陈宁
1 , 2 , 张扬建
1 , 3 , 4 , 朱军涛
, , 1 , * , 李军祥
5 , 刘瑶杰
1 , 2 , 俎佳星
1 , 2 , 丛楠
1 , 黄珂
1 , 王荔
5 1 中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室, 拉萨高原生态试验站, 北京 100101 2 中国科学院大学, 北京 100190 3 中国科学院青藏高原地球科学卓越创新中心, 北京 100101 4 中国科学院大学资源与环境学院, 北京 100190 5 北京大学深圳研究生院, 深圳 518055 Nonlinear responses of productivity and diversity of alpine meadow communities to degradation CHEN Ning
1 , 2 , ZHANG Yang-Jian
1 , 3 , 4 , ZHU Jun-Tao
, , 1 , * , LI Jun-Xiang
5 , LIU Yao-Jie
1 , 2 , ZU Jia-Xing
1 , 2 , CONG Nan
1 , HUANG Ke
1 , WANG Li
5 1 Lhasa Plateau Ecosystem Research Station, Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101 China 2 University of Chinese Academy of Sciences, Beijing 100190, China 3 CAS Center for Excellence in Tibetan Plateau Earth Sciences, Beijing 100101, China 4 College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100190, China and
5 Peking University Shenzhen Graduate School, Shenzhen 518055, China 通讯作者: * 通信作者Corresponding author ( zhujt@igsnrr.ac.cn ) 基金资助: 国家重点研发计划 .2016YFC0501802 国家重点研发计划 .2017YFA0604802 国家自然科学基金 .41571195 国家自然科学基金 .41501103
Online: 2018-01-20
Fund supported: Supported by the National Key Research and Development Project of China .2016YFC0501802 Supported by the National Key Research and Development Project of China .2017YFA0604802 the National Natural Science Foundation of China .41571195 the National Natural Science Foundation of China .41501103
摘要 为了阐释青藏高原高寒草甸退化的关键生态过程, 该研究依托藏北高原草地生态系统研究站(那曲站), 设置不同退化梯度实验, 即对照、轻度退化、中度退化、重度退化和极度退化5个梯度, 探究群落生产力和物种多样性对不同退化强度的响应机制。结果表明: 1)随着退化程度不断加剧, 地上生物量呈现线性或非线性增加趋势, 在重度退化处理下, 地上生物量显著高于对照32.3%, 其中高山嵩草(Kobresia pygmaea )地上生物量呈非线性下降趋势, 而矮火绒草(Leontopodium nanum )地上生物量呈非线性增加趋势; 2)与地上生物量的响应模式相反, 随着退化程度加剧, 地下生物量与总生物量均呈现非线性降低趋势; 3)高寒草甸退化过程中, 物种辛普森指数、丰富度指数、香农多样性指数和均匀度指数均呈现非线性上升趋势。结构等式方程结果表明, 土壤碳含量和体积含水量与地下生物量均呈现显著的正相关关系。土壤碳含量、体积含水量和砾石质量比对地上生物量无显著影响, 土壤碳、氮含量与物种多样性指数呈现显著的负相关关系。研究认为地上生产力的变化不能很好地指示草地的退化程度, 建议今后研究应以可食性牧草和毒杂草等植物功能群的变化来衡量草地退化。 关键词: 高寒草甸 ;
退化梯度 ;
物种多样性 ;
生产力 ;
非线性响应 Abstract Aims Methods Important findings p < 0.05) and volume water content (p < 0.1). However, soil nutrient and soil physical properties had no significant impact on aboveground biomass (p < 0.1). Compared with soil physical properties, soil nutrition is an important factor influencing the diversity index. In our study, the nonlinear responses of productivity and diversity of alpine meadow were described by using the multiple levels of degradation in space. The results suggested that aboveground productivity cannot interpret the degree of degradation of alpine meadow, and by contrast, alpine meadow degradation should be measured by the change of plant functional groups, such as edible grasses and poisonous forbs.Keywords: alpine meadow ;
degeneration gradients ;
species diversity ;
productivity ;
nonlinear response PDF (1907KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 本文引用格式 陈宁, 张扬建, 朱军涛, 李军祥, 刘瑶杰, 俎佳星, 丛楠, 黄珂, 王荔. 高寒草甸退化过程中群落生产力和物种多样性的非线性响应机制研究.
植物生态学报 [J], 2018, 42(1): 50-65 doi:10.17521/cjpe.2017.0252
CHEN Ning.
Nonlinear responses of productivity and diversity of alpine meadow communities to degradation .
