 ,1,2,*1
,1,2,*1 2
3
Effects of short-term experimental warming on soil microbes in a typical alpine steppe
WANG Jun1,2, WANG Guan-Qin1,2, LI Fei1,2, PENG Yun-Feng1, YANG Gui-Biao1,2, YU Jian-Chun1,2, ZHOU Guo-Ying3, YANG Yuan-He,1,2,*1 2
3
通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1207KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王军, 王冠钦, 李飞, 彭云峰, 杨贵彪, 郁建春, 周国英, 杨元合. 短期增温对紫花针茅草原土壤微生物群落的影响. 植物生态学报[J], 2018, 42(1): 116-125 doi:10.17521/cjpe.2017.0297
WANG Jun.
当前全球正经历着快速的增温过程。自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (IPCC, 2013)。显著的气候变暖会改变陆地生态系统碳循环(Smith & Fang, 2010; Wolf et al., 2016)。气候变暖会加速土壤呼吸(Hicks et al., 2017), 导致生态系统向大气的碳排放增加, 进而引起生态系统碳循环对气候变暖的正反馈(Luo et al., 2009; Friedlingstein, 2015)。然而, 关于正反馈假说的实验证据存在较大争议, 这些争议很大程度上是因为学术界对生态系统碳循环对气候变暖的反馈机制认识不足, 没有充分考虑土壤微生物的作用。土壤微生物是碳(C)、氮(N)、磷(P)、硫(S)等生物地球化学循环的重要参与者和调控者, 在生态系统碳循环中起着关键作用(沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016), 地球系统模型中加入土壤微生物因素能有效地改进模型对碳循环及其对气候变暖反馈关系的预测能力(Lehmann & Kleber, 2015)。鉴于此, 研究土壤微生物生物量及其群落组成对气候变暖的响应, 对准确认识未来气候变暖背景下陆地碳循环的演变规律及其与气候变暖之间的反馈关系具有重要意义(Schmidt et al., 2011)。
国内外****已广泛开展了增温对土壤微生物影响的研究。以往研究显示, 气候变暖可能会影响土壤微生物群落结构。例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(Zhou et al., 2011; Xue et al., 2016b), 也可能单独提高土壤细菌或真菌群落的丰度, 而不影响其他群落(杨林等, 2016; Cheng et al., 2017)。此外, 有些研究发现增温可能并不改变土壤微生物的群落结构(Weedon et al., 2012; Hopkins et al., 2014)。这些研究很大程度上促进了学术界对土壤微生物群落对气候变暖响应的认识, 然而, 以往的研究主要集中在温带地区, 来自寒冷地区的证据较为缺乏。最新的一项整合分析显示, 在全球11类生态系统的25个原位增温实验中, 仅有两个实验来自低温地区(Romero-Olivares et al., 2017)。实际上, 低温条件下土壤微生物群落对气候变暖十分敏感(Margesin et al., 2009; Xue et al., 2016a)。例如, 最近的一项研究指出, 在北极冻土区, 增温1.5年显著改变了土壤中与碳氮循环相关的微生物功能类群(Xue et al., 2016a)。但是, 温度上升会导致土壤含水量下降, 尤其是在低温并且干旱的环境中, 这可能会限制土壤微生物对气候变暖的响应。按照现有的报道, 由于土壤干旱对微生物活性的抑制, 土壤微生物的生物量在气候变暖背景下可能并不会提高(李世清等, 2004), 甚至可能会出现下降(Sheik et al., 2011)。同样, 在土壤干旱的条件下, 土壤微生物的群落结构也可能并不会响应气候变暖, 而是保持相对稳定的状态(Zhang et al., 2015a)。因此, 关于低温干旱地区土壤微生物如何响应气候变暖这一问题值得进一步研究。
青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”。自20世纪以来, 青藏高原地区的气温显著上升(Liu & Chen, 2000; Zhang et al., 2015b)。高海拔导致青藏高原具有低温的气候特征, 进而使得这一地区的生态过程对气候变暖十分敏感(Chen et al., 2013)。高寒草原是青藏高原面积最大的草地生态系统, 面积达6.4 × 105 km2 (Ding et al., 2016), 而紫花针茅(Stipa purpurea)是青藏高原地区的特有植物(Yang et al., 2015), 作为建群种构成了青藏高原高寒草地中分布广、最具代表性的紫花针茅草原(Miller, 2005; 段敏杰等, 2010)。因此, 紫花针茅草原成为研究高寒干旱生态系统中土壤微生物群落对增温响应的理想平台。本研究依托在青海省刚察县三角城种羊场建立的紫花针茅草原增温控制实验平台, 试图揭示紫花针茅草原土壤微生物生物量及群落结构对增温的响应及其调控因素。
1 材料和方法
1.1 样地概况和实验设计
本研究在青海省刚察县三角城种羊场开展, 详细的样地情况见Peng等(2017)的描述。简单来讲, 研究区域过去30年的年平均气温为0.08 ℃, 年降水量为387 mm, 属于高原大陆性气候, 植被类型为青藏高原高寒草原中最具代表性的紫花针茅草原。我们于2013年建立了增温实验平台, 每个4 m × 4 m的样方内设置增温与对照两种配对处理, 共设10个重复。采用开顶箱(OTC)装置实现增温, 于2013年7月开始全年连续增温处理。同时, 在每个实验处理中安装土壤温湿度探头, 采用EM50数据采集器(Decagon Devices, Pullman, USA)以0.5 h为间隔持续记录5 cm深度的土壤温湿度。1.2 研究方法
1.2.1 土壤样品采集与测定我们于2015和2016年的生长季旺盛期(8月)采集了土壤样品。在每个样方的对照、增温处理中随机选3个点, 去除表层凋落物, 采用直径3 cm的土钻取表层0-10 cm的土壤, 将3钻样品混匀, 挑根, 并过2 mm筛后分为两份。一份储存于-20 ℃冰箱内用于微生物生物量碳、氮以及磷脂脂肪酸(PLFA)等指标的测定; 另一份自然风干, 用于分析土壤理化性质。此外, 在每年8月的生长季旺期每隔7-10天使用多光谱数码相机(Tetracam, Chatsworth, Los Angeles, USA)测定归一化植被指数(NDVI)。
为了揭示短期增温对土壤性质的影响, 我们测定了土壤碳(C)、氮(N)元素含量。其中, 土壤总碳含量(TC)和全氮含量(TN)采用元素分析仪(Vario EL III Elementar, Elementar Analysensysteme GmbH, Langenselbold, Germany)测定。土壤无机碳含量(SIC)采用碳酸盐分析仪(Eijelkamp 08.53, Eijkelkamp, Giesbeek, Netherlands)测定, 土壤有机碳含量(SOC)则通过计算TC减去SIC的差值获得。SOC与TN的比值作为土壤碳氮比(C:N)。
1.2.2 土壤微生物生物量碳、氮含量的测定
为了揭示短期增温对土壤微生物生物量的影响, 我们基于氯仿熏蒸法(Vance et al., 1987)测定了微生物生物量碳(MBC)、氮(MBN)。首先, 称取20 g的新鲜土样在25 ℃条件下预培养7天, 然后放入真空干燥器, 常温下用氯仿熏蒸24 h, 用0.5 mol·L-1的K2SO4浸提。同时称取同等质量的土壤样品, 不经氯仿熏蒸, 直接用0.5 mol·L-1的K2SO4浸提(吴金水等, 2006)。浸提液采用碳氮分析仪(multi-N/C 3100, Analytik Jena AG, Jena, Germany)测定C、N含量, 通过计算熏蒸和未熏蒸浸提液的C、N浓度差值获得MBC和MBN, 转换系数采用0.45和0.54 (Brookes et al., 1985; Tischer et al., 2014)。
1.2.3 土壤微生物群落结构的测定
为了揭示短期增温对土壤微生物群落结构的影响, 我们参照Bossio等(1998)的方法测定了土壤微生物PLFA。首先利用单相氯仿-甲醇-磷酸缓冲液提取土壤微生物PLFA, 然后将提取的磷脂通过固相萃取柱分离糖脂和中性脂, 并将极性脂甲酯化形成PLFA甲基脂。最后将样品用含有正十九酸甲酯 (19:0)作为内标的正己烷重新溶解, 用气相色谱仪(Agilent 6850, Agilent Technologies, Santa Clara, USA)进行测定。分析过程中, 依据MIDI微生物鉴别系统进行脂肪酸鉴定, 并采用以下指示脂肪酸标准: 丛枝菌根真菌(AMF): 16:1ω5c (Olsson, 1999); 真菌: 18:2ω6, 9c 18:1ω9 (Frosteg?rd et al., 1993); 革兰氏阳性菌(G+): i14:0, i15:0, a15:0, i16:0, a17:0, i17:0; 革兰氏阴性菌(G-): 16:1ω7c, cy-17:0, 18:1ω7, cy19:0 (Frosteg?rd & B??th, 1996), 阳性菌与阴性菌加和得到细菌的丰度; 放线菌: 10Me-16:0, 10Me-17:0, 10Me-18:0 (Kroppenstedt, 1985)。饱和脂肪酸: 14:0, 15:0, 16:0, 17:0, 18:0; 不饱和脂肪酸: 16:1ω7c, 16:1ω5c, 17:1ω8c, a17:1, 18:1ω9c, 18:1ω7c, 18:1ω5c。土壤微生物的生理状态通过饱和与不饱和脂肪酸之比(S/M)表征(Moore-Kucera & Dick, 2008)。土壤微生物总PLFA为各类群微生物PLFA的总和, 各类群PLFA占总PLFA的比例为相应类群的相对丰度。
1.3 数据分析
数据分析按照以下步骤进行。首先, 采用配对t检验来检验环境变量(土壤温度、土壤含水量、SOC、TC、TN、C:N及NDVI)和微生物生物量及群落对增温的响应。其次, 采用双因素方差分析检验增温处理和环境因子的年际波动对不同土壤微生物群落的影响, 并利用主成分分析法(PCA)检验增温对微生物群落结构的影响,冗余分析法(RDA)分析微生物群落结构与环境变量(土壤温度、土壤含水量、SOC、TC、TN、C:N)之间的关系。最后, 利用混合线性回归分析了土壤温度的变化量(ΔST, 增温-对照)、土壤水分变化量(ΔSM, 增温-对照)与微生物生物量碳的变化量(ΔMBC, 增温-对照)之间的关系。上述统计分析利用软件R 3.4.0 (R Development Core Team, 2017)及CANOCO 5.0 (Microcomputer Power, Ithaca, New York)完成。2 研究结果
2.1 短期增温对土壤环境和地上植被的影响
4年增温处理显著改变了土壤微环境(表1)。增温显著提高了土壤表层0-5 cm的温度, 2015和2016年的土壤平均温度分别为14.4和15.4 ℃, 相对于对照均上升了1.6 ℃ (2015年: p < 0.01; 2016年: p < 0.01; 表1)。同时, 增温处理导致土壤含水量显著下降, 2015和2016年的土壤平均含水量分别为9.8%和13.3%, 相对于对照分别下降了3.4%和2.4% (体积分数; 2015年: p < 0.05, 2016年: p < 0.01; 表1)。然而, 增温并没有改变土壤SOC、TC、TN和C:N等化学性质, 也没有改变NDVI (表1)。需要说明的是, NDVI在2015和2016年间略有差异, 但土壤属性在两年实验期间较为稳定(表1)。Table 1
表1
表1短期增温对土壤理化性质及地上植被的影响
Table 1
| 土壤因子 Soil factors | 2015年 | 2016年 | |||
|---|---|---|---|---|---|
| 对照 Control | 增温 Warming | 对照 Control | 增温 Warming | ||
| 土壤温度 Soil temperature (℃) | 12.80 ± 0.30 | 14.40 ± 0.20** | 13.90 ± 0.20 | 15.40 ± 0.30** | |
| 土壤含水量(体积分数) Soil moisture (volume fraction) | 13.20 ± 1.80 | 9.80 ± 1.10** | 15.60 ± 1.80 | 13.30 ± 1.20* | |
| 有机碳含量 Soil organic carbon content (%) | 3.20 ± 0.20 | 3.20 ± 0.20 | 3.30 ± 0.10 | 3.32 ± 0.10 | |
| 总碳含量 Total carbon content (%) | 4.50 ± 0.20 | 4.40 ± 0.20 | 4.20 ± 0.10 | 4.50 ± 0.10 | |
| 全氮含量 Total nitrogen content (%) | 0.38 ± 0.02 | 0.38 ± 0.02 | 0.38 ± 0.01 | 0.38 ± 0.01 | |
| 碳氮比 | 8.50 ± 0.20 | 8.60 ± 0.20 | 8.70 ± 0.20 | 8.80 ± 0.20 | |
| 归一化植被指数 Normalized difference vegetation index | 0.27 ± 0.01 | 0.26 ± 0.01 | 0.18 ± 0.02 | 0.19 ± 0.02 | |
新窗口打开|下载CSV
2.2 短期增温对土壤微生物群落的影响
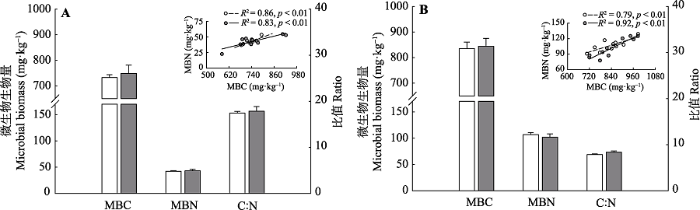
4年增温并没有改变MBC和MBN (图1)。短期增温处理后, 2015、2016年的MBC分别为749.0、 844.3 mg·kg-1 , MBN分别为43.1和102.1 mg·kg-1 , 二者相对于对照没有显著差异(图1A、1B)。类似地, 增温处理中2015、2016年的微生物C:N分别为17.9和8.4, 并且和对照相比没有显著差异(图1A、1B)。此外, MBC和MBN之间呈现显著的线性关系(2015年, 对照: R2 = 0.86, p < 0.01; 增温: R2 = 0.83, p <0.01; 2016年, 对照: R2 = 0.79, p < 0.01; 增温: R2 = 0.92, p < 0.01; 图1A、1B), 且这种关系并没有受到短期增温的影响(2015年: p = 0.08, 2016年: p = 0.13)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12015(A)和2016年(B)生长季(8月)对照与增温处理下的土壤微生物生物量碳(MBC)、氮(MBN)、微生物生物量碳氮比(C:N)及MBC与MBN之间的关系(平均值±标准误差)。灰色柱条和圆圈为增温处理; 白色柱条和圆圈为对照处理。
Fig. 1Microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), and microbial biomass C:N under control and warming treatment during the growing season (August) in 2015 (A) and 2016 (B), as well as the relationships between MBC and MBN (mean ± SD). The gray bars and cycles represent warming treatments. The white bars and cycles represent control treatments.
