
Relationships between species diversity or community structure and productivity of woody-plants in a broad-leaved Korean pine forest in Jiaohe, Jilin, China
TANLing-Zhao通讯作者:
收稿日期:2016-10-17
接受日期:2017-10-23
网络出版日期:2017-11-10
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Methods We used the data of 10 973 woody-plants in a 11.76 hm2 large sample plot and analyzed the correlations between 7 different indices of species diversity or community structure and productivity. Structural equation model was used to compare the effects of species diversity and community structure on productivity.
Important findings The results showed that: (1) Both species diversity and community structure had significant effects on productivity when they were considered separately in linear regression analysis, i.e. species evenness was negatively correlated with productivity, the Shannon index of community structure was positively correlated with productivity and the Gini index was negatively correlated with productivity. (2) In the structural equation model, when simultaneously considered, community structure had stronger influence on productivity than species diversity. Our research suggests that, the effects of community structure on productivity are greater than species diversity and it is important to increase community structure complexity to improve forest productivity during forest management.
Keywords:
-->0
PDF (926KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
多样性-生产力关系是森林可持续经营的重要论题(贺金生等, 2003; Naeem et al., 2009)。大量研究表明混交林比纯林有更高的生产力(Zhang et al., 2012; Maire et al., 2013; Pretzsch, 2014; Forrester & Bauhus, 2016), 增加物种多样性能提高生产力水平(Tilman et al., 1996, 2006; Loreau & Hector, 2001; Hooper et al., 2005; Ruijven & Berendse, 2005)。对于这样的结果, 生态位互补效应是一个主要解释(Loreau & Hector, 2001)。生态位互补效应理论认为不同物种在资源利用上存在差异(Tilman et al., 1997), 或者物种之间具有相互促进作用(Bertness & Leonard, 1997), 因此增加物种多样性可以优化生态系统功能。然而在实际调查中, 物种多样性并不能直接反映不同物种在生态位利用上的差异, 并且生态位互补效应成因复杂, 不仅与种间差异有关, 而且受到多种生物因素(种间作用、种内作用)和非生物因素(土壤、光照等)的影响, 因而导致验证生态位互补效应的证据不足(Chesson, 2000; Clark, 2010)。
近年来, 群落结构对生产力的影响逐渐成为研究热点(Lei et al., 2008; Zhang & Chen, 2015; D?nescu et al., 2016)。群落结构变异反映了研究区内个体的大小组成变化, 其可能来自于不同物种间的固有差异, 也可能由同种或异种个体间的不对称竞争产生(Coomes et al., 2009; Clark, 2010; Morin et al., 2011; Zhang et al., 2012)。在群落水平, 复杂的结构组成使得树木在空间中形成多元配置, 从而提高了资源利用效率(Yachi & Loreau, 2007)。因此, 可以认为群落结构变异在一定程度上反映了物种在资源利用上的生态位互补, 这是促进生产力的一个重要 机制。
在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(Lei et al., 2008; Zhang & Chen, 2015; D?nescu et al., 2016), 但也有结果表现出不相关或负相关(Liang et al., 2007; Long & Shaw, 2010; Ryan et al., 2010; Bourdier et al., 2016): Lei等(2008)发现在以云杉(Picea sp.)为主的森林中, 群落结构变异和树木生长存在正相关关系。D?nescu等(2016)验证了群落结构变异对德国西南部异龄混交林的生长有促进作用。Liang等(2007)发现在以Pseudotsuga menziesii和Tsuga heterophylla为主的群落中, 结构变异大的样方呈现出与生产力的负相关关系。Long和Shaw (2010)发现在Pinus ponderosa林中, 群落结构与生产力并没有显著的相关关系。甚至有研究直接指出群落结构对生产力的影响比物种多样性更强(D?nescu et al., 2016)。综上来看, 群落结构与生产力的关系仍需进一步讨论, 且物种多样性和群落结构对生产力影响的相对大小究竟如何, 亟待阐明。
阔叶红松林作为东北地区典型的森林植被类型, 对维持东北地区生态系统功能具有重要价值。其中丰富的物种和结构组成, 对研究生产力维持机制具有重要意义。本研究以吉林蛟河阔叶红松林大样地为研究对象, 试图分析物种多样性和群落结构变异对阔叶红松林生产力的影响, 为东北地区天然林经营指导提供科学依据。研究主要关注以下两个问题: 1)物种多样性和群落结构二者是否均与生产力存在相关关系? 其影响是否显著? 2)物种多样性和群落结构哪个对生产力的作用更大?
为解决上述两个问题, 首先构建了线性回归模型, 以生产力作为响应变量, 以物种多样性和群落结构变异分别作为解释变量, 单独分析物种多样性和群落结构对生产力的影响。进一步利用线性回归模型筛选出与生产力相关性最强的物种多样性和群落结构变异指标, 由此构建结构方程模型, 从而比较物种多样性和群落结构对生产力的作用路径, 验证二者对生产力的相对作用大小。
1 材料和方法
1.1 研究样地概况
研究样地建于2010年, 面积11.76 hm2 (420 m × 280 m), 地属吉林省蛟河市林业实验管理局辖区(43.85°-44.08° N, 127.58°-127.85° E), 位于长白山系张广才岭西南坡余脉, 属于受季风影响的温带大陆性气候区。研究区年平均气温3.8 ℃, 最热月7月平均气温21.7 ℃, 最冷月1月平均气温-18.6 ℃。年降水量约696 mm, 主要集中在夏季。样地中土壤类型单一, 为暗棕色森林土。样地地形平坦, 海拔变化范围在495.4-504.1 m之间, 最大高差不超过10 m。植被类型属长白山植物区系, 为典型的天然针阔混交林。研究样地中共调查到16 373棵木本植株, 隶属于19科25属36种。主要乔木树种包括红松(Pinus koraiensis)、紫椴(Tilia amurensis)、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、千金榆(Carpinus cordata)、色木槭(Acer pictum subsp. mono)等; 主要灌木树种包括髭脉槭(Acer barbinerve)、暴马丁香(Syringa reticulata subsp. amurensis)、毛榛(Corylus mandshurica)等。1.2 数据调查与计算
以20 m × 20 m连续小样方为取样单元, 于2010年对样地中所有胸径≥1 cm的木本植物的物种、胸径、树高、冠幅和相对位置进行记录并挂牌监测。2015年进行第一次复测调查。本研究以样地内胸径≥4 cm的活立木为研究对象, 共10 973株, 隶属于14科20属31种, 2010年平均胸径为15.31 cm, 2015年平均胸径为16.27 cm。计算两次调查间样方内胸高断面积年均增长量作为样方生产力。考虑到树木生长遵循幂次定律, 树木越大胸高断面积增长越多, 为改善数据正态性(D?nescu et al., 2016), 在R软件中对胸高断面积增长量作Box-Cox转换(Box & Cox, 1964)。
2012年用冠层分析仪(WinSCANOPY, Quebec, Canada)在每个小样方中心距离地面1.5 m处拍摄一张半球状照片, 用对应的WinSCANOPY和XLScanopy软件对照片进行处理, 得到林冠开阔度, 表示小样方的光照环境。同年在每个小样方中心20- 50 cm处采集500 g土壤样品, 于实验室内测定全氮、全磷、全钾、速效氮、速效磷、速效钾、有机碳含量和pH值共8个变量代表每个样方的土壤环境。考虑到群落生产力与林分密度密切相关, 计算胸高断面积密度。样地较为平坦, 地形差异对生产力的影响在此不作考虑。各样方数据统计如表1。
Table 1
表1
表1样地内基础变量信息统计表
Table 1The statistical information of basic variables in the sampling plot
| 变量 Variables | 范围 Range | 平均值 Mean | 标准偏差 Standard deviation |
|---|---|---|---|
| 胸高断面积年均增长量 Annual growth of basal area (cm2·a-1) | 76.53-543.60 | 272.22 | 83.62 |
| 林冠开阔度 Canopy openness | 1.12-3.99 | 1.87 | 0.38 |
| 土壤全氮 Soil total nitrogen (g·kg-1) | 0.65-1.60 | 0.95 | 0.16 |
| 土壤全磷 Soil total phosphorus (g·kg-1) | 0.33-1.09 | 0.79 | 0.12 |
| 土壤全钾 Soil total potassium (g·kg-1) | 20.40-20.90 | 20.70 | 0.01 |
| 土壤速效氮 Soil available nitrogen (mg·kg-1) | 29.93-68.81 | 53.65 | 8.48 |
| 土壤速效磷 Soil available phosphorus (mg·kg-1) | 0.09-22.01 | 11.94 | 5.59 |
| 土壤速效钾 Soil available potassium (mg·kg-1) | 206.80-236.20 | 226.30 | 5.74 |
| 土壤有机碳 Soil organic carbon (%) | 5.51-13.33 | 9.64 | 1.60 |
| 土壤pH值 Soil pH value | 4.38-5.32 | 4.88 | 0.25 |
| 胸高断面积密度 Basal area density (cm2·m-2) | 9.73-46.31 | 27.97 | 5.96 |
新窗口打开
1.3 物种多样性和群落结构变异计算
计算物种丰富度、香农指数和均匀度指数代表样地的物种多样性情况。群落结构变异的计算采用胸径香农指数和均匀度指数, 具体方法如下: 将初测胸径以1 cm为间隔划分等级, 统计每个样方内出现的胸径等级数以及每个等级内的个体数, 按照香农指数和均匀度指数的计算公式, 用胸径等级数替换物种数即可计算相应的群落结构变异, 其意义表示不同大小的树木间个体分配的均匀性(D?nescu et al., 2016)。另外, 计算胸径变异系数、胸径基尼系数表示群落结构变化。变异系数度量了胸径大小的离散程度, 其值越大离散程度越大。基尼系数表示胸径分布偏离绝对均匀状况的程度, 其值越大不同径级间个体分配越不均匀。各指标计算公式及结果见表2。Table 2
表2
表2物种多样性和群落结构变异计算公式及结果
Table 2The formulas and results of species diversity and community structure of woody plants in the studied forest
| 指数 Index | 计算公式 Formula | 平均值 Mean value | 范围 Range | |
|---|---|---|---|---|
| 物种多样性 Species diversity | 物种丰富度 Species richness | $S=Ns$ | 8.34 | 4-15 |
| 物种香农指数 Species Shannon index | $Hs=-\sum\limits_{i=1}^{Ns}{\frac{{{n}_{i}}}{N}\ \times \ ln\left( \frac{{{n}_{i}}}{N} \right)}$ | 1.81 | 1.18-2.55 | |
| 物种均匀度 Species evenness | $Es={Hs}/{ln\left( Ns \right)}\;$ | 0.87 | 0.65-0.97 | |
| 群落结构变异 Variation of community structure | 胸径香农指数 DBH Shannon index | $Hd=-\sum\limits_{j=1}^{Nd}{\frac{{{n}_{j}}}{N}\ \times \ ln\left( \frac{{{n}_{j}}}{N} \right)}$ | 2.77 | 2.24-3.11 |
| 胸径均匀度 DBH evenness | $Es={Hd}/{\text{l}n\left( Nd \right)}\;$ | 0.94 | 0.70-1.11 | |
| 胸径变异系数 Coefficient of DBH variation | $VarD\,=\,100%\,\ \times \ \frac{\sqrt{\frac{1}{N}{{\left( DB{{H}_{k}}-\mu \right)}^{2}}}}{\mu }$ | 77.36 | 44.14-112.70 | |
| 胸径基尼系数 DBH Gini index | $GiniD=\frac{\sum\limits_{k=2}^{N}{\left( 2\times \ k-N-\ 1 \right)\,\ \times \ b{{a}_{k}}}}{\sum\limits_{k=2}^{N}{\left( N-\ 1 \right)\,\ \times \ b{{a}_{k}}}}$ | 0.39 | 0.24-0.51 |
新窗口打开
1.4 回归模型构建
进行回归分析前, 用Z-score方法对解释变量进行标准化处理。利用转换后的响应变量和解释变量构建线性回归模型, 步骤如下:1.4.1 多重共线性检验
多重共线性会影响解释变量对响应变量的解释和预测能力。利用方差膨胀因子(VIF)检验变量间的多重共线性, 根据以往经验, VIF < 10时认为变量间没有多重共线性(Fox & Monette, 1992; Fox, 2008)。验证结果显示, 本研究解释变量VIF均小于10, 没有多重共线性。
1.4.2 模型构建
构建回归模型基本形式如下:
$ABAI={{b}_{0}}+{{b}_{1}}\times ENV+{{b}_{2}}\times BAD+{{b}_{3}}\times Diversity+\varepsilon $
式中ABAI为样方生产力, ENV为9个环境变量, 包括林冠开阔度, 土壤全氮、全磷、全钾、速效氮、速效磷、速效钾、有机碳、pH值, BAD为胸高断面积密度, Diversity为物种多样性或群落结构指数, b0表示截距项, b1、b2、b3为相应解释变量系数, ?表示随机误差。基于7个物种多样性和群落结构变异指数分别构建模型, 计算其与生产力的相关性。
1.4.3 模型评价
对于包含不同指数的7个模型, 均利用逐步回归法确定显著变量作为解释变量。计算调整决定系数(adjusted R2)和AIC信息量准则(Akaike information criterion)判断模型优度, 观察和比较物种多样性和群落结构与生产力的相关性。同时, 筛选与生产力最为相关的物种多样性和群落结构变异指数, 用来构建结构方程模型。
1.5 结构方程模型构建
为比较物种多样性和群落结构对生产力的相对作用大小, 研究构建物种多样性和群落结构变异同时影响生产力的结构方程模型。根据线性回归的结果, 结构方程模型中不仅考虑了物种多样性和群落结构变异的作用, 而且考虑了环境变量和胸高断面积密度对生产力的作用路径, 以及其对物种多样性和群落结构变异的影响。由于研究中环境变量较多, 通过比较模型整体优度保留最为相关的环境因子。利用模型中各路径标准化系数, 定量表示不同因素对生产力的相对作用大小, 从而比较物种多样性和群落结构与生产力的关系。2 结果
通过构建物种多样性和群落结构与生产力之间的多元回归模型(包含了环境因子和BAD作为解释变量), 比较了不同指数对生产力的作用。结果发现, 物种多样性和群落结构均对生产力有显著影响(表3)。7个多样性指标中, 物种均匀度、胸径香农指数和胸径基尼系数在模型中与生产力相关性显著(表3), 而物种丰富度、物种香农指数、胸径均匀度和胸径变异系数对生产力没有显著解释力(模型未列出)。Table 3
