Main factors driving changes in soil respiration under altering precipitation regimes and the controlling processes
YANG Qing-Xiao1,2, TIAN Da-Shuan2, ZENG Hui1,4,*,, NIU Shu-Li2,3 1School of Urban Planning and Design, Peking University Shenzhen Graduate School, Shenzhen 518055, China2Key Laboratory of Ecosystem Network Observation and Modelling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China3College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, Chinaand 4Key Laboratory for Earth Surface Processes of the Ministry of Education, Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China 通讯作者:* 通信作者Author for correspondence (E-mail: zengh@pkusz.edu.cn) 责任编辑: 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(31470528和31625006)
关键词:土壤呼吸;碳循环;整合分析;降水变化 Abstract Aims Our objective was to determine the effects of changes in global pattern of precipitation on soil respiration and the controlling factors. Methods Data were collected from literature on precipitation manipulation experiments globally and a meta-analysis was conducted to synthesize the responses of soil respiration to changes in precipitation regimes. Important findings We found that an increased precipitation stimulated soil respiration while a decreased precipitation suppressed it. When changes in rainfall were normalized to the average treatment level (41% of the current annual precipitation), the level of increases in soil respiration with increased precipitation (49%) were higher than that of decreases with decreased precipitation (21%), showing an asymmetric responses of soil respiration to increases and decreases in precipitation. Soil moisture occurred as the most predominant factor driving the changes in soil respiration under altered precipitation. Changes in soil moisture affected soil respiration directly and indiscreetly by changing aboveground/belowground net primary productivity and microbial biomass carbon, which collectively contributed 98% of variations in soil respiration. In addition, the responses of soil respiration to altered precipitation varied with background temperature and precipitation. The sensitivity of soil respiration increased with local mean annual temperature when precipitation was reduced, while remaining unchanged when precipitation was increased. Meanwhile, the sensitivity of soil respiration to either increases or decreases in precipitation decreased with increasing local mean annual precipitation. Under future altered precipitation regimes, the sensitivity of soil respiration to changes in precipitation is likely dependent of local environment conditions. Keywords:soil respiration;carbon cycle;meta-analysis;precipitation change -->0 PDF (1290KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 杨青霄, 田大栓, 曾辉, 牛书丽. 降水格局改变背景下土壤呼吸变化的主要影响因素及其调控过程. 植物生态学报, 2017, 41(12): 1239-1250 https://doi.org/10.17521/cjpe.2017.0208 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013)。多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015)。未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005)。同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013)。全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013)。作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013)。 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014)。土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用。 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016)。为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017)。由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生。Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用。Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用。但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚。一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变。因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑。
将土壤湿度对土壤呼吸的影响分为增雨和减雨处理来看。增雨处理下, 土壤湿度对土壤呼吸变化的贡献率较高(图5A, R2 = 0.46), 并且, 土壤呼吸对水分的敏感性在增雨处理下(图5A, S = 0.74, S是斜率, 表示土壤湿度的响应值每变化一个单位引起的土壤呼吸响应值变化量, 以此类推)高于减雨处理(图5A, S = 0.58), 呈现出土壤呼吸对降水变化的非对称响应, 这进一步验证了图2的结果。 显示原图|下载原图ZIP|生成PPT 图1降水变化下土壤湿度(A)和土壤呼吸(B)响应值随降水改变量梯度的变化。黑色点表示增加降水处理, 白色点表示减少降水处理。p < 0.01表示统计结果极显著, p < 0.05表示统计结果显著。 -->Fig. 1Regressional relationships of soil moisture (A) and soil respiration (B) with percentage changes of precipitation. The filled circles represent increased precipitation, and the open circles represent decreased precipitation. p < 0.01, statistically highly significant; p < 0.05, statistically significant. -->
显示原图|下载原图ZIP|生成PPT 图2土壤呼吸对增雨和减雨的总响应及不同生态系统类型对增雨和减雨的响应差异(效应值±95%置信区间)。白色的条形柱表示减少降水处理引起的土壤呼吸降低幅度, 黑色代表增加降水处理引起的土壤呼吸增加幅度。数字表示用于整合分析的样本量。*表示响应的统计结果显著。 -->Fig. 2Changes of normalized soil respiration in overall and different ecosystems under increased or decreased precipitation (effect size ± 95% confidence interval). The white bars represent the negative effects of decreased precipitation while the black bars represent the positive effects of increased precipitation. The values beside the bars indicate sample sizes used in meta-analysis.* means statistically significant. -->
显示原图|下载原图ZIP|生成PPT 图3降水变化下土壤湿度(A)地上净初级生产力(B)、地下净初级生产力(C)和土壤微生物碳含量(D)变化对土壤呼吸的影响。黑色点代表增加降水处理, 白色点代表减少降水处理。p < 0.01表示统计结果极显著, p < 0.05表示统计结果显著。 -->Fig. 3Regressional relationships of soil respiration with soil moisture (A), aboveground net primary productivity (B), belowground net primary productivity (C), and microbial biomass carbon (D). The filled circles represent increased precipitation, and the open circles represent decreased precipitation. p < 0.01, statistically highly significant; p < 0.05, statistically significant. -->
显示原图|下载原图ZIP|生成PPT 图4降水变化下土壤湿度、地上净初级生产力、地下净初级生产力和土壤微生物碳含量变化影响土壤呼吸变化机制的结构方程模型。灰色和黑色的箭头分别代表显著的正效应和负效应。箭头旁边的数字代表回归系数。箭头的粗细代表回归关系的强度。R2表示有关变量的共同解释度。***, < 0.001; **, p < 0.01; *, p < 0.05。χ2 = 0.49, p = 0.48, 比较拟合指数(CFI) = 1.00, 近似误差均方根(RMSEA) = 0.00, 赤池信息准则(AIC) = 38.49。 -->Fig. 4A structural equation model of the effects of soil moisture, aboveground net primary productivity, belowground net primary productivity, and microbial biomass carbon on soil respiration. Gray and black arrows represent significant positive and negative pathways, respectively. Values beside the arrows indicate the standard path coefficients. Arrow width is proportional to the strength of the relationship. R2 values represent the proportion of variance explainable by each variable in the model. ***, p < 0.001; **, p < 0.01; *, p < 0.05. χ2 = 0.49, p = 0.48, comparative fit index (CFI) = 1.00, root mean square error of approximation (RMSEA) = 0.00, Akaike information criteria (AIC) = 38.49. -->
土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小。因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010)。本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸。这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致。并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016)。 显示原图|下载原图ZIP|生成PPT 图5增加降水和减少降水下土壤呼吸变化与土壤湿度变化的关系(A, D)及在不同本底年平均气温(℃; B, E)和年降水量(mm; C, F)下土壤呼吸随土壤湿度变化的敏感性差异。黑色实线、虚线、灰色实线分别表示3个降水或温度梯度下的土壤呼吸变化和土壤湿度变化的回归关系。S1、S2、S3分别是3条回归线的斜率。A, B, C, 增加降水处理。D, E, F, 减少降水处理。p < 0.01表示统计结果极显著, p < 0.05表示统计结果显著。RR, 响应值。 -->Fig. 5Regressional relationships between response ratio of soil respiration and response ratio of soil moisture under increased or decreased precipitation (A, D). Different sensitivities of changes in soil respiration to soil moisture under increased or decreased precipitation in different conditional mean annual temperature (℃; B, E) and mean annual precipitation (mm; C, F). Black line, dash line and gray line represent the regression relationships between response ratio of soil respiration and response ratio of soil moisture under three precipitation or three temperature gradients, respectively. S1, S2, and S3 represent slopes of the three regression lines, respectively. A, B, and C, Increased precipitation treatment. D, E, and F, Decreased precipitation treatments. p < 0.01, statistically highly significant; p < 0.05, statistically significant. RR, response ratio. -->
AanderudZT, SchoolmasterDR, LennonJT (2011). Plants mediate the sensitivity of soil respiration to rainfall variability .Ecosystems, 14, 156-167.https://doi.org/10.1007/s10021-010-9401-yURL [本文引用: 2]摘要 Soil respiration from grasslands plays a critical role in determining carbon dioxide (CO 2 ) feedbacks between soils and the atmosphere. In these often mesic systems, soil moisture and temperature tend to co-regulate soil respiration. Increasing variance of rainfall patterns may alter aboveground–belowground interactions and have important implications for the sensitivity of soil respiration to fluctuations in moisture and temperature. We conducted a set of field experiments to evaluate the independent and interactive effects of rainfall variability and plant–soil processes on respiration dynamics. Plant removal had strong effects on grassland soils, which included altered CO 2 flux owing to absence of root respiration; increased soil moisture and temperature; and reduced availability of dissolved organic carbon (DOC) for heterotrophic respiration by microorganisms. These plant-mediated effects interacted with our rainfall variability treatments to determine the sensitivity of soil respiration to both moisture and temperature. Using time-series multiple regression, we found that plants dampened the sensitivity of respiration to moisture under high variability rainfall treatments, which may reflect the relative stability of root contributions to total soil respiration. In contrast, plants increased the sensitivity of respiration to temperature under low variability rainfall treatment suggesting that the environmental controls on soil CO 2 dynamics in mesic habitats may be context dependent. Our results provide insight into the aboveground–belowground mechanisms controlling respiration in grasslands under variable rainfall regimes, which may be important for predicting CO 2 dynamics under current and future climate scenarios.
BaiYF, HanXG, WuJG, ChenZZ, LiLH (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland .Nature, 431, 181-184.https://doi.org/10.1038/nature02850URLPMID:202020202020202020202020 [本文引用: 1]摘要 Numerous studies have suggested that biodiversity reduces variability in ecosystem productivity through compensatory effects109“6;that is, a species increases in its abundance in response to the reduction of another in a fluctuating environment1,7. But this view has been challenged on several grounds809“10. Because most studies have been based on artificially constructed grass- lands with short duration, long-term studies of natural ecosystems are needed. On the basis of a 24-year study of the Inner Mongolia grassland, here we present three key findings. First, that January-July precipitation is the primary climatic factor causing fluctuations in community biomass production; second, that ecosystem stability (conversely related to variability in community biomass production) increases progressively along the hierarchy of organizational levels (that is, from species to functional group to whole community); and finally, that the community-level stability seems to arise from compensatory interactions among major components at both species and functional group levels. From a hierarchical perspective, our results corroborate some previous findings of compensatory effects1,4,7,11. Undisturbed mature steppe ecosystems seem to culminate with high biodiversity, productivity and ecosystem stability concurrently. Because these relationships are correlational, further studies are necessary to verify the causation among these factors. Our study provides new insights for better management and restoration of the rapidly degrading Inner Mongolia grassland.
[4]
BaiW, WanS, NiuS, LiuW, ChenQ, WangQ, ZhangW, HanX, LiL (2010). Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling .Global Change Biology, 16, 1306-1316.https://doi.org/10.1111/gcb.2010.16.issue-4URL
[5]
BaoF, ZhouGS (2010). Review of research advances in soil respiration of grassland in China .Chinese Journal of Plant Ecology, 34, 713-726.(in Chinese with English abstract) [鲍芳, 周广胜 (2010). 中国草原土壤呼吸作用研究进展 . 植物生态学报, 34, 713-726.]https://doi.org/10.3773/j.issn.1005-264x.2010.06.011 [本文引用: 1]摘要 中国草原面积约占国土面积的40%,且大都位于生态脆弱区,对气候和环境变化十分敏感,在未来大气CO2调控中有着重要的作用。为增进对中国草原土壤呼吸作用的理解,该文综述了近10年来中国草原土壤呼吸作用的最新研究进展,指出中国草原土壤呼吸作用的研究主要集中在东北平原、内蒙古高原和青藏高原。草原土壤呼吸作用日动态的主导控制因子是温度,季节动态的主导控制因子可以是温度、水分或二者的交互作用,取决于研究地点的限制性环境因子,而年际动态的主导控制因子为水分。草原土壤呼吸作用还存在着巨大的空间变异,年降水和土壤全氮含量是不同类型草原土壤呼吸作用空间异质性的主导控制因子。土壤呼吸作用对全球变化的响应比较复杂,取决于各因子之间相互影响的贡献。现有的土壤呼吸作用模型大多只考虑了水热因子,很少包含土壤因子和生物因子及其协同作用的影响。在此基础上,指出未来中国草原土壤呼吸作用拟加强的研究重点:1)温带荒漠草原土壤呼吸作用研究;2)非生长季土壤呼吸作用研究;3)多时空尺度草原土壤呼吸作用的比较研究;4)草原土壤呼吸作用过程模拟研究;5)草原土壤呼吸作用的遥感监测评估研究。
[6]
BeierC, BeierkuhnleinC, WohlgemuthT, PenuelasJ, EmmettB, KornerC, de BoeckH, ChristensenJH, LeuzingerS, JanssensIA, HansenK (2012). Precipitation manipulation experiments—Challenges and recommendations for the future .Ecology Letters, 15, 899-911.https://doi.org/10.1111/j.1461-0248.2012.01793.xURL [本文引用: 2]
[7]
BorkenW, XuYJ, DavidsonEA, BeeseA (2002). Site and temporal variation of soil respiration in European beech, Norway spruce, and Scots pine forests .Global Change Biology, 8, 1205-1216.https://doi.org/10.1046/j.1365-2486.2002.00547.xURL [本文引用: 2]
[8]
ChenSP, LinGH, HuangJH, HeM (2008). Responses of soil respiration to simulated precipitation pulses in semiarid steppe under different grazing regimes .Journal of Plant Ecology, 1, 237-246.https://doi.org/10.1093/jpe/rtn020URL [本文引用: 1]
[9]
ChenSP, LinGH, HuangJH, JeneretteGD (2009). Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe .Global Change Biology, 15, 2450-2461.https://doi.org/10.1111/j.1365-2486.2009.01879.xURL [本文引用: 1]摘要 Precipitation pulses play an important role in regulating ecosystem carbon exchange and balance of semiarid steppe ecosystems. It has been predicted that the frequency of extreme rain events will increase in the future, especially in the arid and semiarid regions. We hypothesize that large rain pulses favor carbon sequestration, while small ones cause more carbon release in the semiarid steppes. To understand the potential response in carbon sequestration capacity of semiarid steppes to the changes in rain pulse size, we conducted a manipulative experiment with five simulated rain pulse sizes (0, 5, 10, 25, and 75 mm) in Inner Mongolia steppe. Our results showed that both gross ecosystem productivity (GEP) and ecosystem respiration ( R e ) responded rapidly (within 24 h) to rain pulses and the initial response time was independent of pulse size. However, the time of peak GEP was 1 3 days later than that of R e , which depended on pulse size. Larger pulses caused greater magnitude and longer duration of variations in GEP and R e . Differences in the response time of microbes and plants to wetting events constrained the response pattern of heterotrophic ( R h ) and autotrophic ( R a ) components of R e following a rain event. R h contributed more to the increase of R e in the early stage of rain pulse response, while R a played an more important role later, and determined the duration of pulse response, especially for large rain events of >10 mm. The distinct responses of ecosystem photosynthesis and respiration to increasing pulse sizes led to a threshold in rain pulse size between 10 and 25 mm, above which post wetting responses favored carbon sequestration. The disproportionate increase of the primary productivity of higher plants, compared with those in the activities of microbial decomposers to larger pulse events suggests that the carbon sequestration capacity of Inner Mongolia steppes will be sensitive to changes in precipitation size distribution rather than just precipitation amount.
