 ,1,2,*1
,1,2,*1 2
3
Responses of exchangeable base cations to continuously increasing nitrogen addition in alpine steppe: A case study of Stipa purpurea steppe
QIN Shu-Qi1,2, FANG Kai1,2, WANG Guan-Qin1,2, PENG Yun-Feng1, ZHANG Dian-Ye1,2, LI Fei1,2, ZHOU Guo-Ying3, YANG Yuan-He,1,2,*1 2
and 3
通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1266KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
秦书琪, 房凯, 王冠钦, 彭云峰, 张典业, 李飞, 周国英, 杨元合. 高寒草原土壤交换性盐基离子对氮添加的响应: 以紫花针茅草原为例. 植物生态学报[J], 2018, 42(1): 95-104 doi:10.17521/cjpe.2017.0100
QIN Shu-Qi.
土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(Lucas et al., 2011)。一方面, Ca2+、Mg2+、K+是植物生长的必需营养元素, 发挥着重要的生理功能。其中, Ca2+参与组成细胞壁(Kinzel, 1989), 维持细胞膜结构与功能, 并起到调节生物酶活性以及植物对环境变化响应的作用(Poovaiah & Reddy, 1993; Marschner, 1995; 蒋廷惠等, 2005; 姬飞腾等, 2009); Mg2+位于叶绿素分子中心, 在叶绿体结构维持、酶活化、蛋白质合成等过程中扮演重要角色(汪洪和褚天铎, 1999); K+同样参与酶活性调节与蛋白质合成, 此外还可维持细胞电荷平衡、调节细胞膨压(王毅和武维华, 2009); Na+虽不是所有植物生长必需的营养元素, 但它有利于某些特殊植物类群如C4植物、景天酸代谢途径(CAM)植物及盐生植物的生长, 同时由于Na+与K+的相似性, 它可在缺K+的环境中代替K+ (Pilon-Smits et al., 2009)。鉴于Ca2+、Mg2+、K+、Na+在植物中的重要作用, 土壤中这些营养元素的缺失将影响植物的吸收利用, 降低植物生产力, 并使植物对干旱、冰冻及病虫害的抗性减弱(DeHayes et al., 1999; Demchik & Sharpe, 2000; Watmough & Dillon, 2003)。另一方面, 作为土壤胶体吸附的可交换性阳离子, 4种盐基离子均可为H+提供交换位点, 从而交换土壤酸化过程中产生的H+, 缓解土壤急剧酸化对生态系统造成的危害(Brady & Weil, 2002)。一般来讲, 盐基离子对酸化的缓冲主要发生在pH值4.5-7.5范围内(Bowman et al., 2008; Yang et al., 2012b), 其含量决定了此阶段的缓冲能力, 即高盐基离子含量代表更多H+交换位点与更强的缓冲能力(Lu et al., 2015)。当盐基离子随土壤pH值降低而逐渐耗尽时, 土壤进入Al3+、Mn2+缓冲阶段, 这些元素进入土壤溶液将对植物生长产生毒害作用(Blake et al., 1999; Horswill et al., 2008; Lieb et al., 2011)。因此, 土壤交换性盐基离子在维持土壤养分与缓冲土壤酸化中的作用不容忽视。
工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(Galloway et al., 2008; Bodirsky et al., 2014)。按照目前的估计, 2000-2010年全球氮沉降超过50 kg·hm-2 (Penuelas et al., 2013; Tian & Niu, 2015), 随之产生的土壤酸化、NO3-淋溶、NH4+交换以及植物加速生长等过程使土壤交换性盐基离子含量降低(Lucas et al., 2011), 已经引起了学术界的广泛关注。已有的研究表明, 温带与寒温带生态系统中氮沉降导致的土壤酸化使盐基离子含量显著降低(Saarsalmi & Malkonen, 2001; Huntington, 2005; H?gberg et al., 2006)。已有的研究还发现, 与温带相比, 热带生态系统土壤交换性盐基离子对氮沉降的响应更为剧烈。热带地区土壤高度风化, 酸性强, 盐基离子含量低, 且植物生长不受氮限制, 持续氮输入导致的酸化将耗竭盐基离子并使土壤进入非盐基离子(Al3+、Mn2+)缓冲阶段(Lu et al., 2014; Cusack et al., 2016)。这些研究很大程度上促进了学术界对土壤盐基离子对氮沉降响应的认识, 然而目前的相关研究绝大多数集中在酸性土中。与酸性土不同, 碱性土中主要由碳酸盐缓冲氮沉降引起的土壤酸化(Bowman et al., 2008)。这一过程并不会造成盐基离子的大量损失, 使得学术界在很长一段时间内忽略了在碱性土中开展相关研究。但是, 目前尚不清楚氮沉降是否会通过其他途径对碱性土中盐基离子造成影响, 进而改变土壤养分供应与缓冲能力。因此, 了解全球变化尤其是氮沉降背景下碱性土中交换性盐基离子的含量与动态对准确评估生态系统结构与功能对全球变化的响应具有重要的科学意义, 也在草原生态系统可持续管理等方面具有重要的实践价值。
青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元。由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(中国科学院中国植被图编辑委员会, 2001; 张蓓蓓等, 2016)。并且, 高寒草地分布区土壤大多呈碱性, pH值处于6.0-10.1之间(附件I; Ding et al., 2016)。此外, 高寒草地分布区经历了自西向东逐渐增强的氮沉降, 平均年沉降量达8.36 kg·hm-2 (Zhu et al., 2016), 为研究碱性土中盐基离子对氮沉降的响应提供了理想平台。为此, 本研究选择青海湖北岸典型的高寒草原生态系统作为研究对象, 依托氮添加控制实验平台, 通过测定不同施氮水平下盐基离子含量, 并分析其与各土壤、植物因素之间的相关关系, 从而探讨土壤交换性盐基离子对氮添加的响应及其可能原因, 以期为认识大气氮沉降背景下碱性土中土壤交换性盐基离子的动态变化提供直接证据。
1 材料和方法
1.1 样地概况
试验地点位于青海省刚察县三角城种羊场。地理位置为37.30° N, 100.25° E, 海拔3β290 m。该地区寒冷干燥, 1980-2012年年平均气温为0.08 ℃, 年降水量为387 mm。植被类型为青藏高原典型的高寒草原, 优势种为紫花针茅(Stipa purpurea), 细叶薹草(Carex duriuscula subsp. stenophylloides), 其他伴生物种包括冷地早熟禾(Poa crymophila), 冰草(Agropyron cristatum), 以及阿尔泰狗娃花(Heteropappus altaicus)等。土壤类型为栗钙土, 其中黏粒含量4.9%, 粉粒含量33.4%, 砂粒含量61.7% (Peng et al., 2017b)。试验开始前0-30 cm土壤总氮含量为2.5 g·kg-1, NO3--N含量11.5 mg·kg-1, NH4+-N含量为0.1 mg·kg-1 (Peng et al., 2017a)。1.2 试验设计
为了模拟连续氮输入对高寒草原生态系统结构和功能的影响, 于2013年5月在上述研究点建立了氮添加控制实验平台。正式氮处理开始前, 用围栏围封试验样地以减少放牧等干扰对实验造成的影响。施肥处理于2013年5月开始, 所施氮肥形态为NH4NO3。试验中共设置8个氮水平, 施氮量依次为:0、1、2、4、8、16、24、32 g·m-2·a-1, 分别用N0、N1、N2、N4、N8、N16、N24、N32表示。试验采用随机区组设计, 共设置5个重复。每个小区大小为6 m × 6 m, 小区之间间隔1 m, 区组之间间隔2 m。施肥时间为每年5、6、7、8及9月月初。施肥主要使用肩背式喷雾器进行: 将每个样地所需的NH4NO3溶于10 L水中, 不断搅拌至全部溶解。每个小区来回均匀喷施两次, 对照小区(N0)喷洒等量清水(Peng et al., 2017a)。需要说明的是, 本试验中添加的10 L水相当于约0.3 mm降雨, 对植物生长的影响基本可以忽略。
1.3 样品采集与分析
于2014、2015与2016年8月中旬采集土壤与植物样品。在每个小区内随机选择3个25 cm × 25 cm的小样方, 采集植物地上部分。然后, 在收获植物地上部分后的小样方中用直径3 cm的土钻采集0-10 cm的土壤样品。将3个点的土壤样品混合均匀, 挑去根系等, 并过2 mm筛。植物样品于65 ℃烘干至恒质量用于估算地上生物量。土壤pH值依据电位法选择1:2.5的土水比测定(Lu et al., 2014)。将土壤用1 mol·L-1 KCl溶液浸提后, 采用连续流动分析仪(Autoanalyzer 3 SEAL, Bran and Luebbe, Norderstedt, Germany)测定土壤NO3--N与NH4+-N含量, 二者之和表示土壤总无机氮(TIN)含量(Peng et al., 2017a)。依据全国农业技术推广服务中心(2006)报道的方法, 用70%的乙醇溶液洗去土壤中易溶氯化物和硫酸盐等, 然后用氯化铵-乙醇交换液提取, 制得交换性Ca2+、Mg2+、K+、Na+的待测液, 用电感耦合等离子体光谱仪(ICP-OES; iCAP 6300, Thermo Scientific, Waltham, USA)测定其含量。4种交换性盐基离子的和表示总的交换性盐基离子。
1.4 数据处理
数据处理主要包括以下步骤: 首先采用非参数K-S检验法检验各变量的正态性, 然后采用单因素方差分析与最小显著差异法(LSD)比较不同氮处理下各生物与非生物因素的差异, 最后, 采用回归分析分析盐基离子与各生物、非生物因素之间的关系。显著水平为0.05。所有的统计分析均在SPSS 20.0 (IBM SPSS, Chicago, Illinois, USA)进行, 图表绘制均在Origin 9.1 (OriginLab, Northampton, Massachusetts, USA)中完成。2 研究结果
2.1 氮添加对各生物与非生物因素的影响
随着施氮量的增加, 土壤理化性质与植物生物量发生了显著变化(表1)。2014年, 土壤pH值在各处理间无显著差异, 但从2015年开始, 施氮显著降低了pH值。与对照相比, N32处理分别使2015与2016年pH值下降了0.2与0.3。在实验进行的3年期间, 施氮显著增加了土壤中无机氮含量。从2014到2016年, 土壤无机氮含量分别从对照的34.3、9.5、32.0 mg·kg-1增加至N32处理下的45.5、69.6、41.1 mg·kg-1。同时, 随着施氮量的增加, 2014-2016年植物地上生物量均显著增加。当施氮量达到32 g·m-2·a-1时, 3年的地上生物量比对照分别增加了124.5%、155.5%和108.9%。Table 1