表3
表3与生产力显著相关的多样性线性回归结果
Table 3The correlation coefficient between the examined variables and forest productivity using different models
| 解释变量 Explanatory variables | 估计值 Estimate | ||
|---|---|---|---|
| 物种多样性模型 Species diversity model | 群落结构变异模型 Community structural variation model | ||
| A | B | ||
| 物种均匀度 Species evenness | -0.211* | - | - |
| 胸径香农指数 DBH Shannon index | - | 0.400*** | - |
| 胸径基尼系数 DBH Gini index | - | - | -0.337*** |
| 胸高断面积密度 Basal area density | 0.914*** | 0.756*** | 0.964*** |
| 林冠开阔度 Canopy openness | 0.289*** | 0.275** | 0.298*** |
| 有机碳 Soil organic carbon | 0.517*** | 0.221* | 0.293*** |
| 速效钾 Available potassium | 0.991*** | 0.193* | 0.241** |
| 速效氮 Available nitrogen | 0.413*** | - | - |
| pH | -0.520*** | - | - |
| 速效磷 Available phosphorus | - | 0.413*** | - |
| 全磷 Total phosphorus | - | -0.244* | - |
| 调整决定系数 Adjusted R2 | 34.67% | 35.95% | 31.93% |
| AIC值 AIC value | 1 080.385 | 1 074.56 | 1 090.52 |
新窗口打开
物种均匀度与生产力具有显著的负相关关系 (r = -0.211, p = 0.028), 即物种间个体分配越不均匀生产力越高。胸径香农指数与生产力具有显著的正相关关系(r = 0.400, p = 0.001), 胸径基尼系数与生产力具有显著负相关关系(r = -0.337, p < 0.001), 均表明个体在不同径级间分配越均匀时生产力水平 越高。
物种多样性的3个指标中, 只有均匀度指数与生产力具有显著的相关性, 因此选用均匀度指数代表物种多样性。群落结构变异指标中, 胸径基尼系数和香农指数与生产力具有显著相关性。比较发现, 此二者回归模型的调整决定系数香农指数(adjustedR2 = 35.95%)高于基尼系数(adjusted R2 = 31.93%), 而AIC值香农指数(AIC = 1 074.56)显著低于基尼系数(AIC = 1 090.52), 因此选用香农指数代表群落结构变异。
以均匀度指数代表物种多样性, 以胸径香农指数代表群落结构变异, 构建结构方程模型。变量较多时, 结构方程模型不易收敛, 故逐个加入环境变量构建结构方程模型, 最优结果(图1)如下:
群落结构变异对生产力有显著影响, 路径系数(反映作用大小)为0.329, 而物种多样性对生产力的影响不显著(路径系数为-0.158)。同时, 生产力还受到环境(主要包括速效磷、速效钾、有机碳以及光照)和林分密度的显著影响, 作用大小分别为0.694和0.773。研究结果表明, 在同时考虑物种多样性和群落结构变异对生产力的影响时, 群落结构变异的作用要大于物种多样性。
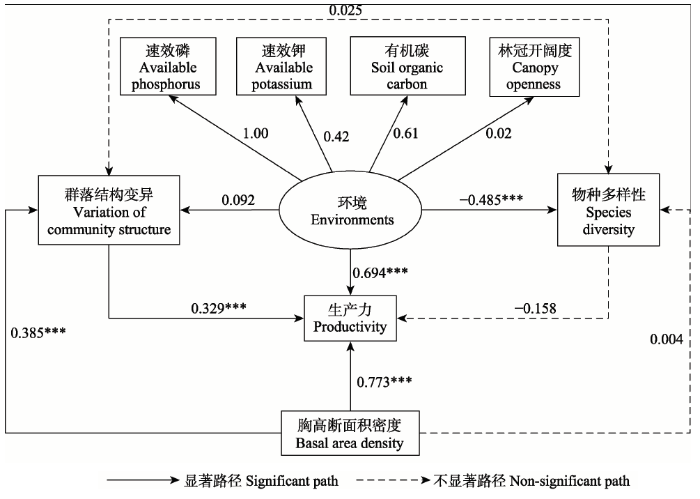 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1物种多样性和群落结构对生产力的作用路径图。图中实线表示作用路径显著, 虚线表示作用路径不显著。***, p < 0.001。
-->Fig. 1The impact paths of species diversity and community structure on productivity. Solid line indicates significant path, while dashed line indicates insignificant path. ***, p < 0.001.
-->
3 讨论
研究首先通过构建物种多样性及群落结构变异与生产力之间的回归模型, 分析了7个不同指标对生产力的影响, 并筛选了与生产力密切相关的物种多样性和群落结构变异指标, 进而通过结构方程模型, 验证了物种多样性和群落结构对生产力的相对作用大小。结果表明: 1)分别考虑物种多样性和群落结构作用时, 二者均对生产力具有显著影响; 2)同时考虑物种多样性和群落结构的影响时, 群落结构对生产力的作用更强。与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强。研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(Fahey et al., 2015)。在最近的研究中, 复杂冠层结构促进光吸收利用的重要性越发凸显(Parker et al., 2004; Christopherm et al., 2010; Hardiman et al., 2013)。具体而言, 群落结构复杂时, 在相同的叶面积条件下, 植物可以通过冠层间隙吸收更多的光能, 相比于结构单一的群落具有更高的光利用效率。在总叶面积和生产力关系达到一个相对稳定的状态后, 生产力会随冠层间隙的增多而增加(Hardiman et al., 2011, 2013)。同时, 结构复杂的群落更容易产生异质性光环境, 使得耐阴性物种存活率较高且枝叶更浓密(Valladares & Niinemets, 2008), 促进了下林层对生产力的贡献(Parker et al., 2002)。对于我们的研究对象而言, 针阔混交林具有更复杂的冠层结构, 红松圆锥形的树冠周围易形成 “光通道” (Pretzsch, 2014), 在以红松为优势种的群落中“光通道”广布, 相邻个体由此获得更多的光资源而生长较快, 促进对生产力的贡献。总之, 不同大小的个体具有特定的环境需求, 维持和增加群落结构变异有助于满足不同个体的特殊需求, 相反, 结构单一的群落中个体大小相近, 竞争相对激烈, 降低了生态位互补作用, 无法有效地促进生产力。
在其他群落结构与生产力关系的研究中, Liang等(2007)发现群落结构(胸径香农指数)与个体生长以及群落更新呈负相关关系, 而死亡率与群落结构呈正相关关系, 但此项研究针对的是10年的生长变化, 因此10年内的短期更新没有被监测到或死亡率被高估。Kuehne等(2015)通过不同强度的采伐使群落具有不同梯度的结构异质性(11个胸径结构变异指标), 发现群落结构对生产力的影响十分微弱。此外, Ryan等(2010)也发现结构异质性高(胸径分布不均匀)的群落相比结构均匀的群落生产力平均降低了约14%。造成以上结果与本研究不一致的原因, 有可能是方法上的差异, 包括多样性计算方法(比如选用不同的多样性指数结果不同)或者是样地监测方法(比如样地监测周期不同)(D?nescu et al., 2016), 也有可能是森林系统本身的差异造成: 一方面, 群落结构与发展阶段有关, 随着群落年龄增加, 群落结构更复杂, 在光资源利用上的互补更强, 对生产力的促进作用更强, 遵循生态位互补效应随群落发展阶段增强的规律(Reich et al., 2012; Zhang et al., 2012; Zhang & Chen, 2015); 另一方面, 群落结构与生产力的关系可能还与物种组成有关, 当群落中绝大多数都是不耐阴物种时, 透过冠层进入林下的光能并不能被当作生长所需的资源而吸收, 只有当群落中具有一定数量的耐阴性物种时, 由冠层结构差异造成的光异质环境才能被充分有效利用从而促进生产力(Parker et al., 2002; Valladares & Niinemets, 2008)。
关于物种多样性与生产力的关系, 虽然本研究得到了与大多数观点(即物种多样性促进生产力) (贺金生等, 2003; Forrester & Bauhus, 2016)不一致的结果, 但是长期以来关于物种多样性和生产力关系的激烈争论也证明物种多样性不能完美统一地解释现实世界中所有的多样性-生产力关系。本研究认为, 植物的生长受到所处环境及群落格局的影响, 而群落结构反映了群落在水平结构和垂直结构上的变化, 可以在一定程度上表示群落个体资源利用效率的差异, 因而更准确地反映了多物种多径级天然林中生态位互补效应对生产力的作用。
4 结论
在吉林蛟河阔叶红松林中, 物种多样性与生产力显著负相关, 群落结构与生产力显著正相关, 且对生产力的作用比物种多样性更强, 这主要是由于增加了冠层结构的复杂性影响群落对光能的捕获和利用从而影响了生产力。因此, 在森林经营管理中, 增加森林群落结构变异对有效增加生产力具有重要 意义。致谢 感谢北京林业大学郝珉辉博士和徐卫博士在数据处理分析过程中给予的帮助, 感谢李晓宇博士在论文修改过程中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . Plant structural diversity is usually considered as beneficial for ecosystem functioning. For instance, numerous studies have reported positive species diversity-productivity relationships in plant communities. However, other aspects of structural diversity such as individual size inequality have been far less investigated. In forests, tree size inequality impacts directly tree growth and asymmetric competition, but consequences on forest productivity are still indeterminate. In addition, the effect of tree size inequality on productivity is likely to vary with species shade-tolerance, a key ecological characteristic controlling asymmetric competition and light resource acquisition. Using plot data from the French National Geographic Agency, we studied the response of stand productivity to size inequality for ten forest species differing in shade tolerance. We fitted a basal area stand production model that included abiotic factors, stand density, stand development stage and a tree size inequality index. Then, using a forest dynamics model we explored whether mechanisms of light interception and light use efficiency could explain the tree size inequality effect observed for three of the ten species studied. Size inequality negatively affected basal area increment for seven out of the ten species investigated. However, this effect was not related to the shade tolerance of these species. According to the model simulations, the negative tree size inequality effect could result both from reduced total stand light interception and reduced light use efficiency. Our results demonstrate that negative relationships between size inequality and productivity may be the rule in tree populations. The lack of effect of shade tolerance indicates compensatory mechanisms between effect on light availability and response to light availability. Such a pattern deserves further investigations for mixed forests where complementarity effects between species are involved. When studying the effect of structural diversity on ecosystem productivity, tree size inequality is a major facet that should be taken into account. |
| [3] | . |
| [4] | . The focus of most ideas on diversity maintenance is species coexistence, which may be stable or unstable. Stable coexistence can be quantified by the long-term rates at which community members recover from low density. Quantification shows that coexistence mechanisms function in two major ways: They may be (a) equalizing because they tend to minimize average fitness differences between species, or (b) stabilizing because they tend to increase negative intraspecific interactions relative to negative interspecific interactions. Stabilizing mechanisms are essential for species coexistence and include traditional mechanisms such as resource partitioning and frequency-dependent predation, as well as mechanisms that depend on fluctuations in population densities and environmental factors in space and time. Equalizing mechanisms contribute to stable coexistence because they reduce large average fitness inequalities which might negate the effects of stabilizing mechanisms. Models of unstable coexitence, in which species diversity slowly decays over time, have focused almost exclusively on equalizing mechanisms. These models would be more robust if they also included stabilizing mechanisms, which arise in many and varied ways but need not be adequate for full stability of a system. Models of unstable coexistence invite a broader view of diversity maintenance incorporating species turnover. |