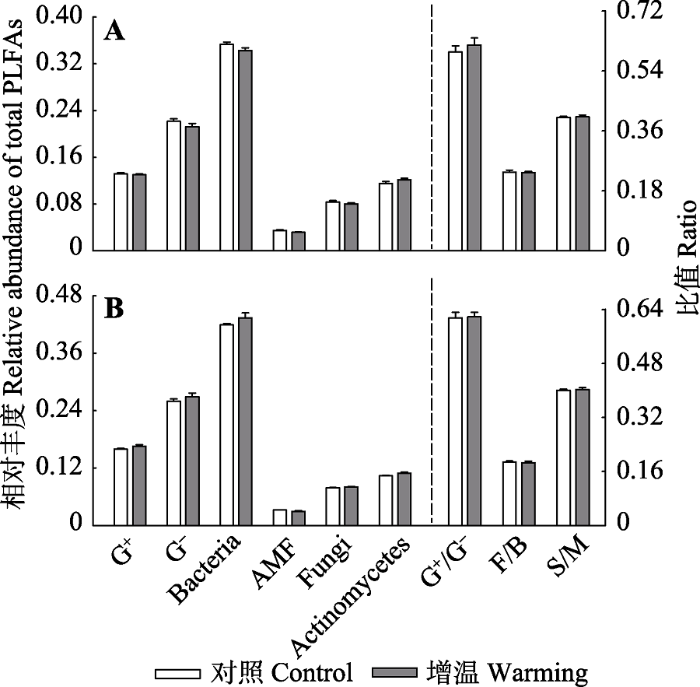
基于PLFA分析的结果显示, 细菌在紫花针茅草原生态系统的土壤微生物群落中占据主导地位, 其他类群的微生物占群落总量的35%-43%; 而AMF的占比为3%, 是相对量最少的微生物类群(图2)。并且, 短期增温并没有改变紫花针茅草原土壤微生物的群落结构。2015、2016年增温处理中, 土壤革兰氏阳性菌(G+)、革兰氏阴性菌(G-)、细菌、AMF、真菌以及放线菌的相对丰度均没有发生变化。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2短期增温对不同土壤微生物类群的影响(平均值±标准误差)。A, 2015年。B, 2016年。Actinomycetes, 放线菌; AMF, 丛枝菌根真菌; Bacteria, 细菌; Fungi, 真菌; G+, 革兰氏阳性菌; G-, 革兰氏阴性菌; F/B, 真菌与细菌比值; G+/G-, 革兰氏阳性菌与阴性菌比值; S/M, 饱和脂肪酸与不饱和脂肪酸比值。
Fig. 2Effects of short-term experimental warming on different soil microbial groups (mean ± SD). A, In 2015. B, In 2016. AMF, arbuscular mycorrhizal fungi; G+, gram-positive bacteria; G-, gram-negative bacteria; F/B, ratio of fungi and bacteria; G+/G-, ratio of gram-positive bacteria and gram-negative bacteria; S/M, ratio of saturated PLFAs and monosaturated PLFAs.
基于双因素方差分析的结果表明, 细菌、G+、G-、AMF、放线菌的相对丰度以及土壤总PLFA、F/B、S/M出现显著的年际差异(p < 0.05), 而真菌和G+/G-在两年间保持不变(表2)。两年综合分析中, 除了AMF和放线菌, 增温对其他各个土壤微生物类群及F/B、G+/G-、S/M没有显著影响。短期增温处理并没有显著改变S/M, 表明短期增温可能并没有对微生物群落的生理状态产生影响。此外, 除了细菌群落, 增温和环境因子年际波动的交互作用对土壤微生物群落也没有显著影响(表2)。
Table 2
表2
表2基于双因素方差分析得到的短期增温(W)、取样时间(T)及其交互作用(W × T)对微生物群落的影响(F值)
Table 2
| 微生物群落 Microbial community | T | W | W × T |
|---|---|---|---|
| 细菌 Bacteria | 159.90** | 0.11 | 4.30* |
| 革兰氏阳性菌 Gram-positive bacteria | 173.66** | 0.93 | 2.56 |
| 革兰氏阴性菌 Gram-negative bacteria | 67.70** | 0.00 | 2.50 |
| 丛枝菌根真菌 Arbuscular mycorrhizal fungi | 7.20* | 8.49* | 0.03 |
| 真菌 Fungi | 2.12 | 0.18 | 2.13 |
| 放线菌 Actinomycetes | 24.28** | 5.37* | 0.02 |
| 真菌细菌比 Fungi/Bacteria | 123.85** | 0.15 | 0.01 |
| G+/G- Gram-positive bacteria/Gram-negative bacteria | 0.27 | 0.45 | 0.28 |
| 饱和与不饱和脂肪酸比 Saturated PLFAs / Monosaturated PLFAs | 7.13* | 2.24 | 0.86 |
| 磷脂脂肪酸总量 Total phospholipid fatty acids | 157.39** | 1.67 | 1.08 |
新窗口打开|下载CSV
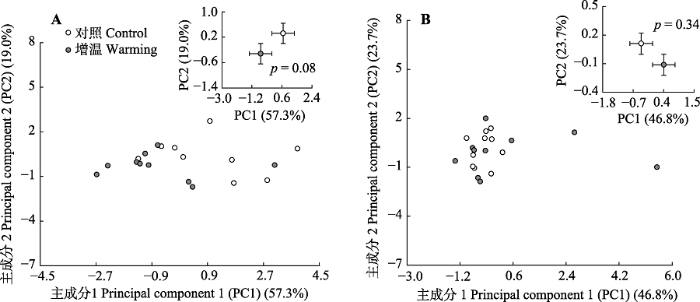
PCA分析的结果进一步证实增温没有改变土壤微生物的群落结构。2015年PC1轴和PC2轴分别能够解释土壤微生物群落结构变异的57.3%和19.0%, 总解释率为76.3% (图3A); 2016年PC1轴和PC2轴分别解释了群落结构变异的46.8%和23.7%, 总解释率为70.5% (图3B)。两年中增温处理与对照的微生物群落相互重叠, 没有显著差异(2015年, p = 0.08; 2016年, p = 0.34; 图3A、3B)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3短期增温对土壤微生物群落结构的影响(平均值±标准误差)。A, 2015年。B, 2016年。
Fig. 3Effects of short-term experimental warming on soil microbial community structure (mean ± SE). A, In 2015. B, In 2016.
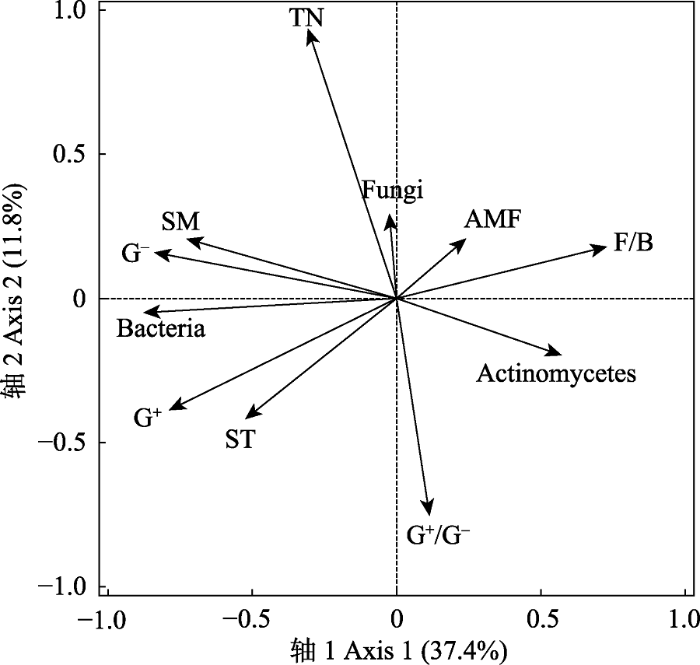
RDA分析结果显示, 排序轴能够解释微生物群落结构变异的49.2% (轴1: 37.4%; 轴2: 11.8%, 图4)。土壤温度、含水量、TN分别解释22.8% (p < 0.01)、20.7% (p < 0.01)、8.7% (p < 0.01)的微生物群落结构变异。土壤细菌生物量与土壤温度和含水量之间显著正相关, 进一步证明土壤温度和含水量对土壤微生物群落的调控作用。此外,TN能够解释8.7%的微生物群落结构变异(p < 0.01)。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4基于冗余分析得到的2015和2016年微生物群落与环境因子之间的关系。Actinomycetes, 放线菌; AMF, 丛枝菌根真菌; Bacteria, 细菌; Fungi, 真菌; G+, 革兰氏阳性菌; G-, 革兰氏阴性菌; F/B, 真菌与细菌比值; G+/G-, 革兰氏阳性菌与阴性菌比值; SM, 土壤含水量; ST, 土壤温度; TN, 全氮含量。
Fig. 4Redundancy analysis of soil microbial community to soil environmental parameters. AMF, arbuscular mycorrhizal fungi; G+, gram-positive bacteria; G-, gram-negative bacteria; F/B, ratio of fungi and bacteria; G+/G-, ratio of gram-positive bacteria and gram-negative bacteria; SM, soil moisture; ST, soil temperature; TN, total nitrogen content.
