 ,*西南民族大学青藏高原研究院, 四川成都 610041
,*西南民族大学青藏高原研究院, 四川成都 610041Effects of nitrogen addition on root dynamics in an alpine meadow, Northwestern Sichuan
ZI Hong-Biao, CHEN Yan, HU Lei, WANG Chang-Ting,*Institute of Qinghai-Tibetan Plateau Research, Southwest Minzu University, Chengdu 610041, China通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (4459KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
字洪标, 陈焱, 胡雷, 王长庭. 氮肥添加对川西北高寒草甸植物群落根系动态的影响. 植物生态学报[J], 2018, 42(1): 38-49 doi:10.17521/cjpe.2017.0255
ZI Hong-Biao.
氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(Han et al., 2005; 宾振钧等, 2014)。近年来, 随着化石燃料的大量燃烧和化肥的过度使用, 陆地生态系统中N元素摄入迅速增加(Galloway et al., 2004; Wang et al., 2015), 从而刺激植物生长, 增加了植物生物量和土壤碳(C)库的固存(Pregitzer et al., 2008; Reay et al., 2008; Thomas et al., 2010)。而植物根系在陆地C循环中扮演着关键的角色, 它们不仅从土壤中吸收水分和养分用于植物生产, 而且通过根系呼吸和死亡后分解进行C排放(Li et al., 2015)。另外, N元素摄入增加会通过影响根系形态(如根长和直径)、生物量和C循环相关的功能(如根系呼吸)来影响植物的根系(Nadelhoffer et al., 2000; Rasse, 2002)。因此, 探讨N添加对根系特征的影响将会更好地解释N素增加对生态系统C循环影响的潜在机制。
N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(Galloway et al., 2004)。而土壤N的有效性在植物根系动态特征中起着重要的作用(Vogt et al., 1995)。已有研究表明, N添加会降低森林生态系统的细根生物量(Nadelhoffer et al., 2000; Hendricks et al., 2006), 缩短(Pregitzer et al., 1995)或延长(Vogt et al., 1986)根系寿命, 降低根系死亡率, 提高根系生产量(Majdi & Andersson, 2005), 增强根系吸收水分和养分的能力(Gaudinski et al., 2001)。然而, 草地生态系统中草本植物是否具有相似变化机制的研究依然缺乏(Bai et al., 2008)。Bai等(2008)研究发现, N添加会降低0-10 cm和20-30 cm土层的根系寿命, 但是10-20 cm土层根系寿命并未受到影响。Martin和Chambers (2002)研究发现, N肥显著增加河岸边草甸的地下生物量。Rytter和Rytter (2012)研究发现营养供给受限时, 根密度高于原来的3倍, 施N减少了根系活力和入土深度, 但对根系组织和根系分解没有影响。然而, 青藏高原高寒草甸植物根系生产、死亡等的动态特征对于N肥添加的响应依然不清楚。
青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区。因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(MacDonald et al., 2002), 成为观测研究N沉降对生态系统影响的理想区域。一系列N添加实验结果表明, N添加促进植物生长(Song et al., 2012), 增加植物群落的初级生产力(Ren et al., 2010; 张杰琦等, 2010; 杨晓霞等, 2014)和叶片N含量(宾振钧等, 2014), 降低植物群落的物种丰富度和多样性(Wang et al., 2010)。但N添加对于陆地生态系统地下部分(尤其是根系)的影响了解甚少。为了分析青藏高原高寒草甸植物群落根系动态、空间分布格局和C分配特征对N添加的响应, 本研究采用微根管法对不同N添加梯度下高寒草甸植物群落根系现存量、生产量、死亡量和周转率进行研究, 旨在验证: (1) N添加提高根系生产量, 而降低根系死亡量, 从而提高根系周转率; (2) N添加增加了土壤速效养分, 从而提高了深层根系生产量。
1 材料和方法
1.1 研究区域概况
研究区位于四川省阿坝藏族羌族自治州红原县, 地理坐标32.83° N, 102.60° E, 海拔3 494 m。该区属于大陆性高原寒温带半湿润季风气候, 日温差大, 霜冻期长, 四季变化不明显, 年平均气温1.1 ℃, 最冷月(1月)平均气温为-10.3 ℃, 最热月(7月)平均气温为10.9 ℃, 平均5 ℃以下的月份长达7个月。干湿季节分明, 年降水量650-800 mm, 主要集中在5-9月; 日照时间长, 太阳辐射强, 年日照时间2 417.9 h, 年平均相对湿度71%。草甸类型主要是矮嵩草草甸, 植被盖度80%以上。其中四川嵩草(Kobresia setchwanensis)和高山嵩草(Kobresia pygmaea)是主要的莎草科植物, 三颖披碱草(Elymus nutans)和剪股颖(Agrostis matsumurae)是主要的禾本科植物, 而杂类草主要是匙叶银莲花(Anemone trullifolia)、蕨麻(Potentilla anserina)、钝苞雪莲(Saussurea nigrescens)、美丽龙胆(Gentiana formosa)和高山唐松草(Thalictrum alpinum)占优。该地区土壤类型是亚高山草甸土, 其土层深度达40 cm以上, 表层(0-10 cm)土壤SOM含量110.92 g·kg-1, TN含量6.38 g·kg-1, AN含量343.00 mg·kg-1, pH值为5.74 (表1)。该研究区地处青藏高原东缘, 是青藏高原的重要组成部分, 已有研究证实青藏高原大部分草地受到N限制(Yang et al., 2011)。早在20世纪90代初期就有****在青藏高原地区开始施肥试验, 我们综合前人研究发现, 施加100 kg·hm-2尿素时对地下生物量无显著影响(杨晓霞等, 2014), 但是施加200-300 kg·hm-2时高寒草甸群落生物量最高(王长庭等, 2013)。因此, 我们以此为依据进行N添加梯度的试验设计来研究高寒草甸植物群落根系动态特征对N添加的响应。Table 1
表1
表1氮添加梯度对寒草甸土壤养分的影响(平均值±标准偏差, n = 6)
Table 1
| 土层 Soil layer (cm) | 处理 Treatment | pH值 pH value | 有机质 SOM (g·kg-1) | 速效氮 AN (mg·kg-1) | 全氮 TN (g·kg-1) |
|---|---|---|---|---|---|
| 0-10 cm | CK | 5.74 ± 0.10Aa | 110.92 ± 10.48Aa | 343.00 ± 9.40Ac | 6.38 ± 0.84Aa |
| N10 | 5.42 ± 0.14Bb | 119.61 ± 19.21Aa | 323.40 ± 9.96Ad | 7.01 ± 0.81Aa | |
| N20 | 5.50 ± 0.25Ab | 121.68 ± 25.26Aa | 392.00 ± 2.46Ab | 6.95 ± 0.70Aa | |
| N30 | 5.44 ± 0.08Ab | 124.58 ± 11.78Aa | 418.95 ± 9.15Aa | 7.27 ± 0.97Aa | |
| 10-20 cm | CK | 5.73 ± 0.08Aa | 94.60 ± 11.23Aa | 290.33 ± 7.99Bb | 5.34 ± 0.93Aa |
| N10 | 5.69 ± 0.27Aa | 88.10 ± 24.43Ba | 276.85 ± 8.20Bb | 5.20 ± 0.71Aa | |
| N20 | 5.62 ± 0.19Ab | 91.91 ± 17.75Ba | 328.30 ± 6.12Ba | 5.40 ± 0.83Aa | |
| N30 | 5.48 ± 0.16Ab | 95.30 ± 16.64Ba | 316.05 ± 4.85Ba | 5.64 ± 0.85Aa |
新窗口打开|下载CSV
1.2 试验设计
1.2.1 试验样地设置于2012年5月下旬在研究区内选择地势相对平坦, 植被分布均匀, 面积为100 m × 100 m的未退化矮嵩草草甸, 用围栏进行保护, 作为试验样地。同时把样地分成6个30 m × 20 m的实验样区, 4个角用木桩进行标记, 样区间距为3 m。每个样区内设置4个3 m × 3 m小样方, 共计24 (4 × 6)个小样方, 小样方之间间隔不小于2 m作为缓冲区。采用随机区组试验设计把0 (CK)、10 (N10)、20 (N20)、30 g·m-2 (N30) 4个N添加水平随机安排在小区内。所施肥为尿素(CO(NH2)2, 46% N)。并于每年5月下旬选择多云的阴天, 用手将N肥均匀地撒在小区内, 每年施肥1次。
于2013年5月在上述的每个样方内安装一根聚碳酸酯材料微根管, 微根管长度为100 cm, 内径 5 cm, 外径6 cm。首先用特制的土钻在样方内钻取一个与地面呈30°夹角, 深度为70-75 cm, 直径为 7 cm的微根管洞, 然后把微根管密封的一头缓慢插入孔中。露出地面的微根管用黑色棉布包裹, 避免光线射入影响根系生长, 同时用海绵和胶带缠绕, 避免微根管因撞击而受损, 顶端盖上不透光的黑盖子。此外, 在微根管周围用钻出的土填平, 使微根管与土壤紧密接触, 同时尽量减少对微根管周围的土壤扰动, 在不读取数据时用塑料袋包裹并用胶带缠紧以减少微根管的热传导。
1.2.2 样品采集
1.2.2.1 土壤样品采集与测定 于2015年8月中旬, 在3 m × 3 m样方内, 用内径5 cm的土钻分层(0-10 cm和10-20 cm)按“S”形钻取5钻, 混合为1个土壤样品, 每个处理重复6次, 并做好标记。样品风干后过筛用于测定土壤pH值、SOM、TN和AN含量。其中, 用电极法测定土壤pH值, 丘林法测定SOM含量, 凯氏定氮法测定TN含量, 康惠法测定AN含量(中国科学院南京土壤研究所, 1983)。
1.2.2.2 根系生物量采集 于2015年8月中旬在3 m × 3 m样方内, 用内径 5 cm的土钻分层(0-10 cm和10-20 cm)取根系样品, 每层取3钻混合为1个样品, 每个处理6次重复。把样品带回实验室过筛去掉土壤, 并用清水冲洗得到根系样品, 并测量所获得根系的长度, 装入信封放入烘箱在65 ℃烘干至恒质量(48 h), 称质量后计算比根长。
1.2.2.3 根系图片采集和分析 于2015年5-9月对根系进行连续取样, 首先将微根管外包裹的塑料袋打开, 打开顶端盖子, 用棉布将微根管内的水蒸气擦干, 等待10 min后, 待微根管内外温度相同时, 将标定好的摄像机放入微根管内, 并在微根管上标记好第一次摄像头放入的位置, 同时采用CI-600根系扫描仪(CID Bio-Science, Camas, USA)进行图像采集。每个微根管采集两个斜坡深, 分别为0-20 cm和20-40 cm, 根据根管与地面的夹角把深度换算成0-10 cm和10-20 cm垂直土层深度(D), 每次测定结束后, 将露出地面的根管按第一次的方法包裹好, 每2周采集一次图像, 共取10次图像, 总共追踪标记到12β219条根系。
将CI-600扫描的图像带回实验室用WinRHZIO Tron MF (CID Bio-Science, Camas, USA)软件进行分析处理, 测定每张图像中根系的直径、长度等各形态特征指标的数据, 并通过图像中根系的颜色来区分活根和死根, 其中白色和褐色的根定义为活根, 黑色和消失的根定义为死根。全部活根和死根根系的图像和数据采集都必须完整保存。
根系的估算: 首先通过根系图像分析得到根系的长度, 并利用如下的公式计算单位体积的根长密度(RLD, m·m-3):
RLDv = L/(A × DOF)
式中, L (m)为微根窗中观察到的细根根长; A (m2)为观测窗面积(图像大小); DOF (m)为微根管到周围土壤的距离, DOF一般为0.002-0.003 m (Taylor et al., 1970; Sanders & Brown, 1978)。由于本研究中的草地群落的细根平均直径为0.254 mm, 计算中DOF取0.002 m (吴伊波等, 2010)。
其次, 通过比根长(SRL, m·g-1)将RLDV转化为单位体积的生物量(RBD, g·m-3)。
RBD = RLDv/SRL
式中, SRL为土钻法所得每克根(直径<1 mm)生物量的根长(m·g-1)。
最后, 通过单位体积生物量(RDB)乘以取样土壤剖面深度(D)将RDB转换成单位面积根系生物量(RLDA, g·cm-2):
RLDA = RBD/D
根系周转运用上述介绍的方法, 计算出单位面积根系生物量(RLDA), 在每次微根管取样时能够作为细根现存量的估计值, 每次测定的细根生产量累加来估算年生物量产量(Norby et al., 2004; Majdi & Andersson, 2005):
T = P/Y
式中, T为根系周转率(a-1); P为细根年生物量产量; Y为活细根现存量平均根系生物量密度。
根系生产量是在t时不存在而在t + 1时所增加的新鲜根长总和, 再加上所有在t时原有根系上延长的长度来估算。根系死亡量是t时所有活根和新鲜根在t + 1时死亡和消失掉的根长总和, 根系现存量根据季节性生产和死亡的差异来计算(Majdi & Andersson, 2005), 根系生产、死亡和现存量都是用单位面积根系生物量(RLDA)计算得出。
1.3 统计分析
采用SPSS 19.0对土壤理化性质(土壤pH值, SOM、TN和AN含量)、根系特征(根系现存量、生产量、死亡量和周转率)进行单因素方差分析, 用最小显著极差法(LDS)进行不同处理之间的显著性检验(p < 0.05)。不同土层(0-10 cm和10-20 cm)之间的差异则采用独立样本t检验。采用重复测量方差方法分析取样时间、施肥梯度及交互作用对根系现存量、生产量和死亡量动态特征的影响; 用Turkey HSD法进行施肥梯度间的多重比较。此外, 采用Pearson相关性分析0-10 cm和10-20 cm土层土壤理化性质和根系累积生产量、死亡量、现存量和年周转率(n = 6)的相关性(p = 0.05)。所有图表绘制均在Origin 8.5中 进行。2 结果
2.1 土壤理化性质变化
在0-10 cm土层, 土壤pH值在N添加后显著降低; AN的含量在N10处理显著降低, 但N20和N30处理显著增加; SOM和TN含量对N添加无显著响应(表1)。在10-20 cm土层, 土壤pH值在N20和N30处理下显著降低, AN则显著提高(p < 0.05); SOM和TN含量对N添加则无显著响应(表1)。此外, 土壤pH值在N10处理0-10 cm土层显著小于10-20 cm, 其他处理均无显著变化; SOM、AN和TN含量则随土层深度的增加而降低(表1)。2.2 根系动态变化
重复测量方差分析表明, 在0-10 cm和10-20 cm土层, 取样时间和N添加梯度均显著影响根系现存量(p < 0.05), 但它们的交互作用对根系现存量无显著影响(p > 0.05; 表2)。在0-10 cm土层, N10和N20处理仅在生长初期(5月份)显著降低根系现存量, 但是N30处理在整个生长季均显著降低根系现存量 (p < 0.05; 图1A)。在10-20 cm土层, N添加对生长初期(5月份)根系现存量无显著影响(p > 0.05), 但是6月到9月根系现存量则表现出CK显著大于N添加处理(p < 0.05; 图1B)。此外, 根系现存量随土层深度增加而显著降低(图1)。图1
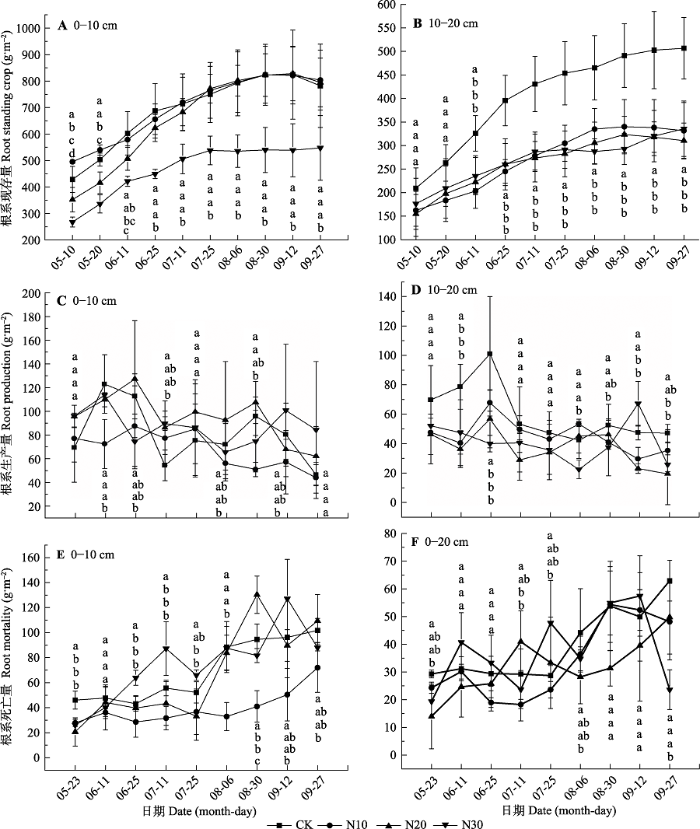 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同氮添加梯度根系的季节动态(平均值±标准偏差, n = 6)。不同小写字母表示不同处理间差异显著(p < 0.05)。CK、N10、N20、N30同
Fig. 1Seasonal patterns of root for different nitrogen addition gradients (mean ± SD, n = 6). Different lowercase letters indicate significant differences in different treatment (p < 0.05). CK, N10, N20, N30, see
Table 2
表2
表2不同N添加梯度根系动态的重复测量方差分析(n = 6)
Table 2