| [5] | . The mixed deciduous forests of the upper Midwest, USA are approaching an ecological threshold in which early successional canopy trees are reaching maturity and beginning to senesce, giving way to a more diverse canopy of middle and late successional species. The net primary production (NPP) of these forests is generally considered past peak and in decline, but recent studies show a striking resilience in the NPP trajectories of some middle and late successional forests; yet, the mechanisms controlling such temporal changes in NPP are largely unknown. At the University of Michigan Biological Station in northern Michigan, we used a 鈮9-year continuous record of wood net primary production (NPP), leaf area index (LAI), canopy composition, and stem mortality in 30 forested plots to identify the constraints on wood NPP as a mixed forest transitions from early to middle succession. Although wood NPP decreased over time in most stands, the rate of decline was attenuated when the canopy comprised a more diverse assemblage of early and middle/late successional species. The mechanism for sustained NPP in stands with more species diverse canopies was the proliferation of LAI by intact later successional tree species, even as stem mortality rates of early successional trees increased. We conclude that projections of carbon sequestration for the aging mixed forests of the upper Midwest should account for species composition shifts that affect the resilience wood NPP. |
| [6] | . In the past, explanations for high species diversity have been sought at the species level. Theory shows that coexistence requires substantial differences between species, but species-level data rarely provide evidence for such differences. Using data from forests in the southeastern United States, I show here that variation evident at the individual level provides for coexistence of large numbers of competitors. Variation among individuals within populations allows species to differ in their distributions of responses to the environment, despite the fact that the populations to which they belong do not differ, on average. Results are consistent with theory predicting that coexistence depends on competition being stronger within than between species, shown here by analysis of individual-level responses to environmental fluctuation. |
| [7] | Summary Top of page Summary Introduction Methods Results Discussion Acknowledgements References Supporting Information 1 A central concept in forest ecology is that differences in the growth rates and shade tolerances of tree species determine patterns of secondary succession. The most shade-tolerant tree species are the competitive dominants in late-successional forests, while species with fast growth rates persist through rapid establishment after disturbance. There is ample support for niche differentiation along the shade-tolerance axis, at least for temperate forests, but less thought has been given to the range of shade tolerances and growth rates encountered within a community and to how it might vary along environmental gradients. 2 We hypothesized that a wider range of growth rates and shade tolerances are found on nutrient-rich soils, because such soils not only support fast-growing species with high metabolic rates, but also species capable of tolerating the very deep shade cast by forest canopies growing where nutrients are plentiful. We test our hypothesis by quantifying light transmission through two neighbouring forests in southern New Zealand, one on phosphorus-rich alluvial soil and one on phosphorus-depleted marine-terrace soil, and comparing the growth rates of saplings on these contrasting sites. 3 Less light was transmitted to the forest floor on alluvial sites than on marine terraces (2.5% vs. 7.5% daylight, on average) and neighbourhood analyses within mapped stands indicated that large-leaved subcanopy species were responsible for intercepting that extra light. Sapling growth was strongly inhibited by shade in the understorey of the alluvial forests, but was less inhibited under the terrace forests. 4 Fast-growing subcanopy species were common on the alluvial sites and these species had characteristically soft leaves and high foliar-nutrient concentrations. Slow-growing shade-tolerant species were also abundant on these sites. Therefore, the interspecific variance in growth rates was greater on nutrient-rich sites, supporting our hypothesis of a greater range of shade tolerance niches on better soils. Of the five species found on both forest sites, all five had greater high-light growth rate on the alluvial sites. 5 Synthesis : A wider range of growth rates was observed in the nutrient-rich forests. This wider range may translate into a greater number of shade tolerance niches and thereby provides an explanation for the greater numbers of species commonly found on nutrient-rich soils when compared with neighbouring nutrient-poor sites. |
| [8] | . Abstract Forest diversity-productivity relationships have been intensively investigated in recent decades. However, few studies have considered the interplay between species and structural diversity in driving productivity. We analyzed these factors using data from 52 permanent plots in southwestern Germany with more than 53,000 repeated tree measurements. We used basal area increment as a proxy for productivity and hypothesized that: (1) structural diversity would increase tree and stand productivity, (2) diversity-productivity relationships would be weaker for species diversity than for structural diversity, and (3) species diversity would also indirectly impact stand productivity via changes in size structure. We measured diversity using distance-independent indices. We fitted separate linear mixed-effects models for fir, spruce and beech at the tree level, whereas at the stand level we pooled all available data. We tested our third hypothesis using structural equation modeling. Structural and species diversity acted as direct and independent drivers of stand productivity, with structural diversity being a slightly better predictor. Structural diversity, but not species diversity, had a significant, albeit asymmetric, effect on tree productivity. The functioning of structurally diverse, mixed forests is influenced by both structural and species diversity. These sources of trait diversity contribute to increased vertical stratification and crown plasticity, which in turn diminish competitive interferences and lead to more densely packed canopies per unit area. Our research highlights the positive effects of species diversity and structural diversity on forest productivity and ecosystem dynamics. |
| [9] | . Abstract The regrowing forests of eastern North America have been an important global C sink over the past 100+ years, but many are now transitioning into late succession. The consequences of this transition are unclear due to uncertainty around the C dynamics of old-growth forests. Canopy structural complexity (CSC) has been shown to be an important source of variability in C dynamics in younger forests (e.g., in productivity and resilience to disturbance), but its role in late-successional forests has not been widely addressed. We investigated patterns of CSC in two old-growth forest landscapes in the Upper Peninsula of Michigan, USA, to assess factors associated with CSC and its influence on productivity and disturbance resilience (to moderate-severity windstorm). CSC was quantified using a portable below-canopy LiDAR (PCL) system in 65 plots that also had long-term (50 70+ years) inventory data, which were used to quantify aboveground net primary productivity (ANPP), disturbance history, and stand characteristics. We found high and variable CSC relative to younger forests across a suite of PCL-derived metrics. Variation in CSC was driven by species composition and size structure, rather than disturbance history or site characteristics. Recent moderate severity wind disturbance decreased plot-scale CSC, but increased stand-scale variation in CSC. The strong positive correlation between CSC and productivity illustrated in younger forests was not present in undisturbed portions of these late-successional ecosystems. Moderate severity disturbance appeared to reestablish the positive link between CSC and productivity, but this relationship was scale and severity dependent. A positive CSC roductivity relationship was evident at the plot scale with low-severity, dispersed disturbance, but only at a patch scale in more severely disturbed areas. CSC does not appear to strongly correlate with variation in productivity in undisturbed old-growth forests, but may play a very important (and scale/severity-dependent) role in their response to disturbance. Understanding potential drivers and consequences of CSC in late-successional forests will inform management focused on promoting complexity and old-growth conditions, and illustrate potential impacts of such treatments on regional C dynamics. |
