Accumulation of humic acid and fulvic acid during root humification of three diameters of two dominant subalpine trees in western Sichuan, China
LIU Qun1, ZHUANG Li-Yan1, YANG Wan-Qin1,2, NI Xiang-Yin1, LI Ting-Ting1, XU Zhen-Feng1,2,*, 1Key Laboratory of Ecological Forestry Engineering of Sichuan Province, Institute of Ecology & Forest, Sichuan Agricultural University, Chengdu 611130, China;and 2Long-Term Research Station of Alpine Forest Ecosystems, Collaborative Innovation Center of Ecological Security in the Upper Reaches of Yangtze River, Chengdu 611130, China 通讯作者:* 通信作者Author for correspondence (E-mail: sicauxzf@163.com) 责任编辑: 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(31570601和31570445)、国家重点研发计划(2016YFC0502505和2017YFC0505003)、四川省教育厅重点基金(17ZA0321)和中国博士后科学基金项目(2014T70880)
关键词:胡敏酸;富里酸;径级;根系腐殖化 Abstract Aims Plant roots store large amount of terrestrial carbon, but little is known about humus formation processes during the decomposing root litter. Compared with coarse roots, fine roots have greater nutrients, which may be favorable to humus formation. The objective of the study was to examine how root diameters affect their humus formation processes. Methods In this study, in order to examine the accumulation of humic acid and fulvic acid of three root diameter classes (0-2, 2-5 and 5-10 mm) of two subalpine tree species (Abies faxoniana and Picea asperata) on the eastern Qinghai-Xizang Plateau of China, a two-year field experiment was conducted using a litter-bag method. Air-dried roots of A. faxoniana and P. asperata were placed in litterbags and incubated at 10 cm of soil depth in October 11th, 2013. Duplicate litter bags were collected in May (late winter) and October (late in the growing season) of 2014 and 2015, respectively. Concentrations of humic acid and fulvic acid were measured, and net accumulations were calculated for different periods. Important findings The concentrations of humic acid and fulvic acid were significantly influenced by root diameter that humic acid and fulvic acid decreased with increase in root diameter. Root diameter had significant effects on the net accumulation of humic acid, but not for the accumulation of fulvic acid. However, there were no significant differences in both humic acid and fulvic acid between A. faxoniana and P. asperata roots. Regardless of tree species, humic acid degraded during the winter but accumulated during the growing season. After two years of decomposition, the net accumulations of humic acid in 0-2, 2-5 and 5-10 mm roots were 8.0, 10.8 and 7.6 g·kg-1 for P. asperata and 15.2, 8.0 and 7.8 g·kg-1 for A. faxoniana, respectively. Conversely, the degradation of fulvic acid in 0-2, 2-5 and 5-10 mm roots were 178.0, 166.0 and 118.0 g·kg-1 for P. asperata and 170.0, 160.0 and 128.0 g·kg-1 for A. faxoniana, respectively. Our results suggest that diameter-associated variations in substrate quality could be an important driver for root litter humification in this subalpine forest. Moreover, diameter effect is dependent on decomposition period in this specific area. Keywords:humic acid;fulvic acid;diameter size;root humification -->0 PDF (1094KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 刘群, 庄丽燕, 杨万勤, 倪祥银, 李婷婷, 徐振锋. 川西亚高山两种树木不同径级根系腐殖化过程中胡敏酸和富里酸的累积特征. 植物生态学报, 2017, 41(12): 1251-1261 https://doi.org/10.17521/cjpe.2017.0169 腐殖质是一种结构复杂的高分子有机化合物, 是生物残体在微生物作用下经降解、络合形成的产物(Stevenson, 1994)。植物残体腐殖化过程中腐殖质的累积是土壤有机质形成和碳吸存的一个重要途径。已有研究发现不同有机物料(如稻草)能不同程度地提高土壤腐殖质含量(Chien et al., 2006; 窦森, 2010)。但这些研究集中于地表植物残体, 而对地下根系的研究较少。研究表明, 在微生物作用下, 植物根系木质素及其降解产物(如醌类)氧化生成的化合物构成腐殖质的芳香环骨架(Amir et al., 2006), 促进腐殖质累积, 影响土壤腐殖质的性质和结构(Asli & Neumann, 2010), 对土壤形成、发育具有重要意义(Abakumov et al., 2013; Berg & McClaugherty, 2014)。 由于提取方法的限制, 目前腐殖质的研究主要是胡敏酸和富里酸的研究(Lehmann & Kleber, 2015)。已有研究表明, 由于胡敏酸溶于碱而富里酸溶于酸的性质, 导致腐殖质累积受土壤环境、季节变化、基质品质和微生物等综合因素的影响(Prescott et al., 2000; Ponge & Chevalier, 2006)。在同一气候条件下, 根系分解主要受自身化学性质影响(Silver & Miya, 2001)。不同物种、不同直径的根系化学组分差异显著(靳贝贝和国庆喜, 2013; Makita et al., 2015)。随着林木根系直径的增加, 养分浓度(如N、P含量)降低, 导致不同径级根系分解速率不同(Silver & Miya, 2001; Ludovici & Kress, 2006), 这可能影响后续腐殖化进程。细根分解产物是一种潜在的碳源, 而粗根分解主要取决于气候变化, 尤其是温度的影响(Zhang & Wang, 2015)。在高寒地区, 碳周转较慢, 且季节性雪被和冻融作用对根系分解有重要影响(Henry, 2008), 这可能影响植物不同直径根系腐殖化过程。虽然一些研究已经表明, 细根和粗根形态、养分含量、功能和分解机制显著不同(Guo et al., 2004; Aulen et al., 2011; Goebel et al., 2011; Olajuyigbe et al., 2012), 但细根和粗根在不同气候条件下的腐殖质累积特征在很大程度上仍然是未知的。 川西亚高山森林地处青藏高原东缘, 在调节区域气候、保持水土和保育生物多样性等方面具有重要的作用(Yang et al., 2005; 邓仁菊等, 2009)。常年的低温和频繁的地质活动, 造成川西亚高山森林土壤发育缓慢(Yang et al., 2005)。已有研究表明, 川西亚高山植物根系分布主要集中于表层土壤(胡建利等, 2009), 该层土壤是有机质形成和腐殖化最为活跃的层次。因此, 根系腐殖化对该区域森林土壤有机质形成具有重要意义。前期研究表明, 冬季雪被显著影响凋落叶腐殖质累积(倪祥银等, 2014b), 且这种影响在冬季和生长季节明显不同(Ni et al., 2015), 但并未关注根系腐殖化过程中不同根系直径的胡敏酸和富里酸累积的变化格局。因此, 本研究在前期研究基础上, 初步研究了川西亚高山森林两种优势林木粗枝云杉(Picea asperata)和岷江冷杉(Abies faxoniana)不同径级根系(0-2、2-5和5-10 mm)在两年腐殖化过程中胡敏酸和富里酸在冬季和生长季节的累积特征, 为进一步了解川西亚高山森林生态系统地下碳循环过程提供基础数据。
川西亚高山森林根系腐殖化过程中不同径级对胡敏酸含量影响显著(p < 0.05), 而径级和树种差异对胡敏酸净累积量均无显著影响, 但不同径级间有一定的波动, 时期与径级的交互作用显著(p < 0.05) (表2)。经过两年的腐殖化分解, 两树种不同径级根系胡敏酸含量表现出在各个时期均随径级的增大而显著减少(p < 0.05)的趋势(图2A-2B)。而胡敏酸均净累积, 每5 g粗枝云杉细根、中根和粗根中两年总的净累积量为0.040、0.054 、0.038 g; 岷江冷杉为0.076、0.040、0.039 g (图3A-3B)。第一年冬季, 两树种不同径级根系胡敏酸表现出不同程度的降解, 且粗枝云杉根系胡敏酸净累积量随径级的增大而显著增加(p < 0.05); 生长季节, 两树种根系的胡敏酸均累积, 随根系径级增大, 岷江冷杉胡敏酸净累积量显著减少(p < 0.05)。第二年, 两树种根系的胡敏酸均表现出不同程度的降解, 但在生长季节胡敏酸表现出不同程度的累积, 且不同径级间根系胡敏酸净累积量差异不显著。经过两年的腐质化, 两树种不同径级根系胡敏酸在冬季均降解, 生长季节均累积。岷江冷杉和粗枝云杉在生长季节的累积量(0.25 g和0.26 g)分别大于冬季的降解量(0.10 g和0.13 g), 且在第一年生长季节的累积量(0.25 g和0.20 g)分别大于第二年生长季节的累积量(0.05 g和0.01 g)。 Table 1 表1 表1粗枝云杉和岷江冷杉不同径级根系的初始品质特征(平均值±标准误差, n = 3) Table 1Initial chemical quality in three root diameter classes of Picea asperata and Abies faxoniana (mean ± SE, n = 3)