3 讨论
本文研究了紫花针茅草原土壤微生物群落对短期增温的响应, 发现尽管短期增温导致土壤温度显著增加, 土壤微生物的生物量和群落结构并没有发生改变。这种现象可能由以下3个原因造成。首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定。一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(Griffiths & Philippot, 2013; 贺纪正等, 2013)。本研究的结果表明, 尽管短期增温显著地影响了AMF和放线菌的生物量(表2), 然而这两个类群的微生物在土壤总微生物群落中所占比例很小, 不足以影响微生物的群落结构(图3)。因此, 持续4年的增温并没有整体上改变微生物的生物量及群落结构。另一方面, 除了土壤微生物本身的特性, 土壤性质和地上植被对微生物也有重要的影响, 三者间存在密切的交互作用(Pailler et al., 2014; Birgander et al., 2017)。最近的研究显示, 土壤性质和地上植物是调控青藏高原高寒草地土壤微生物群落的主要因素, 具体表现为SOC、土壤C:N、NDVI等环境因子与土壤微生物群落的显著相关性(Chen et al., 2016)。本研究中, 增温处理并没有改变土壤SOC、TC、TN以及土壤C:N, 同时NDVI也没有发生显著变化。因此, 土壤性质和地上植被在短期增温背景下的相对稳定性可能导致土壤微生物群落没有受到短期土壤增温的显著影响。
其次, 土壤温度和水分等环境因素会对土壤微生物群落产生不同影响, 进而抑制土壤微生物对气候变暖的响应。大量研究表明, 土壤温度与土壤含水量对微生物的生理状态、生物量及群落结构有重要影响(Brockett et al., 2012; Zeglin et al., 2013)。气候变暖能够提高微生物代谢过程的酶活性(Nazaries et al., 2015), 促进微生物的生长。本研究中, 我们发现土壤温度和水分对微生物群落结构有显著影响(图4), 同时土壤温度的变化量与微生物生物量碳的变化量之间存在显著的正相关关系(图5A), 在土壤温度显著上升的背景下(图5A), 这一结果意味着气候变暖也可能会对紫花针茅草原土壤微生物产生促进作用。然而, 如果生态系统受到水分的限制, 微生物的变化将有所不同。例如, Sheik等(2011)在温带草原的研究发现, 正常降水年份中, 增温使土壤微生物生物量提高40%-150%; 而在干旱年份, 增温处理的土壤微生物生物量会下降50%-80%, 表明干旱会抵消增温对微生物的促进效应。本研究中, 我们发现土壤含水量的变化量与微生物生物量碳的变化量同样存在显著的正相关关系(图5B), 说明高寒草地生态系统存在水分限制, 而由于增温处理导致土壤含水量显著下降(图5B), 土壤的干旱将抵消增温对土壤微生物的促进作用, 进而使得微生物群落没有显著改变。
图5
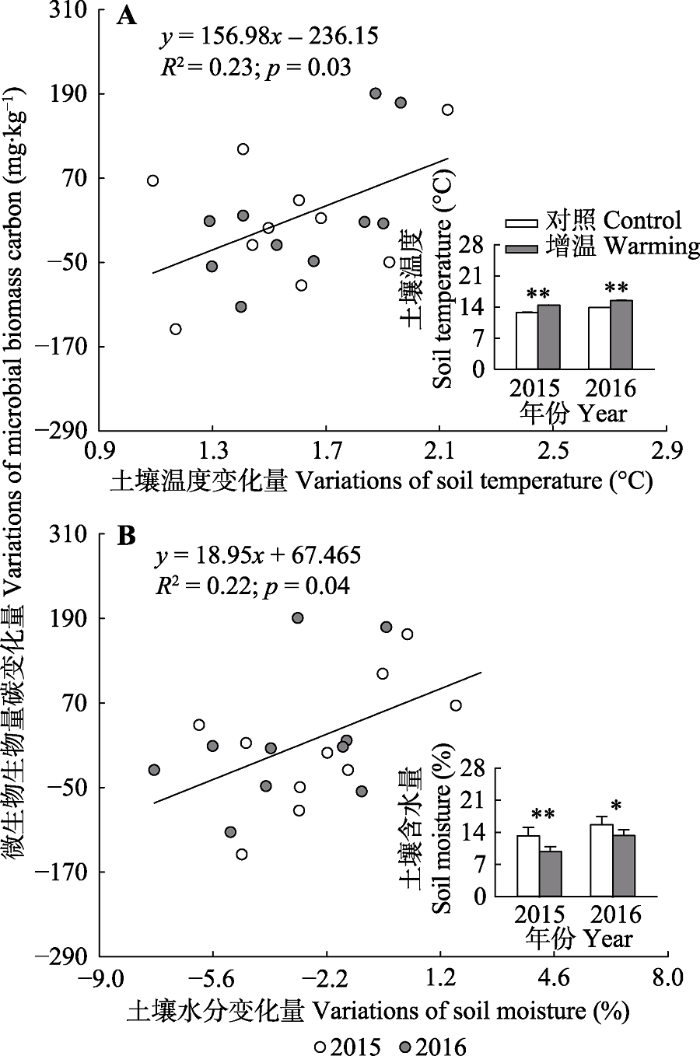 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5土壤微生物生物量碳变化量(增温-对照)与土壤温度的变化量(增温-对照, A)、土壤水分的变化量(增温-对照, B)之间的关系及短期增温对土壤温度和土壤含水量的影响。
Fig. 5Relationships among warming-induced changes (warming-control) in microbial biomass carbon, soil temperature (A) and soil moisture (B), and the effects of short-term warming on soil temperature and moisture.
此外, 增温处理中土壤微生物群落特征保持相对稳定可能与增温时间较短以及增温幅度较低有关。以往的增温实验表明, 土壤微生物对增温的响应取决于增温年限。例如, Weedon等(2012)发现9年的增温实验不足以改变微生物群落, 然而15年的增温导致微生物群落发生了显著变化(Rinnan et al., 2007), 说明土壤微生物群落对增温的响应可能存在时间滞后, 也就是说微生物群落对短时间增温不会做出及时的响应。在我们的研究中, 4年的增温对于微生物群落来说可能时间较短, 不足以产生变化。除了增温的年限, 增温的幅度可能也会限制土壤微生物对增温的响应。王文立等(2015)通过整合分析证实, 土壤微生物的响应比与增温幅度之间显著正相关。即增温幅度越高, 微生物群落的变化越大。本实验采用开顶箱被动增温, 该装置对表层土壤的增温能力有限(< 2 ℃), 因此可能尚未达到显著影响土壤微生物群落的温度阈值。
值得注意的是, 本研究采用PLFA作为生物标记物指示微生物群落, PLFA法能够准确地反映不同类群微生物的生物量, 并且能反映微生物表现型和活性的信息(Frosteg?rd et al., 2011), 因此该方法能够确定相对宏观的微生物群落结构特征。然而, 该标记物的检测精度有限, 很难得到微生物种属一级的精确信息(Frosteg?rd et al., 2011), 无法检测更精细的微生物群落组成。鉴于此, 未来研究应考虑采用基于分子分析的定量PCR、高通量测序以及宏基因组等手段, 以揭示土壤微生物群落的精细结构及其对增温的响应。
4 结论
本研究利用青藏高原高寒草原生态系统的增温平台, 通过熏蒸法和PLFA法测定了土壤微生物生物量及群落结构, 在此基础上揭示了紫花针茅草原土壤微生物群落对短期增温的响应。结果表明, 土壤干旱抵消了温度上升对土壤微生物群落的促进作用, 导致其生物量与群落结构在4年增温中均保持不变。这个结果表明, 未来预测增温对土壤微生物群落的影响时, 应当考虑干旱对土壤微生物群落响应增温的限制作用。尽管如此, 由于土壤微生物群落对外界干扰具有一定程度的抵抗力, 4年增温可能并不会产生显著影响, 同时PLFA法的检测精度有限, 难以区分更精细的微生物群落组成。因此, 未来的研究应当延长增温处理期限, 并采用诸如高通量测序等更高精度的手段揭示土壤微生物群落结构及其对气候变暖的响应。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/gcb.13803URLPMID:28675677 [本文引用: 1]

Abstract A decisive set of steps in the terrestrial carbon (C) cycle is the fixation of atmospheric C by plants and the subsequent C-transfer to rhizosphere microorganisms. With climate change winters are expected to become milder in temperate ecosystems. Although the rate and pathways of rhizosphere C input to soil could be impacted by milder winters, the responses remain unknown. To address this knowledge-gap, a winter-warming experiment was established in a semi-natural temperate grassland in order to follow the C flow from atmosphere, via the plants, to different groups of soil microorganisms. In situ 13 CO 2 pulse labelling was used to track C into signature fatty acids of microorganisms. The winter warming did not result in any changes in biomass of any of the groups of microorganisms. However, the C flow from plants to arbuscular mycorrhizal (AM) fungi, increased substantially by winter warming. Saprotrophic fungi also received large amounts of plant-derived C - indicating a higher importance for the turnover of rhizosphere C than biomass estimates would suggest - still, C flow was unaffected by winter warming. AM fungi was the only microbial group positively affected by winter warming - the group with the closest connection to plants. Winter warming resulted in higher plant productivity earlier in the season, and this aboveground change likely induced plant nutrient limitation in warmed plots, thus stimulating the plant dependence on, and C allocation to, belowground nutrient acquisition. The preferential C allocation to AM fungi was at the expense of C flow to other microbial groups, which were unaffected by warming. Our findings imply that warmer winters may shift rhizosphere C-fluxes to become more AM fungal-dominated. Surprisingly, the stimulated rhizosphere C flow was matched by increased microbial turnover, leading to no accumulation of soil microbial biomass. This article is protected by copyright. All rights reserved. This article is protected by copyright. All rights reserved.
DOI:10.1007/s002489900087URLPMID:9622559
Phospholipid fatty acid (PLFA) profiles were measured in soils from organic, low-input, and conventional farming systems that are part of the long term Sustainable Agriculture Farming Systems (SAFS) Project. The farming systems differ in whether their source of fertilizer is mineral or organic, and in whether a winter cover crop is grown. Sustained increases in microbial biomass resulting from high organic matter inputs have been observed in the organic and low-input systems. PLFA profiles were compared to ascertain whether previously observed changes in biomass were accompanied by a change in the composition of the microbial community. In addition, the relative importance of environmental variables on PLFA profiles was determined. Redundancy analysis ordination showed that PLFA profiles from organic and conventional systems were significantly different from April to July. On ordination plots, PLFA profiles from the low-input system fell between organic and conventional systems on most sample dates. A group of fatty acids (i14:0, a15:0, 16:1 7c, 16:1 5c, 14:0, and 18:2 6c) was enriched in the organic plots throughout the sampling period, and another group (10Me16:0, 2OH 16:1 and 10Me17:0) was consistently lower in relative abundance in the organic system. In addition, another group (15:0, a17:0, i16:0, 17:0, and 10Me18:0) was enriched over the short term in the organic plots after compost incorporation. The relative importance of various environmental variables in governing the composition of microbial communities could be ranked in the order: soil type > time > specific farming operation (e.g., cover crop incorporation or sidedressing with mineral fertilizer) > management system > spatial variation in the field. Measures of the microbial community and soil properties (including microbial biomass carbon and nitrogen, substrate induced respiration, basal respiration, potentially mineralizable nitrogen, soil nitrate and ammonium, and soil moisture) were seldom associated with the variation in the PLFA profiles.