| [10] | . Through complementarity interactions, mixed-species forests can be more productive than monocultures, and this effect can increase with tree-species richness. However, this is not always the case. This review examines the processes and stand structural attributes that can influence diversity-productivity relationships (DPRs); how they influence resource availability, resource uptake, and resource-use efficiency; and also describes some important differences between tree-diversity versus grassland-diversity experiments. The size of the complementarity effects caused by these processes and stand structures varies along spatial and temporal gradients in resource availability and climate. These spatial and temporal dynamics have now been examined in many studies, and the general patterns are summarized using a simple framework; complementarity is predicted to increase as the availability of resource declines (or climatic condition X becomes harsher) if the species interactions improve the availability, uptake, or use efficiency of resource X (or interactions improve climatic condition X). Importantly, this framework differs from the stress-gradient hypothesis to account for a wider range of inter-specific plant interactions (not only facilitation) by considering contrasting methods used to quantify species interactions while accounting for stand structure. In addition, complementarity (as opposed to facilitation) for a given species combination can increase as growing conditions improve in forests, contrary to predictions of the stress-gradient hypothesis with regards to facilitation. This review indicates that while the effects of tree-species diversity on growth and other forest functions are now receiving a lot of attention, far less is known about the effects of structural diversity on growth or forest functioning. Direct measurements of the processes, as opposed to focusing mainly on growth responses, could greatly contribute to our understanding of structural diversity effects. |
| [11] | , |
| [12] | . |
| [13] | . The even-aged northern hardwood forests of the Upper Great Lakes Region are undergoing an ecological transition during which structural and biotic complexity is increasing. Early-successional aspen (Populus spp.) and birch (Betula papyrifera) are senescing at an accelerating rate and are being replaced by middle-successional species including northern red oak (Quercus rubra), red maple (Acer rubrum), and white pine (Pinus strobus). Canopy structural complexity may increase due to forest age, canopy disturbances, and changing species diversity. More structurally complex canopies may enhance carbon (C) sequestration in old forests. We hypothesize that these biotic and structural alterations will result in increased structural complexity of the maturing canopy with implications for forest C uptake. At the University of Michigan Biological Station (UMBS), we combined a decade of observations of net primary productivity (NPP), leaf area index (LAI), site index, canopy tree-species diversity, and stand age with canopy structure measurements made with portable canopy lidar (PCL) in 30 forested plots. We then evaluated the relative impact of stand characteristics on productivity through succession using data collected over a nine-year period. We found that effects of canopy structural complexity on wood NPP (NPPw) were similar in magnitude to the effects of total leaf area and site quality. Furthermore, our results suggest that the effect of stand age on NPPw is mediated primarily through its effect on canopy structural complexity. Stand-level diversity of canopy-tree species was not significantly related to either canopy structure or NPPw. We conclude that increasing canopy structural complexity provides a mechanism for the potential maintenance of productivity in aging forests. |
| [14] | . Recent observations demonstrate that, against expectations, some forests maintain high carbon (C) storage rates for centuries, though the underlying mechanisms remain poorly understood. To test the hypothesis that age-related increases in canopy structural complexity improve resource-use efficiency and to evaluate canopy structural influences on forest C storage over successional timescales, we measured the fraction of photosynthetically active radiation absorbed by the canopy (fAPAR), foliar N mass , and aboveground net primary production (ANPP) in a chronosequence of 39 passively managed stands spanning >160 years of forest development in Northern Lower Michigan, USA. We used ground-based portable canopy LiDAR to quantify canopy structural complexity as rugosity, an integrated metric of 3D heterogeneity in canopy leaf area arrangement. Here, we describe a mechanism capable of maintaining high rates of ANPP over nearly two centuries of forest development. Results support our hypothesis that increasing canopy complexity over the course of forest development mediates greater resource-use efficiency in these forests. Forest stands with more structurally complex canopies had higher light and nitrogen use efficiencies (LUE & NUE) and higher ANPP. LAI was stable across stands older than 50 years, while canopy complexity (rugosity) increased with age through >160 years of stand development. Rugosity had a bigger influence on ANPP across all stands than did LAI, demonstrating the greater long-term influence of leaf area arrangement, rather than quantity within the canopy on forest C storage. We conclude that canopy structural complexity may facilitate greater resource use efficiency (RUE) in aging forests and so increase ANPP compared to structurally simpler canopies in young forests, thus maintaining significant C storage potential in aging forests. We suggest that forest managers should incorporate canopy structural complexity as a robust proxy of stand C storage potential in forests differing widely in age and disturbance history. |
| [15] | 人们担心生物多样性的空前丧失会危及到生态系统的服务功能,因此有关生物多样性-生态系统生产力这一古老命题的讨论成为当今生态学的热点议题之一.20世纪90年代以David Tilman和Shahid Naeem为代表的生态学家利用大规模的受控实验,对物种多样性与生态系统功能的诸多方面进行了研究,结果表明物种多样性导致生态系统生产力的增加.这一结果遭到以David Wardle, Michael Huston和Phillip Grime为代表的生态学家的质疑,认为这样的结果可能是由抽样效应引起的,而不是生物多样性本身的作用.争论的本身在很大程度上是由于受控实验结果和野外观测不一致造成的.结合作者的实验以及文献综述,该文分析了生物多样性实验群落和自然发育的群落之间的异同,认为野外观测和受控实验结果不一致的原因,可能源于群落密度、均匀度及土壤营养状况的不同,以及由此改变了生物多样性-生态系统生产力之间的关系.这些因子在自然状况下是多变的,而在生物多样性实验条件下则相对一致.最后,我们提出生物多样性-生态系统生产力之间的关系还可能受到土壤元素可利用性的负反馈作用的影响. |
| [16] | . Humans are altering the composition of biological communities through a variety of activities that increase rates of species invasions and species extinctions, at all scales, from local to global. These changes in components of the Earth's biodiversity cause concern for ethical and aesthetic reasons, but they also have a strong potential to alter ecosystem properties and the goods and services they provide to humanity. Ecological experiments, observations, and theoretical developments show that ecosystem properties depend greatly on biodiversity in terms of the functional characteristics of organisms present in the ecosystem and the distribution and abundance of those organisms over space and time. Species effects act in concert with the effects of climate, resource availability, and disturbance regimes in influencing ecosystem properties. Human activities can modify all of the above factors; here we focus on modification of these biotic controls.The scientific community has come to a broad consensus on many aspects of the relationship between biodiversity and ecosystem functioning, including many points relevant to management of ecosystems. Further progress will require integration of knowledge about biotic and abiotic controls on ecosystem properties, how ecological communities are structured, and the forces driving species extinctions and invasions. To strengthen links to policy and management, we also need to integrate our ecological knowledge with understanding of the social and economic constraints of potential management practices. Understanding this complexity, while taking strong steps to minimize current losses of species, is necessary for responsible management of Earth's ecosystems and the diverse biota they contain.Based on our review of the scientific literature, we are certain of the following conclusions:1)Species' functional characteristics strongly influence ecosystem properties. Functional characteristics operate in a variety of contexts, includin |