[10]
ChimnerRA, WelkerJM, MorganJ, LecainD, ReederJ (2010). Experimental manipulations of winter snow and summer rain influence ecosystem carbon cycling in a mixed-grass prairie, Wyoming, USA .Ecohydrology, 3, 284-293.https://doi.org/10.1002/eco.106URL [本文引用: 1]摘要 Water-limited grasslands may be exceedingly vulnerable to changes in the timing or amount of precipitation which may result in shifts in the magnitudes and patterns of carbon cycling. Shifts in CO2 exchange may lead to alterations in carbon sequestration or net losses and could accentuate the rising CO2 concentrations in the atmosphere or ameliorate the increases. The objective of our project was to quantify how changes in winter, summer, and combined winter and summer precipitation may alter rates of ecosystem C cycling in the mixed-grass prairie of the United States. Three replicated 50 m snow fences were installed to increase winter snow while summer precipitation was manipulated by either increasing (+50%) or decreasing ( 50%) precipitation amounts. Measurements of net ecosystem exchange (NEE), gross ecosystem photosynthesis (GEP), and ecosystem respiration (ER) and plant biomass were conducted throughout the snow-free period. Deeper snow in winter increased summer ER by an average of 27%, GEP by 45%, NEE by 90% and plant biomass by 50% compared to ambient snow conditions. Average plant biomass increased 44% with the addition of summer rain and decreased by 18% where rainfall was excluded under ambient snow conditions. Average NEE was also lower where rain was excluded. Our findings indicate that seasonality changes of precipitation may be important in regulating the future extent of C sequestration and C cycling in one of the most extensive, intact grasslands of North America. Copyright 2010 John Wiley & Sons, Ltd.
[11]
ClevelandCC, WiederWR, ReedSC, TownsendAR (2010). Experimental drought in a tropical rain forest increases soil carbon dioxide losses to the atmosphere .Ecology, 91, 2313-2323.https://doi.org/10.1890/09-1582.1URLPMID:20836453 [本文引用: 2]摘要 Climate models predict precipitation changes for much of the humid tropics, yet few studies have investigated the potential consequences of drought on soil carbon (C) cycling in this important biome. In wet tropical forests, drought could stimulate soil respiration via overall reductions in soil anoxia, but previous research suggests that litter decomposition is positively correlated with high rainfall fluxes that move large quantities of dissolved organic matter (DOM) from the litter layer to the soil surface. Thus, reduced rainfall could also limit C delivery to the soil surface, reducing respiration rates. We conducted a throughfall manipulation experiment to investigate how 25% and 50% reductions in rainfall altered both C movement into soils and the effects of those DOM fluxes on soil respiration rates. In response to the experimental drought, soil respiration rates increased in both the -25% and -50% treatments. Throughfall fluxes were reduced by 26% and 55% in the-25% and-50% treatments, respectively. However, total DOM fluxes leached from the litter did not vary between treatments, because the concentrations of leached DOM reaching the soil surface increased in response to the simulated drought. Annual DOM concentrations averaged 7.7 ?? 0.8, 11.2 ?? 0.9, and 15.8 ?? 1.2 mg C/L in the control, -25%, and -50% plots, respectively, and DOM concentrations were positively correlated with soil respiration rates. A laboratory incubation experiment confirmed the potential importance of DOM concentration on soil respiration rates, suggesting that this mechanism could contribute to the increase in CO2 fluxes observed in the reduced rainfall plots. Across all plots, the data suggested that soil CO2 fluxes were partially regulated by the magnitude and concentration of soluble C delivered to the soil, but also by soil moisture and soil oxygen availability. Together, our data suggest that declines in precipitation in tropical rain forests could drive higher CO2 fluxes to the atmosphere both via increased soil O2 availability and through responses to elevated DOM concentrations. ?? 2010 by the Ecological Society of America.
[12]
CoxPM, BettsRA, JonesCD, SpallSA, TotterdellIJ (2000). Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model .Nature, 408, 184-187.https://doi.org/10.1038/35041539URLPMID:11089968 [本文引用: 1]摘要 The continued increase in the atmospheric concentration of carbon dioxide due to anthropogenic emissions is predicted to lead to significant changes in climate. About half of the current emissions are being absorbed by the ocean and by land ecosystems, but this absorption is sensitive to climate as well as to atmospheric carbon dioxide concentrations, creating a feedback loop. General circulation models have generally excluded the feedback between climate and the biosphere, using static vegetation distributions and CO2 concentrations from simple carbon-cycle models that do not include climate change. Here we present results from a fully coupled, three-dimensional carbon-climate model, indicating that carbon-cycle feedbacks could significantly accelerate climate change over the twenty-first century. We find that under a 'business as usual' scenario, the terrestrial biosphere acts as an overall carbon sink until about 2050, but turns into a source thereafter. By 2100, the ocean uptake rate of 5 Gt C yr(-1) is balanced by the terrestrial carbon source, and atmospheric CO2 concentrations are 250 p.p.m.v. higher in our fully coupled simulation than in uncoupled carbon models, resulting in a global-mean warming of 5.5 K, as compared to 4 K without the carbon-cycle feedback.
[13]
DaiAG (2013). Increasing drought under global warming in observations and models .Nature Climate Change, 3, 52-58.https://doi.org/10.1038/nclimate1633URL [本文引用: 1]摘要 Historical records of precipitation, streamflow and drought indices all show increased aridity since 1950 over many land areas(1,2). Analyses of model-simulated soil moisture(3,4), drought indices(1,5,6) and precipitation-minus-evaporation(7) suggest increased risk of drought in the twenty-first century. There are, however, large differences in the observed and model-simulated drying patterns(1,2,6). Reconciling these differences is necessary before the model predictions can be trusted. Previous studies(8-12) show that changes in sea surface temperatures have large influences on land precipitation and the inability of the coupled models to reproduce many observed regional precipitation changes is linked to the lack of the observed, largely natural change patterns in sea surface temperatures in coupled model simulations(13). Here I show that the models reproduce not only the influence of El Nino-Southern Oscillation on drought over land, but also the observed global mean aridity trend from 1923 to 2010. Regional differences in observed and model-simulated aridity changes result mainly from natural variations in tropical sea surface temperatures that are often not captured by the coupled models. The unforced natural variations vary among model runs owing to different initial conditions and thus are irreproducible. I conclude that the observed global aridity changes up to 2010 are consistent with model predictions, which suggest severe and widespread droughts in the next 30-90 years over many land areas resulting from either decreased precipitation and/or increased evaporation.
[14]
DengQ, HuiDF, ChuGW, HanX, ZhangQF (2017). Rain- induced changes in soil CO2 flux and microbial community composition in a tropical forest of China .Scientific Reports, 7, 5539. doi: 10.1038/s41598-017-06345-2.https://doi.org/10.1038/s41598-017-06345-2URL [本文引用: 1]
[15]
EstiarteM, ViccaS, PenuelasJ, BahnM, BeierC, EmmettBA, FayPA, HansonPJ, HasibederR, KigelJ, Kroel-DulayG, LarsenKS, Lellei-KovacsE, LimousinJM, OgayaR, OurcivalJM, ReinschS, SalaOE, SchmidtIK, SternbergM, TielborgerK, TietemaA, JanssensIA (2016). Few multiyear precipitation-reduction experiments find a shift in the productivity-precipitation relationship .Global Change Biology, 22, 2570-2581.https://doi.org/10.1111/gcb.13269URLPMID:26946322 [本文引用: 1]摘要 Abstract Well defined productivity-precipitation relationships of ecosystems are needed as benchmarks for the validation of land-models used for future projections. The productivity-precipitation relationship may be studied in two ways: the spatial approach relates differences in productivity to those in precipitation among sites along a precipitation gradient (the spatial fit, with a steeper slope); the temporal approach relates inter-annual productivity changes to variation in precipitation within sites (the temporal fits, with flatter slopes). Precipitation-reduction experiments in natural ecosystems represent a complement to the fits, because they can reduce precipitation below the natural range and are thus well suited to study potential effects of climate drying. Here, we analyze the effects of dry treatments in eleven multi-year precipitation-manipulation experiments, focusing on changes in the temporal fit. We expected that structural changes in the dry treatments would occur in some experiments, thereby reducing the intercept of the temporal fit and displacing the productivity-precipitation relationship downward the spatial fit. The majority of experiments (72%) showed that dry treatments did not alter the temporal fit. This implies that current temporal fits are to be preferred over the spatial fit to benchmark land-model projections of productivity under future climate within the precipitation ranges covered by the experiments. Moreover, in two experiments, the intercept of the temporal fit unexpectedly increased due to mechanisms that reduced either water- or nutrient losses. The expected decrease of the intercept was observed in only one experiment, and only when distinguishing between the late and the early phases of the experiment. This implies that we currently do not know at which precipitation-reduction level or at which experimental duration structural changes will start to alter ecosystem productivity. Our study highlights the need for experiments with multiple, including more extreme, dry treatments, to identify the precipitation boundaries within which the current temporal fits remain valid. This article is protected by copyright. All rights reserved. This article is protected by copyright. All rights reserved.
[16]
FialaK, T?maI, HolubP (2009). Effect of manipulated rainfall on root production and plant belowground dry mass of different grassland ecosystems .Ecosystems, 12, 906-914.https://doi.org/10.1007/s10021-009-9264-2URL [本文引用: 1]摘要 A field experiment was established to quantify the effects of different amounts of rainfall on root growth and dry mass of belowground plant parts in three types of grassland ecosystems. Mountain ( Nardus grassland), highland (wet Cirsium grassland), and lowland grassland (dry Festuca grassland) ecosystems were studied in 2006 and 2007. Roofs constructed above the canopy of grass stands and gravity irrigation systems simulated three climate scenarios: (1) rainfall reduced by 50%, (2) rainfall enhanced by 50%, and (3) the full natural rainfall of the current growing season. Experimentally reduced amounts of precipitation significantly affected both yearly root increments and total root dry mass in the highland grassland. Dry conditions in 2007 resulted in considerable reduction of total belowground dry mass in highland and mountain grasslands. Although not all differences in root biomass of studied grasslands were statistically significantly, some also showed a decrease in root increment and in the amount of belowground dry mass in dry conditions.