Chinese Journal of Plant Ecology [J], 2018, 42(1): 50-65 doi:10.17521/cjpe.2017.0252
高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分。据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%。特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 )。自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 )。以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 )。
近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010)。与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等。气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 )。但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 )。过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 )。过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 )。研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大。
草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 )。草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 )。即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式。
群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化。在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 )。随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 )。然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式。
综上所述, 以往的研究多关注不同退化阶段生产力和多样性的响应程度及其影响因素, 而关于群落生产力和物种多样性在退化过程中的动态响应, 即线性或者非线性响应研究较少。本研究依托中国科学院地理科学与资源研究所高原生态系统研究中心藏北高原草地生态系统研究站(那曲站), 采用熵权法划分出5个退化等级, 即对照、轻度退化、中度退化、重度退化和极度退化等, 从2016年开始, 获取两个生长季的连续群落调查和取样数据, 分析在高寒草甸退化过程中生产力和物种多样性的非线性响应, 揭示青藏高原高寒草甸退化的关键生态过程及影响因素。
1 材料和方法 1.1 研究区域概况 研究区域位于藏北高原核心地带, 那曲地区那曲县那曲镇曲果仁毛村境内的藏北高原草地生态系统研究站(那曲站)(31.64° N, 92.02° E, 海拔4 585 m), 隶属于中国科学院地理科学与资源研究所。那曲县属于高原亚寒带季风半湿润气候区。全年大风日100天左右, 平均风速2.7 m·s
-1 , 最大风速可达26.3 m·s
-1 ; 年平均气温-1.16 ℃, 全年日照时间2 788 h; 年降水量430 mm, 集中在6-9月, 占全年降水量的85% (数据来源于中国气象科学数据共享服务网那曲站点)。全年无绝对无霜期, 每年10月至次年5月为风雪期和土壤冻结期(
朱军涛, 2016 )。植被类型为典型的高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 常见伴生种有钉柱委陵菜(
Potentilla saundersiana )、无茎黄鹌菜(
Youngia simulatrix )、藏西风毛菊(
Saussurea stoliczkai )、矮羊茅(
Festuca coelestis )、楔叶委陵菜(
Potentilla cuneata )、矮火绒草(
Leontopodium nanum )等。土壤类型为高寒草甸土(
朱军涛, 2016 )。
1.2 实验设计 根据研究区高寒草甸的退化程度, 采用草地退化五级梯度标准, 即对照、轻度退化、中度退化、重度退化和极度退化。在5个退化梯度上各选取坡度<10°较为平缓的6个(即6个重复) 6 m × 6 m的典型样地(
图1 ), 共计30块实验样地。在每个样地中选取一个4 m × 4 m的核心样地, 将核心样地分割成4个2 m × 2 m的子区域(A, B, C, D), 在A区域内随机选取1个1 m × 1 m的永久样方, 用于调查植物的高度、盖度和多度; B区域用于测定地上和地下生物量; C区域用于测定碳通量; D区域用于未来的相关研究 (
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1样地布设图。A, 群落调查; B, 测定生物量; C, 测定碳通量; D, 未来的相关研究。 Fig. 1Plot layout. A, community survey; B, measure biomass; C, measure carbon flux; D, future related researches. 1.3 野外调查和取样 在2016年和2017年生长季, 对永久样方进行植物群落调查; 然后在B区域随机选取0.5 m × 0.5 m的样方分种剪草, 带至室内在烘箱烘干至恒质量后称量(65 ℃; 大约48 h); 同时, 在剪去植物地上部分后, 使用直径为7.0 cm的土钻采集0-10 cm、10-20 cm和20-30 cm的土壤样品, 为了避免土壤样品的分析误差, 所有土壤样品的采集均在晴朗的上午集中进行; 并将收集到的土壤样品使用1 mm的土壤筛将根系从土壤中分离, 过筛后的根系样品及时洗净后烘干至恒质量(65 ℃; 大约48 h)后称质量。
1.4 相关指标的测定 1.4.1 全碳(C)、全氮(N)分析
将采集到的3层土壤样品过1 mm的网筛, 自然风干、磨碎并过60目(0.27 mm)网筛。土壤全C、全N利用干烧法进行测量, 在纯氧环境下, 样品在相应试剂中燃烧或在惰性气体中高温裂解, 转化为CO
2 、N
2 , 然后通过色谱柱分离后, 分别进行热导检测, 得到C、N的质量分数, 上述分析是在元素分析仪(Elementar Analysensysteme GmbH, Hesse, Germany)中进行, 本文测量的全C、全N是表层0-10 cm土壤的C、N含量。
1.4.2 土壤体积含水量