| [17] | . |
| [18] | . Relationships between stand growth and structural diversity were examined in spruce-dominated forests in New Brunswick, Canada. Net growth, survivor growth, mortality, and recruitment represented stand growth, and tree species, size, and height diversity indices were used to describe structural diversity. Mixed-effects second-order polynomial regressions were employed for statistical analysis. ... |
| [19] | . The objective of this study was to determine the relationship, or lack thereof, between growth and diversity of tree species and size in conifer stands of western North America. Growth was measured by net basal area growth and its components: survivor growth, recruitment, and mortality. The analysis used inventory data from permanent plots in the Douglas-fir/western hemlock forest type in Oregon and Washington, and in the mixed-conifer forest type in California. The methods consisted of generalized least square regression with spatial autocorrelation, controlling for the effect of other stand characteristics. Other things being equal, in the two forest types under study there was a strong positive relationship between net basal area growth and tree-species diversity. This effect was associated with higher recruitment in stands of higher tree-species diversity. Neither mortality nor growth of survivors was related to tree-species diversity. The relationship between growth and tree-size diversity was less clear. For Douglas-fir/western hemlock, net basal area growth was negatively correlated with tree-size diversity, essentially because recruitment was lower on plots of high tree-size diversity. For mixed conifers, net basal area growth tended also to be lower in plots of high tree-size diversity, but this was mostly because mortality was higher in plots of higher tree-size diversity. |
| [20] | . Data from ~1500 ponderosa pine (Pinus ponderosa C. Lawson) stands in the western United States were used to examine the potential influence of compositional and structural diversity on forest productivity. Relative density, height and site quality were combined in a conceptually sound expression of the relationship between growth and growing stock for ponderosa pine-dominated stands of relative... |
| [21] | . |
| [22] | . |
| [23] | . Understanding the link between biodiversity and ecosystem functioning (BEF) is pivotal in the context of global biodiversity loss. Yet, long-term effects have been explored only weakly, especially for forests, and no clear evidence has been found regarding the underlying mechanisms. We explore the long-term relationship between diversity and productivity using a forest succession model. Extensive simulations show that tree species richness promotes productivity in European temperate forests across a large climatic gradient, mostly through strong complementarity between species. We show that this biodiversity effect emerges because increasing species richness promotes higher diversity in shade tolerance and growth ability, which results in forests responding faster to small-scale mortality events. Our study generalises results from short-term experiments in grasslands to forest ecosystems and demonstrates that competition for light alone induces a positive effect of biodiversity on productivity, thus providing a new angle for explaining BEF relationships. |
| [24] | |
| [25] | . Abstract We measured vertical and horizontal variation in canopy transmittance of photosynthetically active radiation in five Pseudotsuga menziesii (Mirb.) Franco-Tsuga heterophylla (Raf.) Sarg. (Douglas-fir-western hemlock) stands in the central Cascades of southern Washington to determine how stand structure and age affect the forest light environment. The shape of the mean transmittance profile was related to stand height, but height of mean maximum transmittance was progressively lower than maximum tree height in older stands. The vertical rate of attenuation declined with stand age in both the overstory and understory. A classification of vertical light zones based on the mean and variance of transmittance showed a progressive widening of the bright (low variance and high mean) and transition (high variance and rapid vertical change) zones in older stands, whereas the dim zone (low variance and mean) narrowed. The zone of maximum canopy surface area in height profiles, estimated by inversion of transmittance profiles, changed from relatively high in the canopy in most young stands ("top-heavy") to lower in the canopy in older stands ("bottom-heavy"). In the understory, all stands had similar mean transmittances, but the spatial scale of variation increased with stand age and increasing crown size. The angular distribution of openness was similar in all stands, though the older stands were less open at all angles than the younger stands. Understory openness was generally unrelated to transmittance in the canopy above. Whole-canopy leaf area indices, estimated using three methods of inverting light measurements, showed little correspondence across methods. The observed patterns in light environment are consistent with structural changes occurring during stand development, particularly the diversification of crowns, the creation of openings of various sizes and the elaboration of the outer canopy surface. The ensemble of measurements has potential use in distinguishing forests of differing ages that have similar stature. |
| [26] | . We describe the three-dimensional structure of an old-growth Douglas-fir/western hemlock forest in the central Cascades of southern Washington, USA. We concentrate on the vertical distribution of foliage, crowns, external surface area, wood biomass, and several components of canopy volume. In addition, we estimate the spatial variation of some aspects of structure, including the topography of the outer surface, and of microclimate, including the within-canopy transmittance of photosynthetically active radiation (PAR). The crowns of large stems, especially of Douglas-fir, dominate the structure and many aspects of spatial variation. The mean vertical profile of canopy surfaces, estimated by five methods, generally showed a single maximum in the lower to middle third of the canopy, although the height of that maximum varied by method. The stand leaf area index was around 9 m 2 m 2 , but also varied according to method (from 6.3 to 12.3). Because of the deep narrow crowns and numerous gaps, the outer canopy surface is extremely complex, with a surface area more than 12 times that of the ground below. The large volume included below the outer canopy surface is very porous, with spaces of several qualitatively distinct environments. Our measurements are consistent with emerging concepts about the structure of old-growth forests, where a high degree of complexity is generated by diverse structural features. These structural characteristics have implications for various ecosystem functions. The height and large volume of the stand indicate a large storage component for microclimatic variables. The high biomass influences the dynamics of those variables, retarding rates of change. The complexity of the canopy outer surface influences radiation balance, particularly in reducing short-wave reflectance. The bottom-heaviness of the foliage profile indicates much radiation absorption and gas exchange activity in the lower canopy. The high porosity contributes to flat gradients of most microclimate variables. Most stand respiration occurs within the canopy and is distributed over a broad vertical range. |