FlanaganLB, WeverLA, CarlsonPJ (2002). Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland .Global Change Biology, 8, 599-615.https://doi.org/10.1051/0004-6361:200810276URL [本文引用: 3]摘要 Net ecosystem carbon dioxide (CO2 ) exchange (NEE) was measured in a northern temperate grassland near Lethbridge, Alberta, Canada for three growing seasons using the eddy covariance technique . The study objectives were to document how NEE and its major component processes-gross photosynthesis (GPP) and total ecosystem respiration (TER)-vary seasonally and interannually, and to examine how environmental and physiological factors influence the annual C budget. The greatest difference among the three study years was the amount of precipitation received. The annual precipitation for 1998 (481.7 mm) was significantly above the 1971-2000 mean (+/- SD, 377.9 +/- 97.0 mm) for Lethbridge, whereas 1999 (341.3 mm) was close to average, and 2000 (275.5 mm) was significantly below average. The high precipitation and soil moisture in 1998 allowed a much higher GPP and an extended period of net carbon gain relative to 1999 and 2000. In 1998, the peak NEE was a gain of 5 g C m(-2) d(-1) (day 173). Peak NEE was lower and also occurred earlier in the year on days 161 (3.2 g C m(-2) d(-1) ) and 141 (2.4 g C m(-2) d(-1) ) in 1999 and 2000, respectively. Change in soil moisture was the most important ecological factor controlling C gain in this grassland ecosystem. Soil moisture content was positively correlated with leaf area index (LAI). Gross photosynthesis was strongly correlated with changes in both LAI and canopy nitrogen (N) content. Maximum GPP (A (max) : value calculated from a rectangular hyperbola fitted to the relationship between GPP and incident photosynthetic photon flux density (PPFD)) was 27.5, 12.9 and 8.6 mumol m(-2) s(-1) during 1998, 1999 and 2000, respectively. The apparent quantum yield also differed among years at the time of peak photosynthetic activity, with calculated values of 0.0254, 0.018 and 0.018 during 1998, 1999 and 2000, respectively. The ecosystem accumulated a total of 111.9 g
[19]
GuanC, ZhangP, LiXR (2017). Responses of respiration with biocrust cover to water and temperature in the southeastern edge of Tengger Desert, Northwest China .Chinese Journal of Plant Ecology, 41, 301-310.(in Chinese with English abstract) [管超, 张鹏, 李新荣 (2017). 腾格里沙漠东南缘生物结皮土壤呼吸对水热因子变化的响应 . 植物生态学报, 41, 301-310.]https://doi.org/10.17521/cjpe.2016.0326URL [本文引用: 2]摘要 生物结皮土壤呼吸是干旱区碳循环的重要参与者,是了解荒漠生态系统碳循环的重要过程之一,但有关生物结皮土壤呼吸对水热因子的响应还存在许多不确定性,难以在区域尺度上准确评估生物结皮土壤系统碳排放对水热因子变化的响应及反馈方向和程度。该文以腾格里沙漠东南缘天然植被区藓类和藻-地衣结皮土壤为研究对象,利用开顶式生长室模拟增温,采用全自动土壤碳通量测定系统研究了模拟增温及降水格局变化对不同类型生物结皮土壤呼吸的影响。结果表明:观测期间(2016年4月1日到7月31日),不同自然降水事件下(降水量在0.3–30.0 mm间),藓类结皮土壤呼吸速率在–0.16–4.69μmol·m^–2·s^–1之间变动,藻-地衣结皮土壤呼吸速率在–0.21–5.72μmol·m^–2·s^–1之间变动。藓类结皮土壤呼吸速率平均为1.09μmol·m^–2·s^–1,高于藻-地衣结皮土壤呼吸速率的0.94μmol·m^–2·s^–1,是藻-地衣结皮土壤呼吸速率的1.2倍。生物结皮土壤呼吸在不同的降水事件下具有明显的时空异质性,且生物结皮土壤呼吸速率与降水量有显著正相关关系。对照下两类结皮土壤呼吸速率平均为1.24μmol·m^–2·s^–1,增温条件下为0.79μmol·m^–2·s^–1,增温显著降低了其呼吸速率,增温主要是通过加速土壤水分的散失而降低两类结皮土壤呼吸。大多数情况下,土壤温度和生物结皮土壤呼吸呈现类似的单峰曲线,但土壤温度峰值出现的时间滞后于生物结皮土壤呼吸峰值出现的时间,滞后时间一般为2 h。
[20]
GutschickVP, BassiriRadH (2003). Extreme events as shaping physiology, ecology, and evolution of plants: Toward a unified definition and evaluation of their consequences .New Phytologist, 160, 21-42.https://doi.org/10.1046/j.1469-8137.2003.00866.xURL [本文引用: 1]摘要 Abstract Top of page Abstract I.68Introduction II.68Moving to an organismally based definition of extreme events III.68Features to discern in extreme events 1.68Cascades of phenomena over time 2.68The importance of concurrent changes in several environmental variables 3.68Evolutionary constraints to responses in populations IV.68Additional challenges in the study of extreme events 1.68Shortcomings in statistical descriptions 2.68Scaling up in space and time 3.68Quantifying stochastic costs and benefits 4.68Difficulties in describing biotic dimensions in process-based models V.68Evolutionary dimensions 1.68Responses depend upon evolutionary history 2.68Are extreme events themselves strong(est) agents of natural selection? 3.68Arguments for functional neutrality of most trait variations in normal times 4.68Generalized neutrality: cancellation among fitness contributions in time or among different performance measures, and overdominance 5.68Some traits, and trait combinations, do get strongly selected and fixed 6.68Which traits are likely to be selected by extreme events? VI.68The mandate for new conceptual tools for ecological and evolutionary prediction VII.68Tools in hand, and tools needed, to study extreme events 1.68Tools for identifying extreme events in terms of organismal responses 2.68Tools for sensing driving variables of extreme events 3.68Testing neutrality and nonneutrality of trait variations 4.68Predicting ecological and evolutionary trajectories VIII.68Conclusions Acknowledgements References Contents I. Introduction000 II. Moving to an organismally based definition of extreme events000 III. Features to discern in extreme events000 IV. Additional challenges in the study of extreme events000 V. Evolutionary dimensions000 VI. The mandate for new conceptual tools for ecological and evolutionary prediction000 VII. Tools in hand, and tools needed, to study extreme events000 VIII. Conclusions000 Acknowledgements000 References000 Summary Here we consider how extreme events, particularly climatic and biotic, affect the physiology, development, ecology and evolution of organisms, focusing on plants. The marked effects on organisms are of increasing interest for ecological prediction, given the natural and anthropogenic changes in spectra of extreme events being induced by global change. Yet there is currently a paucity of knowledge or even a common world-view of how extreme events shape individuals, communities and ecosystems. We propose that extreme events need be defined in terms of organismal responses of acclimation and of de-acclimation or hysteresis. From this definition we proceed to develop a number of hypotheses, including that fitness effects of extreme events occur primarily during recovery. We review evidence that, on the evolutionary time scale, selection is virtually absent except during extreme events; these drive strong directional selection, even to trait fixation and speciation. We describe a number of new tools, both conceptual and technological, that are now at hand or that merit rapid development.
[21]
HagedornF, JoosO (2014). Experimental summer drought reduces soil CO2 effluxes and DOC leaching in Swiss grassland soils along an elevational gradient .Biogeochemistry, 117, 395-412.https://doi.org/10.1007/s10533-013-9881-xURL [本文引用: 1]
[22]
HansonPJ, EdwardsNT, GartenCT, AndrewsJA (2000). Separating root and soil microbial contributions to soil respiration: A review of methods and observations .Biogeochemistry, 48, 115-146.https://doi.org/10.1023/A:1006244819642URL [本文引用: 1]
[23]
HarperCW, BlairJM, FayPA, KnappAK, CarlisleJD (2005). Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem . Global Chang Biology, 11, 322-334.https://doi.org/10.1021/jp0017543URL [本文引用: 2]摘要 Predicted climate changes in the US Central Plains include altered precipitation regimes with increased occurrence of growing season droughts and higher frequencies of extreme rainfall events. Changes in the amounts and timing of rainfall events will likely affect ecosystem processes, including those that control C cycling and storage. Soil carbon dioxide (CO2) flux is an important component of C cycling in terrestrial ecosystems, and is strongly influenced by climate. While many studies have assessed the influence of soil water content on soil CO2 flux, few have included experimental manipulation of rainfall amounts in intact ecosystems, and we know of no studies that have explicitly addressed the influence of the timing of rainfall events. In order to determine the responses of soil CO2 flux to altered rainfall timing and amounts, we manipulated rainfall inputs to plots of native tallgrass prairie (Konza Prairie, Kansas, USA) over four growing seasons (1998-2001). Specifically, we altered the amounts and/or timing of growing season rainfall in a factorial combination that included two levels of rainfall amount (100% or 70% of naturally occurring rainfall quantity) and two temporal patterns of rain events (ambient timing or a 50% increase in length of dry intervals between events). The size of individual rain events in the altered timing treatment was adjusted so that the quantity of total growing season rainfall in the ambient and altered timing treatments was the same (i.e. fewer, but larger rainfall events characterized the altered timing treatment). Seasonal mean soil CO2 flux decreased by 8% under reduced rainfall amounts, by 13% under altered rainfall timing, and by 20% when both were combined (P<0.01). These changes in soil CO2 flux were consistent with observed changes in plant productivity, which was also reduced by both reduced rainfall quantity and altered rainfall timing. Soil CO2 flux was related to both soil temperature and soil water content in re
HertelD, StreckerT, Müller-HauboldH, LeuschnerC, GuoD (2013). Fine root biomass and dynamics in beech forests across a precipitation gradient — Is optimal resource partitioning theory applicable to water-limited mature trees? Journal of Ecology, 101, 1183-1200.https://doi.org/10.1111/1365-2745.12124URL [本文引用: 1]摘要 Optimal resource partitioning theory predicts that plants should increase the ratio between water absorbing and transpiring surfaces under short water supply. An increase in fine root mass and surface area relative to leaf area has frequently been found in herbaceous plants, but supporting evidence from mature trees is scarce and several results are contradictory.In 12 mature Fagus sylvatica forests across a precipitation gradient (820 540 mm yr 1), we tested several predictions of the theory by analysing the dependence of standing fine root biomass, fine root production and fine root morphology on mean annual precipitation (MAP), the precipitation of the study year, and stand structural and edaphic variables. The water storage capacity of the soil (WSC) was included as a covariable by comparing pairs of stands on sandy (lower WSC) and loam-richer soils (higher WSC).Fine root biomass, total fine root surface area, fine root production and the fine root : leaf biomass production ratio markedly increased with reduced MAP and precipitation in the study year, while WSC was only a secondary factor and stand structure had no effect.The precipitation effect on fine root biomass and production was more pronounced in stands on sandy soil with lower WSC, which had, at equal precipitation, a higher fine root biomass and productivity than stands on loam-richer soil.The high degree of allocational plasticity in mature F. sylvatica trees contrasts with a low morphological plasticity of the fine roots. On the more extreme sandy soils, a significant decrease in mean fine root diameter and increase in specific root area with decreasing precipitation were found; a similar effect was absent on the loam-richer soils.Synthesis. In support of optimal partitioning theory, mature Fagus sylvatica trees showed a remarkable allocational plasticity as a long-term response to significant precipitation reduction with a large increase in the size and productivity of the fine root system, while only minor adaptive modifications occurred in root morphology. More severe summer droughts in a future warmer climate may substantially alter the above-/below-ground C partitioning of this tree species with major implications for the forest C cycle.
[27]
HungateBA, van GroenigenKJ, SixJ, JastrowJJ, LuoYQ, GraaffMA, KesselC, OsenbergCW (2009). Assessing the effect of elevated CO2 on soil carbon: A comparison of four meta-analysis .Global Change Biology, 15, 2020-2034.https://doi.org/10.1016/j.amjcard.2004.01.038URL [本文引用: 1]摘要 Soil is the largest reservoir of organic carbon (C) in the terrestrial biosphere and soil C has a relatively long mean residence time. Rising atmospheric carbon dioxide (CO) concentrations generally increase plant growth and C input to soil, suggesting that soil might help mitigate atmospheric CO rise and global warming. But to what extent mitigation will occur is unclear. The large size of the soil C pool not only makes it a potential buffer against rising atmospheric CO, but also makes it difficult to measure changes amid the existing background. Meta-analysis is one tool that can overcome the limited power of single studies. Four recent meta-analyses addressed this issue but reached somewhat different conclusions about the effect of elevated CO on soil C accumulation, especially regarding the role of nitrogen (N) inputs. Here, we assess the extent of differences between these conclusions and propose a new analysis of the data. The four meta-analyses included different studies, derived different effect size estimates from common studies, used different weighting functions and metrics of effect size, and used different approaches to address nonindependence of effect sizes. Although all factors influenced the mean effect size estimates and subsequent inferences, the approach to independence had the largest influence. We recommend that meta-analysts critically assess and report choices about effect size metrics and weighting functions, and criteria for study selection and independence. Such decisions need to be justified carefully because they affect the basis for inference. Our new analysis, with a combined data set, confirms that the effect of elevated CO on net soil C accumulation increases with the addition of N fertilizers. Although the effect at low N inputs was not significant, statistical power to detect biogeochemically important effect sizes at low N is limited, even with meta-analysis, suggesting the continued need for long-term experiments.