物种 Species
径级 Diameter (mm)
碳 Carbon (C) (g·kg-1)
氮 Nitrogen (N) (g·kg-1)
磷 Phosphorus (P) (g·kg-1)
碳氮比 C/N
碳磷比 C/P
木质素 Lignin (%)
纤维素 Cellulose (%)
木质素/N Lignin/N
木质素/P Lignin/P
木质素/纤维素 Lignin/Cellulose
粗枝云杉 Picea asperata
0-2
496.4 ± 7.1
5.6 ± 0.4
0.34 ± 0.01
89 ± 4
1 450 ± 33
16 ± 0.6
28.9 ± 6.9
20.8 ± 3.3
51.7 ± 3.5
843 ± 40
2-5
496.6 ± 6.5
4.2 ± 0.2
0.33 ± 0.04
118 ± 5
1 538 ± 139
13 ± 1.3
26.1 ± 3.4
23.0 ± 1.6
62.0 ± 2.3
808 ± 76
5-10
512.1 ± 7.8
2.6 ± 0.2
0.26 ± 0.04
195 ± 18
2 022 ± 291
10 ± 1.7
25.9 ± 6.6
32.3 ± 9.6
98.7 ± 6.3
1 022 ± 128
岷江冷杉 Abies faxoniana
0-2
498.8 ± 20.0
5.0 ± 0.4
0.32 ± 0.01
99 ± 5
1 559 ± 74
16 ± 1.5
38.8 ± 6.1
16.2 ± 5.7
77.2 ± 4.9
1 214 ± 3 8
2-5
533.9 ± 7.3
3.8 ± 0.6
0.30 ± 0.01
144 ± 19
1 760 ± 44
12 ± 1.9
34.7 ± 10.8
18.9 ± 5.7
93.6 ± 12.9
1 142 ± 19
5-10
540.9 ± 13.3
2.4 ± 0.3
0.23 ± 0.04
231 ± 21
2 397 ± 360
10 ± 0.8
33.0 ± 6.6
32.8 ± 9.7
141.1 ± 14.6
1 465 ± 230
物种 Species (S)
***
***
NS
***
***
***
***
***
***
*
径级 Diameter (D)
**
***
***
***
***
***
***
***
**
**
S × D
*
NS
NS
NS
***
*
***
NS
NS
NS
Two-way ANOVA, Significant effects (NS, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001).双因素方差分析(NS, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001)。 新窗口打开 Table 2 表2 表2不同分解时期、物种、径级对胡敏酸含量和净累积量、富里酸含量和净累积量以及胡敏酸/富里酸影响的重复测量方差分析结果 Table 2Results of repeated measures ANOVA testing for the effects for date, tree species and root diameter on concentrations and net accumulations of humic acid and fulvic acid as well as on humic acid to fulvic acid ratio
因子 Factor
胡敏酸含量的F值 F value for humic acid concentration
胡敏酸净累积量的F值 F value for net accumulations of humic acid
富里酸含量的F值 F value for fulvic acid concentration
富里酸净累积量的F值 F value for net accumulations of fulvic acid
胡敏酸/富里酸的F值 F value for humic acid to fulvic acid ratio
时期 Date (T)
97.950***
46.690***
366.440***
82.531***
146.677***
树种 Species (S)
0.067
0.356
0.154
0.001
1.123
径级 Diameter (D)
63.535***
1.079
37.853***
19.003***
1.146
时期×物种 T × S
1.270
0.899
4.519**
4.622**
2.266
时期×径级 T × D
5.054**
2.704*
4.704**
1.578
0.322
物种×径级 S × D
3.751
1.871
4.609*
0.710
1.145
时期×物种×径级 T × S × D
1.642
1.272
2.141
2.348
0.532
*, p < 0.05; **, p < 0.01; ***, p < 0.001. 新窗口打开 显示原图|下载原图ZIP|生成PPT 图2粗枝云杉和岷江冷杉不同径级根系在不同采样日期的胡敏酸和富里酸含量(平均值±标准误差, n = 3)。*, p < 0.05。 -->Fig. 2Concentrations of humic acid and fulvic acid in three root diameter classes of Picea asperata and Abies faxoniana on different sampling dates (mean ± SE, n = 3). *, p < 0.05. -->
川西亚高山森林根系腐殖化过程中胡敏酸/富里酸总体上表现出随径级增大而减小的趋势, 除第二年生长季节以外的所有时期胡敏酸/富里酸均小于1 (图4)。第一年, 两树种根系胡敏酸/富里酸在不同径级间差异不显著, 且冬季比生长季节小。第二年生长季节, 粗枝云杉和岷江冷杉根系胡敏酸/富里酸随径级增大而减少, 粗枝云杉细根、中根和粗根胡敏酸/富里酸值分别为1.58、1.62、1.18, 岷江冷杉为1.92、1.68、1.80, 远远高于冬季。就整个研究期而言, 第一年冬季和生长季胡敏酸/富里酸分别小于第二年冬季和生长季。 显示原图|下载原图ZIP|生成PPT 图3粗枝云杉和岷江冷杉不同径级根系不同分解时期的胡敏酸、富里酸净累积量(平均值±标准误差, n = 3)。W1, 第一年冬季; Gs1, 第一年生长季节; W2, 第二年冬季; Gs2, 第二年生长季节; 1st yr, 第一年; 2nd yr, 第二年; two yr, 两年。不同小写字母表示在同一树种不同径级之间差异显著(p < 0.05)。 -->Fig. 3Net accumulation of humic acid and fulvic acid in three root diameter classes of Picea asperata and Abies faxoniana during the winter and growing season (mean ± SE, n = 3). W1, the first winter; Gs1, the first growing season; W2, the second winter; Gs2, the second growing season; 1st yr, the first year; 2nd yr, the second year; two yr, two years. Different lowercase letters show significant differences among root diameter classes for the same tree species (p < 0.05). -->
已有研究表明, 植物根系分解与腐殖化密切相关(Stevenson, 1994), Ciarkowska和Miech Wka (2017)认为高C/N影响腐殖质的形成, 腐殖化程度与N含量增加密切相关(Kramer et al., 2003), Melillo等(1982)发现分解速率与木质素/N呈极显著的负相关关系。而细根(直径<2 mm)和粗根(直径>2 mm)的分解对土壤C及养分的输入贡献存在显著差异(Zhang & Wang, 2015), 因此根系腐殖化过程中腐殖质含量可能受植物根系径级影响。对川西亚高山森林粗枝云杉和岷江冷杉根系腐殖化研究发现, 胡敏酸和富里酸含量在各时期均随径级的增大而显著减少(图2)。这可能是因为本研究中两树种根系N含量随径级增大而降低(表1), 促进微生物作用生成腐殖质(Cotrufo et al., 2013), 因此细根腐殖质含量更高。本研究还发现, 胡敏酸净累积量在细根、中根和粗根间差异不显著, 但富里酸净累积量差异显著(表2)。这可能是因为在根系腐殖化过程中, 较细的根含有大量可溶性组分, 能被土壤微生物更有效地利用(Cotrufo et al., 2013), 增强微生物聚合作用, 易形成分子量较小的富里酸(张夫道和Fokina, 1994), 导致细根、中根和粗根的富里酸净累积量差异显著。其次, 随腐殖化程度的加深, 富里酸转化为分子量更大、结构更复杂的胡敏酸, 其所需前体物质木质素及其降解产物(窦森, 2010)增多。而前期研究发现, 根系径级在短期内对木质素降解的影响不明显(唐仕姗等, 2015), 导致细根、中根和粗根供应给富里酸转化为胡敏酸的底物浓度无显著差异, 因此根系径级对胡敏酸净累积量的影响不显著。 显示原图|下载原图ZIP|生成PPT 图4粗枝云杉和岷江冷杉不同径级根系在不同分解时期的胡敏酸/富里酸(平均值±标准误差, n = 3)。i, 初始值; W1, 第一年冬季; Gs1, 第一年生长季节; W2, 第二年冬季; Gs2, 第二年生长季节。不同小写字母表示在同一树种不同径级之间差异显著(p < 0.05)。 -->Fig. 4Humic acid and fulvic acid in three root diameter classes of Picea asperata and Abies faxoniana during winter and growing season (mean ± SE, n = 3). i, initial; W1, the first winter; Gs1, the first growing season; W2, the second winter; Gs2, the second growing season. Different lowercase letters show significant differences among root diameter classes for the same tree species (p < 0.05). -->
Table 3 表3 表3分解两年后胡敏酸和富里酸净累积量与初始基质品质的相关关系 Table 3Relationships between the net accumulation of humic acid and fulvic acid and the initial root quality after 2-year decomposition
川西亚高山森林中岷江冷杉和粗枝云杉的根系在两年的腐殖化过程中, 胡敏酸、富里酸在不同时期均表现出随根系径级的增大而减少的趋势, 富里酸净累积量在不同径级间差异显著, 而胡敏酸净累积量差异不显著; 胡敏酸、富里酸的含量和净累积量在岷江冷杉和粗枝云杉间差异不显著。同时, 两树种不同径级根系中胡敏酸在冬季降解在生长季累积, 且第一年生长季胡敏酸累积量显著大于第二年生长季, 但富里酸总体表现为降解。分解两年后, 胡敏酸净累积量与根系基质初始品质无显著相关关系, 而富里酸净累积量与根系基质初始品质显著相关。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
AbakumovEV, CajthamlT, BrusJ, FrouzJ (2013). Humus accumulation, humification, and humic acid composition in soils of two post-mining chronosequences after coal mining .Journal of Soils and Sediments, 13, 491-500.https://doi.org/10.1007/s11368-012-0579-9URL [本文引用: 3]