表1
表1不同氮水平下土壤与植物基本理化性质(平均值±标准误差)
Table 1
| 年 Year | N0 | N1 | N2 | N4 | N8 | N16 | N24 | N32 | |
|---|---|---|---|---|---|---|---|---|---|
| pH值 pH value | 2014 | 8.9 ± 0.10b | 9.2 ± 0.06a | 9.2 ± 0.08a | 9.1 ± 0.07ab | 9.1 ± 0.08ab | 9.1 ± 0.07ab | 9.0 ± 0.03ab | 9.1 ± 0.10ab |
| 2015 | 9.0 ± 0.07bc | 9.1 ± 0.06a | 9.0 ± 0.03ab | 9.0 ± 0.05ab | 8.9 ± 0.03cd | 9.0 ± 0.04ab | 8.8 ± 0.04d | 8.8 ± 0.05d | |
| 2016 | 8.5 ± 0.11a | 8.5 ± 0.05a | 8.5 ± 0.09a | 8.4 ± 0.07ab | 8.3 ± 0.07bc | 8.6 ± 0.02a | 8.2 ± 0.05c | 8.2 ± 0.05c | |
| TIN (mg·kg-1) | 2014 | 34.3 ± 2.6cd | 37.8 ± 3.1bcd | 35.9 ± 3.7bcd | 33.1 ± 0.9d | 52.6 ± 9.1a | 46.8 ± 3.4ab | 43.5 ± 1.8abcd | 45.5 ± 1.4abc |
| 2015 | 9.5 ± 0.3e | 11.4 ± 1.5e | 11.8 ± 1.1e | 11.9 ± 0.7e | 22.4 ± 2.5d | 41.8 ± 2.5c | 56.7 ± 5.0b | 69.6 ± 7.1a | |
| 2016 | 32.0 ± 0.9d | 32.3 ± 1.5cd | 31.2 ± 2.0d | 32.9 ± 2.0cd | 37.4 ± 1.9bc | 41.4 ± 2.8ab | 43.2 ± 3.0a | 41.1 ± 2.1ab | |
| AGB (g·m-2) | 2014 | 126.9 ± 10.0d | 142.2 ± 17.8d | 152.4 ± 13.9cd | 186.7 ± 13.9c | 256.6 ± 14.3ab | 241.2 ± 21.4b | 271.7 ± 26.1ab | 284.9 ± 27.6a |
| 2015 | 146.0 ± 11.6c | 146.8 ± 8.3c | 175.7 ± 14.7c | 180.4 ± 13.6c | 265.1 ± 23.5b | 287.5 ± 21.0b | 350.4 ± 13.3a | 373.0 ± 18.6a | |
| 2016 | 93.7 ± 9.3e | 117.0 ± 17.8de | 127.0 ± 19.3cde | 144.2 ± 9.3bcd | 181.2 ± 18.4ab | 161.9 ± 8.3abc | 190.2 ± 21.1a | 195.7 ± 7.1a |
新窗口打开|下载CSV
2.2 氮添加对土壤交换性盐基离子的影响
2014-2016年间, 土壤总交换性盐基离子对氮添加的响应并不完全相同(图1)。2014年沿施氮梯度总交换性盐基离子含量呈降低趋势, 但并不显著。2015年盐基离子含量沿施氮梯度显著降低, N32处理下盐基离子含量比对照降低了27.1%。2016年各施氮量下盐基离子含量虽存在差异, 但主要表现为N1、N2处理下盐基离子含量高于对照, 其余各处理与对照无显著差异。图1
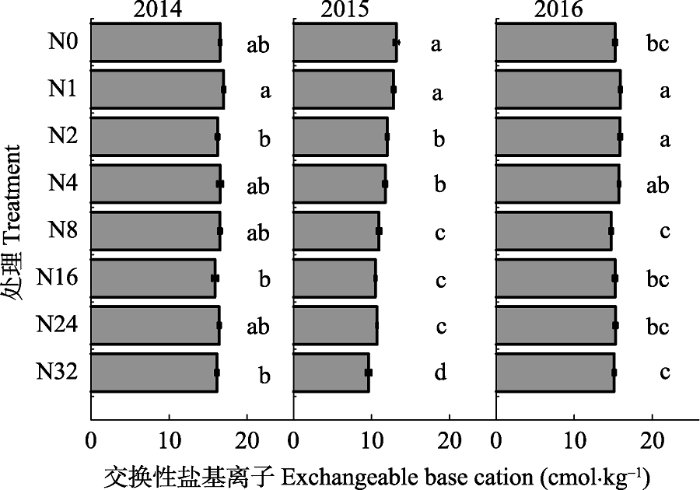 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1氮添加对土壤交换性盐基离子含量的影响(平均值±标准误差)。N0-N32, 氮添加量分别为: 0、1、2、4、8、16、24、32 g·m-2·a-1。不同字母表示各施氮量下差异显著(p < 0.05)。
Fig. 1Effects of nitrogen addition on soil exchangeable base cations (mean ± SE). N0-N32, nitrogen addition 0, 1, 2, 4, 8, 16, 24, 32 g·m-2·a-1, respectively. Different letters indicate significant differences among treatments (p < 0.05).
不同年份中4种交换性盐基离子(Ca2+、Mg2+、K+、Na+)对氮添加的响应也存在差异(图2)。其中, 交换性Mg2+在3年中都随施氮量增加显著降低(图2B)。交换性Na+在2014与2015年沿施氮梯度显著降低, 2016年降低并不显著(图2D)。交换性Ca2+在2014与2016年随施氮量增加显著增加, 而在2015年却显著降低(图2A), 这可能是由于在2014与2016年, CaCO3与施氮后产生的H+反应释放大量Ca2+, 同时Ca2+在这两年的损失量小于生成量, 而在2015年的损失量大于生成量。对于交换性K+, 2014年N1处理下K+显著低于N4到N32处理, 其余各处理间均无显著差异, 2015年沿施氮梯度K+显著降低, 2016年各施氮量下K+则无显著差异(图2C)。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2氮添加对土壤交换性Ca2+、Mg2+、K+、Na+的影响(平均值±标准误差)。A, Ca2+。B, Mg2+。C, K+。D, Na+。N0-N32, 氮添加量分别为: 0、1、2、4、8、16、24、32 g·m-2·a-1。不同字母表示各施氮量下差异显著(p < 0.05)。
Fig. 2Effects of nitrogen addition on soil exchangeable Ca2+, Mg2+, K+, Na+ (mean ± SE)。A, Ca2+. B, Mg2+. C, K+. D, Na+. N0-N32, nitrogen addition 0, 1, 2, 4, 8, 16, 24, 32 g·m-2·a-1, respectively. Different letters indicate significant differences among treatments (p < 0.05).
2.3 氮添加背景下土壤交换性盐基离子变化的影响因素
2015年总交换性盐基离子与植物地上生物量(R2 = 0.69, p < 0.001, 图3A)、土壤无机氮含量(R2 = 0.56, p < 0.001, 图3B)均呈显著负相关关系。与此类似, 3年的交换性Mg2+与植物地上生物量(2014: R2 = 0.16, p < 0.05, 图4A; 2015: R2 = 0.48, p < 0.001, 图4C; 2016: R2 = 0.20, p < 0.01, 图4E )、土壤无机氮含量(2014: R2 = 0.12, p < 0.05, 图4B; 2015: R2 = 0.35, p < 0.001, 图4D; 2016: R2 = 0.29, p < 0.001, 图4F)呈显著负相关关系。2014年(R2 = 0.21, p < 0.01, 图5A)与2015年(R2 = 0.35, p < 0.001, 图5B)的交换性Na+与土壤无机氮含量也呈显著负相关关系。图3
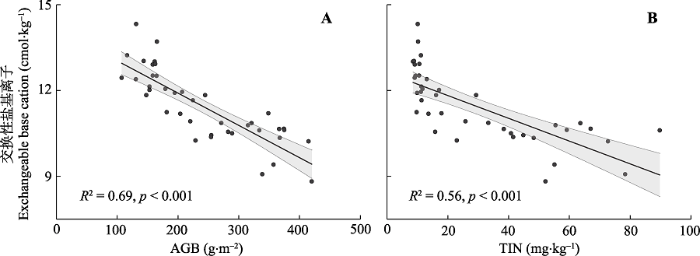 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图32015年交换性盐基离子与地上生物量和土壤总无机氮含量的关系。A, 地上生物量(AGB)。B, 土壤总无机氮(TIN)。直线与阴影部分分别表示拟合曲线与95%置信区间。
Fig. 3Relationships of soil exchangeable base cations with above-ground biomass and soil total inorganic nitrogen in 2015. A, above-ground biomass (AGB). B, soil total inorganic nitrogen (TIN). The black lines represent the fitted curves and shades for 95% confidence intervals.
图4
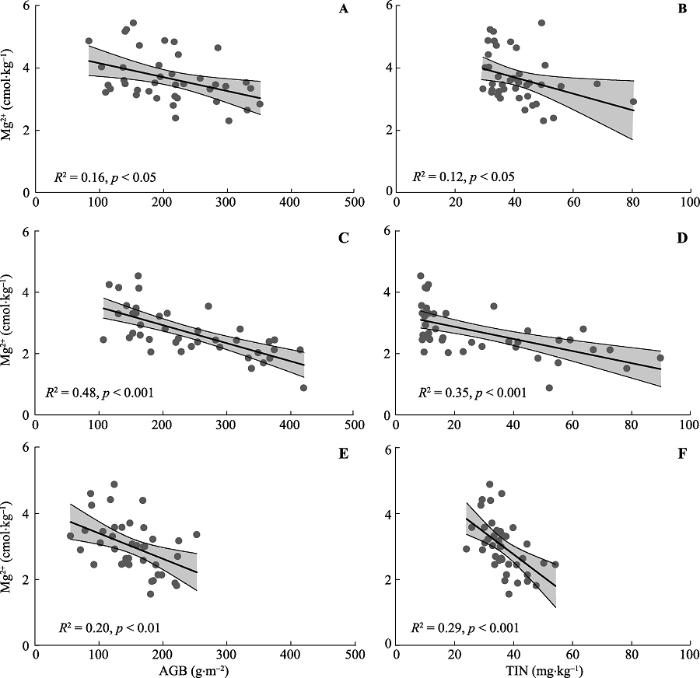 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同年份土壤交换性Mg2+与地上生物量以及土壤总无机氮含量的关系。A, 2014年AGB。B, 2014年TIN。C, 2015年AGB。D, 2015年TIN。E, 2016年AGB。F, 2016年TIN。AGB, 地上生物量, TIN, 土壤总无机氮。直线与阴影部分分别表示拟合曲线与95%置信区间。
Fig. 4Relationships of soil exchangeable Mg2+ with above-ground biomass and soil total inorganic nitrogen in different years. A, AGB in 2014. B, TIN in 2014. C, AGB in 2015. D, TIN in 2015. E, AGB in 2016. F, TIN in 2016. AGB, above-ground biomass, TIN, soil total inorganic nitrogen. The black lines represent the fitted curves and shades for 95% confidence intervals.
图5
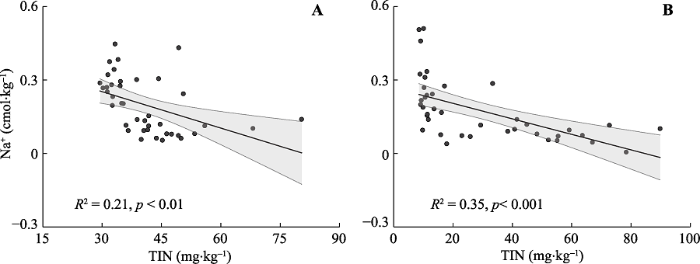 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同年份土壤交换性Na+与土壤总无机氮含量的关系。A, 2014年。B, 2015年。TIN, 土壤总无机氮。直线与阴影部分分别表示拟合曲线与95%置信区间。
Fig. 5Relationships of soil exchangeable Na+ with soil total inorganic nitrogen in different years. A, 2014. B, 2015. TIN, soil total inorganic nitrogen. The black lines represent the fitted curves and shades for 95% confidence intervals.
