
海南大学热带农林学院,环南海陆域生物多样性研究中心, 海口 570228
Effects of monsoon on distribution patterns of tropical plants in Asia
JIANGChao, TANKe, RENMing-Xun通讯作者:
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (404KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
季风(monsoon)是由于太阳高度角变化引起海陆热力性质差异或气压带随季节移动导致盛行风向和降水随季节改变的现象(An et al., 2001)。季风的发生以盛行风向的改变和剧烈降水为特征, 往往伴随着大气能量和水分循环的急速加强(吴国雄等, 2013), 直接影响着全球水热循环和热量分布(Webster et al., 1998; An et al., 2000), 调控着全球的气候变化。
亚洲地区是全球季风气候最为典型的区域, 最主要的特征就是降水多发生在高温的夏季(Tao & Chen, 1987; 周浙昆等, 2017)。根据发生时间, 亚洲的季风可分为冬季风、夏季风(表1)。夏季风根据盛行风向、发生范围与成因可分为3支: 东亚季风、南亚季风和西北太平洋季风(Wang, 2002; 高雅和王会军, 2012; Salinger et al., 2014)。其中, 南亚季风和西北太平洋季风发源于赤道一带的暖湿气流, 可称为“热带季风”。
目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群。亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(孙航和周浙昆, 1996; Ren, 2015; 周浙昆等, 2017)。中国-缅甸生物多样性热点地区(包括中南半岛和中国的云南南部、广西西南部以及海南岛)的植物多样性也有很大比例为热带植物属性(Myers et al., 2000; 朱华, 2011; Zhu, 2013)。这些位于热带北缘的地区有着如此高水平的热带植物多样性, 其中一个关键因素可能就是热带季风的作用(Wurster et al., 2010; Liu et al. 2013; Ren, 2015; 周浙昆等, 2017)。
本文拟着重分析亚洲热带季风气候对热带植物迁移特别是北向迁移的作用, 揭示季风对热带北缘部分物种局域隔离分化的影响, 并解释特殊地形局域地带性与非地带性植被如热带季节性雨林、热带季雨林、中国西南干旱河谷及海南岛西部的稀树灌丛与草原等的形成原因; 并以4个典型热带植物类群苦苣苔亚科(Cyrtandroideae)、秋海棠属(Begonia)、金虎尾科(Malpighiaceae)风筝果属(Hiptage)和盾翅藤属(Aspidopterys)为例, 分析热带植物物种丰富度中心的地理分布, 揭示季风气候对亚洲热带植物分布格局及物种多样性与特有种分布中心形成的影响。
1 亚洲热带季风概况
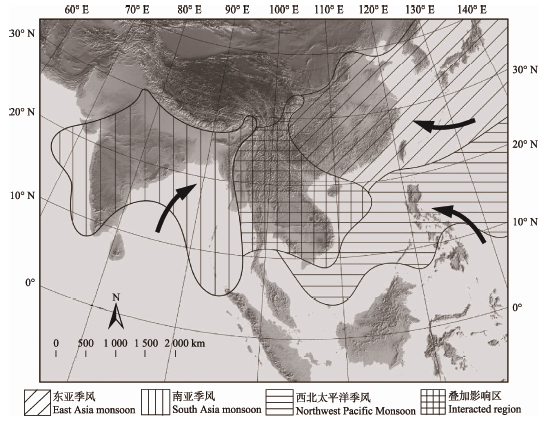
亚洲的热带季风包括南亚季风(影响南亚和东南亚)和西北太平洋季风(影响菲律宾、中国南海及华南沿海地区), 与东亚季风(主要影响中国长江流域、日本和韩国等地)同为夏季风(表1)。这3支夏季风的大气环流和水汽输送联系紧密, 相互影响, 直接决定着整个南亚、东南亚和东亚地区夏季的降雨和气温(高雅和王会军, 2012)。其中, 中国西南-中南半岛一带是这三大季风交汇的叠加区域(图1), 是亚洲降水最为丰沛的地区之一(Salinger et al., 2014)。Table 1
表1
表1亚洲季风基本情况
Table 1Main characteristics of monsoons in Asia
| 季风类型 Monsoon type | 发生月份 Occurrence month | 影响区域 Affected area | 历史形成时间 Age (Ma) | 参考文献 Reference | |
|---|---|---|---|---|---|
| 夏季风 Summer monsoon | 东亚季风(东南季风) East Asia monsoon; (Southeast monsoon) | 6-8月 June to August | 中国中部与南部; 日本南部; 朝鲜半岛 Center and South China; South Japan; Korea Peninsula | 25-22 | Sun & Wang, 2005 |
| 9-8 | An et al., 2001 | ||||
| 西北太平洋季风 Northwest Pacific monsoon | 5-9月 May to September | 中国南海及其沿岸地区; 东南亚含中南半岛 South China Sea and its coastal areas; Southeast Asia including Indo-China Peninsula | 未知 Unknown | Li et al., 2014 | |
| 南亚季风(西南季风) South Asia monsoon (Southwest monsoon) | 5-7月 May to July | 中国海南岛, 云南与广西南部; 印度和中南半岛 Hainan Island, Southern Yunnan and Guangxi of China; India and Indo-China Peninsula | 20 | Wu et al., 2013 | |
| 9-8 | An et al., 2001 | ||||
| 冬季风 Winter monsoon | 10月到次年2月 October to next February | 中国北方与中部; 蒙古 North and Center China; Mongolia | 2.6 | An et al., 2001 | |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1亚洲夏季风及其影响区域(箭头示主要风向)。
-->Fig. 1Asia summer monsoons and their affected areas (arrow indicates the main wind direction).
-->
南亚季风的成因既有印度洋与印度半岛的海陆热力性质差异的驱动, 也受东南亚及大洋洲一带气压带在夏季北移的影响(Tao & Chen, 1987; 高雅和王会军, 2012); 同时, 高耸的青藏高原也影响着南亚季风的风向和风力(Tao & Chen, 1987; 周晓霞等, 2008; 高雅和王会军, 2012)。南亚季风在5-7月强度最大, 风向以西南向东北为主, 主要影响印度、孟加拉湾、中南半岛, 以及我国西南地区如横断山区和云贵高原(周晓霞等, 2008)。
西北太平洋季风发生在每年的5-9月, 发源于太平洋西部和新几内亚东岸150° E附近区域的一条跨越赤道的气流通道(Tao & Chen, 1987)。由于地球自转力以及南亚季风的影响, 西北太平洋季风的风向由东南向西北转而吹向东北方向, 影响着南海沿岸地区包括菲律宾、中国华南沿海地区和中南半岛部分区域(图1)。西北太平洋季风有时伴随着热带风暴和台风(Li & Wang, 2005; 李肖雅等, 2014), 对南海及周边地区植物生长和迁移影响巨大。
2 季风气候对亚洲热带植物分布的影响
对古植物化石的研究发现, 热带植物曾经在东亚特别是中国南部有更广阔的分布区, 热带北界比现今的热带北界更靠北, 如中新世时期中国的东南部和24° N地区存在着龙脑香科植物(Shi & Li, 2010)、中国中部一带分布有热带和亚热带常绿阔叶林(Yu et al., 2000)。这说明, 历史上季风气候(主要是形成于25-22 Ma的东亚季风)确实促进和维持了热带植物分布区在较北区域的分布(孙湘君和汪品先, 2005)。之后, 随着冬季风的出现, 第四季冰期和造山运动造成的压力, 东亚季风的影响逐渐南退, 难以对中国北部和西部地区产生显著影响, 热带植物大幅南撤(孙航和周浙昆, 1996; Yu et al., 2000)。在热带植物南退的历史过程中, 部分热带植物在一些地区特殊地形地貌的“避难所”作用下得以存活, 再加上后期(不早于20 Ma; 表1)形成的南亚季风和西北太平洋季风的强大影响, 一些热带植物类群在热带北缘地带保存至今。现代亚洲热带植物分布的北界位于中国西南, 整体处于云南南部、广西西南部、海南岛、台湾岛南部一线(Zhu, 2013), 并在当地复杂地形的影响下形成了多种多样的局域小气候和非地带性植被(Zhu, 2013; Ren, 2015)。
2.1 季风与热带季节性雨林
亚洲的热带季节性雨林是热带雨林在热带边缘零星分布的常绿或半常绿森林, 主要出现在受到热带季风气候深刻影响的热带边缘区域, 如亚洲热带南缘的澳大利亚北领地、热带北缘的中国云南西双版纳、海南岛及广西与越南接壤处。从优势种、植被外貌来看, 季节性雨林与热带雨林相差不大, 建群种和优势种都以龙脑香科的望天树(Parashorea chinensis)和婆罗双(Shorea assamica)(Tuyet, 2001)、椴树科的蚬木(Excentrodendron hsienmu)、大戟科的肥牛树(Cephalomappa sinensis)、大风子科的海南大风子(Hydnocarpus hainanensis)、梧桐科的苹婆(Sterculia monosperma)等为主(王献溥, 1984; 朱华, 2011)。热带季节性雨林能够在热带边缘出现和维持, 根本原因就是热带季风席裹而来的温热水汽的强大作用。热带季风带来的湿热水汽在热带北缘地区的部分地点如河谷两侧和矮山沟谷能够维持长期的良好水热条件, 保证了热带物种在这些地方的长期续存。西藏东南部的雅鲁藏布江在29° N的地方出现了呈南北走向、海拔差达5 000 m左右的深切峡谷, 成为南亚季风向北输送水汽与热量的天然通道(孙航和周浙昆, 1996)。雅鲁藏布江峡谷年降水量达600-800 mm, 是我国第二大降水中心(冯蕾和周天军, 2015)。这里分布的热带植物以泛热带分布和热带亚洲分布最多, 是印度-马来植物区系在南亚季风作用下沿雅鲁藏布江河谷北进的结果(孙航和周浙昆, 1996)。雅鲁藏布江中下游河谷(墨脱县背崩乡及以南)的建群乔木大多是热带起源成分, 如龙脑香科的婆罗双(张金泉和王兰州, 1985)、金缕梅科的细青皮(Altingia excelsa)和使君子科的千果榄仁(Terminalia myriocarpa)(刘冰, 2016)、金虎尾科的风筝果(Hiptage benghalensis)(任明迅, 未发表数据), 板根现象、老茎生花、附生植物、云雾林或苔藓林等现象也较典型。孙航等(1997)将雅鲁藏布江中下游河谷植被称之为“热带低山半常绿雨林”, 本文将之归为“热带季节性雨林”。
在云南东南部的文山古林箐(刘颖颖和朱华, 2014)、广西西南部的弄岗(吴春林, 1991; 吴望辉, 2011)、海南岛西部的俄贤岭与尖峰岭(李意德等, 2006; 秦新生等, 2014)等地, 山峰中下部及山脚沟谷分布着常绿的“沟谷雨林”和“山地雨林”。这些以热带植物为主的植被虽然有着一定的干湿季节更替, 但植被外貌基本常绿, 优势种与建群种也是亚洲热带雨林特征成分——龙脑香科植物, 如青皮(Vatica mangachapoi)和坡垒(Hopea hainanensis)等(李意德等, 2006; 张永夏等, 2007; 李丹, 2016), 应属“热带季节性雨林”。在海南岛东侧的西北太平洋季风的迎风面沿海一带的万宁市石梅湾就分布着一片绵延约16 km、宽约400 m的青皮林单优自然群落(张永夏等, 2007; 李丹, 2016), 便是这类热带季节性雨林的典型代表。