AmirS, HafidiM, LemeeL, MerlinaG, GuiresseM, PinelliE, RevelJC, BaillyJR, AmblesA (2006). Structural characterization of humic acids, extracted from sewage sludge during composting, by thermochemolysis-gas chromatography- mass spectrometry .Process Biochemistry, 41, 410-422.https://doi.org/10.1016/j.procbio.2005.07.005URL [本文引用: 3]
[4]
AsliS, NeumannPM (2010). Rhizosphere humic acid interacts with root cell walls to reduce hydraulic conductivity and plant development .Plant and Soil, 336, 313-322.https://doi.org/10.1007/s11104-010-0483-2URL [本文引用: 1]摘要 Humic acids are ubiquitous, organic-end-products of the chemical and microbial degradation of dead biota in soils throughout the world. Humic acids can be transported in soil water as heterogeneous, supra-molecular, colloidal-agglomerates. Humic acid accumulation in the rhizosphere of transpiring plants may chemically stimulate development by increasing root availability of mineral nutrients and/or growth regulatory biomolecules. This report introduces novel, physical mechanisms by which humic acid can also reduce plant development. Effects of humic acid addition to the root media of intact maize plants (Zea mays L.) on their growth, transpiration and resistance to water deficits were assayed, as were the effects of external humic acid on the hydraulic conductivity of excised primary-roots. Humic acid reduced shoot growth, transpiration and resistance to water stress but not root growth. Root hydraulic conductivity was reduced by up to 44% via a time-, concentration- and size-dependent fouling mechanism resulting from humic acid accumulation at root cell-walls. Thus, humic acid is shown, apparently for the first time, to be able to exert novel physical effects in addition to its known chemical effects on plant development.
[5]
AulenM, ShipleyB, BradleyR (2011). Prediction of in situ root decomposition rates in an interspecific context from chemical and morphological traits . Annals of Botany, 109, 287-297.https://doi.org/10.1093/aob/mcr259URLPMID:22003237 [本文引用: 1]摘要 Abstract BACKGROUND AND AIMS: We quantitatively relate in situ root decomposition rates of a wide range of trees and herbs used in agroforestry to root chemical and morphological traits in order to better describe carbon fluxes from roots to the soil carbon pool across a diverse group of plant species. METHODS: In situ root decomposition rates were measured over an entire year by an intact core method on ten tree and seven herb species typical of agroforestry systems and were quantified using decay constants (k values) from Olson's single exponential model. Decay constants were related to root chemical (total carbon, nitrogen, soluble carbon, cellulose, hemicellulose, lignin) and morphological (specific root length, specific root length) traits. Traits were measured for both absorbing and non-absorbing roots. KEY RESULTS: From 61 to 77 % of the variation in the different root traits and 63 % of that in root decomposition rates was interspecific. N was positively correlated, but total carbon and lignin were negatively correlated with k values. Initial root traits accounted for 75 % of the variation in interspecific decomposition rates using partial least squares regressions; partial slopes attributed to each trait were consistent with functional ecology expectations. CONCLUSIONS: Easily measured initial root traits can be used to predict rates of root decomposition in soils in an interspecific context.
ChienSW, WangMC, HsuJH, SeshaiahK (2006). Influence of fertilizers applied to a paddy-upland rotation on characteristics of soil organic carbon and humic acids .Journal of Agricultural and Food Chemistry, 54, 6790-6799.https://doi.org/10.1021/jf061275mURLPMID:16939341 [本文引用: 1]摘要 The qualitative and quantitative characteristics of soil organic carbon (SOC) and related humic acids (HAs) extracted from the soils of field plots were investigated after 8 years of annual paddy (Oryza sativa L.) and upland maize (Zea mays L.) rotation with various fertilizations. Seven fertilization treatments were selected: Ck (no inputs); Chem (chemical fertilizer of NPK); Comp (swine compost); Comp + 33% of Chem N rate; Comp + 67% of Chem N rate; GM (legume green manure) + 33% of Chem N rate; and peat + 33% of Chem N rate. Organic and inorganic nitrogen inputs of six treatments were equivalent with respect of nitrogen content, but Comp, GM, and peat treatments were complemented with various amounts of inorganic N. After harvest of the eighth paddy crop, surface soil samples collected from the plots were subjected to soil characterizations and extraction of humic substances, which were used for chemical, spectroscopic (FTIR, 13C NMR, ESR, X-ray diffractometry), delta13C, and 14C dating analyses. The yields of HAs extracted from the seven treatments were significantly different. Treatment containing persistent organic compound such as the peat + 33% N treatment increased the humification process in topsoils and produced higher yield of HA. Spectroscopic analyses revealed that fertilization treatments changed the functional groups, alkyl C, crystalline characteristics, and delta13C ratios of HAs and turnover rate of SOC considerably. The SOC of the peat + 33% N treatment had the highest mean residence time of 3100 years. Various fertilizer treatments are correlated with turnover rate of SOC and related HAs, which are associated with concerned carbon sequestration as well as mitigation of CO2 emission in the soil environment.