3 讨论
本研究结果显示, 氮添加造成土壤交换性盐基离子, 尤其是Mg2+与Na+的减少。造成这种现象的原因主要有以下两点。首先, 氮添加促进植物生长, 加速其对土壤中营养元素的吸收, 进而导致盐基离子含量下降。作为植物生长所需的营养元素, 氮素是生态系统生产力的重要限制因素(LeBauer & Treseder, 2008)。氮添加可增加土壤中氮的可利用性, 提高植物生产力(Xia & Wan, 2008)。和以往的研究类似, 紫花针茅(Stipa purpurea)草原地上生物量沿施氮梯度显著增加(表1), 且这种现象在2015年尤为明显。植物生长加速使其对营养元素的需求增大, 进而导致土壤中交换性Ca2+、K+的含量在2015年沿施氮梯度显著降低, Mg2+含量在3年间均显著下降(图2)。有趣的是, 我们的结果显示盐基离子(总量与Mg2+)与植物地上生物量呈显著负相关关系(图3A、图4A, 4C, 4E), 同时植物地上部分Mg库随施氮量增加而显著增大(附件II), 进一步说明生物量增加促进了植物对盐基离子的吸收。需要指出的是, 不同年份元素响应的差异可能由生物量及植物需求的年际差异所致。并且, 由于植物对Na+的吸收主要发生在C4植物、CAM植物及盐生植物中(Pilon-Smits et al., 2009), 植物吸收并不是本研究中观察到的Na+损失的主要原因。此外, 我们还发现2016年总盐基离子含量高于2015年(图1), 这可能是因为盐基离子可随植物凋落物的分解重新进入土壤, 而2016年的植物生物量不及2015年高, 对盐基离子的吸收也相对较少, 导致其含量升高。由此也可看出, 在围封保护的草原生态系统中, 植物吸收能够引起盐基离子在土壤与植物间的重新分配, 但不会造成其大量损失(Lucas et al., 2011)。然而, 在植物生长迅速且有收获的生态系统中, 植物吸收可能造成盐基离子的大量损失且不可逆。例如Zhang等(2016)发现, 重庆地区烟草地上生物量的收割从土壤中移除的盐基离子量高达339.23 kg·hm-2·a-1。其次, 氮添加导致土壤无机氮含量增加, NH4+与NO3-通过不同途径造成盐基离子损失。Liu等(2013)的研究表明, NH4+-N是氮沉降的主要形式, 但是自20世纪80年代以来, NO3--N的沉降速率也在持续增加, 两者共同导致了日益加剧的氮沉降。由此可以看出, 本研究所选氮肥形态(NH4NO3)能较好地模拟大气氮沉降。随着施氮量增加, 土壤中无机氮含量显著增加(表1)。增加的NH4+与土壤有更强的吸附性(Matschonat & Matzner, 1996), 可以交换土壤吸附的盐基离子, 使盐基离子进入土壤溶液。由于土壤对4种盐基离子的吸附性大小依次为Ca2+ > Mg2+ > K+ > Na+ (Chapin III et al., 2011), 使得Na+最容易被交换, 这也可能是在2年中均观察到Na+含量随施氮量增加显著降低的原因(图2D)。同时, 以往的研究表明当生态系统中氮添加量超过2.5 g·m-2·a-1时, NO3-淋溶显著增加(Dise & Wright, 1995)。为了揭示氮添加是否引起土壤NO3-淋溶量的变化, 我们于2014与2015年测定了实验地深层土壤中的NO3-含量(Peng et al., 2017a), 并且于2016年用树脂法测定了表层0-10 cm的NO3-淋溶(附件III), 发现当氮添加量超过16 g·m-2·a-1时, NO3-淋溶显著增加。为保持土壤溶液电荷平衡, NO3-淋溶时将带走包括4种盐基离子在内的等电荷的阳离子(Gundersen et al., 2006), 从而造成盐基离子的损失。与上述推断一致, 本实验的结果表明盐基离子含量与无机氮含量呈显著负相关(图3B、图4B, 4D, 4F、图5), 进一步证明无机氮增加促进了盐基离子的损失。该结果也得到以往研究的支持, Cusack等(2016)发现, 氮添加使土壤中可交换性NH4+的含量与土壤溶液中NO3-含量增加,证实NH4+交换与NO3-淋溶能造成土壤盐基离子的损失。
虽然我们在碱性土中观察到的“土壤交换性盐基离子含量因氮添加显著下降”的现象与酸性土一致, 但导致两类土壤中盐基离子动态变化的驱动因素并不完全相同。这主要是因为两类土壤对土壤酸化的缓冲机制存在差异(Bowman et al., 2008)。在不同生态系统中, 氮沉降可通过以下途径引起土壤酸化: (1)伴随NO3-发生的H+沉降; (2)植物吸收NH4+释放等量H+; (3) NH4+在硝化作用过程中被氧化, 产生H+; (4)氮添加促进植物吸收盐基离子, 释放更多H+ (Guo et al., 2010; Chen et al., 2016)。一般而言, 土壤初始pH值决定了其所处的缓冲阶段。当初始pH值大于7.5时, 土壤酸化主要由碳酸盐缓冲; pH值为4.5-7.5时, 土壤表面吸附的交换性盐基离子与H+交换, 中和土壤中增加的H+。随酸化加剧, 盐基离子逐渐被消耗, 土壤吸附的Al3+开始起缓冲作用(pH值小于4.5)(Bowman et al., 2008; Yang et al., 2012b)。本研究中, 紫花针茅草原土壤初始pH值约达到9, 持续的氮添加下, pH值仍大于8, 一直处于碳酸盐主导缓冲的阶段, 因此氮添加导致的土壤酸化并非造成盐基离子损失的主要途径。而在酸性土中, 盐基离子须参与交换氮沉降产生的H+, 缓冲土壤酸化, 是其损失的主要途径。例如, Chen等(2016)在内蒙古草地生态系统中的研究结果显示, 长期氮添加导致的酸化使土壤进入盐基离子缓冲阶段而造成盐基离子损失。Bowman等(2008)在斯洛伐克高寒生态系统中的研究也表明, 持续氮添加在使土壤从Al3+向Fe3+缓冲过渡的同时, 造成交换性盐基离子的损失, 由此增强土壤毒性, 降低养分可利用性。总之, 在不同的pH值条件下盐基离子的损失途径不同。
4 结论和意义
本研究揭示了碱性土中盐基离子对连续氮输入的响应及其损失途径。结果发现, 碱性土中氮输入增加了植物对盐基离子的吸收, 同时导致NH4+交换作用与NO3-淋溶增强, 进而加速盐基离子损失。与酸性土不同, 氮沉降背景下土壤酸化造成的盐基离子损失在碱性土中的作用相对较弱。这一发现对认识高寒草原生态系统对全球变化的响应等方面具有重要启示。按照目前的预测, 在未来40年中我国的氮沉降仍将维持在较高水平(Galloway et al., 2008), 预计将造成草地土壤的持续酸化(Yang et al., 2012b; Tian & Niu, 2015)。在酸化过程中, 碳酸盐缓冲释放的碳会改变区域碳平衡(Yang et al., 2012a)。同时, 若连续氮添加造成的盐基离子的损失得不到及时补充, 将减弱土壤养分供应能力, 限制植被生产力(H?gberg et al., 2006), 在较长的时间尺度上还将影响土壤缓冲能力。因此, 在未来草原生态系统可持续管理中, 应当重视氮沉降引起的土壤盐基离子变化。附件
附件I
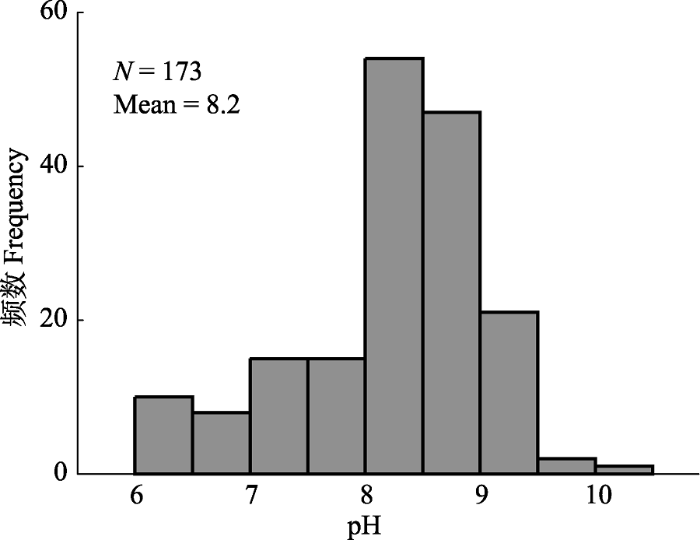 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附件I青藏高原高寒草地表层土壤pH值的频度分布。数据来自杨元合课题组于2013-2014年间在青藏高原草地样带调查的173个样地
Appendix IFrequency distribution of pH values on the Qinghai-Xizang Plateau. Data from 173 sampling sites along a grassland transect across Qinghai-Xizang alpine grasslands during 2013-2014, which were collected by members from Dr. Yuanhe Yang’s group
附件II
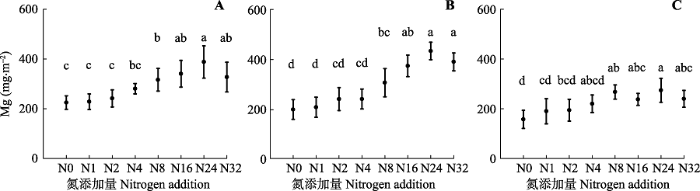 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附件II氮添加对植物地上部分Mg库的影响(平均值±标准误差)。A, 2014年。B, 2015年。C, 2016年。N0-N32, 氮添加量分别为: 0、1、2、4、8、16、24、32 g·m-2·a-1。不同字母表示各施氮量下差异显著(p < 0.05)
Appendix IIEffects of nitrogen addition on Mg pool in aboveground plant (mean ± SE). A, 2014. B, 2015. C, 2016. N0-N32, nitrogen addition 0, 1, 2, 4, 8, 16, 24, 32 g·m -2·a -1, respectively. Different letters indicate significant differences among treatments (p < 0.05)
附件III
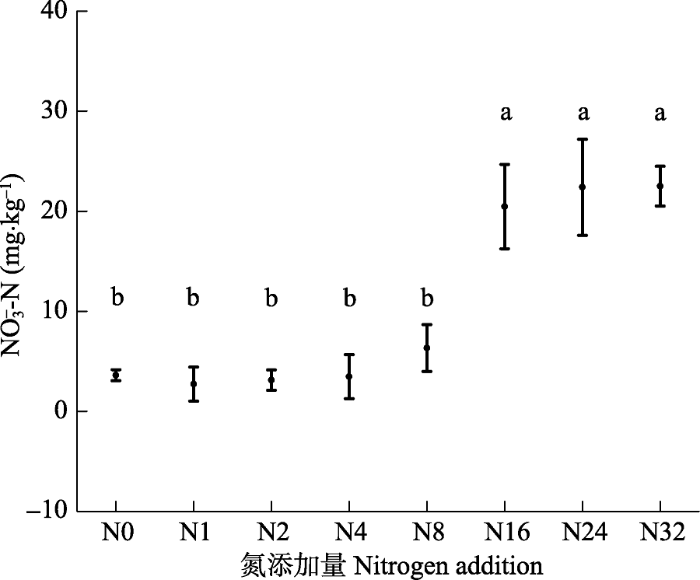 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附件III2016年氮添加对表层0-10 cm土壤NO3-淋溶量的影响(平均值±标准误差)。N0-N32, 氮添加量分别为: 0、1、2、4、8、16、24、32 g·m-2·a-1。不同字母表示各施氮量下差异显著(p < 0.05)
Appendix IIIEffects of nitrogen addition on NO3- leaching in top 0-10 cm soil in 2016 (mean ± SE). N0-N32, nitrogen addition 0, 1, 2, 4, 8, 16, 24, 32 g·m -2·a -1, respectively. Different letters indicate significant differences among treatments (p < 0.05)

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1046/j.1365-2389.1999.00253.xURL [本文引用: 1]

Summary We have examined the effect that acid deposition and other sources of acidity have had over the last 110鈥140 years on soil under woodland (Broadbalk and Geescroft Wildernesses) and grassland (Park Grass) comprising some of the Classical Experiments at Rothamsted Experimental Station. Changes in soil chemistry have been followed by analysing some of the unique archive of stored samples for pH, water-soluble and exchangeable base cations, aluminium, iron and manganese, exchangeable acidity, cation exchange capacity (CEC) and soluble anions. Proton balances and historical data show the importance of acid deposition to acidification and concomitant changes in the chemistry of the soil. The pH of the surface soil of Geescroft Wilderness has fallen from 6.2 to 3.8 since 1883. The decrease in the pH of the unlimed, unfertilized plot on Park Grass was less over a similar period (from pH 5.2 to 4.2), illustrating the significant effect of the woodland canopy on the interception of acidifying pollutants. The effect of increasing acidity on the soil chemistry of Geescroft Wilderness is seen in its decreasing base saturation and CEC, with base cations moving down the soil profile. Clay minerals are being irreversibly weathered, and Mn and Al progressively mobilized, so that today Al occupies 70% of the exchange complex in the surface soil. Even with present reductions in sulphur deposition critical loads for sulphur, nitrogen and acidity are still exceeded. Such semi-natural ecosystems are unsustainable under the current climate of pollution.
