关键词:多样性海拔格局;谱系alpha多样性;谱系beta多样性;谱系结构主坐标分析;草本植物 Abstract Aims Incorporating phylogenetic data in the studies of species diversity patterns along elevational gradients can bridge the gap between ecological and evolutionary processes, and thus shed light on the issues related to community assemblage. We aim is to explore the elevational patterns of phylogenetic relatedness and phylodiversity in the herbaceous angiosperm assemblages of alpine conifer forest within mountain ecosystem, and to quantify the relationship between the patterns and habitat factors. Methods We sampled 17 plots (20 m × 30 m) in the coniferous forest communities at ca. 50 m altitudinal intervals along the elevation gradient. In each plot, we documented all species encountered and the environmental conditions. We used the Net Relatedness Index (NRI) and Nearest Taxon Index (NTI) to quantify the phylogenetic structures of each herbaceous assemblage, and used the Mean Pairwise Distance and Mean Nearest Taxon Distance to quantify phylogenetic beta diversity. Ordinary least square regression and multiple regression on distance matrices were employed respectively to explore the elevational trends of phylogenetic structure and phylobetadiversity. We analyzed the taxonomic composition of the herbaceous assemblages within forests, and demonstrated the relationship between the clustering of clades and the significant habitat descriptors using principal coordinates of phylogenetic structure (PCPS). Important findings The result showed that the herbaceous communities tended to be more phylogenetically overdispersed at lower elevations, suggesting that intraspecific competition potentially influences the local assemblages. In contrast, species occurring at high-elevation sites tended to be more closely related, implying that these communities are structured primarily by environmental filtering. However, we found that all of the NRI (or NTI) were confined within 95% confidence intervals, suggesting strong contributions of stochastic processes on species assembly. Phylogenetic beta diversity significantly increased with the elevational distance between community pairs, also suggesting that habitat filtering probably played an important role on structuring the herbaceous communities. The first two axes of PCPS contained 55.9% of total variation in phylogeny-weighted species composition, and were both significantly related with elevation and arboreal basal area. We found that species of large families, including Asteraceae and Poaceae, were phylogenetically clustered at high elevations, whereas the others were phylogenetically overdispersed at low elevation region. Our findings suggest that exploring the patterns of phylogenetic structures across elevational gradients is important, which can provide insights into the underlying mechanisms shaping community composition within montane ecosystems.