[9]
CiarkowskaK, Miech WkaA (2017). The role of bilberry and alpine lady-fern in soil formation within the Carpathian subalpine spruce forest stands .Geoderma, 305, 162-172.https://doi.org/10.1016/j.geoderma.2017.06.004URL
[10]
CotrufoMF, WallensteinMD, BootCM, DenefK, PaulE (2013). The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Global Change Biology, 19, 988-995.https://doi.org/10.1111/gcb.12113URL [本文引用: 4]
DengRJ, YangWQ, FengRF, HuJL, QinJL, QiongXJ (2009). Mass loss and element release of litter in the subalpine forest over one freeze-thaw season .Acta Ecologica Sinica, 29, 5730-5735.(in Chinese with English abstract) [邓仁菊, 杨万勤, 冯瑞芳, 胡建利, 秦嘉励, 熊雪晶 (2009). 季节性冻融期间亚高山森林凋落物的质量损失及元素释放 . 生态学报, 29, 5730-5735.]https://doi.org/10.3321/j.issn:1000-0933.2009.10.065URL [本文引用: 2]摘要 季节性冻融期间的凋落物分解对季节性冻土区的森林生态系统过程可能具有重要的影响,但已有的研究报道很少。因此,采用凋落物分解袋法研究了岷江冷杉(Abies faxoniana Rehder&amp;E.H.Wilson)林和白桦(Betula platyphylla Sukaczev)林凋落叶的分解。一个季节性冻融期间,冷杉林和白桦林凋落物的质量损失率分别为(19.4±2.0)%和(21.5±3.5)%,约为1a中凋落物分解的64.5%和65.6%,表明季节性冻融对亚高山森林凋落物分解影响显著。冷杉凋落物中C、N、P、K、Ca和Mg的释放率为(15.0±1.0)%、(34.1±3.6)%、(17.0±0.9)%、(22.8±5.9)%、(20.1±0.1)%和(36.3±2.1)%,白桦凋落物中C、N、P、K、Ca和Mg的释放率为(20.7±0.1)%、(29.4±3.4)%、(15.7±1.3)%、(16.8±5.1)%、(21.3±1.8)%和(20.5±2.8)%。结合叶凋落物产量可以推断,冷杉林凋落物在一个季节性冻融期间释放到土壤的N、P、K、Ca、Mg为(10.17±1.14)kg·hm^-2、(0.68±0.08)kg·hm^-2、(4.08±0.46)kg·hm^-2、(0.46±0.05)kg·hm^-2、(0.09±0.01)kg·hm^-2,白桦林为(5.61±1.12)kg·hm^-2、(0.34±0.07)kg·hm^-2、(1.21-0.24)kg·hm^-2、(0.300±0.059)kg·hm^-2、(0.051±0.010)kg·hm^-2,这对于春季亚高山森林植物生长具有重要的生态学意义。
DouS, YuSQ, ZhangJJ (2007). Effects of carbon dioxide concentration on humus formation in cornstalk decomposition .Acta Pedologica Sinica, 44, 458-466.(in Chinese with English abstract) [窦森, 于水强, 张晋京 (2007). 不同CO2浓度对玉米秸秆分解期间土壤腐殖质形成的影响 . 土壤学报, 44, 458-464.]https://doi.org/10.3321/j.issn:0564-3929.2007.03.012URL [本文引用: 1]摘要 以往人们较多的研究了土壤有机碳与温室气体的源、汇关系问题,却很少研究温室气体对土壤有机碳本身分解转化和各组分性质的影响。为了探讨CO2浓度升高对腐殖物质形成转化的作用,本文通过室内人为高浓度CO2的培养试验,用腐殖质组成修改法研究了玉米秸秆分解期间(1~180d)土壤总有机碳(TOC)、水溶性物质(WSS)、碱提取腐殖物质(HE)、胡敏酸(HA)和胡敏素(HM)数量的动态变化规律及不同CO2浓度对有机碳各组分形成与转化的影响。结果表明:玉米秸秆分解期间,“新形成”的净TOC逐渐降低,净HE表现为先增加而后下降的趋势,HM的绝对数量逐渐下降。可提取腐殖物质中HA的比例(PQ)先增加后下降,最终趋于平稳,说明最初富里酸(FA)的形成速度大于HA,随培养时间的延长,FA和HA经历了一段相互转化的过程,最终达到动态平衡。30%CO2浓度(V/V)处理的TOC、WSS、HE和HM数量明显高于3%CO2浓度处理和正常CO2浓度(0.0375%)处理,其中WSS和HM反应更为敏感。但3%CO2浓度处理与正常CO2浓度处理的差异不明显。可提取腐殖物质的PQ的顺序是30%CO2浓度处理3%CO2浓度处理正常CO2浓度处理,说明CO2浓度增加更有利于FA而不利于HA的形成和稳定。
[15]
ElliottJ (2013). Evaluating the potential contribution of vegetation as a nutrient source in snowmelt runoff .Canadian Journal of Soil Science, 93, 435-443.https://doi.org/10.4141/cjss2012-050URL [本文引用: 1]
[16]
GoebelM, HobbieSE, BulajB, ZadwornyM, ArchibaldDD, OleksynJ, ReichPB, EissensstatDM (2011). Decomposition of the finest root branching orders, linking belowground dynamics to fine-root function and structure .Ecological Monographs, 81, 89-102.https://doi.org/10.1890/09-2390.1URL [本文引用: 1]
[17]
GuoDL, MitchellRJ, HendricksJJ (2004). Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest .Oecologia, 140, 450-456.https://doi.org/10.1007/s00442-004-1596-1URL [本文引用: 1]
[18]
HaoXD, ZhouP, CaoYL (2017). Origins and evolution processes of humic substances in wastewater treatment .Chinese Journal of Environmental Engineering, 11, 1-11.(in Chinese with English abstract) [郝晓地, 周鹏, 曹亚莉 (2017). 污水处理中腐殖质的来源及其演变过程 . 环境工程学报, 11, 1-11.]
[19]
Henry (2008). Climate change and soil freezing dynamics: Historical trends and projected changes .Climatic Change, 87, 421-434.https://doi.org/10.1007/s10584-007-9322-8URL摘要 Changes to soil freezing dynamics with climate change can modify ecosystem carbon and nutrient losses. Soil freezing is influenced strongly by both air temperature and insulation by the snowpack, and it has been hypothesized that winter climate warming may lead to increased soil freezing as a result of reduced snowpack thickness. I used weather station data to explore the relationships between winter air temperature, precipitation and soil freezing for 31 sites in Canada, ranging from the temperate zone to the high Arctic. Inter-annual climate variation and associated soil temperature variation over the last 40/years were examined and used to interpolate the effects of projected climate change on soil freezing dynamics within sites using linear regression models. Annual soil freezing days declined with increasing mean winter air temperature despite decreases in snow depth and cover, and reduced precipitation only increased annual soil freezing days in the warmest sites. Annual soil freeze haw cycles increased in both warm and dry winters, although the effects of precipitation were strongest in sites that experience low mean winter precipitation. Overall, it was projected that by 2050, changes in winter temperature will have a much stronger effect on annual soil freezing days and freeze haw cycles than changes in total precipitation, with sites close to but below freezing experiencing the largest changes in soil freezing days. These results reveal that experimental data relevant to the effects of climate changes on soil freezing dynamics and changes in associated soil physical and biological processes are lacking.
[20]
HobbieSE (2008). Nitrogen effects on decomposition: A five-year experiment in eight temperate sites .Ecology, 89, 2633-2644.https://doi.org/10.1890/07-1119.1URLPMID:18831184 [本文引用: 1]摘要 Abstract The influence of inorganic nitrogen (N) inputs on decomposition is poorly understood. Some prior studies suggest that N may reduce the decomposition of substrates with high concentrations of lignin via inhibitory effects on the activity of lignin-degrading enzymes, although such inhibition has not always been demonstrated. I studied the effects of N addition on decomposition of seven substrates ranging in initial lignin concentrations (from 7.4% to 25.6%) over five years in eight different grassland and forest sites in central Minnesota, USA. I predicted that N would stimulate the decomposition of lignin-poor substrates but retard the decomposition of lignin-rich substrates. Across these sites, N had neutral or negative effects on decomposition rates. However, in contrast to my hypothesis, effects of N on decomposition were independent of substrate initial lignin concentrations, and decomposition of the lignin fraction was unaffected by N fertilization. Rather, substrate-site combinations that exhibited more rapid decomposition rates in the control treatment were affected more negatively by addition of N fertilization. Taken together, these results suggest that decreased decomposition with added N did not result from inhibition of lignin-degrading enzyme activity, but may have resulted from abiotic interactions between N fertilizer and products of microbial degradation or synthesis or from N effects on the decomposer community. Low initial substrate N concentrations and N fertilization both stimulated N immobilization, but the differences among substrates were generally much larger than the effects of fertilization. This study suggests that atmospheric N addition could stimulate ecosystem carbon sequestration in some ecosystems as a result of reduced rates of forest floor decomposition.