| 土层 Soil layer | 根系指标 Root index | 取样时间 Sampling time | N添加梯度 N addition gradient | 取样时间×N添加梯度 Sampling time × N addition gradient | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| 0-10 cm | 根系现存量 Root standing crop | 81.262 | < 0.001 | 7.468 | 0.010 | 1.579 | 0.239 |
| 根系生产量 Root production | 63.809 | < 0.001 | 1.078 | 0.412 | 9.53 | < 0.001 | |
| 根系死亡量 Root mortality | 131.752 | < 0.001 | 4.477 | 0.04 | 9.693 | 0.001 | |
| 10-20 cm | 根系现存量 Root standing crop | 104.472 | < 0.001 | 7.793 | 0.009 | 3.918 | 0.250 |
| 根系生产量 Root production | 60.200 | < 0.001 | 3.460 | 0.071 | 12.932 | < 0.001 | |
| 根系死亡量 Root mortality | 138.540 | < 0.001 | 0.482 | 0.704 | 14.224 | < 0.001 | |
新窗口打开|下载CSV
根系生产量主要受到取样时间以及取样时间与N添加梯度交互作用的显著影响, 但N添加梯度对根系生产量无显著影响(表2)。在0-10 cm土层, 根系生产量仅在观测初期(6月11日)表现为N10处理显著低于CK, 其余生长期N添加对根系生产量均无显著影响(p > 0.05; 图1C)。10-20 cm土层, N添加对5月份和7月份根系生产量无显著影响(p > 0.05), 但N添加显著降低了6月份和9月底的根系生产量, 而8月份N30处理显著降低了根系生产量(p < 0.05; 图1D)。
根系死亡量在0-10 cm土层受到取样时间和N添加梯度及二者交互作用的影响, 但是10-20 cm土层主要受到取样时间及二者交互作用的影响(表2)。在0-10 cm土层, N添加显著降低生长初期(5月份)的根系死亡量, 生长中期(6月初到7月底) N30处理显著提高根系死亡量, 生长后期(8月初到9月底) N10处理显著降低根系死亡量(p < 0.05, 图1E)。在10- 20 cm土层, 生长初期(5月份) N20处理显著提高根系死亡量, 生长季末期(9月底) N30处理显著降低根系死亡量(p < 0.05), 其他时期添加对根系死亡量无显著影响(图1F)。
2.3 累积根系生产量和死亡量、平均现存量和周转率的变化
对N添加梯度和土层深度进行双因素分析发现, 施肥梯度显著影响累积根系死亡量, 土层深度显著影响累积根系生产量、平均根系现存量和周转率, 但是二者的交互作用对根系特征无显著影响(表3)。在0-10 cm土层, 累积根系生产量和平均根系现存量对N添加无显著响应; 根系死亡量在N10处理 显著降低; 根系周转率表现为N30 (1.48) > N20 (1.38) > CK (1.26) > N10 (1.17), 而且在N10处理显著降低, N30处理显著提高(图2)。在10-20 cm土层, N添加显著降低了累积根系生产量和平均根系现存量, 但对根系死亡量无显著影响; 根系周转率变化较大且表现为N20 (1.77) > N10 (1.44) > N30 (1.24) > CK (1.21), 但各处理间均无显著差异(图2)。此外, 累积根系生产量和平均现存量均表现为0-10 cm显著高于10-20 cm土层; 累积根系死亡量在N10处理不同土层间无显著变化, 其余处理均表现为0- 10 cm土层显著高于10-20 cm土层; 根系周转率除N10处理10-20 cm土层显著高于0-10 cm土层外, 其余处理均无显著差异(图2)。图2
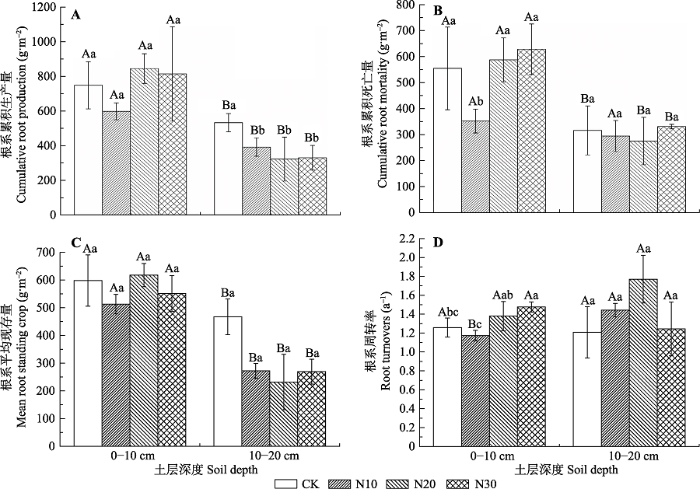 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同氮添加梯度根系年生产量和死亡量、平均现存量和周转率的变化(平均值±标准差, n = 6)。不同小写字母表示不同处理间差异显著(p < 0.05); 不同大写字母表示不同土层间差异显著(p < 0.05)。CK、N10、N20、N30同
Fig. 2Cumulative length production, cumulative length mortality, mean root standing crop and Annual root turnovers over the growing season different nitrogen addition gradients (mean ± SD, n = 6). Different lowercase letters indicate significant differences in different treatment (p < 0.05). Different capital letters indicate significant different in different soil layers (p < 0.05). CK, N10, N20, N30, see
Table 3
表3
表3不同N添加梯度和土层深度对根系特征的方差分析(n = 6)
Table 3
| 因子 Factor | 处理 Treatment | 深度 Depth | 处理×深度 Treatment × Depth | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| 累积根系生产量 Cumulative root production | 1.328 | 0.300 | 47.14 | < 0.001 | 2.661 | 0.083 |
| 平均根系现存量 Mean root standing crop | 1.770 | 0.190 | 29.93 | < 0.001 | 1.237 | 0.329 |
| 根系周转 Root turnovers | 1.655 | 0.220 | 0.689 | 0.420 | 1.573 | 0.235 |
| 累积根系死亡量 Cumulative root mortality | 3.280 | 0.050 | 38.11 | < 0.001 | 2.538 | 0.093 |
新窗口打开|下载CSV
2.4 土壤理化性质与根系特征间的相关性
在0-10 cm土层, 土壤pH值和SOM含量与根系现存量、死亡量、生产量和周转率均无显著相关性, 但是AN含量与根系生产量呈显著负相关关系, 与死亡量和周转率呈显著正相关关系; TN含量与根系现存量和周转率呈显著正相关关系, 与根系死亡量和周转率无显著相关性。在10-20 cm土层, 除根系现存量与SOM和AN含量呈显著负相关关系, 与TN呈显著正相关关系外, 其余根系指标与土壤理化性质均无显著相关性(表4)。Table 4
表4
表4土壤理化性质与根系特征的相关性(n = 6)
Table 4
| 土层 Soil layer | 项目名称 Item | pH值 pH value | 有机质 SOM (g·kg-1) | 速效氮 AN (mg·kg-1) | 全氮 TN (mg·kg-1) |
|---|---|---|---|---|---|
| 0-10 cm | 根系现存量 Root standing crop | -0.27 | -0.414 | 0.487 | 0.622* |
| 根系生产量 Root production | 0.280 | -0.421 | -0.843** | -0.167 | |
| 根系死亡量 Root mortality | 0.315 | -0.044 | 0.716** | 0.021 | |
| 根系周转率 Root turnover | -0.459 | -0.225 | 0.673* | 0.691* | |
| 10-20 cm | 根系现存量 Root standing crop | -0.075 | -0.590* | -0.684* | 0.624* |
| 根系生产量 Root production | 0.371 | 0.005 | -0.413 | 0.095 | |
| 根系死亡量 Root mortality | -0.095 | -0.038 | -0.404 | 0.300 | |
| 根系周转率 Root turnover | -0.193 | -0.476 | 0.287 | 0.293 |
新窗口打开|下载CSV
3 讨论
3.1 不同N添加梯度下土壤理化性质的变化
青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (彭琴等, 2008), 据报道,该地区N限制的草地面积已达5.17 × 105 km2, 占该地区草地总面积的34% (Yang et al., 2008), 而且牧草生产主要受N限制, 但是施N肥会造成土壤酸化(Chen & Bai, 2013; Wei et al., 2014), 本研究结果同样证实这一观点, 即在N20和N30处理下土壤pH值显著降低。一方面, N添加促进了植物生长, 积累了大量有机物质, 增加了表层SOM的含量(表1), 而有机物质的大量累积会产生有机酸, 导致土壤pH值的下降(Cammeraat & Risch, 2008); 另一方面, 由于土壤微生物的硝化作用产生NH4+和H+, 从而增加了土壤酸度(Birkhofer et al., 2008; Luo et al., 2015)。此外, 青藏高原高寒草甸地处高海拔地区, 虽然土壤中储藏着大量SOM, 但是寒冷气候条件导致土壤矿化速率较低, 而草地施肥有效地解除了土壤速效养分的限制(孔彬彬等, 2016), 使得土壤速效养分(速效N)含量显著提高(如N20和N30 (表1))。土壤TN含量并未随着N肥添加而显著改变, 说明TN含量相对稳定, 其含量变化需要一个长期扰动, 短期施肥(施肥期3年)不足以引起TN含量改变。3.2 不同N添加梯度下根系动态的变化
根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(Bessler et al., 2009), 这些功能的稳定发挥主要取决于根系的分布格局和现存量(Kurz et al., 2006)。而根系的分布格局和现存量又会受到外界环境因素(增温、降雨和N沉降等)的影响(Majdi & Ohrvik, 2004; Wan et al., 2004; Bai et al., 2010)。本研究发现, N添加显著降低深层根系现存量。这一结果与梅莉等(2008)的研究结果相类似, 即N添加降低根系现存量。其主要原因是N添加提高了N的有效性(表1), 而随N的有效性的提高, 分配给细根的碳的相对比例下降, 从而降低根系生物量(Nadelhoffer et al., 2000; Hendricks et al., 2006)。相关性分析结果也同样证实, 根系现存量与N的有效性呈负相关关系(表3)。另外, N添加会显著降低土壤pH值(表1), 从而改变生产木质素降解酶的真菌环境, 导致相关酶活性降低, 抑制根系生长(涂利华等, 2014)。根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面。先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (Jackson et al., 1997), 是理解陆地生态系统C和养分循环必不可少的指标(Woodward & Osborne, 2000)。N添加对表层根系生产量无显著影响, 但是显著降低深层根系生产量。这一结果与Majdi和Ohrvik (2004)的研究结果相似。因为N添加显著提高了土壤中速效养分含量, 缓解了高寒草甸植物的养分限制, 促进了植物地上部分生长, 植物将会拥有更多的地上生物量分配, 从而减缓地下根系生长(Bai et al., 2008; 杨晓霞等, 2014)。另外, 养分资源丰富的环境中, 根系分布空间相对较浅(熊淑萍等, 2011), 从而导致深层根系生产量显著低于CK。根系死亡量仅在N10处理的表层显著降低, 其他处理均无显著变化。这是由于土壤可利用养分含量差异所致, 在低养分含量的环境中植物通过降低根系死亡来储存其所需要的营养物质(van der Krift & Berendse 2002), 但提高土壤养分资源, 植物将会分配更多C在地上部分, 降低根系C的累积从而缩短根系寿命(Bai et al., 2008)。本研究我们发现N10处理降低土壤AN含量, 而N20和N30处理显著提高AN含量(表1), 而且根系死亡量与AN含量呈显著正相关关系(表2)。因此, 根系死亡量显著降低。
3.3 不同N添加梯度下根系年周转率的变化
根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(Gill & Jackson, 2000)。根据已有的报道, 随着土壤可利用N的增加, 根系周转率主要表现为提高或降低(郭大立和范萍萍, 2007)。我们的研究表明, 随N添加, 根系周转率均呈增加趋势, 这与Li等(2015)通过Meta分析得到的结果相一致。主要机理是: 1) N添加增加N的有效性, 导致根系代谢活性增加(Gill & Jackson, 2000)、呼吸加快(Li et al., 2015), 从而缩短了植物群落根系寿命(Eissenstat et al., 2000; Bai et al., 2008), 而根系寿命决定细根周转, 细根寿命越短, 周转越快(Schoettle & Fahey, 1994; Pregitzer et al., 1995); 2)菌根能够延长根系寿命, 降低根系的死亡率和周转率(Tierney & Fahey, 2001), 但是N添加降低菌根真菌侵染率(Li et al., 2015), 从而降低了菌根抵御病原菌、土壤动物对细根侵害的能力(Linderman, 1994; Eissenstat et al., 2000), 导致根系寿命降低, 加快根系周转。同时, 在陆地生态系统中细根年周转率的范围在0.019-2.644, 平均年周转率为0.56 (Jackson et al., 1997; Gill & Jackson, 2000)。本研究中细根年周转率在1.0-1.8, 施N后根系周转率明显提高, 这种相对较高的细根周转率说明N添加促进高寒草甸植物生命活动, 使它们可以不断产生新的细根以代替旧的细根来吸收水分和营养物质, 增加根系对水分和营养物质利用效率(赵学春等, 2014)。同时, 周转率的提高会导致根系生产消耗的C增加, 从而改变了土壤C库。另外, 细根的周转可以将大量营养物质和养分带进土壤, 促进根系微生物活动, 改善土壤的理化性质(裴智琴等, 2011)。因此, 根系周转可能是控制土壤养分有效性的一个重要指标。3.4 N添加梯度和土层深度对根系特征的影响
许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(张杰琦等, 2010), 而地上和地下部分是相互联系的, 在养分条件较好时, 植物倾向于将生物量较多地分配于地上部分, 在养分条件较差时则倾向于分配到地下部分(Müller et al., 2000)。本研究随施N量的增加AN含量增加, 但是根系特征对N添加梯度增加无显著响应。这可能是由于土壤中N浓度的增加, 导致可利用的P受到限制, 而N在青藏高原倾向于降低地下生物量, P则倾向于增加地下生物量(杨晓霞等, 2014)。因此, 不同N肥梯度间根系特征无显著差异。此外, 高寒草甸根系特征还受到土层深度的影响, 随着土层深度增加根系生产量显著降低。这是因为随土层深度增加, 土壤温度、含水量、有效养分和通气条件逐渐恶劣, 植物为了适应生态环境的变化采用减少根量的生存策略(周兴民, 2001)。同时, 土层深度增加会延长根系寿命(于水强等, 2007), 土层深度每增加1 cm, Vitis labruscana细根的死亡危险率降低0.3%-0.7% (Anderson et al., 2003)。4 结论
川西北高寒草甸N添加后显著提高土壤速效N含量, 降低土壤pH值, 进而影响根系生长动态。其中, N添加在土壤表层主要影响根系死亡量和周转率, 而在土壤深层主要影响根系现存量和生产量。同时,根系动态特征与土壤速效N含量显著相关。此外, N添加改变了高寒草甸植物根系的生物量分配, 促使根系向表层集中。而根系在空间层次上的差异主要是由土壤资源有效性差异造成, 而资源有效性的差异又会引起根系寿命的变化, 但是本研究并未涉及根系寿命对N添加梯度和土层深度变化的响应。因此, 在今后研究的中, 我们应该关注N添加对高寒草甸植物根系寿命的影响以及根系寿命与根系生产、周转和死亡之间的相互关系, 这将有助于我们更好地认识高寒草甸植物群落根系对N添加的响应机制。
扫码加入读者圈
听语音,看问答
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1046/j.1469-8137.2003.00757.xURL [本文引用: 1]

090004 Minirhizotron techniques were used to examine root lifespan in Vitis labruscana (Concord grape) for roots born in four different years that varied in rainfall.