Keywords:diversity elevational pattern;phylogenetic alpha diversity;phylogenetic beta diversity;principal coordinates of phylogenetic structure;herbaceous plants -->0 PDF (1150KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 赵鸣飞, 薛峰, 王宇航, 王国义, 邢开雄, 康慕谊, 王菁兰. 山西芦芽山针叶林草本层群落谱系结构与多样性的海拔格局. 植物生态学报, 2017, 41(7): 707-715 https://doi.org/10.17521/cjpe.2016.0247 ZHAOMing-Fei, XUEFeng, WANGYu-Hang, WANGGuo-Yi, XINGKai-Xiong, KANGMu-Yi, WANGJing-Lan. Phylogenetic structure and diversity of herbaceous communities in the conifer forests along an elevational gradient in Luya Mountain, Shanxi, China. Chinese Journal of Plant Ecology, 2017, 41(7): 707-715 https://doi.org/10.17521/cjpe.2016.0247 探求群落物种组成以及群落间物种周转格局的形成原因一直以来是生态学的核心问题(Ricklefs, 2004), 学界对关键机制的认识目前尚存争议(Bryant et al., 2008; Tello et al., 2015)。目前, 生态学家已普遍达成共识, 认为物种多样性是生态位过程与中性因素等诸多生态过程共同作用的结果(Oommen & Shanker, 2005; McCain, 2009)。前者包括非生物因子(如气候条件或土壤营养状况等)和生物因子(如竞争、捕食、共生等种间相互作用)等确定性因素; 后者包括生态漂变和扩散限制等随机因素(Gilbert & Lechowicz, 2004; Soininen et al., 2007)。事实上, 基于宏观尺度, 与物种形成、灭绝以及长距离扩散有关的进化过程则是直接决定群落多样性水平的背景性动因(Wiens & Donoghue, 2004; Wiens et al., 2007)。这一点正逐渐受到越来越多生态学工作者的重视(Bryant et al., 2008; Swenson et al., 2011)。 人们很早就发现, 物种的功能性状在演化上普遍具有内在关联(Elton, 1946)。具体来说, 由于物种对不同生境状况所做出的响应主要通过其功能性状来实现, 而演化的生态位保守性(niche conservatism)使近缘物种保持来自共同祖先的功能属性(Vamosi et al., 2009)。因此, 群落成员之间的谱系关系能够体现它们生态位的相似性。例如, 由生境过滤作用所主导的群落构建过程, 有利于筛选出具有耐受特定环境生理性状的物种(这些物种往往来自同一分支), 从而在谱系结构上表现出聚集(phylogenetically clustering)。相反, 如果群落成员间存在强烈的竞争排斥作用, 则有利于具有不同资源利用方式的物种稳定共存。这些物种之间的亲缘关系一般较为疏远, 从而使谱系结构呈现发散(phylogenetically over-dispersion) (Webb et al., 2002; Losos, 2008)。另外, 在生态位保守假设成立的情况下, 若谱系结构并未表现出统计意义上的显著聚集或发散, 则可能意味着物种集聚主要被中性过程所驱动(Kembel & Hubbell, 2006)。由此可见, 整合有谱系信息的多样性研究可为探究物种共存机制等核心生态学问题提供更为深刻的洞见(Webb et al., 2002; Cavender- Bares et al., 2009)。 山地生态系统通常在有限的空间内具备宽广的水热梯度和较高的生境异质性(Gaston, 2000; Körner, 2007; Sheldon et al., 2011)。这些特点一方面为新物种形成创造了有利条件, 使山地成为多样性的热点地域; 另一方面, 由于该系统对全球气候变化十分敏感, 从而增加了物种的灭绝风险。因此, 物种多样性沿海拔梯度的变化情况常被视作大尺度纬向格局的镜像, 长久以来为人们所关注(Stevens, 1992; Lomolino, 2001; Rahbek, 2005; Tang et al., 2012a)。然而, 有关群落谱系结构及其多样性沿海拔梯度变化规律的研究目前还处于起步阶段(Machac et al., 2011), 尚未形成统一的认识。现有经验案例显示, 研究结果会随着山地和生物类群的不同而存在显著差异。一般来说, 高海拔低温可对多数物种构成主要的生理胁迫, 因而群落谱系结构理论上应趋向聚集。这一预测已在蚂蚁(Machac et al., 2011)、蜂鸟(Graham & Mcguire, 2009)、蕨类植物(Kluge & Kessler, 2011)以及高山被子植物(Li et al., 2014)等多个类群得到验证。而Bryant等(2008)对美国落基山脉植物群落和土壤微生物群落谱系结构的海拔格局进行对比研究发现: 与理论预期相反, 该山地被子植物在低海拔地区谱系聚集, 而在高海拔地区谱系发散。卢孟孟等(2014)通过探索哀牢山森林群落谱系结构在海拔梯度上的变化, 发现采用不同指标得到的结论并不一致。 草本层是森林生态系统的关键组分, 对于温带森林植被的多样性尤为重要, 其物种数往往占整个群落物种丰富度的比例最大(Gilliam, 2007)。另外, 草本层在调控乔木物种更新、促进养分循环、增加生境异质性等诸多生态过程中发挥着重要作用(Gilliam, 2007; Cook, 2015; Márialigeti et al., 2016)。地处山西西北部的芦芽山具有较大相对海拔高差, 并存在大面积完好、典型的寒温性针叶林。这些森林属于演替后期相对稳定的顶级群落, 建群种组成简单, 灌木层因林冠郁闭度较高而普遍不发达, 因此为其下的草本层物种创造了相对同质的生境条件(张金屯, 1989)。本文以该地区针叶林草本层为研究对象, 基于厘定有进化时间的谱系树, 结合物种组成与生境因子数据, 通过运用多种统计分析方法, 尝试回答以下科学问题: (1)冠层下草本层物种集聚的谱系结构沿海拔梯度呈怎样的格局? (2)谱系beta多样性与海拔差异的相互关系及解释。(3)群落物种组成、谱系关系与环境因子之间的定量关系如何?