[21]
HuJL, YangWQ, ZhangJ, DengRJ (2009). Characteristics of biomass and carbon stock of fir and birch fine roots in subalpine forest of western Sichuan, China .Chinese Journal of Applied& Environmental Biology, 15, 313-317.(in Chinese with English abstract) [胡建利, 杨万勤, 张健, 邓仁菊 (2009). 川西亚高山冷杉和白桦细根生物量与碳储量特征 . 应用与环境生物学报, 15, 313-317.] [本文引用: 1]
[22]
JinBB, GuoQX (2013). Root decomposition and nutrient dynamics of Quercus mongolica and Betula platyphylla . Acta Ecologica Sinica, 33, 2416-2424.(in Chinese with English abstract) [靳贝贝, 国庆喜 (2013). 蒙古栎、白桦根系分解及养分动态 . 生态学报, 33, 2416-2424.] [本文引用: 2]
[23]
KramerMG, SollinsP, SlettenRS, SwartPK (2003). N isotope fractionation and measures of organic matter alteration during decomposition .Ecology, 84, 2021-2025.https://doi.org/10.1890/02-3097URL [本文引用: 1]
[24]
LehmannJ, KleberM (2015). The contentious nature of soil organic matter .Nature, 528, 60-68.https://doi.org/10.1038/nature16045URLPMID:26633767 [本文引用: 1]摘要 Tuberculosis is a major source of global mortality caused by infection, partly because of a tremendous ongoing burden of undiagnosed disease. Improved diagnostic technology may play an increasingly crucial part in global efforts to end tuberculosis, but the ability of diagnostic tests to curb tuberculosis transmission is dependent on multiple factors, including the time taken by a patient to seek health care, the patient's symptoms, and the patterns of transmission before diagnosis. Novel diagnostic assays for tuberculosis have conventionally been evaluated on the basis of characteristics such as sensitivity and specificity, using assumptions that probably overestimate the impact of diagnostic tests on transmission. We argue for a shift in focus to the evaluation of such tests' incremental value, defining outcomes that reflect each test's purpose (for example, transmissions averted) and comparing systems with the test against those without, in terms of those outcomes. Incremental value can also be measured in units of outcome per incremental unit of resource (for example, money or human capacity). Using a novel, simplified model of tuberculosis transmission that addresses some of the limitations of earlier tuberculosis diagnostic models, we demonstrate that the incremental value of any novel test depends not just on its accuracy, but also on elements such as patient behaviour, tuberculosis natural history and health systems. By integrating these factors into a single unified framework, we advance an approach to the evaluation of new diagnostic tests for tuberculosis that considers the incremental value at the population level and demonstrates how additional data could inform more-effective implementation of tuberculosis diagnostic tests under various conditions.
[25]
LiuH, YangWQ, NiXY, XiaoS, WuFZ (2015). Characters of different type of coarse woody debris humification in an alpine forest .Ecology and Environmental Sciences, 24, 1143-1149.(in Chinese with English abstract) [刘辉, 杨万勤, 倪祥银, 肖洒, 吴福忠 (2015). 高山森林不同类型粗木质残体腐殖化特征 . 生态环境学报, 24, 1143-1149.] [本文引用: 2]
[26]
LiuRL, YangWQ, TanB, WangWJ, NiXY, WuFZ (2013). Effects of soil fauna on N and P dynamics at different stages during the first year of litter decomposition in subalpine and alpine forests of western Sichuan .Chinese Journal of Plant Ecology, 37, 1080-1090.(in Chinese with English abstract) [刘瑞龙, 杨万勤, 谭波, 王文君, 倪祥银, 吴福忠 (2013). 土壤动物对川西亚高山和高山森林凋落叶第一年不同分解时期N和P元素动态的影响 . 植物生态学报, 37, 1080-1090.] [本文引用: 1]
[27]
LiuXH, ZouDY, KangXF, ChengYL, WangHY, ZhouCJ, WangSX (1999). The effect of persistent rotation fertilization on dynamic change in humic acid of brown earth .Chinese Journal of Soil Sciences, 30, 68-70.(in Chinese) [刘小虎, 邹德乙, 康笑峰, 程艳丽, 王洪岩, 周崇峻, 王绍新 (1999). 长期轮作施肥对棕壤腐殖酸动态变化的影响 . 土壤通报, 30, 21-23.]https://doi.org/10.3321/j.issn:0564-3945.1999.02.008URL [本文引用: 1]摘要 在沈阳地区棕壤上设置长期定位试验.结果表明,凡经15年在玉米-大豆-玉米轮作条件下连续施用猪厩肥和NPK化肥,耕层土壤胡敏酸C相对下降54.5%,富里酸C相对提高162.6%,HA/FA比由1979年的2.69降到1993年的0.46,土壤腐殖质组成以胡敏酸为主体转化为以富里酸为主体.本文还讨论了不同施肥处理对腐殖酸消长的影响.
[28]
LudoviciKH, KressLW (2006). Decomposition and nutrient release from fresh and dried pine roots under two fertilizer regimes .Canadian Journal of Forest Research, 36, 105-111.https://doi.org/10.1139/x05-227URL [本文引用: 1]
[29]
MakitaN, KawamuraA, OsawaA (2015). Size-dependent morphological and chemical property of fine root litter decomposition .Plant and Soil, 393, 283-295.https://doi.org/10.1007/s11104-015-2491-8URL [本文引用: 1]摘要 Aims: To estimate cause-and-effect relationships of decomposition rates of fine root litter up to 2 mm in diameter, we explored the relationships between mass loss rates and substrate litter properties with respect to chemical and morphological dynamics. Methods: Mass loss, nitrogen (N) concentration, specific root area (SRA), root tissue density (RTD), and their remaining proportions were measured for initial and residual fine roots of two diameter classes (<0.5 and 0.5-2 mm) of Quercus serrata and Ilex pedunculosa in a temperate forest. Litterbags were sequentially collected after 1 month of decomposition and then every 3 months for 27 months. Results: N concentration dynamics during decomposition showed a significant interaction between species and diameter, and there was a greater convergence between diameter classes for Q. serrata than for I. pedunculosa. The remaining area and volume of the two diameters changed during decomposition in both species, but the degree of change was somewhat different between the species. The remaining surface area and volume in I. pedunculosa roots showed greater differences between diameter classes compared with Q. serrata roots. Remaining mass was positively correlated with the remaining C concentration and surface area and was negatively correlated with the remaining tissue volume and N concentration, according to principal component analysis analysis. Conclusions: Differences in morphological and chemical properties between diameter sizes of two species could indicate differences in decomposition processes. Estimates of fine root decomposition rates are best obtained by a combination of biological and physicochemical species-specific processes.
[30]
MelilloJM, AberJD, MuratoreJF (1982). Nitrogen and lignin control of hardwood leaf litter decomposition dynamics .Ecology, 63, 621-626.https://doi.org/10.2307/1936780URL
[31]
NiXY, YangWQ, LiH, XuLY, HeJ, WuFZ (2014a). Effects of snowpack on early foliar litter humification during winter in a subalpine forest of western Sichuan . Chinese Journal of Plant Ecology, 38, 540-549.(in Chinese with English abstract) [倪祥银, 杨万勤, 李晗, 徐李亚, 何洁, 吴福忠 (2014a). 雪被斑块对川西亚高山森林6种凋落叶冬季腐殖化的影响 . 植物生态学报, 38, 540-549.]