[本文引用: 1]
DOI:10.1111/j.1365-2435.2008.01403.xURL [本文引用: 4]
1. To understand root dynamics of the native, perennial clonal grass Leymus chinensis in Inner Mongolia steppe, we examined spatial and temporal effects of N addition on root survivorship and longevity traits of L. chinensis by studying responses of survivorship of roots distributed in three soil layers (009000910, 1009000920 and 2009000930 cm) and born in three seasons (spring, summer and autumn) to N addition with modified rhizotron technique during the growing seasons of 200409000905. Kaplan090009Meier analysis was used to generate survival functions and to estimate mean root life spans. 2. Roots in the three soil layers displayed similar life spans with mean overall value of 81 days in the control plots. N addition reduced life span of roots in the soil layers of 009000910 cm and 2009000930 cm, but it had no effect on life span of roots in the soil layer of 1009000920 cm. 3. Roots born in the three seasons differed in their life spans with the life span values of 2109000958, 60090009153 and 101090009151 days for those roots born in spring, summer and autumn, respectively. N addition markedly reduced life span of roots born in summer in the three soil layers with the greatest reduction occurring in roots present in soil layer of 2009000930 cm, but it had marginal effect on life span of the spring-born and autumn-born roots. 4. The differential sensitivity of survivorship and longevity of L. chinensis roots occurred in different soil depths and born in different seasons to N addition highlights the importance of spatial and temporal characteristics of roots in response to soil N availability in Inner Mongolia steppe.
DOI:10.1890/08-0867.1URLPMID:19569367 [本文引用: 1]
We investigated effects of plant species richness in experimental grassland plots on annual above- and belowground biomass production estimated from repeated harvests and ingrowth cores, respectively. Aboveground and total biomass production increased with increasing plant species richness while belowground production remained constant. Root to shoot biomass production ratios (R/S) in mixtures were lower than expected from monoculture performance of the species present in the mixtures, showing that interactions among species led to reduced biomass partitioning to belowground organs. This change in partitioning to belowground organs was not confined to mixtures with legumes, but also measured in mixtures without legumes, and correlated with aboveground overyielding in mixtures. It is suggested that species-rich communities invest less in belowground biomass than do monocultures to extract soil resources, thus leading to increased investment into aboveground organs and overyielding.
DOI:10.3724/SP.J.1258.2014.00020URL [本文引用: 2]
以青藏高原高寒草甸为研究对象,通过人工氮肥添加试验,研究6个群落优势种在不同施氮(N)水平下叶片碳(C)、N、磷(P)元素含量的变化以及生态化学计量学特征.结果表明:自然条件下,6个物种叶片N、P质量浓度存在显著的差异,表现为:黄花棘豆(Oxytropis ochrocephala)最高,为24.5和2.51 g.kg-1,其叶片N含量低于而P含量高于我国其他草地的豆科植物;其余5个物种叶片N、P质量浓度分别为11.5-18.1和1.49-1.72 g·kg-1,嵩草(Kobresia myosuroides)叶片N含量最低,垂穗披碱草(Elymus nutans)叶片P含量最低,与我国其他区域的研究结果相比,其叶片N和P含量均低于我国其他草地非豆科植物.随氮素添加量的增大,6种群落优势种叶片的C和P含量保持不变;其他5种植物叶片N含量显著增加,黄花棘豆叶片N含量保持不变.未添加氮肥时,6种植物叶片N:P为7.3-11.2,说明该区植物生长更多地受N限制.随N添加量的增加,除黄花棘豆外,其他5种植物叶片N:P大于16,表现为植物生长受P限制.综合研究表明,青藏草原高寒草甸植物叶片N含量较低,植物受N影响显著,但不同物种对N的添加反应不同,豆科植物黄花棘豆叶片对N添加不敏感,其他5个物种叶片全N含量随着N添加量的升高而增加,该研究结果可为高寒草甸科学施肥提供理论依据.
DOI:10.3724/SP.J.1258.2014.00020URL [本文引用: 2]
以青藏高原高寒草甸为研究对象,通过人工氮肥添加试验,研究6个群落优势种在不同施氮(N)水平下叶片碳(C)、N、磷(P)元素含量的变化以及生态化学计量学特征.结果表明:自然条件下,6个物种叶片N、P质量浓度存在显著的差异,表现为:黄花棘豆(Oxytropis ochrocephala)最高,为24.5和2.51 g.kg-1,其叶片N含量低于而P含量高于我国其他草地的豆科植物;其余5个物种叶片N、P质量浓度分别为11.5-18.1和1.49-1.72 g·kg-1,嵩草(Kobresia myosuroides)叶片N含量最低,垂穗披碱草(Elymus nutans)叶片P含量最低,与我国其他区域的研究结果相比,其叶片N和P含量均低于我国其他草地非豆科植物.随氮素添加量的增大,6种群落优势种叶片的C和P含量保持不变;其他5种植物叶片N含量显著增加,黄花棘豆叶片N含量保持不变.未添加氮肥时,6种植物叶片N:P为7.3-11.2,说明该区植物生长更多地受N限制.随N添加量的增加,除黄花棘豆外,其他5种植物叶片N:P大于16,表现为植物生长受P限制.综合研究表明,青藏草原高寒草甸植物叶片N含量较低,植物受N影响显著,但不同物种对N的添加反应不同,豆科植物黄花棘豆叶片对N添加不敏感,其他5个物种叶片全N含量随着N添加量的升高而增加,该研究结果可为高寒草甸科学施肥提供理论依据.
DOI:10.1016/j.soilbio.2008.05.007URL [本文引用: 1]
Organic farming may contribute substantially to future agricultural production worldwide by improving soil quality and pest control, thereby reducing environmental impacts of conventional farming. We investigated in a comprehensive way soil chemical, as well as below and aboveground biological parameters of two organic and two conventional wheat farming systems that primarily differed in fertilization and weed management strategies. Contrast analyses identified management related differences between erbicide-free bioorganic (BIOORG) and biodynamic (BIODYN) systems and conventional systems with (CONFYM) or without manure (CONMIN) and herbicide application within a long-term agricultural experiment (DOK trial, Switzerland). Soil carbon content was significantly higher in systems receiving farmyard manure and concomitantly microbial biomass (fungi and bacteria) was increased. Microbial activity parameters, such as microbial basal respiration and nitrogen mineralization, showed an opposite pattern, suggesting that soil carbon in the conventional system (CONFYM) was more easily accessible to microorganisms than in organic systems. Bacterivorous nematodes and earthworms were most abundant in systems that received farmyard manure, which is in line with the responses of their potential food sources (microbes and organic matter). Mineral fertilizer application detrimentally affected enchytraeids and Diptera larvae, whereas aphids benefited. Spider abundance was favoured by organic management, most likely a response to increased prey availability from the belowground subsystem or increased weed coverage. In contrast to most soil-based, bottom-up controlled interactions, the twofold higher abundance of this generalist predator group in organic systems likely contributed to the significantly lower abundance of aboveground herbivore pests (aphids) in these systems. Long-term organic farming and the application of farmyard manure promoted soil quality, microbial biomass and fostered natural enemies and ecosystem engineers, suggesting enhanced nutrient cycling and pest control. Mineral fertilizers and herbicide application, in contrast, affected the potential for top-down control of aboveground pests negatively and reduced the organic carbon levels. Our study indicates that the use of synthetic fertilizers and herbicide application changes interactions within and between below and aboveground components, ultimately promoting negative environmental impacts of agriculture by reducing internal biological cycles and pest control. On the contrary, organic farming fosters microbial and faunal decomposers and this propagates into the aboveground system via generalist predators thereby increasing conservation biological control. However, grain and straw yields were 23% higher in systems receiving mineral fertilizers and herbicides reflecting the trade-off between productivity and environmental responsibility.
DOI:10.1111/j.1439-0418.2008.01281.xURL [本文引用: 1]
Abstract Soil dwelling ants are important soil engineers that have a large impact on the soil ecosystem. This is reflected in the alteration of soil properties by ants due to burrowing activities, the accumulation of organic matter and other nutrients in the soil, which, in turn, alters soil physical, chemical and (micro) biological processes. A review of recent literature on these fine scale soil changes resulting from ant activity is presented in this paper. However, ants do not only affect soil properties at the fine scale but also have effects at the hillslope and catchment scales with respect to surface hydrological processes and ecosystem functioning. At the finest scale, most studies showed changed soil textures, lowered bulk densities and increased infiltration where ant burrows or mineral mounds are prominent. Only a few studies investigated the effects of ants on soil properties such as soil structure and aggregate stability, which were improved by ant activity. Nutrients were generally considered to be concentrated in nests, and increased nutrient cycling involving microbial processes were reported. On a broader scale, the information on nest densities and patterns is crucial to understand the impacts of ants, but this information is scarcely available. It has been reported that the activity of ants increases the heterogeneity of soil infiltration rates and nutrient concentrations at the landscape scale, which in turn has consequences for the redistribution of sediments and nutrients by geomorphological and biotic processes. A hierarchical descriptive model focusing on dry land ecosystems is presented to incorporate small-scale ant activity into broader scale hillslope and catchment processes. However, this model has to be tested and more research is needed, especially on these broader scale aspects, to be able to fully understand the role of ants on the ecosystem and landscape scale.
DOI:10.1111/1365-2745.12119URL [本文引用: 1]
Anthropogenic acid deposition nduced soil acidification is one of the major threats to biodiversity, ecosystem functioning and services. Few studies, however, have explored in detail how above-ground changes in plant species richness and productivity resulting from soil acidification are mediated by effects on below-ground biota and soil properties.To increase our understanding of this linkage, we collected data on below- and above-ground communities and soil properties in a 3-year field experiment with seven levels of acid addition rate to build-up broad intensities of soil acidification in the semi-arid Inner Mongolian grassland.Acid addition directly elevated concentrations of soil Al3+ ions, decreased the base cations Ca2+, Mg2+ and Na+, and increased soil moisture and available phosphorus. Acid addition also appears to have altered the soil microbial community via changes in H+ and Al3+ ions and altered the nematode community via changes in H+ ions and soil moisture.The observed changes in soil N availability (i.e. net N mineralization, -N and -N) could be explained by mediating changes in the H+ and Al3+ ions, microbial community (i.e. community structure, bacteria and fungi/bacteria as indicated by phospholipid fatty acids analysis) and the nematode community (i.e. total abundance, taxa richness and maturity index).Declines in plant species richness and productivity were greater at high intensities of soil acidification in the second sampling year than in the first sampling year. The changes in plant community observed were mostly explained by soil nutrient pathways (e.g. N availability or base mineral cations), which were in turn regulated by the soil microbial or nematode communities as well as by the direct effects of the increase in H+ or Al3+ ions.Synthesis. Our results suggest that the below-ground microbial and nematode communities are more sensitive to soil acidification than the plant communities are, and further that soil acidification nduced changes in plants are mediated by changes in below-ground communities and soil nutrients. These findings improve our understanding of the links between below- and above-ground communities in the Inner Mongolia grassland, especially in the context of anthropogenic acid enrichment.
DOI:10.1046/j.1469-8137.2000.00686.xURL [本文引用: 2]
Abstract Root turnover is important to the global carbon budget as well as to nutrient cycling in ecosystems and to the success of individual plants. Our ability to predict the effects of environmental change on root turnover is limited by the difficulty of measuring root dynamics, but emerging evidence suggests that roots, like leaves, possess suites of interrelated traits that are linked to their life span. In graminoids, high tissue density has been linked to increased root longevity. Other studies have found root longevity to be positively correlated with mycorrhizal colonization and negatively correlated with nitrogen concentration, root maintenance respiration and specific root length. Among fruit trees, apple roots (which are of relatively small diameter, low tissue density and have little lignification of the exodermis) have much shorter life spans than the roots of citrus, which have opposite traits. Likewise, within the branched network of the fine root system, the finest roots with no daughter roots tend to have higher N concentrations, faster maintenance respiration, higher specific root length and shorter life spans than secondary and tertiary roots that bear daughter roots. Mycorrhizal colonization can enhance root longevity by diverse mechanisms, including enhanced tolerance of drying soil and enhanced defence against root pathogens. Many variables involved in building roots might affect root longevity, including root diameter, tissue density, N concentration, mycorrhizal fungal colonization and accumulation of secondary phenolic compounds. These root traits are highly plastic and are strongly affected by resource supply (CO 2 , N, P and water). Therefore the response of root longevity to altered resource availability associated with climate change can be estimated by considering how changes in resource availability affect root construction and physiology. A cost enefit approach to predicting root longevity assumes that a plant maintains a root only until the efficiency of resource acquisition is maximized. Using an efficiency model, we show that reduced tissue Nconcentration and reduced root maintenance respiration, both of which are predicted to result from elevated CO 2 , should lead to slightly longer root life spans. Complex interactions with soil biota and shifts in plant defences against root herbivory and parasitism, which are not included in the present efficiency model, might alter the effects of future climate change on root longevity in unpredicted ways.