DOI:10.1038/ncomms4858URLPMID:24819889 [本文引用: 1]
Reactive nitrogen (Nr) is an indispensable nutrient for agricultural production and alimentation. Simultaneously, agriculture is the largest contributor to Nr pollution, causing severe damages to health and ecosystem services. The trade-off between food availability and Nr pollution can be attenuated by several key mitigation options, including Nr efficiency improvements in crop and animal production systems, food waste reduction in households and lower consumption of Nr-intensive animal products. However, their quantitative mitigation potential remains unclear, especially under the added pressure of population growth and changes in food consumption. Here we show by model simulations, that under baseline conditions, Nr pollution in 2050 can be expected to rise to 102-156% of the 2010 value. Only under ambitious mitigation, does pollution possibly decrease to 36-76% of the 2010 value. Air, water and atmospheric Nr pollution go far beyond critical environmental thresholds without mitigation actions. Even under ambitious mitigation, the risk remains that thresholds are exceeded.
DOI:10.1038/ngeo339URL [本文引用: 4]
Anthropogenic nitrogen deposition over the past half century has had a detrimental impact on temperate ecosystems in Europe and North America, resulting in soil acidification and a reduction in plant biodiversity. During the acidification process, soils release base cations, such as calcium and magnesium, neutralizing the increase in acidity. Once these base cations have been depleted, aluminium is released from the soils, often reaching toxic levels. Here, we present results from a nitrogen deposition experiment that suggests that a long legacy of acid deposition in the Western Tatra Mountains of Slovakia has pushed soils to a new threshold of acidification usually associated with acid mine drainage soils. We show that increases in nitrogen deposition in the region result in a depletion of both base cations and soluble aluminium, and an increase in extractable iron concentrations. In conjunction with this, we observe a nitrogen-deposition-induced reduction in the biomass of vascular plants, associated with a decrease in shoot calcium and magnesium concentrations. We suggest that this site, and potentially others in central Europe, have reached a new and potentially more toxic level of soil acidification in which aluminium release is superseded by iron release into soil water.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/1365-2435.12525URL [本文引用: 1]
Summary Terrestrial ecosystems worldwide are receiving increasing amounts of biologically reactive nitrogen (N) as a consequence of anthropogenic activities. This intended or unintended fertilization can have a wide-range of impacts on biotic communities and hence on soil respiration. Reduction in below-ground carbon (C) allocation induced by high N availability has been assumed to be a major mechanism determining the effects of N enrichment on soil respiration. In addition to increasing available N, however, N enrichment causes soil acidification, which may also affect root and microbial activities. The relative importance of increased N availability vs. soil acidification on soil respiration in natural ecosystems experiencing N enrichment is unclear. We conducted a 12-year N enrichment experiment and a 4-year complementary acid addition experiment in a semi-arid Inner Mongolian grassland. We found that N enrichment had contrasting effects on root and microbial respiration. N enrichment significantly increased root biomass, root N content and specific root respiration, thereby promoting root respiration. In contrast, N enrichment significantly suppressed microbial respiration likely by reducing total microbial biomass and changing the microbial community composition. The effect on root activities was due to both soil acidity and increased available N, while the effect on microbes primarily stemmed from soil acidity, which was further confirmed by results from the acid addition experiment. Our results indicate that soil acidification exerts a greater control than soil N availability on soil respiration in grasslands experiencing long-term N enrichment. These findings suggest that N-induced soil acidification should be included in predicting terrestrial ecosystem C balance under future N deposition scenarios.
DOI:10.1007/s10533-016-0195-7URL [本文引用: 1]
Rates of atmospheric nitrogen (N) deposition are increasing rapidly in tropical regions, which are projected to receive some of the greatest deposition levels globally in the coming decades. Tropical...
DOI:10.2307/1313570URL [本文引用: 1]
Forest ecosystems throughout the world are exposed to acid rain, a complex solution consisting largely of H+, SO42-, NH4+, and NO3- pollutant ions derived from sulfur and nitrogen oxides. Although the public in the United States may view acid deposition as a localized pollution issue specific to certain regions of North America and northern Europe, recent evidence of acidic deposition throughout much of Asia, including tropical forest regions, has demonstrated that acid deposition is actually a global phenomenon (Abate 1995).
DOI:10.1016/S0378-1127(99)00307-2URL [本文引用: 1]
Unexplained mortality of northern red oak ( Quercus rubra L.) has occurred on the Laurel Hill in southwestern Pennsylvania. We attempted to characterize this mortality and determine its relationship to soil acidity, soil nutrient status and droughts. Three blocks with high and low mortality of northern red oak were selected. Basal area increment, terminal elongation and leaf mass were measured along with soil chemistry. Basal area growth patterns were compared to historic drought records. Both living and standing dead trees on high mortality plots had been responsive to most of these droughts. After 2 years of droughts in the 1960s, basal area growth rate of trees on high mortality plots did not recover. In addition, the standing dead trees on high mortality plots were historically a less vigorous subset. After this mortality, the living trees did not respond with increased growth due to thinning of competitors. Basal area growth on low mortality plots did recover after these same droughts and mortality was limited. In addition, the trees on the high mortality plots showed foliar deficiency of K and low levels of Ca while trees on the low mortality plots did not. Furthermore, the A- and B- horizons of the soils on the high mortality plots had lower levels of soil bases, higher levels of available Al and lower Ca/Al ratios. The results of this study suggest that nutrient deficiency and associated Al toxicity may be causing stress for northern red oaks and is related to the mortality and reduced growth of northern red oak in this region.
DOI:10.1111/gcb.13257URLPMID:26913840 [本文引用: 1]
Abstract The permafrost organic carbon (OC) stock is of global significance because of its large pool size, and the potential positive feedback to climate warming. However, due to the lack of systematic field observations and appropriate upscaling methodologies, substantial uncertainties exist in the permafrost OC budget, which limits our understanding of the fate of frozen carbon in a warming world. In particular, the lack of comprehensive estimates of OC stocks across alpine permafrost means that current knowledge on this issue remains incomplete. Here we evaluated the pool size and spatial variations of permafrost OC stock to 3 metres depth on the Tibetan Plateau by combining systematic measurements from a substantial number of pedons (i.e., 342 three-metre-deep cores and 177 50-cm-deep pits) with a machine learning technique (i.e., support vector machine, SVM). We also quantified uncertainties in permafrost carbon budget by conducting a Monte Carlo simulation. Our results revealed that the combination of systematic measurements with the SVM model allowed spatially explicit estimates to be made. The OC density (OC amount per unit area, OCD) exhibited a decreasing trend from the southeastern to the northwestern plateau, with the exception that OCD in the swamp meadow was substantially higher than that in surrounding regions. Our results also demonstrated that Tibetan permafrost stored a large amount of OC in the top 3 metres, with the median OC pool size being 15.31 Pg C (interquartile range: 13.03-17.77 Pg C). 44% of OC occurred in deep layers (i.e., 100-300 cm), close to the proportion observed across the northern circumpolar permafrost region. The large carbon pool size, together with significant permafrost thawing suggests a risk of carbon emissions and positive climate feedback across the Tibetan alpine permafrost region. This article is protected by copyright. All rights reserved. This article is protected by copyright. All rights reserved.
DOI:10.1016/0378-1127(94)06092-WURL [本文引用: 1]
Forests in Europe currently receive inorganic nitrogen deposition (wet and dry) ranging from less than 1 to more than 75 kg N ha 611 year 611 . A survey of nitrogen output from 65 forested plots and catchments throughout Europe indicated a three-stage response to these inputs. Below a deposition threshold of about 10 kg N ha 611 year 611 no significant nitrogen leaching occurred from the forests; at intermediate levels of 10–25 kg N ha 611 year 611 leaching occurred at some sites; above 25 kg ha 611 year 611 significant leaching occurred at all sites. NITREX sites followed this larger European pattern. Among 41 different variables tested in the survey, output-N (seepage or runoff) was most highly correlated with input-N ( r 2 =0.69), followed by input-SO 4 , soil pH (negative correlation), percent slope, bedrock type and latitude (negative correlation). The latter three, however, were highly intercorrelated with input-N. A combination of input-N and soil pH explained 87% of the variability in output-N ( N = 20) and predicted output-N successfully from NITREX sites and other European test sites.
DOI:10.1126/science.1136674URLPMID:18487183 [本文引用: 2]
Abstract Humans continue to transform the global nitrogen cycle at a record pace, reflecting an increased combustion of fossil fuels, growing demand for nitrogen in agriculture and industry, and pervasive inefficiencies in its use. Much anthropogenic nitrogen is lost to air, water, and land to cause a cascade of environmental and human health problems. Simultaneously, food production in some parts of the world is nitrogen-deficient, highlighting inequities in the distribution of nitrogen-containing fertilizers. Optimizing the need for a key human resource while minimizing its negative consequences requires an integrated interdisciplinary approach and the development of strategies to decrease nitrogen-containing waste.
DOI:10.1139/a05-015URL [本文引用: 1]
DOI:10.1126/science.1182570URL [本文引用: 1]
DOI:10.1111/j.1365-2486.2006.01102.xURL [本文引用: 2]
Abstract Relations among nitrogen load, soil acidification and forest growth have been evaluated based on short-term (<15 years) experiments, or on surveys across gradients of N deposition that may also include variations in edaphic conditions and other pollutants, which confound the interpretation of effects of N per se . We report effects on trees and soils in a uniquely long-term (30 years) experiment with annual N loading on an un-polluted boreal forest. Ammonium nitrate was added to replicated ( N =3) 0.09ha plots at two doses, N1 and N2, 34 and 68kgNha 611 yr 611 , respectively. A third treatment, N3, 108kgNha 611 yr 611 , was terminated after 20 years, allowing assessment of recovery during 10 years. Tree growth initially responded positively to all N treatments, but the longer term response was highly rate dependent with no gain in N3, a gain of 50m 3 ha 611 stemwood in N2 and a gain of 100m 3 ha 611 stemwood in excess of the control (N0) in N1. High N treatments caused losses of up to 70% of exchangeable base cations (Ca 2+ , Mg 2+ , K + ) in the mineral soil, along with decreases in pH and increases in exchangeable Al 3+ . In contrast, the organic mor-layer (forest floor) in the N-treated plots had similar amounts per hectare of exchangeable base cations as in the N0 treatment. Magnesium was even higher in the mor of N-treated plots, providing evidence of up-lift by the trees from the mineral soil. Tree growth did not correlate with the soil Ca/Al ratio (a suggested predictor of effects of soil acidity on tree growth). A boron deficiency occurred on N-treated plots, but was corrected at an early stage. Extractable NH 4 + and NO 3 61 were high in mor and mineral soils of on-going N treatments, while NH 4 + was elevated in the mor only in N3 plots. Ten years after termination of N addition in the N3 treatment, the pH had increased significantly in the mineral soil; there were also tendencies of higher soil base status and concentrations of base cations in the foliage. Our data suggest the recovery of soil chemical properties, notably pH, may be quicker after removal of the N-load than predicted. Our long-term experiment demonstrated the fundamental importance of the rate of N application relative to the total amount of N applied, in particular with regard to tree growth and C sequestration. Hence, experiments adding high doses of N over short periods do not mimic the long-term effects of N deposition at lower rates.
