
Effects of short-term nitrogen addition on fine root biomass, lifespan and morphology of Castanopsis platyacantha in a subtropical secondary evergreen broad-leaved forest
CHENGuan-Tao通讯作者:
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (450KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(Jackson et al., 1997; Poorter et al., 2011), 因此, 细根周转推动着陆地生态系统碳和养分循环。影响细根周转速率的主导因子是细根寿命(Majdi et al., 2005)。根系是植物和土壤交互的界面, 对土壤环境变化高度敏感, 根系不断调节自身特性以适应土壤环境变化, 进而支持植物正常生长(Eissenstat &Yanai, 1997; Eissenstat et al., 2000; Wurzburger & Wright, 2015)。这类调节涉及生物量、形态、生理过程等多方面, 且相互紧密联系, 进而影响包括根系周转在内的诸多生态系统过程(Bardgett et al., 2014)。
植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(Nacry et al., 2013)。大多数陆地生态系统的净初级生产力受到氮元素的限制(LeBauer & Treseder, 2008)。根据“成本-效益”理论, 植物根系总是以尽可能少的碳消耗来满足植物对氮素的需求(Eissenstat & Yanai, 1997)。因此, 土壤氮素可利用性对根系寿命和形态具有重要影响(Eissenstat et al., 2000), 这在氮沉降梯度研究和元素添加试验中得到了广泛证明(Ostonen et al., 2007; Li et al., 2015)。
近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(Galloway & Cowling, 2002; Galloway et al., 2004), 对森林生态系统物质循环造成了潜在的巨大影响(吕超群等, 2007)。大量研究表明, 日益增加的氮沉降会增加土壤氮素可利用性, 多数情况会下提高植被地上部分生产力(Bedison & Mcneil, 2009)。然而森林植物根系对土壤氮素增加的响应方向和程度却存在很大的不确定性。有研究表明氮添加增加了根系寿命, 减缓了细根周转速率(Jourdan et al., 2008; 于水强等, 2009; Mei et al., 2010), 但也有研究观察到与之相反的结果(Johnson et al., 2000; Wang et al. 2012)。现有的少量氮添加对森林植物根系影响的相关研究往往在人工林中开展(于水强等, 2009)。人工林林分简单, 为根系研究提供了很大便利(史建伟等, 2006)。然而作为全球森林主体的天然林(Chirici et al., 2013), 往往与人工林在土壤特性、林分结构、稳定性、物种多样性等方面存在极大的差异(Hou et al., 2010; 李丹等, 2011)。因此, 在天然林中开展相关研究对于更全面地了解氮沉降对森林生态系统的影响具有重要意义。
根系生长于地下, 不易接近和直接观察, 对于根系的认识很大程度上受限于研究手段。传统挖掘法、土钻法、内生长法研究根系, 往往费时费力, 对根系破坏严重, 且不能连续观察, 很难实现对根寿命的准确测定(李俊英等, 2007)。近年来, 微根管技术以其特有的原位性、非破坏性等特点在根系研究中得到快速发展。通过微根管技术能直接观察单个根段从出生到死亡的全过程(Chen & Brassard, 2013; Balogianni et al., 2016), 在细根生产、寿命、周转、细根生物量的估算等方面应用越来越广泛, 并得到普遍认可(Addo-Danso et al., 2016)。
为此, 本研究于2013年5月到2015年9月在四川瓦屋山次生常绿阔叶林中开展每月一次的氮添加试验, 并在氮添加处理1年后(2014年4月), 每月使用微根管技术对该林分建群种扁刺栲(Castanopsis platyacantha)根系影像进行收集, 连续收集18个月, 研究扁刺栲细根生物量、寿命和形态对短期氮添加的响应, 以期为深入理解氮沉降对森林地下生态系统过程的影响提供基础数据和参考。
1 材料和方法
1.1 研究区概况
试验地位于四川洪雅县南部的瓦屋山国家森林公园中山部的木姜杠林班(29.54° N, 103.26° E), 海拔1β600 m, 属于中亚热带季风性湿润气候。年平均气温10-14 ℃, 年日照时间不足800 h, 年降水量2 398 mm, 降水日199-226天, 年平均相对空气湿度85%-90%。该区域植被在1956年遭到砍伐破坏, 遗留下一些幼树幼苗, 后基本无进一步的干扰破坏而得以自然恢复。目前已形成次生常绿阔叶成林, 优势种为扁刺栲和中华木荷(Schima sinensis), 主要伴生树种为硬壳柯(Lithocarpus hancei)、野梧桐(Mallotus japonicus)、小叶青冈(Cyclobalanopsis myrsinifolia)等阔叶树种。该区域成土母质是晚古生界二叠纪至元古界的砂岩、玄武岩、花岗岩等古老岩层风化的残积物, 土壤类型为山地黄壤, 土壤厚约1 m, 其中腐殖质层厚约10 cm。土壤基本理化性质见表1。Table 1
表1
表1瓦屋山次生常绿阔叶林土壤理化性质(平均值±标准误差)
Table 1Soil physicochemical properties in a secondary evergreen broad-leaved forest, Wawu Mountain (mean ± SE)
| 土层深度 Soil depth (cm) | pH | 土壤容重 Soil bulk density (g·cm-3) | 有机碳含量 Organic carbon content (g·kg-1) | 全氮含量 Total nitrogen content (g·kg-1) | 全磷含量 Total phosphorus content (g·kg-1) | 全钾含量 Total potassium content (g·kg-1) |
|---|---|---|---|---|---|---|
| 0-10 | 3.19 ± 0.03 | 0.41 ± 0.03 | 121.9 ± 11.7 | 5.80 ± 0.30 | 0.50 ± 0.01 | 13.89 ± 0.92 |
| 10-40 | 3.76 ± 0.03 | 0.66 ± 0.02 | 26.6 ± 2.5 | 1.51 ± 0.08 | 0.26 ± 0.03 | 17.07 ± 0.82 |
| 40-70 | 3.97 ± 0.02 | 0.89 ± 0.03 | 12.6 ± 1.2 | 0.82 ± 0.05 | 0.18 ± 0.01 | 19.02 ± 1.17 |
| 70-100 | 4.03 ± 0.02 | 0.99 ± 0.03 | 7.8 ± 0.8 | 0.57 ± 0.04 | 0.21 ± 0.01 | 19.31 ± 1.26 |
新窗口打开
1.2 试验设计
2012年10月, 根据典型性和代表性原则在木姜杠林班设置9块20 m × 20 m的样地。各样地坡向均为北向, 坡度0-5°, 样地间缓冲距离大于20 m。使用NH4NO3进行氮添加处理, 共设3个水平: 对照(CK, 0 g·m-2·a-1), 低氮(LN, 5 g·m-2·a-1), 高氮(HN, 15 g·m-2·a-1), 每个处理设置3个重复。将各处理的NH4NO3年添加量均分为12份, 即得到各处理每月添加量, 从2013年5月到2015年9月, 每月下旬对各样地进行定量氮添加处理。具体方法是将各处理所需的NH4NO3溶解于10 L自来水中, 用喷雾器在该样方中来回均匀喷洒, CK只喷洒等量的自来水。1.3 微根管安装
2013年12月初, 于各样地内选择3株胸径大致相同(25 cm左右)、生长较好、相对独立的扁刺栲作为细根动态观测的目标树。在树干周围1/2冠幅的位置(到树干基部的距离约1 m)对称地安装两根由有机玻璃制成的微根管(Tingey et al., 2003)。微根管内径为6.4 cm, 长度为1 m。具体安装过程为: 先用外径略大于微根管外径的特制土钻朝向树干基部与垂直方向成45°角钻一个通直的洞, 深度约为80 cm (换算成垂直深度约为56 cm), 然后沿洞将微根管插入土壤中, 插入深度80 cm。露出地表的微根管部分先用黑色胶布缠绕, 以防光线透入, 然后再用白色胶布覆盖, 以防吸热, 最后在顶端加盖密封, 以防灰尘、水或降雨和其他物体落入管中。在每根管的外壁刻划一条细线, 以保证每次取样时都以该位置为起点进行影像数据收集。1.4 图像采集和土样根样收集
从2014年4月到2015年9月, 每月下旬采用CI-600扫描仪(CID Inc., Vancouver, Washington, USA)进行根系影像收集。每次扫描时, 从地表开始按20 cm (换算成垂直深度约为15 cm)的间距每管收集3张图片。即分别获取0-15 cm土层(I层)、15-30 cm土层(II层)、30-45 cm土层(III层)的根系图片。每次的影像收集工作在1天内完成。在观察期间共收集到了2β916张图片(3层× 6管× 9样地× 18次)。2014年11月(氮处理1.5年后), 在每个样地内随机选取5个采样点, 使用土钻钻取0-10 cm层(腐殖质层)土壤样品充分混合后带回实验室进行养分测定。为了计算扁刺栲细根比根长(SRL)以计算细根生物量, 于2016年4月用土块法获取每个样地的扁刺栲根系, 具体操作为: 在每个样地的每株目标树基部50-150 cm的圆环区域内用自制取样器挖取一个30 cm × 30 cm × 20 cm (长×宽×深)大小的土块; 由于试验区域土壤疏松, 在样地内将土块中的根系小心挑出, 将土壤分层回填到挖取的区域; 取出的根系存放于低温保温箱内, 迅速带回实验室; 用自来水冲洗掉附着在根系表面较多的土壤, 再用低温去离子水将根系完全清洗干净, 装袋编号后冷藏储存以保持其活性。1.5 图像分析和土样及比根长测定