整体而言, NRI反映了芦芽山林下草本群落谱系结构随着海拔梯度的上升呈现由发散到聚集的变化趋势, 且回归系数达到统计显著水平(R2 = 0.447, p = 0.002, 图1A); 但NTI与海拔并无显著线性关系(R2 = 0.001, p = 0.702, 图2B)。具体来讲, 每个群落的谱系结构均处于随机置信区间内(NRI或NTI绝对值均小于1.96, 图1)。17个样方中有4个NRI大于0, 且均为高海拔地区群落; 而对于NTI, 虽然多数群落(76.5%)取值大于0, 但并无明显的海拔分布趋势。 显示原图|下载原图ZIP|生成PPT 图1芦芽山针叶林草本群落谱系结构的海拔格局。 -->Fig. 1Elevational patterns of the phylogenetic structures of herbaceous assemblages in the coniferous forest plots in Luya Mountain. -->
回归分析发现, 代表群落间谱系近缘关系的betaMNTD与海拔差异存在极显著的正相关关系(R2 = 0.691, p < 0.001, 图2B)。而群落间平均谱系关系的betaMPD与样地海拔差异之间并不存在显著线性关系(R2 = 0.001, p = 0.668, 图2A)。 显示原图|下载原图ZIP|生成PPT 图2芦芽山针叶林草本层谱系beta多样性与海拔差异之间的关系。 -->Fig. 2The relationships of phylogenetic beta diversity with elevational distance for the herbaceous plants assemblages in the coniferous forest plots in Luya Mountain. -->
PCPS模型全局F检验达到显著水平(F-pseudo = 1.173, p = 0.014, 999次置换)。前两轴(特征值分别为0.301和0.090)累积解释了55.9%的总变异。PCPS二维排序图显示, 环境因子中只有海拔、胸高断面积和对草本层物种集聚具有显著影响(图3; 表1), 而土壤、地形等其他局地因子的作用均不显著。由图3看出, 排序前两轴代表了全局种库谱系关系中演化早期分支的变异情况(例如单子叶分支和双子叶分支, 图1)。群落中物种丰富度大的科级分支(物种数大于5)多与海拔或胸高断面积和密切相关, 如菊科和禾本科存在高海拔聚集趋势, 而蔷薇科、石竹科、毛茛科等物种数少的科更多分布于中、低海拔地区。 显示原图|下载原图ZIP|生成PPT 图3草本群落物种分布、谱系结构与环境因子之间相互关系的谱系结构主坐标分析(PCPS)排序图。●, 草本植物种, 包含5种以上的科以彩色显示; Ele, 海拔; BA, 胸高断面积和。 -->Fig. 3Scatter diagram between the first two axes of the principal coordinates of phylogenetic structure (PCPS) for herbaceous plants occurring in the coniferous forest plots in Luya Mountain. Color points represent large families (>5 species) grouped in monocots and dicotyledon clades. Ele, elevation; BA, total basal area of breast height. -->
Table 1 表1 表1环境因子与谱系结构主坐标分析(PCPS)前两排序轴的关系 Table 1Correlations between the first two constrained ordination axes (PCPS 1 and PCPS 2) and environmental factors
环境因子 Environmental variables
第1排序轴 PCPS 1
第2排序轴 PCPS 2
R2
p
海拔 Elevation
-0.495
0.869
0.653 8
0.001
坡向 Aspect
-0.620
0.784
0.203 9
0.224
坡度 Slope
0.305
-0.952
0.290 0
0.109
冠层高 Delta height
0.296
-0.955
0.145 6
0.349
胸高断面积和 Total basal area of breast height
-0.701
0.713
0.359 0
0.049
立木密度 Stem density
0.300
0.954
0.156 9
0.325
土壤深度 Soil depth
-0.436
-0.900
0.175 0
0.289
凋落物厚度 Litter thickness
-0.762
0.648
0.010 6
0.928
土壤有机碳 Soil organic carbon
-0.914
-0.406
0.183 4
0.267
土壤总氮 Soil total nitrogen
-0.384
-0.923
0.134 8
0.389
土壤总磷 Soil total phosphorus
0.165
0.986
0.045 5
0.733
Statistical significance is based on permutation test (999 times).统计显著性基于999次置换检验。 新窗口打开
综上所述, 本研究揭示了芦芽山针叶林冠层下草本群落谱系结构随海拔升高而呈现由发散到聚集的趋势; 海拔差异是驱动群落间谱系beta多样性递减格局的主因; 通过PCPS方法综合展示了群落谱系分支构成及其与主要生境因子之间的相互关系。上述结果表明研究区草本群落构建过程可能是生境过滤、竞争排斥以及随机因素等生态因素共同作用的结果。将谱系理念引入群落生态学研究, 可为全球气候变化背景下的生物多样性保育与维持工作带来全新的理论视角和重要的应用价值。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
APGIII (2009). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society, 161, 105-121.
[2]