DOI:10.1016/j.envpol.2007.11.006URLPMID:18164110 [本文引用: 1]
Pollutant nitrogen deposition effects on soil and foliar element concentrations were investigated in acidic and limestone grasslands, located in one of the most nitrogen and acid rain polluted regions of the UK, using plots treated for 8–10 years with 35–140 kg N ha 612 y 611 as NH 4NO 3. Historic data suggests both grasslands have acidified over the past 50 years. Nitrogen deposition treatments caused the grassland soils to lose 23–35% of their total available bases (Ca, Mg, K, and Na) and they became acidified by 0.2–0.4 pH units. Aluminium, iron and manganese were mobilised and taken up by limestone grassland forbs and were translocated down the acid grassland soil. Mineral nitrogen availability increased in both grasslands and many species showed foliar N enrichment. This study provides the first definitive evidence that nitrogen deposition depletes base cations from grassland soils. The resulting acidification, metal mobilisation and eutrophication are implicated in driving floristic changes.
DOI:10.1139/x05-034URL [本文引用: 1]
Forest harvesting and acidic deposition can cause substantial decreases in the calcium (Ca) inventory of forest soils if such losses are not replenished through mineral weathering, atmospheric deposition, or fertilization. The net balance between losses and gains defines the forest Ca status. Site-specific studies have measured Ca pools and fluxes in Maine forests, but no synthesis has been published. In this paper, I review the literature on forest Ca and assess the current status and potential future trends. Forest soils in Maine are currently at lesser risk of Ca depletion compared with many forest soils in the central and southeastern United States, because levels of acidic deposition and rates of Ca accumulation in trees are lower in Maine. The rate of Ca accumulation in trees is reduced in Maine as a result of lower growth rates and a higher proportion of conifer trees that require less Ca than hardwoods. However, field-scale biogeochemical studies in Maine and New Hampshire, and regional estimates of harvest removals and soil inventories coupled with low weathering estimates, indicate that Ca depletion is a realistic concern in Maine. The synthesis of site-specific and regional data for Maine in conjunction with the depletion measured directly in surrounding areas indicates that the Ca status of many forest soils in Maine is likely declining. Ca status could decrease further in the future if forest growth rates increase in response to climate trends and recovery from insect-induced mortality and excessive harvesting in recent years. Proposed climate change induced reductions in spruce and fir and increases in hardwoods would also increase the risk of Ca depletion. ?? 2005 NRC.
DOI:10.3773/j.issn.1005-264x.2009.05.012URL [本文引用: 1]
喀斯特地区土壤的高钙含量是影 响该地区植物生理特征的最重要环境因素之一。高钙影响植物的光合作用、生长速率及磷代谢,从而限制了许多物种在该地区的分布。选取贵州4个石漠化程度不同 的地区,测定采集地内45种优势种或常见种的地上部分和地下部分的全钙含量以及土壤的交换性钙含量。通过分析喀斯特地区植物与土壤钙含量的特征发现:喀斯 特地区植物具有较高的钙含量平均值;土壤交换性钙含量对植物地上部分钙含量的影响总体上不显著,对植物地下部分钙含量的影响显著;不同类别植物的钙含量存 在显著差异,蕨类植物地上部分钙含量平均值明显低于被子植物;不同类别植物钙的分布部位也存在显著差异,在蕨类植物和单子叶植物中地上部分和地下部分的钙 含量相近,而双子叶植物的地上部分钙含量明显高于地下部分。分析了喀斯特地区14种优势灌木和草本植物地上部分与地下部分钙含量的差异性以及与土壤交换性 钙含量的相关关系,以此为根据将14种优势植物对土壤高钙的适应方式分为3种类型:随遇型、高钙型和低钙型。随遇型植物的钙含量主要受土壤交换性钙含量影 响,其地上部分和地下部分的钙含量均与土壤交换性钙含量成显著正相关关系;高钙型植物具有较强的钙富集能力,其地上部分即使在低钙含量的土壤中也可维持较 高的钙含量;低钙型植物的地上部分即使在高钙含量的土壤中亦可维持较低的钙含量。对植物适应钙的不同方式的研究可用于筛选退化生态系统恢复所需的植物资 源。
DOI:10.3773/j.issn.1005-264x.2009.05.012URL [本文引用: 1]
喀斯特地区土壤的高钙含量是影 响该地区植物生理特征的最重要环境因素之一。高钙影响植物的光合作用、生长速率及磷代谢,从而限制了许多物种在该地区的分布。选取贵州4个石漠化程度不同 的地区,测定采集地内45种优势种或常见种的地上部分和地下部分的全钙含量以及土壤的交换性钙含量。通过分析喀斯特地区植物与土壤钙含量的特征发现:喀斯 特地区植物具有较高的钙含量平均值;土壤交换性钙含量对植物地上部分钙含量的影响总体上不显著,对植物地下部分钙含量的影响显著;不同类别植物的钙含量存 在显著差异,蕨类植物地上部分钙含量平均值明显低于被子植物;不同类别植物钙的分布部位也存在显著差异,在蕨类植物和单子叶植物中地上部分和地下部分的钙 含量相近,而双子叶植物的地上部分钙含量明显高于地下部分。分析了喀斯特地区14种优势灌木和草本植物地上部分与地下部分钙含量的差异性以及与土壤交换性 钙含量的相关关系,以此为根据将14种优势植物对土壤高钙的适应方式分为3种类型:随遇型、高钙型和低钙型。随遇型植物的钙含量主要受土壤交换性钙含量影 响,其地上部分和地下部分的钙含量均与土壤交换性钙含量成显著正相关关系;高钙型植物具有较强的钙富集能力,其地上部分即使在低钙含量的土壤中也可维持较 高的钙含量;低钙型植物的地上部分即使在高钙含量的土壤中亦可维持较低的钙含量。对植物适应钙的不同方式的研究可用于筛选退化生态系统恢复所需的植物资 源。
URL [本文引用: 1]
随着酸雨的蔓延和化学氮肥用量的增加,土壤酸化日趋严重,钙流失不断加剧,而含钙肥料却因高浓度复合肥比重的增加(磷铵取代含钙磷肥)而不断下降,使钙供应与植物需要的矛盾日渐突出.文中简述了钙对植物抗酸、盐、氨、重金属、渍害、冷害、热害、病害及酸雨等能力的影响,以及维持土壤钙供应的措施,指出植物生长过程中分泌的酸是施肥引起土壤酸化的主要原因.根系分泌酸能力强的田菁改良盐碱土的重要机理之一是促进土壤钙的活化,增强后季作物的抗盐能力.降水中氮素含量的增加,促进了林木的生长,并使其他元素缺乏,易加剧根际酸化,可能是一些工业化地区森林退化的重要机制.基于钙在植物抗逆中的作用,建议在肥料试验设计中增加施钙的空白对照区,以便将钙的效应从其它处理效应中区分出来.磷铵不应作为我国磷肥工业的发展方向.
URL [本文引用: 1]
随着酸雨的蔓延和化学氮肥用量的增加,土壤酸化日趋严重,钙流失不断加剧,而含钙肥料却因高浓度复合肥比重的增加(磷铵取代含钙磷肥)而不断下降,使钙供应与植物需要的矛盾日渐突出.文中简述了钙对植物抗酸、盐、氨、重金属、渍害、冷害、热害、病害及酸雨等能力的影响,以及维持土壤钙供应的措施,指出植物生长过程中分泌的酸是施肥引起土壤酸化的主要原因.根系分泌酸能力强的田菁改良盐碱土的重要机理之一是促进土壤钙的活化,增强后季作物的抗盐能力.降水中氮素含量的增加,促进了林木的生长,并使其他元素缺乏,易加剧根际酸化,可能是一些工业化地区森林退化的重要机制.基于钙在植物抗逆中的作用,建议在肥料试验设计中增加施钙的空白对照区,以便将钙的效应从其它处理效应中区分出来.磷铵不应作为我国磷肥工业的发展方向.
DOI:10.1016/S0367-2530(17)30398-5URL [本文引用: 1]
Abstract Current knowledge of Ca uptake and modes of transport in higher plants were discussed. The relationships of Ca both within the cell and in relation to metabolic movement along transport paths throughout the plant were considered. It was stressed that Ca flows into the plant for the most part passively with the water flow of transpiration. The deposition of this often surplus amount of Ca is accomplished by various means in different physiological types of plant.
DOI:10.1890/06-2057.1URLPMID:18409427 [本文引用: 1]
Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R ANPPN/ANPPctrl) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within cerlain biome types.
DOI:10.1016/j.geoderma.2011.06.013URL [本文引用: 1]
78 N deposition decreased soil pH and acid neutralizing capacity of alpine soils. 78 N deposition decreased extractable Mg and increased extractable Mn and Al. 78 Small-scale variation in soil geochemistry influenced the responses to nitrogen deposition.
DOI:10.1038/nature11917URLPMID:23426264
Abstract China is experiencing intense air pollution caused in large part by anthropogenic emissions of reactive nitrogen. These emissions result in the deposition of atmospheric nitrogen (N) in terrestrial and aquatic ecosystems, with implications for human and ecosystem health, greenhouse gas balances and biological diversity. However, information on the magnitude and environmental impact of N deposition in China is limited. Here we use nationwide data sets on bulk N deposition, plant foliar N and crop N uptake (from long-term unfertilized soils) to evaluate N deposition dynamics and their effect on ecosystems across China between 1980 and 2010. We find that the average annual bulk deposition of N increased by approximately 8090009kilograms of nitrogen per hectare (P090009<0900090.001) between the 1980s (13.2090009kilograms of nitrogen per hectare) and the 2000s (21.1090009kilograms of nitrogen per hectare). Nitrogen deposition rates in the industrialized and agriculturally intensified regions of China are as high as the peak levels of deposition in northwestern Europe in the 1980s, before the introduction of mitigation measures. Nitrogen from ammonium (NH4(+)) is the dominant form of N in bulk deposition, but the rate of increase is largest for deposition of N from nitrate (NO3(-)), in agreement with decreased ratios of NH3 to NOx emissions since 1980. We also find that the impact of N deposition on Chinese ecosystems includes significantly increased plant foliar N concentrations in natural and semi-natural (that is, non-agricultural) ecosystems and increased crop N uptake from long-term-unfertilized croplands. China and other economies are facing a continuing challenge to reduce emissions of reactive nitrogen, N deposition and their negative effects on human health and the environment.
DOI:10.1111/gcb.12665URLPMID:24953639 [本文引用: 2]
Abstract Elevated anthropogenic nitrogen (N) deposition has greatly altered terrestrial ecosystem functioning, threatening ecosystem health via acidification and eutrophication in temperate and boreal forests across the northern hemisphere. However, response of forest soil acidification to N deposition has been less studied in humid tropics compared to other forest types. This study was designed to explore impacts of long-term N deposition on soil acidification processes in tropical forests. We have established a long-term N-deposition experiment in an N-rich lowland tropical forest of Southern China since 2002 with N addition as NH4 NO3 of 0, 50, 100 and 150 kg N ha(-1) yr(-1) . We measured soil acidification status and element leaching in soil drainage solution after 6-year N addition. Results showed that our study site has been experiencing serious soil acidification and was quite acid-sensitive showing high acidification (pH(H2O) <4.0), negative water-extracted acid neutralizing capacity (ANC) and low base saturation (BS,< 8%) throughout soil profiles. Long-term N addition significantly accelerated soil acidification, leading to depleted base cations and decreased BS, and further lowered ANC. However, N addition did not alter exchangeable Al(3+) , but increased cation exchange capacity (CEC). Nitrogen addition-induced increase in SOC is suggested to contribute to both higher CEC and lower pH. We further found that increased N addition greatly decreased soil solution pH at 20 cm depth, but not at 40 cm. Furthermore, there was no evidence that Al(3+) was leaching out from the deeper soils. These unique responses in tropical climate likely resulted from: exchangeable H(+) dominating changes of soil cation pool, an exhausted base cation pool, N-addition stimulating SOC production, and N saturation. Our results suggest that long-term N addition can contribute measurably to soil acidification, and that shortage of Ca and Mg should receive more attention than soil exchangeable Al in tropical forests with elevated N deposition in the future. 2014 John Wiley & Sons Ltd.