土壤体积含水量利用TDR300土壤水分仪(Spertrum, New York, USA)测定, 分别测定7.6 cm、12 cm深处土壤含水量, 每10天测定一次。
1.4.3 土壤物理特征指标
使用容重测定后的土壤样品进行土壤砾石含量(土石比)的测定。基于0-5、5-10、10-20、20-30、30-50、50-70、70-100 cm不同土壤深度对每层容重样品烘干称质量后, 计环刀内土壤样品干质量为
m 1, 体积为
V1 。将环刀内土壤取出后放入塑料盆中, 注入清水, 用手搅拌土体, 使得砾石和土壤分离, 待砾石沉底后, 倒掉泥水。如此反复几次, 使土壤和砾石彻底分离, 用吸水纸吸干砾石表面水分。将砾石放入到孔径为2 mm的网筛内过筛。反复摇晃后, 筛内的砾石即为粒径>2 mm的砾石, 并称量其质量
m 2。选择适当大小的量筒, 注入一定体积的水, 记下水的刻度
H 1。将粒径>2 mm的砾石放入量筒内, 记下水的刻度
H 2。粒径>2 mm的砾石体积为
V 2 =
H 2-
H 1。在试验中, 需要选择适当的量筒, 如果量筒过大, 精确度不够, 量筒过小,则水容易溢出。砾石含量体积比和质量比计算公式如下:
1.5 物种多样性的计算 通过选用物种丰富度指数(
C )、辛普森指数(
D )、香农多样性指数(
H )和均匀度指数(
J ) 4类多样性指数来计算群落的物种多样性(
左小安等, 2007 ), 上述4种指数的计算均使用孔凡洲等(2012)的计算方法。
物种丰富度指数(
C ):
$C=\frac{(S-1)}{\ln N}$
辛普森指数(
D ):
$D=1-\sum\limits_{i=1}^{S}{{{P}^{2}}_{i}}$
其中,${{P}_{i}}^{2}=\frac{{{n}_{i}}({{n}_{i}}-1)}{{{N}_{i}}({{N}_{i}}-1)}$
香农多样性指数(
H ):
$H=-\sum\limits_{i=1}^{S}{{{P}_{i}}}\ln {{P}_{i}}$
均匀度指数(
J ):
$J=H/\ln S$
式中:
S 为群落中物种数;
N 为所有物种的个体数之和;
n i 为第
i 个种的个体数。
1.6 数据分析 采用单因素方差分析检验不同退化程度间的物种盖度、生物量、多样性指数等指标的差异; 使用Pearson相关分析判定多样性指数的影响因素, 以上分析在SPSS 20.0 (SPSS, Chicago, USA)中实现。在R 3.3.0中分别使用线性函数和非线性函数拟合物种盖度、生物量、多样性指数对高寒草甸退化的线性和非线性响应。本文在选取有关退化指标的基础上, 在Excel中使用熵权法评价不同退化梯度的综合得分。
在Amos 21.0中使用结构等式模型(SEM)分析生物量的直接和间接影响因素, 具体步骤如下:
1.6.1 结构方程和测量方程设立
结构等式模型假定一组隐变量之间存在相关关系, 隐变量是几个显变量的线性组合(
侯杰泰和成子娟, 1999 )。通过检验显变量之间的相关系数或协方差, 进而估计模型中的通径系数, 因此可以在统计上检验所假设的模型对所研究的过程是否合适。
结构等式模型的表达式如下:
$\eta =\beta \eta +\alpha \xi +\varsigma $ (1)
$x={{\Lambda }_{X}}\xi +\delta $ (2)
$y={{\Lambda }_{Y}}\eta +\varepsilon $ (3)
公式(1)是结构等式模型中的结构模型, 阐述隐变量与显变量之间的关系。其中,$\eta $是隐变量向量,$\beta $是隐变量$\eta $的系数矩阵, 同时也是隐变量间通径系数矩阵;$\xi $是显变量向量;
a 是显变量$\xi $的系数矩阵, 同时也是显变量对应内生潜变量的通径系数矩阵;$\varsigma $是残差向量。
公式(2)和(3)是结构等式模型中的测量模型。测量模型通常由两个方程构成, 分别表示显变量向量$\xi $和观测变量
x 之间, 及隐变量向量$\eta $和内生的观测变量
y 之间的联系。${{\Lambda }_{X}}$为观测变量
x 在显变量向量$\xi $上的因子载荷矩阵,$\delta $是公式(2)的误差向量;${{\Lambda }_{Y}}$为观测变量
y 在隐变量向量$\eta $上的因子载荷矩阵,$\varepsilon $是公式(3)的误差向量。
1.6.2 变量的选取
由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化。基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响。
1.6.3 SEM整体模型适配度评价
本文选取
χ 2 、绝对适配度指数(
RMSEA, <0.05)、增值适配度指数(
NFI, >0.9)和最小信息准则(
AIC )(理论模型的
AIC 值要小于独立模型和饱和模型的
AIC 值)衡量SEM的拟合优度(
吴明隆, 2010 )。
2 结果分析 2.1 不同退化阶段高寒草甸群落盖度 随着草地退化程度加剧, 群落盖度呈现下降趋势, 但是线性与非线性的拟合结果没有显著差异(
R 2 值一致)。其中与对照相比, 轻度退化、中度退化、重度退化与极度退化的群落盖度分别显著降低11.5% (
p < 0.01)、26.4% (
p < 0.01)、64.3% (
p < 0.01)和50.7% (
p < 0.01,
图2A )。从功能群角度看, 高山嵩草盖度变化显著, 与对照相比, 轻度退化、中度退化、重度退化和极度退化处理下的高山嵩草盖度分别显著降低24.6% (
p < 0.01)、52.8% (
p < 0.01)、89.7% (
p < 0.01)和97.7% (
p < 0.01), 说明在高寒草
甸退化过程中高山嵩草的响应较敏感(
图2B )。与高山嵩草变化趋势相反, 随着退化程度加剧, 矮火绒草盖度呈现非线性(
R 2 nonlinear >
R 2 linear )上升趋势, 其中在中度退化和极度退化中, 矮火绒草盖度分别显著增加371.5% (
p = 0.05)和656.0% (
p < 0.01,
图2D ), 而且矮火绒草盖度占总盖度的比重在极度退化处理中超过高山嵩草。委陵菜属的盖度并未随着退化程度的加剧而显著改变(
p > 0.1,
图2C )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同退化阶段高寒草甸的群落盖度。图中不同字母表示处理之间有显著差异(p < 0.1)。实线是非线性拟合结果, 虚线是线性拟合结果。Control, 对照, 无退化草甸; D1, 轻度退化草甸; D2, 中度退化草甸; D3, 重度退化草甸; D4, 极度退化草甸。 Fig. 2The community coverage of alpine meadow in different degradation stages. Different letters in figures indicate significant difference (p < 0.1). The solid lines are nonlinear fitting results. Dotted lines are linear fitting results. Control, non-degraded meadow; D1, mild degraded meadow; D2, moderate degraded meadow; D3, severe degraded meadow; D4, serious sandy meadow. 2.2 不同退化阶段高寒草甸群落生产力 随着退化程度加剧, 地上生物量呈现上升趋势, 与对照相比, 重度退化处理下地上生物量显著提高32.3% (