BryantJA, LamannaC, MorlonH, KerkhoffAJ, EnquistBJ, GreenJL (2008). Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proceedings of the National Academy of Sciences of the United States of American, 105, 11505-11511. [本文引用: 3]
[3]
Cavender-BaresJ, KozakKH, FinePV, KembelSW (2009). The merging of community ecology and phylogenetic biology. Ecology Letters, 12, 693-715. [本文引用: 1]
[4]
CookJE (2015). Structural effects on understory attributes in second-growth forests of northern Wisconsin, USA. Forest Ecology and Management, 347, 188-199. [本文引用: 1]
[5]
DonoghueMJ (2008). Phylogenetic perspective on the distribution of plant diversity. Proceedings of the National Academy of Sciences of the United States of American, 105, 11549-11555. [本文引用: 1]
EltonC (1946). Competition and the structure of ecological communities. Journal of Animal Ecology, 15, 54-68. [本文引用: 1]
[8]
FinePVA, KembelSW (2011). Phylogenetic community structure and phylogenetic turnover across space and edaphic gradients in western Amazonian tree communities. Ecography, 34, 552-565. [本文引用: 1]
[9]
GastonKJ (2000). Global patterns in biodiversity. Nature, 405, 220-227. [本文引用: 1]
[10]
GilbertB, LechowiczMJ (2004). Neutrality, niches, and dispersal in a temperate forest understory. Proceedings of the National Academy of Sciences of the United States of America, 101, 7651-7656. [本文引用: 1]
[11]
GilliamFS (2007). The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience, 57, 845-858. [本文引用: 2]
[12]
GrahamCH, FinePV (2008). Phylogenetic beta diversity: Linking ecological and evolutionary processes across space in time. Ecology Letters, 11, 1265-1277. [本文引用: 1]
[13]
GrahamCH, McguireJA (2009). Phylogenetic structure in tropical hummingbird communities. Proceedings of the National Academy of Sciences of the United States of American, 106, 19673-19678. [本文引用: 1]
[14]
HardyOJ, SenterreB (2007). Characterizing the phylogenetic structure of communities by an additive partitioning of phylogenetic diversity. Journal of Ecology, 95, 493-506. [本文引用: 1]
[15]
HawkinsBA, RuedaM, RangelTF, FieldR, Diniz-FilhoJAF (2014). Community phylogenetics at the biogeographical scale: Cold tolerance, niche conservatism and the structure of North American forests. Journal of Biogeography, 41, 23-38. [本文引用: 1]
[16]
HelmusM, SavageK, DiebelMJ, IvesA (2007). Separating the determinants of phylogenetic community structure. Ecology Letters, 10, 917-925. [本文引用: 1]
[17]
KembelSW, HubbellSP (2006). The phylogenetic structure of a neotropical forest tree community. Ecology, 87, 86-99. [本文引用: 2]
[18]
KlugeJ, KesslerM (2011). Phylogenetic diversity, trait diversity and niches: Species assembly of ferns along a tropical elevational gradient. Journal of Biogeography, 38, 394-405. [本文引用: 1]
[19]
KörnerC (2007). The use of “altitude” in ecological research. Trends in Ecology & Evolution, 22, 569-574. [本文引用: 1]
[20]
LathamRE, RicklefsRE (1993). Global patterns of tree species richness in moist forests: Energy-diversity theory does not account for variation in species richness. Oikos, 67, 325-333. [本文引用: 1]