微根管影像带回实验室后, 使用Adobe Photoshop CS6软件调节图片亮度和对比度, 并且对每段根系进行编号并记录每张图片的根段数量。用根系分析系统WinRHIZOTron MF 2012b软件对每个影像中的根系长度、表面积、直径进行量化, 以及根系类型的区分, 由于扁刺栲新生根表皮呈白色, 之后很快变为红色, 由此可以将其与其他树种根系进行区分, 并且因此将白色和红色根定义为活根, 黑色的根、皮层脱落或表皮褶皱的根定义为死根; 另外, 根系在没表现出来死亡形态时的突然消失, 也定义为死根。根系寿命定义为第一次在影像中出现时刻到第一次被定义为死亡或消失的时间。土壤样品带回实验室后挑出土壤中的杂质, 过2 mm筛后冷藏保存, 测定土壤pH值、铵态氮(NH4+-N)含量、硝态氮(NO3--N)含量, 取部分鲜土风干后过0.25 mm筛用于测定土壤全氮(TN)含量。使用电位法(LY/T 1239-1999)测定土壤pH值, TN用半微量凯氏法(LY/T 1228-1999)测定, NO3--N用紫外分光光度法(宋歌等, 2007)测定, NH4+-N用2 mol·L-1的KCl浸提-靛酚蓝比色法(鲍士旦, 2000)测定。土块法获取的扁刺栲根系清洗干净, 挑除死根后取直径<1 mm细根, 使用Epson数字化扫描仪(Expression 10000XL, Epson Electronics, San Jose, USA)扫描。用WinRHIZO (pro 2007b)根系图像分析软件对根扫描图像进行根长的定量分析。扫描分析完成后将细根在65 ℃烘箱中烘干至恒质量, 记录干质量。计算不同氮处理下的比根长(SRL), SRL =根长/干质量。
1.6 数据处理和分析
1.6.1 根系寿命建立根系寿命数据库, 将影像中相连但不存在分叉的一段根系(即根段)作为统计的基本单位。每一段根系都赋予指定ID号, 按照土壤层次、氮添加水平分别归类。在Excel中统计各根段的存活天数, 使用Kaplan-Meier方法进行生存分析, 估算细根平均寿命及中值寿命(即存活率达到50%时所用的时间), 并做存活曲线; 使用对数秩检验(log-rank test)比较不同氮处理对存活曲线的影响。
1.6.2 细根生物量
参考吴伊波等(2014)的计算方法, 估算单位土壤体积扁刺栲细根生物量。
首先, 利用每张影像中的根系长度, 计算单位土壤体积的根长密度(RLD, mm·cm-3)。计算公式 如下:
RLD = RL/(A × DOF) (1)
式中, RL (mm)为每张影像中的根系长度; A (cm2)为影像的面积; DOF (cm)为微根管观察可代表的土壤距离, 本研究中取0.25 cm。
然后, 通过SRL (m·g-1)将RLD转化为单位土壤体积的细根生物量(RBD, g·m-3)。
RBD = RLD/SRL × 1000 (2)
式中, SRL为利用土块法取样测定的直径小于1 mm的扁刺栲细根比根长。对照处理SRL为10.0 g·m-3, 低氮处理SRL为10.6 g·m-3, 高氮处理SRL为9.4 g·m-3。
1.6.3 根段形态
影像分析过程中发现, 影像中的根系绝大多数属于扁刺栲的1级或2级细根, 因此我们使用根段为单位来量化根系的形态特征。利用每张影像中的根系长度(RL, cm)、根系表面积(RA, cm2)和根段数量(N)计算根段长(RSL, cm)和根段表面积(RSA, cm2)。计算公式如下
RSL = RL/N(3)
RSA = RA/N(4)
1.6.4 统计分析
计算观察期内各个指标的样地平均值, 并使用平均值进行统计分析。使用单因素方差分析和LSD多重比较(α = 0.05)检验氮添加处理对土壤pH值、TN、NO3--N、NH4+-N、根段数量、根段长、根段表面积、根直径和生物量影响的显著性。对0-10 cm土壤养分指标和I土层根形态指标及中值寿命数据进行Pearson相关分析(两尾, α = 0.05, n = 9)。所有统计分析均在SPSS 20.0 (IBM SPSS, Chicago, USA)软件中执行。
2 结果和分析
2.1 氮添加对土壤化学指标的影响
与CK相比, HN处理显著降低了土壤pH值(p < 0.05)。土壤TN、NO3--N含量均表现出随氮添加梯度增加而增加的趋势, 但未达到统计学显著水平(表2)。相关分析结果表明土壤pH值与NH4+-N、NO3--N含量呈显著负相关关系(表3)。
Table 2
表2
表2氮添加对表层土壤(0-10 cm)化学养分和pH值的影响(平均值±标准误差)
Table 2The effect of N addition on surface soil (0-10 cm) chemical element and pH value (mean ± SE)
| pH | TN (g·kg-1) | NH4+-N (mg·kg-1) | NO3--N (mg·kg-1) | |
|---|---|---|---|---|
| CK | 3.99 ± 0.05a | 6.62 ± 0.50 | 25.62 ± 5.79 | 25.47 ± 3.90 |
| LN | 3.82 ± 0.02ab | 7.19 ± 0.57 | 21.63 ± 6.02 | 26.27 ± 4.65 |
| HN | 3.78 ± 0.08b | 7.23 ± 0.35 | 40.15 ± 12.39 | 36.69 ± 4.61 |
| 单因素方差分析 One-way ANOVA analysis | p = 0.04 | p = 0.62 | p = 0.33 | p = 0.21 |
新窗口打开
Table 3
表3
表3I层(0-15 cm)扁刺栲细根形态指标、中值寿命和表层土壤(0-10 cm)理化指标相关分析结果
Table 3Layer I (0-15 cm) correlation analysis results of Castanopsis platyacantha fine root morphology index, median lifespan, and soil physicochemical property (0-10 cm)
| 中值寿命 Median lifespan | pH | 总氮 Total nitrogen | NH4+-N | NO3--N | 根段长 Root segment length | 根段表面积 Root segment surface-area | |
|---|---|---|---|---|---|---|---|
| pH | 0.84** | ||||||
| 总氮 Total nitrogen | -0.54 | -0.57 | |||||
| NH4+-N | -0.34 | -0.75* | 0.45 | ||||
| NO3--N | -0.41 | -0.67* | 0.54 | 0.72* | |||
| 根段长 Root segment length | 0.37 | 0.35 | -0.15 | 0.01 | -0.18 | ||
| 根段表面积 Root segment surface-area | 0.73* | 0.74* | -0.50 | -0.24 | -0.50 | 0.76* | |
| 根直径 Root diameter | 0.48 | 0.47 | -0.32 | -0.27 | -0.14 | -0.48 | 0.15 |
新窗口打开
2.2 氮添加对扁刺栲细根生物量的影响
扁刺栲细根(<1 mm)在I、II和III土层生物量分别是65.01 g·m-3、40.79 g·m-3和22.50 g·m-3, 随土层深度增加而不断降低(图1)。CK、LN、HN处理下0- 45 cm土层细根生物量分别为128.30 g·m-3、 125.91 g·m-3、143.87 g·m-3, 无显著差异。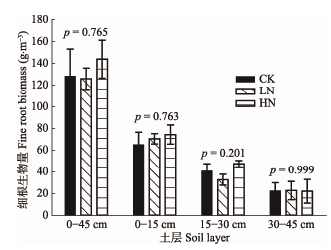 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1不同土层细根生物量(平均值±标准误差)。CK, 对照 (0 g·m-2·a-1); LN, 低氮(5 g·m-2·a-1); HN, 高氮(15 g·m-2·a-1)。p值为不同土层单因素方差分析的结果。
-->Fig. 1Fine root biomass in different soil layer (mean ± SE). CK, control (0 g·m-2·a-1); LN, low nitrogen addition (5 g·m-2·a-1); HN, high nitrogen addition (15 g·m-2·a-1). p value is the results of one-way ANOVA analysis in different soil layer.
-->
2.3 氮添加对扁刺栲细根寿命的影响
扁刺栲细根的平均寿命与土层深度有关, 表现为: I层> III层> II层。对照处理中, I、II、III层细根的平均寿命分别为: 186、113、130天(表4)。生存曲线的对数秩检验结果表明, 3个土层中氮添加处理对细根寿命的影响均未达到统计学显著水平(图2)。Pearson相关分析表明, I层土壤扁刺栲细根的中值寿命与表层土壤的pH值、根段表面积呈显著正相关关系(表3)。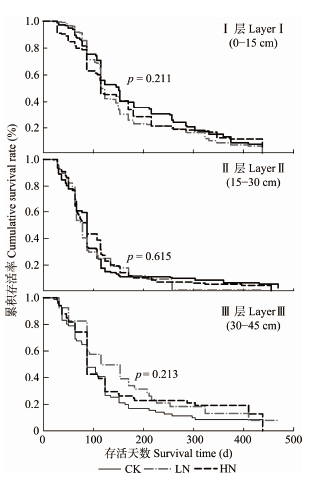 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2氮添加对不同土层细根生存曲线的影响。p值为氮添加处理对同一土层细根生存曲线的显著性检验结果。CK, 对照; LN, 低氮; HN, 高氮。
-->Fig. 2Effects of nitrogen additions on fine root survival curve. p values are the survival curves significance test result of N addition in the same soil layers. CK, control; LN, low nitrogen; HN, high nitrogen.