DOI:10.1016/j.soilbio.2011.09.003URL [本文引用: 1]
Although soil microorganisms play a central role in the soil processes that determine nutrient availability and productivity of forest ecosystems, we are only beginning to understand how microbial communities are shaped by environmental factors and how the structure and function of soil microbial communities in turn influence rates of key soil processes. Here we compare the structure and function of soil microbial communities in seven mature, undisturbed forest types across a range of regional climates in British Columbia and Alberta, and examine the variation in community composition within forest types. We collected the forest floor fermentation (F) and humus (H) layers and upper 10 cm of mineral soil at 3 sites in each of seven forest types (corresponding to seven Biogeoclimatic zones) in both spring and summer. Phospholipid fatty acid analysis was used to investigate the structure of soil microbial communities and total soil microbial biomass; potential activities of extra-cellular enzymes indicated the functional potential of the soil microbial community in each layer at each site. Multivariate analysis indicated that both structure and enzyme activities of soil microbial communities differed among the forest types, and significantly separated along the regional climate gradient, despite high local variation. Soil moisture and organic matter contents were most closely related to microbial community characteristics. Forests in the Ponderosa Pine and Mountain Hemlock zones were distinct from other forests and from each other when comparing potential enzyme activities and had the most extreme moisture and temperature values. Forest floors from the hot and dry Ponderosa Pine forests were associated with enzymes characteristic of water-stress and high concentrations of phenols and other recalcitrant compounds. The wet and cold Mountain Hemlock forests were associated with low enzyme activity. An influence of tree species was apparent at the three sites within the Coastal Western Hemlock zone; high bacterial:fungal biomass ratios were found under western redcedar (Thuja plicata) which also had high pH and base-cation levels, and under Douglas-fir (Pseudotsuga menziesii), which had high N availability. Potential activities enzymes differed among soil layers: potential activities of phenol oxidase and peroxidase were highest in mineral soil, whereas phosphatase, betaglucosidase, NAGase, sulfatase, xylosidase and cellobiohydrolase were highest in the forest floors.
DOI:10.1016/0038-0717(85)90144-0URL [本文引用: 1]
A new “direct extraction” method for measuring soil microbial biomass nitrogen (biomass N) is described. The new method (fumigation-extraction) is based on CHC1 3 fumigation, followed by immediate extraction with 0.5 M K 2SO 4 and measurement of total N released by CHC1 3 in the soil extracts. The amounts of NH 4-N and total N extracted by K 2SO 4 immediately after fumigation increased with fumigation time up to 5 days. Total N released by CHC1 3 after 1 day fumigation (1 day CHC1 3-N) and after 5 days fumigation (5 day CHC1 3-N) were positively correlated with the flush of mineral N (F N) in 37 soils that had been fumigated, the fumigant removed and the soils incubated for 10 days (fumigation-incubation). The regression equations were 1 day CHC1 3-N = (0.79 ± 0.022) F N and 5 day CHC1 3-N = (1.01 ± 0.027) F N, both regressions accounting for 92% of the variance in the data. In field soils previously treated with 15N-labelled fertilizer, the amounts of labelled N, measured after fumigation-extraction, were very similar to the amounts of labelled N mineralized during fumigation-incubation; both were about 4 times as heavily labelled as the soil N as a whole. These results suggest that fumigation-extraction and fumigation-incubation both measure the same fraction of the soil organic N (probably the cytoplasmic component of the soil microbial biomass) and that measurement of the total N released by CHC1 3 fumigation for 24 h provides a rapid method for measuring biomass N.
DOI:10.1111/gcb.12277URLPMID:23744573 [本文引用: 1]
With a pace of about twice the observed rate of global warming, the temperature on the Qinghai-Tibetan Plateau (Earth's hird pole ) has increased by 0.2 C per decade over the past 50 years, which results in significant permafrost thawing and glacier retreat. Our review suggested that warming enhanced net primary production and soil respiration, decreased methane (CH4) emissions from wetlands and increased CH4 consumption of meadows, but might increase CH4 emissions from lakes. Warming-induced permafrost thawing and glaciers melting would also result in substantial emission of old carbon dioxide (CO2) and CH4. Nitrous oxide (N2O) emission was not stimulated by warming itself, but might be slightly enhanced by wetting. However, there are many uncertainties in such biogeochemical cycles under climate change. Human activities (e.g. grazing, land cover changes) further modified the biogeochemical cycles and amplified such uncertainties on the plateau. If the projected warming and wetting continues, the future biogeochemical cycles will be more complicated. So facing research in this field is an ongoing challenge of integrating field observations with process-based ecosystem models to predict the impacts of future climate change and human activities at various temporal and spatial scales. To reduce the uncertainties and to improve the precision of the predictions of the impacts of climate change and human activities on biogeochemical cycles, efforts should focus on conducting more field observation studies, integrating data within improved models, and developing new knowledge about coupling among carbon, nitrogen, and phosphorus biogeochemical cycles as well as about the role of microbes in these cycles.
DOI:10.1111/jbi.12806URL [本文引用: 1]
Abstract AimSoil microorganisms play key roles in regulating many important ecosystem processes. However, our understanding of the patterns and drivers of soil microbial communities remains limited. This study aims to test the hypothesis that edaphic factors are more important in explaining variations in soil microbial communities than climatic and biotic factors, as soils directly provide substrates and environment for soil microbial communities. LocationTibetan alpine grasslands and global terrestrial biomes. MethodsOn the basis of phospholipid fatty acid (PLFA) analysis, we investigated large-scale patterns and drivers of soil microbial communities using data obtained from 196 sites within two major grassland types (alpine steppe and alpine meadow) on the Tibetan Plateau. We also explored global patterns of soil microbial communities by analysing published data from around the world. ResultsSoil microbial PLFAs in Tibetan alpine grasslands were positively associated with mean annual temperature and mean annual precipitation (MAP), contradicting patterns previously observed across global biomes. A combined stepwise regression and variation partitioning analysis revealed that soil microbial community variations in Tibetan alpine grasslands were mainly explained by edaphic factors, such as soil organic carbon, C : N ratio, pH and soil texture, then by biotic factors, such as aboveground biomass and plant species richness, and further by climatic factors, such as MAP. The global analysis confirmed that edaphic factors accounted for a greater portion of the variation in soil microbial communities than did climatic and biotic variables. Main conclusionsThese results provide strong support for the hypothesis that edaphic factors are the dominant drivers of spatial variations in soil microbial communities across regional and global scales.
DOI:10.1038/ismej.2017.48URLPMID:28430189 [本文引用: 1]
Abstract Soil organic matter (SOM) stocks contain nearly three times as much carbon (C) as the atmosphere and changes in soil C stocks may have a major impact on future atmospheric carbon dioxide concentrations and climate. Over the past two decades, much research has been devoted to examining the influence of warming on SOM decomposition in topsoil. Most SOM, however, is old and stored in subsoil. The fate of subsoil SOM under future warming remains highly uncertain. Here, by combining a long-term field warming experiment and a meta-analysis study, we showed that warming significantly increased SOM decomposition in subsoil. We also showed that a decade of warming promoted decomposition of subsoil SOM with turnover times of decades to millennia in a tall grass prairie and this effect was largely associated with shifts in the functional gene structure of microbial communities. By coupling stable isotope probing with metagenomics, we found that microbial communities in warmed soils possessed a higher relative abundance of key functional genes involved in the degradation of organic materials with varying recalcitrance than those in control soils. These findings suggest warming may considerably alter the stability of the vast pool of old SOM in subsoil, contributing to the long-term positive feedback between the C cycle and climate.
DOI:10.1038/ncomms10541URLPMID:4738359 [本文引用: 1]
Despite the importance of microbial communities for ecosystem services and human welfare, the relationship between microbial diversity and multiple ecosystem functions and services (that is, multifunctionality) at the global scale has yet to be evaluated. Here we use two independent, large-scale databases with contrasting geographic coverage (from 78 global drylands and from 179 locations across Scotland, respectively), and report that soil microbial diversity positively relates to multifunctionality in terrestrial ecosystems. The direct positive effects of microbial diversity were maintained even when accounting simultaneously for multiple multifunctionality drivers (climate, soil abiotic factors and spatial predictors). Our findings provide empirical evidence that any loss in microbial diversity will likely reduce multifunctionality, negatively impacting the provision of services such as climate regulation, soil fertility and food and fibre production by terrestrial ecosystems. The role of microbial diversity in ecosystems is less well understood than, for example, that of plant diversity. Analysing two independent data sets at a global and regional scale, Delgado-Baquerizoet al. show positive effects of soil diversity on multiple terrestrial ecosystem functions.
DOI:10.1111/gcb.13257URLPMID:26913840 [本文引用: 1]
Abstract The permafrost organic carbon (OC) stock is of global significance because of its large pool size, and the potential positive feedback to climate warming. However, due to the lack of systematic field observations and appropriate upscaling methodologies, substantial uncertainties exist in the permafrost OC budget, which limits our understanding of the fate of frozen carbon in a warming world. In particular, the lack of comprehensive estimates of OC stocks across alpine permafrost means that current knowledge on this issue remains incomplete. Here we evaluated the pool size and spatial variations of permafrost OC stock to 3 metres depth on the Tibetan Plateau by combining systematic measurements from a substantial number of pedons (i.e., 342 three-metre-deep cores and 177 50-cm-deep pits) with a machine learning technique (i.e., support vector machine, SVM). We also quantified uncertainties in permafrost carbon budget by conducting a Monte Carlo simulation. Our results revealed that the combination of systematic measurements with the SVM model allowed spatially explicit estimates to be made. The OC density (OC amount per unit area, OCD) exhibited a decreasing trend from the southeastern to the northwestern plateau, with the exception that OCD in the swamp meadow was substantially higher than that in surrounding regions. Our results also demonstrated that Tibetan permafrost stored a large amount of OC in the top 3 metres, with the median OC pool size being 15.31 Pg C (interquartile range: 13.03-17.77 Pg C). 44% of OC occurred in deep layers (i.e., 100-300 cm), close to the proportion observed across the northern circumpolar permafrost region. The large carbon pool size, together with significant permafrost thawing suggests a risk of carbon emissions and positive climate feedback across the Tibetan alpine permafrost region. This article is protected by copyright. All rights reserved. This article is protected by copyright. All rights reserved.
[本文引用: 1]
[本文引用: 1]
DOI:10.1098/rsta.2014.0421URLPMID:26438284 [本文引用: 1]
Climate and carbon cycle are tightly coupled on many timescales, from interannual to multi-millennial timescales. Observations always evidence a positive feedback, warming leading to release of carbon to the atmosphere; however, the processes at play differ depending on the timescales. State-of-the-art Earth System Models now represent these climate-carbon cycle feedbacks, always simulating a positive feedback over the twentieth and twenty-first centuries, although with substantial uncertainty. Recent studies now help to reduce this uncertainty. First, on short timescales, El Ni o years record larger than average atmospheric CO2 growth rate, with tropical land ecosystems being the main drivers. These climate-carbon cycle anomalies can be used as emerging constraint on the tropical land carbon response to future climate change. Second, centennial variability found in last millennium records can be used to constrain the overall global carbon cycle response to climatic excursions. These independent methods point to climate-carbon cycle feedback at the low-end of the Earth System Models range, indicating that these models overestimate the carbon cycle sensitivity to climate change. These new findings also help to attribute the historical land and ocean carbon sinks to increase in atmospheric CO2 and climate change.
DOI:10.1007/BF00384433 [本文引用: 1]
The cell content of 12 bacterial phospholipid fatty acids (PLFA) was determined in bacteria extracted from soil by homogenization/centrifugation. The bacteria were enumerated using acridine orange direct counts. An average of 1.40×10 -17 mol bacterial PLFA cell -1 was found in bacteria extracted from 15 soils covering a wide range of pH and organic matter contents. With this factor, the bacterial biomass based on PLFA analyses of whole soil samples was calculated as 1.0–4.8 mg bacterial C g -1 soil C. The corresponding range based on microscopical counts was 0.3–3.0 mg bacterial C g -1 soil C. The recovery of bacteria from the soils using homogenization/centrifugation was 2.6–16% (mean 8.7%) measured by PLFA analysis, and 12–61% (mean 26%) measured as microscopical counts. The soil content of the PLFA 18:2ω6 was correlated with the ergosterol content ( r =0.92), which supports the use of this PLFA as an indicator of fungal biomass. The ratio 18:2ω6 to bacterial PLFA is therefore suggested as an index of the fungal:bacterial biomass ratio in soil. An advantage with the method based on PLFA analyses is that the same technique and even the same sample is used to determine both fungi and bacteria. The fungal:bacterial biomass ratio calculated in this way was positively correlated with the organic matter content of the soils ( r =0.94).