2.2 季风与热带季雨林
热带季雨林或称“季风雨林”, 是在干湿季节交替的热带季风条件下形成的一种地带性密闭型森林植被, 其突出的特点是建群种为旱季落叶的热带性阔叶树、生长期存在一个明显的落叶期(朱华, 2011)。热带季雨林是热带季风气候影响植被变化的一个最典型的反映, 普遍分布于印度半岛、中南半岛与东南亚岛屿等受到热带季风深刻影响的地区, 通常分布在海拔1 000 m以下的河流两岸山峰中上部(王献溥, 1984; 朱华, 2011)。热带季雨林被认为是热带雨林(季节性雨林)向热带稀树草原植被过渡的一种特殊植被类型(朱华, 2011)。热带季雨林的群落结构较简单, 乔木多样性低, 但仍以热带属性植物为绝对优势。热带季雨林群落具有明显的季节性变化, 旱季上层乔木多数落叶, 林冠稀疏, 下层草本枯黄; 雨季的时候林冠浓密, 整个季相转为绿色(Elliott et al., 2006; 朱华, 2011)。位于滇黔桂交界区的红水河流域的部分沟谷以木棉(Bombax ceiba)、短穗鱼尾葵(Caryota mitis),以及大型木质藤本植物大喙省藤(Calamus macrorrhynchus)、短柄垂子买麻藤(Gnetum pendulum f. intermedium)、风筝果等组成的旱季落叶植被, 应属较典型的热带季雨林(朱华, 2011; Ren, 2015)。
在东南亚以及中国云南南部、广西西南部及海南岛等地的石灰岩山峰中上部, 还发育着一类特殊的季雨林(吴春林, 1991; 朱华等, 1996; 秦新生等, 2014)。石灰岩山峰海拔较高的地方由于石灰岩漏水特性导致干湿变动极其剧烈(朱华等, 1996), 再加上土层薄、含钙量极大等特点, 发育出了喜钙耐旱植物为主的特殊季雨林(朱华等, 1996; 吴望辉, 2011; 秦新生等, 2014)。位于这些石灰岩山峰中部及下部的不存在明显落叶期的常绿或半常绿森林, 《中国植被》和《云南植被》称之为“石灰山常绿季雨林” (吴春林, 1991), 朱华(2011)则称之为“热带季节性湿润林(tropical seasonal moist forest)”, 本文将之归入前面论述的“热带季节性雨林”。
2.3 季风与中国西南干旱河谷
中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(Wang, 2002; Salinger et al., 2014; 图1)。这里的地形非常复杂, 既包括了横断山区、云贵高原等崇山峻岭, 也有着怒江、澜沧江、南盘江、红河等大河深谷。当三大夏季风共同作用于这块复杂地形时, 便造就了有着“焚风效应”的局域“干旱河谷” (张荣祖, 1992; 金振洲和欧晓昆, 2000; 金振洲, 2002; 刘晔等, 2016)。干旱河谷的主要特征是气温年较差小、干湿季分明(雨季短、旱季长而炎热)(金振洲, 2002; 刘晔等, 2016)。由于纵向岭谷对季风的水热再分配作用(吴绍洪等, 2012), 干旱河谷的气温垂直递减, 降水量随海拔的升高而增长(张荣祖, 1992)。干旱河谷底部植被多为耐旱的稀疏灌丛, 河谷中上部植被变为喜湿热的大冠幅乔木, 有时发展出热带季雨林(朱华, 2011; 刘晔等, 2016)。根据热量的差异, 干旱河谷可以进一步分为干热、干暖和干温河谷三大类型(刘晔等, 2016)。
干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(Bourliere, 1983; 张荣祖, 1992; 金振洲, 2002)。干旱河谷的植被类型复杂多样, 植物特有种比率可高达38% (欧晓昆和金振洲, 1996)。其中, 干热河谷的温度最高, 以热带性质的稀树草原、稀树肉质灌丛植被为主; 干暖河谷植被则以暖性落叶阔叶灌丛、常绿硬叶林、常绿阔叶灌丛为优势类型(张荣祖, 1992; 金振洲和欧晓昆, 2000) ; 暖性针叶林、落叶阔叶林则主要在干温河谷环境占优势(刘晔等, 2016)。这些不同类型的局域植被很有规律地出现在同一河谷季风影响程度不同的上、下游地段, 再次证实了季风对植物分布与局域植被形成的重大影响。
2.4 季风与海南岛西部的稀树草原
海南岛西部从洋浦、昌江直到莺歌海的沿海平地、河流三角洲及部分沙地上, 分布着一片长约170 km、宽3-20 km的沙漠化土地, 这里曾经是我国唯一的热带稀树草原分布区(郑影华等, 2009; 欧先交等, 2013)。海南岛的稀树草原受热带季风气候控制, 雨季和旱季明显, 植被是以热带型的旱生或中生多年生禾草类为主的草本植物群落, 以华三芒草(Aristida chinensis)、丈野古草(Arundinella decempedalia)、艾纳香(Blumea balsamifera)、黄花稔(Sida acuta)等耐旱草本为主。混生灌木有火索麻(Helicteres isora.)、刺篱木(Flacourtia indica)、银柴(Aporosa chinensis)等, 散生乔木有木棉(Bombax malabaricum)和厚皮树(Lannea coromandelica)等, 呈现出独特的群落结构和生态外貌(林培松, 2004; 欧先交等, 2013)。处于热带季风气候区的海南岛缘何在其西部滨海地带出现较大面积的热带稀树草原, 引起了较大的争议, 公认的最主要原因是季风气候(郑影华等, 2009; 欧先交等, 2013)。首先, 海南岛中南部呈东北到西南走向的鹦哥岭与雅加大岭阻断了东面来自西北太平洋季风的暖湿气流, 导致山脉东侧降水丰沛、西侧的洋浦到昌江一带降水稀少(林培松, 2004; 郑影华等, 2009)。其次, 海南岛西侧隔海相望的中南半岛分布着近似南北走向的长山山脉, 这条平均海拔达2 000 m、延绵约1 000 km的山脉阻隔了南亚季风, 导致从西面抵达海南岛的南亚季风水汽含量少; 再加上北部湾冷水流的终年影响, 海南岛西部出现热带半干旱气候(林培松, 2004)。再者, 海南岛西侧每年11月到次年4月, 既是旱季, 又正处风季, 而且该区域分布有海南岛最集中分布的石灰岩地貌(昌江王下到俄贤岭一带), 地表储水力极差, 导致旱季出现尤为严酷的干热生境, 抑制了木本植物生长, 最终形成了稀树草原(林培松, 2004; 欧先交等, 2013)。近10年来, 由于人工造林与旅游开发, 海南岛西部的热带稀树草原大部分已经消失(林培松, 2004; 任明迅, 个人观察)。
3 季风对热带植物北向扩散和物种分化的作用
亚洲的热带边缘以及三大夏季风的叠加影响区, 正是高山大河与石灰岩地貌集中分布的中国西南及中南半岛一带(图1)。由于受到不同方向、不同季节夏季风的共同影响, 这里的热带季风气候更明显、时间更长, 维持了可以满足热带植物生长与繁衍的基本水热条件。这里复杂的地形地貌以及不同植被类型形成的交错区(ecotone), 也为植物提供了丰富多样的生态位。因此, 中国西南及中南半岛是热带植物适应分化, 产生大量特有物种的一个热点地区, 可能是中国-缅甸生物多样性热点地区(Myers et al., 2000)形成与维持的一个主要原因。本文以4个典型热带植物类群的物种多样性与特有中心的地理分布情况为例, 分析季风如何在局域地形共同作用下影响热带植物的地理分布与物种分化。3.1 苦苣苔亚科
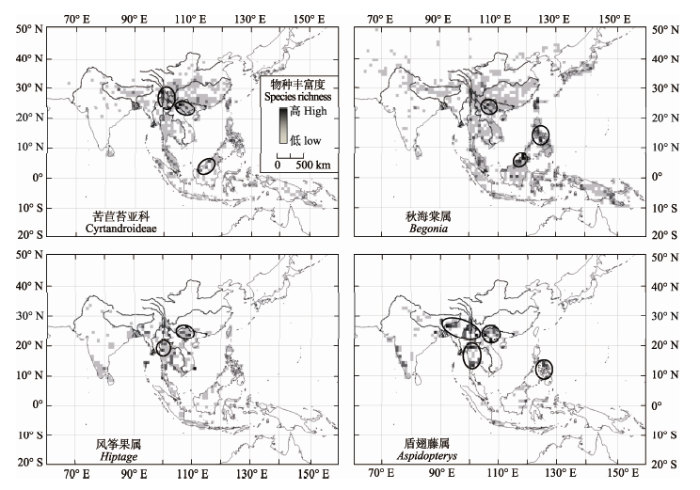
苦苣苔科(Gesneriaceae)可能于7000万年前起源于南美, 在约49 Ma扩散至亚洲的东南亚群岛(Perret et al., 2013), 逐渐演变成子房上位、子叶不等大、无胚乳的苦苣苔亚科(Cyrtandroideae)(李振宇和王印政, 2005)。现今的苦苣苔亚科约有86属1 700 种, 物种多样性中心位于中国西南特别是滇黔桂交界区的石灰岩地区(应俊生和张志松, 1984; 韦毅刚等, 2004; Perret et al., 2013; 图2)。以往的研究强调石灰岩生境的隔离效应对特有类群分化与维持的重要作用(韦毅刚等, 2004; 李振宇和王印政, 2005)。然而, 对于苦苣苔亚科这类典型的热带植物而言, 季风才是它们能够从典型热带地区扩散至热带北缘的中国西南地区并存活下来的先决条件(Ren, 2015)。为更准确地揭示苦苣苔亚科物种的分布与迁移规律, 我们从“全球生物多样性信息站” (http://www.gbif.org)收集了苦苣苔亚科所有的物种已知分布点信息, 按照经纬度1 × 1的精度统计物种数量, 利用软件DIVA-GIS7.5绘制物种多样性分布图。结果证实了苦苣苔亚科的物种丰富度中心并非亚洲典型热带地区和苦苣苔亚科最早分布地点东南亚群岛, 而是位于热带北缘地区的中国西南滇黔桂交界区、横断山区及泰国北部(图2)。这3个地区都位于热带季风叠加影响区(图1), 证实了热带季风对苦苣苔亚科向北扩散的强大作用。
可以推测, 苦苣苔亚科这类早期分布在热带赤道一带的植物, 在热带季风作用下向北扩散至中国西南及中南半岛之后, 受到高山深谷与石灰岩地貌的隔离作用, 这些位于分布区边缘的“先锋个体”与热带地区核心种群的基因流遭到阻断; 再加上局域高度异质性的地形造成的隔离作用, 导致扩散到这里的个体逐渐适应局域生境季节性干湿气候、石灰岩高钙土壤等因素, 慢慢分化出新种(韦毅刚等, 2004; Ren, 2015; 钱贞娜等, 2016)。这种在季风作用下的“扩散、阻断、隔离、分化”过程, 可能是苦苣苔亚科在中国西南与中南半岛一带出现物种丰富度中心与特有种分布中心的主要原因。
3.2 秋海棠属(Begonia)
秋海棠属广布于全球热带区域, 非洲约150种、美洲约600种、亚洲600种左右(Gu et al., 2007)。分子系统学的证据显示, 亚洲的秋海棠属是在上新世从非洲通过长距离扩散而来(de Wilde et al., 2011)。与苦苣苔科类似, 秋海棠属植物对石灰岩生境也有着较高的适应专一性(Chung et al., 2014)。亚洲的秋海棠属植物出现了3个物种分布中心: 加里曼岛北部、菲律宾中部、中国西南-中南半岛北部(图2)。其中, 尤以中国西南-中南半岛北部分布中心面积最大、特有属最多(Gu et al., 2007; Chung et al., 2014)。基于分子系统学的研究发现, 东亚季风气候的形成加速了中国西南-中南半岛北部一带石灰岩地层的侵蚀, 形成峰林、峰丛、洼地与洞穴交错的高度破碎化、异质性的喀斯特生境, 促进了这一地区秋海棠属的物种分化(Chung et al., 2014)。因此, 秋海棠属可能在中国西南-中南半岛一带经历了类似苦苣苔亚科的“扩散、阻断、隔离、分化”过程, 形成了如今的物种多样性分布中心。其中, 最关键的因素正是季风携带植物繁殖体的扩散作用和季风气候带来的强降雨侵蚀石灰岩导致的生境隔离作用。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2亚洲4个代表性典型热带植物类群的物种分布格局。黑色圆圈示物种丰富度与特有比例最高的区域。物种分布信息来自http://www.gbif.org, 分布图制作软件是DIVA-GIS7.5。
-->Fig. 2The distribution pattern of species richness and endemism of four typical tropical plant taxa in Asia. Black circles indicate regions with highest species richness and endemism. The species distribution information is obtained from http://www.gbif.org and the map was drawn using DIVA-GIS7.5.