[28]
HuxmanTE, SnyderKA, TissueD, LefflerAJ, OgleK, PockmanWT, SandquistDR, PottsDL, SchwinningS (2004). Precipitation pulses and carbon fluxes in semiarid and arid ecosystems .Oecologia, 141, 254-268.https://doi.org/10.1007/s00442-004-1682-4URL [本文引用: 1]
[29]
InglimaI, AlbertiG, BertoliniT, VaccariFP, GioliB, MigliettaF, CotrufoMF, PeressottiA (2009). Precipitation pulses enhance respiration of Mediterranean ecosystems: The balance between organic and inorganic components of increased soil CO2 efflux .Global Change Biology, 15, 1289-1301.https://doi.org/10.1111/gcb.2009.15.issue-5URL [本文引用: 1]
[30]
IPCC (Intergovernmental Panel on Climate Change) (2013). Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In: Stocker TF, Qin DPlattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y eds. Climate Change 2013: The Physical Science Basis . Cambridge University Press, Cambridge, UK. [本文引用: 2]
[31]
JiangH, DengQ, ZhouG, HuiD, ZhangD, LiuS, ChuG, LiJ (2013). Responses of soil respiration and its temperature/moisture sensitivity to precipitation in three subtropical forests in southern China .Biogeosciences, 10, 3963-3982.https://doi.org/10.5194/bg-10-3963-2013URL [本文引用: 1]摘要 Both long-term observation data and model simulations suggest an increasing chance of serious drought in the dry season and extreme flood in the wet season in southern China, yet little is known about how changes in precipitation pattern will affect soil respiration in the region. We conducted a field experiment to study the responses of soil respiration to precipitation manipulations - precipitation exclusion to mimic drought, double precipitation to simulate flood, and ambient precipitation as control (abbr. EP, DP and AP, respectively) - in three subtropical forests in southern China. The three forest sites include Masson pine forest (PF), coniferous and broad-leaved mixed forest (MF) and monsoon evergreen broad-leaved forest (BF). Our observations showed that altered precipitation strongly influenced soil respiration, not only through the well-known direct effects of soil moisture on plant and microbial activities, but also by modification of both moisture and temperature sensitivity of soil respiration. In the dry season, soil respiration and its temperature sensitivity, as well as fine root and soil microbial biomass, showed rising trends with precipitation increases in the three forest sites. Contrarily, the moisture sensitivity of soil respiration decreased with precipitation increases. In the wet season, different treatments showed different effects in three forest sites. The EP treatment decreased fine root biomass, soil microbial biomass, soil respiration and its temperature sensitivity, but enhanced soil moisture sensitivity in all three forest sites. The DP treatment significantly increased soil respiration, fine root and soil microbial biomass in the PF only, and no significant change was found for the soil temperature sensitivity. However, the DP treatment in the MF and BF reduced soil temperature sensitivity significantly in the wet season. Our results indicated that soil respiration would decrease in the three subtropical forests if soil moisture continues to decrease in the future. More rainfall in the wet season could have limited effect on the response of soil respiration to the rising of temperature in the BF and MF.
[32]
JiaoM, ShenWJ (2014). Effects of seasonal precipitation variation on litter-fall in lower subtropical evergreen broad-leaved forest .Journal of Tropical and Subtropical Botany, 22, 549-557.(in Chinese with English abstract) [焦敏, 申卫军 (2014). 模拟降水分配季节变化对南亚热带常绿阔叶林凋落物的影响 . 热带亚热带植物学报, 22, 549-557.] [本文引用: 3]
[33]
KnappAK, BeierC, BriskeDD, ClassenAT, LuoY, ReichsteinM, SmithMD, SmithSD, BellJE, FayPA, HeislerJL, LeavittSW, SherryR, SmithB, WengE (2008). Consequences of more extreme precipitation regimes for terrestrial ecosystems .Bioscience ,58, 811-821.https://doi.org/10.1641/B580908URL [本文引用: 3]摘要 Amplification of the hydrological cycle as a consequence of global warming is forecast to lead more extreme intra-annual precipitation regimes. characterized by larger rainfall events and longer intervals between events. We present a conceptual framework, based on past investigations and ecological theory, for predicting the consequences of this underappreciated aspect of climate change. We consider a brand range of terrestrial ecosystems that vary in their over-all mater balance. More extreme rainfall regimes are expected to increase the duration and severity of soil water water stress in mesic ecosystems as intervals between rainfall events increase. In contrast, xeric ecosystems may exhibit the opposite response to extreme events. Larger but less frequent rainfall events may result in proportional reductions in evaporative losses in xenic systems, and thus may lead to greater soil water availability Hydric (wetland) ecosystems are predicted to experience reduced periods of anoxia in response to prolonged intervals between rainfall events. Understanding these contingent effects of ecosystem canter balance is necessary for predicting how nun e intervals precipitation regimes will modify ecosystem processes and alter interactions with related global change drivers.
[34]
KnappAK, CiaisP, SmithMD (2017). Reconciling inconsistencies in precipitation-productivity relationships: Implications for climate change .New Phytologist, 214, 41-47.https://doi.org/10.1111/nph.14381URL [本文引用: 1]
[35]
KnappAK, HooverDL, WilcoxKR, AvolioML, KoernerSE, La PierreKJ, LoikME, LuoY, SalaOE, SmithMD (2015). Characterizing differences in precipitation regimes of extreme wet and dry years: Implications for climate change experiments .Global Change Biology, 21, 2624-2633.https://doi.org/10.1111/gcb.12888URLPMID:25652911 [本文引用: 3]摘要 Abstract Climate change is intensifying the hydrologic cycle and is expected to increase the frequency of extreme wet and dry years. Beyond precipitation amount, extreme wet and dry years may differ in other ways, such as the number of precipitation events, event size, and the time between events. We assessed 1614 long-term (100 /year) precipitation records from around the world to identify key attributes of precipitation regimes, besides amount, that distinguish statistically extreme wet from extreme dry years. In general, in regions where mean annual precipitation (MAP) exceeded 1000 /mm, precipitation amounts in extreme wet and dry years differed from average years by ~40% and 30%, respectively. The magnitude of these deviations increased to >60% for dry years and to >150% for wet years in arid regions (MAP99th percentile of all events); these occurred twice as often in extreme wet years compared to average years. In contrast, these large precipitation events were rare in extreme dry years. Less important for distinguishing extreme wet from dry years were mean event size and frequency, or the number of dry days between events. However, extreme dry years were distinguished from average years by an increase in the number of dry days between events. These precipitation regime attributes consistently differed between extreme wet and dry years across 12 major terrestrial ecoregions from around the world, from deserts to the tropics. Thus, we recommend that climate change experiments and model simulations incorporate these differences in key precipitation regime attributes, as well as amount into treatments. This will allow experiments to more realistically simulate extreme precipitation years and more accurately assess the ecological consequences. 2015 John Wiley & Sons Ltd.
KorichevaJ, GurevitchJ, MengersenK (2013). Handbook of Meta-Analysis in Ecology and Evolution . Princeton University Press, Princeton, USA. [本文引用: 1]
[38]
LiLH, ChenZZ (1998). Soil respiration in grassland communities in the world .Chinese Journal of Ecology, 17(4), 45-51.(in Chinese with English abstract) [李凌浩, 陈佐忠 (1998). 草地群落的土壤呼吸 . 生态学杂志, 17(4), 45-51.] [本文引用: 5]
[39]
LiuL, WangX, LajeunesseMJ, MiaoG, PiaoS, WanS, WuY, WangZ, YangS, LiP, DengM (2016). A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes .Global Change Biology, 22, 1394-1405.https://doi.org/10.1111/gcb.2016.22.issue-4URL [本文引用: 3]
[40]
LiuT, ZhangYX, XuZZ, ZhouGS, HouYH, LinL (2012). Effects of short-term warming and increasing precipitation on soil respiration of desert steppe of Inner Mongolia .Chinese Journal of Plant Ecology, 36, 1043-1053.(in Chinese with English abstract) [刘涛, 张永贤, 许振柱, 周广胜, 侯彦会, 林琳 (2012). 短期增温和增加降水对内蒙古荒漠草原土壤呼吸的影响 . 植物生态学报,36, 1043-1053.] [本文引用: 1]
[41]
LuM, ZhouX, YangQ, LiH, LuoYQ, FangCM, ChenJK, YangX, LiB (2013). Responses of ecosystem carbon cycle to experimental warming: A meta-analysis .Ecology, 94, 726-738.https://doi.org/10.1890/12-0279.1URL [本文引用: 1]摘要 Abstract Background/Question/Methods Global warming potentially alters terrestrial ecosystem carbon (C) cycle, feeding back to further climate warming. However, how ecosystem C cycle responds and feed backs to warming remain unclear. We here used a meta-analysis to quantify response ratios of 18 variables of ecosystem C cycle to experimental warming. Results/Conclusions Our results showed that warming stimulated gross ecosystem photosynthesis (GEP) by 15.7%, net primary production (NPP) by 4.4%, and above- and belowground plant C pools by 6.8% and 7.0%, respectively. The experimental warming also accelerated litter mass loss by 6.8%, soil respiration by 9.0%, and dissolved organic C leaching by 12.1%. In addition, the responses of some of those variables to experimental warming differed among the ecosystem types. The warming effects on C influx and efflux are roughly counterbalanced with each other, resulting in insignificant changes in litter and soil C contents, and net ecosystem exchange (NEE). The minor changes in soil C storage and NEE across ecosystems suggest that climate warming might not trigger strong carbon-climate feedback from terrestrial ecosystems. Our results are also potentially useful for parameterizing and benchmarking land surface models in terms of C cycle responses to climate warming.
[42]
LüWQ, WangSJ, LiuXM, RongL (2011). Influence of rainfall on soil respiration in Karst Urban green space .Earth and Environment, 39(2), 174-180.(in Chinese with English abstract) [吕文强, 王世杰, 刘秀明, 容丽 (2011). 喀斯特地区城市绿地土壤呼吸对降水变化的响应研究 . 地球与环境, 39(2), 174-180.] [本文引用: 1]
[43]
PengQ, QiYC, DongYS, HeYT, LiuXC, SunLJ, JiaJQ, JinZ (2012). Responses of carbon cycling key processes to precipitation changes in arid and semiarid grassland ecosystems: A review .Process in Geography, 31, 1510-1518.(in Chinese with English abstract) [彭琴, 齐玉春, 董云社, 何亚婷, 刘欣超, 孙良杰, 贾军强, 金钊 (2012). 干旱半干旱地区草地碳循环关键过程对降雨变化的响应 . 地理科学进展, 31, 1510-1518.] [本文引用: 1]
[44]
PollC, MarhanS, BackF, NiklausPA, KandelerE (2013). Field-sacle manipulation of soil temperature and precipitation change soil CO2 flux in a temperate agricultural ecosystem .Agriculture, Ecosystems and Environment, 165, 88-97.https://doi.org/10.1016/j.agee.2012.12.012URL [本文引用: 1]
[45]
RaichJW, SehlesingerWH (1992). The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate .Tullus, 44B, 81-99.
[46]
ReichmannLG, SalaOE, WhiteheadD (2014). Differential sensitivities of grassland structural components to changes in precipitation mediate productivity response in a desert ecosystem .Functional Ecology, 28, 1292-1298.https://doi.org/10.1111/1365-2435.12265URL [本文引用: 1]摘要 Summary In arid and semi-arid ecosystems, there are legacies of previous-year precipitation on current-year above-ground net primary production. We hypothesized that legacies of past precipitation occur through changes in tiller density, stolon density, tiller growth, axillary bud density and percentage of viable axillary buds. We examined the sensitivity to current- and previous-year precipitation of these grassland structural components in Bouteloua eriopoda, the dominant grass in the northern Chihuahuan Desert. We conducted a rainfall manipulation experiment consisting in 6180% reduced precipitation, ambient, +80% increased precipitation treatments that were subjected to one of five precipitation levels in the previous two years (6180% and 6150% reduced precipitation, ambient, +50% and +80% increased precipitation). The first two years preconditioned the experimental plots for year three, in which we created wet-to-dry and dry-to-wet transitions. Measurements were taken in year 3. We found that stolon density was the most sensitive to changes in precipitation and that percent-active buds were insensitive. We also found that past precipitation had a significant legacy on grassland structural components regardless of the precipitation received in the current year, and that the legacy occurs mostly through changes in stolon density. Here, we showed that there is a differential sensitivity of structural components to current and past precipitation and supported previous findings that vegetation structure is one of the controls of productivity during precipitation transitions.
[47]
ReichsteinM, BahnM, CiaisP, FrankD, MahechaMD, SeneviratneSI, ZscheischlerJ, BeerC, BuchmannN, FrankDC, PapaleD, RammigA, SmithP, ThonickeK, van der VeldeM, ViccaS, WalzA, WattenbachM (2013). Climate extremes and the carbon cycle .Nature, 500, 287-295.https://doi.org/10.1038/nature12350URL [本文引用: 1]
[48]
RischAC, FrankDA (2007). Effects of increased soil water avaulability on grassland ecosystem carbon dioxide fluxes .Biogeochemistry, 86, 91-103.https://doi.org/10.1007/s10533-007-9148-5URL摘要 There is considerable interest in how ecosystems will respond to changes in precipitation. Alterations in rain and snowfall are expected to influence the spatio-temporal patterns of plant and soil processes that are controlled by soil moisture, and potentially, the amount of carbon (C) exchanged between the atmosphere and ecosystems. Because grasslands cover over one third of the terrestrial landscape, understanding controls on grassland C processes will be important to forecast how changes in precipitation regimes will influence the global C cycle. In this study we examined how irrigation affects carbon dioxide (CO 2 ) fluxes in five widely variable grasslands of Yellowstone National Park during a year of approximately average growing season precipitation. We irrigated plots every 2/weeks with 25% of the monthly 30-year average of precipitation resulting in plots receiving approximately 150% of the usual growing season water in the form of rain and supplemented irrigation. Ecosystem CO 2 fluxes were measured with a closed chamber-system once a month from May-September on irrigated and unirrigated plots in each grassland. Soil moisture was closely associated with CO 2 fluxes and shoot biomass, and was between 1.6% and 11.5% higher at the irrigated plots (values from wettest to driest grassland) during times of measurements. When examining the effect of irrigation throughout the growing season (Mayeptember) across sites, we found that water additions increased ecosystem CO 2 fluxes at the two driest and the wettest sites, suggesting that these sites were water-limited during the climatically average precipitation conditions of the 2005 growing season. In contrast, no consistent responses to irrigation were detected at the two sites with intermediate soil moisture. Thus, the ecosystem CO 2 fluxes at those sites were not water-limited, when considering their responses to supplemental water throughout the whole season. In contrast, when we explored how the effect of irrigation varied temporally, we found that irrigation increased ecosystem CO 2 fluxes at all the sites late in the growing season (September). The spatial differences in the response of ecosystem CO 2 fluxes to irrigation likely can be explained by site specific differences in soil and vegetation properties. The temporal effects likely were due to delayed plant senescence that promoted plant and soil activity later into the year. Our results suggest that in Yellowstone National Park, above-normal amounts of soil moisture will only stimulate CO 2 fluxes across a portion of the ecosystem. Thus, depending on the topographic location, grassland CO 2 fluxes can be water-limited or not. Such information is important to accurately predict how changes in precipitation/soil moisture will affect CO 2 dynamics and how they may feed back to the global C cycle.