-->
2.4 氮添加对扁刺栲细根形态的影响
在18个月的观察期内, 共观察到1β179条根(直径<1 mm, 均属细根范畴), 其中I层548条, II层459条, III层172条。I、II、III层扁刺栲根系的直径分别为(0.86 ± 0.13) mm、(0.61 ± 0.01) mm和(0.67 ± 0.06) mm。与CK相比, LN处理对I层细根根段表面积的影响达到了显著水平(p = 0.049, 降低了48.8%)。氮添加未显著影响各土层扁刺栲根段长、根直径。相关分析结果表明, I层扁刺栲根段表面积与土壤pH值呈显著的正相关关系(表3)。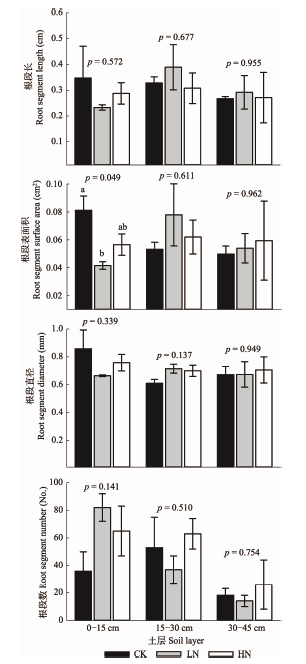 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3氮添加对不同土层细根直径、根段数、根段长、根段表面积的影响(平均值±标准误差)。CK, 对照; LN, 低氮; HN, 高氮。图中文字为对应土层的单因素方差分析结果; 图中字母表示氮添加处理对同一土层各形态指标的单因素方差分析结果。
-->Fig. 3Effects of nitrogen additions on fine root diameter, length and superficial area at different soil layers (mean ± SE). CK, control; LN, low nitrogen; HN, high nitrogen. Text and letters in figure are the result of one-way ANOVA analysis on same morphology indexes in the same soil layers.
-->
3 讨论
3.1 氮添加对土壤无机氮含量和土壤pH值的影响
氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势。土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(Huang et al., 2014)。一方面, 当NH4+被植物根系所吸收时, 一个H+将会释放到土壤溶液中, 增加土壤溶液酸度。另一方面, 当土壤中的无机氮含量增加, 特别是超过了植物和微生物的需求时, 土壤中的硝化作用则会加强, 其结果使得土壤溶液中的H+增加(Evans et al., 2008)。H+由于具有较强的阳离子交换能力, 可将Na+、K+、Mg2+、Ca2+等盐基阳离子从胶体颗粒中交换到土壤溶液中。这一过程加速了湿润地区土壤盐基离子通过淋溶作用的损失(Lucas et al., 2011), 并导致土壤对酸的缓冲能力减弱(Tian & Niu, 2015)。在本研究区域, 由于背景氮沉降量很高, 达到90 kg·hm-2·a-1 (Xu et al., 2013)。进一步的氮添加处理(尤其是高氮处理)使得无机氮含量接近或超过了植物和微生物的需求, 可能导致了土壤接近或超过氮饱和, 因此在高氮处理后几年内就观察到土壤pH值显著降低。有较多研究观察到氮添加导致土壤酸化, 并且往往伴随着土壤盐基离子的损失, 严重的会导致土壤中Al3+等金属离子活化(Lucas et al., 2011)。例如, 本课题组前期研究发现, 持续每月一次的氮添加试验6年后, 苦竹(Pleioblastus amarus)人工林土壤交换性Al含量显著增加, 其中高氮处理下增幅达70% (Chen et al., 2017)。而土壤酸化和金属离子活化会对物种多样性、植被生产力、森林健康、土壤生物群落造成严重影响(肖辉林, 2001)。3.2 氮添加对扁刺栲细根寿命的影响
细根寿命由植物的本身特性包括根形态、分布深度、菌根共生、植物生命形态等和外在特性包括土壤氮和水分可利用性、温度等因素共同控制, 因此不同树种及同一树种生长在不同环境中均表现出不同的根寿命(Chen & Brassard, 2013)。本研究结果表明I层扁刺栲根系的平均寿命和中值寿命分别为186天和151天。与中亚热带区域的树种相比, 扁刺栲细根中值寿命低于常绿阔叶树种细柄蕈树(Altingia gracilipes, 184天)和米槠(Castanopsis carlesii, 212天)的中值寿命, 以及针叶树种杉木(Cunninghamia lanceolata, 236天)的中值寿命(袁一丁等, 2009; 黄锦学等, 2012); 高于闽楠(Phoebe bournei, 89天)的中值寿命(郑金兴等, 2012)。在本研究中, 虽然低氮和高氮处理均使I层扁刺栲细根的中值寿命降低了24%, 但是未达到统计学显著水平。传统理论认为, 土壤氮可利用性增加, 通常提高植物地上部分的生长速率, 进而对养分元素的需求也相应增加。植物为了支持地上部分对养分的需求, 需要提高细根对土壤养分的吸收能力, 而新鲜的细根具有更强的养分获取能力, 因此植物会提高根系统中年轻细根的比例, 降低细根的寿命, 加快整个根系统的周转速率(Eissenstat & Yanai, 1997)。因此, 氮添加可能通过降低扁刺栲根系的中值寿命, 加快根系周转速率以提高其吸收能力。
但是, 氮添加试验(Mei et al., 2010)和背景氮梯度研究结果(Burton et al., 2000)并不总支持这一理论。细根对氮素添加的不同响应可能归因于两方面。首先是区域土壤养分可利用性差异(Chen & Brassard, 2013)。当土壤氮限制比较严重, 长寿命的细根可能只能给予植物较少的养分回报, 这时当土壤氮可利用性提高时, 可能会极大地改善目前细根所处环境的养分状况, 植物提高细根寿命对植物养分获取更加有利。而土壤氮限制较小的状况下, 氮添加可能会促进根系向根寿命更短的方向转变, 因为与维持根系的碳投入相比, 发育新的细根可能对植物更为有利。其次是树种差异。叶和根经济谱的研究表明不同的树种具有不同的资源获取策略, 有的树种采取“快速投资-收益”策略, 有的树种采取“保守投资-收益”策略。不同的策略在适应土壤环境和养分含量变化时, 会有不同的表现(陈莹婷和许振柱, 2014; Weemstra et al., 2016)。于水强等(2009)同时研究了水曲柳(Fraxinus mandschurica)和落叶松(Larix gmelinii)细根寿命对土壤氮添加的响应, 结果表明水曲柳细根中值寿命与土壤氮可利用性表现出正相关关系, 而落叶松细根中值寿命对土壤氮可利用性的变化没有显著响应。因此, 土壤初始养分状况和树种对于细根寿命对氮添加的响应有重要的影响。对于本研究试验样地, 由于土壤氮素含量较高, 并且具有很高的背景氮沉降量。区域天然林植被可能处于氮饱和状态, 因此进一步的氮添加并未对扁刺栲的根系养分吸收策略产生显著影响, 导致氮添加未对扁刺栲根寿命产生显著影响。
3.3 氮添加对扁刺栲细根生物量和形态特征的影响
氮添加处理未影响3个土层中扁刺栲的细根生物量。目前, 氮添加对细根生物量的影响存在减少、增加、不影响3种情况(Wang et al., 2012; Noguchi et al., 2013; Kou et al., 2015)。细根生物量对氮添加的不同响应方向可能与不同气候区域有关。Li等(2015)整合全球相关研究表明, 氮添加显著增加了北方森林的细根生物量, 而没有改变热带、亚热带森林的细根生物量, 分析这可能与区域土壤养分状况有关; 另外氮添加对细根生物量的影响也与氮添加量以及氮处理时间有关。在本研究中, 氮添加使I层扁刺栲细根的根段长和根段表面积呈下降趋势, 其中低氮处理对根段表面积的影响达到显著(p = 0.049)水平。植物依靠根系获取自身需要的多种元素, 由于土壤各种养分具有较强的空间异质性。植物根系利用其强大的可塑性予以应对, 形成了“成本-效益”策略来适应各种土壤养分分布状况(Bardgett et al., 2014)。然而在本研究中, 虽然氮添加使表层土壤的无机氮增加了8.6%-9.2%, 但相关分析结果表明I层扁刺栲根形态指标与表层土壤NH4+-N、NO3--N含量之间并无显著的相关关系, 而根段表面积与土壤pH值之间存在显著的正相关关系。这说明在该次生常绿阔叶林中, 与无机氮可利用性相比, 扁刺栲形态特征与土壤pH值有更为紧密的关系。
4 结论
高氮处理显著降低了该次生林表层土壤pH值, 低氮处理显著降低了0-15 cm土层扁刺栲细根的根段表面积。根段表面积与土壤pH值之间存在显著的正相关关系。氮添加处理未影响扁刺栲细根寿命和生物量。本研究表明氮添加不会影响扁刺栲细根的寿命和生物量。仅可能通过改变该次生林表层土壤无机氮含量, 导致土壤一定程度的酸化, 进而对表层土壤的扁刺栲根系形态特征造成一定的影响。The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | Fine and coarse roots are key contributors to belowground net primary productivity, and play critical roles in the biogeochemical cycling of forest and woodland ecosystems. Despite their critical roles, roots have been understudied mainly due to methodological challenges. There is currently no consensus on which methods are most suitable to accurately study root biomass and production. Critical evaluation of the assumptions, strengths and inherent limitations of methods to study root biomass and production are necessary to help investigators decide which method is best for their purposes. This synthesis compares existing methods for root biomass and production estimation based on relevant criteria that include cost, labor requirements, time efficiency and accuracy and, also compares fine- and coarse-root biomass and production estimates from different methods measured at the same sites. Root excavation and soil-pit methods are commonly used to estimate coarse-root biomass, despite the high cost and labor required. Ground-Penetrating Radar is a very promising indirect approach to estimate coarse-root biomass, but may not be suitable for ecosystems with dense understory and soils with high organic matter and ion contents. Soil-core remains the most preferred method to estimate fine-root biomass. Empirical models are accepted as fast and cost-effective indirect approach to predict fine- and coarse-root biomass and production. Fine-root production is usually estimated with the (mini) rhizotrons, sequential-coring and ingrowth-core methods. Coarse-root biomass estimates were not significantly different between soil-pit and soil-core methods. There was a significant positive correlation ( r 2 02=020.91, p 02<020.0001) between fine-root biomass estimates obtained from soil-pit and soil-core methods. Fine-root production estimates were lower in the ingrowth-core (2.0602±020.2302Mg02ha 611 02year 611 ) compared to the (mini) rhizotrons (3.8102±020.4602Mg02ha 611 02year 611 ) and sequential-coring (3.8402±020.9302Mg02ha 611 02year 611 ) methods. Based on the reviewed literature and comparative analysis we propose that (mini) rhizotrons should be preferred over the others in estimating fine-root production. In situations where cost and site conditions preclude their use, the sequential-coring and ingrowth-core methods are suitable. The ingrowth-core should be used with caution in sites where root growth is slow and root biomass may be influenced by strong seasonal fluctuations. Multiple methods are still recommended for yielding realistic estimates of fine- and coarse-root production, and more comparative studies of different methods should be conducted on the same sites. |
| [2] | Despite their critical role in every terrestrial ecosystem, fine root production and mortality have not been widely compared among systems due to the practical difficulties of belowground research. We examined fine root production and mortality among five contrasting sites: native and invaded grassland in eastern Montana, USA, aspen forest in southern Saskatchewan, Canada, and birch forest and tundra in northern Sweden. Additionally, we investigated the importance of minirhizotron sampling interval on measures of root production and mortality by comparing measures produced from 1-, 7-, 14-, and 21-day sample intervals. Root length and mortality varied significantly among sites, with invaded grassland having the greatest root length (> 2 x than any other site) and significantly greater root mortality than native grassland (54 %). In contrast, there were no significant differences in root production among the sites. Sample interval had no significant influence on root production or mortality. Minirhizotron sampling intervals up to 3 weeks did not underestimate the measures of root production and mortality in comparison to measures derived from shorter sampling intervals, regardless of the site studied. The results suggest that 3 weeks can be an accurate and efficient sample interval when studying root production and mortality with minirhizotrons. |
| [3] | |
| [4] | Ecologists are increasingly adopting trait-based approaches to understand how community change influences ecosystem processes. However, most of this research has focussed on aboveground plant traits, whereas it is becoming clear that root traits are important drivers of many ecosystem processes, such as carbon (C) and nutrient cycling, and the formation and structural stability of soil. Here, we synthesise emerging evidence that illustrates how root traits impact ecosystem processes, and propose a pathway to unravel the complex roles of root traits in driving ecosystem processes and their response to global change. Finally, we identify research challenges and novel technologies to address them. |
| [5] | The extent to which atmospheric N deposition is enhancing primary production and CO sequestration along the ambient N deposition gradients found within many regional temperate forest ecosystems remains unknown. We used tree diameter measurements from 1984 and 2004, allometric equations, and estimates of wet N deposition from 32 permanent plots located along an ambient N deposition gradient in the Adirondack Park, New York, USA, to determine the effects of N deposition on the basal area and woody biomass increments (BAI and WBI, respectively) of individual stems from all the major tree species. Nitrogen deposition had either a neutral or positive effect on BAI and WBI, with the positive effects especially apparent within the smaller size classes of several species. The nature of these growth responses suggests that other co-varying factors (e.g., temperature, tropospheric ozone, soil acidification) may be partially counteracting the species-dependent fertilization effect of N deposition that was suggested by recent foliar N data across this gradient. Nevertheless, in documenting a fertilization effect from chronic, low-level, ambient rates of N deposition, this study underscores the need for more research on how N deposition is affecting rates of primary production, CO鈧 sequestration, and even vegetation dynamics in many forests worldwide. |
