桦幼苗偏好吸收也是亚高山森林植物长期适应该区域土壤环境的结果.另外, NO3–-N在土壤中具有更高的可移动性也可导致植物对NO3–-N表现出明显的偏好吸收现象(Warren, 2009; Li et al., 2015).比如, NO3–-N在土壤溶液中的扩散系数比NH4+-N高10-100倍(Gessler et al., 1998), 因此前者能够通过土壤溶液更快地扩散到根系从而为植物提供更大的吸收几率.尽管植物对NH4+-N的吸收消耗更少的能量, 但许多植物对NH4+-N具有较高的敏感性(Chen et al., 2013).与NO3–-N相比, NH4+-N在植物体积累易产生毒性效应, 抑制K+、Ca2+、Mg2+等阳离子的吸收, 从而不利于植物的生长(Britto & Kronzucker, 2002; Coskun et al., 2013), 这可能也是植物偏好吸收NO3–-N的原因之一.然而, 关于不同生态系统、不同树种对NO3–-N和NH4+-N的偏好吸收与利用的影响因素与调控机制并不清楚, 需要进一步开展深入的研究. ...
Soluble organic nitrogen pools in adjacent native and plantation forests of subtropical Australia. 1 2007
... 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009).比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001).然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006).造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014).因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制. ...
Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. 1 2002
... 在同位素标记物无丢失的前提下, 根据外源添加N浓度、细根样品中15N丰度值以及% N计算植物根系对土壤不同形态N的吸收速率(McKane et al., 2002), 公式如下: ...
Plant uptake of inorganic and organic nitrogen: Neighbor identity matters. 1 2007
... 氮(N)是植物需求量最大的一种矿质营养元素, 是植物体内核酸、蛋白质、酶等物质的主要组成成分, 同时也是陆地生态系统植物生长发育的主要限制因子(Jones et al., 2005), 对陆地生态系统净初级生产力和功能具有极其重要的作用(Elser et al., 2007).土壤中的N有多种赋存形态和性质, 主要可分为无机N (以NH4+-N和NO3–-N为主)和有机N (如氨基酸、酰胺、尿素等)两大类.土壤无机N是土壤N的主要组成部分, 占土壤总N的90%以上(李合生, 2002).传统的矿质营养学说通常认为植物只能吸收无机N (如NH4+-N和NO3–-N), 有机N必须被分解为无机N后才能被植物吸收.然而, 随着同位素技术的广泛运用和不断发展, 越来越多的研究表明: 许多植物(包括菌根植物和一些无菌根植物)能够直接吸收利用土壤中的多种氨基酸、简单蛋白质等小分子和大分子有机N化合物(Bennett & Prescott, 2004; Miller et al., 2007; Rothstein, 2014).在一些常年低温、N矿化受到强烈限制的地区, 植物对有机N的吸收速率甚至超过无机N, 此现象引发了人们对有机N在陆地生态系统营养流动与N循环模型的重新思考和评价(Gallet-Budynek et al., 2009). ...
Effects of nutrient heterogeneity and competition on root architecture of spruce seedlings: implications for an essential feature of root foraging. 1 2013
... 本研究地位于四川省理县境内杂谷脑镇胆扎木沟(31.41° N, 103.21° E, 海拔约2 300 m), 该区位于青藏高原东缘, 四川省西北部, 阿坝藏族羌族自治州东南缘, 地质结构属于龙门山断裂带中段, 山峦起伏, 地形复杂.该区气候类型属于山地季风气候, 夏季温凉多雨, 冬季寒冷干燥, 1月平均气温-8 ℃, 7月平均气温12.6 ℃, 年平均气温4.8-6.9 ℃, 无霜期115-150天, >10 ℃年有效积温1 400-2 500 ℃, 年降水量700-900 mm, 年干燥度0.59-1.18.生长季长度大概为6个月, 从4月下旬到10月下旬.冬季大概有4个月为冰雪覆盖期, 最大雪被厚度约为30 cm.土壤类型主要为山地棕壤(Nan et al., 2013). ...
Boreal forest plants take up organic nitrogen. 2 1998
... 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009).比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001).然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006).造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014).因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制. ...
Soil nitrogen form and plant nitrogen uptake along a boreal forest productivity gradient. 2001
Growth of conifer seedlings on organic and inorganic nitrogen sources. 1 2001
... 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009).比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001).然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006).造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014).因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制. ...