-->
3.3 风筝果属(Hiptage)与盾翅藤属(Aspidopterys)
风筝果属和盾翅藤属都属于金虎尾科, 是热带亚洲特有的大型木质藤本。这两个属都有较为特殊的果实传播方式, 风筝果属果具3条果翅; 盾翅藤属果皮全部伸展成薄翅状附属物。翅果使得这两个属能够随着季风长距离迁移(Ren, 2015; 钱贞娜等, 2016)。风筝果属物种多样性与特有种分布中心主要有两个: 滇黔桂交界区的红水河流域、泰国北部的湄平河上游(图2; Ren, 2015), 两者都位于河流的河谷或河谷两侧山坡。盾翅藤属的物种分布中心也位于河流河谷上, 如红水河、红河、澜沧江、湄平河、布拉马普特拉河(雅鲁藏布江下游印度境内河段)等(图2)。这表明, 季风沿着河谷向北迁移促进热带植物繁殖体扩散, 并使之在河谷生境中得以生存与繁衍是导致热带植物沿河谷呈带状分布的关键因素。
综合以上4个典型热带植物类群的物种分布格局可以发现, 中国西南的滇黔桂交界区是这些属或亚科共同的物种丰富度中心(图2)。这可能是由于以下两个方面的原因: (1)滇黔桂交界区是三大夏季风的交汇区, 是很多植物扩散的交汇带; (2)滇黔桂交界区有着极其破碎化的石灰岩地貌, 形成了大量散布的隔离生境。那么, 为什么物种丰富度中心出现在滇黔桂交界区, 而非同为季风重叠区、同样有着大量岩溶地貌的云南南部或西南部呢?滇黔桂交界区的河流如右江、左江与红水河等水量小、河谷窄, 季风通过性较弱(Ren, 2015), 一旦热带植物类群抵达这些河流河谷及上游, 很难与下游的种群保持基因流, 容易导致隔离物种形成(Ren, 2015; 钱贞娜等, 2016)。云南西南部、南部和中南半岛的河流与山脉则多为南北走向, 澜沧江、怒江与红河等大江大河的河谷非常宽阔, 与中南半岛的纵向连通性较好, 使得南北方向的植物繁殖体扩散和基因流得以维持在一个较高水平, 物种不易分化(吴绍洪等, 2012)。
4 结语
东亚和东南亚地区一直被认为是现代被子植物的起源与分化中心(吴征镒等, 2005; Buerki et al., 2014), 热带季风可能在其中起着重要作用。季风气候不仅促使一部分热带植物的繁殖体得以向北迁移至热带地区北缘, 还通过强烈的降雨和剧烈的干湿交替加剧了局域生境的隔离程度, 与当地高山大河等隔离生境一起导致部分热带植物逐渐适应局域特殊地形与气候而分化出新的类群, 使得三大夏季风叠加影响区(中国西南的滇黔桂交界区和中南半岛)成为亚洲热带植物的物种多样性和特有种分布中心之一。随着全球气候变暖, 亚洲季风对热带植物向北迁移的作用可能更明显, 对东亚和中国植物多样性将产生更深远的影响。在今后的相关研究中, 应进一步重视季风气候的变动规律及其对热带植物长距离扩散与物种分化的作用, 重视我国西南和海南岛与中南半岛植物区系的联系, 更准确地理解这些区域生物多样性形成与维持机制和亚洲热带植物区系的演变趋势。The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | The spatial and temporal distribution of summer monsoon precipitation (or effective moisture) during the Holocene has been reconstructed on the basis of geological data, including lake levels, pollen profiles, and loess/paleosol records. In addition, the summer (July) precipitation increment, effective precipitation, and monsoon strength index have been obtained from numerical modeling experiments. Both geological data and numerical modeling indicate that the Holocene optimum, as defined by peak East Asian summer monsoon precipitation, was asynchronous in central and eastern China, reaching a maximum at different times in different regions, e.g., ca. 10,000–800002yr ago in northeastern China, 10,000–700002yr ago in north-central and northern east-central China, ca. 7000–500002yr ago in the middle and lower reaches of the Yangtze River, and ca. 300002yr ago in southern China. In southwestern China the maximum appeared ca. 11,00002yr ago, but probably was related to the maximum landward extension of the Indian summer monsoon. The regional shift in the maximum precipitation belt from northwest to southeast over the past 10,00002yr is interpreted as a response to changing seasonality related to orbital forcing of the climate. The southeastward shift of the East Asian summer monsoon maximum is consistent with the progressive weakening of the summer monsoon as the summer solar radiation anomaly decreased progressively through the Holocene and the East Asian monsoon index declined, while the early maximum in southwestern China matches the maximum of the Indian monsoon index. |
| [3] | ,. |
| [4] | Darwin described as an ‘abominable mystery’ the abrupt origin of angiosperms in the mid-Cretaceous and the high diversification rates in their early history. The father of evolutionary theory could not fathom this rapid diversification and rather invoked that ‘there was during long ages a small isolated continent in the S. hemisphere, which served as the birthplace of the higher plants’. In this essay, we comment on the spatial origin of angiosperms, but focus primarily on understanding the abiotic factors that promoted the early diversification of angiosperms by reviewing palaeobotanical, palaeogeographical, phylogenetics and biogeographical evidence. We argue that islands located in the region today occupied by South-East Asia played a major role in angiosperm diversification during the Late Jurassic and Early Cretaceous. 08 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2014, 174, 326–333. |
| [5] | The picturesque limestone karsts across the Sino-Vietnamese border are renowned biodiversity hotspot, distinguished for extremely high endemism of calciphilous plants restricted to caves and cave-like |
| [6] | |
| [7] | Aim Most deciduous species of dry monsoon forests in Thailand and India form new leaves 1–2 months before the first monsoon rains, during the hottest and driest part of the year around the spring equinox. Here we identify the proximate causes of this characteristic and counterintuitive 'spring-flushing' of monsoon forest trees. Location Trees of 20 species were observed in semi-deciduous dry monsoon forests of northern Thailand with a 5–6-month-long severe dry season and annual rainfall of 800–1500 mm. They were growing on dry ridges (dipterocarp–oak forest) or in moist gullies (mixed deciduous–evergreen forest) at 680–750 m altitude near Chiang Mai and in a dry lowland stand of Shorea siamensis in Uthai Thani province. Methods Two novel methods were developed to analyse temporal and spatial variation in vegetative dry-season phenology indicative of differences in root access to subsoil water reserves. Results Evergreen and leaf exchanging species at cool, moist sites leafed soon after partial leaf shedding in January–February. Drought-resistant dipterocarp species were evergreen at moist sites, deciduous at dry sites, and trees leafed soon after leaf shedding whenever subsoil water was available. Synchronous spring flushing of deciduous species around the spring equinox, as induced by increasing daylength, was common in Thailand's dipterocarp–oak forest and appears to be prevalent in Indian dry monsoon forests of the Deccan peninsula with its deep, water-storing soils. Main conclusions In all observed species leafing during the dry season relied on subsoil water reserves, which buffer trees against prolonged climatic drought. Implicitly, rainfall periodicity, i.e. climate, is not the principal determinant of vegetative tree phenology. The establishment of new foliage before the summer rains is likely to optimize photosynthetic gain in dry monsoon forests with a relatively short, wet growing season. |
| [8] | . 本文使用MRI模式在不同分辨率下(180km、120km、 60 km、20 km)的AMIP试验结果,分析了该模式对青藏高原夏季降水及水汽输送通量的模拟,并考察模式分辨率的影响.结果表明:MRI模式能够较为合理地模拟出青 藏高原夏季气候平均的降水空间分布,但对气候平均水汽输送通量以及降水年际变化的模拟却存在较大的误差.随着分辨率的提高,该模式对青藏高原气候平均降水 的模拟有明显改进,包括降水年循环以及夏季降水的空间分布等.分辨率为180km、120km、60 km、20 km的MRI模式模拟的青藏高原7月平均降水绝对误差分别为2.2 mm/d、1.2 mm/d、0.7 mm/d、0.2 mm/d.另外,高分辨率模式模拟的青藏高原夏季水汽输送通量的年际变化也更接近观测.当分辨率达到20 km时,MRI模式模拟的西风水汽输送指数与观测的相关系数达到0.43,通过了0.1显著性水平的显著性检验.但MRI模式对青藏高原夏季降水的年际变 化以及气候平均水汽输送通量的模拟技巧并不随分辨率的增加有明显提高.低分辨率模式中模拟降水量偏大、印度季风槽偏强的现象在高分辨率模式中仍然存在. |
| [9] | . |
| [10] | |
| [11] | |
| [12] | |
| [13] | . . 龙脑香科(Dipterocarpaceae)植物为亚洲热带雨林的代表科,成为亚洲热带雨林的表征科。海南岛地处中国的最南端,以热带季风气候为主。热带雨林是最复杂的生态系统,拥有非常丰富的植物资源,在海南分布有青梅(Vatica mangachapoi)、坡垒(Hopea hainanensis)和无翼坡垒(Hopea exalata)3个龙脑香科(Dipterocarpaceae)植物,为海南热带低地雨林的重要组成部分,在海南整个生态系统中起到重要的调节和指示作用。近些年来,海南自然保护区的建立和扩大为保护植物的生存环境提供了一定程度的保障,但是,非保护区的野生植物尤其是珍稀濒危植物的生存依旧受到严重威胁。主要原因还是人为干扰严重,破坏了植物的生长发育环境,阻碍了种群的发展。为了探讨目前整个海南岛生态环境的保护现状及存在的问题,本文以龙脑香科植物为研究对象,采用“样方法”完成野外数据调查,了解海南岛的龙脑香科植物资源分布情况,然后通过“空间分布格局”及“龄级结构划分”、“静态生命表”、“存活曲线”、“种群动态量化”等方法对比分析非自然保护区和自然保护区两种不同环境下龙脑香科植物种群的空间分布格局和种群特征,研究结果表明:(1)对比非自然保护区的环境,在自然保护区内龙脑香科植物分布点较多,种群数量较大,海拔梯度更宽,植株生长发育状况更好,说明自然保护区的环境更适合龙脑香科植物的生长和发育。无翼坡垒仅出现在一个保护区内,情况特殊,为分布区夹窄种群,其原因有待进一步研究。青梅种群在自然保护区内可调查到113个点,频率为0.496,非自然保护区可调查到51个点,频率为0.584,为海南广布种群,目前种群数量还相对较多,但坡垒种群在自然保护区内仅调查到27个点,频率为0.259,在非自然保护区3个点,且频率仅有0.333,坡垒已经处于极度濒危。(2)无论在保护区内外,青梅、无翼坡垒种群均为集群分布,支持濒危植物基本呈现集群分布的结论。从各龄级的种群分布格局动态变化进行分析,随着龄级的增加,种群均表现出扩散的趋势,这是种群适应环境完成自我更新的必然过程。对比非自然保护区,自然保护区内的青梅种群和坡垒种群进入均匀分布的龄级更大,说明自然保护区对种群的保护力度更强,种群自我更新能力更强。(3)尽管青梅、坡垒和无翼坡垒3个种群在分布区域上有较大的差异,种群大小、濒危程度与原因也不一样,但在植物种群的径级结构、静态生命表和存活曲线等3个方面的研究结果表明,无论非自然保护区还是自然保护区的青梅种群接近DeeveyⅡ型,坡垒种群DeeveyⅢ型,而甘什岭自然保护区的无翼坡垒种群接近DeeveyⅢ型。由植物种群动态指数对种群进一步对比分析,发现无论非自然保护区还是自然保护区,龙脑香科植物种群各龄级虽然均有不同程度的波动,但是种群整体表现为增长型,而且Vpi,趋近于0,说明所发现的龙脑香科植物,其种群基本趋于稳定。但是,对比非自然保护区,自然保护区内的青梅种群和坡垒种群的Vpi,数值更接近于0,说明自然保护区内的种群结构更加稳定。通过种群时间序列预测未来龙脑香科种群结构,与上述结果一致,总体表现为非自然保护区的物种受威胁程度明显强于自然保护区。综上结果,对比分析非自然保护区与自然保护区的植物种群分布情况和种群特征,发现自然保护区内植物种群生长发育、自我更新情况均优于非自然保护区,说明建立自然保护区可以为植物创造更有力的生存环境,保证植物种群的演替发展。但是野生植物处境依旧不乐观,面临灭绝的危险,需要加强保护力度,改善保护措施。 |
| [14] | In this paper we review the observed structure and evolution characteristics of the western North Pacific monsoon on various time scales, including its annual cycle, synoptic wave activity, intraseasonal oscillations, and interannual variabilities. On the synoptic (2-10-day) timescale, summertime synoptic waves and equatorial symmetric and anti-symmetric modes are often observed, and they may be responsible for triggering tropical cyclone genesis. On the intraseasonal scale, there are significant spectrum peaks at bi-weekly (10-20-day) and lower-frequency (20-70-day) bands. On the interannual time scale, the monsoon is greatly modulated by and possibly feeds back to the El Nio-Southern Oscillation (ENSO). The paper reviews our current understanding of physical mechanisms that give rise to the synoptic-scale, intraseasonal and interannual variabilities, and multi- scale interactions among these motions. The comparison between the Indian monsoon and the western North Pacific monsoons in terms of their differences in precipitation and circulation patterns, dominant time scales, and global teleconnection is also illustrated. Finally we discuss some remaining issues related to the western North Pacific monsoon variabilities. |