| [6] | Minirhizotrons were used to observe fine root (09‰¤1 mm) production, mortality, and longevity over 2 years in four sugar-maple-dominated northern hardwood forests located along a latitudinal temperature gradient. The sites also differed in N availability, allowing us to assess the relative influences of soil temperature and N availability in controlling fine root lifespans. Root production and mortality occurred throughout the year, with most production occurring in the early portion of the growing season (by mid-July). Mortality was distributed much more evenly throughout the year. For surface fine roots (009“10 cm deep), significant differences in root longevity existed among the sites, with median root lifespans for root cohorts produced in 1994 ranging from 405 to 540 days. Estimates of fine root turnover, based on the average of annual root production and mortality as a proportion of standing crop, ranged from 0.50 to 0.68 year09“1 for roots in the upper 30 cm of soil. The patterns across sites in root longevity and turnover did not follow the north to south temperature gradient, but rather corresponded to site differences in N availability, with longer average root lifespans and lower root turnover occurring where N availability was greater. This suggests the possibility that roots are maintained as long as the benefit (nutrients) they provide outweighs the C cost of keeping them alive. Root N concentrations and respiration rates (at a given temperature) were also higher at sites where N availability was greater. It is proposed that greater metabolic activity for roots in nitrogen-rich zones leads to greater carbohydrate allocation to those roots, and that a reduction in root C sink strength when local nutrients are depleted provides a mechanism through which root lifespan is regulated in these forests. |
| [7] | Plant roots play a critical role in ecosystem underground processes, and are potentially influenced by elevated atmospheric nitrogen (N) deposition. However, the effects of N deposition or long-term N |
| [8] | Although fine roots play an integral role in biogeochemical cycling and supporting plant function, fundamental understanding of the mechanisms that control fine root life span is limited. Based on literature, we examined how intrinsic plant characteristics including root diameter, root branching order, rooting depth, and mycorrhizal symbiosis affect fine root life span, and how fine root life span differs with plant life form and foliar habit and between early versus late seral species. We also examined how soil nitrogen and water availability, temperature, and atmospheric carbon dioxide concentration influence fine root life span. We focused on evidence from rhizotron and minirhizotron observations which allow for individual roots to be directly monitored in situ. Fine root life span increased with increasing root diameter, was shorter for more distal than proximal roots, and increased with increasing rooting depth, but was not influenced by mycorrhizal symbiosis. Trees had the longest fine root life spans of all the plant life forms, followed by grasses, lianas, shrubs, and forbs. Among trees, deciduous species had shorter fine root life spans than evergreen species. Fine root life span appears to decrease with increasing temperature and increase with soil water availability, whereas the effects of soil nitrogen availability and atmospheric carbon dioxide concentration on fine root life span were highly inconsistent among studies. Our findings indicate that root morphological characteristics and plant traits are useful predictors of fine root life span. However, environmental influences on fine root life span remain poorly understood due to the limited number of respective studies. Future studies of root demographic processes are needed to better understand environmental controls of fine root life span. It is also critical that research continues into developing more direct and less invasive techniques for studying root demographics. |
| [9] | . The concept of leaf economics spectrum (LES) has attracted much attention and debate since its emergence. It for the first time provides quantitative analysis of plant functional traits and their relationships on the global scale, hence quantifying and generalizing the context and variations of the trade-off strategies. This is of great theoretical value, and provides a useful research method and scientific ideas for subsequent study on plant traits and their functions. In this paper, we try to comprehensively review the meaning, contents, relevant verifications and objections about LES, and to explore its underlying mechanisms. In addition, we emphasize the multi-scale and multidimensional extensions, integration and potential applications of LES. Currently there are still several shortcomings about LES research in China, and we outlook the development of LES theory domestically and abroad. It may be of significance for ecological researchers to establish and exploit jointly a global database on plant traits. |
| [10] | |
| [11] | Abstract Root turnover is important to the global carbon budget as well as to nutrient cycling in ecosystems and to the success of individual plants. Our ability to predict the effects of environmental change on root turnover is limited by the difficulty of measuring root dynamics, but emerging evidence suggests that roots, like leaves, possess suites of interrelated traits that are linked to their life span. In graminoids, high tissue density has been linked to increased root longevity. Other studies have found root longevity to be positively correlated with mycorrhizal colonization and negatively correlated with nitrogen concentration, root maintenance respiration and specific root length. Among fruit trees, apple roots (which are of relatively small diameter, low tissue density and have little lignification of the exodermis) have much shorter life spans than the roots of citrus, which have opposite traits. Likewise, within the branched network of the fine root system, the finest roots with no daughter roots tend to have higher N concentrations, faster maintenance respiration, higher specific root length and shorter life spans than secondary and tertiary roots that bear daughter roots. Mycorrhizal colonization can enhance root longevity by diverse mechanisms, including enhanced tolerance of drying soil and enhanced defence against root pathogens. Many variables involved in building roots might affect root longevity, including root diameter, tissue density, N concentration, mycorrhizal fungal colonization and accumulation of secondary phenolic compounds. These root traits are highly plastic and are strongly affected by resource supply (CO 2 , N, P and water). Therefore the response of root longevity to altered resource availability associated with climate change can be estimated by considering how changes in resource availability affect root construction and physiology. A cost enefit approach to predicting root longevity assumes that a plant maintains a root only until the efficiency of resource acquisition is maximized. Using an efficiency model, we show that reduced tissue Nconcentration and reduced root maintenance respiration, both of which are predicted to result from elevated CO 2 , should lead to slightly longer root life spans. Complex interactions with soil biota and shifts in plant defences against root herbivory and parasitism, which are not included in the present efficiency model, might alter the effects of future climate change on root longevity in unpredicted ways. |
| [12] | This chapter discusses the competing theories of root lifespan and reviews the evidence available to support them. New methods of root observation and analysis produce data appropriate to testing these theories, but the results till date are few and often conflicting. Tentative generalizations include a suggestion that small diameter roots with low tissue density tend to have short lifespan. Root lifespan appears to be longest in cold environments, but data are lacking for tropical species. There is a strong seasonal variation in lifespan, with roots produced in the fall surviving longest, at least in temperate climates. Species differences are difficult to quantify because of seasonal and interannual variation, but root lifespan of deciduous fruit crops seems to be shorter than those of temperate deciduous forest trees or citrus, a broadleaf evergreen. The current model of root efficiency omits some important factors that may exert control over root lifespan. Fine roots have other functions in addition to absorption, including transport of water and nutrients. Furthermore, seasonality of climate and the need for carbon and nutrient storage could constrain the root lifespan that optimizes plant fitness to differ from that which maximizes root efficiency. |
| [13] | Nitrate (NO 3 61) is often observed in surface waters draining terrestrial ecosystems that remain strongly nitrogen (N) limited. It has been suggested that this occurs due to hydrological bypassing of soil or vegetation N retention, particularly during high flows. To test this hypothesis, artificial rain events were applied to 12 replicate soil blocks on a Welsh podzolic acid grassland hillslope, labelled with 15N-enriched NO 3 61 and a conservative bromide (Br 61) tracer. On average, 31% of tracer-labelled water was recovered within 4 h, mostly as mineral horizon lateral flow, indicating rapid vertical water transfer through the organic horizon via preferential flowpaths. However, on average only 6% of 15N-labelled NO 3 61 was recovered. Around 80% of added NO 3 61 was thus rapidly immobilised, probably by microbial communities present on the surfaces of preferential flowpaths. Transitory exceedance of microbial N-uptake capacity during periods of high water and N flux may therefore provide a mechanism for NO 3 61 leaching. |