DOI:10.1016/0038-0717(93)90113-PURL [本文引用: 1]
The effects of lime and wood-ash on the microbial community structure were evaluated by analyzing the phospholipid fatty acid (PLFA) composition of soils from two areas in the south of Sweden. A pine forest was amended with lime or ash at two concentrations, and a spruce forest was limed at one concentration. The treatments were carried out 5–6 years before sampling and raised the pH from approx. 4.0 to values between 4.8 and 7.0. At both sites there was a difference in the PLFA composition between the treated plots and the controls. The changes found were similar at both sites and correlated to the pH changes. No difference was found between limed plots and those treated with wood-ash. The methyl-branched fatty acids i15:0, i16:0 and 10Me16:0, the monounsaturated fatty acids 16: 1ω 7t and 18: 1ω 9, the cyclopropane fatty acid cy 19:0, and the saturated fatty acid 20:0 were more abundant in the control plots. In the plots with the highest pH there was a three-fold increase in the fatty acid 16: lω 5. An increase was also found for the fatty acids i14:0, 16:lω9, 16:lω 7c, cy17:0, 18:lω 7 and 10Me18:0. No effect on 18:2ω6 was found. The changes in PLFA pattern indicated that the increased pH caused a shift in the bacterial community to more Gram-negative and fewer Gram-positive bacteria, while the amount of fungi was unaffected. The increase in 10Me18:0 in limed soils indicated an increase in actinomycetes.
DOI:10.1016/j.soilbio.2010.11.021URL [本文引用: 2]
The determination of the phospholipid fatty acid (PLFA) pattern of soil organisms has become one of the most commonly used methods to study microbial community structure. Here we recapitulate the background of our work applying the PLFA method to soil in the early 1990s. We also stress that although the PLFA method was, and still is, a rapid and sensitive method to detect changes in the microbial community in soil, as with all popular methods it can be misused. We discuss problems in PLFA interpretation, the extent of turn-over of PLFAs in soil, and the flawed use of diversity indices to evaluate PLFA patterns.
DOI:10.1111/j.1574-6976.2012.00343.xURLPMID:22568555 [本文引用: 1]
Soil is increasingly under environmental pressures that alter its capacity to fulfil essential ecosystem services. To maintain these crucial soil functions, it is important to know how soil microorganisms respond to disturbance or environmental change. Here, we summarize the recent progress in understanding the resistance and resilience (stability) of soil microbial communities and discuss the underlying mechanisms of soil biological stability together with the factors affecting it. Biological stability is not solely owing to the structure or diversity of the microbial community but is linked to a range of other vegetation and soil properties including aggregation and substrate quality. We suggest that resistance and resilience are governed by soil physico-chemical structure through its effect on microbial community composition and physiology, but that there is no general response to disturbance because stability is particular to the disturbance and soil history. Soil stability results from a combination of biotic and abiotic soil characteristics and so could provide a quantitative measure of soil health that can be translated into practice.
DOI:10.3724/SP.J.1003.2013.10033URL [本文引用: 1]
自20世纪50年代以来,生物多样性与生态系统稳定性的关系一直是生态学中重点讨论的理论问题之一。在当今人类活动对自然生态系统产生重大影响的情况下,全面理解生态系统多样性与稳定性的关系,有助于我们更好地应对环境变化和生物多样性丧失等生态问题。在陆地生态系统中,关注重点多集中在地上植物生态系统;而对地下生态系统,尤其是对微生物多样性与系统稳定性关系的研究尚重视不够。事实上,土壤微生物作为生命元素循环的驱动者,主导和参与地下生态系统中一系列重要生态过程,对土壤能否正常有序地执行各项生态功能至关重要。对土壤微生物多样性的研究,能使我们明确土壤中微生物对各种环境条件(包括自然和人为因素)变化的响应机制,更好地维持土壤生态系统的稳定性及其生态服务功能。本文在介绍土壤微生物多样性概念、研究方法、地下生态系统稳定性的基础上,重点讨论了土壤微生物多样性对土壤生态系统稳定性的影响,对多样性一稳定性关系在土壤微生物生态学中的应用进行了较为深入和全面的思考。作者提出,土壤微生物系统是一个动态变化的自组织系统,通过遗传来维持其组成和结构的相对稳定性,通过变异而适应外界干扰,共同构成土壤微生物系统的抵抗力(resistance)~恢复力(resilience),维护土壤生态系统的稳定性。今后土壤微生物多样性一稳定性关系的研究,需要注重地上与地下生态系统的结合与统一,借鉴宏观生态学理论来构建微生物生态学的理论框架,建立微生物多样性一稳定性关系的机理模型,从定性描述向定量表征方向发展。
DOI:10.3724/SP.J.1003.2013.10033URL [本文引用: 1]
自20世纪50年代以来,生物多样性与生态系统稳定性的关系一直是生态学中重点讨论的理论问题之一。在当今人类活动对自然生态系统产生重大影响的情况下,全面理解生态系统多样性与稳定性的关系,有助于我们更好地应对环境变化和生物多样性丧失等生态问题。在陆地生态系统中,关注重点多集中在地上植物生态系统;而对地下生态系统,尤其是对微生物多样性与系统稳定性关系的研究尚重视不够。事实上,土壤微生物作为生命元素循环的驱动者,主导和参与地下生态系统中一系列重要生态过程,对土壤能否正常有序地执行各项生态功能至关重要。对土壤微生物多样性的研究,能使我们明确土壤中微生物对各种环境条件(包括自然和人为因素)变化的响应机制,更好地维持土壤生态系统的稳定性及其生态服务功能。本文在介绍土壤微生物多样性概念、研究方法、地下生态系统稳定性的基础上,重点讨论了土壤微生物多样性对土壤生态系统稳定性的影响,对多样性一稳定性关系在土壤微生物生态学中的应用进行了较为深入和全面的思考。作者提出,土壤微生物系统是一个动态变化的自组织系统,通过遗传来维持其组成和结构的相对稳定性,通过变异而适应外界干扰,共同构成土壤微生物系统的抵抗力(resistance)~恢复力(resilience),维护土壤生态系统的稳定性。今后土壤微生物多样性一稳定性关系的研究,需要注重地上与地下生态系统的结合与统一,借鉴宏观生态学理论来构建微生物生态学的理论框架,建立微生物多样性一稳定性关系的机理模型,从定性描述向定量表征方向发展。
DOI:10.1126/science.aal1319URLPMID:28280251 [本文引用: 1]
Soil organic carbon harbors three times as much carbon as Earth’s atmosphere, and its decomposition is a potentially large climate change feedback and major source of uncertainty in climate projections. The response of whole-soil profiles to warming has not been tested in situ. In a deep warming experiment in mineral soil, we found that CO2 production from all soil depths increased with 4°C warming; annual soil respiration increased by 34 to 37%. All depths responded to warming with similar temperature sensitivities, driven by decomposition of decadal-aged carbon. Whole-soil warming reveals a larger soil respiration response than many in situ experiments (most of which only warm the surface soil) and models. Authors: Caitlin E. Hicks Pries, C. Castanha, R. C. Porras, M. S. Torn
DOI:10.1016/j.soilbio.2014.04.028URL [本文引用: 1]
61Increased carbon inputs in laboratory cause a positive priming of SOC decomposition.61Increased carbon inputs to soil from 1002y of eCO2 similarly altered SOC decomposition.61Warming increased the rate, not amount, of SOC primed by increased carbon inputs.61Microbial substrate use efficiency for added sucrose was unchanged by eCO2 or warming.61The priming effect did not change the proportion of different ages of C in respiration.
[本文引用: 2]
[本文引用: 1]
DOI:10.1038/nature16045URLPMID:26633767 [本文引用: 1]
Tuberculosis is a major source of global mortality caused by infection, partly because of a tremendous ongoing burden of undiagnosed disease. Improved diagnostic technology may play an increasingly crucial part in global efforts to end tuberculosis, but the ability of diagnostic tests to curb tuberculosis transmission is dependent on multiple factors, including the time taken by a patient to seek health care, the patient's symptoms, and the patterns of transmission before diagnosis. Novel diagnostic assays for tuberculosis have conventionally been evaluated on the basis of characteristics such as sensitivity and specificity, using assumptions that probably overestimate the impact of diagnostic tests on transmission. We argue for a shift in focus to the evaluation of such tests' incremental value, defining outcomes that reflect each test's purpose (for example, transmissions averted) and comparing systems with the test against those without, in terms of those outcomes. Incremental value can also be measured in units of outcome per incremental unit of resource (for example, money or human capacity). Using a novel, simplified model of tuberculosis transmission that addresses some of the limitations of earlier tuberculosis diagnostic models, we demonstrate that the incremental value of any novel test depends not just on its accuracy, but also on elements such as patient behaviour, tuberculosis natural history and health systems. By integrating these factors into a single unified framework, we advance an approach to the evaluation of new diagnostic tests for tuberculosis that considers the incremental value at the population level and demonstrates how additional data could inform more-effective implementation of tuberculosis diagnostic tests under various conditions.
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/(ISSN)1097-0088URL [本文引用: 1]
DOI:10.1111/gcbb.2009.1.issue-1URL [本文引用: 1]
DOI:10.1111/j.1574-6941.2008.00620.xURLPMID:19049494 [本文引用: 1]
Soil samples were collected along two slopes (south and north) at subalpine (15000900091900 m, under closed vegetation, up to the forest line) and alpine altitudes (23000900092530, under scattered vegetation, above the forest line) in the Grossglockner mountain area (Austrian central Alps). Soils were analyzed for a number of properties, including physical and chemical soil properties, microbial activity and microbial communities that were investigated using culture-dependent (viable heterotrophic bacteria) and culture-independent methods (phospholipid fatty acid analysis, FISH). Alpine soils were characterized by significantly ( P <0.01) colder climate conditions, i.e. lower mean annual air and soil temperatures, more frost and ice days and higher precipitation, compared with subalpine soils. Microbial activity (soil dehydrogenase activity) decreased with altitude; however, dehydrogenase activity was better adapted to cold in alpine soils compared with subalpine soils, as shown by the lower apparent optimum temperature for activity (30 vs. 37 00°C) and the significantly ( P <0.010900090.001) higher relative activity in the low-temperature range. With increasing altitude, i.e. in alpine soils, a significant ( P <0.050900090.01) increase in the relative amount of culturable psychrophilic heterotrophic bacteria, in the relative amount of the fungal population and in the relative amount of Gram-negative bacteria was found, which indicates shifts in microbial community composition with altitude.
[本文引用: 1]
DOI:10.1007/s00248-007-9295-1URL [本文引用: 1]
The impact and frequency of forest harvesting could significantly affect soil microbial community (SMC) structure and functioning. The ability of soil microorganisms to perform biogeochemical processes is critical for sustaining forest productivity and has a direct impact on decomposition dynamics and carbon storage potential. The Wind River Canopy Crane Research Forest in SW, WA, provided a unique opportunity to study a forest chronosequence and the residual effects of harvesting on the SMC in comparison to old-growth forests. The objective of this study was to determine the effect of clear-cutting and stand age on temporal dynamics of SMC and physiological stress markers using phospholipid fatty acid (PLFA) profiling. Soil microbial PLFA profiles were determined seven times over 22 months (Nov. 02 to Sep. 04) in old-growth coniferous forest stands (300-500 years) and 8 (CC8)- or 25 (CC25)-year-old replanted clear-cuts. PLFA patterns of the SMC shifted because of clear-cutting, but seasonal temporal changes had greater shifts than differences among stand age. The microbial biomass (total PLFA) and bacterial, fungal, and selected other PLFAs were significantly reduced in CC8 but not in CC25 sites relative to the old-growth sites. An increase in stress indicators [PLFA ratios of saturated/monsaturated and (cy17:0 + cy19:0)/(16:1 omega 7 + 18:1 omega 7)] in late summer was related to water stress. Although the canopy and litter input are quite different for a 25-year clear-cut compared to virgin old-growth forest, we conclude that the composition of the microbial communities, 25 years after clear-cutting, has recovered sufficiently to be much more similar to old-growth forests than a recent clear-cut at this Pacific Northwest forest site. The study shows the potential of PLFA analysis for profiling microbial communities and their stress status under field conditions, but wide temporal shifts emphasize the need for sampling over seasons to fully interpret ecosystem man
DOI:10.1016/j.soilbio.2015.06.027URL [本文引用: 1]
61Afforestation induced shift in soil respiration is linked to microbial community.61Elevated temperature consistently increase soil respiration.61Metabolic rate and substrate quality explain soil respiration response to warming.