DOI:10.1021/es5047233URLPMID:25741588 [本文引用: 1]
Elevated anthropogenic nitrogen (N) deposition has become an important driver of soil acidification at both regional and global scales. It remains unclear, however, how long-term N deposition affects soil buffering capacity in tropical forest ecosystems and in ecosystems of contrasting land-use history. Here, we expand on a long-term N deposition experiment in three tropical forests that vary in land-use history (primary, secondary, and planted forests) in Southern China, with N addition as NH4NO3 of 0, 50, 100, and 150 kg N ha(-1) yr(-1), respectively. Results showed that all three forests were acid-sensitive ecosystems with poor soil buffering capacity, while the primary forest had higher base saturation and cation exchange capacity than others. However, long-term N addition significantly accelerated soil acidification and decreased soil buffering capacity in the primary forest, but not in the degraded secondary and planted forests. We suggest that ecosystem N status, influenced by different land-use history, is primarily responsible for these divergent responses. N-rich primary forests may be more sensitive to external N inputs than others with low N status, and should be given more attention under global changes in the future, because lack of nutrient cations is irreversible.
DOI:10.1016/j.foreco.2011.03.018URL [本文引用: 3]
The dominant base cations (BC; i.e., Ca 2+, Mg 2+, K +, and Na +) are important in buffering soil and water acidity in both terrestrial and aquatic ecosystems. Ca 2+, Mg 2+, and K + are also important in many plant physiological functions. Because BC availability is affected by changes in the nitrogen (N) cycle, we conducted a meta-analysis of previously published data to determine if N fertilization alters the availability of BC in terrestrial and stream ecosystems across biomes. We include data from 107 independent studies published in 62 different articles, taking a holistic perspective on BC by examining their responses to added N in plant foliage, bulk soil, soil solution, and stream water. Our results suggest N fertilization may accelerate BC loss from terrestrial ecosystems over time periods less than five years. We found that N additions resulted in an overall 24% decrease in the availability of exchangeable Ca 2+, Mg 2+, and K + in the bulk soil of boreal forest, temperate forest, and grassland biomes. Collectively, responses of BC in boreal forest, temperate forest, tropical forest, and grassland biomes increased following N fertilization by about 71% in soil solution and 48% in stream waters. Additionally, BC responses in foliage decreased in boreal forest and temperate forest biomes following N additions over time periods less than five years, but there were no significant changes over longer time periods. Despite large short-term shifts in BC responses following N additions, we did not find evidence of widespread negative impacts on ecosystems over time periods greater than five years. This analysis suggests effects of N addition on the availability of exchangeable BC may diminish over time. Although the effects on BC can be substantial over periods less than five years, there is little available evidence that N fertilization has had large-scale detrimental effects on the availability of BC needed for plant growth within terrestrial or aquatic ecosystems.
[本文引用: 1]
DOI:10.1002/jpln.1996.3581590514URL [本文引用: 1]
Abstract Fourteen European forest soils from the boreal to the mediterranean climate on different parent materials were investigated with respect to their ability to store NH4+ in exchangeable form, using sorption isotherms. Distribution coefficients for NH4+ sorption per unit weight of soil were in the range of 0.02 to 0.77. NH4+ sorption coefficients were usually highest in the forest floor of a given soil. NH4+ sorption behaviour of mineral soil horizons was correlated to soil parameters that are determined during routine soil analysis. A combination of CEC and base saturation explained up to 95% of the variability Of NH4+ sorption. In the forest floors, variability in NH4+ sorption could not be explained quantitatively from independent soil parameters. The affinity of the sorption sites for NH4+ was the most important factor for explanation of the variability in NH4+ sorption in the forest floors but was of low importance in mineral soil horizons. As NH4+ exchanges predominantly base cations, susceptibility of NH4+ to transport through the soil profile increases with Iowbase saturation of a soil as well as with low CEC values. Einflu08 bodenchemischer Parameter auf die NH4+ -Sorption in Waldb02den 14 europ01ische Waldb02den des borealen bis mediterranen Klimas auf verschiedenen Ausgangsgesteinen wurden hinsichtlich ihrer Speicherf01higkeit für austauschbar gebundenes NH4+ durch die Erstellung von Sorptionsisothermen untersucht. Die Verteilungskoefllzienten für zugegebenes NH4+ zwischen Boden und L02sung bezogen auf das Bodengewicht (Masse) lagen zwischen 0.02 and 0.77. Die NH4+ -Sorpti- onskoeffizienten waren in der Regel im Auflagehumus eines gegebenen Bodens am h02chsten. Die NH4+ -Sorptionskoeffizienten der Mineralbodenhorizonte korrelierten mit Bodenparametern, die bei der routinem0108igen Bodenuntersuchung bestimmt werden. Die KAK in Kombination mit der Basens01ttigung der B02den erkl01rte bis zu 95% der Variabilit01t der NH4+ -Sorptionskoeffizienten. In den Auflagehumus-Horizonten konnte die Variabilit01t der NH4+ -Sorption nicht auf einzelne bodenchemische Parameter zurückgeführt werden. Im Auflagehumus war die Affinit01t der Sorptionspl01tze für NH4+ die wichtigste Einflu08gr0208e zur Erkl01rung der Streuung der Sorptionskoeffizienten; sie war jedoch von untergeordneter Bedeutung in den Mineralbodenhorizonten. Da NH4+ vorwiegend basische Kationen austauscht, steigt die Wahrscheinlichkeit für eine Verlagerung von NH4+ in B02den sowohl mit abnehmender KAK als auch mit abnehmender Basens01ttigung.
[本文引用: 1]
[本文引用: 1]
DOI:10.1088/1748-9326/? aa5ba6URL [本文引用: 4]
Nitrogen (N) availability is a key regulator of carbon (C) cycling in terrestrial ecosystems. Anthropogenic N input, such as N deposition and fertilization, increases N availability in soil, which has important implications for an ecosystem's C storage and loss. Soil respiration (Rs), which is the second largest C flux from terrestrial ecosystems to the atmosphere, plays an important role in terrestrial C cycles. The direction and magnitude of the responses of Rs and its components to N addition have been widely evaluated, but it remains unclear how these processes change across multiple N addition levels. Here we conducted a two-year field experiment to examine the changes of Rs and its autotrophic respiration (Ra) and heterotrophic respiration (Rh) components along a gradient of eight N levels (0, 1 2, 4, 8, 16, 24, 32 g m(-2) yr(-1)) in a Tibetan alpine steppe, and used structural equation modeling (SEM) to explore the relative contributions of biotic and abiotic variables and their direct and indirect pathways regulating the Ra and Rh. Our results indicated that both Rs and Ra exhibited first increasing and then subsequent decreasing trends at the threshold of 8 g N m(-2) yr(-1). In contrast, the Rh declined linearly with the N addition rate continuously increasing. SEM analysis revealed that, among various environmental factors, soil temperature was the most important one modulating Rs, which not only had a direct effect on the two Rs components, but also indirectly regulated the Ra and Rh via root and microbial biomass. These findings suggest that the nonlinear response patterns of Rs should be considered for better predicting terrestrial C balance, given that anthropogenic N input to the terrestrial ecosystems is increasing continuously.
DOI:10.1111/gcb.13789URLPMID:28614594 [本文引用: 1]
Abstract Unprecedented levels of nitrogen (N) have entered terrestrial ecosystems over the past century, which substantially influences the carbon (C) exchange between the atmosphere and biosphere. Temperature and moisture are generally regarded as the0002major controllers over the N effects on ecosystem C uptake and release. N-phosphorous (P) stoichiometry regulates the growth and metabolisms of plants and soil organisms, thereby affecting many ecosystem C processes. However, it remains unclear how the N-induced shift in the plant N:P ratio affects ecosystem production and C fluxes and its relative importance. We conducted a field manipulative experiment with eight N addition levels in a Tibetan alpine steppe and assessed the influences of N on aboveground net primary production (ANPP), gross ecosystem productivity (GEP), ecosystem respiration (ER), and net ecosystem exchange (NEE); we used linear mixed-effects models to further determine the relative contributions of various factors to the N-induced changes in these parameters. Our results showed that the ANPP, GEP, ER, and NEE all exhibited nonlinear responses to increasing N additions. Further analysis demonstrated that the plant N:P ratio played a dominate role in shaping these C exchange processes. There was a positive relationship between the N-induced changes in ANPP (0200ANPP) and the plant N:P ratio (0200N:P), whereas the 0200GEP, 0200ER, and 0200NEE exhibited quadratic correlations with the 0200N:P. In contrast, soil temperature and moisture were only secondary predictors for the changes in ecosystem production and C fluxes along the N addition gradient. These findings highlight the importance of plant N:P ratio in regulating ecosystem C exchange, which is crucial for improving our understanding of C cycles under the scenarios of global N enrichment. 0008 2017 John Wiley & Sons Ltd.
DOI:10.1038/ncomms3934URLPMID:24343268 [本文引用: 1]
Abstract The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen:phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
DOI:10.1016/j.pbi.2009.04.009URLPMID:19477676 [本文引用: 2]
Abstract Aluminum (Al), cobalt (Co), sodium (Na), selenium (Se), and silicon (Si) are considered beneficial elements for plants: they are not required by all plants but can promote plant growth and may be essential for particular taxa. These beneficial elements have been reported to enhance resistance to biotic stresses such as pathogens and herbivory, and to abiotic stresses such as drought, salinity, and nutrient toxicity or deficiency. The beneficial effects of low doses of Al, Co, Na and Se have received little attention compared to toxic effects that typically occur at higher concentrations. Better understanding of the effects of beneficial elements is important to improve crop productivity and enhance plant nutritional value for a growing world population.
DOI:10.1080/07352689309701901URLPMID:11540065 [本文引用: 1]
Environmental and hormonal signals control diverse physiological processes in plants. The mechanisms by which plant cells perceive and transduce these signals are poorly understood. Understanding biochemical and molecular events involved in signal transduction pathways has become one of the most active areas of plant research. Research during the last 15 years has established that Ca2+ acts as a messenger in transducing external signals. The evidence in support of Ca2+ as a messenger is unequivocal and fulfills all the requirements of a messenger. The role of Ca2+ becomes even more important because it is the only messenger known so far in plants. Since our last review on the Ca2+ messenger system in 1987, there has been tremendous progress in elucidating various aspects of Ca2+-signaling pathways in plants. These include demonstration of signal-induced changes in cytosolic Ca2+, calmodulin and calmodulin-like proteins, identification of different Ca2+ channels, characterization of Ca2+-dependent protein kinases (CDPKs) both at the biochemical and molecular levels, evidence for the presence of calmodulin-dependent protein kinases, and increased evidence in support of the role of inositol phospholipids in the Ca2+-signaling system. Despite the progress in Ca2+ research in plants, it is still in its infancy and much more needs to be done to understand the precise mechanisms by which Ca2+ regulates a wide variety of physiological processes. The purpose of this review is to summarize some of these recent developments in Ca2+ research as it relates to signal transduction in plants.
DOI:10.1080/02827580152699358URL [本文引用: 1]
DOI:10.1088/1748-9326/10/2/024019URL [本文引用: 2]
Nitrogen (N) deposition-induced soil acidification has become a global problem. However, the response patterns of soil acidification toNaddition and the underlyingmechanisms remain far from clear. Here, we conducted ameta-analysis of 106 studies to reveal global patterns of soil acidification in responses toNaddition.We found thatNaddition significantly reduced soil pHby 0.26 on average globally. However, the responses of soil pH varied with ecosystem types,Naddition rate,Nfertilization forms, and experimental durations. Soil pH decreasedmost in grassland, whereas boreal forest was not observed a decrease toNaddition in soil acidification. Soil pH decreased linearly withNaddition rates. Addition of urea and NH4NO3 contributedmore to soil acidification thanNH4-formfertilizer. When experimental duration was longer than 20 years,Naddition effects on soil acidification diminished. Environmental factors such as initial soil pH,soil carbon and nitrogen content, precipitation, and temperature all influenced the responses of soil pH. Base cations of Ca2+,Mg2+ and K+ were critical important in buffering againstN-induced soil acidification at the early stage. However,Naddition has shifted global soils into the Al3+ buffering phase. Overall, this study indicates that acidification in global soils is very sensitive to N deposition, which is greatlymodified by biotic and abiotic factors. Global soils are now at a buffering transition from base cations (Ca2+,Mg2+ and K+) to non-base cations (Mn2+ and Al3+). This calls our attention to care about the limitation of base cations and the toxic impact of nonbase cations for terrestrial ecosystems withNdeposition.