[32]
NiXY, YangWQ, TanB, HeJ, XuLY, LiH, WuFZ (2015). Accelerated foliar litter humification in forest gaps: Dual feedbacks of carbon sequestration during winter and the growing season in an alpine forest . Geoderma, s241-242, 136-144. [本文引用: 2]
[33]
NiXY, YangWQ, XuLY, HeJ, LiH, WuFZ (2014b). Effects of winter snowpack on accumulation of humic acid and fulvic acid during humification of foliar litters in an alpine forest .Acta Pedologica Sinica, 51, 1138-1152.(in Chinese with English abstract) [倪祥银, 杨万勤, 徐李亚, 何洁, 李晗, 吴福忠 (2014b). 雪被斑块对高山森林凋落叶腐殖化过程中胡敏酸和富里酸累积的影响 . 土壤学报, 51, 1138-1152.]
[34]
OlajuyigbeS, TobinB, HawkinsM, NieuwenhuisM (2012). The measurement of woody root decomposition using two methodologies in a Sitka spruce forest ecosystem .Plant and Soil ,360, 77-91.https://doi.org/10.1007/s11104-012-1222-7URL [本文引用: 1]
SchimelJP, BilbroughC, WelkerJM (2004). Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities .Soil Biology & Biochemistry, 36, 217-227.https://doi.org/10.1016/j.soilbio.2003.09.008URL [本文引用: 1]摘要 Microbial activity in Arctic tundra ecosystems continues through the winter and is an important component of the annual C budget. This activity is sensitive to climatic variation, particularly snow depth because that regulates soil temperature. The influence of winter conditions on soil N cycling is poorly understood. In this study, we used intact core incubations sampled periodically through the winter and following growing season to measure net N mineralization and nitrification in dry heath and in moist tussock tundra under ambient and experimentally increased snow depths (by use of a snowfence). In dry heath, we sampled soils under Dryas octopetela or Arctostaphylos alpine , while in tussock tundra, we sampled Eriophorum vaginatum tussocks and Sphagnum dominated areas between tussocks. Our objectives were to: (1) examine how different winter snow regimes influenced year-round N dynamics in the two tundra types, and (2) evaluate how these responses are affected by dominant species present in each system. In tussock tundra, soils with increased winter snow cover had high net N mineralization rates during the fall and winter, followed by immobilization during thaw. In contrast, N mineralization only occurred during the autumn in soils with ambient snow cover. During the growing season when N immobilization dominated in areas with ambient snow cover, soils with increased winter snow cover had positive net mineralization and nitrification rates. In dry heath tundra, soils with increased snow depth had high late winter net N mineralization rates, but these rates were: (a) comparable to early winter rates in soils under Arctostaphylos plants with ambient snow cover; (b) greater in soils under Arctostaphylos plants than in soils under Dryas plants; and (c) less than the rates found in tussock tundra. Our findings suggest under ambient snow conditions, low soil temperatures limit soil N mineralization, but that deeper snow conditions with the associated warmer winter soil temperatures dramatically increase over-winter N mineralization and thereby alter the amount and timing of plant-available N in tundra ecosystems.
[38]
SilverWL, MiyaRK (2001). Global patterns in root decomposition: Comparisons of climate and litter quality effects .Oecologia, 129, 407-419.https://doi.org/10.1007/s004420100740URLPMID:28547196 [本文引用: 2]摘要 Root decomposition represents a significant C flux in terrestrial ecosystems. Roots are exposed to a different decomposition environment than aboveground tissues, and few general principles exist regarding the factors controlling rates of root decay. We use a global dataset to explore the relative importance of climate, environmental variables, and litter quality in regulating rates of root decomposition. The parameters that explained the largest amount of variability in root decay were root Ca concentrations and C:N ratios, with a smaller proportion explained by latitude, mean annual temperature, mean annual precipitation, and actual evapotranspiration (AET). Root chemistry and decay rates varied by plant life form (conifer, broadleaf, or graminoid). Conifer roots had the lowest levels of Ca and N, the highest C:N and lignin:N ratios, and decomposed at the slowest rates. In a stepwise multiple linear regression, AET, root Ca, and C:N ratio accounted for approximately 90% of the variability in root decay rates. Root chemistry appeared to be the primary controller of root decomposition, while climate and environmental factors played secondary roles, in contrast to previously established leaf litter decomposition models.
[39]
SmitAL, GeorgeE, GroenwoldJ (1999). Root observations and measurements at (transparent) interfaces with soil. In: Smit AL, Bengough AG, Engels C, Noordwijk MV, Pellerin S, vande Geijn SC eds. Root Methods . Springer- Verlag, New York.236-266.
[40]
StevensonFJ (1994). Humus Chemistry: Genesis, Composition, Reactions. 2nd edn . John Wiley & Sons, New York. 17. [本文引用: 2]
[41]
TangSH (2016). Root Decomposition of Different Diameters of Three Dominant Subalpine Trees and Its Responses to Experimental Warming in the Western Sichuan . Master degree dissertation, Sichuan Agricultural University, Chengdu.(in Chinese with English abstract) [唐仕姗 (2016). 川西亚高山3种优势林木不同径级根系分解及其对模拟增温的响应 . 硕士学位论文, 四川农业大学, 成都.]URL [本文引用: 1]
[42]
TangSH, YangWQ, HeW, WangHP, XiongL, NieFY, XuZF (2015). Root decomposition, lignin and cellulose degradation of three dominant subalpine trees of different diameters in western Sichuan .Chinese Journal of Applied&Environmental Biology, 21, 754-761.(in Chinese with English abstract) [唐仕姗, 杨万勤, 何伟, 王海鹏, 熊莉, 聂富育, 徐振锋 (2015). 川西亚高山3种优势林木不同径级根系分解及木质素、纤维素降解特征 . 应用与环境生物学报,21, 754-761.]https://doi.org/10.3724/sp.j.1145.2015.01043URL [本文引用: 2]摘要 采用尼龙网袋法研究川西亚高山3种优势林木(岷江冷杉、粗枝云杉和红桦)不同径级(≤2 mm,2-5 mm,≥5mm)的根系在生长季和非生长季的质量损失及木质素和纤维素降解特征.结果显示,红桦根系的质量损失率往往高于岷江冷杉和粗枝云杉;根系质量损失率随径级增加而下降;非生长季的质量损失率占全年质量损失率的52%-65%;方差分析表明,木质素降解在树种之间有显著差异(P0.05),而径级影响不显著(P0.05),但二者交互作用有显著影响(P0.05);3个树种木质素总体表现为非生长季降解生长季累积,且红桦降解或累积程度均显著高于岷江冷杉和粗枝云杉.树种、径级及其交互作用对纤维素降解均产生显著影响(P0.05).除了红桦根系以外,岷江冷杉和粗枝云杉纤维素降解没有明显的季节特征.综上所述,川西亚高山森林根系分解及木质素和纤维素降解在物种之间有所不同,而根系径级效应与物种及季节有一定关联.图3表2参39
WangH, HongYT, LinQH, HongB, ZhuYX, WangY, XuH (2010). Response of humification degree to monsoon climate during the Holocene from the Hongyuan peat bog, eastern Tibetan Plateau .Palaeogeography, 286, 171-177.https://doi.org/10.1016/j.palaeo.2009.12.015URL
[45]
WetterstedtJ?M, PerssonT, ?grenGRI (2010). Temperature sensitivity and substrate quality in soil organic matter decomposition: Results of an incubation study with three substrates .Global Change Biology, 16, 1806-1819. [本文引用: 1]
[46]
WuY, OnipcenkoVG (2007). The impact of snow-cover on alpine vegetation type of different aspects in the west of Sichuan Province .Acta Ecologica Sinica, 27, 5120-5129.(in Chinese with English abstract) [吴彦, OnipcenkoVG (2007). 雪被对川西高山植被坡向性分异的影响 . 生态学报, 27, 5120-5129.] [本文引用: 1]
[47]
YangWQ, WangKY, KellomkiS, GongHD (2005). Litter dynamics of three subalpine forests in western Sichuan .Pedosphere, 15, 653-659. [本文引用: 2]
[48]
ZhangFD, ForkinAD (1994). Study on the decomposition and transformation of crop straws C in soils (in Chinese) . Plant Nutrition and Fertilizer Science, 1, 27-38.(in Chinese with English abstract) [张夫道, Fokina D (1994). 作物秸秆碳在土壤中分解和转化规律的研究 . 植物营养与肥料学报, 1, 27-38.]