DOI:10.1007/s10533-004-0370-0URL [本文引用: 2]
DOI:10.1007/s004420100746URL [本文引用: 1]
Using a new approach involving one-time measurements of radiocarbon ( 14 C) in fine (<202mm diameter) root tissues we have directly measured the mean age of fine-root carbon. We find that the carbon making up the standing stock of fine roots in deciduous and coniferous forests of the eastern United States has a mean age of 3–1802years for live fine roots, 10–1802years for dead fine roots, and 3–1802years for mixed live+dead fine roots. These 14 C-derived mean ages represent the time C was stored in the plant before being allocated for root growth, plus the average lifespan (for live roots), plus the average time for the root to decompose (for dead roots and mixtures). Comparison of the 14 C content of roots known to have grown within 102year with the 14 C of atmospheric CO 2 for the same period shows that root tissues are derived from recently fixed carbon, and the storage time prior to allocation is <202years and likely <102year. Fine-root mean ages tend to increase with depth in the soil. Live roots in the organic horizons are made of C fixed 3–802years ago compared with 11–1802years in the mineral B horizons. The mean age of C in roots increases with root diameter and also is related to branching order. Our results differ dramatically from previous estimates of fine-root mean ages made using mass balance approaches and root-viewing cameras, which generally report life spans (mean ages for live roots) of a few months to 1–202years. Each method for estimating fine-root dynamics, including this new radiocarbon method, has biases. Root-viewing approaches tend to emphasize more rapidly cycling roots, while radiocarbon ages tend to reflect those components that persist longest in the soil. Our 14 C-derived estimates of long mean ages can be reconciled with faster estimates only if fine-root populations have varying rates of root mortality and decomposition. Our results indicate that a standard definition of fine roots, as those with diameters of <202mm, is inadequate to determine the most dynamic portion of the root population. Recognition of the variability in fine-root dynamics is necessary to obtain better estimates of belowground C inputs.
DOI:10.1046/j.1469-8137.2000.00681.xURL [本文引用: 3]
Abstract Root turnover is a critical component of ecosystem nutrient dynamics and carbon sequestration and is also an important sink for plant primary productivity. We tested global controls on root turnover across climatic gradients and for plant functional groups by using a database of 190 published studies. Root turnover rates increased exponentially with mean annual temperature for fine roots of grasslands ( r 2 = 0.48) and forests ( r 2 = 0.17) and for total root biomass in shrublands ( r 2 = 0.55). On the basis of the best-fit exponential model, the Q 10 for root turnover was 1.4 for forest small diameter roots (5 mm or less), 1.6 for grassland fine roots, and 1.9 for shrublands. Surprisingly, after accounting for temperature, there was no such global relationship between precipitation and root turnover. The slowest average turnover rates were observed for entire tree root systems (10% annually), followed by 34% for shrubland total roots, 53% for grassland fine roots, 55% for wetland fine roots, and 56% for forest fine roots. Root turnover decreased from tropical to high-latitude systems for all plant functional groups. To test whether global relationships can be used to predict interannual variability in root turnover, we evaluated 14 yr of published root turnover data from a shortgrass steppe site in northeastern Colorado, USA. At this site there was no correlation between interannual variability in mean annual temperature and root turnover. Rather, turnover was positively correlated with the ratio of growing season precipitation and maximum monthly temperature ( r 2 = 0.61). We conclude that there are global patterns in rates of root turnover between plant groups and across climatic gradients but that these patterns cannot always be used for the successful prediction of the relationship of root turnover to climate change at a particular site.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1469-8137.2005.01530.xURLPMID:16219077 [本文引用: 1]
Leaf nitrogen and phosphorus stoichiometry of Chinese terrestrial plants was studied based on a national data set including 753 species across the country. Geometric means were calculated for functional groups based on life form, phylogeny and photosynthetic pathway, as well as for all 753 species. The relationships between leaf N and P stoichiometric traits and latitude (and temperature) were analysed. The geometric means of leaf N, P, and N : P ratio for the 753 species were 18.6 and 1.21 mg g(-1) and 14.4, respectively. With increasing latitude (decreasing mean annual temperature, MAT), leaf N and P increased, but the N : P ratio did not show significant changes. Although patterns of leaf N, P and N : P ratios across the functional groups were generally consistent with those reported previously, the overall N : P ratio of China's flora was considerably higher than the global averages, probably caused by a greater shortage of soil P in China than elsewhere. The relationships between leaf N, P and N : P ratio and latitude (and MAT) also suggested the existence of broad biogeographical patterns of these leaf traits in Chinese flora.
DOI:10.1111/j.1365-2745.2005.01067.xURL [本文引用: 2]
Summary 1 Elucidation of the patterns and controls of forest net primary production at ecosystem scales has been hindered by a poor understanding of fine root production, due largely to technical limitations. 2 Fine root (≤0.5mm diameter) production was assessed using minirhizotron, soil core, ingrowth core, nitrogen budget and carbon budget techniques in three longleaf pine-wiregrass forest ecosystem types (hydric, mesic and xeric) forming an edaphic resource availability and above-ground productivity gradient. 3 Fine root production estimates differed substantially in magnitude, e.g. values ranged from 0 to 4618kgha 611 year 611 for the soil core and minirhizotron techniques, respectively, in the hydric site. 4 Minirhizotron production estimates in the hydric, mesic and xeric sites were 4618, 1905 and 2295kgha 611 year 611 , respectively. 5 Soil core and ingrowth core root production estimates were on average 81 and 54% lower, respectively, than corresponding minirhizotron production estimates, and minirhizotron estimates were negatively related to soil core and ingrowth core estimates across the resource gradient. 6 The N budget method yielded unreliable root production estimates, presumably due to the underestimation of N availability for plant assimilation. 7 C budget estimates of total below-ground C allocation (6773, 5646 and 4647kgCha 611 year 611 ) were positively related to minirhizotron production estimates, but negatively related to soil core and ingrowth core production estimates. 8 Critical evaluations of the assumptions, potential errors and results for each method suggest that the minirhizotron technique yielded the most reliable root production estimates, and that the negative relationship between minirhizotron and core-based estimates may be attributed to the inherent deficiency of the core techniques in assessing root production when mortality and production occur simultaneously. 9 Minirhizotron root production estimates were positively related to foliage production estimates, supporting the hypothesis of constant proportional allocation of production to foliage, wood and fine roots across resource availability gradients in temperate forests. 10 These results suggest that fine root production is not negatively correlated with soil resource availability and foliage production as is commonly perceived in the ecological community and represented in ecosystem computer models.
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.94.14.7362URLPMID:11038557 [本文引用: 2]
Global biogeochemical models have improved dramatically in the last decade in their representation of the biosphere. Although leaf area data are an important input to such models and are readily available globally, global root distributions for modeling water and nutrient uptake and carbon cycling have not been available. This analysis provides global distributions for fine root biomass, length, and surface area with depth in the soil, and global estimates of nutrient pools in fine roots. Calculated root surface area is almost always greater than leaf area, more than an order of magnitude so in grasslands. The average C:N:P ratio in living fine roots is 450:11:1, and global fine root carbon is more than 5% of all carbon contained in the atmosphere. Assuming conservatively that fine roots turn over once per year, they represent 33% of global annual net primary productivity.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.agee.2005.10.004URL [本文引用: 1]
The loss of nutrients from agricultural land to water bodies is a serious concern in many countries. To gain information on the contribution of grazing animals to diffuse nutrient losses from pasture areas to water, this study looked at the impact of cattle on nutrient concentrations in overland flow and on soil hydrology (bulk density, macroporosity and resistance to penetration). Rainfall simulations to produce overland flow were conducted and soil physical measurements were taken on experimental plots assigned to one of two treatments: (1) cattle had unrestricted access to the plot; (2) cattle could graze the plot but they could neither walk on the plot area nor deposit excrements on it. Areas to which the cattle had free access were characterised by 57–83% lower macroporosity, by 8–17% higher bulk density and by 27–50% higher resistance to penetration than areas from which the cattle were excluded. The nutrient losses from grassland that can specifically be attributed to the presence of grazing animals were found mainly in the particulate nitrogen, the organic phosphorus and in the potassium exports. Overall, the presence of cattle had a longer lasting effect on the soil hydrological parameters measured than on the nutrient concentrations in overland flow.
DOI:10.1016/j.soilbio.2015.01.001URL [本文引用: 3]
61We evaluated the responses of plant root traits to simulated nitrogen deposition.61Simulated nitrogen deposition elevated the ratio of carbon sequestered in plant.61Simulated nitrogen deposition increased carbon and nutrient retention in the soil.61The amount of N-addition of many simulated experiments was excessive.
[本文引用: 1]
DOI:10.1007/s13213-014-0889-9URLPMID:4331610 [本文引用: 1]
Abstract In this study, the effect of mineral fertilizer and organic manure were evaluated on soil microbial biomass, dehydrogenase activity, bacterial and fungal community structure in a long-term (33 years) field experiment. Except for the mineral nitrogen fertilizer (N) treatment, long-term fertilization greatly increased soil microbial biomass carbon (SMBC) and dehydrogenase activity. Organic manure had a significantly greater impact on SMBC and dehydrogenase activity, compared with mineral fertilizers. Bacterial and fungal community structure was analyzed by polymerase chain reaction (PCR)-denaturing gradient gel electrophoresis (DGGE). Long-term fertilization increased bacterial and fungal ribotype diversity. Total soil nitrogen (TN) and phosphorus (TP), soil organic carbon (SOC) and available phosphorus (AP) had a similar level of influence on bacterial ribotypes while TN, SOC and AP had a larger influence than alkali-hydrolyzable nitrogen (AHN) on fungal ribotypes. Our results suggested that long-term P-deficiency fertilization can significantly decrease soil microbial biomass, dehydrogenase activity and bacterial diversity. N-fertilizer and SOC have an important influence on bacterial and fungal communities.
DOI:10.1046/j.1365-2486.2002.00532.xURL [本文引用: 1]
The IFEF database (Indicators of Forest Ecosystem Functioning), consisting of nitrogen deposition, nitrate leaching fluxes, and soil and ecosystem characteristics, is analysed to evaluate the C/N ratio in the organic horizon as an indicator of nitrate leaching. One hundred and eighty one forests are examined, from countries across Europe ranging from boreal to Mediterranean regions, encompassing broadleaf and coniferous sites and plot and catchment studies. N input in throughfall ranges from less than 1 kg N ha 611 y 611 in northern Norway and Finland to greater than 60 kg N ha 611 y 611 in the Netherlands and Czech Republic. The amount of NO 3 – leached covers a smaller range, between 1 and 40 kg N ha 611 y 611 . Nitrate leaching is strongly dependent on the amount of nitrogen deposited in throughfall (N input) and simply adding the C/N ratio in the organic horizon to a regression equation does not improve this relationship. However, when the data are stratified based on C/N ratios less than or equal to 25 and greater than 25, highly significant relationships ( P < 0.05) are observed between N input and NO 3 – leached. The slope of the relationship for those sites where C/N ratio is ≤ 25 ('nitrogen enriched' sites) is twice that for those sites where C/N ratio is > 25. These empirical relationships may be used to identify which forested ecosystems are likely to show elevated rates of nitrate leaching under predicted future nitrogen deposition scenarios. Elevated NO 3 – leaching also shows a relationship with soil pH, with high rates of NO 3 – leaching only observed at sites with a pH < 4.5 and N inputs > 30 kg N ha 611 y 611 . Tree age and species have no significant impact on the ecosystem response to N input at a regional scale.
URL [本文引用: 3]
DOI:10.1111/j.1365-2486.2004.00733.xURL [本文引用: 1]
Abstract The effects of soil warming and nitrogen availability on root production, longevity and mortality were studied using minirhizotrons in irrigation (C), fertilized (F), heated (H), and heated-fertilized (HF) plots in a Norway spruce stand in northern Sweden from October 1996 to October 1997. Irrigation was included in all treatment plots. Heating cables were used to maintain the soil temperature in heated plots at 5 C above that in unheated plots during the growing season. A Kaplan eier approach was used to estimate the longevity of fine roots and Cox proportional hazards regression to analyze the effects of the H, F, and HF treatments on the risk of root mortality. The proportion of annual root length production contributed by winter pring production amounted to 52% and 49% in heated plots and heated-fertilized plots, respectively. The annual root length production in C plots was significantly higher than in other treatments, while the HF treatment gave significantly greater production compared with the F treatment. The risk of mortality (hazard ratio) relative to C plots was higher in H plots (358%) and F plots (191%). The interaction between heating and fertilizing was strongly significant. The increase in the risk of root mortality in combined fertilization and heating (103%) was lower than that in the H or F plots. The results show that nitrogen addition combined with warmer temperatures decreases the risk of root mortality, and fine root production is a function of the length of the growing season. In the future, fertilization combined with the warmer temperatures expected to follow predicted climatic change may increase root production in boreal forests at low fertility sites.
DOI:10.1023/A:1020372220163URL
Riparian meadows in the western US are highly productive areas withinthe riparian corridor that have been extensively utilized for livestock grazingand that are often degraded. This study aimed to increase our understanding ofboth the functional responses and restoration potential of mesic meadow systemsin central Nevada, USA. Nitrogen addition and clipping treatments were used toexamine grazing effects, while aeration and revegetation were included toevaluate potential restoration treatments. Belowground response was determinedby examining gravimetric soil water content and rooting depth and activity.Aboveground response was determined for two key mesic meadow species, Carex nebrascensis and Poa pratensis ,by quantifying gas exchange and water relations. Large yearly and seasonaldifferences among water tables largely determined the rooting activity anddepthin these mesic meadows and influenced treatment responses. Little rootingactivity occurred within or at the surface of the water table and rooting depthincreased as water table level lowered during the growing season. In general,nitrogen addition decreased rooting activity and depth relative to controls. Itresulted in less negative water potentials and photosynthetic rates that werehigher early in the growing season, but lower later in the growing season. Theeffects of nitrogen addition could be attributed to accelerated phenology andearlier senescence. Clipping resulted in less negative plant water potentialsin Carex and Poa , and Poa had higher photosynthetic rates immediately afterclipping but only for certain dates and comparisons. Aeration increased rootingactivity and depth where there was no confounding effect of water table. Also,predawn and midday water potentials were generally less negative for Carex and Poa in aerated plots.Establishment on the revegetation plots was dominated by annual and early seralspecies and, consequently, these plots exhibited reduced rooting depth andactivity early in the growing season and high root turnover. Results indicatethat water table depth and its effects on soil water are dominant factors indetermining the functional processes and recovery potentials of these riparianmeadows. Clipping had minimal effects on above- and belowground responses,perhaps because clipping was performed late in the growing season after plantshad begun to senescence. Nitrogen addition decreased rooting activity andaltered phenology, indicating deleterious effects. Aeration was effective atovercoming some of the negative affects of overgrazing and may be an effectiverestoration treatment. However, revegetation may have limited potential due toestablishment constraints of the dominant species.