| [15] | . Studies have shown that large-scale monsoon gyre activity is closely associated with tropical cyclogenesis over the western North Pacific.In this study,two cases of monsoon gyre activities in 2002 and 2009 were first examined.It was found that a monsoon gyre can be linked to the formation of one or more tropical cyclones,which usually occur near or to the east of the gyre center.Further analysis of the monsoon gyre activity during the period of 2000 2009 indicates that tropical cyclogenesis mainly occurs near or to the east of the gyre center,although the definition of a monsoon gyre depends on its duration and the circulation intensity.It is suggested that the tropical cyclogensis may be associated with the Rossby wave energy dispersion of monsoon gyres. |
| [16] | . <p>三分区位于尖峰岭国家级自然保护区,目前区内有海南岛保存最好的青皮林。根据1.38hm<sup>2</sup>的样地资料,林分平均蓄积量高达469.773m<sup>3</sup>·hm<sup>-2</sup>;平均胸径28.6cm,最大胸径143.3cm;林木组成复杂,记载种类300多种。青皮为热带常绿季雨林的表征种和建群种,重要值为16.8,该种群的径级和年龄结构合理,具有反J曲线的正常增长型种群。聚类分析结果表明:属于典型热带常绿季雨林类型的青皮林群落分布在海拔300~650m处,伴生树种有白茶、细子龙、野生荔枝等。调查区内有8种国家重点保护植物,其中坡垒为国家一级保护植物,青皮是关键种,该区的森林生态系统具有极高的保护价值。</p> |
| [17] | |
| [18] | . . 海南岛西部是中国唯一的热带稀树干草原沙漠化地区,是热带沙漠化土地类型、沙漠化过程十分典型的区域.该研究以RS与GIS为支持,在研究区土地沙漠化现状野外调查的基础上,根据土地沙漠化分类原则、监测对象和遥感图像的分辨率,建立了海南岛西部沙漠化土地分类分级指标体系和相应的沙漠化土地解译标志.参考其他研究资料,利用Arc/View软件对海南岛西部1986年、2003年TM遥感图像进行解译,获得海南岛西部土地沙漠化的两期矢量数据,并运用Arc/View地理分析程序对解译结果进行空间分析和数据处理.根据分析与处理的结果,对海南岛西部近20年来沙漠化土地面... |
| [19] | . |
| [20] | The interplay of orographic uplift and climatic changes in the Himalaya-Hengduan Mountains region (HHM) have had a key role in speciation and population demography. To gain further insight into these processes, we investigated their effects on Taxus wallichiana by combining molecular phylogeography and species distribution modeling.Molecular data were obtained from 43 populations of T. wallichiana. Nineteen climatic variables were analyzed alongside genetic discontinuities. Species distribution modeling was carried out to predict potential past distribution ranges.Two distinct lineages were identified, which diverged c. 4.2 (2.0–6.5) million years ago (Ma), a timescale that corresponds well with the recent uplift of the Qinghai-Tibet Plateau and subsequent climatic changes of the region. Correlations with climatic variables also suggest that ecological factors may have further reinforced the separation of the two lineages. Both lineages experienced population expansion during the last glaciation.The high genetic divergence, long-term isolation and ecological differentiation suggest a scenario of cryptic speciation in T. wallichiana associated with geological and climatic changes in the HHM. Our findings also challenge the notion of general population ‘contraction’ during the last glaciation in the HHM. |
| [21] | . 中国西南干旱河谷植被是我国西南横断山区特有的植被类型, 目前关于西南干旱河谷植被还没有整体性的群落类型划分研究。根据对甘肃、四川、云南三省九条主要河流的干旱河谷段野外调查和文献来源的1,339个植物群落样方数据, 采用自适应仿射传播聚类方法, 对我国西南干旱河谷的植物群落进行数量分类, 并采用典范对应分析方法进行排序分析。结果表明: (1)调查样方的植物群落分为7个植被型(稀树草原、肉质灌丛、常绿阔叶灌丛、暖性落叶阔叶灌丛、常绿硬叶林、落叶阔叶林和暖性针叶林), 24个群系, 31个群丛类型。暖性落叶阔叶灌丛是本植被区的代表性植被类型; 分布最广的群系为鞍叶羊蹄甲灌丛(Form. Bauhinia brachycarpa, 样方比例50.9%)、黄茅灌草丛(Form. Heteropogon contortus, 样方比例11.9%)、孔颖草灌草丛(Form. Bothriochloa pertusa, 样方比例5.6%)、黄荆灌丛(Form. Vitex negundo, 样方比例4.2%)、知风草灌草丛(Form. Eragrostis ferruginea, 样方比例3.8%)、车桑子灌丛(Form. Dodonaea viscosa, 样方比例3.4%)、云南松疏林(Form. Pinus yunnanensis, 样方比例3.3%)。(2)冬季低温和降水的季节性是限制干旱河谷植物群落分布的主要气候因子。稀树草原、肉质灌丛是典型的干热河谷植被类型; 暖性落叶阔叶灌丛、常绿硬叶林、常绿阔叶灌丛是干暖河谷植被的优势类型; 暖性针叶林、落叶阔叶林则主要在干温河谷环境占优势。 |
| [22] | . |
| [23] | Conservationists are far from able to assist all species under threat, if only for lack of funding. This places a premium on priorities: how can we support the most species at the least cost? One way is to identify 'biodiversity hotspots' where exceptional concentrations of endemic species are undergoing exceptional loss of habitat. As many as 44% of all species of vascular plants and 35% of all species in four vertebrate groups are confined to 25 hotspots comprising only 1.4% of the land surface of the Earth. This opens the way for a 'silver bullet' strategy on the part of conservation planners, focusing on these hotspots in proportion to their share of the world's species at risk. |
| [24] | . |
| [25] | . |
| [26] | Gesneriaceae are represented in the New World (NW) by a major clade (c. 1000 species) currently recognized as subfamily Gesnerioideae. Radiation of this group occurred in all biomes of tropical America and was accompanied by extensive phenotypic and ecological diversification. Here we performed phylogenetic analyses using DNA sequences from three plastid loci to reconstruct the evolutionary history of Gesnerioideae and to investigate its relationship with other lineages of Gesneriaceae and Lamiales. Our molecular data confirm the inclusion of the South Pacific Coronanthereae and the Old World (OW) monotypic genus Titanotrichum in Gesnerioideae and the sister-group relationship of this subfamily to the rest of the OW Gesneriaceae. Calceolariaceae and the NW genera Peltanthera and Sanango appeared successively sister to Gesneriaceae, whereas Cubitanthus, which has been previously assigned to Gesneriaceae, is shown to be related to Linderniaceae. Based on molecular dating and biogeographical reconstruction analyses, we suggest that ancestors of Gesneriaceae originated in South America during the Late Cretaceous. Distribution of Gesneriaceae in the Palaeotropics and Australasia was inferred as resulting from two independent long-distance dispersals during the Eocene and Oligocene, respectively. In a short time span starting at 3465Mya, ancestors of Gesnerioideae colonized several Neotropical regions including the tropical Andes, Brazilian Atlantic forest, cerrado, Central America and the West Indies. Subsequent diversification within these areas occurred largely in situ and was particularly extensive in the mountainous systems of the Andes, Central America and the Brazilian Atlantic forest. Only two radiations account for 90% of the diversity of Gesneriaceae in the Brazilian Atlantic forest, whereas half of the species richness in the northern Andes and Central America originated during the last 10 Myr from a single radiation. 08 2012 The Linnean Society of London, Botanical Journal of the Linnean Society, 2012, ●●, ●●–●●. |
| [27] | . |
| [28] | . 【Objective】The purpose of this study was to reveal the floristic composition and characteristics of seed plants in limestone areas in Hainan.【Method】Extensive field work, specimens collection and identification were conducted to collect data for the analysis.【Result and conclusion】A total of 1 176 species, 58 varieties and 1 forma belonging to 645 genera and 142 families were recorded. The tropical and subtropical distribution elements, especially tropical Asia ones, were dominant in the limestone flora in Hainan. The tropical and subtropical distribution families accounted for 73.24% of the total and the genera accounted for 87.75%. The proportions of temperate distribution families and genera accounted for 11.97% and 7.29%, respectively. The limestone areas contributed about 27.26% to the total number of species with less than 2% land area of the whole island. However, many genera include only one or very few species. There are 113 single and few species families, accounting for 79.58% of the total families. There are 2 genera endemic to China. |
| [29] | . The biodiversity hotspot at the Guizhou–Yunnan–Guangxi borders is a distribution centre of tropical plants in China. It spans the whole upper reaches of Zhujiang River, the largest river in Southern China. In this paper, I aimed to explore the roles of the river in the spread and diversification of tropical plants in this area, using the Asia-endemic genus Hiptage Gaertn. (Malpighiaceae) as an example. Two diversity and endemism centres of Hiptage area recognized: Indo-China Peninsula and upper reaches of Zhujiang River (UZJ). The area-adjusted endemism index further indicates UZJ as the most important distribution region of endemic species since UZJ has a very small area (~210,000 km2) but six out of the total seven species are narrow endemics. UZJ is located at the northern edge of distribution ranges of Hiptage, which resulted mainly from the north-west–south-east river systems of UZJ promoting northward spreads of this tropical genus. The highly-fragmented limestone landscapes in this region may promote habitat isolation and tends to be the main driving factor for origins of these endemic species. Hiptage is also distinctive for its highly-specialized pollination system, mirror-image flowers, which probably facilitates species diversification via floral and pollination isolation. Other studies also found UZJ as a major diversification centre of the tropical plant families Gesneriaceae and Begoniaceae. Thereafter, it is concluded that UZJ is an “evolutionary front” of tropical plants in China, which contributes significantly to the origin and maintenance of the unique biodiversity in the area.El hotspot de biodiversidad en las fronteras de las provincias Guizhou-Yunnan- Guangxi es un centro de distribución de plantas tropicales en China. Se extiende por toda la cuenca alta del río Zhujiang, el mayor río del sur de China. En este artículo, se