[49]
RochetteP, DesjardinsRL, PatteyE (1991). Spatial and temporal variability of soil respiration in agricultural fields .Canadian Journal of Soil Science, 71, 189-196.https://doi.org/10.4141/cjss91-018URL [本文引用: 1]摘要 ABSTRACT Chamber measurements of CO2 evolution were made on bare soil, and in maize (1988) and wheat (1989) crops in order to study the spatial and temporal variability of soil respiration (Rsoil). Semivariograms showed no definite structure of spatial auto-correlation on bare soil when measurements were made along transects. Spatial variability was shown to occur at a scale smaller than 15 cm. In a maize crop, Rsoil in the row was significantly higher than in the interrow when the soil surface was dry due to the contribution of plant roots. Under wet soil conditions, Rsoil in the interrow compacted by the tractor wheel was lower than on noncompacted soil and no significant difference was found between rows and interrows. This was due to the role of microbial activity which was dominant except in the compacted interrow where lower airfilled porosity caused lower Rsoil. The maximum Rsoil during the growing season coincided with the period of maximum growth of both crops. A post-rainfall Rsoil burst is also described. -from Authors
SchuurEAG, MatsonPA (2001). Net primary productivity and nutrient cycling across a mesic to wet precipitation gradient in Hawaiian montane forest .Oecologia, 128, 431-442.https://doi.org/10.1007/s004420100671URLPMID:24549913 [本文引用: 1]摘要 Abstract Variation in rainfall in humid tropical forests has the potential to alter patterns of primary productivity andnutrient cycling. Net primary productivity (NPP) and nutrient cycling were measured at six sites similar in temperature regime, parent material, ecosystem age, vegetation and topographical relief, while mean annual precipitation (MAP) varied from 2,200 toover 5,000 mm/year. Aboveground NPP declined by a factor of 2.2 with increased MAP across the precipitation gradient. Increased water availability in excess of plant demand is likely to have decreased the other resources for plant growth. Patterns of nutrient cycling and other factors that affect plant growth suggest that increased nutrient limitation in wetter sites could be the direct cause of the decline in NPP. Foliar nitrogen (N) and soil N availability decreased with increased precipitation, corresponding with the decrease in forest growth. In contrast, patterns of foliar and soil phosphorus (P) did not correspond with the decrease in growth; P availability was highest at either end of the precipitation gradient and lowest across the middle. Natural abundance of (15)N in foliage and soils decreased with increased precipitation, further supporting the idea that N availability declined. Decreased N availability was associated with a decrease in soil reduction-oxidation potentials. Oxygen limitation in soil microsites was a factor at all sites, but became increasingly widespread at higher MAP regimes. There was no strong evidence that soil oxygen availability, expressed in foliar (13)C values, directly limited plant growth. In addition foliar micronutrients either showed no change (Ca, Mg) or declined (Al, Fe) with increased MAP while soil pH was low but constant, suggesting that toxic elements in the soil solution were also not direct factors in decreased plant growth across the gradient. Thus, the decline in NPP with associated MAP appeared to be most directly associated with decreased N availability in these humid forests. Fluctuating anaerobic conditions that increased in intensity and duration with increased rainfall could be a mechanism that slows decomposition and N mineralization while concurrently increasing P solubility from soil mineral-bound pools.
[52]
SmithNG, RodgersVL, BrzostekER, KulmatiskiA, AvolioML, HooverDL, KoernerSE, GrantK, JentschA, FatichiS, NiyogiD (2014). Toward a better integration of biological data from precipitation manipulation experiments into Earth system models .Reviews of Geophysics, 52, 412-434.https://doi.org/10.1002/2014RG000458URL [本文引用: 1]
[53]
SottaED, VeldkampE, SchwendenmannL, GuimaraesBR, PaixaoRK, RuivoMDLP, Da CostaACL, MeirP (2007). Effects of an induced drought on soil carbon dioxide (CO2) efflux and soil CO2 production in an Eastern Amazonian rainforest, Brazil .Global Change Biology, 13, 2218-2229.https://doi.org/10.1111/j.1365-2486.2007.01416.xURL [本文引用: 1]摘要 Abstract In the next few decades, climate of the Amazon basin is expected to change, as a result of deforestation and rising temperatures, which may lead to feedback mechanisms in carbon (C) cycling that are presently unknown. Here, we report how a throughfall exclusion (TFE) experiment affected soil carbon dioxide (CO 2 ) production in a deeply weathered sandy Oxisol of Caxiuan00 (Eastern Amazon). Over the course of 2 years, we measured soil CO 2 efflux and soil CO 2 concentrations, soil temperature and moisture in pits down to 3m depth. Over a period of 2 years, TFE reduced on average soil CO 2 efflux from 4.3±0.1μmolCO 2 m 612 s 611 (control) to 3.2±0.1μmolCO 2 m 612 s 611 (TFE). The contribution of the subsoil (below 0.5m depth) to the total soil CO 2 production was higher in the TFE plot (28%) compared with the control plot (17%), and it did not differ between years. We distinguished three phases of drying after the TFE was started. The first phase was characterized by a translocation of water uptake (and accompanying root activity) to deeper layers and not enough water stress to affect microbial activity and/or total root respiration. During the second phase a reduction in total soil CO 2 efflux in the TFE plot was related to a reduction of soil and litter decomposers activity. The third phase of drying, characterized by a continuing decrease in soil CO 2 production was dominated by a water stress-induced decrease in total root respiration. Our results contrast to results of a drought experiment on clay Oxisols, which may be related to differences in soil water retention characteristics and depth of rooting zone. These results show that large differences exist in drought sensitivity among Amazonian forest ecosystems, which primarily seem to be affected by the combined effects of texture (affecting water holding capacity) and depth of rooting zone.
SuseelaA, DukesJ (2013). The responses of soil and rhizosphere respiration to simulated climatic changes vary by season .Ecology, 94, 403-413.https://doi.org/10.1890/12-0150.1URLPMID:23691659 [本文引用: 2]摘要 Responses of soil respiration (Rs) to anthropogenic climate change will affect terrestrial storage and, thus, feed back to warming. To provide insight into how warming and changes in precipitation regimes affect the rate and temperature sensitivity of Rs and rhizosphere respiration (Rr) across the year, we subjected a New England old-field ecosystem to four levels of warming and three levels of precipitation (ambient, drought, and wet treatments). We measured Rs and heterotrophic respiration (Rh) monthly (in areas of the plots with and without , respectively) and estimated Rr by calculating the difference in respiration between Rs and Rh. Even in this mesic ecosystem, Rs and Rr responded strongly to the precipitation treatments. Drought reduced Rs and Rr, both annually and during the growing season. Annual cumulative Rs responded nonlinearly to precipitation treatments; both drought and supplemental precipitation suppressed Rs compared to the ambient treatment. Warming increased Rs and Rr in spring and winter when soil moisture was optimal but decreased these rates in summer when moisture was limiting. Cumulative winter Rr increased by about 200% in the high warming (approximately 3.5 degrees C) treatment. The effect of climate treatments on the temperature sensitivity of Rs depended on the season. In the fall, the drought treatment decreased apparent Q10 relative to the other precipitation treatments. The responses of Rs to warming and altered precipitation were largely driven by changes in Rr. We emphasize the importance of incorporating realistic soil moisture responses into simulations of soil fluxes; the long-term effects of warming on --climate feedback will depend on future precipitation regimes. Our results highlight the nonlinear responses of soil respiration to soil moisture and, to our knowledge, quantify for the first time the loss of through winter rhizosphere respiration due to warming. While this additional loss is small relative to the cumulative annual flux in this system, such increases in rhizosphere respiration during the non-growing season could have greater consequences in ecosystems where they offset or reduce subsequent warming-induced gains in plant .
[56]
ThomeyML, CollinsSL, VargasR, JohnsonJE, BrownRF, NatvigDO, FriggensMT (2011). Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland .Global Change Biology, 17, 1505-1515.https://doi.org/10.1111/j.1365-2486.2010.02363.xURL [本文引用: 1]摘要 Abstract Precipitation regimes are predicted to become more variable with more extreme rainfall events punctuated by longer intervening dry periods. Water-limited ecosystems are likely to be highly responsive to altered precipitation regimes. The bucket model predicts that increased precipitation variability will reduce soil moisture stress and increase primary productivity and soil respiration in aridland ecosystems. To test this hypothesis, we experimentally altered the size and frequency of precipitation events during the summer monsoon (July through September) in 2007 and 2008 in a northern Chihuahuan Desert grassland in central New Mexico, USA. Treatments included (1) ambient rain, (2) ambient rain plus one 20/mm rain event each month, and (3) ambient rain plus four 5/mm rain events each month. Throughout two monsoon seasons, we measured soil temperature, soil moisture content, soil respiration ( R s), along with leaf-level photosynthesis ( A net), predawn leaf water potential (pd), and seasonal aboveground net primary productivity (ANPP) of the dominant C4 grass, Bouteloua eriopoda . Treatment plots receiving a single large rainfall event each month maintained significantly higher seasonal soil which corresponded with a significant increase in R s and ANPP of B. eriopoda when compared with plots receiving multiple small events. Because the strength of these patterns differed between years, we propose a modification of the bucket model in which both the mean and variance of soil water change as a consequence of interannual variability from 1 year to the next. Our results demonstrate that aridland ecosystems are highly sensitive to increased precipitation variability, and that more extreme precipitation events will likely have a positive impact on some aridland ecosystem processes important for the carbon cycle.
[57]
VargasR, CollinsSL, ThomeyM, JohnsonJE, BrownRF, NatvigDO, FriggensMT (2012). Precipitation variability and fire influence the temporal dynamics of soil CO2 efflux in an arid grassland . Global Change Biology, 18, 1401-1411.https://doi.org/10.1111/j.1365-2486.2011.02628.xURL [本文引用: 4]摘要 Climate models suggest that extreme rainfall events will become more common with increased atmospheric warming. Consequently, changes in the size and frequency of rainfall will influence biophysical drivers that regulate the strength and timing of soil CO2 efflux a major source of terrestrial carbon flux. We used a rainfall manipulation experiment during the summer monsoon season (July eptember) to vary both the size and frequency of precipitation in an arid grassland 2 years before and 2 years after a lightning-caused wildfire. Soil CO2 efflux rates were always higher under increased rainfall event size than under increased rainfall event frequency, or ambient precipitation. Although fire reduced soil CO2 efflux rates by nearly 70%, the overall responses to rainfall variability were consistent before and after the fire. The overall sensitivity of soil CO2 efflux to temperature (Q10) converged to 1.4, but this value differed somewhat among treatments especially before the fire. Changes in rainfall patterns resulted in differences in the periodicity of soil CO2 efflux with strong signals at 1, 8, and 30 days. Increased rainfall event size enhanced the synchrony between photosynthetically active radiation and soil CO2 efflux over the growing season before and after fire, suggesting a change in the temporal availability of substrate pools that regulate the temporal dynamics and magnitude of soil CO2 efflux. We conclude that arid grasslands are capable of rapidly increasing and maintaining high soil CO2 efflux rates in response to increased rainfall event size more than increased rainfall event frequency both before and after a fire. Therefore, the amount and pattern of multiple rain pulses over the growing season are crucial for understanding CO2 dynamics in burned and unburned water-limited ecosystems.