DOI:10.17521/cjpe.2004.0094URL
细根周转要消耗大量的C,它影响森林生态系统C分配格局与过程和养分循环,对生态系统生产力具有重要意义.细根的周转取决于细根的寿命,细根寿命越短,周转越快,根系对C的消耗也越多.大量研究表明,细根的寿命与地上部分C向根系供应的多少有密切关系,同时也与细根直径大小、土壤中N和水分的有效性、土壤温度以及根际周围的土壤动物和微生物的活动有关.本文综述了国外近年来在该领域里的研究进展,特别是对控制细根寿命的机理和主要影响因子进行了评述,目的是引起国内研究者的关注,促进我国根系生态学的研究与发展.
DOI:10.17521/cjpe.2004.0094URL
细根周转要消耗大量的C,它影响森林生态系统C分配格局与过程和养分循环,对生态系统生产力具有重要意义.细根的周转取决于细根的寿命,细根寿命越短,周转越快,根系对C的消耗也越多.大量研究表明,细根的寿命与地上部分C向根系供应的多少有密切关系,同时也与细根直径大小、土壤中N和水分的有效性、土壤温度以及根际周围的土壤动物和微生物的活动有关.本文综述了国外近年来在该领域里的研究进展,特别是对控制细根寿命的机理和主要影响因子进行了评述,目的是引起国内研究者的关注,促进我国根系生态学的研究与发展.
URL
细根周转与土壤养分密切相关,但由于根系研究方法的差异以及研究对象的不同,土壤养分对细根周转影响的研究存在不一致的结论。本文以水曲柳(Fraxinus mandshurica)人工林为对象,应用3种方法研究施氮肥对细根生产和周转的影响。结果表明:施肥降低了活细根现存生物量,但施氮肥样地细根年生产量平均值(93.105g·m-2·a-1)与对照样地(93.505g·m-2·a-1)没有差异,不同方法得出施氮肥样地细根平均周转率(0.917次·a-1)大于对照样地(0.710次·a-1);不同土层内细根的生产量显著不同,表层生产量最大,土层越深细根生产量越低,但细根周转率一般随土壤加深而加快;不同的研究方法得出细根的年生产量和周转率差异较大,分室模型法最高,其次是内生长土芯法,极差法和积分法最低。
URL
细根周转与土壤养分密切相关,但由于根系研究方法的差异以及研究对象的不同,土壤养分对细根周转影响的研究存在不一致的结论。本文以水曲柳(Fraxinus mandshurica)人工林为对象,应用3种方法研究施氮肥对细根生产和周转的影响。结果表明:施肥降低了活细根现存生物量,但施氮肥样地细根年生产量平均值(93.105g·m-2·a-1)与对照样地(93.505g·m-2·a-1)没有差异,不同方法得出施氮肥样地细根平均周转率(0.917次·a-1)大于对照样地(0.710次·a-1);不同土层内细根的生产量显著不同,表层生产量最大,土层越深细根生产量越低,但细根周转率一般随土壤加深而加快;不同的研究方法得出细根的年生产量和周转率差异较大,分室模型法最高,其次是内生长土芯法,极差法和积分法最低。
DOI:10.1078/1433-8319-00007URL [本文引用: 1]
We investigated allocation to roots, stems and leaves of 27 species of herbaceous clonal plants grown at two nutrient levels. Allocation was analyzed as biomass ratios and also allometrically. As in other studies, the fraction of biomass in stems and, to a lesser extent, in leaves, was usually higher in the high-nutrient treatment than in the low-nutrient treatment, and the fraction of biomass in roots was usually higher under low-nutrient conditions. The relationship between the biomass of plant structures fits the general allometric equation, with an exponent 1 in most of the species. The different biomass ratios under the two nutrient conditions represented points on simple allometric trajectories, indicating that natural selection has resulted in allometric strategies rather than plastic responses to nutrient level. In other words, in most of the species that changed allocation in response to the nutrient treatment, these changes were largely a consequence of plant size. Our data suggest that some allocation patterns that have been interpreted as plastic responses to different resource availabilities may be more parsimoniously explained as allometric strategies.
DOI:10.1046/j.1469-8137.2000.00677.xURL [本文引用: 3]
DOI:10.1073/pnas.0403491101URL [本文引用: 1]
Fine-root production and turnover are important regulators of the biogeochemical cycles of ecosystems and key components of their response to global change. We present a nearly continuous 6-year record of fine-root production and mortality from minirhizotron analysis of a closed-canopy, deciduous sweetgum forest in a free-air CO(2) enrichment experiment. Annual production of fine roots was more than doubled in plots with 550 ppm CO(2) compared with plots in ambient air. This response was the primary component of the sustained 22% increase in net primary productivity. Annual fine-root mortality matched annual production, and the mean residence time of roots was not altered by elevated CO(2), but peak fine-root standing crop in midsummer was significantly higher in CO(2)-enriched plots, especially deeper in the soil profile. The preferential allocation of additional carbon to fine roots, which have a fast turnover rate in this species, rather than to stemwood reduces the possibility of long-term enhancement by elevated CO(2) of carbon sequestration in biomass. However, sequestration of some of the fine-root carbon in soil pools is not precluded, and there may be other benefits to the tree from a seasonally larger and deeper fine-root system. Root-system dynamics can explain differences among ecosystems in their response to elevated atmospheric CO(2); hence, accurate assessments of carbon flux and storage in forests in a globally changing atmosphere must account for this unseen and difficult-to-measure component.
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1001-8166.2008.08.011URL [本文引用: 1]
碳氮作为陆地生态系统最关键的两大生源要素,它们在自然界的循环过程中不仅各自对全球变暖做出重要贡献,而且两者的循环过程显著耦合,互相影响各自的作用和效果。从氮元素对植物光合作用、呼吸作用以及土壤呼吸作用影响的角度入手,综述了氮输入对陆地生态系统碳固定和碳排放这两个碳循环关键过程的影响特征和机理,分析了陆地生态系统碳源汇对氮素变化响应的不确定性,在此基础上对未来的相关重点研究方向进行了探讨和展望。
DOI:10.3321/j.issn:1001-8166.2008.08.011URL [本文引用: 1]
碳氮作为陆地生态系统最关键的两大生源要素,它们在自然界的循环过程中不仅各自对全球变暖做出重要贡献,而且两者的循环过程显著耦合,互相影响各自的作用和效果。从氮元素对植物光合作用、呼吸作用以及土壤呼吸作用影响的角度入手,综述了氮输入对陆地生态系统碳固定和碳排放这两个碳循环关键过程的影响特征和机理,分析了陆地生态系统碳源汇对氮素变化响应的不确定性,在此基础上对未来的相关重点研究方向进行了探讨和展望。
DOI:10.1111/j.1365-2486.2007.01465.xURL [本文引用: 1]
Abstract High levels of atmospheric nitrogen (N) deposition in Europe and North America were maintained throughout the 1990s, and global N deposition is expected to increase by a factor of 2.5 over the next century. Available soil N limits primary production in many terrestrial ecosystems, and some computer simulation models have predicted that increasing atmospheric N deposition may result in greater terrestrial carbon (C) storage in woody biomass. However, empirical evidence demonstrating widespread increases in woody biomass C storage due to atmospheric N deposition is uncommon. Increased C storage in soil organic matter due to chronic N inputs has rarely been reported and is often not considered in computer simulation models of N deposition effects. Since 1994, we have experimentally simulated chronic N deposition by adding 3gNm 612 yr 611 to four different northern hardwood forests, which span a 500km geographic gradient in Michigan. Each year we measured tree growth. In 2004, we also examined soil C content to a depth of 70cm. When we compared the control treatment with the NO 3 61 deposition treatment after a decade of experimentation, ecosystem C storage had significantly increased in both woody biomass (500gCm 612 ) and surface soil (0–10cm) organic matter (690gCm 612 ). The increase in surface soil C storage was apparently driven by altered rates of organic matter decomposition, rather than an increase in detrital inputs to soil. Our results, for study locations stretching across hundreds of kilometers, support the hypothesis that chronic N deposition may increase C storage in northern forests, potentially contributing to a sink for anthropogenic CO 2 in the northern Hemisphere.
DOI:10.1111/j.1469-8137.1995.tb03025.xURL [本文引用: 2]
In most natural ecosystems a significant portion of carbon fixed through photosynthesis is allocated to the production and maintenance of fine roots, the ephemeral portion of the root system that absorbs growth-limiting moisture and nutrients. In turn, senescence of fine roots can be the greatest source of C input to forest soils. Consequently, important questions in ecology entail the extent to which increasing atmospheric CO 2 may alter the allocation of carbon to, and demography of, fine roots. Using microvideo and image analysis technology, we demonstrate that elevated atmospheric CO 2 increases the rates of both fine root production and mortality. Rates of root mortality also increased substantially as soil nitrogen availability increased, regardless of CO 2 concentration. Nitrogen greatly influenced the proportional allocation of carbon to leaves vs. fine roots. The amount of available nitrogen in the soil appears to be the most important factor regulating fine root demography in Populus trees.
DOI:10.1046/j.1365-2486.2002.00481.xURL [本文引用: 1]
Abstract Fine root turnover is a critical component of below-ground forest ecology, which regulates nutrient dynamics, forest net primary productivity, carbon input to soils, and soil respiration. Understanding fine root responses to changing environmental conditions is critical for predicting the productivity and carbon sequestration potential of forest ecosystems during the 21st century. The first objective of this study is to demonstrate that a mechanistic model can realistically simulate spatial and temporal fine root demography in temperate forests on the basis of two hypotheses: (1) absorption of mineral N (N) stimulates the production of new roots, and (2) fine root longevity decreases with increasing N availability. Based on this model, my second objective is to predict fine root responses to changing atmospheric CO 2 levels and N deposition rates. To meet these objectives, an extensive description of the N cycle and the new fine root module were implemented in the ASPECTS model. In agreement with a wide body of literature information, the new model predicted: (1) a preferential colonisation by fine roots of the uppermost soil layer, and (2) a flush of fine root growth in the spring. The simulations indicate that fine root biomass will increase in response to elevated CO 2 under the double effect of (1) an increase in root longevity due to increased N stress, and (2) larger amounts of assimilates available to the growth of plant tissue due to increased photosynthesis. Although the simulated total fine root biomass increased under both increasing N deposition rates and atmospheric CO 2 concentrations, the model predicts that the distribution of fine roots among soil layers will be altered. This suggests that experimental studies must consider the full depth of the root system in order to accurately assess effects of environmental changes on fine root dynamics. The model also suggests that fine root longevity is a plastic parameter, which varied from less than 1year to more than 3years depending on forcing values of N deposition rates and atmospheric CO 2 concentrations. Finally, the model indicates that the increase in net ecosystem exchange (NEE) and soil respiration in temperate forests under elevated CO 2 will be proportional to the amount of available N, with little to no response in low N conditions and up to +28% for both NEE and soil respiration under the highest deposition rate (7.0gNm 2 y 1 ).
DOI:10.1038/ngeo230URL [本文引用: 1]
Land and ocean uptake of carbon dioxide plays a critical role in determining atmospheric carbon dioxide levels. Future increases in nitrogen deposition have been predicted to increase the size of these terrestrial and marine carbon sinks, but although higher rates of nitrogen deposition might enhance carbon uptake in northern and tropical forests, they will probably have less of an impact on ocean sink strength. Combined, the land and ocean sinks may sequester an additional 10% of anthropogenic cabon emissions by 2030 owing to increased nitrogen inputs, but a more conservative estimate of 1 to 2% is more likely. Thus nitrogen-induced increases in the strength of land and ocean sinks are unlikely to keep pace with future increases in carbon dioxide.
DOI:10.1093/jpe/rtp034URL [本文引用: 1]
[Aims] Theories based on resource additions indicate that plant species richness is mainly determined by the number of limiting resources. However, the individual effects of various limiting resources on species richness and aboveground net primary productivity (ANPP) are less well understood. Here, we analyzed potential linkages between additions of limiting resources, species loss and ANPP increase and further explored the underlying mechanisms. [Methods] Resources (N, P, K and water) were added in a completely randomized block design to alpine meadow plots in the Qinghai-Tibetan Plateau....
DOI:10.1007/s11104-011-0896-6URL
Aims A key issue related to the usefulness of the minirhizotron technique is whether root presence and behaviour in the soil zone at the minirhizotron interface are consistent with those in the bulk soil. We wanted to test the null hypotheses that there were no differences in root densities or specific root length (SRL) between those positions. The effects of different nutrient and water regimes, and different tree species (grey alder and basket willow) on the root variables were also examined. Methods We quantified root mass and length densities, and calculated SRL, of three diameter classes from cores sampled around minirhizotrons and in the bulk soil, i.e. quartz sand. Fine root dynamics were monitored in the minirhizotrons during three consecutive years prior to sampling. The study was conducted on individual root systems within buried and covered lysimeters, placed in a stand structure, and with different water and nutrient regimes. Results Significant discrepancies in root densities were found between positions. Plants subjected to limited water or nutrient supply had up to three times higher densities at the minirhizotrons. However, effects of species and treatments showed a similar pattern for the diameter class <1 mm between the two sampling positions and minirhizotron observations. The pulses of fine root growth and decay were coordinated in time for both species and treatments. The SRL was not affected by the tube in any diameter class. Conclusions We concluded that the minirhizotron technique alone was of limited use for estimating root densities in the bulk soil. But the results showed that minirhizotrons could be useful, for example, combined with soil coring or in comparative studies. The patterns of fine root growth and decay were similar for species and treatments, and SRL was unaffected, suggesting that minirhizotrons can be used in studies of root dynamics and morphology. There is, however, a need for further studies concerning the influence on root survival and decay.
DOI:10.2134/agronj1978.00021962007000060043xURL [本文引用: 1]
An experimental procedure, described in this paper, involved the use of a highly refined fiber optic duodenoscope for observing and photographing root development patterns within a soil profile. The root was monitored by inserting the unit into a transparent Plexiglas tube imbedded within the root zone. This technique permitted the observation and quantitative characterization of the root system throughout the entire growing season, which, until this time, has been excessively expensive, laborious, time consuming, and nonreproducible. Root lengths for soybeans [(L.) Merr. ‘Lee-68’] at the R-7 stage of maturity were calculated and compared by the duodenoscope and the Newman-Core methods at four depth-intervals. The depth intervals were reported as 0–18, 18–36, 36–54, and 54–72 cm in a Peridge silt loam (Typic Paleudalf; fine silty, mixed, mesic). Total root lengths estimated by the Newman-Core method for the 0 to 72 cm depth-intervals was 56.36 m, while the estimate by the scope was 52.84 m. Although both methods gave similar results, the scope tended to estimate a greater root length than did the soil cores at all depths except the 0 to 18 cm. Both methods showed that the majority of the root system was in the 0 to 18 cm depth; 68% for the soil cores and 47% for the scope. Root density (cm/cm) decreased with soil depth from 10.58 to 1.52 by the soil core method and from 6.77 to 2.83 for the scope measurement. The coefficient of variation was 52.64 and 14.10 percent for the core and scope measurements, respectively. The study indicated that the duodenoscope method can be used as a nondestructive technique for quantitatively measuring soybean root systems throughout the growing season.