explora el papel del río en la propagación y la diversificación de las plantas tropicales en este área, usando el género endémico de Asia Hiptage Gaertn. (Malpighiaceae) como ejemplo. Se reconocen dos centros de diversidad y endemismo de Hiptage: la Península Indochina y el curso superior del río Zhujiang (UZJ). El índice de endemismo ajustado al área indica UZJ como la región más importante de distribución de especies endémicas, ya que, aunque UZJ tiene un área muy peque09a (~210.000 km2), seis de un total de siete especies son estrictamente endémicas. UZJ está situado en el extremo norte del área de distribución de Hiptage, lo que resultó principalmente de la disposición noroeste-sureste de los sistemas fluviales de UZJ, que facilitaron la expansión y diferenciación hacia el norte de este género tropical. Los paisajes de piedra caliza altamente fragmentados en esta región han contribuido al aislamiento de hábitat y pueden ser el principal factor para el origen de estas especies endémicas. Hiptage también se distingue por su sistema de polinización altamente especializado, con flores de imagen especular, lo que probablemente facilita la diversificación de las especies a través del aislamiento de la polinización. Otros estudios también encontraron que UZJ es un importante centro de diversificación de las familias de plantas tropicales Begoniaceae y Gesneriaceae. Por consiguiente, se concluye que UZJ es un 00frente evolutivo03 de plantas tropicales en China, lo que contribuye de manera significativa al origen y mantenimiento de la biodiversidad única en la zona中国西南的云南-贵州-广西交界区是全球性的生物多样性热点地区,也是中国热带植物 的分布中心。这一区域横跨南中国最大河流—珠江的整个上游地区。这里,我试图以典型的热带植物类群金 虎尾科 (Malpighiaceae) 亚洲特有的风筝果属(Hiptage Gaertn.)为例,从河流的作用来解释这一地区热带 植物的扩散与物种分化及其生物多样性热点地区的形成。风筝果属具有两个物种多样性与特有中心:一个位 于中南半岛南部,一个则是珠江上游地区。面积校正后的特有性指数进一步证实,珠江上游地区是风筝果属 最集中的特有种分布地区,其大约210,000 km2的面积上有7个种,其中6个为狭域分布的地方特有种。珠江上 游地区位于风筝果属整个分布区域的北缘;风筝果属植物能扩散到这一地区,主要得益于该地的河流走向基 本都呈西北-东南走向。西北-东南走向的河道可以促进东南亚暖湿气流北进,从而允许风筝果属等热带植物 得以扩散至珠江上游地区的红水河、南盘江以及右江等地。这些地方分布着高度破碎化的岩溶地貌,又进一 步促进了局部生境隔离与物种分化。此外,风筝果属还具有极其特化的传粉系统“镜像花柱”(mirror-image flowers),可能加速了繁殖隔离与物种分化。其它研究也发现,珠江上游地区是一些典型热带植物类群如苦 苣苔科和秋海棠科的物种多样化中心。因此,珠江上游地区(滇黔桂交界区)很可能就是中国热带植物的一 个“进化前沿”,对这一地区独特生物多样性的形成与维持具有重要作用 |
| [30] | |
| [31] | Dipterocarpus zhengae sp. nov. is described from the middle Miocene Fotan Group of Zhangpu county, Fujian Province, Southeast China on the basis of a fruit wing. Three kinds of venation of the calyx longer lobes (enlarged sepals) occur in the winged fruits of extant Dipterocarpus . The fossil species is referred to the kind having three primary veins and within that category is most similar to extant Dipterocarpus gracilis Blume in the size of the longer lobe as well as its venation. The occurrence of the fruit wing of Dipterocarpus , together with palynological evidence, indicates unequivocally that a tropical climate and tropical rain forest occurred in Zhangpu during the middle Miocene, warmer and more humid than at the present day. The palaeobiogeography of Dipterocarpus is reviewed. |
| [32] | . |
| [33] | . |
| [34] | . 我国最近发现的早中新世季风记录,对东亚季风系统何时形成又提出了疑问.现代季风系统的一个明显特征,在于它打乱了行星气候系统纬向分布的带状模式,因而地质记录中季风气候地理模式的出现,就应当意味着季风系统的建立.据此汇总了中国大陆125个地点所获得的古植物和岩性资料,揭示了两种完全不同的气候分带模式:早第三纪宽阔的干旱带由西向东横跨中国大陆,而晚第三纪至今的干旱带仅局限在中国西北部.渐新世/中新世交界时气候系统的大改组,就是现代东亚季风建立的证据.其后,季风系统在晚第三纪又经历了巨大变化,包括15~13 Ma前、8 Ma、3 Ma前干旱气候的加剧和季风系统的加强.所获得的新资料并不支持亚洲季风系统起源于约8 Ma前这一观点,而认为东亚大约是在晚渐新世向季风气候转变这一假设更为合适. |
| [35] | . |
| [36] | Abstract Karst in Vietnam covers an area of about 60,000 km 2 , i.e. 18 % of the surface of the country. The country has an annual average temperature of 24C, an annual average rainfall of 2300 mm and a relative humidity of about 90%. Karst in Vietnam is typified by peak cluster-depression landscapes ranging in elevation from 200 to over 2000 m. Tower and coastal karst landscapes also exit. Because of naturally favourable conditions, karst ecosystems are diverse and very rich. Higher plants (cormophytes) are abundant. They are represented by approximately 2000 species, 908 genera, 224 families, 86 orders and 7 phyla. They form a thick vegetation cover of evergreen tropical rainforest. Knowledge about lower plants is limited. The fauna is rich and diverse. Phyla such as Protozoa, Vermes, Mollusca and Arthropoda are yet ill known. Preliminary results show that the phylum Chordata is represented by 541 species from 80 families, 40 orders and 5 classes. There exist many precious and rare mammals, in particular some endemic species such as Trachypithecus poliocephalus, T. delacouri, Rhinopithecus avanculus, Rhinolophus rouxi, Seotoma dineties and Silurus cuephuongensis. The class Insecta has about 2000 species. The fast population growth, particularly in the mountainous areas of the country, triggers an increasing demand for land and therefore threatens the ecosystem. To obtain land for farming, people have cut, burned and destroyed natural forest cover; resulting in occurrence of hazards such as soil-loss, water-loss, flash floods, mud-rock flows, rock-falls, severe drought, water logging and changes of karstic aquifers etc. Poaching precious animals and illegal logging are increasing. In contrast to other natural systems, karst ecosystems cannot be reestablished once damaged. Living karst landscapes will become rocky desert ones without life. Conservation of karstic environmental systems in general and karstic ecosystems in particular should not be the sole vocation of scientists but also a duty and responsibility of authorities and people from all levels. A good example of a multidisciplinary approach to karst-related problems is the implementation of the Vietnamese-Belgian Karst Project (VBEKAP): ural development in the mountain karst area of NW Vietnam by sustainable water and land management and social learning: its conditions and facilitation . The aim of this project is to improve living conditions of local people and sustained protection and management of the karst environment and ecosystem. |
| [37] | |
| [38] | . |
| [39] | The Tropical Ocean-Global Atmosphere (TOGA) program sought to determine the predictability of the coupled ocean-atmosphere system. The World Climate Research Programme's (WCRP) Global Ocean-Atmosphere-Land System (GOALS) program seeks to explore predictability of the global climate system through investigation of the major planetary heat sources and sinks, and interactions between them. The Asian-Australian monsoon system, which undergoes aperiodic and high amplitude variations on intraseasonal, annual, biennial and interannual timescales is a major focus of GOALS. Empirical seasonal forecasts of the monsoon have been made with moderate success for over 100 years. More recent modeling efforts have not been successful. Even simulation of the mean structure of the Asian monsoon has proven elusive and the observed ENSO-monsoon relationships has been difficult to replicate. Divergence in simulation skill occurs between integrations by different models or between members of ensembles of the same model. This degree of spread is surprising given the relative success of empirical forecast techniques. Two possible explanations are presented: difficulty in modeling the monsoon regions and nonlinear error growth due to regional hydrodynamical instabilities. It is argued that the reconciliation of these explanations is imperative for prediction of the monsoon to be improved. To this end, a thorough description of observed monsoon variability and the physical processes that are thought to be important is presented. Prospects of improving prediction and some strategies that may help achieve improvement are discussed. |
| [40] | . 滇黔桂及其邻近地区是我国苦苣苔科植物的分布和特有中心,广西正 处于这个中心的位置上,种类十分丰富,共计有38属、166种(含种下等级,下同),属和种的分布区类型不太复杂,特有现象极为突出,其中仅产广西的特有 属有5个,特有种达81个.广西苦苣苔科植物区系与相邻的贵州、云南两省属的相似性系数较高,分别为75.76%和71.42%,但与相邻省份苦苣苔科植 物种的相似性系数却较低,从35.48%至6.49%不等.苦苣苔科植物在广西全境分布较广泛,但各地种类分布很不均衡.广西苦苣苔科植物的天然分布对基 质有较严格的专一性,种群植株数量一般较少,同一种类不同的居群间形态变异较大. |
| [41] | . Thirty samples of tropical limestone forests in Guangxi were classified into six formations using Two-Way Indicator Species Analysis (TWINSPAN) and these six formations constitute two vegetation types of higher class. The classification results are: Ⅰ. Valley seasonal rain forest:( 1 ) Form. Saraca chinensis + Sterculia nobilis ( 2 ) Form. Horsfieldia hainanensis +Dracontomelon duperreanum. Ⅱ.Limestone hill seasonal forest: (1)Form. Deutzianthus tonkinensis + Acrocarpus fraxinifolius var. guangxiensis( 2 )Form. Cleistanthus petolottii + Vitex annamensis ( 3 ) Form. Cephalomappa sinensis ( 4 ) Form. Cephalomappa sinensis + Burretiodendron hsienmu. The samples were also ordinated using DCA (Detrend Correspondence Analysis) . The first axis of ordination reflected water gradient from wet valley to dry hill. The second axis was shade gradient. |
| [42] | . |