[21]
LiXH, ZhuXX, NiuY, SunH (2014). Phylogenetic clustering and overdispersion for alpine plants along elevational gradient in the Hengduan Mountains Region, southwest China. Journal of Systematics and Evolution, 52, 280-288. [本文引用: 2]
[22]
LiYJ, WangSY, NiuJJ, FangKY, LiXL, LiY, BuWL, LiYH (2016). Climate-adial growth relationship of Larix principis-rupprechtii at different altitudes on Luya Mountain. Acta Ecologica Sinica, 36, 1608-1618. (in Chinese with English abstract)[李颖俊, 王尚义, 牛俊杰, 方克艳, 李晓岚, 栗燕, 布文丽, 李玉晗 (2016). 芦芽山华北落叶松(Larix principis-rupprechtii)树轮宽度年表对气候因子的响应 . 生态学报, 36, 1608-1618.] [本文引用: 1]
[23]
LichsteinJW (2007). Multiple regression on distance matrices: A multivariate spatial analysis tool. Plant Ecology, 188, 117-131. [本文引用: 1]
[24]
LomolinoMV (2001). Elevation gradients of species-density: Historical and prospective views. Global Ecology Biogeography, 10, 3-13. [本文引用: 1]
[25]
LososJB (2008). Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecology Letters, 11, 995-1003. [本文引用: 1]
[26]
LozuponeCA, HamadyM, KelleyST, RobK (2007). Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Applied & Environmental Microbiology, 73, 1576-1585. [本文引用: 1]
[27]
LuMM, HuangXC, CiXQ, YangGP, LiJ (2014). Phylogenetic community structure of subtropical forests along elevational gradients in Ailao Mountains of southwest China. Biodiversity Science, 22, 438-448. (in Chinese with English abstract)[卢孟孟, 黄小翠, 慈秀芹, 杨国平, 李捷 (2014). 沿海拔梯度变化的哀牢山亚热带森林群落谱系结构 . 生物多样性, 22, 438-448.]
[28]
MachacA, JandaM, DunnRR, SandersNJ (2011). Elevational gradients in phylogenetic structure of ant communities reveal the interplay of biotic and abiotic constraints on diversity. Ecography, 34, 364-371. [本文引用: 4]
[29]
MárialigetiS, TinyaF, BidlóA, ÓdorP (2016). Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in Hungary. Plant Ecology, 217, 549-563. [本文引用: 1]
[30]
McCainCM (2009). Global analysis of bird elevational diversity. Global Ecology and Biogeography, 18, 346-360. [本文引用: 1]
[31]
OommenMA, ShankerK (2005). Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants. Ecology, 86, 3039-3047. [本文引用: 1]
[32]
PillarVD, DuarteLDS (2010). A framework for metacommunity analysis of phylogenetic structure. Ecology Letters, 13, 587-596. [本文引用: 1]
[33]
QianH, HaoZ, ZhangJ (2014). Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, China. Journal of Plant Ecology, 7, 154-165. [本文引用: 1]
[34]
RahbekC (2005). The role of spatial scale and the perception of large-scale species-richness patterns. Ecology Letters, 8, 224-239. [本文引用: 1]
[35]
RicklefsRE (2004). A comprehensive framework for global patterns in biodiversity. Ecology Letters, 7, 1-15. [本文引用: 1]
[36]