DOI:10.1111/j.1574-6941.1999.tb00621.xURL [本文引用: 1]
Abstract Mycorrhizal fungi form extensive mycelia in soil and play significant roles in most soil ecosystems. The estimation of their biomasses is thus of importance in order to understand their possible role in soil nutrient processes. For arbuscular mycorrhizal (AM) fungi the signature fatty acid 16:1 5 provides a new and promising tool for the estimation of AM fungal biomass in soil and roots. For ectomycorrhizal fungi 18:2 6,9 dominates among the fatty acids and can be used as an indicator of mycelial biomass of these fungi in soil in experimental systems. In biomass estimation primarily the phospholipid fatty acids (PLFAs) are suitable. Through the use of specific PLFAs it is possible to study interactions between mycorrhizal mycelia and bacteria in soil as well as between AM fungal mycelia and mycelia of saprophytic and parasitic fungi in soil and in roots. AM fungi, in particular, store a large proportion of their energy as lipids and by using the signature fatty acids it is possible to determine the relation between membrane and storage lipids, which could be an indication of energy storage levels. Various aspects of how the fatty acid signatures can be used for studies related to questions of biomass distribution and nutritional status of mycorrhizal fungi are discussed.
DOI:10.1016/j.soilbio.2013.12.003URL
Little is known about how spatial and environmental patterns structure soil microbial activities. We investigated, on 47 soil samples collected in Mediterranean forests, the net and interaction effects of climatic-geographic and edaphic variables as well as vegetation cover and composition on soil microbial community-level physiological profiles (CLPPs) assessed by Microresp64. The effects of these variables were also analyzed on CLPP response to an experimental drought treatment. CLPPs were shown to be mainly driven by climate–plant–soil and plant–soil interactions; even after drought treatment, there was a decrease in microbial activity but no change in CLPPs. Our findings highlight the robustness of these relationships, which need to be assessed within different ecosystems considering various spatial scales to reliably predict climate change effects on terrestrial ecosystems.
DOI:10.1111/gcb.13789URLPMID:28614594 [本文引用: 2]
Abstract Unprecedented levels of nitrogen (N) have entered terrestrial ecosystems over the past century, which substantially influences the carbon (C) exchange between the atmosphere and biosphere. Temperature and moisture are generally regarded as the0002major controllers over the N effects on ecosystem C uptake and release. N-phosphorous (P) stoichiometry regulates the growth and metabolisms of plants and soil organisms, thereby affecting many ecosystem C processes. However, it remains unclear how the N-induced shift in the plant N:P ratio affects ecosystem production and C fluxes and its relative importance. We conducted a field manipulative experiment with eight N addition levels in a Tibetan alpine steppe and assessed the influences of N on aboveground net primary production (ANPP), gross ecosystem productivity (GEP), ecosystem respiration (ER), and net ecosystem exchange (NEE); we used linear mixed-effects models to further determine the relative contributions of various factors to the N-induced changes in these parameters. Our results showed that the ANPP, GEP, ER, and NEE all exhibited nonlinear responses to increasing N additions. Further analysis demonstrated that the plant N:P ratio played a dominate role in shaping these C exchange processes. There was a positive relationship between the N-induced changes in ANPP (0200ANPP) and the plant N:P ratio (0200N:P), whereas the 0200GEP, 0200ER, and 0200NEE exhibited quadratic correlations with the 0200N:P. In contrast, soil temperature and moisture were only secondary predictors for the changes in ecosystem production and C fluxes along the N addition gradient. These findings highlight the importance of plant N:P ratio in regulating ecosystem C exchange, which is crucial for improving our understanding of C cycles under the scenarios of global N enrichment. 0008 2017 John Wiley & Sons Ltd.
DOI:10.1111/gcb.2007.13.issue-1URL [本文引用: 1]
DOI:10.1016/j.soilbio.2016.12.026URL [本文引用: 1]
Numerous field studies have found changes in soil respiration and microbial abundance under experimental warming. Yet, it is uncertain whether the magnitude of these responses remains consistent over the long-term. We performed a meta-analysis on 25 field experiments to examine how warming effects on soil respiration, microbial biomass, and soil microbial C respond to the duration of warming. For each parameter, we hypothesized that effect sizes of warming would diminish as the duration of warming increased. In support of our hypothesis, warming initially increased soil respiration, but the magnitude of this effect declined significantly as warming progressed as evidenced by the two longest studies in our meta-analysis. In fact, after 10 years of warming, soil respiration in warmed treatments was similar to controls. In contrast, warming effect sizes for fungal biomass, bacterial biomass, and soil microbial C did not respond significantly to the duration of warming. Microbial acclimation, community shifts, adaptation, or reductions in labile C may have ameliorated warming effects on soil respiration in the long-term. Accordingly, long-term soil C losses might be smaller than those suggested by short-term warming studies.
DOI:10.1038/nature10386URLPMID:21979045 [本文引用: 1]
Abstract Globally, soil organic matter (SOM) contains more than three times as much carbon as either the atmosphere or terrestrial vegetation. Yet it remains largely unknown why some SOM persists for millennia whereas other SOM decomposes readily--and this limits our ability to predict how soils will respond to climate change. Recent analytical and experimental advances have demonstrated that molecular structure alone does not control SOM stability: in fact, environmental and biological controls predominate. Here we propose ways to include this understanding in a new generation of experiments and soil carbon models, thereby improving predictions of the SOM response to global warming.
DOI:10.1038/ismej.2011.32URLPMID:3176507 [本文引用: 1]
The soil microbiome is responsible for mediating key ecological processes; however, little is known about its sensitivity to climate change. Observed increases in global temperatures and alteration to rainfall patterns, due to anthropogenic release of greenhouse gases, will likely have a strong influence on soil microbial communities and ultimately the ecosystem services they provide. Therefore, it is vital to understand how soil microbial communities will respond to future climate change scenarios. To this end, we surveyed the abundance, diversity and structure of microbial communities over a 2-year period from a long-term in situ warming experiment that experienced a moderate natural drought. We found the warming treatment and soil water budgets strongly influence bacterial population size and diversity. In normal precipitation years, the warming treatment significantly increased microbial population size 40-150% but decreased diversity and significantly changed the composition of the community when compared with the unwarmed controls. However during drought conditions, the warming treatment significantly reduced soil moisture thereby creating unfavorable growth conditions that led to a 50-80% reduction in the microbial population size when compared with the control. Warmed plots also saw an increase in species richness, diversity and evenness; however, community composition was unaffected suggesting that few phylotypes may be active under these stressful conditions. Our results indicate that under warmed conditions, ecosystem water budget regulates the abundance and diversity of microbial populations and that rainfall timing is critical at the onset of drought for sustaining microbial populations.
URL [本文引用: 1]
土壤是地球表层最为重要的碳库也是温室气体的源或汇。自丁业革命以来,对土壤温室气体的容量、收支平衡和通量等已有较多研究和估算,但对关键过程及其源/汇的研究却十分有限。微生物是土壤碳氮转化的主要驱动者,在生态系统碳氮循环过程中扮演重要的角色,对全球气候变化有着响应的响应、适应及反馈,然而其个体数量,群落结构和多样性如何与气候扰动相互关联、进而怎样影响生态系统过程的问题仍有待进一步探索。从微生物介导的碳氮循环过程人手,重点讨论微生物对气候变化包括温室气体(CO2,CH4,N20)增加、全球变暖、大气氮沉降等的响应和反馈,并由此提出削减温室气体排放的可能途径和今后发展的方向。
URL [本文引用: 1]
土壤是地球表层最为重要的碳库也是温室气体的源或汇。自丁业革命以来,对土壤温室气体的容量、收支平衡和通量等已有较多研究和估算,但对关键过程及其源/汇的研究却十分有限。微生物是土壤碳氮转化的主要驱动者,在生态系统碳氮循环过程中扮演重要的角色,对全球气候变化有着响应的响应、适应及反馈,然而其个体数量,群落结构和多样性如何与气候扰动相互关联、进而怎样影响生态系统过程的问题仍有待进一步探索。从微生物介导的碳氮循环过程人手,重点讨论微生物对气候变化包括温室气体(CO2,CH4,N20)增加、全球变暖、大气氮沉降等的响应和反馈,并由此提出削减温室气体排放的可能途径和今后发展的方向。
DOI:10.1038/464499aURLPMID:20336128 [本文引用: 1]
The flux of carbon from soils to the atmosphere has apparently increased with climate warming. But does this reflect a net loss of carbon to the atmosphere that could exacerbate climate change?
DOI:10.1007/s00442-014-2894-xURLPMID:24532178 [本文引用: 1]
Global change phenomena, such as forest disturbance and land-use change, significantly affect elemental balances as well as the structure and function of terrestrial ecosystems. However, the importance of shifts in soil nutrient stoichiometry for the regulation of belowground biota and soil food webs have not been intensively studied for tropical ecosystems. In the present account, we examine the effects of land-use change and soil depth on soil and microbial stoichiometry along a land-use sequence (natural forest, pastures of different ages, secondary succession) in the tropical mountain rainforest region of southern Ecuador. Furthermore, we analyzed (PLFA-method) whether shifts in the microbial community structure were related to alterations in soil and microbial stoichiometry. Soil and microbial stoichiometry were affected by both land-use change and soil depth. After forest disturbance, significant decreases of soil C:N:P ratios at the pastures were followed by increases during secondary succession. Microbial C:N ratios varied slightly in response to land-use change, whereas no fixed microbial C:P and N:P ratios were observed. Shifts in microbial community composition were associated with soil and microbial stoichiometry. Strong positive relationships between PLFA-markers 18:2n6,9c (saprotrophic fungi) and 20:4 (animals) and negative associations between 20:4 and microbial N:P point to land-use change affecting the structure of soil food webs. Significant deviations from global soil and microbial C:N:P ratios indicated a major force of land-use change to alter stoichiometric relationships and to structure biological systems. Our results support the idea that soil biotic communities are stoichiometrically flexible in order to adapt to alterations in resource stoichiometry.
DOI:10.1016/0038-0717(87)90052-6URL
The effects of fumigation on organic C extractable by 0.5 M K 2SO 4 were examined in a contrasting range of soils. E C (the difference between organic C extracted by 0.5 M K 2SO 4 from fumigated and non-fumigated soil) was about 70% of F C (the flush of CO 2-C caused by fumigation during a 10 day incubation), meaned for ten soils. There was a close relationship between microbial biomass C, measured by fumigation-incubation (from the relationship Biomass C = F C/0.45) and E C given by the equation: Biomass C = (2.64 0.060) E C that accounted for 99.2% of the variance in the data. This relationship held over a wide range of soil pH (3.9 8.0). ATP and microbial biomass N concentrations were measured in four of the soils. The (ATP) (E C) ratios were very similar in the four soils, suggesting that both ATP and the organic C rendered decomposable by CHCl 3 came from the soil microbial biomass. The C:N ratio of the biomass in a strongly acid (pH 4.2) soil was greater (9.4) than in the three less-acid soils (mean C:N ratio 5.1). We propose that the organic C rendered extractable to 0.5 m K 2SO 4 after a 24 h CHCl 3-fumigation ( E C) comes from the cells of the microbial biomass and can be used to estimate soil microbial biomass C in both neutral and acid soils.