[58]
ViccaS, BahnM, EstiarteM, van LoonEE, VargasR, AlbertiG, AmbusP, ArainMA, BeierC, BentleyLP, BorkenW, BuchmannN, CollinsSL, de DatoG, DukesJS, EscolarC, FayP, GuidolottiG, HansonPJ, KahmenA, Kr?el-DulayG, Ladreiter-KnaussT, LarsenKS, Lellei-KovacsE, Lebrija-TrejosE, MaestreFT, MarhanS, MarshallM, MeirP, MiaoY, MuhrJ, NiklausPA, OgayaR, Pe?uelasJ, PollC, RustadLE, SavageK, SchindlbacherA, SchmidtIK, SmithAR, SottaED, SuseelaV, TietemaA, van GestelN, van StraatenO, WanS, WeberU, JanssensIA (2014). Can current moisture responses predict soil CO2 efflux under altered precipitation regimes? A synthesis of manipulation experiments .Biogeosciences, 11, 2991-3013.https://doi.org/10.5194/bg-11-2991-2014URL [本文引用: 1]
[59]
WangX, LiuLL, PiaoSL, JanssensIA, TangJW, LiuWX, ChiYG, WangJ, XuS (2014). Soil respiration under climate warming: Differential response of heterotrophic and autotrophic respiration .Global Change Biology, 20, 3229-3237.https://doi.org/10.1111/gcb.2014.20.issue-10URL [本文引用: 1]
[60]
WilcoxKR, BlairJM, SmithMD, KnappAK (2016). Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions? Ecology, 97, 561-568.URL [本文引用: 1]
[61]
WilcoxKR, ShiZ, GherardiLA, LemoineNP, KoernerSE, HooverDL, BorkE, ByrneKM, CahillJJ, CollinsSL, EvansS, GilgenAK, HolubP, JiangL, KnappAK, LeCainD, LiangJ, Garcia-PalaciosP, PenuelasJ, PockmanWT, SmithMD, SunS, WhiteSR, YahdjianL, ZhuK, LuoYQ (2017). Asymmetric responses of primary productivity to precipitation extremes: A synthesis of grassland precipitation manipulation experiments .Global Change Biology, 23, 4376-4385.https://doi.org/10.1111/gcb.13706URLPMID:28370946 [本文引用: 2]摘要 Abstract Climatic changes are altering Earth's hydrological cycle, resulting in altered precipitation amounts, increased inter-annual variability of precipitation, and more frequent extreme precipitation events. These trends will likely continue into the future, having substantial impacts on net primary productivity (NPP) and associated ecosystem services such as food production and carbon sequestration. Frequently, experimental manipulations of precipitation have linked altered precipitation regimes to changes in NPP. Yet, findings have been diverse and substantial uncertainty still surrounds generalities describing patterns of ecosystem sensitivity to altered precipitation. Additionally, we do not know whether previously observed correlations between NPP and precipitation remain accurate when precipitation changes become extreme. We synthesized results from 83 case studies of experimental precipitation manipulations in grasslands worldwide. We used meta-analytical techniques to search for generalities and asymmetries of aboveground NPP (ANPP) and belowground NPP (BNPP) responses to both the direction and magnitude of precipitation change. Sensitivity (i.e., productivity response standardized by the amount of precipitation change) of BNPP was similar under precipitation additions and reductions, but ANPP was more sensitive to precipitation additions than reductions; this was especially evident in drier ecosystems. Additionally, overall relationships between the magnitude of productivity responses versus the magnitude of precipitation change were saturating in form. The saturating form of this relationship was likely driven by ANPP responses to very extreme precipitation increases, although there were limited studies imposing extreme precipitation change and there was considerable variation among experiments. This highlights the importance of incorporating gradients of manipulations, ranging from extreme drought to extreme precipitation increases into future climate change experiments. Additionally, policy and land management decisions related to global change scenarios should consider how ANPP and BNPP responses may differ, and that ecosystem responses to extreme events might not be predicted from relationships found under moderate environmental changes. This article is protected by copyright. All rights reserved.
[62]
WuZ, DijkstraP, KochGW, Pe?uelasJ, HungateBA (2011). Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation .Global Change Biology, 17, 927-942.https://doi.org/10.1111/gcb.2010.17.issue-2URL [本文引用: 1]
[63]
XuMJ, WangHM, WenXF, ZhangT, DiYB, WangYD, WangJL, ChengCP, ZhangWJ (2017). The full annual carbon balance of a subtropical coniferous plantation is highly sensitive to autumn precipitation .Scientific Reports, 7, 10025. doi: 10.1038/s41598-017-10485-w.https://doi.org/10.1038/s41598-017-10485-wURL [本文引用: 1]
[64]
XuX, ShiZ, ChenX, LinY, NiuS, JiangL, LuoR, LuoY (2016). Unchanged carbon balance driven by equivalent responses of production and respiration to climate change in a mixed-grass prairie .Global Change Biology, 22, 1857-1866.https://doi.org/10.1111/gcb.13192URL [本文引用: 2]
[65]
ZhangLH, ChenYN, ZhaoRF, LiWH (2009). Impact of temperature and soil water content on soil respiration in temperate deserts, China .Chinese Journal of Plant Ecology, 33, 936-949.(in Chinese with English abstract) [张丽华, 陈亚宁, 赵锐峰, 李卫红 (2009). 温带荒漠中温度和土壤水分对土壤呼吸的影响 . 植物生态学报, 33, 936-949.]https://doi.org/10.3773/j.issn.1005-264x.2009.05.013URL [本文引用: 1]摘要 荒漠对气候变化具有高度敏感性,深刻认识和量化非生物因子对荒漠 生态系统土壤呼吸的影响具有重要意义.采用自动CO2通量系统(Li-8100)监测了梭梭(Haloxylon ammodendron)、假木贼(Anabasis aphylla)和盐穗木(Halostachys caapica)群落生长季土壤呼吸及温度、土壤含水量等,深入分析了水热因子对土壤呼吸的影响.土壤呼吸具有不对称的日格局,最小值出现在8:00,最 大值在12:00~14:00.土壤呼吸的季节格局与气温变化基本同步,最小值在生长季末期(10月),最大值在生长季中期(6~7月).梭梭、假木贼和 盐穗木群落生长季平均十壤呼吸速率分别为0.76、0.52和0.46~μmol CO2·m-2·s-1.气温对假木贼(51%)和盐穗木群落(65%)土壤呼吸季节变化的解释率高于梭梭(35%).梭梭、假木贼和盐穗木群落土壤呼吸 温度敏感性(Q10)逐渐增大,基础呼吸速率(R10)逐渐减小.剔除温度影响后,梭梭、假木贼群落土壤呼吸与土壤含水量呈显著的幂二次方函数关系,盐穗 木群落两者关系却明显减弱,未达到显著水平.气温、土壤含水量的二元方程均能解释群落土壤呼吸大部分的时间变异:梭梭群落71%~93%、假木贼群落 79%~82%、盐穗木群落70%~80%.人工模拟降水后土壤呼吸速率表现出降水后10 min减小、180 min时明显增加、达到最大值后再次衰减的现象.5和2.5 mm降水处理下的土壤呼吸速率最大值和其后的递减值高于对照处理,土壤呼吸增加、达到峰值和其后递减过程与5 cm土壤温度变化基本同步.
[66]
ZhouGS, JiaBR, HanGX, ZhouL (2008). Toward a general evaluation model for soil respiration (GEMSR) .Science in China Series C: Life Sciences, 51, 254-262.https://doi.org/10.1007/s11427-008-0030-zURLPMID:18246313 [本文引用: 1]摘要 Soil respiration is an important component of terrestrial carbon budget. Its accurate evaluation is essential to the study of terrestrial carbon source/sink. Studies on soil respiration at present mostly focus on the temporal variations and the controlling factors of soil respiration, but its spatial variations and controlling factors draw less attention. Moreover, the evaluation models for soil respiration at present include only the effects of water and heat factors, while the biological and soil factors controlling soil respiration and their interactions with water and heat factors have not been considered yet. These models are not able to accurately evaluate soil respiration in different vegetation/terrestrial ecosystems at different temporal and spatial scales. Thus, a general evaluation model for soil respiration (GEMSR) including the interacting meteorological (water and heat factors), soil nutrient and biological factors is suggested in this paper, and the basic procedure developing GEMSR and the research tasks of soil respiration in the future are also discussed.
[67]
ZhouX, ZhouL, NieY, FuY, DuZ, ShaoJ, ZhengZ, WangX (2016). Similar responses of soil carbon storage to drought and irrigation in terrestrial ecosystems but with contrasting mechanisms: A meta-analysis .Agriculture, Ecosystems & Environment, 228, 70-81.URL
Plants mediate the sensitivity of soil respiration to rainfall variability 2 2011
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
... 土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小.因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010).本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸.这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致.并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016). ...
Atmospheric warming and the amplification of precipitation extremes 1 2008
... 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013).多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015).未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005).同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013).全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013).作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013). ...
Ecosystem stability and compensatory effects in the Inner Mongolia grassland 1 2004
... 土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小.因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010).本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸.这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致.并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016). ...
Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling 2010
中国草原土壤呼吸作用研究进展 1 2010
... 土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小.因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010).本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸.这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致.并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016). ...
Precipitation manipulation experiments—Challenges and recommendations for the future 2 2012
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
... 土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小.因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010).本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸.这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致.并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016). ...
Site and temporal variation of soil respiration in European beech, Norway spruce, and Scots pine forests 2 2002
... 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013).多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015).未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005).同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013).全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013).作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013). ...
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
Responses of soil respiration to simulated precipitation pulses in semiarid steppe under different grazing regimes 1 2008
Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe 1 2009
Experimental manipulations of winter snow and summer rain influence ecosystem carbon cycling in a mixed-grass prairie, Wyoming, USA 1 2010
... 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013).多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015).未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005).同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013).全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013).作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013). ...
Experimental drought in a tropical rain forest increases soil carbon dioxide losses to the atmosphere 2 2010
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
... 降水变化主要通过改变土壤水分来改变生态系统碳循环过程, 结合图3和图4的结果我们发现, 土壤湿度是降水变化下驱动土壤呼吸变化的主导因子, 土壤湿度及由其引发的BNPP和土壤微生物碳库变化对土壤呼吸变化的解释度高达98%.土壤湿度一方面直接影响土壤呼吸, 另一方面通过影响ANPP、BNPP及MBC间接影响土壤呼吸(图4).ANPP并不直接影响土壤呼吸, 但其对地下根系及土壤微生物都有重要作用, 降水变化下ANPP对土壤呼吸的解释度高达59% (图3B).ANPP代表植物光合作用积累有机物的能力, 增加降水可以促进植物光合作用, 这一方面会直接导致根系活动加剧, 另一方面光合产物的增加导致土壤凋落物增加, 同时降水增加又会促进凋落物进入土壤(Knapp & Smith, 2001; Sponseller, 2007; Inglima et al., 2009; Cleveland et al., 2010; Smith et al., 2014).这为土壤微生物活动提供了底物, 促进土壤异养呼吸.同时, 土壤湿度增加会导致土壤微生物碳库增加, 进一步促进异养呼吸.尽管土壤湿度增加会抑制BNPP, 但ANPP增加又会促进BNPP及土壤微生物活力, 最终导致土壤总呼吸增加.反之, 减少降水会抑制植物光合作用及微生物活动进而抑制土壤呼吸, 并且过度干旱会造成植物根系死亡(Harper et al., 2005; Sotta et al., 2007), 这一方面直接导致根系呼吸降低, 另一方面会通过影响根系对水分的吸收利用来影响植物光合作用, 形成一系列正反馈过程降低土壤呼吸. ...
Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model 1 2000
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
Increasing drought under global warming in observations and models 1 2013
... 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013).多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015).未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005).同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013).全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013).作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013). ...
Rain- induced changes in soil CO2 flux and microbial community composition in a tropical forest of China 1 2017
... 尽管meta分析在定量的综述研究中具有极其重要的意义, 但有一定的局限性.纵观本研究的数据库, 我们可以发现, 多数降水控制实验开展于草地生态系统, 由于草地生态系统较为干旱, 水分是主要限制因子, 故对增加降水的响应可能会表现得较为敏感(Knapp et al., 2017; Wilcox et al., 2017).而森林生态系统由于降水量较高, 对降水变化的响应较弱, 所以不同类型生态系统所占比例的不同会导致整体结果的变化.另外, 热带、亚热带森林有明显的干湿季(焦敏和申卫军, 2014), 季节性降水格局差异会导致土壤呼吸对降水变化的差异性响应(Deng et al., 2017; Xu et al., 2017), 这些差异很难在meta分析中捕捉到.因此全球meta分析的主要目的是提供普遍规律, 具体气候条件下和生态系统类型的独特响应过程机理还需实验验证. ...
Few multiyear precipitation-reduction experiments find a shift in the productivity-precipitation relationship 1 2016
... 土壤呼吸对降水变化的响应受土壤本身的理化性质、土壤结构、植被类型、气候类型等影响(Vicca et al., 2014).在较为干旱的荒漠、草地或灌丛生态系统中, 降水是主要的限制因子, 增加降水土壤呼吸增加40%以上, 而减少降水仅降低了20%左右土壤呼吸; 而在较为湿润森林生态系统中, 增加降水引起的土壤呼吸增量很小(<10%), 而减少降水降低了土壤呼吸的16%.已有研究结果表明, 在较为干旱的地带, NEE和植物生产力等碳循环过程对增加降水的响应敏感性高于减少降水(Flanagan et al., 2002; Knapp & Smith, 2001; Knapp et al., 2017), 本文的研究结果与之一致.而在考虑总体土壤呼吸对降水的响应时, 增雨增加了土壤呼吸量的49%, 减雨减少了土壤呼吸量的21%, 并且根据图5, 土壤呼吸对增雨的敏感性高于减雨处理, 这与Wu等(2011)和Zhou等(2016)的研究结果一致.导致这种规律的原因可能是在较为干旱的环境中, 土壤以及植物对干旱有一定的抵抗力, 且继续减少降水会提高植物及土壤微生物的水分利用效率(Gutschick & BassiriRad, 2003; Huxman et al., 2004; Liu et al., 2016), 从而使得减雨的效应略小; 而增加降水引起土壤湿度增加的同时, 导致土壤微生物及其他养分元素的变化, 进而更大程度地促进土壤呼吸的增长(Flanagan et al., 2002; Knapp et al., 2017).而在较为湿润的环境中, 水分不是主要的限制因子, 增雨不能进一步提高土壤呼吸, 甚至过多降水会导致土壤透气性下降(Knapp et al., 2008; Estiarte et al., 2016)及养分淋溶流失严重(Schuur & Matson, 2001; Schuur, 2003), 导致土壤呼吸下降. ...