DOI:10.3969/j.issn.1674-3466.1999.03.007URL [本文引用: 1]
总结近些年来有关镁素营养研究应用。详细综述植物对镁的吸收、镁在植物体内的分配与运输、镁的生理作用、植物体内镁与一些元素间的相互作用。
DOI:10.3969/j.issn.1674-3466.1999.03.007URL [本文引用: 1]
总结近些年来有关镁素营养研究应用。详细综述植物对镁的吸收、镁在植物体内的分配与运输、镁的生理作用、植物体内镁与一些元素间的相互作用。
DOI:10.3969/j.issn.1674-3466.2009.01.003URL [本文引用: 1]
钾是植物生长发育所必需的矿质 营养元素之一。不同种类植物的钾营养效率存在差异,已有证据表明这种差异是受遗传基因控制的。植物细胞依靠细胞膜上的各种钾转运体和通道蛋白吸收和转运钾 离子,这些膜蛋白的活性调控是植物钾营养效率调控的关键和基础。本文对植物钾营养高效性状分子遗传机制以及相关基因的分子功能和调控机制的研究进展进行了 简要评述,并讨论了改善作物钾营养高效性状的可能途径。
DOI:10.3969/j.issn.1674-3466.2009.01.003URL [本文引用: 1]
钾是植物生长发育所必需的矿质 营养元素之一。不同种类植物的钾营养效率存在差异,已有证据表明这种差异是受遗传基因控制的。植物细胞依靠细胞膜上的各种钾转运体和通道蛋白吸收和转运钾 离子,这些膜蛋白的活性调控是植物钾营养效率调控的关键和基础。本文对植物钾营养高效性状分子遗传机制以及相关基因的分子功能和调控机制的研究进展进行了 简要评述,并讨论了改善作物钾营养高效性状的可能途径。
DOI:10.1007/s10021-002-0164-yURL [本文引用: 1]
There is growing concern that available base cation pools in soil are declining in eastern North America and that some forests are approaching nitrogen (N) saturation due to the combined effects of acid deposition and harvesting. To assess these concerns, elemental mass balances for calcium (Ca), magnesium (Mg), potassium (K), and N were conducted over a 17-year period in a representative mixed hardwood forest (HP4) in the Muskoka-Haliburton region in central Ontario, Canada. On average, 76% of the N measured in bulk deposition, which is a conservative estimate of total N deposition, was retained in HP4, with tree uptake accounting for over half of the retained N. Year-to-year variations in annual NO3export were affected by climate variations, although the low annual NO3- N concentrations (80-156 g/L) suggest that HP4 is not approaching N saturation. Losses of Ca, Mg, and K in stream export plus accumulation in trees (more than 12 cm in diameter at breast height) exceeded inputs in deposition by 296, 76.2, and 53.6 kg/ha, respectively, over the 17-year period. Inclusion of mineral weathering estimates obtained using PROFILE, zirconium (Zr) depletion, and total analysis correlation failed to balance Ca losses from HP4, and calculations indicate that between 98 and 145 kg/ha (depending on mineral weathering estimate) was lost from the soil exchangeable pool between 1983 and 1999. These losses were supported by repeated field measurements, which showed that the exchangeable Ca concentrations and soil pH decreased over the 17-year period, particularly in the upper soil horizons. When mineral weathering estimates are included, mass balance calculations generally indicated that there was no net loss of Mg and K from HP4, which was confirmed by our soil measurements. At present, there is sufficient Ca in the soil exchangeable pool to sustain forest growth at HP4; however, continued losses of Ca due to leaching and harvesting at the present rate may ultimately threaten the health and productivity of the forest within just a few decades.
DOI:10.1111/j.1469-8137.2008.02488.xURLPMID:19086179 [本文引用: 1]
Summary 6168 Better understanding of the responses of terrestrial plant species under global nitrogen (N) enrichment is critical for projection of changes in structure, functioning, and service of terrestrial ecosystems. 6168 Here, a meta-analysis of data from 304 studies was carried out to reveal the general response patterns of terrestrial plant species to the addition of N. 6168 Across 456 terrestrial plant species included in the analysis, biomass and N concentration were increased by 53.6 and 28.5%, respectively, under N enrichment. However, the N responses were dependent upon plant functional types, with significantly greater biomass increases in herbaceous than in woody species. Stimulation of plant biomass by the addition of N was enhanced when other resources were improved. In addition, the N responses of terrestrial plants decreased with increasing latitude and increased with annual precipitation. 6168 Dependence of the N responses of terrestrial plants on biological realms, functional types, tissues, other resources, and climatic factors revealed in this study can help to explain changes in species composition, diversity, community structure and ecosystem functioning under global N enrichment. These findings are critical in improving model simulation and projection of terrestrial carbon sequestration and its feedbacks to global climate change, especially when progressive N limitation is taken into consideration.
DOI:10.1111/gcb.12025URL [本文引用: 1]
Soil carbon (C) stocks consist of inorganic and organic components, 1.7 times larger than the total of the C stored in vegetation and the atmosphere together. Significant soil C losses could thus offset any C sink in vegetation, creating a positive feedback to climate change. However, compared with the susceptible sensitivity of organic matter decay to climate warming, soil inorganic carbon (SIC) stocks are often assumed to be relatively stable. Here, we evaluated SIC changes across China';s grasslands over the last two decades using data from a recent regional soil survey during 20012005 and historical national soil inventory during the 1980s. Our results showed that SIC stocks in the top 10 cm decreased significantly between the two sampling periods, with a mean rate of 26.8 (95% confidence interval: 15.841.7) g C m(-2) yr(-1). The larger decreases in SIC stocks were observed in those regions with stronger soil acidification and richer soil carbonates. The lost SIC could be released to the atmosphere as carbon dioxide, redistributed to the deeper soil layer, and transferred to the nearby regions. The fraction of soil carbonates entering into the atmosphere may diminish the strength of terrestrial C sequestration and amplify the positive C-climate feedback.
DOI:10.1111/j.1365-2486.2012.02694.xURL [本文引用: 3]
Anthropogenic acid deposition may lead to soil acidification, with soil buffering capacity regulating the magnitude of any soil pH change. However, little evidence is available from large-scale observations. Here, we evaluated changes in soil pH across northern China's grasslands over the last two decades using soil profiles obtained from China's Second National Soil Inventory during the 1980s and a more recent regional soil survey during 2001 2005. A transect from the central-southern Tibetan Plateau to the eastern Inner Mongolian Plateau, where Kriging interpolation provided robust predictions of the spatial distribution of soil pH, was then selected to examine pH changes during the survey period. Our results showed that soil pH in the surface layer had declined significantly over the last two decades, with an overall decrease of 0.63 units (95% confidence interval = 0.54 0.73 units). The decline of soil pH was observed in both alpine grasslands on the Tibetan Plateau and temperate grasslands on the Inner Mongolian Plateau. Soil pH decreased more intensively in low soil carbonate regions, while changes of soil pH showed no significant associations with soil cation exchange capacity. These results suggest that grassland soils across northern China have experienced significant acidification from the 1980s to 2000s, with soil carbonates buffering the increase in soil acidity. The buffering process may induce a large loss of carbon from soil carbonates and thus alter the carbon balance in these globally important ecosystems.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11356-015-5673-2URLPMID:26566613
Changes in soil exchangeable cations relative to soil acidification are less studied particularly under long-term cash crop plantation. This study investigated soil acidification in an Ali-Periudic...
DOI:10.1002/2016JG003393URL [本文引用: 1]
Atmospheric wet nitrogen (N) and phosphorus (P) depositions are important sources of bioavailable N and P, and the input of N and P and their ratios significantly influences nutrient availability and balance in terrestrial as well as aquatic ecosystems. Here we monitored atmospheric P depositions by measuring monthly dissolved P concentration in rainfall at 41 field stations in China. Average deposition fluxes of N and P were 13.69 8.69 kg N haa(our previous study) and 0.21 0.17 kg P haa, respectively. Central and southern China had higher N and P deposition rates than northwest China, northeast China, Inner Mongolia, or Qinghai-Tibet. Atmospheric N and P depositions showed strong seasonal patterns and were dependent upon seasonal precipitation. Fertilizer and energy consumption were significantly correlated with N deposition but less correlated with P deposition. The N:P ratios of atmospheric wet deposition (with the average of 77 40, by mass) were negatively correlated with current soil N:P ratios in different ecological regions, suggesting that the imbalanced atmospheric N and P deposition will alter nutrient availability and strengthen P limitation, which may further influence the structure and function of terrestrial ecosystems. The findings provide the assessments of both wet N and P deposition and their N:P ratio across China and indicate potential for strong impacts of atmospheric deposition on broad range of terrestrial ecosystems.
Changes in soil chemistry accompanying acidification over more than 100 years under woodland and grass at Rothamsted Experimental Station, UK
1
1999
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Reactive nitrogen requirements to feed the world in 2050 and potential to mitigate nitrogen pollution
1
2014
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
Negative impact of nitrogen deposition on soil buffering capacity
4
2008
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
... id="C23">虽然我们在碱性土中观察到的“土壤交换性盐基离子含量因氮添加显著下降”的现象与酸性土一致, 但导致两类土壤中盐基离子动态变化的驱动因素并不完全相同.这主要是因为两类土壤对土壤酸化的缓冲机制存在差异(
... 开始起缓冲作用(pH值小于4.5)(
The Nature and Properties of Soils
1
2002
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Principles of Terrestrial Ecosystem Ecology.
1
2011
... id="C22">其次, 氮添加导致土壤无机氮含量增加, NH4+与NO3-通过不同途径造成盐基离子损失.Liu等(2013)的研究表明, NH4+-N是氮沉降的主要形式, 但是自20世纪80年代以来, NO3--N的沉降速率也在持续增加, 两者共同导致了日益加剧的氮沉降.由此可以看出, 本研究所选氮肥形态(NH4NO3)能较好地模拟大气氮沉降.随着施氮量增加, 土壤中无机氮含量显著增加(
Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment
1
2016
... id="C23">虽然我们在碱性土中观察到的“土壤交换性盐基离子含量因氮添加显著下降”的现象与酸性土一致, 但导致两类土壤中盐基离子动态变化的驱动因素并不完全相同.这主要是因为两类土壤对土壤酸化的缓冲机制存在差异(
Nitrogen additions mobilize soil base cations in two tropical forests
1
2016
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
Acid rain impacts on calcium nutrition and forest health— Alteration of membrane-associated calcium leads to membrane destabilization and foliar injury in red spruce
1
1999
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
The effect of soil nutrition, soil acidity and drought on northern red oak (Quercus rubra L.) growth and nutrition on Pennsylvania sites with high and low red oak mortality.