| [14] | This paper examines the impact of food and energy production on the global N cycle by contrasting N flows in the late-19~th century with those of the late-20~th century. We have a good understanding of the amounts of reactive N created by humans, and the primary points of loss to the environment. However, we have a poor understanding of nitrogen's rate of accumulation in environmental reservoirs, which is problematic because of the cascading effects of accumulated N in the environment. The substantial regional variability in reactive nitrogen creation, its degree of distribution, and the likelihood of increased rates of reactive-N formation (especially in Asia) in the future creates a situation that calls for the development of a Total Reactive Nitrogen Approach that will optimize food and energy production and protect environmental systems. |
| [15] | |
| [16] | SHORT-ROTATION; EUCALYPTUS-CAMALDULENSIS; LITTER DECOMPOSITION; ABOVEGROUND BIOMASS; MIXED PLANTATIONS; STORAGE CAPACITY; SOUTHERN CHINA; WATER; INDIA; CONSERVATION |
| [17] | Soil acidification is defined as the process in which exchangeable cations are leaching and soil H + concentration is raising thereby increases soil acidity. Changes in soil pH value and acid neutralizing capacity are mainly indicators of soil acidification. Soil acidification is considered to be a serious ecological and environmental issue, which not only reduces soil quality, but also decreases biodiversity of forest ecosystem and induces forest decline. With nitrogen (N) deposition rapidly increasing, its contribution to soil acidification becomes a major concern in the world. However, the impact of increased N deposition on soil acidification is not well addressed highlighting the need for further attention to the issue. In this paper, the studies on forest soil acidification induced by N deposition were reviewed. The factors related to soil acidification driven by N deposition were classified and discussed, which included soil acidic buffering capacity, N components in atmospheric N deposition, climate, plant species in forests, and N status in ecosystem. Iron (Fe) buffering phase and the consequent Fe toxicity occurring to the acidified soil caused by high N deposition were concerned. The scarcity of phosphorus (P) element induced by soil acidification was particularly emphasized. The research methods used to study soil acidification driven by N deposition were also evaluated. In the end we stressed the importance of the study on soil acidification especially in tropical and subtropical regions driven by N deposition and its mechanisms. This paper can serve for maintaining sustainable forest and agricultural ecosystems. |
| [18] | . 采用微根管技术与挖掘法相结合的方式对福建省万木林自然保护区细柄阿丁枫和米槠天然林细根生产和死亡动态进行了为期两年多的观测,分析细根生产和死亡的季节变化、垂直分布及径级和序级分配,并估计细根的年生产量和年死亡量。结果表明:细柄阿丁枫细根年生产量和年死亡量分别为(230.1±162.8) g·m-2·a-1和(188.8±75.5) g·m-2·a-1,均略大于米槠的(214.5±185.8) g·m-2·a-1和(178.8±26.5) g·m-2·a-1,但两种森林群落的细根年生产量和年死亡量均无显著差异(P>0.05)。两森林群落细根生产均在春季达到高峰,其中米槠细根生产与月降水量呈极显著相关(P<0.01, r=0.566);细根死亡则呈现季节性地波动,米槠细根死亡峰值主要发生于夏季和秋季,而细柄阿丁枫则出现在秋季。两森林群落细根生产和死亡皆主要集中于土壤表层0-40 cm中,而且不同径级细根生产和死亡集中于0-1 mm细根中,其中0.3-0.6 mm细根的生产和死亡在两森林群落中均最大。两森林群落一级根的生产和死亡均大于高级根。;The turnover of fine root, the most dynamic component of root system, play an important role in nutrient cycle and energy flow in forest ecosystems. Fine root production and mortality in two subtropical evergreen broadleaved forests, Altingia grlilipes forest and Castanopsis carlesii forest, in Wanmulin Natural Reserve, Jian-ou, Fujian province, were monitored for more than two consecutive years by using both minirhizotrons and the exvacation methods. The objectives of this research were to: (1) analyze the seasonal patterns of fine root production and mortality; (2) to examine the allocation of both root production and mortality among different soil depths, root diameter classes and orders. In order to extract biomass data of individual root from root image, the relationships between root biomass (M) and root diameter (D) and root length (L) were established: M=0.4671 D2L for Altingia grlilipes, and M=0.5869 D2L for Castanopsis carlesii, respectively. Though there were no significant differences (P>0.05), annual fine root production and mortality in Altingia grlilipes ( (230.1±162.8) g·m-2·a-1 and (188.8±75.5) g·m-2·a-1, respectively) were slightly higher than those in Castanopsis carlesii ((214.5±185.8) g·m-2·a-1 and (178.8±26.5) g·m-2·a-1, respectively). Fine root production reached a peak in the spring, and showed a normal distribution in 2009 in both two communities. There was a significant correlation between fine root production and monthly precipitation (P<0.01, r=0.566) in Castanopsis carlesii . Fine root mortality fluctuated seasonally in these two forests. The peak of fine root mortality occurred mainly in summer and autumn in Castanopsis carlesii and in fall in Altingia grlilipes. The fine root production of these two forest communities appeared mainly in the top soil layer (0-40 cm), accounting for 88.6% and 95.1% of the total production in Altingia grlilipes and Castanopsis carlesii , respectively. Fine root mortality had the same distribution pattern, and made up to 89.7% and 99.2% of total mortality, respectively, in the 0-40 cm layer. In addition, the maximum of fine root production (Altingia grlilipes: (131.7±91.4) g·m-2·a-1; Castanopsis carlesii : (108.6±100.4) g·m-2·a-1) and mortality (Altingia grlilipes: (106.4±69.5) g·m-2·a-1; Castanopsis carlesii : (99.7±2.4) g·m-2·a-1) of these two forest communities appeared in the 2040 cm soil layer. With the root diameter increases, the number of root growth and death decreased, while fine root production and mortality had no corresponding tendency with diameter. Root production occurred mainly in 0-1mm, which accounts for 90.4% of total production in Altingia grlilipes and 96.1% in Castanopsis carlesii . Furthermore, fine root production in 0.3-0.6 mm was (100.0±44.0) g·m-2·a-1 in Altingia grlilipes and (94.8±90.4) g·m-2·a-1 in Castanopsis carlesii , which was the highest among different root diameter classes. Fine root mortality had the same trend with production, which accounted for 95.5% and 100% of the total fine root mortality in 0-1mm in Altingia grlilipes and Castanopsis carlesii , respectively. Root production and mortality of the first root order were much higher than those of the higher order in both two forest communities. |
| [19] | Global biogeochemical models have improved dramatically in the last decade in their representation of the biosphere. Although leaf area data are an important input to such models and are readily available globally, global root distributions for modeling water and nutrient uptake and carbon cycling have not been available. This analysis provides global distributions for fine root biomass, length, and surface area with depth in the soil, and global estimates of nutrient pools in fine roots. Calculated root surface area is almost always greater than leaf area, more than an order of magnitude so in grasslands. The average C:N:P ratio in living fine roots is 450:11:1, and global fine root carbon is more than 5% of all carbon contained in the atmosphere. Assuming conservatively that fine roots turn over once per year, they represent 33% of global annual net primary productivity. |
| [20] | ABSTRACT We used minirhizotrons to assess the effects of elevated CO2, N, and season on the life-span of ponderosa pine (Pinus ponderosa Dougl. ex Laws.) fine roots. CO2 levels were ambient air (A), ambient air + 175 mu mol.mol(-1) (A + 175) and ambient air + 350 mu mol.mol(-1) (A + 350). N treatments were 0, 100, and 200 kg N.ha(-1) per year (N0, N100, and N200, respectively). Fine root survival was strongly influenced by season and seemed to be most strongly linked to soil temperature. Roots born in the fall and winter had longer median root life-span (MRLs) than those born during the spring and summer. Elevated CO2 increased root life-span, but N fertilization decreased it. Under A, MRL was 74 +/- 12 days (mean +/- SE) and was significantly different from the MRL for the A + 350 treatment (102 +/- 14 days). MRL under A + 175 averaged 92 +/- 10 days. MRL was 116 +/- 13 days for the N0 treatment and was significantly greater than MRL for the N100 (70 +/- 10 days) and N200 (62 +/- 14 days) treatments. Assuming that longer lived fine roots continue their resource acquisition functions, then elevated CO2 may have the effect of extending the resource acquisition period. In contrast, fine roots in N-rich environments have shorter life-spans than fine roots in N-poor environments. |
| [21] | |
| [22] | Background and aims Growth, morphological traits, and mycorrhizal colonization of fine roots show high degree of plasticity in response to changes in nutrient availability, causing shifts in root nutrient-foraging strategy. However, little is known about how this plasticity associated with root branching orders respond to atmospheric nitrogen (N) deposition in subtropical coniferous forests. Methods We used soil block sampling method to examine the responses of six key root functional parameters (including three morphological traits (specific root length (SRL), root tissue density (RTD), and root diameter), two growth indices (total root length (TRL) and biomass) on an areal basis across five root orders, and ectomycorrhizal (EM) tip colonization) to different doses and species of N addition in a slash pine (Pinus elliottii) plantation in subtropical China. Results TRL, root biomass in all root orders, and EM tip colonization increased significantly with N addition. However, SRL, RTD, and root diameter did not change in any root orders. In comparison to low doses of N input, high doses of N input exerted greater effects on lower-order roots. In regard to species of N added, stronger responses in lower-order roots were observed under ammonium-based than nitrated-based N input. Foliar P content was significantly decreased and stoichiometric N:P ratio was markedly increased in response to high dose of ammonium-based N input. Conclusions Fine root growth and EM tip colonization displayed higher degree of plasticity than morphological traits in response to N addition. The plastic responses were not root-order dependent, but dependent on both N dose and species, especially for ephemeral lower-order roots that are mostly like to be the main nutrient acquisition structures. Our results imply that while N limitation was alleviated by exogenous N input, P limitation may persist or even be exacerbated, thus causing an increase of absorptive root length, biomass, and dependence on ectomycorrhizae for nutrient acquisition in subtropical slash pine plantation forests. |