Effect of manipulated rainfall on root production and plant belowground dry mass of different grassland ecosystems 1 2009
... 土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小.因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010).本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸.这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致.并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016). ...
Robust spatially aggregated projections of climate extremes 1 2013
... 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013).多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015).未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005).同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013).全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013).作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013). ...
Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland 3 2002
... 土壤呼吸对降水变化的响应受土壤本身的理化性质、土壤结构、植被类型、气候类型等影响(Vicca et al., 2014).在较为干旱的荒漠、草地或灌丛生态系统中, 降水是主要的限制因子, 增加降水土壤呼吸增加40%以上, 而减少降水仅降低了20%左右土壤呼吸; 而在较为湿润森林生态系统中, 增加降水引起的土壤呼吸增量很小(<10%), 而减少降水降低了土壤呼吸的16%.已有研究结果表明, 在较为干旱的地带, NEE和植物生产力等碳循环过程对增加降水的响应敏感性高于减少降水(Flanagan et al., 2002; Knapp & Smith, 2001; Knapp et al., 2017), 本文的研究结果与之一致.而在考虑总体土壤呼吸对降水的响应时, 增雨增加了土壤呼吸量的49%, 减雨减少了土壤呼吸量的21%, 并且根据图5, 土壤呼吸对增雨的敏感性高于减雨处理, 这与Wu等(2011)和Zhou等(2016)的研究结果一致.导致这种规律的原因可能是在较为干旱的环境中, 土壤以及植物对干旱有一定的抵抗力, 且继续减少降水会提高植物及土壤微生物的水分利用效率(Gutschick & BassiriRad, 2003; Huxman et al., 2004; Liu et al., 2016), 从而使得减雨的效应略小; 而增加降水引起土壤湿度增加的同时, 导致土壤微生物及其他养分元素的变化, 进而更大程度地促进土壤呼吸的增长(Flanagan et al., 2002; Knapp et al., 2017).而在较为湿润的环境中, 水分不是主要的限制因子, 增雨不能进一步提高土壤呼吸, 甚至过多降水会导致土壤透气性下降(Knapp et al., 2008; Estiarte et al., 2016)及养分淋溶流失严重(Schuur & Matson, 2001; Schuur, 2003), 导致土壤呼吸下降. ...
... ), 从而使得减雨的效应略小; 而增加降水引起土壤湿度增加的同时, 导致土壤微生物及其他养分元素的变化, 进而更大程度地促进土壤呼吸的增长(Flanagan et al., 2002; Knapp et al., 2017).而在较为湿润的环境中, 水分不是主要的限制因子, 增雨不能进一步提高土壤呼吸, 甚至过多降水会导致土壤透气性下降(Knapp et al., 2008; Estiarte et al., 2016)及养分淋溶流失严重(Schuur & Matson, 2001; Schuur, 2003), 导致土壤呼吸下降. ...
... 同样, 土壤呼吸对水分的敏感性受到本底降雨量的影响.由图5的结果我们发现土壤呼吸对水分的敏感性在增雨和减雨处理时都随着本底降雨量的增加而逐渐下降(图5C、5F), 这一结果与大多数研究结果(Jiang et al., 2013; Liu et al., 2016)一致.一般来说, 在较为湿润(年降水量较高)的地带, 土壤呼吸对水分的敏感性较低.增加降水只能微弱地促进土壤呼吸, 甚至过量的水分添加会降低土壤通透性, 导致土壤氧分不足而抑制土壤呼吸, 此时, 适当地移除部分水分有可能促进土壤呼吸(Rocette et al., 1991; Zhou et al., 2008).而在原本就很干旱(年降水量较少)的地带, 即使微小的降水也会引起土壤呼吸极大的响应(Knapp et al., 2008), 但减雨对较为干旱的生态系统的影响说法并不一致.有的研究认为在干旱环境中继续减少降水会导致土壤呼吸下降(刘涛等, 2012; Hertel et al., 2013); 但也有研究指出, 干旱生态系统对干旱具有一定的抵抗力和适应性, 移除一定的雨量并不一定会影响土壤呼吸(Knapp & Smith, 2001; Flanagan et al., 2002).本研究中, 在减雨处理下, 土壤呼吸对土壤湿度的响应在降水量超过1 000 mm的梯度下并未呈现显著的线性关系.并且, 由于在降水量低于500 mm的环境中开展的减雨控制实验较少, 无法进一步印证以上结论.而Liu等(2016)用干旱指数表征环境湿润度时, 也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱, 对减雨的响应随环境湿润度变化不明显. ...
腾格里沙漠东南缘生物结皮土壤呼吸对水热因子变化的响应 2 2017
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
... 土壤呼吸对降水变化的响应主要是因为降水改变土壤湿度和土壤温度(Aanderud et al., 2011; Beier et al., 2012; Vicca et al., 2014), 而Liu等(2016)通过研究降水改变对土壤呼吸及Q10的影响发现, 降水引起的土壤温度改变很小.因此降水变化对土壤呼吸的改变主要是通过直接改变土壤含水量以及间接影响土壤微生物活性或植物生理作用来影响土壤呼吸(鲍芳和周广胜, 2010).本文结果表明, 在全球水平上土壤呼吸对降水量响应的普遍规律是: 增加降水促进土壤呼吸, 减少降水抑制土壤呼吸.这与大多数单个站点的研究结果(张丽华等, 2009; Wu et al., 2011; 刘涛等, 2012; Zhou et al., 2016; 管超等, 2017)一致.并且, 土壤呼吸增加或减少的量与降水量变化呈现一致的线性关系, 即增加或减少的降水量越多, 引起的土壤呼吸变化越大(Bai et al., 2004; Fiala et al., 2009; Wu et al., 2011; Liu et al., 2016). ...
Extreme events as shaping physiology, ecology, and evolution of plants: Toward a unified definition and evaluation of their consequences 1 2003
... 土壤呼吸对降水变化的响应受土壤本身的理化性质、土壤结构、植被类型、气候类型等影响(Vicca et al., 2014).在较为干旱的荒漠、草地或灌丛生态系统中, 降水是主要的限制因子, 增加降水土壤呼吸增加40%以上, 而减少降水仅降低了20%左右土壤呼吸; 而在较为湿润森林生态系统中, 增加降水引起的土壤呼吸增量很小(<10%), 而减少降水降低了土壤呼吸的16%.已有研究结果表明, 在较为干旱的地带, NEE和植物生产力等碳循环过程对增加降水的响应敏感性高于减少降水(Flanagan et al., 2002; Knapp & Smith, 2001; Knapp et al., 2017), 本文的研究结果与之一致.而在考虑总体土壤呼吸对降水的响应时, 增雨增加了土壤呼吸量的49%, 减雨减少了土壤呼吸量的21%, 并且根据图5, 土壤呼吸对增雨的敏感性高于减雨处理, 这与Wu等(2011)和Zhou等(2016)的研究结果一致.导致这种规律的原因可能是在较为干旱的环境中, 土壤以及植物对干旱有一定的抵抗力, 且继续减少降水会提高植物及土壤微生物的水分利用效率(Gutschick & BassiriRad, 2003; Huxman et al., 2004; Liu et al., 2016), 从而使得减雨的效应略小; 而增加降水引起土壤湿度增加的同时, 导致土壤微生物及其他养分元素的变化, 进而更大程度地促进土壤呼吸的增长(Flanagan et al., 2002; Knapp et al., 2017).而在较为湿润的环境中, 水分不是主要的限制因子, 增雨不能进一步提高土壤呼吸, 甚至过多降水会导致土壤透气性下降(Knapp et al., 2008; Estiarte et al., 2016)及养分淋溶流失严重(Schuur & Matson, 2001; Schuur, 2003), 导致土壤呼吸下降. ...
Experimental summer drought reduces soil CO2 effluxes and DOC leaching in Swiss grassland soils along an elevational gradient 1 2014
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
Separating root and soil microbial contributions to soil respiration: A review of methods and observations 1 2000
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem 2 2005
... 近年来, 人类活动导致大气中的CO2浓度升高, 引起了全球升温和降水格局的改变(Borken et al., 2002; Chimner et al., 2010; IPCC, 2013).多个气候模型预测结果一致显示, 未来气候变化会带来全球或局部降水格局的变化(Fischer et al., 2013; Knapp et al., 2015).未来降水将在高纬度和部分热带地区增加, 而在中低纬度和部分亚热带地区减少(Harper et al., 2005).同时, 极端降水事件的发生频率增加, 降雨强度和降雨时间分布进一步变化(Thomey et al., 2011; 彭琴等, 2012; IPCC, 2013).全球变暖的背景不断促成干旱化趋势加重, 导致未来大多数地区面临严重和大面积的干旱(Allan & Soden, 2008; Dai, 2013).作为驱动生态过程的关键影响因子, 降水量的变化将改变陆地生态系统碳循环, 对陆地生态系统源汇功能带来巨大的影响(Knapp et al., 2008; Reichstein et al., 2013). ...
... 降水变化主要通过改变土壤水分来改变生态系统碳循环过程, 结合图3和图4的结果我们发现, 土壤湿度是降水变化下驱动土壤呼吸变化的主导因子, 土壤湿度及由其引发的BNPP和土壤微生物碳库变化对土壤呼吸变化的解释度高达98%.土壤湿度一方面直接影响土壤呼吸, 另一方面通过影响ANPP、BNPP及MBC间接影响土壤呼吸(图4).ANPP并不直接影响土壤呼吸, 但其对地下根系及土壤微生物都有重要作用, 降水变化下ANPP对土壤呼吸的解释度高达59% (图3B).ANPP代表植物光合作用积累有机物的能力, 增加降水可以促进植物光合作用, 这一方面会直接导致根系活动加剧, 另一方面光合产物的增加导致土壤凋落物增加, 同时降水增加又会促进凋落物进入土壤(Knapp & Smith, 2001; Sponseller, 2007; Inglima et al., 2009; Cleveland et al., 2010; Smith et al., 2014).这为土壤微生物活动提供了底物, 促进土壤异养呼吸.同时, 土壤湿度增加会导致土壤微生物碳库增加, 进一步促进异养呼吸.尽管土壤湿度增加会抑制BNPP, 但ANPP增加又会促进BNPP及土壤微生物活力, 最终导致土壤总呼吸增加.反之, 减少降水会抑制植物光合作用及微生物活动进而抑制土壤呼吸, 并且过度干旱会造成植物根系死亡(Harper et al., 2005; Sotta et al., 2007), 这一方面直接导致根系呼吸降低, 另一方面会通过影响根系对水分的吸收利用来影响植物光合作用, 形成一系列正反馈过程降低土壤呼吸. ...
The meta-analysis of response ratios in experimental ecology 2 1999
... 采用反映降水变化对土壤呼吸效应的反应比值(response ratios, RR)作为效应值(Hedges et al., 1999).单个实验研究的效应值计算公式如下: ...
... 由于反应比的自然对数值偏倚较小、样本近似成正态分布, 故反应比的自然对数值非常适于作为meta分析的测量值(Hedges et al., 1999; Koricheva et al., 2013; Lu et al., 2013).综合所有的单个研究, 运用meta分析方法计算全球水平上土壤呼吸对降水变化的响应, 使用随机效应模型, 模型计算权重采用方差的倒数.总体的RR和置信区间(CI)计算运用软件METAWIN 2.1 (Sinauer Associates, Sunderland, USA).当95%的置信区间(CI)与0不重叠, 认为在统计学意义上显著. ...