1
2000
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
The permafrost carbon inventory on the Tibetan Plateau: A new evaluation using deep sediment cores
1
2016
... id="C6">青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元.由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(
Nitrogen leaching from European forests in relation to nitrogen deposition
1
1995
... id="C22">其次, 氮添加导致土壤无机氮含量增加, NH4+与NO3-通过不同途径造成盐基离子损失.Liu等(2013)的研究表明, NH4+-N是氮沉降的主要形式, 但是自20世纪80年代以来, NO3--N的沉降速率也在持续增加, 两者共同导致了日益加剧的氮沉降.由此可以看出, 本研究所选氮肥形态(NH4NO3)能较好地模拟大气氮沉降.随着施氮量增加, 土壤中无机氮含量显著增加(
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions
2
2008
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
... id="C24">本研究揭示了碱性土中盐基离子对连续氮输入的响应及其损失途径.结果发现, 碱性土中氮输入增加了植物对盐基离子的吸收, 同时导致NH4+交换作用与NO3-淋溶增强, 进而加速盐基离子损失.与酸性土不同, 氮沉降背景下土壤酸化造成的盐基离子损失在碱性土中的作用相对较弱.这一发现对认识高寒草原生态系统对全球变化的响应等方面具有重要启示.按照目前的预测, 在未来40年中我国的氮沉降仍将维持在较高水平(
Leaching of nitrate from temperate forests—Effects of air pollution and forest management
1
2006
... id="C22">其次, 氮添加导致土壤无机氮含量增加, NH4+与NO3-通过不同途径造成盐基离子损失.Liu等(2013)的研究表明, NH4+-N是氮沉降的主要形式, 但是自20世纪80年代以来, NO3--N的沉降速率也在持续增加, 两者共同导致了日益加剧的氮沉降.由此可以看出, 本研究所选氮肥形态(NH4NO3)能较好地模拟大气氮沉降.随着施氮量增加, 土壤中无机氮含量显著增加(
Significant acidification in major Chinese croplands
1
2010
... id="C23">虽然我们在碱性土中观察到的“土壤交换性盐基离子含量因氮添加显著下降”的现象与酸性土一致, 但导致两类土壤中盐基离子动态变化的驱动因素并不完全相同.这主要是因为两类土壤对土壤酸化的缓冲机制存在差异(
Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest
2
2006
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
... id="C24">本研究揭示了碱性土中盐基离子对连续氮输入的响应及其损失途径.结果发现, 碱性土中氮输入增加了植物对盐基离子的吸收, 同时导致NH4+交换作用与NO3-淋溶增强, 进而加速盐基离子损失.与酸性土不同, 氮沉降背景下土壤酸化造成的盐基离子损失在碱性土中的作用相对较弱.这一发现对认识高寒草原生态系统对全球变化的响应等方面具有重要启示.按照目前的预测, 在未来40年中我国的氮沉降仍将维持在较高水平(
Base cation depletion, eutrophication and acidifycation of species-rich grasslands in response to long-term simulated nitrogen deposition
1
2008
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Assessment of calcium status in Maine forests: Review and future projection
1
2005
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
喀斯特地区植物钙含量特征与高钙适应方式分析
1
2009
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
喀斯特地区植物钙含量特征与高钙适应方式分析
1
2009
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
钙对植物抗逆能力的影响及其生态学意义
1
2005
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
钙对植物抗逆能力的影响及其生态学意义
1
2005
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Calcium in the vacuoles and cell walls of plant tissue: Forms of deposition and their physiological and ecological significance
1
1989
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed
1
2008
... id="C21">本研究结果显示, 氮添加造成土壤交换性盐基离子, 尤其是Mg2+与Na+的减少.造成这种现象的原因主要有以下两点.首先, 氮添加促进植物生长, 加速其对土壤中营养元素的吸收, 进而导致盐基离子含量下降.作为植物生长所需的营养元素, 氮素是生态系统生产力的重要限制因素(
Nitrogen deposition decreases acid buffering capacity of alpine soils in the southern Rocky Mountains
1
2011
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Enhanced nitrogen deposition over China
2013
Nitrogen deposition contributes to soil acidification in tropical ecosystems
2
2014
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
... id="C11">植物样品于65 ℃烘干至恒质量用于估算地上生物量.土壤pH值依据电位法选择1:2.5的土水比测定(
Divergent responses of soil buffering capacity to long-term N deposition in three typical tropical forests with different land-use history
1
2015
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams
3
2011
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
... id="C21">本研究结果显示, 氮添加造成土壤交换性盐基离子, 尤其是Mg2+与Na+的减少.造成这种现象的原因主要有以下两点.首先, 氮添加促进植物生长, 加速其对土壤中营养元素的吸收, 进而导致盐基离子含量下降.作为植物生长所需的营养元素, 氮素是生态系统生产力的重要限制因素(
1
1995
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Soil chemical properties affecting NH4 + sorption in forest soils
1
1996
... id="C22">其次, 氮添加导致土壤无机氮含量增加, NH4+与NO3-通过不同途径造成盐基离子损失.Liu等(2013)的研究表明, NH4+-N是氮沉降的主要形式, 但是自20世纪80年代以来, NO3--N的沉降速率也在持续增加, 两者共同导致了日益加剧的氮沉降.由此可以看出, 本研究所选氮肥形态(NH4NO3)能较好地模拟大气氮沉降.随着施氮量增加, 土壤中无机氮含量显著增加(
1
2006
... id="C6">青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元.由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(
1
2006
... id="C6">青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元.由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(
Nonlinear response of soil respiration to increasing nitrogen additions in a Tibetan alpine steppe
4
2017
... id="C7">试验地点位于青海省刚察县三角城种羊场.地理位置为37.30° N, 100.25° E, 海拔3β290 m.该地区寒冷干燥, 1980-2012年年平均气温为0.08 ℃, 年降水量为387 mm.植被类型为青藏高原典型的高寒草原, 优势种为紫花针茅(Stipa purpurea), 细叶薹草(Carex duriuscula subsp. stenophylloides), 其他伴生物种包括冷地早熟禾(Poa crymophila), 冰草(Agropyron cristatum), 以及阿尔泰狗娃花(Heteropappus altaicus)等.土壤类型为栗钙土, 其中黏粒含量4.9%, 粉粒含量33.4%, 砂粒含量61.7% (
... id="C9">0、1、2、4、8、16、24、32 g·m-2·a-1, 分别用N0、N1、N2、N4、N8、N16、N24、N32表示.试验采用随机区组设计, 共设置5个重复.每个小区大小为6 m × 6 m, 小区之间间隔1 m, 区组之间间隔2 m.施肥时间为每年5、6、7、8及9月月初.施肥主要使用肩背式喷雾器进行: 将每个样地所需的NH4NO3溶于10 L水中, 不断搅拌至全部溶解.每个小区来回均匀喷施两次, 对照小区(N0)喷洒等量清水(
... id="C11">植物样品于65 ℃烘干至恒质量用于估算地上生物量.土壤pH值依据电位法选择1:2.5的土水比测定(
... id="C22">其次, 氮添加导致土壤无机氮含量增加, NH4+与NO3-通过不同途径造成盐基离子损失.Liu等(2013)的研究表明, NH4+-N是氮沉降的主要形式, 但是自20世纪80年代以来, NO3--N的沉降速率也在持续增加, 两者共同导致了日益加剧的氮沉降.由此可以看出, 本研究所选氮肥形态(NH4NO3)能较好地模拟大气氮沉降.随着施氮量增加, 土壤中无机氮含量显著增加(
Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe
1
2017
... id="C7">试验地点位于青海省刚察县三角城种羊场.地理位置为37.30° N, 100.25° E, 海拔3β290 m.该地区寒冷干燥, 1980-2012年年平均气温为0.08 ℃, 年降水量为387 mm.植被类型为青藏高原典型的高寒草原, 优势种为紫花针茅(Stipa purpurea), 细叶薹草(Carex duriuscula subsp. stenophylloides), 其他伴生物种包括冷地早熟禾(Poa crymophila), 冰草(Agropyron cristatum), 以及阿尔泰狗娃花(Heteropappus altaicus)等.土壤类型为栗钙土, 其中黏粒含量4.9%, 粉粒含量33.4%, 砂粒含量61.7% (
Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe
1
2013
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
Physiological functions of beneficial elements
2
2009
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
... id="C21">本研究结果显示, 氮添加造成土壤交换性盐基离子, 尤其是Mg2+与Na+的减少.造成这种现象的原因主要有以下两点.首先, 氮添加促进植物生长, 加速其对土壤中营养元素的吸收, 进而导致盐基离子含量下降.作为植物生长所需的营养元素, 氮素是生态系统生产力的重要限制因素(
Calcium and signal transduction in plants
1
1993
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Forest fertilization research in Finland: A literature review
1
2001
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
2001
2001
A global analysis of soil acidification caused by nitrogen addition
2
2015
... id="C5">工业革命以来, 人类活动如化肥施用、工业生产等使进入生态系统的活性氮显著增加(
... id="C24">本研究揭示了碱性土中盐基离子对连续氮输入的响应及其损失途径.结果发现, 碱性土中氮输入增加了植物对盐基离子的吸收, 同时导致NH4+交换作用与NO3-淋溶增强, 进而加速盐基离子损失.与酸性土不同, 氮沉降背景下土壤酸化造成的盐基离子损失在碱性土中的作用相对较弱.这一发现对认识高寒草原生态系统对全球变化的响应等方面具有重要启示.按照目前的预测, 在未来40年中我国的氮沉降仍将维持在较高水平(
植物镁素营养的研究进展
1
1999
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
植物镁素营养的研究进展
1
1999
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
植物钾营养高效分子遗传机制
1
2009
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
植物钾营养高效分子遗传机制
1
2009
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Base cation and nitrogen budgets for a mixed hardwood catchment in South-Central Ontario
1
2003
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
Global response patterns of terrestrial plant species to nitrogen addition
1
2008
... id="C21">本研究结果显示, 氮添加造成土壤交换性盐基离子, 尤其是Mg2+与Na+的减少.造成这种现象的原因主要有以下两点.首先, 氮添加促进植物生长, 加速其对土壤中营养元素的吸收, 进而导致盐基离子含量下降.作为植物生长所需的营养元素, 氮素是生态系统生产力的重要限制因素(
a). Widespread decreases in topsoil inorganic carbon stocks across China’s grasslands during 1980s-2000s
1
2012
... id="C24">本研究揭示了碱性土中盐基离子对连续氮输入的响应及其损失途径.结果发现, 碱性土中氮输入增加了植物对盐基离子的吸收, 同时导致NH4+交换作用与NO3-淋溶增强, 进而加速盐基离子损失.与酸性土不同, 氮沉降背景下土壤酸化造成的盐基离子损失在碱性土中的作用相对较弱.这一发现对认识高寒草原生态系统对全球变化的响应等方面具有重要启示.按照目前的预测, 在未来40年中我国的氮沉降仍将维持在较高水平(
b). Significant soil acidification across northern China’s grasslands during 1980s-2000s
3
2012
... id="C4">土壤交换性盐基离子(Ca2+、Mg2+、K+、Na+)在维持土壤养分与缓冲土壤酸化中起着重要作用(
... id="C23">虽然我们在碱性土中观察到的“土壤交换性盐基离子含量因氮添加显著下降”的现象与酸性土一致, 但导致两类土壤中盐基离子动态变化的驱动因素并不完全相同.这主要是因为两类土壤对土壤酸化的缓冲机制存在差异(
... id="C24">本研究揭示了碱性土中盐基离子对连续氮输入的响应及其损失途径.结果发现, 碱性土中氮输入增加了植物对盐基离子的吸收, 同时导致NH4+交换作用与NO3-淋溶增强, 进而加速盐基离子损失.与酸性土不同, 氮沉降背景下土壤酸化造成的盐基离子损失在碱性土中的作用相对较弱.这一发现对认识高寒草原生态系统对全球变化的响应等方面具有重要启示.按照目前的预测, 在未来40年中我国的氮沉降仍将维持在较高水平(
青藏高原高寒草地3米深度土壤无机碳库及分布特征
1
2016
... id="C6">青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元.由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(
青藏高原高寒草地3米深度土壤无机碳库及分布特征
1
2016
... id="C6">青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元.由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(
Long-term tobacco plantation induces soil acidification and soil base cation loss
2016
Imbalanced atmospheric nitrogen and phosphorus depositions in China: Implications for nutrient limitation
1
2016
... id="C6">青藏高原平均海拔4β000 m以上, 是地球上十分独特的地理单元.由于高原寒旱的气候特征, 研究区拥有独特的高寒草地植被, 主要包括高寒草原与高寒草甸两类(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}