| [43] | . Topographic pattern has important impacts on atmospheric circulation and regional climate.The special terrain of Longitudinal Range-Gorge Region(LRGR) in Southwest China has a "corridor-barrier" effect on monsoons circulation,which led to the redistribution of surface hydrothermal pattern and is the main driving force of the formation and evolution of the ecosystems.Previous studies showed that LRGR was influenced by the combined effect of the Indian monsoon and the Pacific monsoon,which meet at the Ailaoshan mountain ranges.The Ailao Mountains are an important geographical dividing line.In this study,based on proofs of the atmospheric circulation,moisture transport,regional climate,runoff,and plant stable oxygen isotopes compostion,we found that LRGR is affected mainly by the Indian monsoon.The Pacific monsoon only affect this area in August,and the influence is not significant.Under the terrain "corridor-barrier" effect,surface water vapor,precipitation and runoff in LRGR showed significant zonal differences and meridional extension;crowd circulation,moisture transport,regional climate and river runoff.The atmospheric precipitable water,precipitation and stream flow present an apparently different pattern between the west and east sides of Ailao Mountains.The environmental differences are the results of the hydrothermal redistribution by the special terrain,which are the "barrier" and "corridor" effects of the LRGR.These differences are not the results of geographical zonality,but the results of non-zonality.The "corridor-barrier" effects of the longitudial range-gorge on monsoons result in the spatial differences of the geographical elements and the associated ecological effects. |
| [44] | . 在相关文献资料整理、野外实地调查、标本采集及室内标本鉴定的基础上,系统地研究了弄岗国家级自然保护区的植物区系组成、性质以及特有现象;对本区的珍稀濒危植物进行了统计评估,并提出相应的的保护对策。本研究结果如下: 1.弄岗国家级自然保护区共有维管束植物1725种隶属于183科799属(包括种下等级),其中栽培植物64种,隶属于37科59属;归化植物22种,隶属于16科21属;野生维管束植物1639种,隶属于175科753属。 2.弄岗国家级自然保护区共有蕨类植物146种,隶属于29科49属,以水龙骨科、金星蕨科、凤尾蕨科、铁角蕨科、叉蕨科和卷柏科等为优势科,这几个科共含有92种,占本区蕨类植物总种数的63.01%。本区蕨类植物的优势属有凤尾蕨属、毛蕨属、卷柏属、石韦属、铁角蕨属、铁线蕨属、三叉蕨属、线蕨属、鳞盖蕨属和星蕨属,共含有80种,占本区蕨类植物总种数的54.79%。本区蕨类植物以热带、亚热带性质成分占优势。 3.弄岗国家级自然保护区野生裸子植物种类较为贫乏,仅有8种,隶属于4科5属,占本区野生维管束植物科、属、种的2.29%、0.66%、0.49%。 4.弄岗国家级自然保护区的被子植物共有1485种,隶属于142科699属,以大戟科、茜草科、蝶形花科、禾本科、菊科、兰科、桑科、樟科、荨麻科、爵床科、百合科、萝藦科、芸香科、鼠李科、葡萄科、苦苣苔科、马鞭草科、番荔枝科、夹竹桃科、蔷薇科、葫芦科、苏木科、木犀科和唇形科占优势,这些科所含有的属、种分别占本区野生种子植物总属数的56.11%,总种数的59.47%。占优势的属为榕属、素馨属、秋海棠属、爵床属、楼梯草属、铁线莲属、悬钩子属、唇柱苣苔属、山柑属、花椒属、野桐属、沿阶草属、紫金牛属、崖爬藤属、薹草属、木姜子属等。本区种子植物以热带、亚热带成分为主。 5.弄岗国家级自然保护区植物区系具有明显的热带性质,以热带、亚热带成分为主,其中又以泛热带分布成分最多。在蕨类植物中,除世界分布科外其余全为热带、亚热带分布科;热带、亚热带分布的属、种分别占本区蕨类植物属、种总数的73.47%、52.05%,与属、种温带分布成分的比例分别为9.00:1、2.92:1。在种子植物中,热带、亚热带分布的87科561属,分别占本区野生种子植物科、属总数的59.59%、79.69%,与科、属温带分布成分的比例分别为5.44:1、6.10:1。 6.本区多数科、属种类较为贫乏,以单种科(属)和寡种科(属)为主。蕨类植物单种科和寡种科(2-4种)共有20科,占蕨类植物总科数的68.97%;单种属和寡种属(2-4种)共有39属,占蕨类植物总属数的79.59%。种子植物单种科和寡种科(2-10种)共有102科,占本区野生种子植物总科数的69.86%;单种属和寡种属(2-5种)共有662属,占本区野生种子植物总属数的94.03%。 7.在弄岗国家级自然保护区的野外调查中发现新分类群8个,即近林下凤尾蕨、弄岗毛蕨、宁明唇柱苣苔、中越唇柱苣苔、弄岗细筒苣苔、弄岗虾脊兰、岩生沿阶草、弄岗秋海棠:中国新记录种1种,即中越鹤顶兰;广西新记录属1属,即密花藤属;广西新记录种10种。 8.弄岗国家级自然保护区特有植物较为丰富。本区东亚特有的科有猕猴桃科;种子植物中国特有的属有异裂菊属、异裂苣苔属、青檀属、栾树属、瘿椒树属、细筒苣苔属、石山苣苔属、箬竹属等8属;蕨类植物中国特有种有41种,隶属于14科23属,分别占本区蕨类植物科、属、种总数的48.28%、46.94%、28.08%;本区所分布的种子植物广西特有种共有91种,占本区野生种子植物总种数的6.10%,占广西特有种总数的10.34%。弄岗国家级自然保护区分布有众多的石灰岩特有植物,表明了本区具有明显的岩溶区系性质。 9.弄岗国家级自然保护区分布有珍稀濒危植物31种,隶属于24科27属,其中蕨类植物有5种,裸子植物3种,被子植物有23种。被1987年国家环境保护局和中国科学院植物研究所修订的中国稀有濒危植物名录(Ⅰ)收录的有21种,被1999年国家林业局和农业部公布的国家重点保护植物名录(第一批)收录的有19种(国家一级保护3种)。对本区的31种珍稀濒危植物及其他维管植物进行了初步的IUCN评估,建议将本区分布的171种植物列入国家保护植物名录。 |
| [45] | . 对中国植物区系中的239个特有属,分属67个科,进行了分析研究,列出了这些特有属在种子植物各个科的分布,现代地理分布范围.结果表明含特有属在10个以上的有5个科即:Gesneriaceae,Compositae,Labiatae,Cruiciferae,Umbelliferae;其中以Gesneriaceae居榜首(27属),Compositae位居第二(20属),Labiatae有12属,居第三.含2属的科有15个,含1属的科有30个;其中Ginkgaceae,Davidiaceae,Eucommiaceae,Acanthochlamydaceae组成了中国植物区系最具古老性、特有性和代表性的4个单型科.在此基础上,从特有属在被子植物八纲系统各个纲的分布特点,以及在各个科组成和系统关系及已有地质、化石历史和系统学,形态,分子证据论述了这些特有属的起源、系统关系及在植物地理上的关系.在裸子植物中,特有属最为丰富,几乎皆是地质历史上北极-第三纪成分的残遗,起源时间较早,可追溯到白垩纪或更早.被子植物中,中国特有属存在于八纲被子植物的所有纲中,几乎在现代被子植物各个演化阶段均有古老残遗的特有类群存在,同时也不乏新特有类群尤其是在演化的高级阶段的类群.从起源上看,被子植物的古特有属主要发生于晚白垩纪和早第三纪,地质历史上大都占有广阔的分布区;新特有属多发生在新第三纪以后.其源头主要是北极第三纪、古热带第三纪(冈瓦纳第三纪)和古地中海第三纪的奇妙结合,不少类群是就地起源的;特有性是在第三纪中晚期以后北半球气候变迁,迁移途径(如北大西洋陆桥和白令陆桥)中断后形成的,这一时期是我国特有属形成发展的起始标志. |
| [46] | Today, insular Southeast Asia is important for both its remarkably rich biodiversity and globally significant roles in atmospheric and oceanic circulation. Despite the fundamental importance of environmental history for diversity and conservation, there is little primary evidence concerning the nature of vegetation in north equatorial Southeast Asia during the Last Glacial Period (LGP). As a result, even the general distribution of vegetation during the Last Glacial Maximum is debated. Here we show, using the stable carbon isotope composition of ancient cave guano profiles, that there was a substantial forest contraction during the LGP on both peninsular Malaysia and Palawan, while rainforest was maintained in northern Borneo. These results directly support rainforest "refugia" hypotheses and provide evidence that environmental barriers likely reduced genetic mixing between Borneo and Sumatra flora and fauna. Moreover, it sheds light on possible early human dispersal events. |
| [47] | . |
| [48] | Pollen data from China for 6000 and 18,00014C yr BP were compiled and used to reconstruct palaeovegetation patterns, using complete taxon lists where possible and a biomization procedure that entailed the assignment of 645 pollen taxa to plant functional types. A set of 658 modern pollen samples spanning all biomes and regions provided a comprehensive test for this procedure and showed convincing agreement between reconstructed biomes and present natural vegetation types, both geographically and in terms of the elevation gradients in mountain regions of north-eastern and south-western China. The 600014C yr BP map confirms earlier studies in showing that the forest biomes in eastern China were systematically shifted northwards and extended westwards during the mid-Holocene. Tropical rain forest occurred on mainland China at sites characterized today by either tropical seasonal or broadleaved evergreen/warm mixed forest. Broadleaved evergreen/warm mixed forest occurred further north than today, and at higher elevation sites within the modern latitudinal range of this biome. The northern limit of temperate deciduous forest was shifted c. 800 km north relative to today. The 18,00014C yr BP map shows that steppe and even desert vegetation extended to the modern coast of eastern China at the last glacial maximum, replacing today's temperate deciduous forest. Tropical forests were excluded from China and broadleaved evergreen/warm mixed forest had retreated to tropical latitudes, while taiga extended southwards to c. 43. |
| [49] | . 龙脑香科(Dipterocarpaceae)为一泛热带分布的科。据我们研究统计全世界龙脑香科植物约有3个亚科,16属529种,间断分布于热带亚洲、非洲和美洲(图1)。其中92%的种类分布于热带亚洲,为热带亚洲植物区系中的一个优势种;8%的种类散布于热带非洲和热带美洲。龙脑香科是植物学家和热带林业工作者研究较为深入的类群。自林奈(Carolus Linna-eus)1737年建立瓦特香属(Vateria)到布卢姆( |
| [50] | |
| [51] | . 根据样方调查,对海南万宁青皮林群落的种类组成、外貌、结构特征和物种多样性进行分析.结果表明:(1)海南万宁青皮林自然保护区有维管植物137种,隶属于62科110属,其中热带性分布种达95.5%,群落属于热带雨林植被类型中的海岸单优青皮群落.(2)群落的优势种为青皮,外貌常绿,生活型以小高位芽为主(占35.04%),青皮种群表现为增长趋势.(3)群落的物种丰富度Magarlef指数为34.30,Shannon-Wienner指数为2.31,Simpson指数为0.99,均匀度Pielou指数为1.08.(4)群落各层次的丰富度表现为乔木层>灌木层>藤本植物>草本层,Simpson指数表现为灌木层>草本层>藤本植物>乔木层,Shannon-Wienner指数表现为灌木层>藤本植物>草本层>乔木层,均匀度表现为草本层>灌木层>藤本植物>乔木层,表明乔木层的物种丰富度最高,但分布最不均匀,优势种的优势地位突出. |
| [52] | . <P><FONT face=Verdana>海南岛西部是我国惟一的热带稀树干草原沙漠化地区,选取5期遥感数据,采用室内解译和野外检验修正的方法,获得海南岛西部不同时期土地沙漠化数据库。结果分析表明,海南岛西部土地沙漠化在时间上经历了快速发展—初步逆转—全面逆转—波动反复的变化过程;在空间上由沿岸带向昌化、莺歌海两重心收缩、逆转;同时,其发展/逆转过程以沙漠化土地与林地、耕地相互转化为主,伴随着沙漠化土地内部复杂程度间转化。总之,总体逆转、局部反复波动是近50 a海南岛西部土地沙漠化时空演变的基本趋势。</FONT></P> |
| [53] | . |
| [54] | . 在中国植物学文献中,对热带季雨林的解释和运用是不一致的,特别是易于把季雨林与热带雨林相 混淆。季雨林是在具有明显干、湿季变化的热带季风气候下发育的一种热带落叶森林植被,是介于热带雨林与热带稀树草原(savanna)之间的一个植被类 型。云南的热带季雨林在分布生境、生态外貌特征、植物种类组成和地理成分构成上,均与热带雨林有明显区别,季雨林主要分布在海拔1000m以下的几大河流 开阔河段两岸和河谷盆地,其群落结构相对简单,乔木一般仅有1至2层,上层树种在干季落叶或上层及下层树种在干季都落叶;在生活型组成上,季雨林的木质藤 本相对较少,大高位芽植物及地上芽植物很少,但地面芽植物很丰富,地下芽植物和一年生植物也相对丰富;在叶级和叶型特征上,季雨林植物的小叶和复叶比例相 对较高,分别占到24%和44%;在植物区系地理成分构成上,季雨林的热带分布属合计也占绝对优势,但以泛热带分布属的比例相对较高,约占到总属数的 30%,热带亚洲至热带非洲分布属的比例也较高,约占总属数的12%。季雨林的地理成分更为多样性,起源与发展历史也更复杂和古老。 |
| [55] | The distribution patterns of geographical elements of seed plants from 135 regional floras that cover southern China were used to reassess the extent and boundaries of the tropical zone. The areas for which tropical genera account for >0280% of the total genera in the flora are south of 22°30′N in southern and southeastern China, which corresponds closely to the northern boundary of the tropical monsoon forest and rain forest in southeastern China. The line at c. 22°30′N is therefore suggested to be the northern biogeographical boundary of the tropical zone in south and southeastern China. This line exceeds the northern boundary of marginal tropical climate, which implies that the tropical zone could have extended further north in the geological past than it does today. The study supports the suggestion from palaeoecological studies that tropical and subtropical broadleaved evergreen forests in eastern China shifted north during the mid-Holocene. It also shows that there are climatic and biogeographical disparities between southeastern and southwestern China due to their different topography and geology. |
| [56] | . 西双版纳石灰岩森林植物区系经调查有维管束植物153科,640属,1394种及变种,其中,种子植物占129科558属1269种及变种。种子植物的分布区类型组成是热带和主产热带的科占总科数的71.3%;热带分布属占总属数的90.1%;热带分布种超过总种数的90%。热带分布属中又以热带亚洲分布属最多,占总数的35.3%;热带分布种中则以热带亚洲分布及其变型的种类占总种数的64.5%为特点。这表明该石灰岩 |
1
2001
... 季风(monsoon)是由于太阳高度角变化引起海陆热力性质差异或气压带随季节移动导致盛行风向和降水随季节改变的现象(
Asynchronous holocene optimum of the East Asian monsoon.