图3A ,
p < 0.1), 而其他退化处理地上生物量与对照相比差异不显著(
图3A ,
p > 0.1), 其中线性和非线性模型拟合趋势并没有明显差异。进一步分析不同功能群的变化, 我们发现高山嵩草地上生物量随着退化程度不断加剧, 呈现非线性或线性降低, 其过程大致可以分为两个层次, 第一个层次(对照到轻度退化), 高山嵩草地上生物量较高, 与对照相比, 在轻度退化处理下没有显著改变高山嵩草地上生物量(
图3A ,
p > 0.1); 但是在第二个层次(中度退化到极度退化), 高山嵩草地上生物量急剧下降, 其中中度退化和重度退化处理与对照相比, 高山嵩草地上生物量分别显著降低44.7% (
p < 0.01)和85.5% (
p < 0.01), 其中在极度退化处理中, 高山嵩草地上生物量为0 (
图3B )。与高山嵩草地上生物量的变化相反, 矮火绒草地上生物量随着退化程度加剧呈现非线性上升趋势(
R 2 nonlinear >
R 2 linear ), 同样可以分成两个层次, 第一个层次(对照到轻度退化)矮火绒草地上生物量较低, 对照处理下的矮火绒草地上生物量为0, 与对照处理相比, 轻度退化处理下的矮火绒草地上生物量没有显著变化(
图3D ,
p > 0.1), 但是在第二个层次上矮火绒草地上生物量迅速增加, 其中在重度退化处理中达到最大值62.5 g·m
-2 , 占总生物量的62.5% (
图3D ,
p < 0.01), 意味着从重度退化开始, 矮火绒草对地上生物量的贡献超过高山嵩草。与高山嵩草和矮火绒草地上生物量的变化相比, 委陵菜属的地上生物量并没有显著改变(
图3C ,
p > 0.1)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3不同退化阶段高寒草甸的地上生物量。图中不同字母表示处理之间有显著差异(p < 0.1)。实线是非线性拟合结果, 虚线是线性拟合结果。Control, 对照, 无退化草甸; D1, 轻度退化草甸; D2, 中度退化草甸; D3, 重度退化草甸; D4, 极度退化草甸。 Fig. 3The aboveground biomass of alpine meadow in different degradation stages. Different letters in figures indicate significant difference (p < 0.1). The solid lines are nonlinear fitting results. Dotted lines are linear fitting results. Control, non-degraded meadow; D1, mild degraded meadow; D2, moderate degraded meadow; D3, severe degraded meadow; D4, serious sandy meadow. 与地上生物量相比, 在高寒草甸退化过程中, 地下生物量的响应较地上生物量更加强烈, 呈现出显著的非线性下降趋势(
R 2 nonlinear >
R 2 linear )。其中与对照实验处理相比, 轻度退化、中度退化、重度退化和极度退化处理下的地下生物量显著降低, 分别降低24.4% (
p < 0.01), 68.1% (
p < 0.01), 87.8% (
p < 0.01)和94.4% (
p < 0.01,
图4 )。与地下生物量变化趋势一致, 在高寒草甸退化过程中, 总生物量呈现出显著的非线性下降趋势。其中与对照处理相比, 轻度退化处理降低总生物量的24.2% (
p < 0.01), 中度退化降低67.1% (
p < 0.01), 重度退化降低86.4% (
p < 0.01), 极度退化降低93.1% (
图4 ,
p < 0.01)。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4不同退化阶段高寒草甸的总生物与地下生物量。图中不同字母表示处理之间有显著差异(p < 0.1)。实线是非线性拟合结果, 虚线是线性拟合结果。Control, 对照, 无退化草甸; D1, 轻度退化草甸; D2, 中度退化草甸; D3, 重度退化草甸; D4, 极度退化草甸。 Fig. 4The total biomass and belowground biomass of alpine meadow in different degradation stages. Different letters in figures indicate significant difference (p < 0.1). The solid lines are nonlinear fitting results. Dotted lines are linear fitting results. Control, non-degraded meadow; D1, mild degraded meadow; D2, moderate degraded meadow; D3, severe degraded meadow; D4, serious sandy meadow. 2.3 不同退化阶段高寒草甸物种多样性特征 随着退化程度不断加深, 辛普森指数、丰富度指数、香农多样性指数和均匀度指数均呈现出显著的非线性上升趋势(
R 2 nonlinear >
R 2 linear ), 并且物种多样性测度结果表明, 轻度、中度、重度和极度退化草甸的物种辛普森指数(
p < 0.05)、丰富度指数(
p < 0.05)、香农多样性指数(
p < 0.01)和均匀度指数(
p < 0.05)均显著高于对照处理(
图5 )。四个不同退化阶段的高寒草甸物种多样性指数的变化趋势大致可以分为两个层次: 第一个层次具有较低的物种丰富度、 均匀度和多样性, 这一层次主要是以高山嵩草为优势物种的轻度和中度退化草甸群落; 第二个层次具有较高的物种丰富度、均匀度和多样性, 是逐渐以矮火绒草为优势种的重度和极度退化群落(