RobertsDW, CooperSV (1989). Concepts and techniques of vegetation mapping . In: Ferguson D, Morgan P, Johnson FD eds. Land Classifications Based on Vegetation: Applications for Resource Management, General Technical Report INF-257. Department of Agriculture, Forest Service, Intermountain Research Station, Odgen, USA. 90-96. [本文引用: 1]
[37]
SheldonKS, YangS, TewksburyJJ (2011). Climate change and community disassembly: Impacts of warming on tropical and temperate montane community structure. Ecology Letters, 14, 1191-1200. [本文引用: 1]
[38]
SoininenJ, McDonaldR, HillebrandH (2007). The distance decay of similarity in ecological communities. Ecography, 30, 3-12. [本文引用: 1]
[39]
StevensGC (1992). The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. The American Naturalist, 140, 893-911. [本文引用: 1]
[40]
SwensonNG (2011). Phylogenetic beta diversity metrics, trait evolution and inferring the functional beta diversity of communities. PLOS ONE, 6, e21264. doi: 10.1371/journal. pone.0021264. [本文引用: 2]
[41]
SwensonNG, Anglada-CorderoP, BaroneJA (2011). Deterministic tropical tree community turnover: Evidence from patterns of functional beta diversity along an elevational gradient. Proceedings of the Royal Society of London B: Biological Sciences, 278, 877-884.
[42]
TangZY, FangJY, ChiXL, FengJM, LiuYN, ShenZH,WangXP, WangZH, WuXP, ZhengCY (2012a). Patterns of plant beta-diversity along elevational and latitudinal gradients in mountain forests of China. Ecography, 35, 1083-1091. [本文引用: 1]
[43]
TangZY, FangJY, ChiXL, YangYY, MaWH, MohhamotA, GuoZD, LiuYN, GastonKJ (2012b). Geography, environment, and spatial turnover of species in China’s grasslands. Ecography, 35, 1103-1109. [本文引用: 1]
[44]
TelloJS, MyersJA, MacíaMJ, FuentesAF, CayolaL, ArellanoG, LozaMI, TorrezV, CornejoM, MirandaTB (2015). Elevational gradients in β-diversity reflect variation in the strength of local community assembly mechanisms across spatial scales. PLOS ONE, 10, e0121458. doi: 10.1371/ journal.pone.0121458. [本文引用: 1]
[45]
VamosiSM, HeardSB, VamosiJC, WebbCO (2009). Emerging patterns in the comparative analysis of phylogenetic community structure. Molecular Ecology, 18, 572-592. [本文引用: 2]
[46]
WebbCO (2000). Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. The American Naturalist, 156, 145-155. [本文引用: 1]
[47]
WebbCO, AckerlyDD, KembelSW (2008). Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics, 24, 2098-2100. [本文引用: 1]
[48]
WebbCO, AckerlyDD, McPeekMA, DonoghueMJ (2002). Phylogenies and community ecology. Annual Review of Ecology and Systematics, 33, 475-505. [本文引用: 4]
[49]
WebbCO, DonoghueMJ (2005). Phylomatic: Tree assembly for applied phylogenetics. Molecular Ecology Notes, 5, 181-183. [本文引用: 1]
[50]
WiensJJ, DonoghueMJ (2004). Historical biogeography, ecology and species richness. Trends in Ecology & Evolution, 19, 639-644. [本文引用: 3]
[51]
WiensJJ, Parra-OleaG, García-ParísM, WakeDB (2007). Phylogenetic history underlies elevational biodiversity patterns in tropical salamanders. Proceedings of the Royal Society of London B: Biological Sciences, 274, 919-928. [本文引用: 1]