URL [本文引用: 1]
The success of pines under diverse environmental conditions results, in part, from their ability to acquire limiting resources under highly competitive conditions. Among the important traits that favor pines over other competitors is variation in the longevity of the principal tissues for resource acquisition -- the foliage and fine roots. Among the major tree genera the pines clearly show the greatest interspecific variation in foliage longevity (1.5 to >20 yrs), and some preliminary evidence suggests that the same may be true for fine roots (0.2 to >5 yrs). We have constructed a conceptual model suggesting that leaf longevity is a consequence of the interaction of conditions at the site-, crown-, shoot- and leaf-levels. The applicability of this model to explain the variable response of leaf longevity to environmental conditions is evaluated. In contrast to leaves, fine roots of most plants do not show any active separation from the parent in the form of abscission, and limited information is available on the proximate causes of fine root mortality. The effects of environmental stress and resource availability on fine root longevity and production are discussed. Although the interacting factors controlling foliage longevity appear to have been identified, factors influencing fine root longevity are much less certain.
DOI:10.1111/j.1365-2486.2012.02738.xURLPMID:28741827 [本文引用: 1]
Plant species and functional groups in nitrogen (N) limited communities may coexist through strong eco-physiological niche differentiation, leading to idiosyncratic responses to multiple nutrition and disturbance regimes. Very little is known about how such responses depend on the availability of N in different chemical forms. Here we hypothesize that idiosyncratic year-to-year responses of plant functional groups to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing. We conducted a 6 year N addition experiment in an alpine meadow on the Tibetan Plateau released from grazing by livestock. The experimental design featured three N forms (ammonium, nitrate, and ammonium nitrate), crossed with three levels of N supply rates (0.375, 1.500 and 7.500 g N m 2 yr 1), with unfertilized treatments without and with light grazing as controls. All treatments showed increasing productivity and decreasing species richness after cessation of grazing and these responses were stronger at higher N rates. Although N forms did not affect aboveground biomass at community level, different functional groups did show different responses to N chemical form and supply rate and these responses varied from year to year. In support of our hypothesis, these idiosyncratic responses seemed to enable a substantial diversity and biomass of sedges, forbs, and legumes to still coexist with the increasingly productive grasses in the absence of grazing, at least at low and intermediate N availability regimes. This study provides direct field-based evidence in support of the hypothesis that idiosyncratic and annually varying responses to both N quantity and quality may be a key driver of community structure and species coexistence. This finding has important implications for the diversity and functioning of other ecosystems with spatial and temporal variation in available N quantity and quality as related to changing atmospheric N deposition, land-use, and climate-induced soil warming.
DOI:10.2134/agronj1970.00021962006200060039xURL [本文引用: 1]
Shoot and root growth of maize and tomato in a rhizotron were measured. The root intensity at the transparent panel increased during the season for both species but was always greater for maize. At the end of the season, root density and total root length estimates were 3 times greater for maize than tomato.
DOI:10.1038/ngeo721URL [本文引用: 1]
Human activities have greatly accelerated emissions of both carbon dioxide and biologically reactive nitrogen to the atmosphere. As nitrogen availability often limits forest productivity, it has long been expected that anthropogenic nitrogen deposition could stimulate carbon sequestration in forests. However, spatially extensive evidence for deposition-induced stimulation of forest growth has been lacking, and quantitative estimates from models and plot-level studies are controversial. Here, we use forest inventory data to examine the impact of nitrogen deposition on tree growth, survival and carbon storage across the northeastern and north-central USA during the 1980s and 1990s. We show a range of growth and mortality responses to nitrogen deposition among the region's 24 most common tree species. Nitrogen deposition (which ranged from 3 to 11kghayr) enhanced the growth of 11 species and decreased the growth of 3 species. Nitrogen deposition enhanced growth of all tree species with arbuscular mycorrhizal fungi associations. In the absence of disturbances that reduced carbon stocks by more than 50%, above-ground biomass increment increased by 61kg of carbon per kg of nitrogen deposited, amounting to a 40% enhancement over pre-industrial conditions. Extrapolating to the globe, we estimate that nitrogen deposition could increase tree carbon storage by 0.31Pg carbon yr.
DOI:10.1023/A:1004829423160URL [本文引用: 1]
The minirhizotron technique (MR) for in situ measurement of fine root dynamics offers the opportunity to obtain accurate and unbiased estimates of root production in perennial vegetation only if MR tubes do not affect the longevity of fine roots. Assuming fine root biomass is near steady-state, fine root production (g m 612 yr 611 ) can be estimated as the ratio of fine root biomass (g m 612 ) to median fine root longevity (yr). This study evaluates the critical question of whether MR access tubes affect the longevity of fine roots, by comparing fine root survivorship obtained using MR with those from a non-intrusive in situ screen method in the forest floor horizons of a northern hardwood forest in New Hampshire, USA. Fine root survivorship was measured in 380 root screens during 1993–1997 and in six horizontal minirhizotron tubes during 1996–1997. No statistically significant difference was found between estimates of survivorship of fine roots (<1 mm dia.) at this site from MR versus from in situ screens, suggesting that MR tubes do not substantially affect fine root longevity in the forest floor of this northern hardwood forest and providing greater confidence in measurements of fine root production using the MR technique. Furthermore, the methodology for estimating fine root production from MR longevity data was evaluated by comparison of fine root longevity and production estimates made using single vs. multiple root cohorts, and using root-number, root-length, and root-mass weighted methods. Our results indicate that fine root-length longevity estimates based on multiple root cohorts throughout the year can be used to approximate fine root biomass production. Using this method, we estimated fine root longevity and production in the forest floor at this site to be 314 days (or 0.86 yr) and 303 g m 612 yr 611 , respectively. Fine root production in this northern hardwood forest is approximately equivalent to standing biomass and was previously underestimated by root in-growth cores. We conclude that the use of MR to estimate fine root longevity and production as outlined here may result in improved estimates of fine root production in perennial vegetation.
URL [本文引用: 1]
森林细根分解是陆地生态系统碳循环的重要过程之一,其分解速率受到大气氮沉降增加的潜在影响.2007年11月至2013年1月,对华西雨屏区苦竹人工林进行每月1次的模拟氮沉降试验,设对照(CK,0)、低氮(5gN·m-2·a-1)、中氮(15gN·hm-2·a-1)和高氮(30gN·m-2·a-1)4个处理.2011年1月起,采用分解袋法研究苦竹细根分解.结果表明:苦竹细根分解呈现出先快后慢的趋势,在分解第1年质量损失达60%,分解第2年质量残留率变化较为平缓.对照处理细根质量损失50%和95%分别需要1.20和5.17a.模拟氮沉降显著抑制了苦竹细根分解,相对于对照,高氮处理细根在分解2a后残留量增加51.0%.模拟氮沉降显著增加了凋落物碳、氮和磷元素的残留率.与对照相比,模拟氮沉降处理4.5a后,中氮和高氮处理土壤pH值显著降低,高氮处理土壤有机碳、总氮、铵态氮和硝态氮含量以及苦竹细根生物量显著增加.
URL [本文引用: 1]
森林细根分解是陆地生态系统碳循环的重要过程之一,其分解速率受到大气氮沉降增加的潜在影响.2007年11月至2013年1月,对华西雨屏区苦竹人工林进行每月1次的模拟氮沉降试验,设对照(CK,0)、低氮(5gN·m-2·a-1)、中氮(15gN·hm-2·a-1)和高氮(30gN·m-2·a-1)4个处理.2011年1月起,采用分解袋法研究苦竹细根分解.结果表明:苦竹细根分解呈现出先快后慢的趋势,在分解第1年质量损失达60%,分解第2年质量残留率变化较为平缓.对照处理细根质量损失50%和95%分别需要1.20和5.17a.模拟氮沉降显著抑制了苦竹细根分解,相对于对照,高氮处理细根在分解2a后残留量增加51.0%.模拟氮沉降显著增加了凋落物碳、氮和磷元素的残留率.与对照相比,模拟氮沉降处理4.5a后,中氮和高氮处理土壤pH值显著降低,高氮处理土壤有机碳、总氮、铵态氮和硝态氮含量以及苦竹细根生物量显著增加.
DOI:10.1046/j.1365-2435.2002.00611.xURL [本文引用: 1]
Summary 1 n grass species that occur in pastures or hay meadows, life spans of roots determine much of the carbon and nutrient loss from the plant in addition to the amounts that are lost by mowing or grazing. We hypothesized that grass species from nutrient-poor habitats had longer root life spans and consequently lost smaller quantities of nutrients through root turnover. 2 In a garden experiment, root life spans and root diameters were measured by repeated observations in minirhizotrons placed in monocultures of Lolium perenne L. and Arrhenatherum elatius L. (characteristic of fertile soils) and Molinia caerulea L. and Nardus stricta L. (preferring nutrient-poor soils). 3 Average root life spans were 14weeks in L. perenne , 40weeks in A. elatius , 53weeks in M. caerulea and 58weeks in N. stricta . Root life spans of species from fertile habitats were significantly shorter than the root life spans of species from low fertility habitats. 4 In addition, there were significant differences in root diameter among species, root diameter being positively correlated to root life spans. Root diameter decreased during root ageing in all species, while the decline in diameter occurred more slowly in N. stricta than in A. elatius and M. caerulea . 5 n increase in the abundance of plant species adapted to fertile habitats will, because of the greater C and nutrient returns to the soil in root turnover, increase soil fertility. This effect may constitute a positive feedback between changes in plant species composition and nutrient cycling.
DOI:10.1016/S0065-2504(08)60122-1URL [本文引用: 1]
DOI:10.1007/BF00017088URL [本文引用: 1]
DOI:10.1111/j.1469-8137.2004.01034.xURL [本文引用: 1]
Summary 6168 Fine roots are the key link for plant water and nutrient uptake, soil carbon (C) input and soil microbial activity in forest ecosystems, and play a critical role in regulating ecosystem C balance and its response to global change. 6168 Red maple ( Acer rubrum ) and sugar maple ( Acer saccharum ) seedlings were grown for four growing seasons in open-top chambers and exposed to ambient or elevated carbon dioxide concentration [CO 2 ] in combination with ambient or elevated temperature. Fine-root production and mortality were monitored using minirhizotrons, and root biomass was determined from soil cores. 6168 Both elevated [CO 2 ] and temperature significantly enhanced production and mortality of fine roots during spring and summer of 1996. At the end of the experiment in September 1997, fine root biomass was significantly lower in elevated temperature chambers, but there were no effects of elevated [CO 2 ] or the interactions between elevated [CO 2 ] and temperature. 6168 Deciduous trees have dynamic root systems, and their activity can be enhanced by CO 2 enrichment and climatic warming. Static measures of root response, such as soil core data, obscure the dynamic nature, which is critical for understanding the response of forest C cycling to global change.
DOI:10.1007/s11104-009-0259-8URL [本文引用: 1]
We conducted a field experiment in two alpine meadows to investigate the short-term effects of nitrogen enrichment and plant litter biomass on plant species richness, the percent cover of functional groups, soil microbial biomass, and enzyme activity in two alpine meadow communities. The addition of nitrogen fertilizer to experimental plots over two growing seasons increased plant production, as indicated by increases in both the living plant biomass and litter biomass in the Kobresia humilis meadow community. In contrast, fertilization had no significant effect on the amounts of living biomass and litter biomass in the K. tibetica meadow. The litter treatment results indicate that litter removal significantly increased the living biomass and decreased the litter biomass in the K. humilis meadow; however, litter-removal and litter-intact treatments had no impact on the amounts of living biomass and litter biomass in the K. tibetica meadow. Litter production depended on the degree of grass cover and was also influenced by nitrogen enrichment. The increase in plant biomass reflects a strong positive effect of nitrogen enrichment and litter removal on grasses in the K. humilis meadow. Neither fertilization nor litter removal had any impact on the grass biomass in the K . tibetica meadow. Sedge biomass was not significantly affected by either nutrient enrichment or litter removal in either alpine meadow community. The plant species richness decreased in the K. humilis meadow following nitrogen addition. In the K. humilis meadow, microbial biomass C increased significantly in response to the nitrogen enrichment and litter removal treatments. Enzyme activities differed depending on the enzyme and the different alpine meadow communities; in general, enzyme activities were higher in the upper soil layers (0–1002cm and 10–2002cm) than in the lower soil layers (20–4002cm). The amounts of living plant biomass and plant litter biomass in response to the different treatments of the two alpine meadow communities affected the soil microbial biomass C, soil organic C, and soil fertility. These results suggest that the original soil conditions, plant community composition, and community productivity are very important in regulating plant community productivity and microbial biomass and activity.
DOI:10.5846/stxb201202200232URL [本文引用: 1]
在三江源区研究了不同施肥梯度对高寒矮嵩草草甸群落结构、功能;土壤全量养分、速效养分;土壤有机碳和微生物生物量碳的影响,以揭示矮嵩草草甸群落特征;土壤养分和土壤微生物活性对施肥梯度的响应。结果表明:1)随着施肥量的增加,不同功能群的盖度响应各异,其中禾本科植物的响应较大,而豆科和杂类草植物盖度明显降低,莎草科盖度变化不明显;施肥量增加到一定程度,如施氮40 g/m2时,各功能群植物的盖度逐渐降低。生物量随施肥梯度呈单峰曲线变化,不施肥时生物量最低,施肥20 g/m2或32 g/m2时生物量最高。2)土壤全量养分和速效养分在施肥量为20 g/m2或32 g/m2时较高,施肥量增加到40 g/m2时土壤资源逐渐降低。3)不同施肥梯度矮嵩草草甸土壤有机碳和微生物生物量碳在0—10 cm土层明显较高,且随着施肥量的增加,分布在0—40 cm土层的土壤有机碳含量呈单峰曲线变化。施肥20 g/m2或32 g/m2时土壤有机碳和微生物量碳含量最高。4)30 g/m2施肥量可作为高寒草甸最佳施氮水平。施肥梯度下土壤有机碳和微生物量碳含量可作为衡量土壤肥力和土壤质量变化的重要指标。高施肥量(≥40 g/m2)视为影响高寒草甸生态系统结构与功能、土壤养分及土壤微生物活性的阈值。
DOI:10.5846/stxb201202200232URL [本文引用: 1]
在三江源区研究了不同施肥梯度对高寒矮嵩草草甸群落结构、功能;土壤全量养分、速效养分;土壤有机碳和微生物生物量碳的影响,以揭示矮嵩草草甸群落特征;土壤养分和土壤微生物活性对施肥梯度的响应。结果表明:1)随着施肥量的增加,不同功能群的盖度响应各异,其中禾本科植物的响应较大,而豆科和杂类草植物盖度明显降低,莎草科盖度变化不明显;施肥量增加到一定程度,如施氮40 g/m2时,各功能群植物的盖度逐渐降低。生物量随施肥梯度呈单峰曲线变化,不施肥时生物量最低,施肥20 g/m2或32 g/m2时生物量最高。2)土壤全量养分和速效养分在施肥量为20 g/m2或32 g/m2时较高,施肥量增加到40 g/m2时土壤资源逐渐降低。3)不同施肥梯度矮嵩草草甸土壤有机碳和微生物生物量碳在0—10 cm土层明显较高,且随着施肥量的增加,分布在0—40 cm土层的土壤有机碳含量呈单峰曲线变化。施肥20 g/m2或32 g/m2时土壤有机碳和微生物量碳含量最高。4)30 g/m2施肥量可作为高寒草甸最佳施氮水平。施肥梯度下土壤有机碳和微生物量碳含量可作为衡量土壤肥力和土壤质量变化的重要指标。高施肥量(≥40 g/m2)视为影响高寒草甸生态系统结构与功能、土壤养分及土壤微生物活性的阈值。
DOI:10.1038/ngeo2324URL [本文引用: 1]
Atmospheric phosphorus fertilizes plants and contributes to Earth's biogeochemical phosphorus cycle. However, calculations of the global budget of atmospheric phosphorus have been unbalanced, with global deposition exceeding estimated emissions from dust and sea-salt transport, volcanic eruptions, biogenic sources and combustion of fossil fuels, biofuels and biomass, the latter of which thought to contribute about 5% of total emissions. Here we use measurements of the phosphorus content of various fuels and estimates of the partitioning of phosphorus during combustion to calculate phosphorus emissions to the atmosphere from all combustion sources. We estimate combustion-related emissions of 1.8 Tg P yr, which represent over 50% of global atmospheric sources of phosphorus. Using these estimates in atmospheric transport model simulations, we find that the total global emissions of atmospheric phosphorus (3.5 Tg P yr) translate to a depositional sink of 2.7 Tg P yrover land and 0.8 Tg P yrover the oceans. The modelled spatial patterns of phosphorus deposition agree with observations from globally distributed measurement stations, and indicate a near balance of the phosphorus budget. Our finding suggests that the perturbation of the global phosphorus cycle by anthropogenic emissions is larger thanpreviously thought.