图5 )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5不同退化阶段高寒草甸的生物多样性。图中不同字母表示处理之间有显著差异(p < 0.1)。实线是非线性拟合结果, 虚线是线性拟合结果。Control, 对照, 无退化草甸; D1, 轻度退化草甸; D2, 中度退化草甸; D3, 重度退化草甸; D4, 极度退化草甸。 Fig. 5The species diversity of alpine meadow in different degradation stages. Different letters in figures indicate significant difference (p < 0.1). The solid lines are nonlinear fitting results. Dotted lines are linear fitting results. Control, non-degraded meadow; D1, mild degraded meadow; D2, moderate degraded meadow; D3, severe degraded meadow; D4, serious sandy meadow. 2.4 不同退化阶段高寒草甸土壤理化性质变化特征 随着退化程度不断加剧, 土壤C、N含量呈现出显著的非线性下降趋势(
R 2 nonlinear >
R 2 linear ,
表2 ), 并且变化趋势较为一致, 从轻度退化到极度退化可以清晰地划分为两个阶段, 第一阶段(对照到中度退化)中不同退化程度的高寒草甸土壤N和C含量并没有发生显著改变(
表1 ,
p > 0.1); 第二阶段中重度退化和极度退化与对照, 轻度退化和中度退化相比均显著降低土壤中的N和C含量, 与对照处理相比, N含量分别降低28.5% (
p < 0.05)和23.6% (
p < 0.1), C含量分别降低39.2% (
p < 0.01)和32.5% (
p < 0.01) (
表1 )。
Table 1 表1 表1 不同退化阶段高寒草甸的土壤理化特征
Table 1
The soil physicochemical properties of alpine meadow in different degradation stages 草甸类型 土壤容重-3 ) 砾石质量比 砾石体积比 土壤氮含量 土壤碳含量 对照 Non-degraded 1.23 ± 0.04a 29.77 ± 7.96a 16.80 ± 7.45a 0.45 ± 0.06a 5.29 ± 0.95a 轻度退化 Mild degraded 1.26 ± 0.04a 36.51 ± 1.34a 22.19 ± 4.00a 0.50 ± 0.04a 5.76 ± 0.59a 中度退化 Moderate degraded 1.50 ± 0.02b 37.43 ± 6.04a 28.24 ± 2.60ab 0.44 ± 0.11a 5.10 ± 1.30a 重度退化 Severe degraded 1.47 ± 0.02b 47.60 ± 4.52b 32.15 ± 2.41b 0.32 ± 0.05b 3.21 ± 0.69b 极度退化 Serious degraded 1.56 ± 0.11b 49.62 ± 4.94b 33.52 ± 0.68b 0.34 ± 0.02b 3.57 ± 0.23b
Different letters indicate significant difference (p < 0.1). 不同字母表示处理之间有显著差异(p < 0.1)。 新窗口打开 |
下载CSV Table 2 表2 表2 高寒草甸退化过程中土壤理化性质线性和非线性响应比较
Table 2
Comparison between linear and nonlinear responses of soil physicochemical properties to alpine meadow degradation R 2 拟合模型 Fitted model 土壤容重 Soil bulk density 线性模型 Linear model 0.66*** y = -0.006x + 1.695非线性模型 Nonlinear model 0.76*** y = 0.0002x 2 + 0.012x + 1.326砾石质量比 Mass ratio of gravel 线性模型 Linear model 0.13 y = -0.155x + 47.310非线性模型 Nonlinear model 0.53* y = 0.023x 2 -2.326x + 89.838砾石体积比 Volume ratio of gravel 线性模型 Linear model 0.44** y = 0.240x + 37.607非线性模型 Nonlinear model 0.58** y = 0.011x 2 -1.300x + 58.371土壤氮含量 Soil nitrogen content 线性模型 Linear model 0.42*** y = 0.003x + 0.264非线性模型 Nonlinear model 0.46*** y = -0.0001x 2 + 0.011x + 0.115土壤碳含量 Soil carbon content 线性模型 Linear model 0.50*** y = 0.049x + 2.35非线性模型 Nonlinear model 0.55*** y = -0.001x 2 + 0.159x + 0.183
*, **, *** represent statistically significant at the 10%, 5%, 1% confidence level, respectively. *、**、***分别表示在10%、5%、1%的置信水平上具有差异显著性。 新窗口打开 |
下载CSV 我们的研究发现, 随着退化程度加剧, 土壤容重、砾石质量比、砾石体积比呈现出显著的非线性上升趋势(
R 2 nonlinear >
R 2 linear ), 其中只有非线性模型才能拟合砾石质量比对高寒草甸退化的响应(
表2 )。土壤的物理特征表明, 土壤容重、砾石质量比和砾石体积比整体呈现上升趋势, 并且变化趋势较为一致, 从轻度退化到极度退化可以大致划分为两个阶段, 其中土壤容重在第一阶段(对照到轻度退化), 没有随着退化程度的加剧而显著改变。 在第二阶段(中度退化到极度退化), 与对照和轻度退化相比,中度退化、重度退化和极度退化均显著增加土壤容重, 其中与对照相比, 分别显著增加土壤容重20.6% (
p < 0.05)、19.1% (
p < 0.05)和26.4% (
p < 0.05) (
表1 )。与土壤容重不同, 土壤砾石质量比和砾石体积比的第一阶段是从对照到中度退化处理, 其中在第一阶段土壤砾石质量比和体积比没有显著改变; 但是在第二阶段(重度退化到极度退化), 与对照相比, 重度退化和极度退化土壤砾石质量比分别显著增加59.9% (
p < 0.05)和66.7% (
p < 0.05), 显著增加土壤砾石体积比91.4% (
p < 0.01)和99.6% (
p < 0.01) (
表1 )。
在高寒草甸退化过程中, 土壤中0-7.6 cm和0-12 cm两个层次中土壤含水量并没有呈现出明显的非线性响应趋势(