Aboveground production and N and P cycling along a nitrogen mineralization gradient on Blackhawk Island, Wisconsin. 1 1984
... 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009).比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001).然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006).造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014).因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制. ...
Uptake, metabolism and distribution of organic and inorganic nitrogen sources by 1 2006
... 尽管植物可直接吸收有机N已被普遍认识和接受, 但目前关于植物与N关系的研究更多聚焦于无机N的产生、周转和调控机理等方面(Pastor et al., 1984; Binkley et al., 1986; Lajtha, 1994; 陆景陵, 2003), 而对植物吸收有机N的特征了解甚少, 使得土壤中无机N源和有机N源对植物N养分贡献大小及其生态重要性结论不一致(N?sholm et al., 2009).比如, 一些证据表明植物偏好有机N而不是无机N, 或者对两者具有相同的吸收量, 尤其是在低温、N受限制的极地、高山和北方生态系统(Chapin et al., 1993; N?sholm et al., 1998; Ohlund & N?sholm, 2001).然而, 一些研究表明土壤无机N仍是植物最主要的N源(Finzi & Berthrong, 2005; Persson et al., 2006).造成这种差异的原因可能与植物种类及其所处的生长环境、土壤性质有关(Wei et al., 2014).因此, 需要深入开展不同生态系统土壤有机N和无机N对植物N源的贡献大小及其生态反馈效应的研究, 以便更好地认识森林生态系统N循环过程与调控机制. ...
In-situ root uptake and soil transformations of glycine, glutamine and ammonium in two temperate deciduous forests of contrasting N availability. 1 2014
... 氮(N)是植物需求量最大的一种矿质营养元素, 是植物体内核酸、蛋白质、酶等物质的主要组成成分, 同时也是陆地生态系统植物生长发育的主要限制因子(Jones et al., 2005), 对陆地生态系统净初级生产力和功能具有极其重要的作用(Elser et al., 2007).土壤中的N有多种赋存形态和性质, 主要可分为无机N (以NH4+-N和NO3–-N为主)和有机N (如氨基酸、酰胺、尿素等)两大类.土壤无机N是土壤N的主要组成部分, 占土壤总N的90%以上(李合生, 2002).传统的矿质营养学说通常认为植物只能吸收无机N (如NH4+-N和NO3–-N), 有机N必须被分解为无机N后才能被植物吸收.然而, 随着同位素技术的广泛运用和不断发展, 越来越多的研究表明: 许多植物(包括菌根植物和一些无菌根植物)能够直接吸收利用土壤中的多种氨基酸、简单蛋白质等小分子和大分子有机N化合物(Bennett & Prescott, 2004; Miller et al., 2007; Rothstein, 2014).在一些常年低温、N矿化受到强烈限制的地区, 植物对有机N的吸收速率甚至超过无机N, 此现象引发了人们对有机N在陆地生态系统营养流动与N循环模型的重新思考和评价(Gallet-Budynek et al., 2009). ...
Plasticity in nitrogen form uptake and preference in response to long-term nitrogen fertilization. 1 2015