| [27] | . Mixed-species forest stands are well explored in their favourable ecological, economical, and socio-economical functions and services compared with pure stands, but still poorly understood in their structure and functioning. Canopy structure and tree morphology affect the environmental conditions within the stand, the tree growth, and by this most forest functions and services. Here, I review how canopy structure and crown morphology in mixed stands can differ from pure stands and how this depends on the selection of tree species and interactions between them. The focus is on the macrostructure of canopy and crowns derived from the trees’ positions, their convex crown hulls, and their space filling with branches. In mixed canopies the sum of the crown projection area, but not the ground coverage by crowns, mostly exceeds pure stands due to multiple crown overlaps. The interspecific differences in crown shape and allometric scaling cause a ‘selection effect’ when complementary species are combined. In interspecific environment furthermore ‘true mixing effects’ like intraspecific shifts in size, shape, and inner space filling of crowns may occur. The much denser and more plastic canopy space filling in mixed stands may increase light interception, stand density, productivity, and growth resilience to disturbances. I discuss the relevance of interspecific interactions for forest management, model building, and theory development and draw perspectives of further research into stand canopy and crown structure. |
| [28] | . Abstract Plant diversity generally promotes biomass production, but how the shape of the response curve changes with time remains unclear. This is a critical knowledge gap because the shape of this relationship indicates the extent to which loss of the first few species will influence biomass production. Using two long-term ( 13 years) biodiversity experiments, we show that the effects of diversity on biomass productivity increased and became less saturating over time. Our analyses suggest that effects of diversity-dependent ecosystem feedbacks and interspecific complementarity accumulate over time, causing high-diversity species combinations that appeared functionally redundant during early years to become more functionally unique through time. Consequently, simplification of diverse ecosystems will likely have greater negative impacts on ecosystem functioning than has been suggested by short-term experiments. |
| [29] | . Abstract A common pattern emerging from studies on the relationship between plant diversity and ecosystem functioning is that productivity increases with diversity. Most of these studies have been carried out in perennial grasslands, but many lasted only two growing seasons or reported data from a single year. Especially for perennial plant communities, however, the long-term effects of diversity are important. The question whether interactions between few species or among many species lead to increased productivity remained largely unanswered. So far, the main mechanism addressed is the increased input of nitrogen by nitrogen-fixing legumes. We report that other mechanisms can also generate strong increases of productivity with diversity. Results from 4 consecutive years of a plant diversity experiment without legumes show that a positive relationship between plant species richness and productivity emerged in the second year and strengthened with time. We show that increased nutrient use efficiency at high species richness is an important underlying mechanism. This mechanism had not been discussed in earlier studies. Furthermore, our results suggest that complementary nutrient uptake in space and time is important. Together, these mechanisms sustain consistently high productivity at high diversity. |
| [30] | . Wood production varies substantially with resource availability, and the variation in wood production can result from several mechanisms: increased photosynthesis, and changes in partitioning of photosynthesis to wood production, belowground flux, foliage production or respiration. An understanding of the mechanistic basis for patterns in wood production within a stand and across landscapes requires a complete annual carbon budget. We measured annual carbon flows to wood production, foliage production and total belowground carbon flux (the sum of root production, root respiration, and mycorrhizal production and respiration) from ages three to five years in clonal Eucalyptus plantations at four sites in Brazil to test if fertility, water availability and stand structure changed wood production and by what mechanism. We also quantified the patterns in light interception and the efficiency of light use to provide additional mechanistic insights into growth responses and to determine if light-use efficiency was related to changes in flux and partitioning. The routine level of forest fertilization at these four sites was high enough that further increases in nutrient supply did not increase wood growth. Irrigation increased wood net primary productivity (age three to five) from 1.45 to 1.84 kg m 612 year 611 of C (27%), because of increases in light interception (5%), photosynthetic efficiency (from 0.028 to 0.031 mol C/mol photons absorbed, 11%), gross primary productivity (from 3.62 to 4.28 m 612 year 611 of C, 18%), and partitioning to wood (from 0.397 to 0.430 of photosynthesis, 8%). These changes increased light-use efficiency by 20%. Annual flux belowground varied among sites from 0.43 to 1.0 m 612 year 611 of C but did not vary with water availability. Across the four sites for the irrigated and unirrigated treatments, light-use efficiency was positively correlated with gross primary productivity and partitioning to wood production. Increasing heterogeneity of stand structure (resulting from staggered timing of planting within plots) led to a 14% loss in wood biomass relative to uniform stand structure at age six. Light-use efficiency, gross primary productivity, and wood net primary productivity were lower, but not significantly so, in heterogeneous compared to uniform stands. |
| [31] | . |
| [32] | . Abstract Human-driven ecosystem simplification has highlighted questions about how the number of species in an ecosystem influences its functioning. Although biodiversity is now known to affect ecosystem productivity, its effects on stability are debated. Here we present a long-term experimental field test of the diversity-stability hypothesis. During a decade of data collection in an experiment that directly controlled the number of perennial prairie species, growing-season climate varied considerably, causing year-to-year variation in abundances of plant species and in ecosystem productivity. We found that greater numbers of plant species led to greater temporal stability of ecosystem annual aboveground plant production. In particular, the decadal temporal stability of the ecosystem, whether measured with intervals of two, five or ten years, was significantly greater at higher plant diversity and tended to increase as plots matured. Ecosystem stability was also positively dependent on root mass, which is a measure of perenniating biomass. Temporal stability of the ecosystem increased with diversity, despite a lower temporal stability of individual species, because of both portfolio (statistical averaging) and overyielding effects. However, we found no evidence of a covariance effect. Our results indicate that the reliable, efficient and sustainable supply of some foods (for example, livestock fodder), biofuels and ecosystem services can be enhanced by the use of biodiversity. |