Terrestrial ecosystem carbon dynamics and climate feedbacks 1 2008
... 土壤呼吸是指未扰动土壤中产生CO2的所有代谢, 主要包括自养呼吸(植物根系呼吸)和异养呼吸(土壤微生物和土壤动物呼吸)两个组分(Raich & Sehlesinger, 1992; Hanson et al., 2000; Wang et al., 2014).土壤呼吸是土壤中有机质输出的主要形式, 是土壤碳库向大气释放CO2的重要途径, 占全球陆地生态系统总呼吸的60%-90% (Aanderud et al., 2011; Hagedorn & Joos, 2014), 在调控区域和全球尺度的碳循环上起着十分关键的作用. 土壤呼吸同时受到土壤孔隙度、温度、湿度、氧气含量和土壤pH值等非生物因子和土壤微生物、植物根系等生物因子共同影响(李凌浩和陈佐忠, 1998; 刘涛等, 2012; Vargas et al., 2012; Vicca et al., 2014), 全球降水格局的改变对土壤呼吸的作用至关重要, 甚至会进一步影响全球气候变化的进程(Cox et al., 2000; Heimann & Reichstein, 2008; Wilcox et al., 2016; Xu et al., 2016).为了探究土壤呼吸对降水变化的响应情况, 全球各地开展了大量改变降水的控制实验, 覆盖不同气候带、生态系统类型等, 充分模拟了降水变化对土壤呼吸的影响, 并逐渐取得了一些重要的成果(Borken et al., 2002; Risch & Frank, 2007; 张丽华等, 2009; Cleveland et al., 2010; 吕文强等, 2011; Poll et al., 2013; Suseela & Dukes, 2013; 管超等, 2017).由于不同研究的区域条件和研究方法等的差异, 土壤呼吸对降水变化的响应研究并未得出一致结论, 所以, 一些关于全球降水控制实验的整合分析应运而生.Wu等(2011)和Zhou等(2016)整合了全球降水控制实验对主要陆地生态系统碳循环过程的影响, 发现增加降水对全球碳循环过程普遍促进, 而减少降水产生相反的作用.Liu等(2016)则针对性地整合了全球降水格局变化对土壤呼吸的两个组分和温度敏感性的影响, 探究了土壤湿度、土壤呼吸及土壤温度敏感性(Q10)对降水变化的响应, 发现了土壤湿度和干旱指数的主导作用.但并没有进一步探究降水变化下土壤呼吸响应的过程机理, 且对于背景降水量和温度怎样影响土壤呼吸的水分敏感性也不清楚.一般来说, 降水变化下, 土壤湿度通过直接影响根和微生物的生理学过程间接影响底物供应和CO2扩散来影响土壤呼吸(Beier et al., 2012; Vicca et al., 2014; Liu et al., 2016), 且土壤呼吸对水分的依赖性会随着环境中的水热限制因子的变化而改变.因此, 本研究在前人的研究成果上搜集了更为全面的关于土壤呼吸的降水控制实验的数据库, 共129篇文献, 利用整合分析的方法, 探究了降水变化下驱动土壤呼吸的主要因子及过程机理, 以期为未来全球气候变化背景下陆地生态系统碳平衡研究奠定理论基础和数据支撑. ...
Fine root biomass and dynamics in beech forests across a precipitation gradient — Is optimal resource partitioning theory applicable to water-limited mature trees? 1 2013
... 同样, 土壤呼吸对水分的敏感性受到本底降雨量的影响.由图5的结果我们发现土壤呼吸对水分的敏感性在增雨和减雨处理时都随着本底降雨量的增加而逐渐下降(图5C、5F), 这一结果与大多数研究结果(Jiang et al., 2013; Liu et al., 2016)一致.一般来说, 在较为湿润(年降水量较高)的地带, 土壤呼吸对水分的敏感性较低.增加降水只能微弱地促进土壤呼吸, 甚至过量的水分添加会降低土壤通透性, 导致土壤氧分不足而抑制土壤呼吸, 此时, 适当地移除部分水分有可能促进土壤呼吸(Rocette et al., 1991; Zhou et al., 2008).而在原本就很干旱(年降水量较少)的地带, 即使微小的降水也会引起土壤呼吸极大的响应(Knapp et al., 2008), 但减雨对较为干旱的生态系统的影响说法并不一致.有的研究认为在干旱环境中继续减少降水会导致土壤呼吸下降(刘涛等, 2012; Hertel et al., 2013); 但也有研究指出, 干旱生态系统对干旱具有一定的抵抗力和适应性, 移除一定的雨量并不一定会影响土壤呼吸(Knapp & Smith, 2001; Flanagan et al., 2002).本研究中, 在减雨处理下, 土壤呼吸对土壤湿度的响应在降水量超过1 000 mm的梯度下并未呈现显著的线性关系.并且, 由于在降水量低于500 mm的环境中开展的减雨控制实验较少, 无法进一步印证以上结论.而Liu等(2016)用干旱指数表征环境湿润度时, 也发现土壤呼吸对增雨的响应随着环境湿润度的增加而减弱, 对减雨的响应随环境湿润度变化不明显. ...
Assessing the effect of elevated CO2 on soil carbon: A comparison of four meta-analysis 1 2009
Precipitation pulses and carbon fluxes in semiarid and arid ecosystems 1 2004
... 土壤呼吸对降水变化的响应受土壤本身的理化性质、土壤结构、植被类型、气候类型等影响(Vicca et al., 2014).在较为干旱的荒漠、草地或灌丛生态系统中, 降水是主要的限制因子, 增加降水土壤呼吸增加40%以上, 而减少降水仅降低了20%左右土壤呼吸; 而在较为湿润森林生态系统中, 增加降水引起的土壤呼吸增量很小(<10%), 而减少降水降低了土壤呼吸的16%.已有研究结果表明, 在较为干旱的地带, NEE和植物生产力等碳循环过程对增加降水的响应敏感性高于减少降水(Flanagan et al., 2002; Knapp & Smith, 2001; Knapp et al., 2017), 本文的研究结果与之一致.而在考虑总体土壤呼吸对降水的响应时, 增雨增加了土壤呼吸量的49%, 减雨减少了土壤呼吸量的21%, 并且根据图5, 土壤呼吸对增雨的敏感性高于减雨处理, 这与Wu等(2011)和Zhou等(2016)的研究结果一致.导致这种规律的原因可能是在较为干旱的环境中, 土壤以及植物对干旱有一定的抵抗力, 且继续减少降水会提高植物及土壤微生物的水分利用效率(Gutschick & BassiriRad, 2003; Huxman et al., 2004; Liu et al., 2016), 从而使得减雨的效应略小; 而增加降水引起土壤湿度增加的同时, 导致土壤微生物及其他养分元素的变化, 进而更大程度地促进土壤呼吸的增长(Flanagan et al., 2002; Knapp et al., 2017).而在较为湿润的环境中, 水分不是主要的限制因子, 增雨不能进一步提高土壤呼吸, 甚至过多降水会导致土壤透气性下降(Knapp et al., 2008; Estiarte et al., 2016)及养分淋溶流失严重(Schuur & Matson, 2001; Schuur, 2003), 导致土壤呼吸下降. ...
Precipitation pulses enhance respiration of Mediterranean ecosystems: The balance between organic and inorganic components of increased soil CO2 efflux 1 2009
... 降水变化主要通过改变土壤水分来改变生态系统碳循环过程, 结合图3和图4的结果我们发现, 土壤湿度是降水变化下驱动土壤呼吸变化的主导因子, 土壤湿度及由其引发的BNPP和土壤微生物碳库变化对土壤呼吸变化的解释度高达98%.土壤湿度一方面直接影响土壤呼吸, 另一方面通过影响ANPP、BNPP及MBC间接影响土壤呼吸(图4).ANPP并不直接影响土壤呼吸, 但其对地下根系及土壤微生物都有重要作用, 降水变化下ANPP对土壤呼吸的解释度高达59% (图3B).ANPP代表植物光合作用积累有机物的能力, 增加降水可以促进植物光合作用, 这一方面会直接导致根系活动加剧, 另一方面光合产物的增加导致土壤凋落物增加, 同时降水增加又会促进凋落物进入土壤(Knapp & Smith, 2001; Sponseller, 2007; Inglima et al., 2009; Cleveland et al., 2010; Smith et al., 2014).这为土壤微生物活动提供了底物, 促进土壤异养呼吸.同时, 土壤湿度增加会导致土壤微生物碳库增加, 进一步促进异养呼吸.尽管土壤湿度增加会抑制BNPP, 但ANPP增加又会促进BNPP及土壤微生物活力, 最终导致土壤总呼吸增加.反之, 减少降水会抑制植物光合作用及微生物活动进而抑制土壤呼吸, 并且过度干旱会造成植物根系死亡(Harper et al., 2005; Sotta et al., 2007), 这一方面直接导致根系呼吸降低, 另一方面会通过影响根系对水分的吸收利用来影响植物光合作用, 形成一系列正反馈过程降低土壤呼吸. ...
Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In: Stocker TF, Qin DPlattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y eds. Climate Change 2013: The Physical Science Basis 2 2013
 , 牛书丽
, 牛书丽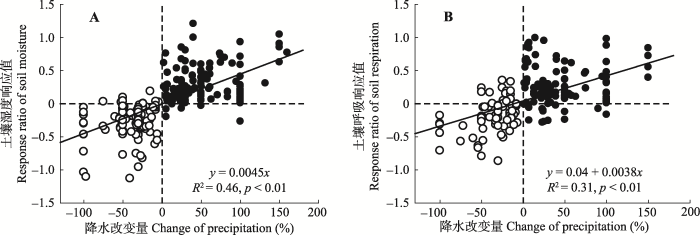 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT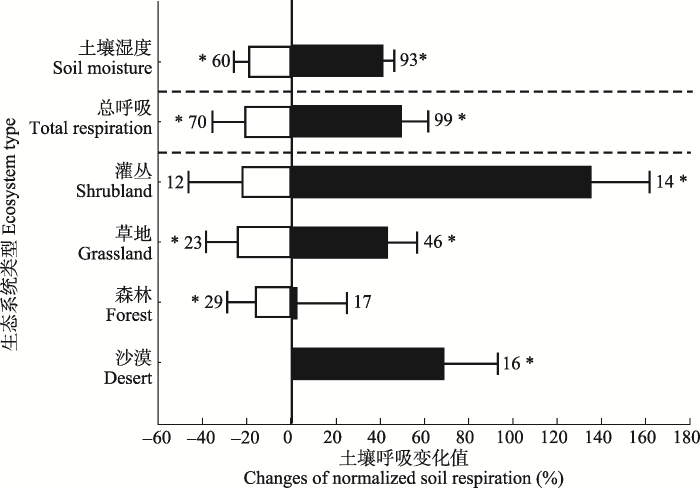 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT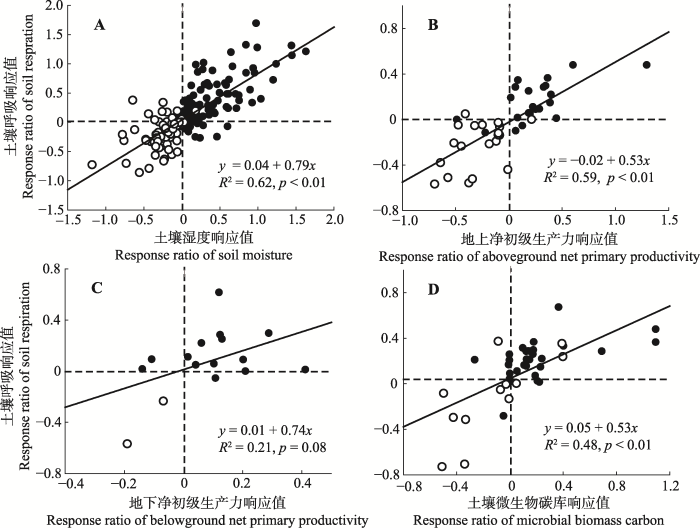 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT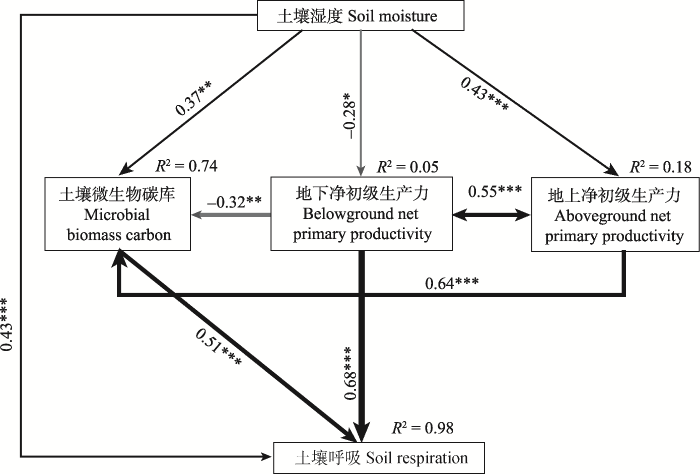 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT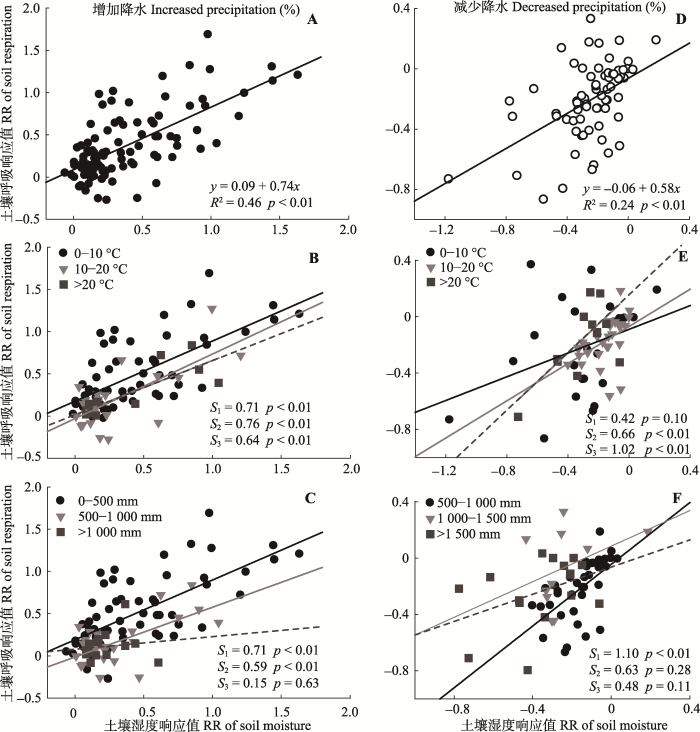 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}