表3 )。不同退化阶段高寒草甸土壤含水量特征表明, 土壤中0-7.6 cm和0-12 cm两个层次中土壤含水量呈现下降趋势, 变化趋势一致, 从轻度退化到极度退化可以清晰地划分为两个阶段, 在第一阶段(对照到中度退化), 不同退化程度的高寒草甸土壤中0-7.6 cm和0-12 cm含水量并没有发生显著改变(
图6 ,
p > 0.1); 但是与第一阶段相比, 第二阶段中重度退化和极度退化与对照、轻度退化和中度退化相比均显著降低土壤中0-7.6 cm和0-12 cm含水量, 与对照相比, 土壤中0-7.6 cm水分含量分别显著降低了19.6% (
p < 0.01)和15.5% (
p < 0.01), 土壤中0-12 cm水分含量分别显著降低了21.9% (
p < 0.01)和17.0% (
p < 0.05) (
图6 )。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6不同退化阶段高寒草甸的土壤含水量特征(平均值±标准误差)。不同字母表示处理之间有显著差异(p < 0.1)。Control, 对照, 无退化草甸; D1, 轻度退化草甸; D2, 中度退化草甸; D3, 重度退化草甸; D4, 极度退化草甸。 Fig. 6The soil water content of alpine meadow in different degradation stages (mean ± SE ). Different letters indicate significant difference (p < 0.1). Control, non-degraded meadow; D1, mild degraded meadow; D2, moderate degraded meadow; D3, severe degraded meadow; D4, serious sandy meadow. Table 3 表3 表3 高寒草甸退化过程中土壤含水量的线性和非线性响应比较
Table 3
Comparison between linear and nonlinear responses of soil water content to alpine meadow degradation R 2 拟合模型 Fitted model 0-7.6 cm土层 0-7.6 cm soil layer 线性模型 Linear model 0.61*** y = 0.044x + 7.396非线性模型 Nonlinear model 0.61*** y = 7.557e0.005x 0-12 cm土层 0-12 cm soil layer 线性模型 Linear model 0.39*** y = 0.066x + 10.148非线性模型 Nonlinear model 0.41*** y = -0.001x 2 + 0.169x + 8.136
*, **, *** represent statistically significant at the 10%, 5%, 1% confidence level, respectively. *、**、***分别表示在10%、5%、1%的置信水平上具有差异显著性。 新窗口打开 |
下载CSV 2.5 物种多样性、群落生产力与环境因子的关系 相关性分析结果表明, 土壤C和N含量与均匀度指数、辛普森指数及香农多样性指数存在显著负相关关系(
p < 0.05), 但是与砾石质量比、砾石体积比及体积含水量无显著相关关系(
p > 0.1,
图7 ), 其中物种丰富度指数与土壤C、N和土壤物理性质均没有显著相关关系(
p > 0.1)。说明在高寒草甸退过程中, 物种的多样性主要受到土壤营养成分的影响。
图7 新窗口打开 |
下载原图ZIP |
生成PPT 图7不同退化阶段高寒草甸群落物种多样性与土壤理化性质之间相关分析。黑色和从左下指向右上的斜杠表示两个变量正相关, 黑色和从左上指向右下的斜杠表示变量之间负相关。颜色越深, 饱和度越高, 说明变量相关性越大; *、**、***分别表示在10%、5%、1%的置信水平上具有统计显著性。 Fig. 7Correlation analysis among soil physico-chemical properties and species diversity of alpine meadow community. Black and from lower left to upper right of the slash said two variables are related, and black from the upper left to lower right slash said negative correlation between variables. The deeper the color, the higher the degree of saturation, explanatory variable correlation; *, **, *** represent statistically significant at the 10%, 5%, 1% confidence level, respectively. SEM结果表明土壤C含量对高山嵩草盖度、矮火绒草盖度和群落总盖度均有显著的直接影响, 之间的通径系数分别为0.71 (
p < 0.01), -0.29 (
p < 0.1)和0.16 (
p < 0.05), 矮火绒草盖度和高山嵩草盖度对群落总盖度存在显著的直接影响, 通径系数为0.25 (
p < 0.01)和0.88 (
p < 0.01), 体积含水量和群落总盖度对地下生物量有显著的直接影响, 通径系数为0.14 (
p < 0.1)和0.99 (
p < 0.01)。碎石质量比和体积含水量之间的通径系数为-0.44 (
p < 0.01)), 因此土壤C含量和碎石质量比对地下生物量有间接的影响, 间接通径系数为0.76 (
p < 0.05)和0.07 (
p > 0.1)。地下生物量主要受高山嵩草盖度和体积含水量直接影响, 土壤C含量通过影响高山嵩草盖度间接影响地下生物量, 但是土壤理化性质对地上生物量没有显著的直接或间接影响。
图8 新窗口打开 |
下载原图ZIP |