DOI:10.1016/j.electacta.2014.10.051URL [本文引用: 1]
DOI:10.1046/j.1469-8137.2000.00691.xURL [本文引用: 1]
The representation of root activity in models is here confined to considerations of applications assessing the impacts of changes in climate or atmospheric [CO 2 ]. Approaches to modelling roots can be classified into four major types: models in which roots are not considered, models in which there is an interplay between only selected above-ground and below-ground processes, models in which growth allocation to all parts of the plants depends on the availability and matching of the capture of external resources, and models with explicit treatments of root growth, architecture and resource capture. All models seem effective in describing the major root activities of water and nutrient uptake, because these processes are highly correlated, particularly at large scales and with slow or equilibrium dynamics. Allocation models can be effective in providing a deeper, perhaps contrary, understanding of the dynamic underpinning to observations made only above ground. The complex and explicit treatment of roots can be achieved only in small-scale highly studied systems because of the requirements for many initialized variables to run the models.
DOI:10.5846/stxb201307031831URL [本文引用: 1]
植物根系是陆地生态系统重要的碳汇和养分库,细根周转过程是陆地生态系统地下部分碳氮循环的核心环节,在 陆地生态系统如何响应全球变化中起着关键作用.在全球变化敏感地区之一的青藏高原,对该地区的主要植被类型矮嵩草草甸同时采用根钻法、内生长袋法和微根管 法3种观测方法研究细根生产和周转速率,并探讨了极差法、积分法、矩阵法和Kaplan-Meier法等数据处理方法对计算值的影响.研究结果显示:在估 算细根净初级生产力时,根钻法宜采用积分法,内生长袋法宜选用矩阵法;由此进一步以最大细根生物量为基础,根钻法和内生长袋法估测的细根年周转速率分别为 0.36 a-1和0.52 a-1,内生长袋法的估算结果是根钻法的1.44倍.对于微根管法,将其观测得到的细根长度转换为单位面积的生物量值后,采用积分法计算出细根周转速率为 0.84 a-1,远高于传统方法的估算结果;若采用Kaplan-Meier生存分析方法,则计算出的细根周转速率更高达3.41 a-1.
DOI:10.5846/stxb201307031831URL [本文引用: 1]
植物根系是陆地生态系统重要的碳汇和养分库,细根周转过程是陆地生态系统地下部分碳氮循环的核心环节,在 陆地生态系统如何响应全球变化中起着关键作用.在全球变化敏感地区之一的青藏高原,对该地区的主要植被类型矮嵩草草甸同时采用根钻法、内生长袋法和微根管 法3种观测方法研究细根生产和周转速率,并探讨了极差法、积分法、矩阵法和Kaplan-Meier法等数据处理方法对计算值的影响.研究结果显示:在估 算细根净初级生产力时,根钻法宜采用积分法,内生长袋法宜选用矩阵法;由此进一步以最大细根生物量为基础,根钻法和内生长袋法估测的细根年周转速率分别为 0.36 a-1和0.52 a-1,内生长袋法的估算结果是根钻法的1.44倍.对于微根管法,将其观测得到的细根长度转换为单位面积的生物量值后,采用积分法计算出细根周转速率为 0.84 a-1,远高于传统方法的估算结果;若采用Kaplan-Meier生存分析方法,则计算出的细根周转速率更高达3.41 a-1.
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1258.2014.00014URL [本文引用: 4]
青藏高原正经历着明显的温暖化过程,由此引起的土壤温度的升高促进了土壤中微生物的活性,同时青藏高原东缘地区大气氮沉降十分明显,并呈逐年增加的趋势,这些环境变化均促使土壤中可利用营养元素增加,因此深入了解青藏高原高寒草甸植物生物量对可利用营养元素增加的响应,是准确预测未来全球变化背景下青藏高原高寒草甸碳循环过程的重要基础。该研究基于在青藏高原高寒草甸连续4年(2009-2012年)氮、磷添加后对不同功能群植物地上生物量、群落地上和地下生物量的测定,探讨高寒草甸生态系统碳输入对氮、磷添加的响应。结果表明:(1)氮、磷添加均极显著增加了禾草的地上绝对生物量及其在群落总生物量中所占的比例,同时均显著降低了杂类草在群落总生物量中的比例,此外磷添加极显著降低了莎草地上绝对生物量及其在群落总生物量中所占的比例。(2)氮、磷添加均显著促进了青藏高原高寒草甸的地上生物量增加,分别增加了24%和52%。(3)氮添加对高寒草甸地下生物量无显著影响,而磷添加后地下生物量有增加的趋势。(4)氮添加对高寒草甸植物总生物量无显著影响,而磷添加后植物总生物量显著增加。研究表明,氮、磷添加可缓解青藏高原高寒草甸植物生长的营养限制,促进植物地上部分的生长,然而高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制。
DOI:10.3724/SP.J.1258.2014.00014URL [本文引用: 4]
青藏高原正经历着明显的温暖化过程,由此引起的土壤温度的升高促进了土壤中微生物的活性,同时青藏高原东缘地区大气氮沉降十分明显,并呈逐年增加的趋势,这些环境变化均促使土壤中可利用营养元素增加,因此深入了解青藏高原高寒草甸植物生物量对可利用营养元素增加的响应,是准确预测未来全球变化背景下青藏高原高寒草甸碳循环过程的重要基础。该研究基于在青藏高原高寒草甸连续4年(2009-2012年)氮、磷添加后对不同功能群植物地上生物量、群落地上和地下生物量的测定,探讨高寒草甸生态系统碳输入对氮、磷添加的响应。结果表明:(1)氮、磷添加均极显著增加了禾草的地上绝对生物量及其在群落总生物量中所占的比例,同时均显著降低了杂类草在群落总生物量中的比例,此外磷添加极显著降低了莎草地上绝对生物量及其在群落总生物量中所占的比例。(2)氮、磷添加均显著促进了青藏高原高寒草甸的地上生物量增加,分别增加了24%和52%。(3)氮添加对高寒草甸地下生物量无显著影响,而磷添加后地下生物量有增加的趋势。(4)氮添加对高寒草甸植物总生物量无显著影响,而磷添加后植物总生物量显著增加。研究表明,氮、磷添加可缓解青藏高原高寒草甸植物生长的营养限制,促进植物地上部分的生长,然而高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制。
DOI:10.1111/j.1365-2486.2008.01591.xURL [本文引用: 1]
The soils of the Qinghai-Tibetan Plateau store a large amount of organic carbon, but the magnitude, spatial patterns and environmental controls of the storage are little investigated. In this study, using data of soil organic carbon (SOC) in 405 profiles collected from 135 sites across the plateau and a satellite-based dataset of enhanced vegetation index (EVI) during 2001鈥2004, we estimated storage and spatial patterns of SOC in the alpine grasslands. We also explored the relationships between SOC density (soil carbon storage per area) and climatic variables and soil texture. Our results indicated that SOC storage in the top 1 m in the alpine grasslands was estimated at 7.4 Pg C (1 Pg=1015 g), with an average density of 6.5 kg m?2. The density of SOC decreased from the southeastern to the northwestern areas, corresponding to the precipitation gradient. The SOC density increased significantly with soil moisture, clay and silt content, but weakly with mean annual temperature. These variables could together explain about 72% of total variation in SOC density, of which 54% was attributed to soil moisture, suggesting a key role of soil moisture in shaping spatial patterns of SOC density in the alpine grasslands.
DOI:10.1007/s11104-011-0784-0URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2010.10.001URL [本文引用: 2]
植物种群对有限资源的竞争是决定植物群落物种组成、多样性和生产力等群落结构和功能的主要因素。该文以青藏高原高寒草甸为研究对象,研究了短期内不同水平的氮素添加对高寒草甸植物群落的影响。结果表明:1)氮素添加提高了土壤中NO3–-N等可利用资源的含量,增加了植物群落植被的盖度,减小了植被的透光率,随着施氮量的增加,群落中物种丰富度显著降低(p0.001);2)氮素添加显著改变了植物群落的地上生产力(p0.05),随着施氮量的增加,地上生产力呈先增加后降低的变化趋势,各功能群中禾草生物量显著增加,而杂类草和豆科植物生物量随施氮量的增加逐渐减少;3)物种多样性与植被透光率呈线性正相关(p0.05);地上生产力与土壤NO3–-N含量呈线性正相关(p0.05);物种丰富度与地上生产力之间呈负相关关系。这说明短期内氮素添加通过改变土壤中NO3–-N等可利用资源的含量而对植物群落物种组成和地上生产力产生影响。
DOI:10.3773/j.issn.1005-264x.2010.10.001URL [本文引用: 2]
植物种群对有限资源的竞争是决定植物群落物种组成、多样性和生产力等群落结构和功能的主要因素。该文以青藏高原高寒草甸为研究对象,研究了短期内不同水平的氮素添加对高寒草甸植物群落的影响。结果表明:1)氮素添加提高了土壤中NO3–-N等可利用资源的含量,增加了植物群落植被的盖度,减小了植被的透光率,随着施氮量的增加,群落中物种丰富度显著降低(p0.001);2)氮素添加显著改变了植物群落的地上生产力(p0.05),随着施氮量的增加,地上生产力呈先增加后降低的变化趋势,各功能群中禾草生物量显著增加,而杂类草和豆科植物生物量随施氮量的增加逐渐减少;3)物种多样性与植被透光率呈线性正相关(p0.05);地上生产力与土壤NO3–-N含量呈线性正相关(p0.05);物种丰富度与地上生产力之间呈负相关关系。这说明短期内氮素添加通过改变土壤中NO3–-N等可利用资源的含量而对植物群落物种组成和地上生产力产生影响。
DOI:10.5846/stxb201212071765URL [本文引用: 1]
细根对植物群落功能的发挥和土壤碳库及全球碳循环具有重要意义.利用连续土钻取样法和分解袋法,于2010年5-10月整个生长季节内,对三工河流域两处长势不同的琵琶柴群落的细根(φ<2mm)生物量、分解与周转规律及其与土壤环境的关系进行研究.结果表明,群落1和群落2土壤容重、土壤含水量、pH和电导率等土壤因子差异显著.两群落的细根生物量表现出相同的季节和垂直变化趋势,即在5-8月逐渐增加,8月达到最大值,9-10月份逐渐下降.平均月细根生物量分别为51.55g/m2和133.93g/m2.群落1的活细根和死细根分别占总细根生物量的69.68%和30.32%,群落2活细根和死细根分别占总细根生物量的72.61%和27.39%.在垂直变化上,随土壤深度增加细根生物量先增加后逐渐降低,其中10-20cm土壤层次细根生物量比例最大,群落1和群落2分别占46.48%和29.15%.群落1和群落2的细根年分解率分别为34.82%、42.91%.达到半分解和95%分解时,群落1需要630 d和2933 d,群落2需要467 d和2238 d.群落1和群落2的细根净生产力分别为50.67 g/m2和178.15g/m2,细根年周转率分别为1.41次、1.69次.逐步回归分析结果显示细根动态受土壤水分、pH值、电导度等土壤因子的显著影响,琵琶柴细根具有相对较低的分解速率和较高的周转速率.
DOI:10.5846/stxb201212071765URL [本文引用: 1]
细根对植物群落功能的发挥和土壤碳库及全球碳循环具有重要意义.利用连续土钻取样法和分解袋法,于2010年5-10月整个生长季节内,对三工河流域两处长势不同的琵琶柴群落的细根(φ<2mm)生物量、分解与周转规律及其与土壤环境的关系进行研究.结果表明,群落1和群落2土壤容重、土壤含水量、pH和电导率等土壤因子差异显著.两群落的细根生物量表现出相同的季节和垂直变化趋势,即在5-8月逐渐增加,8月达到最大值,9-10月份逐渐下降.平均月细根生物量分别为51.55g/m2和133.93g/m2.群落1的活细根和死细根分别占总细根生物量的69.68%和30.32%,群落2活细根和死细根分别占总细根生物量的72.61%和27.39%.在垂直变化上,随土壤深度增加细根生物量先增加后逐渐降低,其中10-20cm土壤层次细根生物量比例最大,群落1和群落2分别占46.48%和29.15%.群落1和群落2的细根年分解率分别为34.82%、42.91%.达到半分解和95%分解时,群落1需要630 d和2933 d,群落2需要467 d和2238 d.群落1和群落2的细根净生产力分别为50.67 g/m2和178.15g/m2,细根年周转率分别为1.41次、1.69次.逐步回归分析结果显示细根动态受土壤水分、pH值、电导度等土壤因子的显著影响,琵琶柴细根具有相对较低的分解速率和较高的周转速率.