1
2000
... 季风(monsoon)是由于太阳高度角变化引起海陆热力性质差异或气压带随季节移动导致盛行风向和降水随季节改变的现象(
Tropical Savannas (Ecosystems of the World). Elsevier
1
1983
... 干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(
Proto-South-East Asia as a trigger of early angiosperm diversification.
1
2014
... 东亚和东南亚地区一直被认为是现代被子植物的起源与分化中心(
Phylogenetic analyses of Begonia sect. Coelocentrum and allied limestone species of China shed light on the evolution of Sino-Vietnamese karst flora.
3
2014
... 秋海棠属广布于全球热带区域, 非洲约150种、美洲约600种、亚洲600种左右(
... 亚洲的秋海棠属植物出现了3个物种分布中心: 加里曼岛北部、菲律宾中部、中国西南-中南半岛北部(
... ).基于分子系统学的研究发现, 东亚季风气候的形成加速了中国西南-中南半岛北部一带石灰岩地层的侵蚀, 形成峰林、峰丛、洼地与洞穴交错的高度破碎化、异质性的喀斯特生境, 促进了这一地区秋海棠属的物种分化(
Pliocene intercontinental dispersal from Africa to Southeast Asia highlighted by the new species
1
2011
... 秋海棠属广布于全球热带区域, 非洲约150种、美洲约600种、亚洲600种左右(
Leaf flushing during the dry season: The paradox of Asian monsoon forests.
1
2006
... 热带季雨林被认为是热带雨林(季节性雨林)向热带稀树草原植被过渡的一种特殊植被类型(
高分辨率MRI模式对青藏高原夏季降水及水汽输送通量的模拟
1
2015
... 西藏东南部的雅鲁藏布江在29° N的地方出现了呈南北走向、海拔差达5 000 m左右的深切峡谷, 成为南亚季风向北输送水汽与热量的天然通道(
泛亚洲季风区: 定义, 降水主模态及其变异特征
4
2012
... 亚洲地区是全球季风气候最为典型的区域, 最主要的特征就是降水多发生在高温的夏季(
... 亚洲的热带季风包括南亚季风(影响南亚和东南亚)和西北太平洋季风(影响菲律宾、中国南海及华南沿海地区), 与东亚季风(主要影响中国长江流域、日本和韩国等地)同为夏季风(
... 南亚季风的成因既有印度洋与印度半岛的海陆热力性质差异的驱动, 也受东南亚及大洋洲一带气压带在夏季北移的影响(
... ;
2
... 秋海棠属广布于全球热带区域, 非洲约150种、美洲约600种、亚洲600种左右(
... 亚洲的秋海棠属植物出现了3个物种分布中心: 加里曼岛北部、菲律宾中部、中国西南-中南半岛北部(
3
... 中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(
... 干旱河谷的主要特征是气温年较差小、干湿季分明(雨季短、旱季长而炎热)(
... 干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(
2
... 中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(
... 干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(
海南省野生龙脑香科植物分布格局及种群特征研究
2
2016
... 在云南东南部的文山古林箐(
... ;
A review on the western North Pacific monsoon: Synoptic-to-interannual variabilities.
1
2005
... 西北太平洋季风发生在每年的5-9月, 发源于太平洋西部和新几内亚东岸150° E附近区域的一条跨越赤道的气流通道(
季风涡旋影响西北太平洋台风生成初步分析
1
2014
... 西北太平洋季风发生在每年的5-9月, 发源于太平洋西部和新几内亚东岸150° E附近区域的一条跨越赤道的气流通道(
海南尖峰岭国家级保护区青皮林资源与乔木层群落学特征
2
2006
... 在云南东南部的文山古林箐(
... )等(
2
... 苦苣苔科(Gesneriaceae)可能于7000万年前起源于南美, 在约49 Ma扩散至亚洲的东南亚群岛(
... ;
近20年来海南岛西部土地沙漠化与气候变化关联度的研究
5
2004
... 海南岛西部从洋浦、昌江直到莺歌海的沿海平地、河流三角洲及部分沙地上, 分布着一片长约170 km、宽3-20 km的沙漠化土地, 这里曾经是我国唯一的热带稀树草原分布区(
... 处于热带季风气候区的海南岛缘何在其西部滨海地带出现较大面积的热带稀树草原, 引起了较大的争议, 公认的最主要原因是季风气候(
... ).其次, 海南岛西侧隔海相望的中南半岛分布着近似南北走向的长山山脉, 这条平均海拔达2 000 m、延绵约1 000 km的山脉阻隔了南亚季风, 导致从西面抵达海南岛的南亚季风水汽含量少; 再加上北部湾冷水流的终年影响, 海南岛西部出现热带半干旱气候(
... ).再者, 海南岛西侧每年11月到次年4月, 既是旱季, 又正处风季, 而且该区域分布有海南岛最集中分布的石灰岩地貌(昌江王下到俄贤岭一带), 地表储水力极差, 导致旱季出现尤为严酷的干热生境, 抑制了木本植物生长, 最终形成了稀树草原(
... ).近10年来, 由于人工造林与旅游开发, 海南岛西部的热带稀树草原大部分已经消失(
墨脱海拔600米到4000米的美丽植物
1
2016
... 西藏东南部的雅鲁藏布江在29° N的地方出现了呈南北走向、海拔差达5 000 m左右的深切峡谷, 成为南亚季风向北输送水汽与热量的天然通道(
Geological and ecological factors drive cryptic speciation of yews in a biodiversity hotspot.
1
2013
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
中国西南干旱河谷植物群落的数量分类和排序分析
5
2016
... 中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(
... 干旱河谷的主要特征是气温年较差小、干湿季分明(雨季短、旱季长而炎热)(
... ;
... ).根据热量的差异, 干旱河谷可以进一步分为干热、干暖和干温河谷三大类型(
... 干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(
云南不同地区和生境代表性热带植物区系的物种组成比较
1
2014
... 在云南东南部的文山古林箐(
Biodiversity hotspots for conservation priorities.
2
2000
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
... 亚洲的热带边缘以及三大夏季风的叠加影响区, 正是高山大河与石灰岩地貌集中分布的中国西南及中南半岛一带(
琼西热带稀树草原成因初探
4
2013
... 海南岛西部从洋浦、昌江直到莺歌海的沿海平地、河流三角洲及部分沙地上, 分布着一片长约170 km、宽3-20 km的沙漠化土地, 这里曾经是我国唯一的热带稀树草原分布区(
... ;
... 处于热带季风气候区的海南岛缘何在其西部滨海地带出现较大面积的热带稀树草原, 引起了较大的争议, 公认的最主要原因是季风气候(
... ;
金沙江干热河谷植物区系和生态多样性的初步研究
1
1996
... 干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(
Temporal and spatial origin of Gesneriaceae in the New World inferred from plastid DNA squences.
2
2013
... 苦苣苔科(Gesneriaceae)可能于7000万年前起源于南美, 在约49 Ma扩散至亚洲的东南亚群岛(
... ;
风筝果镜像花的雌雄异位变化及传粉生态型的形成
3
2016
... 可以推测, 苦苣苔亚科这类早期分布在热带赤道一带的植物, 在热带季风作用下向北扩散至中国西南及中南半岛之后, 受到高山深谷与石灰岩地貌的隔离作用, 这些位于分布区边缘的“先锋个体”与热带地区核心种群的基因流遭到阻断; 再加上局域高度异质性的地形造成的隔离作用, 导致扩散到这里的个体逐渐适应局域生境季节性干湿气候、石灰岩高钙土壤等因素, 慢慢分化出新种(
... 风筝果属和盾翅藤属都属于金虎尾科, 是热带亚洲特有的大型木质藤本.这两个属都有较为特殊的果实传播方式, 风筝果属果具3条果翅; 盾翅藤属果皮全部伸展成薄翅状附属物.翅果使得这两个属能够随着季风长距离迁移(
... 综合以上4个典型热带植物类群的物种分布格局可以发现, 中国西南的滇黔桂交界区是这些属或亚科共同的物种丰富度中心(
海南石灰岩地区的种子植物区系
3
2014
... 在云南东南部的文山古林箐(
... 在东南亚以及中国云南南部、广西西南部及海南岛等地的石灰岩山峰中上部, 还发育着一类特殊的季雨林(
... ;
The upper reaches of the largest river in Southern China as an ‘evolutionary front’ of tropical plants: Evidences from Asia-endemic genus Hiptage (Malpighiaceae)
11
2015
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
... ;
... 在热带植物南退的历史过程中, 部分热带植物在一些地区特殊地形地貌的“避难所”作用下得以存活, 再加上后期(不早于20 Ma;
... 热带季雨林被认为是热带雨林(季节性雨林)向热带稀树草原植被过渡的一种特殊植被类型(
... 处于热带季风气候区的海南岛缘何在其西部滨海地带出现较大面积的热带稀树草原, 引起了较大的争议, 公认的最主要原因是季风气候(
... 苦苣苔科(Gesneriaceae)可能于7000万年前起源于南美, 在约49 Ma扩散至亚洲的东南亚群岛(
... 可以推测, 苦苣苔亚科这类早期分布在热带赤道一带的植物, 在热带季风作用下向北扩散至中国西南及中南半岛之后, 受到高山深谷与石灰岩地貌的隔离作用, 这些位于分布区边缘的“先锋个体”与热带地区核心种群的基因流遭到阻断; 再加上局域高度异质性的地形造成的隔离作用, 导致扩散到这里的个体逐渐适应局域生境季节性干湿气候、石灰岩高钙土壤等因素, 慢慢分化出新种(
... 风筝果属和盾翅藤属都属于金虎尾科, 是热带亚洲特有的大型木质藤本.这两个属都有较为特殊的果实传播方式, 风筝果属果具3条果翅; 盾翅藤属果皮全部伸展成薄翅状附属物.翅果使得这两个属能够随着季风长距离迁移(
... 风筝果属物种多样性与特有种分布中心主要有两个: 滇黔桂交界区的红水河流域、泰国北部的湄平河上游(
... 综合以上4个典型热带植物类群的物种分布格局可以发现, 中国西南的滇黔桂交界区是这些属或亚科共同的物种丰富度中心(
... ), 一旦热带植物类群抵达这些河流河谷及上游, 很难与下游的种群保持基因流, 容易导致隔离物种形成(
3
... 亚洲地区是全球季风气候最为典型的区域, 最主要的特征就是降水多发生在高温的夏季(
... 亚洲的热带季风包括南亚季风(影响南亚和东南亚)和西北太平洋季风(影响菲律宾、中国南海及华南沿海地区), 与东亚季风(主要影响中国长江流域、日本和韩国等地)同为夏季风(
... 中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(
A fossil fruit wing of Dipterocarpus from the middle Micocene of Fujiang, China and its palaeoclimatic significance.