DOI:10.11654/jaes.2015.11.019URL [本文引用: 1]
通过对35例2009—2015年初发表的增温控制试验的96组数据进行Meta分析,研究了土壤微生物对增温的响应模式,结果表明:增温提高了土壤微生物碳(1.4%)、土壤微生物氮(1.7%)和土壤真菌量(3.7%),降低了土壤细菌量(-1.9%);土壤微生物碳的增温响应与试验地年平均降水量和增温幅度显著相关;土壤微生物氮的变化受试验地年均温、年均降水量及增温幅度的影响。
DOI:10.11654/jaes.2015.11.019URL [本文引用: 1]
通过对35例2009—2015年初发表的增温控制试验的96组数据进行Meta分析,研究了土壤微生物对增温的响应模式,结果表明:增温提高了土壤微生物碳(1.4%)、土壤微生物氮(1.7%)和土壤真菌量(3.7%),降低了土壤细菌量(-1.9%);土壤微生物碳的增温响应与试验地年平均降水量和增温幅度显著相关;土壤微生物氮的变化受试验地年均温、年均降水量及增温幅度的影响。
DOI:10.1111/j.1365-2486.2011.02548.xURL [本文引用: 1]
Abstract The balance of primary production and decomposition in northern peatlands may shift due to climate change, with potential feedbacks to atmospheric CO2 concentrations. Nitrogen availability will modulate this shift, but little is known about the drivers of soil nitrogen dynamics in these environments. We used a long-term (9 years) open top chamber (OTC) experiment in an ombrotrophic Sphagnum peat bog in sub-arctic Sweden, to test for the interactive effects of spring warming, summer warming and winter snow addition on soil nitrogen fluxes, potential activities of nitrogen cycle enzymes, and soil microbial community composition. These simultaneous measurements allowed us to identify the level of organization at which climate change impacts are apparent, an important requirement for developing truly mechanistic understanding. Organic-N pools and fluxes were an order of magnitude higher than inorganic-N pools and fluxes. Summer warming approximately doubled fluxes of soil organic nitrogen and ammonia over the growing season. Such a large increase under 1 C warming is unlikely to be due to kinetic effects, and we propose that it is linked to an observed seasonal decrease in microbial biomass, suggesting that N flux is driven by a substantial late-season dieback of microbes. This change in N cycle dynamics was not reflected in any of the measured potential peptidase activities. Moreover, the soil microbial community structure was apparently stable across treatments, suggesting a non-specific microbial dieback. Our results show that in these widespread peat bogs, where many plant species are capable of organic-N uptake, organic soil N dynamics are quantitatively far more important than the commonly studied inorganic-N dynamics. Understanding of climate change effects on organic soil N cycling in this system will be advanced by closer investigation of the seasonal dynamics of the microbial biomass and the input of substrates that maintain it.
DOI:10.1073/pnas.1519620113URLPMID:27114518 [本文引用: 1]
Abstract The global terrestrial carbon sink offsets one-third of the world's fossil fuel emissions, but the strength of this sink is highly sensitive to large-scale extreme events. In 2012, the contiguous United States experienced exceptionally warm temperatures and the most severe drought since the Dust Bowl era of the 1930s, resulting in substantial economic damage. It is crucial to understand the dynamics of such events because warmer temperatures and a higher prevalence of drought are projected in a changing climate. Here, we combine an extensive network of direct ecosystem flux measurements with satellite remote sensing and atmospheric inverse modeling to quantify the impact of the warmer spring and summer drought on biosphere-atmosphere carbon and water exchange in 2012. We consistently find that earlier vegetation activity increased spring carbon uptake and compensated for the reduced uptake during the summer drought, which mitigated the impact on net annual carbon uptake. The early phenological development in the Eastern Temperate Forests played a major role for the continental-scale carbon balance in 2012. The warm spring also depleted soil water resources earlier, and thus exacerbated water limitations during summer. Our results show that the detrimental effects of severe summer drought on ecosystem carbon storage can be mitigated by warming-induced increases in spring carbon uptake. However, the results also suggest that the positive carbon cycle effect of warm spring enhances water limitations and can increase summer heating through biosphere-atmosphere feedbacks.
DOI:10.1038/nclimate2940URL [本文引用: 2]
Microbial decomposition of soil carbon in high-latitude tundra underlain with permafrost is one of the most important, but poorly understood, potential positive feedbacks of greenhouse gas emissions from terrestrial ecosystems into the atmosphere in a warmer world. Using integrated metagenomic technologies, we showed that the microbial functional community structure in the active layer of tundra soil was significantly altered after only 1.5 years of warming, a rapid response demonstrating the high sensitivity of this ecosystem to warming. The abundances of microbial functional genes involved in both aerobic and anaerobic carbon decomposition were also markedly increased by this short-term warming. Consistent with this, ecosystem respiration (R) increased up to 38%. In addition, warming enhanced genes involved in nutrient cycling, which very likely contributed to an observed increase (30%) in gross primary productivity (GPP). However, the GPP increase did not offset the extra R, resulting in significantly more net carbon loss in warmed plots compared with control plots. Altogether, our results demonstrate the vulnerability of active-layer soil carbon in this permafrost-based tundra ecosystem to climate warming and the importance of microbial communities in mediating such vulnerability.
DOI:10.1128/ mBio.00976-16URLPMID:27677789 [本文引用: 2]
Abstract Clipping (i.e., harvesting aboveground plant biomass) is common in agriculture and for bioenergy production. However, microbial responses to clipping in the context of climate warming are poorly understood. We investigated the interactive effects of grassland warming and clipping on soil properties and plant and microbial communities, in particular, on microbial functional genes. Clipping alone did not change the plant biomass production, but warming and clipping combined increased the C 4 peak biomass by 47% and belowground net primary production by 110%. Clipping alone and in combination with warming decreased the soil carbon input from litter by 81% and 75%, respectively. With less carbon input, the abundances of genes involved in degrading relatively recalcitrant carbon increased by 38% to 137% in response to either clipping or the combined treatment, which could weaken long-term soil carbon stability and trigger positive feedback with respect to warming. Clipping alone also increased the abundance of genes for nitrogen fixation, mineralization, and denitrification by 32% to 39%. Such potentially stimulated nitrogen fixation could help compensate for the 20% decline in soil ammonium levels caused by clipping alone and could contribute to unchanged plant biomass levels. Moreover, clipping tended to interact antagonistically with warming, especially with respect to effects on nitrogen cycling genes, demonstrating that single-factor studies cannot predict multifactorial changes. These results revealed that clipping alone or in combination with warming altered soil and plant properties as well as the abundance and structure of soil microbial functional genes. Aboveground biomass removal for biofuel production needs to be reconsidered, as the long-term soil carbon stability may be weakened. IMPORTANCE: Global change involves simultaneous alterations, including those caused by climate warming and land management practices (e.g., clipping). Data on the interactive effects of warming and clipping on ecosystems remain elusive, particularly in microbial ecology. This study found that clipping alters microbial responses to warming and demonstrated the effects of antagonistic interactions between clipping and warming on microbial functional genes. Clipping alone or combined with warming enriched genes degrading relatively recalcitrant carbon, likely reflecting the decreased quantity of soil carbon input from litter, which could weaken long-term soil C stability and trigger positive warming feedback. These results have important implications in assessing and predicting the consequences of global climate change and indicate that the removal of aboveground biomass for biofuel production may need to be reconsidered. Copyright 2016 Xue et al.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10142-014-0419-7URLPMID:25471470 [本文引用: 1]
Abstract Natural selection drives species adaptations to biotic and abiotic stresses. Species distributed along a moisture gradient, such as Stipa purpurea, a dominant grass in alpine arid and semi-arid meadows on the Tibetan Plateau, provide an opportunity to evaluate the effects of long-term adaptation to differing degrees of drought stress on gene expression. However, the genetic basis of this divergence remains largely unknown. Next-generation sequencing technologies have provided important genome-wide insights on the evolution of organisms for which genomic information is lacking. To understand how S. purpurea responds to drought stress, we selected five populations distributed along the degressive rainfall line on the northwestern Tibetan Plateau that currently present evolutionary acclimation to localized drought pressure at the physiological and biochemical levels and compared their transcriptome responses. In addition, we performed de novo assembly of the S. purpurea transcriptome using short read sequencing technology and successfully assembled 84,298 unigenes from approximately 51 million sequencing reads. We quantified gene expression level to compare their transcriptome responses using mRNA-Seq and identified differentially expressed transcripts that are involved in primary and secondary plant metabolism, plant hormone synthesis, defense responses, and cell wall synthesis. Furthermore, physiological and biochemical evidence supports that abscisic acid (ABA) accumulation and cell wall strengthening derived from the differential transcripts contribute to the tolerance of S. purpurea to drought stress. The mechanisms by which S. purpurea adapts to drought stress provide new insight into how plants ecologically adapt and evolve.
DOI:10.1890/12-2018.1URLPMID:24358718
Climate change models predict that future precipitation patterns will entail lower-frequency but larger rainfall events, increasing the duration of dry soil conditions. Resulting shifts in microbial C cycling activity could affect soil C storage. Further, microbial response to rainfall events may be constrained by the physiological or nutrient limitation stress of extended drought periods; thus seasonal or multiannual precipitation regimes may influence microbial activity following soil wet-up. We quantified rainfall-driven dynamics of microbial processes that affect soil C loss and retention, and microbial community composition, in soils from a long-term (14-year) field experiment contrasting "Ambient" and "Altered" (extended intervals between rainfalls) precipitation regimes. We collected soil before, the day following, and five days following 2.5-cm rainfall events during both moist and dry periods (June and September 2011; soil water potential = -0.01 and -0.83 MPa, respectively), and measured microbial respiration, microbial biomass, organic matter decomposition potential (extracellular enzyme activities), and microbial community composition (phospholipid fatty acids). The equivalent rainfall events caused equivalent microbial respiration responses in both treatments. In contrast, microbial biomass was higher and increased after rainfall in the Altered treatment soils only, thus microbial C use efficiency (CUE) was higher in Altered than Ambient treatments (0.70 +/- 0.03 > 0.46 +/- 0.10). CUE was also higher in dry (September) soils. C-acquiring enzyme activities (beta-glucosidase, cellobiohydrolase, and phenol oxidase) increased after rainfall in moist (June), but not dry (September) soils. Both microbial biomass C:N ratios and fungal:bacterial ratios were higher at lower soil water contents, suggesting a functional and/or population-level shift in the microbiota at low soil water contents, and microbial community composition also differed following wet-up and between seasons and treatments. Overall, microbial activity may directly (C respiration) and indirectly (enzyme potential) reduce soil organic matter pools less in drier soils, and soil C sequestration potential (CUE) may be higher in soils with a history of extended dry periods between rainfall events. The implications include that soil C loss may be reduced or compensated for via different mechanisms at varying time scales, and that microbial taxa with better stress tolerance or growth efficiency may be associated with these functional shifts.
DOI:10.1016/j.soilbio.2015.06.022URL [本文引用: 1]
Terrestrial ecosystems experience simultaneous shifts in multiple drivers of global change, which can interactively affect various resources. The concept that different resources co-limit plant productivity has been well studied. However, co-limitation of soil microbial communities by multiple resources has not been as thoroughly investigated. Specifically, it is not clearly understood how microbial communities respond to shifts in multiple interacting resources such as water, temperature, and nitrogen (N), in the context of global change. To test the effects of these various resources on soil microorganisms, we established a field experiment with temperature and N manipulation in three grasslands of northern China, where there is a decrease in precipitation from east to west across the region. We found that microbial responses to temperature depended upon seasonal water regimes in these temperate steppes. When there was sufficient water present, warming had positive effects on soil microorganisms, suggesting an interaction between water and increases in temperature enhanced local microbial communities. When drought or alternating wet ry stress occurred, warming had detrimental effects on soil microbial communities. Our results also provide clear evidence for serial co-limitation of microorganisms by water and N at the functional group and community levels, where water is a primary limiting factor and N addition positively affects soil microorganisms only when water is sufficient. We predict that future microbial responses to changes in temperature and N availability could be seasonal or exist only in non-drought years, and will strongly rely on future precipitation regimes.
DOI:10.1016/j.apsoil.2014.11.012URL [本文引用: 2]
Alpine ecosystems at high altitudes and latitudes are notably sensitive to climatic warming and the Tibetan Plateau is a widely distributed alpine ecosystem. The magnitude of climatic warming on the Tibetan Plateau is expected to be considerably greater than the global average. However, a synthesis of the experimental warming soil carbon and nitrogen data is still lacking and whether forest soils are more sensitive to warming than grassland soils remains unclear. In this study, we used a meta-analysis approach to synthesise 196 observations from 25 published studies on the Tibetan Plateau. Warming significantly increased microbial biomass carbon (MBC) by 14.3% (95% CI: 2.9–24.6%), microbial biomass nitrogen (MBN) by 20.1% (95% CI: 2.0–45.1%), net nitrogen mineralization by 49.2% (95% CI: 38.1–62.3%) and net nitrification by 56.0% (95% CI: 51.4–66.1%), but did not significantly affect soil carbon (95% CI: 6113.9 to 2.7%) or nitrogen (95% CI: 6112.4 to 2.6%). The mean annual air temperature was negatively correlated with the warming effects on MBC and MBN. Grasslands exhibited significant MBC and MBN responses to warming. Specifically, soil microbial biomass was more responsive to warming in colder environments. Moreover, forest soils are not always more sensitive to warming than grassland soils as previous studies have suggested. These findings indicate that clarifying the effect of warming on alpine soils need consider ecosystem types and their local climate.