| [23] | |
| [24] | . |
| [25] | . 树木细根(直径〈2mm)在森林生态系统能量流动和物质循环中起着重要的作用。原有的细根生产周转研究中常采用的土钻法、内生长法、挖掘法、根室法和土柱法等,均不能直接观察到细根的动态变化。微根管法是一种非破坏性、可定点直接观察和研究植物根系的方法,为研究细根的生长、衰老、死亡、分解和再生长的过程提供了有效的工具,尤其适用于细根周转、寿命和分解等方面的研究。但该技术不能直接测定单位面积的细根生物量、细根化学组成及细根周转对土壤碳和养分循环的影响,需要与土钻法结合。本文就运用微根管法对细根生物量、生产、周转和寿命等指标的研究方法进行了评述。 |
| [26] | Global atmospheric nitrogen deposition has increased steadily since the 20th century, and has complex effects on terrestrial ecosystems. This work synthesized results from 54 papers and conducted a meta-analysis to evaluate the general response of 15 variables related to plant root traits to simulated nitrogen deposition. Simulated nitrogen deposition resulted in significantly decreasing fine root biomass (<2mm diameter;6112.8%), while significantly increasing coarse root (≥2mm diameter;+56.5%) and total root (+20.2%) biomass, but had no remarkable effect on root morphology. This suggests that simulated nitrogen deposition could stimulate carbon accumulation in root biomass. The root: shoot ratio decreased (6110.7%) suggests that aboveground biomass was more sensitive to simulated nitrogen deposition than root biomass. In addition, simulated nitrogen deposition increased the fine root nitrogen content (+17.6%), but did not affect carbon content, and thus decreased the fine root C:N ratio (6113.5%). These changes delayed the decomposition of roots, combined with increasing of the fine root turnover rate (+21.4%), which suggests that simulated nitrogen deposition could increase carbon and nutrient retention in the soil. Simulated nitrogen deposition also strongly affected the functional traits of roots, which increased root respiration (+20.7%), but decreased fungal colonization (6117.0%). The effects of simulated nitrogen deposition on the plant root systems were dependent on ecosystem and climate zone types, because soil nutrient conditions and other biotic and abiotic factors vary widely. Long-term simulated experiments, in which the experimental N-addition levels were less than twofold of the average of atmospheric nitrogen deposition, would better reflect the response of ecosystems under atmospheric nitrogen deposition. These results provide a synthetic understanding of the effects of simulated nitrogen deposition on plant root systems, as well as the mechanisms underlying the effects of simulated nitrogen deposition on plants and the terrestrial ecosystem carbon cycle. |
| [27] | The dominant base cations (BC; i.e., Ca 2+, Mg 2+, K +, and Na +) are important in buffering soil and water acidity in both terrestrial and aquatic ecosystems. Ca 2+, Mg 2+, and K + are also important in many plant physiological functions. Because BC availability is affected by changes in the nitrogen (N) cycle, we conducted a meta-analysis of previously published data to determine if N fertilization alters the availability of BC in terrestrial and stream ecosystems across biomes. We include data from 107 independent studies published in 62 different articles, taking a holistic perspective on BC by examining their responses to added N in plant foliage, bulk soil, soil solution, and stream water. Our results suggest N fertilization may accelerate BC loss from terrestrial ecosystems over time periods less than five years. We found that N additions resulted in an overall 24% decrease in the availability of exchangeable Ca 2+, Mg 2+, and K + in the bulk soil of boreal forest, temperate forest, and grassland biomes. Collectively, responses of BC in boreal forest, temperate forest, tropical forest, and grassland biomes increased following N fertilization by about 71% in soil solution and 48% in stream waters. Additionally, BC responses in foliage decreased in boreal forest and temperate forest biomes following N additions over time periods less than five years, but there were no significant changes over longer time periods. Despite large short-term shifts in BC responses following N additions, we did not find evidence of widespread negative impacts on ecosystems over time periods greater than five years. This analysis suggests effects of N addition on the availability of exchangeable BC may diminish over time. Although the effects on BC can be substantial over periods less than five years, there is little available evidence that N fertilization has had large-scale detrimental effects on the availability of BC needed for plant growth within terrestrial or aquatic ecosystems. |
| [28] | . 人类活动在全球范围内极大地改变着氮素从大气向陆地生态系统输入的方式和速率,人为固定的氮素正在不断积累,并对生态系统的结构和功能产生显著影响.该文从以下几个方面综述了大气氮沉降增加对陆地生态系统的影响:1)氮输入增加可能影响植物生产力和生态系统碳蓄积能力,生态系统响应的方向和程度取决于系统的初始氮状况(氮限制或氮饱和)以及当地的植被和土壤特征;2)持续氮输入有可能改变土壤氮循环过程,降低土壤固持氮的能力,甚至导致土壤酸化、盐基离子损耗,进而影响到土壤有机碳的分解;3)高的氮沉降速率和持续氮输入都可能加速含氮痕量气体的释放,但其影响程度受生态系统初始状态的影响(例如磷限制和氮限制);4)氮沉降增加会影响生态系统的物种丰富度、植物群落结构和动态,促进森林扩张,改变菌根真菌的物种多样性;5)持续氮输入带来的植物群落结构和植物生理特征的变化可能影响昆虫取食特性,进而通过食物链改变生态系统的营养结构;6)氮沉降增加对生态系统的影响并不是孤立存在的,它与CO2浓度升高和O3浓度变化有协同作用,但难以从其协同效应中区分出各自的影响.最后,该文总结了我国的氮沉降研究现状,并对今后的研究前景提出了展望. |
| [29] | Development of direct and indirect methods for measuring root turnover and the status of knowledge on fine root turnover in forest ecosystems are discussed. While soil and ingrowth cores give estimates of standing root biomass and relative growth, respectively, minirhizotrons provide estimates of median root longevity (turnover time) i.e., the time by which 50% of the roots are dead. Advanced minirhizotron and carbon tracer studies combined with demographic statistical methods and new models hold the promise of improving our fundamental understanding of the factors controlling root turnover. Using minirhizotron data, fine root turnover (y-1) can be estimated in two ways: as the ratio of annual root length production to average live root length observed and as the inverse of median root longevity. Fine root production and mortality can be estimated by combining data from minirhizotrons and soil cores, provided that these data are based on roots of the same diameter class (e.g., <1 mm in diameter) and changes in the same time steps. Fluxes of carbon and nutrients via fine root mortality can then be estimated by multiplying the amount of carbon and nutrients in fine root biomass by fine root turnover. It is suggested that the minirhizotron method is suitable for estimating median fine root longevity. In comparison to the minirhizotron method, the radio carbon technique favor larger fine roots that are less dynamics. We need to reconcile and improve both methods to develop a more complete understanding of root turnover. |
| [30] | |
| [31] | Nitrogen (N) is one of the key mineral nutrients for plants and its availability has a major impact on their growth and development. Most often N resources are limiting and plants have evolved various |
| [32] | |
| [33] | Abstract Variability in short root morphology of the three main tree species of Europe's boreal forest (Norway spruce (Picea abies L. Karst.), Scots pine (Pinus sylvestris L.) and silver birch (Betula pendula Roth)) was investigated in four stands along a latitudinal gradient from northern Finland to southern Estonia. Silver birch and Scots pine were present in three stands and Norway spruce was present in all stands. For three fertile Norway spruce stands, fine root biomass and number of root tips per stand area or unit basal area were assessed from north to south. Principal component analysis indicated that short root morphology was significantly affected by tree species and site, which together explained 34.7% of the total variability. The range of variation in mean specific root area (SRA) was 51-74, 60-70 and 84-124 m(2) kg(-1) for Norway spruce, Scots pine and silver birch, respectively, and the corresponding ranges for specific root length were 37-47, 40-48 and 87-97 m g(-1). The range of variation in root tissue density of Norway spruce, Scots pine and silver birch was 113-182, 127-158 and 81-156 kg m(-3), respectively. Sensitivity of short root morphology to site conditions decreased in the order: Norway spruce > silver birch > Scots pine. Short root SRA increased with site fertility in all species. In Norway spruce, fine root biomass and number of root tips per m(2) decreased from north to south. The differences in morphological parameters among sites were significant but smaller than the site differences in fine root biomass and number of root tips. |