[49]
ZhangXJ, WuC, Mei, L, HanYZ, WangZQ (2006). Root decomposition and nutrient release ofFraxinus manshurica and Larix gmelinii plantations . Chinese Journal of Applied Ecology, 17, 1370-1376.(in Chinese with English abstract) [张秀娟, 吴楚, 梅莉, 韩有志, 王政权 (2006). 水曲柳和落叶松人工林根系分解与养分释放 . 应用生态学报, 17, 1370-1376.] [本文引用: 1]
[50]
ZhangXY, WangW (2015). The decomposition of fine and coarse roots: Their global patterns and controlling factors .Scientific Reports, 5, 9940. doi: 10.1038/srep09940.https://doi.org/10.1038/srep09940URL [本文引用: 2]
[51]
ZhouJ, YangWQ, WuFZ, TanB, DuanF, LiuH (2017). Dynamics on humic acid and fulvic acid in the stump systems with different log years in thePinus massoniana plantations . Journal of University of Chinese Academy of Sciences, 34, 521-528.(in Chinese with English abstract) [周蛟, 杨万勤, 吴福忠, 谭波, 段斐, 刘辉 (2017). 不同采伐年限马尾松人工林伐桩的胡敏酸和富里酸动态特征 . 中国科学院大学学报, 34, 521-528.] [本文引用: 1]
Humus accumulation, humification, and humic acid composition in soils of two post-mining chronosequences after coal mining 3 2013
... 腐殖质是一种结构复杂的高分子有机化合物, 是生物残体在微生物作用下经降解、络合形成的产物(Stevenson, 1994).植物残体腐殖化过程中腐殖质的累积是土壤有机质形成和碳吸存的一个重要途径.已有研究发现不同有机物料(如稻草)能不同程度地提高土壤腐殖质含量(Chien et al., 2006; 窦森, 2010).但这些研究集中于地表植物残体, 而对地下根系的研究较少.研究表明, 在微生物作用下, 植物根系木质素及其降解产物(如醌类)氧化生成的化合物构成腐殖质的芳香环骨架(Amir et al., 2006), 促进腐殖质累积, 影响土壤腐殖质的性质和结构(Asli & Neumann, 2010), 对土壤形成、发育具有重要意义(Abakumov et al., 2013; Berg & McClaugherty, 2014). ...
... 式中, CHC、CHA和CFA分别为腐殖质、胡敏酸和富里酸含量(倪祥银等, 2014b; Ni et al., 2015); 以胡敏酸、富里酸含量与每次采样测得根系质量残留量的乘积表示胡敏酸、富里酸累积量, 以两次采样日期的胡敏酸、富里酸累积量之差表征该时期胡敏酸、富里酸净累积量.以各采样日期胡敏酸/富里酸表征胡敏酸、富里酸的相对含量和形成速度(Abakumov et al., 2013). ...
Structural characterization of humic acids, extracted from sewage sludge during composting, by thermochemolysis-gas chromatography- mass spectrometry 3 2006
... 腐殖质是一种结构复杂的高分子有机化合物, 是生物残体在微生物作用下经降解、络合形成的产物(Stevenson, 1994).植物残体腐殖化过程中腐殖质的累积是土壤有机质形成和碳吸存的一个重要途径.已有研究发现不同有机物料(如稻草)能不同程度地提高土壤腐殖质含量(Chien et al., 2006; 窦森, 2010).但这些研究集中于地表植物残体, 而对地下根系的研究较少.研究表明, 在微生物作用下, 植物根系木质素及其降解产物(如醌类)氧化生成的化合物构成腐殖质的芳香环骨架(Amir et al., 2006), 促进腐殖质累积, 影响土壤腐殖质的性质和结构(Asli & Neumann, 2010), 对土壤形成、发育具有重要意义(Abakumov et al., 2013; Berg & McClaugherty, 2014). ...
Decomposition and nutrient release from fresh and dried pine roots under two fertilizer regimes 1 2006
... 由于提取方法的限制, 目前腐殖质的研究主要是胡敏酸和富里酸的研究(Lehmann & Kleber, 2015).已有研究表明, 由于胡敏酸溶于碱而富里酸溶于酸的性质, 导致腐殖质累积受土壤环境、季节变化、基质品质和微生物等综合因素的影响(Prescott et al., 2000; Ponge & Chevalier, 2006).在同一气候条件下, 根系分解主要受自身化学性质影响(Silver & Miya, 2001).不同物种、不同直径的根系化学组分差异显著(靳贝贝和国庆喜, 2013; Makita et al., 2015).随着林木根系直径的增加, 养分浓度(如N、P含量)降低, 导致不同径级根系分解速率不同(Silver & Miya, 2001; Ludovici & Kress, 2006), 这可能影响后续腐殖化进程.细根分解产物是一种潜在的碳源, 而粗根分解主要取决于气候变化, 尤其是温度的影响(Zhang & Wang, 2015).在高寒地区, 碳周转较慢, 且季节性雪被和冻融作用对根系分解有重要影响(Henry, 2008), 这可能影响植物不同直径根系腐殖化过程.虽然一些研究已经表明, 细根和粗根形态、养分含量、功能和分解机制显著不同(Guo et al., 2004; Aulen et al., 2011; Goebel et al., 2011; Olajuyigbe et al., 2012), 但细根和粗根在不同气候条件下的腐殖质累积特征在很大程度上仍然是未知的. ...
Size-dependent morphological and chemical property of fine root litter decomposition 1 2015
... 由于提取方法的限制, 目前腐殖质的研究主要是胡敏酸和富里酸的研究(Lehmann & Kleber, 2015).已有研究表明, 由于胡敏酸溶于碱而富里酸溶于酸的性质, 导致腐殖质累积受土壤环境、季节变化、基质品质和微生物等综合因素的影响(Prescott et al., 2000; Ponge & Chevalier, 2006).在同一气候条件下, 根系分解主要受自身化学性质影响(Silver & Miya, 2001).不同物种、不同直径的根系化学组分差异显著(靳贝贝和国庆喜, 2013; Makita et al., 2015).随着林木根系直径的增加, 养分浓度(如N、P含量)降低, 导致不同径级根系分解速率不同(Silver & Miya, 2001; Ludovici & Kress, 2006), 这可能影响后续腐殖化进程.细根分解产物是一种潜在的碳源, 而粗根分解主要取决于气候变化, 尤其是温度的影响(Zhang & Wang, 2015).在高寒地区, 碳周转较慢, 且季节性雪被和冻融作用对根系分解有重要影响(Henry, 2008), 这可能影响植物不同直径根系腐殖化过程.虽然一些研究已经表明, 细根和粗根形态、养分含量、功能和分解机制显著不同(Guo et al., 2004; Aulen et al., 2011; Goebel et al., 2011; Olajuyigbe et al., 2012), 但细根和粗根在不同气候条件下的腐殖质累积特征在很大程度上仍然是未知的. ...
Nitrogen and lignin control of hardwood leaf litter decomposition dynamics 1982
雪被斑块对川西亚高山森林6种凋落叶冬季腐殖化的影响 2014a
Accelerated foliar litter humification in forest gaps: Dual feedbacks of carbon sequestration during winter and the growing season in an alpine forest 2 2015
... 川西亚高山森林地处青藏高原东缘, 在调节区域气候、保持水土和保育生物多样性等方面具有重要的作用(Yang et al., 2005; 邓仁菊等, 2009).常年的低温和频繁的地质活动, 造成川西亚高山森林土壤发育缓慢(Yang et al., 2005).已有研究表明, 川西亚高山植物根系分布主要集中于表层土壤(胡建利等, 2009), 该层土壤是有机质形成和腐殖化最为活跃的层次.因此, 根系腐殖化对该区域森林土壤有机质形成具有重要意义.前期研究表明, 冬季雪被显著影响凋落叶腐殖质累积(倪祥银等, 2014b), 且这种影响在冬季和生长季节明显不同(Ni et al., 2015), 但并未关注根系腐殖化过程中不同根系直径的胡敏酸和富里酸累积的变化格局.因此, 本研究在前期研究基础上, 初步研究了川西亚高山森林两种优势林木粗枝云杉(Picea asperata)和岷江冷杉(Abies faxoniana)不同径级根系(0-2、2-5和5-10 mm)在两年腐殖化过程中胡敏酸和富里酸在冬季和生长季节的累积特征, 为进一步了解川西亚高山森林生态系统地下碳循环过程提供基础数据. ...