| [33] | . THE functioning and sustainability of ecosystems may depend on their biological diversity 1–8 . Elton's 9 hypothesis that more diverse ecosystems are more stable has received much attention 1,3,6,7,10–14 , but Darwin's proposal 6,15 that more diverse plant communities are more productive and the related conjectures 4,5,16,17 that they have lower nutrient losses and more sustainable soils, are less well studied 4–6,8,17,18 . Here we use a well-replicated field experiment, in which species diversity was directly controlled, to show that ecosystem productivity in 147 grassland plots increased significantly with plant biodiversity. Moreover, the main limiting nutrient, soil mineral nitrogen, was utilized more completely when there was a greater diversity of species, leading to lower leaching loss of nitrogen from these ecosystems. Similarly, in nearby native grassland, plant productivity and soil nitrogen utilization increased with increasing plant species richness. This supports the diversity–productivity and diversity–sustainability hypotheses. Our results demonstrate that the loss of species threatens ecosystem functioning and sustainability. |
| [34] | . Light gradients are ubiquitous in nature, so all plants are exposed to some degree of shade during their lifetime. The minimum light required for survival, shade tolerance, is a crucial life-history trait that plays a major role in plant community dynamics. There is consensus on the suites of traits that influence shade tolerance, but debate over the relative importance of traits maximizing photosynthetic carbon gain in low light versus those minimizing losses. Shade tolerance is influenced by plant ontogeny and by numerous biotic and abiotic factors. Although phenotypic plasticity tends to be low in shade-tolerant species (e.g., scant elongation in low light), plasticity for certain traits, particularly for morphological features optimizing light capture, can be high. Understanding differential competitive potentials among co-occurring species mediated by shade tolerance is critical to predict ecosystem responses to global change drivers such as elevated CO sub(2), climate change and the spread of invasive species. |
| [35] | . Recent experiments on grassland ecosystems have shown that biodiversity can enhance ecosystem processes such as plant biomass production. Functional complementarity is generally regarded as the main class of mechanisms generating these effects of biodiversity on ecosystem functioning. Although intuitively appealing and supported by some data, the complementarity hypothesis has been little explored theoretically using mechanistic approaches. Here, we present a simple dynamical model for a light-limited terrestrial ecosystem to assess the effects of species diversity on light competition and total biomass in plant communities. Our model shows that competitive relaxation (reduction in average light competition intensity) due to differences in foliar architecture among species enhances total plant biomass in mixtures, but that competitive imbalance (generated by the variance of the average light competition intensity experienced by different species) can either reinforce the effect of competitive relaxation or counteract it and contribute to reducing total plant biomass. Thus, complementary resource use is not enough to increase total plant biomass in species-rich communities; competitive balance among species also plays an important role. We propose an operational measure of light-use complementarity using empirical field data on light absorption to test the presence of complementarity in natural plant communities. |
| [36] | . Summary Despite the mounting evidence for positive diversity–productivity relationships found in controlled experiments, diversity effects on productivity in natural systems remain hotly debated. Understanding the multivariate links between diversity and productivity in natural systems, in particular natural forests that host the majority of terrestrial biodiversity and provide essential services for humanity, remains a critical challenge for ecologists. We analysed data from 448 plots of varying tree species diversity, stand ages and local nutrient availability in Canada's boreal forest (52°30′–55°24′02N latitude and 102°36′–108°02W longitude). We used structural equation models to link multivariate relationships between above-ground biomass, tree species diversity, stand age and soil nutrient availability. Above-ground biomass increased with diversity indirectly via increasing tree size inequality, increased with stand age and was higher on sites of medium soil nutrient regime directly as well as indirectly via increased tree size inequality. Synthesis . Our results demonstrate positive diversity effects on above-ground biomass in natural forests of diverse forest ages and soil resource availability. Furthermore, we show that tree size inequality acts as a mechanism for the positive diversity effects on above-ground biomass and as a mechanism in regulating above-ground biomass and species diversity simultaneously via interactions among individuals in natural forests. |
| [37] | . 1. Although there is ample support for positive species richness–productivity relationships in planted grassland experiments, a recent 48-site study found no diversity–productivity relationship (DPR) in herbaceous communities. Thus, debate persists about diversity effects in natural versus planted systems. Additionally, current knowledge is weak regarding the influence of evenness on the DPRs, how DPRs are affected by the variation in life-history traits among constituent species in polycultures and how DPRs differ among biomes. The impacts of these factors on DPRs in forest ecosystems are even more poorly understood.2. We performed a meta-analysis of 54 studies to reconcile DPRs in forest ecosystems. We quantified the net diversity effect as log effect size [ln(ES)], the log ratio of the productivity in polycultures to the average of those in monocultures within the same type of mixture, site condition and stand age of each study. The first use of a boosted regression tree model in meta-analysis, a useful method to partition the effects of multiple predictors rather than relying on vote-counting of individual studies, unveiled the relative influences of individual predictors.3. Global average ln(ES) was 0.2128, indicating 23.7% higher productivity in polycultures than monocultures. The final model explained 21% of the variation in ln(ES). The predictors that substantially accounted for the explained variation included evenness (34%), heterogeneity of shade tolerance (29%), richness (13%) and stand age (15%). In contrast, heterogeneity of nitrogen fixation and growth habits, biome and stand origin (naturally established versus planted) contributed negligibly (each ≤664%). Log effect size strongly increased with evenness from 0.6 to 1 and with richness from 2 to 6. Furthermore, it was higher with heterogeneity of shade tolerance and generally increased with stand age.4. Synthesis. Our analysis is, to our knowledge, the first to demonstrate the critical role of species evenness, richness and the importance of contrasting traits in defining net diversity effects in forest polycultures. While testing the specific mechanisms is beyond the scope of our analysis, our results should motivate future studies to link richness, evenness, contrasting traits and life-history stage to the mechanisms that are expected to produce positive net biodiversity effects such as niche differentiation, facilitation and reduced Janzen–Connell effects. |
1
1997
... 多样性-生产力关系是森林可持续经营的重要论题(
Tree size inequality reduces forest productivity: An analysis combining inventory data for ten European species and a light competition model
1
2016
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
An analysis of transformations
1
1964
... 计算两次调查间样方内胸高断面积年均增长量作为样方生产力.考虑到树木生长遵循幂次定律, 树木越大胸高断面积增长越多, 为改善数据正态性(
Mechanisms of maintenance of species diversity
1
2000
... 多样性-生产力关系是森林可持续经营的重要论题(
Wood net primary production resilience in an unmanaged forest transitioning from early to middle succession
1
2010
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
Individuals and the variation needed for high species diversity in forest trees
2
2010
... 多样性-生产力关系是森林可持续经营的重要论题(
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
A greater range of shade-tolerance niches in nutrient-rich forests: An explanation for positive richness-productivity relationships?