| [34] | Abstract Contents Summary 30 I. Allocation in perspective 31 II. Topics of this review 32 III. Methodology 32 IV. Environmental effects 33 V. Ontogeny 36 VI. Differences between species 40 VII. Physiology and molecular regulation 41 VIII. Ecological aspects 42 IX. Perspectives 45 Acknowledgements 45 References 45 Appendices A1 揂4 49 Summary We quantified the biomass allocation patterns to leaves, stems and roots in vegetative plants, and how this is influenced by the growth environment, plant size, evolutionary history and competition. Dose 搑esponse curves of allocation were constructed by means of a meta-analysis from a wide array of experimental data. They show that the fraction of whole-plant mass represented by leaves (LMF) increases most strongly with nutrients and decreases most strongly with light. Correction for size-induced allocation patterns diminishes the LMF-response to light, but makes the effect of temperature on LMF more apparent. There is a clear phylogenetic effect on allocation, as eudicots invest relatively more than monocots in leaves, as do gymnosperms compared with woody angiosperms. Plants grown at high densities show a clear increase in the stem fraction. However, in most comparisons across species groups or environmental factors, the variation in LMF is smaller than the variation in one of the other components of the growth analysis equation: the leaf area:leaf mass ratio (SLA). In competitive situations, the stem mass fraction increases to a smaller extent than the specific stem length (stem length:stem mass). Thus, we conclude that plants generally are less able to adjust allocation than to alter organ morphology. |
| [35] | . |
| [36] | . 使用酚二磺酸法、还原蒸馏法、镀铜镉还原-重氮化偶合比色法和改进紫外分光光度法(校正因数法)测定了中国9种不同类型土壤的硝态氮含量,分析了改进紫外分光光度法与其余三种测定方法的差异及其适用性。统计分析表明对于有机质含量低于50 g kg-1的矿质土壤,可以使用2.2作为校正因数,四种分析方法的测定值具有极显著的相关性,尤其是紫外分光光度法与酚二磺酸法的测定结果最为接近,没有极显著差异;对于有机质含量接近和高于50 g kg-1的土壤,校正因数还需要修改。紫外分光光度法具有操作简单、测定速度快等优点,适用于批量快速测定。 |
| [37] | Nitrogen (N) deposition-induced soil acidification has become a global problem. However, the response patterns of soil acidification toNaddition and the underlyingmechanisms remain far from clear. Here, we conducted ameta-analysis of 106 studies to reveal global patterns of soil acidification in responses toNaddition.We found thatNaddition significantly reduced soil pHby 0.26 on average globally. However, the responses of soil pH varied with ecosystem types,Naddition rate,Nfertilization forms, and experimental durations. Soil pH decreasedmost in grassland, whereas boreal forest was not observed a decrease toNaddition in soil acidification. Soil pH decreased linearly withNaddition rates. Addition of urea and NH4NO3 contributedmore to soil acidification thanNH4-formfertilizer. When experimental duration was longer than 20 years,Naddition effects on soil acidification diminished. Environmental factors such as initial soil pH,soil carbon and nitrogen content, precipitation, and temperature all influenced the responses of soil pH. Base cations of Ca2+,Mg2+ and K+ were critical important in buffering againstN-induced soil acidification at the early stage. However,Naddition has shifted global soils into the Al3+ buffering phase. Overall, this study indicates that acidification in global soils is very sensitive to N deposition, which is greatlymodified by biotic and abiotic factors. Global soils are now at a buffering transition from base cations (Ca2+,Mg2+ and K+) to non-base cations (Mn2+ and Al3+). This calls our attention to care about the limitation of base cations and the toxic impact of nonbase cations for terrestrial ecosystems withNdeposition. |
| [38] | 61 When using minirhizotrons to study fine dynamics in natural ecosystems, it is important to determine how sample collection frequency influences estimates of fine root production and mortality. Minirhizotron images were collected twice per week from mature Pseudotsuga menziesii and Tilia cordata trees and analyzed to estimate fine root production and mortality. These data were used to create data sets reflecting sample frequencies of 1, 2, 4 or 8 wk. 61 When the sampling interval is long, fine roots can appear and disappear between samplings, leading to underestimates of production and mortality. For example, with an 8-wk sample frequency, 24 and 35% of the fine root production in P. menziesii and T. cordata, respectively, is not measured. Fine root mortality displays the same sensitivity to sample frequency. 61 Our experimental observations supported the previously published simulation analysis, which provides an estimate of the proportion of fine roots missed at different sample frequencies and is a tool that can be used to select a sample frequency to balance production and mortality accuracy with sampling and analytical effort. |
| [39] | Knowledge of the responses of soil nitrogen (N) availability, fine root mass, production and turnover rates to atmospheric N deposition is crucial for understanding fine root dynamics and functioning in forest ecosystems. Fine root biomass and necromass, production and turnover rates, and soil nitrate-N and ammonium-N in relation to N fertilization (50 kg N ha(-1) year(-1)) were investigated in a temperate forest over the growing season of 2010, using sequential soil cores and ingrowth cores methods. N fertilization increased soil nitrate-N by 16% (P<0.001) and ammonium-N by 6% (P<0.01) compared to control plots. Fine root biomass and necromass in 0-20 cm soil were 13% (4.61 vs. 5.23 Mg ha(-1), P<0.001) and 34% (1.39 vs. 1.86 Mg ha(-1), P<0.001) less in N fertilization plots than those in control plots. The fine root mass was significantly negatively correlated with soil N availability and nitrate-N contents, especially in 0-10 cm soil layer. Both fine root production and turnover rates increased with N fertilization, indicating a rapid underground carbon cycling in environment with high nitrogen levels. Although high N supply has been widely recognized to promote aboveground growth rates, the present study suggests that high levels of nitrogen supply may reduce the pool size of the underground carbon. Hence, we conclude that high levels of atmospheric N deposition will stimulate the belowground carbon cycling, leading to changes in the carbon balance between aboveground and underground storage. The implications of the present study suggest that carbon model and prediction need to take the effects of nitrogen deposition on underground system into account. |
| [40] | Abstract I. II. III. IV. References SUMMARY: The search for a root economics spectrum (RES) has been sparked by recent interest in trait-based plant ecology. By analogy with the one-dimensional leaf economics spectrum (LES), fine-root traits are hypothesised to match leaf traits which are coordinated along one axis from resource acquisitive to conservative traits. However, our literature review and meta-level analysis reveal no consistent evidence of an RES mirroring an LES. Instead the RES appears to be multidimensional. We discuss three fundamental differences contributing to the discrepancy between these spectra. First, root traits are simultaneously constrained by various environmental drivers not necessarily related to resource uptake. Second, above- and belowground traits cannot be considered analogues, because they function differently and might not be related to resource uptake in a similar manner. Third, mycorrhizal interactions may offset selection for an RES. Understanding and explaining the belowground mechanisms and trade-offs that drive variation in root traits, resource acquisition and plant performance across species, thus requires a fundamentally different approach than applied aboveground. We therefore call for studies that can functionally incorporate the root traits involved in resource uptake, the complex soil environment and the various soil resource uptake mechanisms - particularly the mycorrhizal pathway - in a multidimensional root trait framework. 2016 Wageningen University. New Phytologist 2016 New Phytologist Trust. |
| [41] | . |
| [42] | Questions remain as to which soil nutrients limit primary production in tropical forests. Phosphorus (P) has long been considered the primary limiting element in lowland forests, but recent evidence demonstrates substantial heterogeneity in response to nutrient addition, highlighting a need to understand and diagnose nutrient limitation across diverse forests. Fine-root characteristics including their abundance, functional traits, and mycorrhizal symbionts can be highly responsive to changes in soil nutrients and may help to diagnose nutrient limitation. Here, we document the response of fine roots to long-term nitrogen (N), P, and potassium (K) fertilization in a lowland forest in Panama. Because this experiment has demonstrated that N and K together limit tree growth and P limits fine litter production, we hypothesized that fine roots would also respond to nutrient addition. Specifically we hypothesized that N, P, and K addition would reduce the biomass, diameter, tissue density, and mycorrhizal colonization of fine roots, and increase nutrient concentration in root tissue. Most morphological root traits responded to the single addition of K and the paired addition of N and P, with the greatest response to all three nutrients combined. The addition of N, P, and K together reduced fine-root biomass, length, and tissue density, and increased specific root length, whereas root diameter remained unchanged. Nitrogen addition did not alter root N concentration, but P and K addition increased root P and K concentration, respectively. Mycorrhizal colonization of fine roots declined with N, increased with P, and was unresponsive to K addition. Although plant species composition remains unchanged after 14 years of fertilization, fine-root characteristics responded to N, P, and K addition, providing some of the strongest stand-level responses in this experiment. Multiple soil nutrients regulate fine-root abundance, morphological and chemical traits, and their association with mycorrhizal fungi in a species-rich lowland tropical forest. |
| [43] | . <p>因大气污染而不断增加的大气氮沉降量,在许多地区超过了森林生态系统的氮需求。氮在土壤中的化学和生物化学反应对H<sup>+</sup> 离子的产生与消耗有重要影响。NH<sup>+</sup><sub>4</sub>和NO<sup>-</sup><sub>3</sub>输入与输出的平衡状态影响着土壤-土壤溶液系统的酸化速率。过剩的氮沉降将增加NH<sup>+</sup><sub>4</sub>的硝化和NO<sup>-</sup><sub>3</sub>的淋失,加速土壤的酸化。土壤酸化对森林有危害作用。</p> |
| [44] | Comment in Environ Pollut. 2013 Jun;177:203; discussion 204. Comment on Environ Pollut. 2011 Oct;159(10):2251-64. |
| [45] | . |
| [46] | . |
| [47] | . |
1
2016
... 根系生长于地下, 不易接近和直接观察, 对于根系的认识很大程度上受限于研究手段.传统挖掘法、土钻法、内生长法研究根系, 往往费时费力, 对根系破坏严重, 且不能连续观察, 很难实现对根寿命的准确测定(
Root production in contrasting ecosystems: The impact of rhizotron sampling frequency.