[本文引用: 1]
[本文引用: 1]
Multiple risk factors in root survivorship: A 4-year study in Concord grape
1
2003
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling
1
2010
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
Spatial and temporal effects of nitrogen addition on root life span of Leymus chinensis in a typical steppe of Inner Mongolia
4
2008
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
... ), 但提高土壤养分资源, 植物将会分配更多C在地上部分, 降低根系C的累积从而缩短根系寿命(
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
Aboveground overyielding in grassland mixtures is associated with reduced biomass partitioning to belowground organs
1
2009
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响
2
2014
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响
2
2014
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity
1
2008
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
The impact of ants on mineral soil properties and processes at different spatial scales
1
2008
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe
1
2013
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
Building roots in a changing environment: Implications for root longevity
2
2000
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
... ;
Nitrogen cycles: Past, present, and future
2
2004
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
The age of fine root carbon in three forests of the eastern United States measured by radiocarbon
1
2001
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
Global patterns of root turnover for terrestrial ecosystems
3
2000
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
... ).我们的研究表明, 随N添加, 根系周转率均呈增加趋势, 这与Li等(2015)通过Meta分析得到的结果相一致.主要机理是: 1) N添加增加N的有效性, 导致根系代谢活性增加(
... ;
关于氮有效性影响细根生产量和周转率的四个假说
1
2007
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
关于氮有效性影响细根生产量和周转率的四个假说
1
2007
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
Assessing the patterns and controls of fine root dynamics: An empirical test and methodological review
2
2006
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
1
1983
... id="C16">1.2.2.1 土壤样品采集与测定 于2015年8月中旬, 在3 m × 3 m样方内, 用内径5 cm的土钻分层(0-10 cm和10-20 cm)按“S”形钻取5钻, 混合为1个土壤样品, 每个处理重复6次, 并做好标记.样品风干后过筛用于测定土壤pH值、SOM、TN和AN含量.其中, 用电极法测定土壤pH值, 丘林法测定SOM含量, 凯氏定氮法测定TN含量, 康惠法测定AN含量(
1
1983
... id="C16">1.2.2.1 土壤样品采集与测定 于2015年8月中旬, 在3 m × 3 m样方内, 用内径5 cm的土钻分层(0-10 cm和10-20 cm)按“S”形钻取5钻, 混合为1个土壤样品, 每个处理重复6次, 并做好标记.样品风干后过筛用于测定土壤pH值、SOM、TN和AN含量.其中, 用电极法测定土壤pH值, 丘林法测定SOM含量, 凯氏定氮法测定TN含量, 康惠法测定AN含量(
A global budget for fine root biomass, surface area, and nutrient contents
2
1997
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响
1
2016
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响
1
2016
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
Impact of cattle on soil physical properties and nutrient concentrations in overland flow from pasture in Ireland
1
2006
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
The effects of simulated nitrogen deposition on plant root traits: A meta-analysis
3
2015
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
... ), 但是N添加降低菌根真菌侵染率(
1
1994
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China
1
2015
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests
1
2002
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
Fine root production and turnover in a Norway spruce stand in northern Sweden: Effects of nitrogen and water manipulation
3
2005
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
... id="C28">根系周转运用上述介绍的方法, 计算出单位面积根系生物量(RLDA), 在每次微根管取样时能够作为细根现存量的估计值, 每次测定的细根生产量累加来估算年生物量产量(
... id="C31">根系生产量是在t时不存在而在t + 1时所增加的新鲜根长总和, 再加上所有在t时原有根系上延长的长度来估算.根系死亡量是t时所有活根和新鲜根在t + 1时死亡和消失掉的根长总和, 根系现存量根据季节性生产和死亡的差异来计算(
Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden
1
2004
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
Restoration of riparian meadows degraded by livestock grazing: Above- and belowground responses
2002
林木细根寿命及其影响因子研究进展
2004
林木细根寿命及其影响因子研究进展
2004
施氮肥对水曲柳人工林细根生产和周转的影响
2008
施氮肥对水曲柳人工林细根生产和周转的影响
2008
The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants
1
2000
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
The potential effects of nitrogen deposition on fine-root production in forest ecosystems
3
2000
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment
1
2004
... id="C28">根系周转运用上述介绍的方法, 计算出单位面积根系生物量(RLDA), 在每次微根管取样时能够作为细根现存量的估计值, 每次测定的细根生产量累加来估算年生物量产量(
干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献
1
2011
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献
1
2011
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
氮输入对陆地生态系统碳循环关键过程的影响
1
2008
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
氮输入对陆地生态系统碳循环关键过程的影响
1
2008
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
Simulated chronic nitrogen deposition increases carbon storage in northern temperate forests
1
2008
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
Atmospheric CO2, soil nitrogen and turnover of fine roots
2
1995
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
Nitrogen deposition and atmospheric CO2 interactions on fine root dynamics in temperate forests: A theoretical model analysis
1
2002
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
Global nitrogen deposition and carbon sinks
1
2008
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
Effects of resource additions on species richness and ANPP in an alpine meadow community
1
2010
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
Quantitative estimates of root densities at minirhizotrons differ from those in the bulk soil
2012
A new fiber optic technique for measuring root growth of soybeans under field conditions
1
1978
... id="C22">式中, L (m)为微根窗中观察到的细根根长; A (m2)为观测窗面积(图像大小); DOF (m)为微根管到周围土壤的距离, DOF一般为0.002-0.003 m (
Foliage and fine root longevity in pines
1
1994
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
Different inter-annual responses to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing
1
2012
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
Measurement of soil-grown roots in a rhizotronl
1
1970
... id="C22">式中, L (m)为微根窗中观察到的细根根长; A (m2)为观测窗面积(图像大小); DOF (m)为微根管到周围土壤的距离, DOF一般为0.002-0.003 m (
Increased tree carbon storage in response to nitrogen deposition in the US
1
2010
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
Evaluating minirhizotron estimates of fine root longevity and production in the forest floor of a temperate broadleaf forest
1
2001
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
华西雨屏区苦竹细根分解对模拟氮沉降的响应
1
2014
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
华西雨屏区苦竹细根分解对模拟氮沉降的响应
1
2014
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
Root life spans of four grass species from habitats differing in nutrient availability
1
2002
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests
1
1986
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species
1
1995
... id="C5">N增加对植物根系的影响主要通过直接破坏根系组织和间接改变土壤N的有效性来实现(
CO2 enrichment and warming of the atmosphere enhance both productivity and mortality of maple tree fine roots
1
2004
... id="C47">根系是植物的重要器官, 除具有贮藏营养物质、供给植物营养和水分、调节植物生长发育、支撑植物体等基本功能外, 还对植物生产力的形成和生态系统功能(如固C、养分循环和物质分解等)的正常发挥产生重要的影响(
Fertilization and litter effects on the functional group biomass, species diversity of plants, microbial biomass, and enzyme activity of two alpine meadow communities
1
2010
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
施肥梯度对高寒草甸群落结构、功能和土壤质量的影响
1
2013
... id="C7">研究区位于四川省阿坝藏族羌族自治州红原县, 地理坐标32.83° N, 102.60° E, 海拔3 494 m.该区属于大陆性高原寒温带半湿润季风气候, 日温差大, 霜冻期长, 四季变化不明显, 年平均气温1.1 ℃, 最冷月(1月)平均气温为-10.3 ℃, 最热月(7月)平均气温为10.9 ℃, 平均5 ℃以下的月份长达7个月.干湿季节分明, 年降水量650-800 mm, 主要集中在5-9月; 日照时间长, 太阳辐射强, 年日照时间2 417.9 h, 年平均相对湿度71%.草甸类型主要是矮嵩草草甸, 植被盖度80%以上.其中四川嵩草(Kobresia setchwanensis)和高山嵩草(Kobresia pygmaea)是主要的莎草科植物, 三颖披碱草(Elymus nutans)和剪股颖(Agrostis matsumurae)是主要的禾本科植物, 而杂类草主要是匙叶银莲花(Anemone trullifolia)、蕨麻(Potentilla anserina)、钝苞雪莲(Saussurea nigrescens)、美丽龙胆(Gentiana formosa)和高山唐松草(Thalictrum alpinum)占优.该地区土壤类型是亚高山草甸土, 其土层深度达40 cm以上, 表层(0-10 cm)土壤SOM含量110.92 g·kg-1, TN含量6.38 g·kg-1, AN含量343.00 mg·kg-1, pH值为5.74 (
施肥梯度对高寒草甸群落结构、功能和土壤质量的影响
1
2013
... id="C7">研究区位于四川省阿坝藏族羌族自治州红原县, 地理坐标32.83° N, 102.60° E, 海拔3 494 m.该区属于大陆性高原寒温带半湿润季风气候, 日温差大, 霜冻期长, 四季变化不明显, 年平均气温1.1 ℃, 最冷月(1月)平均气温为-10.3 ℃, 最热月(7月)平均气温为10.9 ℃, 平均5 ℃以下的月份长达7个月.干湿季节分明, 年降水量650-800 mm, 主要集中在5-9月; 日照时间长, 太阳辐射强, 年日照时间2 417.9 h, 年平均相对湿度71%.草甸类型主要是矮嵩草草甸, 植被盖度80%以上.其中四川嵩草(Kobresia setchwanensis)和高山嵩草(Kobresia pygmaea)是主要的莎草科植物, 三颖披碱草(Elymus nutans)和剪股颖(Agrostis matsumurae)是主要的禾本科植物, 而杂类草主要是匙叶银莲花(Anemone trullifolia)、蕨麻(Potentilla anserina)、钝苞雪莲(Saussurea nigrescens)、美丽龙胆(Gentiana formosa)和高山唐松草(Thalictrum alpinum)占优.该地区土壤类型是亚高山草甸土, 其土层深度达40 cm以上, 表层(0-10 cm)土壤SOM含量110.92 g·kg-1, TN含量6.38 g·kg-1, AN含量343.00 mg·kg-1, pH值为5.74 (
Significant contribution of combustion-related emissions to the atmospheric phosphorus budget
1
2015
... id="C4">氮(N)是影响植物生长的最基本元素, 同时也是自然陆地生态系统的主要限制性元素, 在植物生长、发育和繁殖过程中具有重要的作用(
Simultaneous electrochemical determination of hydroquinone, catechol and resorcinol at Nafion/multi-walled carbon nanotubes/carbon dots/multi-walled carbon nanotubes modified glassy carbon electrode
1
2014
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
The representation of root processes in models addressing the responses of vegetation to global change
1
2000
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
高寒草甸植被细根生产和周转的比较研究
1
2014
... id="C22">式中, L (m)为微根窗中观察到的细根根长; A (m2)为观测窗面积(图像大小); DOF (m)为微根管到周围土壤的距离, DOF一般为0.002-0.003 m (
高寒草甸植被细根生产和周转的比较研究
1
2014
... id="C22">式中, L (m)为微根窗中观察到的细根根长; A (m2)为观测窗面积(图像大小); DOF (m)为微根管到周围土壤的距离, DOF一般为0.002-0.003 m (
冬小麦根系时空分布动态及产量对不同氮源配施的响应
1
2011
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
冬小麦根系时空分布动态及产量对不同氮源配施的响应
1
2011
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
4
2014
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
... id="C7">研究区位于四川省阿坝藏族羌族自治州红原县, 地理坐标32.83° N, 102.60° E, 海拔3 494 m.该区属于大陆性高原寒温带半湿润季风气候, 日温差大, 霜冻期长, 四季变化不明显, 年平均气温1.1 ℃, 最冷月(1月)平均气温为-10.3 ℃, 最热月(7月)平均气温为10.9 ℃, 平均5 ℃以下的月份长达7个月.干湿季节分明, 年降水量650-800 mm, 主要集中在5-9月; 日照时间长, 太阳辐射强, 年日照时间2 417.9 h, 年平均相对湿度71%.草甸类型主要是矮嵩草草甸, 植被盖度80%以上.其中四川嵩草(Kobresia setchwanensis)和高山嵩草(Kobresia pygmaea)是主要的莎草科植物, 三颖披碱草(Elymus nutans)和剪股颖(Agrostis matsumurae)是主要的禾本科植物, 而杂类草主要是匙叶银莲花(Anemone trullifolia)、蕨麻(Potentilla anserina)、钝苞雪莲(Saussurea nigrescens)、美丽龙胆(Gentiana formosa)和高山唐松草(Thalictrum alpinum)占优.该地区土壤类型是亚高山草甸土, 其土层深度达40 cm以上, 表层(0-10 cm)土壤SOM含量110.92 g·kg-1, TN含量6.38 g·kg-1, AN含量343.00 mg·kg-1, pH值为5.74 (
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
4
2014
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
... id="C7">研究区位于四川省阿坝藏族羌族自治州红原县, 地理坐标32.83° N, 102.60° E, 海拔3 494 m.该区属于大陆性高原寒温带半湿润季风气候, 日温差大, 霜冻期长, 四季变化不明显, 年平均气温1.1 ℃, 最冷月(1月)平均气温为-10.3 ℃, 最热月(7月)平均气温为10.9 ℃, 平均5 ℃以下的月份长达7个月.干湿季节分明, 年降水量650-800 mm, 主要集中在5-9月; 日照时间长, 太阳辐射强, 年日照时间2 417.9 h, 年平均相对湿度71%.草甸类型主要是矮嵩草草甸, 植被盖度80%以上.其中四川嵩草(Kobresia setchwanensis)和高山嵩草(Kobresia pygmaea)是主要的莎草科植物, 三颖披碱草(Elymus nutans)和剪股颖(Agrostis matsumurae)是主要的禾本科植物, 而杂类草主要是匙叶银莲花(Anemone trullifolia)、蕨麻(Potentilla anserina)、钝苞雪莲(Saussurea nigrescens)、美丽龙胆(Gentiana formosa)和高山唐松草(Thalictrum alpinum)占优.该地区土壤类型是亚高山草甸土, 其土层深度达40 cm以上, 表层(0-10 cm)土壤SOM含量110.92 g·kg-1, TN含量6.38 g·kg-1, AN含量343.00 mg·kg-1, pH值为5.74 (
... id="C48">根系的动态变化不仅体现在现存量的变化上, 而且体现在根系生产量、死亡量和周转率等方面.先前的研究认为细根的生产和死亡约占全球年净初级生产的30% (
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
Storage, patterns and controls of soil organic carbon in the Tibetan grassland
1
2008
... id="C46">青藏高原高寒草甸生态系统N素的输入主要来源于自然N沉降和人为施N (
The effects of long-term fertilization on the temporal stability of alpine meadow communities
1
2011
... id="C7">研究区位于四川省阿坝藏族羌族自治州红原县, 地理坐标32.83° N, 102.60° E, 海拔3 494 m.该区属于大陆性高原寒温带半湿润季风气候, 日温差大, 霜冻期长, 四季变化不明显, 年平均气温1.1 ℃, 最冷月(1月)平均气温为-10.3 ℃, 最热月(7月)平均气温为10.9 ℃, 平均5 ℃以下的月份长达7个月.干湿季节分明, 年降水量650-800 mm, 主要集中在5-9月; 日照时间长, 太阳辐射强, 年日照时间2 417.9 h, 年平均相对湿度71%.草甸类型主要是矮嵩草草甸, 植被盖度80%以上.其中四川嵩草(Kobresia setchwanensis)和高山嵩草(Kobresia pygmaea)是主要的莎草科植物, 三颖披碱草(Elymus nutans)和剪股颖(Agrostis matsumurae)是主要的禾本科植物, 而杂类草主要是匙叶银莲花(Anemone trullifolia)、蕨麻(Potentilla anserina)、钝苞雪莲(Saussurea nigrescens)、美丽龙胆(Gentiana formosa)和高山唐松草(Thalictrum alpinum)占优.该地区土壤类型是亚高山草甸土, 其土层深度达40 cm以上, 表层(0-10 cm)土壤SOM含量110.92 g·kg-1, TN含量6.38 g·kg-1, AN含量343.00 mg·kg-1, pH值为5.74 (
水曲柳和落叶松细根寿命的估计
1
2007
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
水曲柳和落叶松细根寿命的估计
1
2007
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响
2
2010
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响
2
2010
... id="C6">青藏高原高寒草甸地处高海拔地区, 是我国重要的草地畜牧业基地, 也是全球气候变化的敏感区域和生物多样性重点保护区.因为它远离工业区,在大气N沉降持续增加背景下, 遭受的人为破坏相对较少, 大气N沉降量远远小于N污染严重的美国东部和西欧(
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
三工河流域两种琵琶柴群落细根生物量、分解与周转
1
2014
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
三工河流域两种琵琶柴群落细根生物量、分解与周转
1
2014
... id="C49">根系周转不仅是生态系统营养动力学和C储量的重要组成部分, 同时也是维持植物初级生产力的重要汇(
1
2001
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(
1
2001
... id="C50">许多研究证明, 高寒草甸随施N量的增加, 地上生产力呈先增加后降低的趋势(

{kind=link}
{kind=link}
{kind=link}
{kind=link}