[52]
ZanneAE, TankDC, CornwellWK, EastmanJM, SmithSA, FitzJohnRG, McGlinnDJ, O’MearaBC, MolesAT, ReichPB (2014). Three keys to the radiation of angiosperms into freezing environments. Nature, 506, 89-92. [本文引用: 1]
[53]
ZhangJT (1989). Vertical zone of vegetation in Luya Mountain in Shanxi Province. Scientia Geographica Sinica, 9, 346-353. (in Chinese with English abstract)[张金屯 (1989). 山西芦芽山植被垂直带的划分 . 地理科学, 9, 346-353.] [本文引用: 2]
[54]
ZhangWT, JiangY, WangMC, ZhangLN, DongMY (2015). Responses of radial growth in Larix principis-rupprechtii to climate change along an elevation gradient on the southern slope of Luya Mountain. Acta Ecologica Sinica, 35, 1-10. (in Chinese with English abstract)[张文涛, 江源, 王明昌, 张凌楠, 董满宇 (2015). 芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应 . 生态学报, 35, 1-10.] [本文引用: 1]
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. 2009
Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. 3 2008
... 探求群落物种组成以及群落间物种周转格局的形成原因一直以来是生态学的核心问题(Ricklefs, 2004), 学界对关键机制的认识目前尚存争议(Bryant et al., 2008; Tello et al., 2015).目前, 生态学家已普遍达成共识, 认为物种多样性是生态位过程与中性因素等诸多生态过程共同作用的结果(Oommen & Shanker, 2005; McCain, 2009).前者包括非生物因子(如气候条件或土壤营养状况等)和生物因子(如竞争、捕食、共生等种间相互作用)等确定性因素; 后者包括生态漂变和扩散限制等随机因素(Gilbert & Lechowicz, 2004; Soininen et al., 2007).事实上, 基于宏观尺度, 与物种形成、灭绝以及长距离扩散有关的进化过程则是直接决定群落多样性水平的背景性动因(Wiens & Donoghue, 2004; Wiens et al., 2007).这一点正逐渐受到越来越多生态学工作者的重视(Bryant et al., 2008; Swenson et al., 2011). ...
... 虽然具体样地并未发现统计显著的谱系结构, 但NRI随着海拔的升高存在由发散向聚集转变的趋势(图1A), 这与多数相似研究案例的结果相一致(Machac et al., 2011; Li et al., 2014; Qian et al., 2014).此意味着, 除了中性过程以外, 种间竞争、生境过滤等生态位过程可能对草本群落的物种聚集也起到关键作用.生态位理论认为, 生境过滤与竞争排斥是促成物种共存且作用相反的两个主要过程.前者推动群落谱系结构趋向于聚集, 后者则利于谱系发散.对于山地生态系统来说, 一般低海拔地区具有相对良好的水热条件(也即生境过滤作用相对较弱), 有利于更多物种共存.亲缘关系越相近的物种, 其演化保守的关键性状也越相似(Wiens & Donoghue, 2004; Bryant et al., 2008).由此带来较高的生态位重叠度, 促进了竞争排斥作用, 从而降低了种间稳定共存的概率, 使得谱系结构趋于发散(Machac et al., 2011).高海拔地区则与此情况相反, 以冬季低温为代表的严峻气候条件可对多数物种形成生理限制.这种强烈的生境过滤将大多数尚未演化出冷适应性状的物种筛选排除, 仅剩少数耐寒分支(Wiens & Donoghue, 2004; Hawkins et al., 2014), 从而在谱系关系上呈现出聚集趋势. ...
Elevation gradients of species-density: Historical and prospective views. 1 2001
... 山地生态系统通常在有限的空间内具备宽广的水热梯度和较高的生境异质性(Gaston, 2000; Körner, 2007; Sheldon et al., 2011).这些特点一方面为新物种形成创造了有利条件, 使山地成为多样性的热点地域; 另一方面, 由于该系统对全球气候变化十分敏感, 从而增加了物种的灭绝风险.因此, 物种多样性沿海拔梯度的变化情况常被视作大尺度纬向格局的镜像, 长久以来为人们所关注(Stevens, 1992; Lomolino, 2001; Rahbek, 2005; Tang et al., 2012a).然而, 有关群落谱系结构及其多样性沿海拔梯度变化规律的研究目前还处于起步阶段(Machac et al., 2011), 尚未形成统一的认识.现有经验案例显示, 研究结果会随着山地和生物类群的不同而存在显著差异.一般来说, 高海拔低温可对多数物种构成主要的生理胁迫, 因而群落谱系结构理论上应趋向聚集.这一预测已在蚂蚁(Machac et al., 2011)、蜂鸟(Graham & Mcguire, 2009)、蕨类植物(Kluge & Kessler, 2011)以及高山被子植物(Li et al., 2014)等多个类群得到验证.而Bryant等(2008)对美国落基山脉植物群落和土壤微生物群落谱系结构的海拔格局进行对比研究发现: 与理论预期相反, 该山地被子植物在低海拔地区谱系聚集, 而在高海拔地区谱系发散.卢孟孟等(2014)通过探索哀牢山森林群落谱系结构在海拔梯度上的变化, 发现采用不同指标得到的结论并不一致. ...
Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. 1 2008
 , 王菁兰
, 王菁兰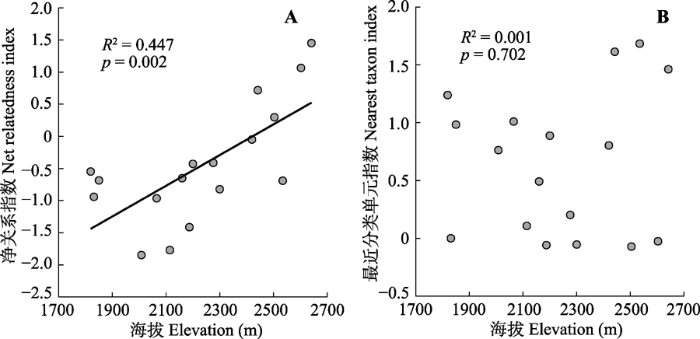 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT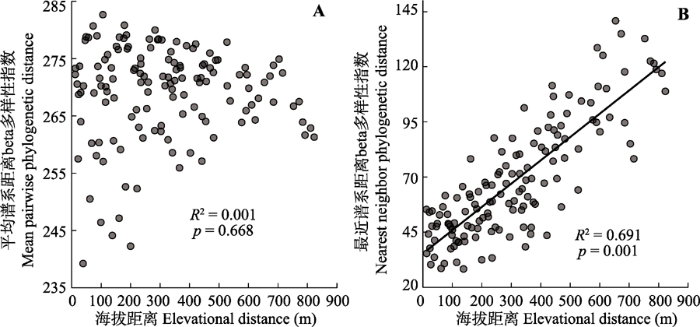 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT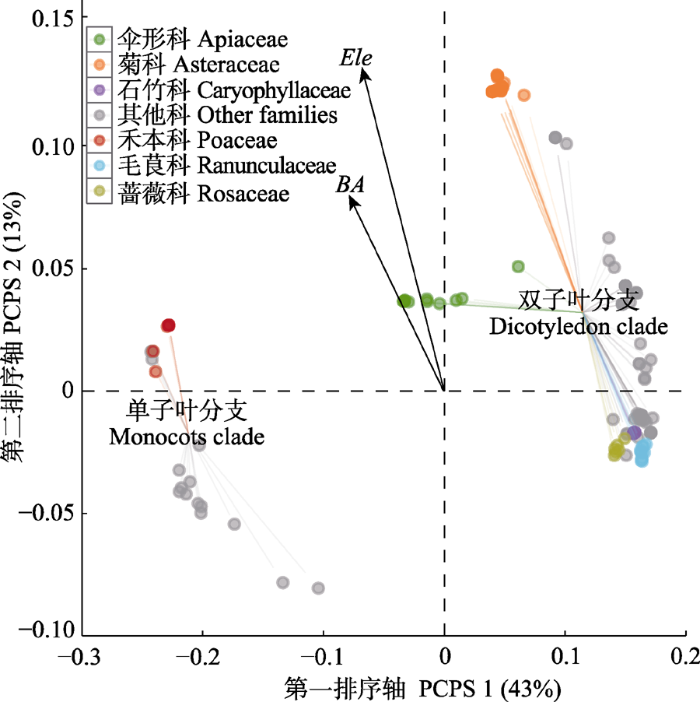 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}