1
2009
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany
6
2016
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
... 林中, 群落结构与生产力并没有显著的相关关系.甚至有研究直接指出群落结构对生产力的影响比物种多样性更强(
... 计算两次调查间样方内胸高断面积年均增长量作为样方生产力.考虑到树木生长遵循幂次定律, 树木越大胸高断面积增长越多, 为改善数据正态性(
... 计算物种丰富度、香农指数和均匀度指数代表样地的物种多样性情况.群落结构变异的计算采用胸径香农指数和均匀度指数, 具体方法如下: 将初测胸径以1 cm为间隔划分等级, 统计每个样方内出现的胸径等级数以及每个等级内的个体数, 按照香农指数和均匀度指数的计算公式, 用胸径等级数替换物种数即可计算相应的群落结构变异, 其意义表示不同大小的树木间个体分配的均匀性(
... 在其他群落结构与生产力关系的研究中,
Quantifying canopy complexity and effects on productivity and resilience in late-successional hemlock-hardwood forests
1
2015
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
A review of processes behind diversity—Productivity relationships in forests
2
2016
... 多样性-生产力关系是森林可持续经营的重要论题(
... 关于物种多样性与生产力的关系, 虽然本研究得到了与大多数观点(即物种多样性促进生产力) (
Applied Regression Analysis and Generalized Linear Models. 2nd edn. Sage Publications, Thousand Oaks
1
2008
... 多重共线性会影响解释变量对响应变量的解释和预测能力.利用方差膨胀因子(VIF)检验变量间的多重共线性, 根据以往经验, VIF < 10时认为变量间没有多重共线性(
Generalized collinearity diagnostics
1
1992
... 多重共线性会影响解释变量对响应变量的解释和预测能力.利用方差膨胀因子(VIF)检验变量间的多重共线性, 根据以往经验, VIF < 10时认为变量间没有多重共线性(
The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest
1
2011
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
Maintaining high rates of carbon storage in old forests: A mechanism linking canopy structure to forest function
2
2013
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
... ,
生物多样性与生态系统生产力: 为什么野外观测和受控实验结果不一致?
2
2003
... 多样性-生产力关系是森林可持续经营的重要论题(
... 关于物种多样性与生产力的关系, 虽然本研究得到了与大多数观点(即物种多样性促进生产力) (
Effects of biodiversity on ecosystem functioning: A consensus of current knowledge
1
2005
... 多样性-生产力关系是森林可持续经营的重要论题(
Effects of thinning-induced changes in structural heterogeneity on growth, ingrowth, and mortality in secondary coastal Douglas-fir forests
2015
Relationships between stand growth and structural diversity in spruce-dominated forests in New Brunswick, Canada
2
2008
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality
2
2007
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
... 在其他群落结构与生产力关系的研究中,
The influence of compositional and structural diversity on forest productivity
1
2010
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
Partitioning selection and complementarity in biodiversity experiments
2
2001
... 多样性-生产力关系是森林可持续经营的重要论题(
... ).对于这样的结果, 生态位互补效应是一个主要解释(
Tree and stand light use efficiencies over a full rotation of single- and mixed- species Eucalyptus grandis, and Acacia mangium plantations
1
2013
... 多样性-生产力关系是森林可持续经营的重要论题(
Tree species richness promotes productivity in temperate forests through strong complementarity between species
1
2011
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
1
2009
... 多样性-生产力关系是森林可持续经营的重要论题(
Canopy light transmittance in Douglas-fir-western hemlock stands
2
2002
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
... 在其他群落结构与生产力关系的研究中,
Three- dimensional structure of an old-growth Pseudotsuga- Tsuga, canopy and its implications for radiation balance, microclimate, and gas exchange
1
2004
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures
2
2014
... 多样性-生产力关系是森林可持续经营的重要论题(
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
Impacts of biodiversity loss escalate through time as redundancy fades
1
2012
... 在其他群落结构与生产力关系的研究中,
Diversity-productivity relationships: Initial effects, long-term patterns, and underlying mechanisms
1
2005
... 多样性-生产力关系是森林可持续经营的重要论题(
Factors controlling Eucalyptus productivity: How water availability and stand structure alter production and carbon allocation
1
2010
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
Plant diversity and ecosystem productivity: Theoretical considerations
1
1997
... 多样性-生产力关系是森林可持续经营的重要论题(
Biodiversity and ecosystem stability in a decade-long grassland experiment
1
2006
... 多样性-生产力关系是森林可持续经营的重要论题(
Productivity and sustainability influenced by biodiversity in grassland ecosystems
1
1996
... 多样性-生产力关系是森林可持续经营的重要论题(
Shade tolerance, a key plant feature of complex nature and consequences
2
2008
... 与以往主要讨论物种多样性作用的研究有所不同, 本研究重点关注了群落结构的影响, 发现群落结构对生产力的作用更强.研究认为, 群落结构变异主要是通过加强光能互补利用促进生产力提高, 其能够直接反映个体大小在水平结构上的变化, 同时间接反映在垂直结构(即树高)上的变化, 群落中个体处在不同径级、不同林层, 增加了冠层结构复杂性(
... 在其他群落结构与生产力关系的研究中,
Does complementary resource use enhance ecosystem functioning? A model of light competition in plant communities
1
2007
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
Individual size inequality links forest diversity and above-ground biomass
3
2015
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
... 在目前探讨群落结构与生产力关系的研究中, 大多研究表明二者之间呈正相关关系(
... 在其他群落结构与生产力关系的研究中,
Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis
3
2012
... 多样性-生产力关系是森林可持续经营的重要论题(
... 近年来, 群落结构对生产力的影响逐渐成为研究热点(
... 在其他群落结构与生产力关系的研究中,


{kind=link}
{kind=link}