1
2010
... 对古植物化石的研究发现, 热带植物曾经在东亚特别是中国南部有更广阔的分布区, 热带北界比现今的热带北界更靠北, 如中新世时期中国的东南部和24° N地区存在着龙脑香科植物(
喜马拉雅东部雅鲁藏布江大峡弯河谷地区植物区系的特点及来源
4
1996
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
... 对古植物化石的研究发现, 热带植物曾经在东亚特别是中国南部有更广阔的分布区, 热带北界比现今的热带北界更靠北, 如中新世时期中国的东南部和24° N地区存在着龙脑香科植物(
... 西藏东南部的雅鲁藏布江在29° N的地方出现了呈南北走向、海拔差达5 000 m左右的深切峡谷, 成为南亚季风向北输送水汽与热量的天然通道(
... ).这里分布的热带植物以泛热带分布和热带亚洲分布最多, 是印度-马来植物区系在南亚季风作用下沿雅鲁藏布江河谷北进的结果(
喜马拉雅东部雅鲁藏布江大峡弯河谷地区植被组成特点
1997
从中国古植被记录看东亚季风年龄
1
2005
... 对古植物化石的研究发现, 热带植物曾经在东亚特别是中国南部有更广阔的分布区, 热带北界比现今的热带北界更靠北, 如中新世时期中国的东南部和24° N地区存在着龙脑香科植物(
A review of recent research on the East Asian summer monsoon in China. In: Chang CP, Krishnamurti TN eds. Monsoon Meteorology
4
... 亚洲地区是全球季风气候最为典型的区域, 最主要的特征就是降水多发生在高温的夏季(
... 南亚季风的成因既有印度洋与印度半岛的海陆热力性质差异的驱动, 也受东南亚及大洋洲一带气压带在夏季北移的影响(
... ); 同时, 高耸的青藏高原也影响着南亚季风的风向和风力(
... 西北太平洋季风发生在每年的5-9月, 发源于太平洋西部和新几内亚东岸150° E附近区域的一条跨越赤道的气流通道(
Characteristics of karst ecosystems of Vietnam and their vulnerability to human impact.
1
2001
... 亚洲的热带季节性雨林是热带雨林在热带边缘零星分布的常绿或半常绿森林, 主要出现在受到热带季风气候深刻影响的热带边缘区域, 如亚洲热带南缘的澳大利亚北领地、热带北缘的中国云南西双版纳、海南岛及广西与越南接壤处.从优势种、植被外貌来看, 季节性雨林与热带雨林相差不大, 建群种和优势种都以龙脑香科的望天树(Parashorea chinensis)和婆罗双(Shorea assamica)(
Rainy season of the Asian-pacific summer monsoon.
2
2002
... 亚洲地区是全球季风气候最为典型的区域, 最主要的特征就是降水多发生在高温的夏季(
... 中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(
雨林和季雨林的主要区别
2
1984
... 亚洲的热带季节性雨林是热带雨林在热带边缘零星分布的常绿或半常绿森林, 主要出现在受到热带季风气候深刻影响的热带边缘区域, 如亚洲热带南缘的澳大利亚北领地、热带北缘的中国云南西双版纳、海南岛及广西与越南接壤处.从优势种、植被外貌来看, 季节性雨林与热带雨林相差不大, 建群种和优势种都以龙脑香科的望天树(Parashorea chinensis)和婆罗双(Shorea assamica)(
... 热带季雨林或称“季风雨林”, 是在干湿季节交替的热带季风条件下形成的一种地带性密闭型森林植被, 其突出的特点是建群种为旱季落叶的热带性阔叶树、生长期存在一个明显的落叶期(
Monsoons: Processes, predictability, and the prospects for prediction.
1
1998
... 季风(monsoon)是由于太阳高度角变化引起海陆热力性质差异或气压带随季节移动导致盛行风向和降水随季节改变的现象(
广西苦苣苔科植物区系和生态特点研究
3
2004
... 苦苣苔科(Gesneriaceae)可能于7000万年前起源于南美, 在约49 Ma扩散至亚洲的东南亚群岛(
... ).以往的研究强调石灰岩生境的隔离效应对特有类群分化与维持的重要作用(
... 可以推测, 苦苣苔亚科这类早期分布在热带赤道一带的植物, 在热带季风作用下向北扩散至中国西南及中南半岛之后, 受到高山深谷与石灰岩地貌的隔离作用, 这些位于分布区边缘的“先锋个体”与热带地区核心种群的基因流遭到阻断; 再加上局域高度异质性的地形造成的隔离作用, 导致扩散到这里的个体逐渐适应局域生境季节性干湿气候、石灰岩高钙土壤等因素, 慢慢分化出新种(
广西热带石灰岩季节雨林分类与排序
3
1991
... 在云南东南部的文山古林箐(
... 在东南亚以及中国云南南部、广西西南部及海南岛等地的石灰岩山峰中上部, 还发育着一类特殊的季雨林(
... ).位于这些石灰岩山峰中部及下部的不存在明显落叶期的常绿或半常绿森林, 《中国植被》和《云南植被》称之为“石灰山常绿季雨林” (
关于亚洲夏季风爆发的动力学研究的若干近期进展
1
2013
... 季风(monsoon)是由于太阳高度角变化引起海陆热力性质差异或气压带随季节移动导致盛行风向和降水随季节改变的现象(
西南纵向岭谷地形对季风的“通道—阻隔”作用
2
2012
... 干旱河谷的主要特征是气温年较差小、干湿季分明(雨季短、旱季长而炎热)(
... 综合以上4个典型热带植物类群的物种分布格局可以发现, 中国西南的滇黔桂交界区是这些属或亚科共同的物种丰富度中心(
广西弄岗国家级自然保护区植物区系地理学研究
2
2011
... 在云南东南部的文山古林箐(
... 在东南亚以及中国云南南部、广西西南部及海南岛等地的石灰岩山峰中上部, 还发育着一类特殊的季雨林(
中国植物区系中的特有性及其起源和分化
1
2005
... 东亚和东南亚地区一直被认为是现代被子植物的起源与分化中心(
Forest contraction in north equatorial Southeast Asia during the Last Glacial Period.
1
2010
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
中国植物区系中的特有现象——特有属的研究
1
1984
... 苦苣苔科(Gesneriaceae)可能于7000万年前起源于南美, 在约49 Ma扩散至亚洲的东南亚群岛(
Palaeovegetation of China: A pollen data-based synthesis for the mid-Holocene and last glacial maximum.
2
2000
... 对古植物化石的研究发现, 热带植物曾经在东亚特别是中国南部有更广阔的分布区, 热带北界比现今的热带北界更靠北, 如中新世时期中国的东南部和24° N地区存在着龙脑香科植物(
... ;
龙脑香科植物的地理分布
1
1985
... 西藏东南部的雅鲁藏布江在29° N的地方出现了呈南北走向、海拔差达5 000 m左右的深切峡谷, 成为南亚季风向北输送水汽与热量的天然通道(
4
... 中国的西南地区是东亚季风与南亚季风的交接处, 也是西北太平洋季风波及到的区域(
... 干旱河谷的主要特征是气温年较差小、干湿季分明(雨季短、旱季长而炎热)(
... 干旱河谷的植被主要包括耐旱的稀树灌丛或灌(禾)草丛, 在外貌和区系成分上类似于非洲稀树草原(Savanna)和地中海气候下的灌木丛(Maquis), 故被称为半萨瓦纳(Semi-savanna)、河谷型Maquis灌丛(
... ).其中, 干热河谷的温度最高, 以热带性质的稀树草原、稀树肉质灌丛植被为主; 干暖河谷植被则以暖性落叶阔叶灌丛、常绿硬叶林、常绿阔叶灌丛为优势类型(
海南万宁青皮林群落多样性特征研究
2
2007
... 在云南东南部的文山古林箐(
... ), 应属“热带季节性雨林”.在海南岛东侧的西北太平洋季风的迎风面沿海一带的万宁市石梅湾就分布着一片绵延约16 km、宽约400 m的青皮林单优自然群落(
RS与GIS支持下近50 a海南岛西部土地沙漠化时空演变过程研究
3
2009
... 海南岛西部从洋浦、昌江直到莺歌海的沿海平地、河流三角洲及部分沙地上, 分布着一片长约170 km、宽3-20 km的沙漠化土地, 这里曾经是我国唯一的热带稀树草原分布区(
... 处于热带季风气候区的海南岛缘何在其西部滨海地带出现较大面积的热带稀树草原, 引起了较大的争议, 公认的最主要原因是季风气候(
... ;
若干重要地质事件对中国植物区系形成演变的影响
5
2017
... 亚洲地区是全球季风气候最为典型的区域, 最主要的特征就是降水多发生在高温的夏季(
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
... ;
... 南亚季风的成因既有印度洋与印度半岛的海陆热力性质差异的驱动, 也受东南亚及大洋洲一带气压带在夏季北移的影响(
... ).南亚季风在5-7月强度最大, 风向以西南向东北为主, 主要影响印度、孟加拉湾、中南半岛, 以及我国西南地区如横断山区和云贵高原(
云南热带季雨林及其与热带雨林植被的比较
8
2011
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
... 亚洲的热带季节性雨林是热带雨林在热带边缘零星分布的常绿或半常绿森林, 主要出现在受到热带季风气候深刻影响的热带边缘区域, 如亚洲热带南缘的澳大利亚北领地、热带北缘的中国云南西双版纳、海南岛及广西与越南接壤处.从优势种、植被外貌来看, 季节性雨林与热带雨林相差不大, 建群种和优势种都以龙脑香科的望天树(Parashorea chinensis)和婆罗双(Shorea assamica)(
... 热带季雨林或称“季风雨林”, 是在干湿季节交替的热带季风条件下形成的一种地带性密闭型森林植被, 其突出的特点是建群种为旱季落叶的热带性阔叶树、生长期存在一个明显的落叶期(
... ;
... 热带季雨林被认为是热带雨林(季节性雨林)向热带稀树草原植被过渡的一种特殊植被类型(
... ;
... )、风筝果等组成的旱季落叶植被, 应属较典型的热带季雨林(
... 干旱河谷的主要特征是气温年较差小、干湿季分明(雨季短、旱季长而炎热)(
Geographical elements of seed plants suggest the boundary of the tropical zone in China.
3
2013
... 目前, 针对亚洲季风气候的研究侧重于温带和亚热带植物, 相对忽略了热带植被类群.亚洲地区是全球热带植物分布最靠北的区域, 如北回归线以北的贵州红水河流域(约25° N)、西藏东南部的雅鲁藏布江河谷(约29° N)都分布有一些典型的热带植被类群(
... 在热带植物南退的历史过程中, 部分热带植物在一些地区特殊地形地貌的“避难所”作用下得以存活, 再加上后期(不早于20 Ma;
... ), 并在当地复杂地形的影响下形成了多种多样的局域小气候和非地带性植被(
西双版纳石灰岩森林的植物区系地理研究
3
1996
... 在东南亚以及中国云南南部、广西西南部及海南岛等地的石灰岩山峰中上部, 还发育着一类特殊的季雨林(
... ).石灰岩山峰海拔较高的地方由于石灰岩漏水特性导致干湿变动极其剧烈(
... ), 再加上土层薄、含钙量极大等特点, 发育出了喜钙耐旱植物为主的特殊季雨林(


{kind=link}
{kind=link}
{kind=link}
{kind=link}