... 式中, CHC、CHA和CFA分别为腐殖质、胡敏酸和富里酸含量(倪祥银等, 2014b; Ni et al., 2015); 以胡敏酸、富里酸含量与每次采样测得根系质量残留量的乘积表示胡敏酸、富里酸累积量, 以两次采样日期的胡敏酸、富里酸累积量之差表征该时期胡敏酸、富里酸净累积量.以各采样日期胡敏酸/富里酸表征胡敏酸、富里酸的相对含量和形成速度(Abakumov et al., 2013). ...
雪被斑块对高山森林凋落叶腐殖化过程中胡敏酸和富里酸累积的影响 2014b
The measurement of woody root decomposition using two methodologies in a Sitka spruce forest ecosystem 1 2012
... 由于提取方法的限制, 目前腐殖质的研究主要是胡敏酸和富里酸的研究(Lehmann & Kleber, 2015).已有研究表明, 由于胡敏酸溶于碱而富里酸溶于酸的性质, 导致腐殖质累积受土壤环境、季节变化、基质品质和微生物等综合因素的影响(Prescott et al., 2000; Ponge & Chevalier, 2006).在同一气候条件下, 根系分解主要受自身化学性质影响(Silver & Miya, 2001).不同物种、不同直径的根系化学组分差异显著(靳贝贝和国庆喜, 2013; Makita et al., 2015).随着林木根系直径的增加, 养分浓度(如N、P含量)降低, 导致不同径级根系分解速率不同(Silver & Miya, 2001; Ludovici & Kress, 2006), 这可能影响后续腐殖化进程.细根分解产物是一种潜在的碳源, 而粗根分解主要取决于气候变化, 尤其是温度的影响(Zhang & Wang, 2015).在高寒地区, 碳周转较慢, 且季节性雪被和冻融作用对根系分解有重要影响(Henry, 2008), 这可能影响植物不同直径根系腐殖化过程.虽然一些研究已经表明, 细根和粗根形态、养分含量、功能和分解机制显著不同(Guo et al., 2004; Aulen et al., 2011; Goebel et al., 2011; Olajuyigbe et al., 2012), 但细根和粗根在不同气候条件下的腐殖质累积特征在很大程度上仍然是未知的. ...
Humus index as an indicator of forest stand and soil properties 2 2006
... 由于提取方法的限制, 目前腐殖质的研究主要是胡敏酸和富里酸的研究(Lehmann & Kleber, 2015).已有研究表明, 由于胡敏酸溶于碱而富里酸溶于酸的性质, 导致腐殖质累积受土壤环境、季节变化、基质品质和微生物等综合因素的影响(Prescott et al., 2000; Ponge & Chevalier, 2006).在同一气候条件下, 根系分解主要受自身化学性质影响(Silver & Miya, 2001).不同物种、不同直径的根系化学组分差异显著(靳贝贝和国庆喜, 2013; Makita et al., 2015).随着林木根系直径的增加, 养分浓度(如N、P含量)降低, 导致不同径级根系分解速率不同(Silver & Miya, 2001; Ludovici & Kress, 2006), 这可能影响后续腐殖化进程.细根分解产物是一种潜在的碳源, 而粗根分解主要取决于气候变化, 尤其是温度的影响(Zhang & Wang, 2015).在高寒地区, 碳周转较慢, 且季节性雪被和冻融作用对根系分解有重要影响(Henry, 2008), 这可能影响植物不同直径根系腐殖化过程.虽然一些研究已经表明, 细根和粗根形态、养分含量、功能和分解机制显著不同(Guo et al., 2004; Aulen et al., 2011; Goebel et al., 2011; Olajuyigbe et al., 2012), 但细根和粗根在不同气候条件下的腐殖质累积特征在很大程度上仍然是未知的. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT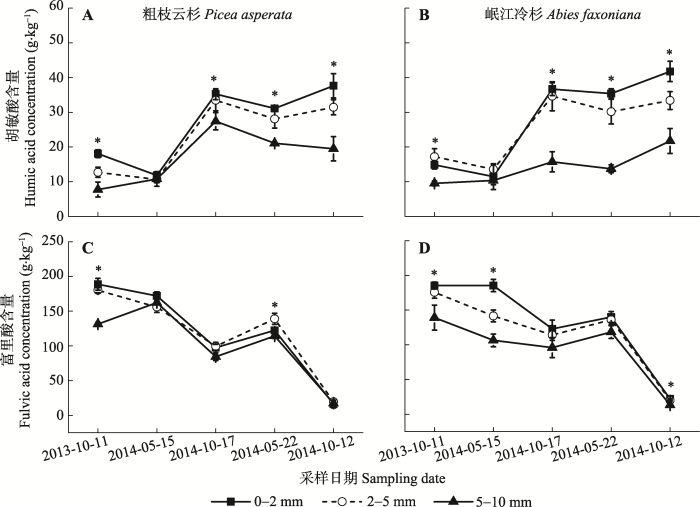 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT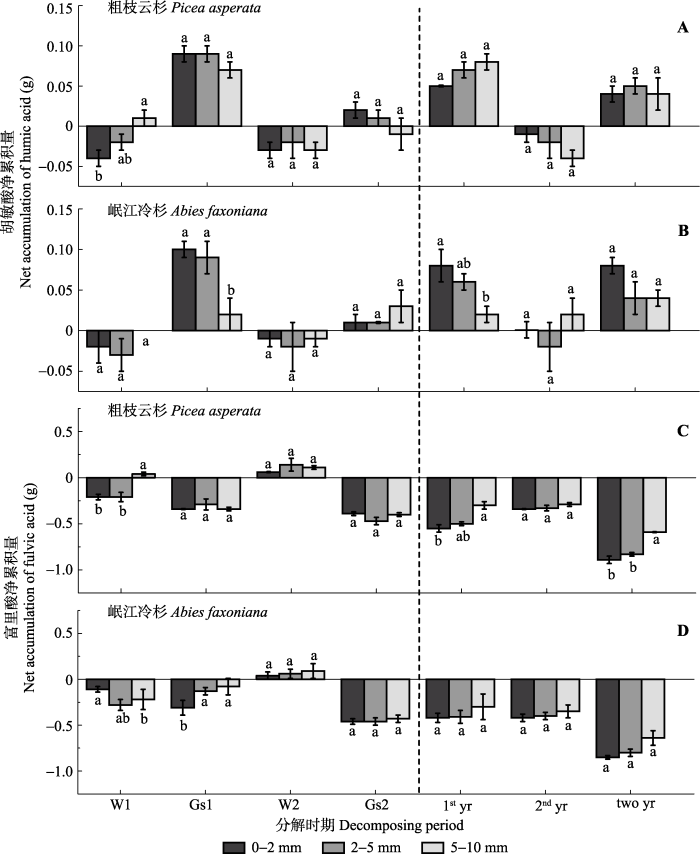 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT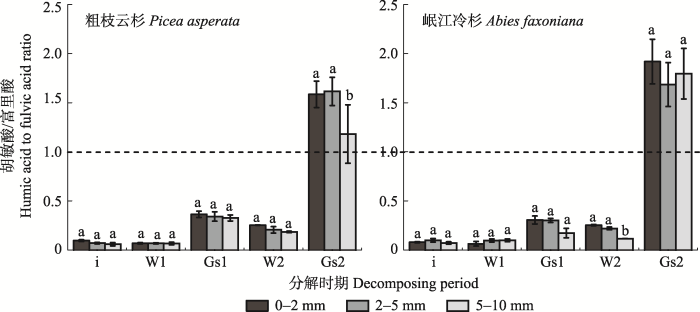 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}