生成PPT 图8影响地上生物量与地下生物量的直接因素和间接因素的结构等式模型。带有箭头的虚线表示关系不显著(p > 0.1), 实线表示关系显著(p < 0.1), 带有箭头线上的数字是标准化路径系数(*, p < 0.1; **, p < 0.05; ***, p < 0.01)。AIC, 最小信息准则; AIC dm , 内定模式; AIC im , 独立模式; AIC sm , 饱和模式; DF, 自由度; NFI, 增值适配度指数; RMSEA, 绝对适配指数。 Fig. 8The results of structural equation models showed the direct and indirect impacts of various factors on biomass and species diversity. A dotted line relationship of the arrow was not significant (p > 0.1), the solid line shows the relationship between significantly (p < 0.1), the degree of thickness line to reflect the degree of a strong relation between variables, with the arrow line Numbers are standardized path coefficient (*, p < 0.1; **, p < 0.05; ***, p < 0.01). AIC, an information criterion; AIC dm , default mode; AIC im , independence model; AIC sm , saturated model; DF, degree of freedom; NFI, normed fit index; RMSEA, root mean square error of approximation. 3 讨论 本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应。研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 )。
3.1 高寒草甸退化过程中群落生产力的非线性响应 我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖。由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能。在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 )。我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加。但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感。
与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致。在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低。与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降。研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势。随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低。由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响。
3.2 高寒草甸退化过程中物种多样性的非线性响应 物种多样性不仅能够度量群落的组成结构和功能的复杂性, 而且也能指示环境状况, 例如土壤养分等(
罗亚勇等, 2014 )。研究结果表明, 在高寒草甸退化过程中群落多样性呈现出非线性增加趋势, 该结果与仁青吉等(2008)的研究结果一致。研究进一步发现, 均匀度指数和多样性指数在重度退化梯度上达到最大, 物种丰富度指数在极度退化梯度上达到最大, 与之前的研究结果(
柳小妮等, 2008 )相似。造成这种差异的主要原因是群落种类数目和种类之间个体分配的均匀性, 过度放牧抑制了优势物种的竞争能力, 导致处于竞争弱势物种的入侵和定居成为可能, 在重度、极度退化阶段物种数增幅较大, 因此物种丰富度显著增加。同时在重度和极度退化阶段, 优势种对其他物种的抑制作用明显减弱, 表现为优势种高山嵩草盖度逐渐降低, 矮火绒草盖度逐渐增加, 因此在重度退化梯度上, 高山嵩草、委陵菜属和矮火绒草在群落中达到相对平衡, 导致均匀度在重度退化阶段最高。
随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化。植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型。这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 )。我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致。由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应。
4 结论 青藏高原的主要草地类型——高寒草甸, 近年来发生了不同程度的退化, 严重威胁高原的畜牧业可持续发展和生态环境。本研究利用空间上的多退化梯度, 采用熵权法为草地退化等级进行评分, 研究群落生产力和多样性在退化过程中的动态响应。我们发现随着高寒草甸退化程度加剧, 地上生物量和群落多样性指数非线性增加, 而地下生物量和总生物量非线性降低。进一步分析我们发现, 草甸退化过程中植物功能群组成发生变化, 例如莎草科植物减少, 矮火绒草等杂类草类增加, 改变了地上和地下生物量的组成, 进而导致地上和地下生物量的不同响应。群落多样性变化的原因比较复杂, 一方面可能是放牧动物的采食和践踏, 引起群落光资源的释放; 另一方面可能是土壤退化过程中, 一些耐瘠薄和耐干旱的物种入侵, 从而提高了群落多样性。本研究刻画了高寒草甸群落生产力和物种多样性的非线性响应模式, 揭示了青藏高原高寒草甸退化的关键生态过程及影响因素。
附件 附件I 基于熵权法的综合评价 Appendix I Comprehensive evaluation based on Entropy Method 1.1 土地退化评价指标的确定 依据邱秋兰(2012)的文献检索结果, 对土地退化进行评价的指标共计83个, 大致可以分为植被、气候、土壤、社会经济、地形地貌及生物6类, 其中土壤类指标较多, 而且植被与社会经济指标所占比例也不少(
附件表1 ;邱秋兰, 2012)。
Appendix table 1 附件表1 附件表1 土地退化中文指标分类(邱秋兰, 2012)
Appendix table 1