1
2016
... 根系生长于地下, 不易接近和直接观察, 对于根系的认识很大程度上受限于研究手段.传统挖掘法、土钻法、内生长法研究根系, 往往费时费力, 对根系破坏严重, 且不能连续观察, 很难实现对根寿命的准确测定(
1
... 土壤样品带回实验室后挑出土壤中的杂质, 过2 mm筛后冷藏保存, 测定土壤pH值、铵态氮(NH4+-N)含量、硝态氮(NO3--N)含量, 取部分鲜土风干后过0.25 mm筛用于测定土壤全氮(TN)含量.使用电位法(LY/T 1239-1999)测定土壤pH值, TN用半微量凯氏法(LY/T 1228-1999)测定, NO3--N用紫外分光光度法(
Going underground: Root traits as drivers of ecosystem processes.
2
2014
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
... 在本研究中, 氮添加使I层扁刺栲细根的根段长和根段表面积呈下降趋势, 其中低氮处理对根段表面积的影响达到显著(p = 0.049)水平.植物依靠根系获取自身需要的多种元素, 由于土壤各种养分具有较强的空间异质性.植物根系利用其强大的可塑性予以应对, 形成了“成本-效益”策略来适应各种土壤养分分布状况(
Is the growth of temperate forest trees enhanced along an ambient nitrogen deposition gradient?
1
2009
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests.
1
2000
... 但是, 氮添加试验(
Effect of nitrogen additions on root morphology and chemistry in a subtropical bamboo forest.
1
2017
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
Intrinsic and extrinsic controls of fine root life span.
3
2013
... 根系生长于地下, 不易接近和直接观察, 对于根系的认识很大程度上受限于研究手段.传统挖掘法、土钻法、内生长法研究根系, 往往费时费力, 对根系破坏严重, 且不能连续观察, 很难实现对根寿命的准确测定(
... 细根寿命由植物的本身特性包括根形态、分布深度、菌根共生、植物生命形态等和外在特性包括土壤氮和水分可利用性、温度等因素共同控制, 因此不同树种及同一树种生长在不同环境中均表现出不同的根寿命(
... 但是, 氮添加试验(
植物叶经济谱的研究进展
1
2014
... 但是, 氮添加试验(
Global forest resources assessment.
1
2013
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Building roots in a changing environment: Implications for root longevity.
2
2000
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
... 植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(
The ecology of root lifespan.
3
1997
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
... 植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(
... 在本研究中, 虽然低氮和高氮处理均使I层扁刺栲细根的中值寿命降低了24%, 但是未达到统计学显著水平.传统理论认为, 土壤氮可利用性增加, 通常提高植物地上部分的生长速率, 进而对养分元素的需求也相应增加.植物为了支持地上部分对养分的需求, 需要提高细根对土壤养分的吸收能力, 而新鲜的细根具有更强的养分获取能力, 因此植物会提高根系统中年轻细根的比例, 降低细根的寿命, 加快整个根系统的周转速率(
Rapid immobilisation and leaching of wet-deposited nitrate in upland organic soils.
1
2008
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
Reactive nitrogen and the world: 200 years of change.
1
2002
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Nitrogen cycles: Past, present, and future.
1
2004
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Nutrient relocation, hydrological functions, and soil chemistry in plantations as compared to natural forests in central Yunnan, China.
1
2010
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Research on acidfication in forest soil driven by atmospheric nitrogen deposition.
1
2014
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
中亚热带细柄阿丁枫和米槠群落细根的生产和死亡动态
1
2012
... 细根寿命由植物的本身特性包括根形态、分布深度、菌根共生、植物生命形态等和外在特性包括土壤氮和水分可利用性、温度等因素共同控制, 因此不同树种及同一树种生长在不同环境中均表现出不同的根寿命(
A global budget for fine root biomass, surface area, and nutrient contents.
1
1997
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
Effects of elevated CO2, N-fertilization, and season on survival of ponderosa pine fine roots.
1
2000
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Fine root production and turnover in
1
2008
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China.
1
2015
... 氮添加处理未影响3个土层中扁刺栲的细根生物量.目前, 氮添加对细根生物量的影响存在减少、增加、不影响3种情况(
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.
1
2008
... 植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(
我国天然林与人工林的比较研究
1
2011
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
应用微根管法测定细根指标方法评述
1
2007
... 根系生长于地下, 不易接近和直接观察, 对于根系的认识很大程度上受限于研究手段.传统挖掘法、土钻法、内生长法研究根系, 往往费时费力, 对根系破坏严重, 且不能连续观察, 很难实现对根寿命的准确测定(
The effects of simulated nitrogen deposition on plant root traits: A meta-analysis.
1
2015
... 植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(
A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams.
1
2011
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
陆地生态系统氮沉降增加的生态效应
1
2007
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
Measuring fine root turnover in forest ecosystems.
1
2005
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
Responses of fine root mass, length, production and turnover to soil nitrogen fertilization in
2
2010
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
... 但是, 氮添加试验(
Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource.
1
2013
... 植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(
Biomass and morphology of fine roots of sugi (
1
2013
... 氮添加处理未影响3个土层中扁刺栲的细根生物量.目前, 氮添加对细根生物量的影响存在减少、增加、不影响3种情况(
Fine root morphological adaptations in scots pine, Norway spruce and silver birch along a latitudinal gradient in boreal forests.
1
2007
... 植物主要通过根系获取维持自身生长发育的多种元素, 其中, 氮元素对植物的意义最为重大(
Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control.
1
2011
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
微根管在细根研究中的应用
1
2006
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
测定土壤硝态氮的紫外分光光度法与其他方法的比较
1
2007
... 土壤样品带回实验室后挑出土壤中的杂质, 过2 mm筛后冷藏保存, 测定土壤pH值、铵态氮(NH4+-N)含量、硝态氮(NO3--N)含量, 取部分鲜土风干后过0.25 mm筛用于测定土壤全氮(TN)含量.使用电位法(LY/T 1239-1999)测定土壤pH值, TN用半微量凯氏法(LY/T 1228-1999)测定, NO3--N用紫外分光光度法(
A global analysis of soil acidification caused by nitrogen addition.
1
2015
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
Optimizing minirhizotron sample frequency for an evergreen and deciduous tree species.
1
2003
... 2013年12月初, 于各样地内选择3株胸径大致相同(25 cm左右)、生长较好、相对独立的扁刺栲作为细根动态观测的目标树.在树干周围1/2冠幅的位置(到树干基部的距离约1 m)对称地安装两根由有机玻璃制成的微根管(
Responses of fine roots and soil N availability to short-term nitrogen fertilization in a broad-leaved Korean pine mixed forest in northeastern China.
2
2012
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
... 氮添加处理未影响3个土层中扁刺栲的细根生物量.目前, 氮添加对细根生物量的影响存在减少、增加、不影响3种情况(
Towards a multidimensional root trait framework: A tree root review.
1
2016
... 但是, 氮添加试验(
高寒草甸植被细根生产和周转的比较研究
1
2014
...
Fine root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest.
1
2015
... 细根占陆地生态系统植物总生物量的比例小于2%, 但由于细根周转快速, 消耗了20%-30%的年净初级生产力(
大气氮沉降对森林土壤酸化的影响
1
2001
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
Implications of greater than average increases in nitrogen deposition on the western edge of the Szechwan basin, China.
1
2013
... 氮添加处理1.5年后, HN处理显著降低了表层土壤pH值, 土壤硝态氮含量表现出随氮添加梯度增加而增加的趋势.土壤pH值与土壤NH4+-N、NO3--N含量呈显著负相关关系, 这与众多氮添加试验中的发现一致(
氮肥对水曲柳和落叶松细根寿命的影响
2
2009
... 近几十年来, 由于工农业发展和化石燃料的大量燃烧导致氮沉降在全球范围增加, 人类活动极大地干扰了全球氮循环(
... ).现有的少量氮添加对森林植物根系影响的相关研究往往在人工林中开展(
杉木人工林细根寿命研究
1
2009
... 细根寿命由植物的本身特性包括根形态、分布深度、菌根共生、植物生命形态等和外在特性包括土壤氮和水分可利用性、温度等因素共同控制, 因此不同树种及同一树种生长在不同环境中均表现出不同的根寿命(
闽楠人工林细根寿命及其影响因素
1
2012
... 细根寿命由植物的本身特性包括根形态、分布深度、菌根共生、植物生命形态等和外在特性包括土壤氮和水分可利用性、温度等因素共同控制, 因此不同树种及同一树种生长在不同环境中均表现出不同的根寿命(


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}