DOI:10.1038/nclimate1331URL
Warmer winters increase the rhizosphere carbon flow to mycorrhizal fungi more than to other microorganisms in a temperate grassland
1
2017
... id="C29">首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定.一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(
Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles
1998
Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogelimatic zones in western Canada
1
2012
... id="C30">其次, 土壤温度和水分等环境因素会对土壤微生物群落产生不同影响, 进而抑制土壤微生物对气候变暖的响应.大量研究表明, 土壤温度与土壤含水量对微生物的生理状态、生物量及群落结构有重要影响(
Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil
1
1985
... id="C12">为了揭示短期增温对土壤微生物生物量的影响, 我们基于氯仿熏蒸法(
The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau
1
2013
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
Patterns and drivers of soil microbial communities in Tibetan alpine and global terrestrial ecosystems
1
2016
... id="C29">首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定.一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(
Warming enhances old organic carbon decomposition through altering functional microbial communities
1
2017
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
Microbial diversity drives multifunctionality in terrestrial ecosystems
1
2016
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
The permafrost carbon inventory on the Tibetan Plateau: A new evaluation using deep sediment cores
1
2016
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
放牧对藏北紫花针茅高寒草原植物群落特征的影响
1
2010
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
放牧对藏北紫花针茅高寒草原植物群落特征的影响
1
2010
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
Carbon cycle feedbacks and future climate change
1
2015
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil
1
1996
... id="C14">为了揭示短期增温对土壤微生物群落结构的影响, 我们参照Bossio等(1998)的方法测定了土壤微生物PLFA.首先利用单相氯仿-甲醇-磷酸缓冲液提取土壤微生物PLFA, 然后将提取的磷脂通过固相萃取柱分离糖脂和中性脂, 并将极性脂甲酯化形成PLFA甲基脂.最后将样品用含有正十九酸甲酯 (19:0)作为内标的正己烷重新溶解, 用气相色谱仪(Agilent 6850, Agilent Technologies, Santa Clara, USA)进行测定.分析过程中, 依据MIDI微生物鉴别系统进行脂肪酸鉴定, 并采用以下指示脂肪酸标准: 丛枝菌根真菌(AMF): 16:1ω5c (
Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis
1
1993
... id="C14">为了揭示短期增温对土壤微生物群落结构的影响, 我们参照Bossio等(1998)的方法测定了土壤微生物PLFA.首先利用单相氯仿-甲醇-磷酸缓冲液提取土壤微生物PLFA, 然后将提取的磷脂通过固相萃取柱分离糖脂和中性脂, 并将极性脂甲酯化形成PLFA甲基脂.最后将样品用含有正十九酸甲酯 (19:0)作为内标的正己烷重新溶解, 用气相色谱仪(Agilent 6850, Agilent Technologies, Santa Clara, USA)进行测定.分析过程中, 依据MIDI微生物鉴别系统进行脂肪酸鉴定, 并采用以下指示脂肪酸标准: 丛枝菌根真菌(AMF): 16:1ω5c (
Use and misuse of PLFA measurements in soils
2
2011
... id="C32">值得注意的是, 本研究采用PLFA作为生物标记物指示微生物群落, PLFA法能够准确地反映不同类群微生物的生物量, 并且能反映微生物表现型和活性的信息(
... ), 因此该方法能够确定相对宏观的微生物群落结构特征.然而, 该标记物的检测精度有限, 很难得到微生物种属一级的精确信息(
Insights into the resistance and resilience of the soil microbial community
1
2013
... id="C29">首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定.一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(
土壤生态系统微生物多样性-稳定性关系的思考
1
2013
... id="C29">首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定.一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(
土壤生态系统微生物多样性-稳定性关系的思考
1
2013
... id="C29">首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定.一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(
The whole-soil carbon flux in response to warming
1
2017
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
Increased belowground carbon inputs and warming promote loss of soil organic carbon through complementary microbial responses
1
2014
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
2
2013
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
... id="C14">为了揭示短期增温对土壤微生物群落结构的影响, 我们参照Bossio等(1998)的方法测定了土壤微生物PLFA.首先利用单相氯仿-甲醇-磷酸缓冲液提取土壤微生物PLFA, 然后将提取的磷脂通过固相萃取柱分离糖脂和中性脂, 并将极性脂甲酯化形成PLFA甲基脂.最后将样品用含有正十九酸甲酯 (19:0)作为内标的正己烷重新溶解, 用气相色谱仪(Agilent 6850, Agilent Technologies, Santa Clara, USA)进行测定.分析过程中, 依据MIDI微生物鉴别系统进行脂肪酸鉴定, 并采用以下指示脂肪酸标准: 丛枝菌根真菌(AMF): 16:1ω5c (
1
1985
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
The contentious nature of soil organic matter
1
2015
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
土壤微生物体氮的季节性变化及其与土壤水分和温度的关系
1
2004
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
土壤微生物体氮的季节性变化及其与土壤水分和温度的关系
1
2004
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
Climatic warming in the Tibetan Plateau during recent decades
1
2000
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
Terrestrial carbon-cycle feedback to climate warming: Experimental evidence on plant regulation and impacts of biofuel feedstock harvest
1
2009
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
Microbial communities and activities in alpine and subalpine soils
1
2009
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
1
2005
... id="C14">为了揭示短期增温对土壤微生物群落结构的影响, 我们参照Bossio等(1998)的方法测定了土壤微生物PLFA.首先利用单相氯仿-甲醇-磷酸缓冲液提取土壤微生物PLFA, 然后将提取的磷脂通过固相萃取柱分离糖脂和中性脂, 并将极性脂甲酯化形成PLFA甲基脂.最后将样品用含有正十九酸甲酯 (19:0)作为内标的正己烷重新溶解, 用气相色谱仪(Agilent 6850, Agilent Technologies, Santa Clara, USA)进行测定.分析过程中, 依据MIDI微生物鉴别系统进行脂肪酸鉴定, 并采用以下指示脂肪酸标准: 丛枝菌根真菌(AMF): 16:1ω5c (
PLFA profiling of microbial community structure and seasonal shifts in soils of a douglas-fir chronosequence
1
2008
... id="C30">其次, 土壤温度和水分等环境因素会对土壤微生物群落产生不同影响, 进而抑制土壤微生物对气候变暖的响应.大量研究表明, 土壤温度与土壤含水量对微生物的生理状态、生物量及群落结构有重要影响(
Shifts in the microbial community structure explain the response of soil respiration to land-use change but not to climate warming
1
2015
... id="C14">为了揭示短期增温对土壤微生物群落结构的影响, 我们参照Bossio等(1998)的方法测定了土壤微生物PLFA.首先利用单相氯仿-甲醇-磷酸缓冲液提取土壤微生物PLFA, 然后将提取的磷脂通过固相萃取柱分离糖脂和中性脂, 并将极性脂甲酯化形成PLFA甲基脂.最后将样品用含有正十九酸甲酯 (19:0)作为内标的正己烷重新溶解, 用气相色谱仪(Agilent 6850, Agilent Technologies, Santa Clara, USA)进行测定.分析过程中, 依据MIDI微生物鉴别系统进行脂肪酸鉴定, 并采用以下指示脂肪酸标准: 丛枝菌根真菌(AMF): 16:1ω5c (
Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil
1
1999
... id="C29">首先, 土壤微生物群落自身及其与土壤、植物组成的互作体系能够在一定程度上保持稳定.一方面, 微生物群落对环境胁迫具有一定的抗性, 使微生物群落得以在气候变暖等环境变化中保持稳定(
Forest soil microbial functional patterns and response to a drought and warming event: Key role of climate-plant-soil interactions at a regional scale
2014
Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe
2
2017
... id="C7">本研究在青海省刚察县三角城种羊场开展, 详细的样地情况见
... id="C31">此外, 增温处理中土壤微生物群落特征保持相对稳定可能与增温时间较短以及增温幅度较低有关.以往的增温实验表明, 土壤微生物对增温的响应取决于增温年限.例如, Weedon等(2012)发现9年的增温实验不足以改变微生物群落, 然而15年的增温导致微生物群落发生了显著变化(
Fifteen years of climate change manipulations alter soil microbial communities in a subarctic heath ecosystem
1
2007
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
Soil microbes and their response to experimental warming over time: A meta-analysis of field studies
1
2017
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
Persistence of soil organic matter as an ecosystem property
1
2011
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
Effect of warming and drought on grassland microbial communities
1
2011
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
微生物介导的碳氮循环过程对全球气候变化的响应
1
2011
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
微生物介导的碳氮循环过程对全球气候变化的响应
1
2011
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
Carbon cycle: A warm response by soils
1
2010
... id="C12">为了揭示短期增温对土壤微生物生物量的影响, 我们基于氯仿熏蒸法(
Land-use and soil depth affect resource and microbial stoichiometry in a tropical mountain rainforest region of southern Ecuador
1
2014
... id="C12">为了揭示短期增温对土壤微生物生物量的影响, 我们基于氯仿熏蒸法(
An extraction method for measuring soil microbial biomass C
1987
土壤微生物对增温响应的Meta分析
1
2015
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
土壤微生物对增温响应的Meta分析
1
2015
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
Summer warming accelerates sub-arctic peatland nitrogen cycling without changing enzyme pools or microbial community structure
1
2012
... id="C4">当前全球正经历着快速的增温过程.自1880年以来, 全球气温已上升0.85 ℃, 预计21世纪末气温将上升1.5-4.8 ℃ (
Warm spring reduced carbon cycle impact of the 2012 US summer drought
1
2016
... id="C12">为了揭示短期增温对土壤微生物生物量的影响, 我们基于氯仿熏蒸法(
2006
2006
a). Tundra soil carbon is vulnerable to rapid microbial decomposition under climate warming
2
2016
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
... ).例如, 最近的一项研究指出, 在北极冻土区, 增温1.5年显著改变了土壤中与碳氮循环相关的微生物功能类群(
b). Annual removal of aboveground plant biomass alters soil microbial responses to warming
2
2016
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
... ), 也可能单独提高土壤细菌或真菌群落的丰度, 而不影响其他群落(
高山森林土壤微生物群落结构和功能对模拟增温的响应
1
2016
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
高山森林土壤微生物群落结构和功能对模拟增温的响应
1
2016
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
Transcriptome analysis reveals diversified adaptation of Stipa purpurea along a drought gradient on the Tibetan Plateau
1
2015
... id="C30">其次, 土壤温度和水分等环境因素会对土壤微生物群落产生不同影响, 进而抑制土壤微生物对气候变暖的响应.大量研究表明, 土壤温度与土壤含水量对微生物的生理状态、生物量及群落结构有重要影响(
Altered precipitation regime affects the function and composition of soil microbial communities on multiple time scales
2013
a). Precipitation modifies the effects of warming and nitrogen addition on soil microbial communities in northern Chinese grasslands
1
2015
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
b). A meta-analysis of the effects of experimental warming on soil carbon and nitrogen dynamics on the Tibetan Plateau
2
2015
... id="C5">国内外****已广泛开展了增温对土壤微生物影响的研究.以往研究显示, 气候变暖可能会影响土壤微生物群落结构.例如, 增温导致温带草原土壤细菌和真菌群落的生物量显著增加(
... id="C6">青藏高原位于我国西南地区, 平均海拔超过4 000 m, 是我国最大、世界最高的高原, 被称为“地球第三极”.自20世纪以来, 青藏高原地区的气温显著上升(
Microbial mediation of carbon-cycle feedbacks to climate warming
2011

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}