... 一般认为, 植物吸收NH4+-N的过程比吸收NO3–-N的过程节省更多能量(Wang & Macko, 2011), 土壤NH4+-N进入植物根系后直接被转化为谷氨酸盐从而被植物进一步利用, 而植物在吸收NO3–-N的过程中必须通过消耗较多能量的硝酸盐还原途径才能被植物吸收利用(Templer & Dawson, 2004).因此, 相对于NO3–-N, 植物通常更偏好吸收土壤中的NH4+-N (Finzi & Berthrong, 2005; N?sholm et al., 2009).然而, 本研究结果表明: 川西亚高山针叶林树种云杉和红桦更偏好吸收土壤NO3–-N.具体而言, 与氨基酸和NH4+-N处理相比, NO3–-N处理的植物细根表现出显著更高的δ15N值和更快的吸收速率(图2, 图3).此外, 云杉和红桦幼苗根系对NO3–-N的平均吸收速率为NH4+-N和氨基酸平均吸收速率的5-10倍(图4).该研究结果与大部分前人研究认为的植物偏好吸收和利用土壤NH4+-N的观点不一致.我们推测一个重要的原因可能与供试土壤有较高的NO3–-N含量有关.前期研究发现, 植物对土壤N的吸收具有可塑性, 当土壤中各种N的有效性发生变化时, 植物会调整自身对N的吸收偏好来维持甚至增加整体N的吸收(Ashton et al., 2010; Song et al., 2015).Breteler和Siegerist (1984)的研究表明土壤中高浓度的NH4+-N抑制植物根系对NO3–-N的吸收, 而当NH4+- N浓度低于1 mmol·L-1时, 这种抑制作用将降至最小, 植物根系不再排斥NO3–-N, 可将其作为N源.在本研究中, 土壤NH4+-N浓度仅有-0.75 mmol·L-1, 因此其对植物吸收NO3–-N的抑制作用可忽略不计.Nordin等(2001)在研究北方森林植物N吸收过程中也发现植物对土壤中的不同形态N的偏好吸收与这些N素在土壤中的有效性呈正相关关系.因此, 我们认为本研究中植物对NO3–-N表现出显著更高的吸收速率(图3), 可能在很大程度上归因于土壤中相对较高的NO3–-N浓度.NO3–-N作为川西亚高山针叶林土壤中最主要的N形态(Xu et al., 2010), 其被云杉和红桦幼苗偏好吸收也是亚高山森林植物长期适应该区域土壤环境的结果.另外, NO3–-N在土壤中具有更高的可移动性也可导致植物对NO3–-N表现出明显的偏好吸收现象(Warren, 2009; Li et al., 2015).比如, NO3–-N在土壤溶液中的扩散系数比NH4+-N高10-100倍(Gessler et al., 1998), 因此前者能够通过土壤溶液更快地扩散到根系从而为植物提供更大的吸收几率.尽管植物对NH4+-N的吸收消耗更少的能量, 但许多植物对NH4+-N具有较高的敏感性(Chen et al., 2013).与NO3–-N相比, NH4+-N在植物体积累易产生毒性效应, 抑制K+、Ca2+、Mg2+等阳离子的吸收, 从而不利于植物的生长(Britto & Kronzucker, 2002; Coskun et al., 2013), 这可能也是植物偏好吸收NO3–-N的原因之一.然而, 关于不同生态系统、不同树种对NO3–-N和NH4+-N的偏好吸收与利用的影响因素与调控机制并不清楚, 需要进一步开展深入的研究. ...
The biology of mycorrhizae in the Ericaceae VII. The relationship between mycorrhizal infection and the capacity to utilize simple and complex organic nitrogen sources. 1 1980
Seasonal variations in the soil amino acid pool and flux following the conversion of a natural forest to a pine plantation on the eastern Tibetan Plateau, China. 2 2017

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT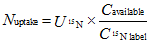 (1)
(1) 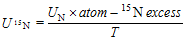 (2)
(2)  显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT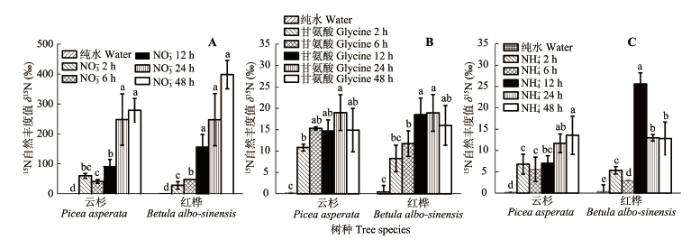 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT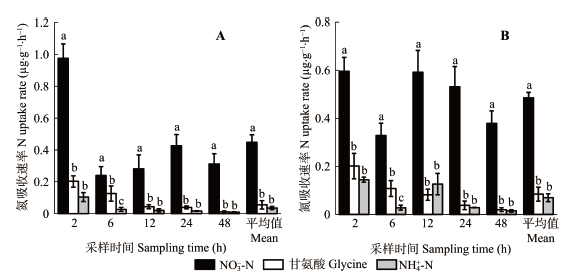 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT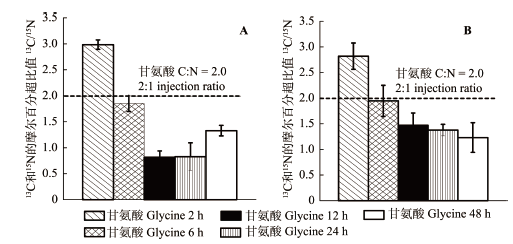 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}