Land degradation index classification in China (Qiu, 2012) 指标分类 指标个数 指标 Indexes 土壤 Soil 41 土壤质地、土层厚度、含水量、团粒结构稳定性等 Soil texture, soil thickness, water content, and stability of aggregates, etc. 植被 Vegetation 11 植被盖度、植被种类、植被均匀度指数、根系深度等 Vegetation cover, vegetation species, vegetation Pielou evenness index, and root depth, etc. 社会经济 Social economy 11 经济收入水平(农民收入、消费水平)、人口数量等 Economic income level (farmer's income, consumption level) and population, etc. 气候 Climate 7 降水量、蒸发量、风速、平均气温、湿润指数等 Precipitation, evaporation, wind speed, average temperature, and wetting index, etc. 生物 Biology 7 生物量、生物多样性指数、群落类型、种群优势度 Biomass, biodiversity index, community type, and population dominance 地形地貌 Topography 6 坡度、坡向、沟谷密度、地貌类型、海拔高度 Slope, slope direction, valley density, landform type, and altitude
新窗口打开 |
下载CSV 其中频次分析结果表明: 应用频次最高的4个指标分别为植被盖度、有机质含量、经济收入水平及坡度, 频次分别为116、48、43和42 (附件
表2 ; 邱秋兰, 2012)。在以上分析基础上, 参考以往相关研究(
周华坤等, 2003 ;
毛飞等, 2008 ), 并结合本研究的数据获取情况, 本文在评价藏北高寒草甸退化过程中选取的评价指标为: 群落总盖度、高山嵩草盖度、土壤碳氮比、地上生物量、地下生物量及土壤砾石比等6个指标。
Appendix table 2 附件表2 附件表2 土地退化中文指标应用频次统计(邱秋兰, 2012)
Appendix table 2
Land degradation index of application frequency statistics (Qiu, 2012) 频次 指标个数 指标 Indexes > 40 41 植被盖度、经济收入水平、坡度、有机质含量 Vegetation cover, economic income level, slope, organic matter content 30-39 11 土壤质地、氮、磷和钾元素含量、含水量、土地利用类型 Soil texture, nitrogen, phosphorus and potassium content, water content, land use type 20-29 11 生产力、降水量、生物多样性、生物量、地貌类型 Productivity, precipitation, biodiversity, biomass, landform type 10-19 7 土壤类型、覆沙盖度、土层厚度、风速、蒸发量 Soil type, cover sand cover, soil thickness, wind speed, evaporation < 10 7 土壤容重、土壤侵蚀模数、电导率、径流量、结皮厚度 Soil bulk density, soil erosion modulus, conductivity, runoff, crust thickness
新窗口打开 |
下载CSV 1.2 综合评价 熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法。值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003)。本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降。
参考文献 Qiu QL (2012).
Assessment of Degraded Land in the Heihe River Basin, Arid Northwestern China . Master degree dissertation, Shandong Normal University, Jinan. [邱秋兰 (2012). 西北干旱区黑河流域土地退化评价研究, 硕士学位论文, 山东师范大学, 济南.]
Shen WS, Cao XZ, Shen FY (2006). Classification and grading of land degradation in China.
Journal of Ecology and Rural Environment, 22(4), 88-93. [沈渭寿, 曹学章, 沈发云 (2006). 中国土地退化的分类与分级. 生态与农村环境学报, 22(4), 88-93.]
Xiao XY (2003). S
tudy on the Appraisal of Enterprise Competitiveness . Master degree dissertation, Jiangxi University of Finance and Economics, Nanchang, Jiangxi. [肖香玉 (2003). 企业竞争力评价研究. 硕士学位论文, 江西财经大学, 江西南昌.]
Zhou HK, Zhou L, Zhao XQ, Liu W, Yan ZL, Shi Y (2003). Degradation process and integrated treatment of “black soil beach” grassland in the source regions of Yangtze and Yellow Rivers.
Chinese Journal of Ecology, 22(5), 51-55. [周华坤, 周立, 赵新全, 刘伟, 严作良, 师燕 (2003). 江河源区“黑土滩”型退化草场的形成过程与综合治理. 生态学杂志, 22(5), 51-55.]
致谢 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。 [1] Bai YF Li LH Wang QB Zhang LX Zhang Y Chen ZZ 2000 ). Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River Basin, Inner MongoliaActa Phytoecologica Sinica, 24, 667 -673 . DOI:
10.1088/0256-307X/17/9/008 URL 用样带法研究了草原群落植物多样性和初级生产力沿海拔和水分梯度的变化,结果表明:在样带梯度上,物种丰富度、多样性和群落初级生产力与海拔高度、年降水量和土壤有机C及全N含量呈正相关,而与年平均气温和干燥度呈负相关。随着海拔高度的降低,降水量的减少,热量和干燥度的增加,以及土壤有机C和全N含量的降低,草原群落的物种丰富度、多样性和初级生产力逐渐降低。典型相关分析的结果揭示出,土壤有机C含量和干燥度是对草
[ 白永飞 , 李凌浩 , 王其兵 , 张丽霞 , 张焱 , 陈佐忠 ( 2000 ). 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究植物生态学报 , 24, 667 -673 .] DOI:
10.1088/0256-307X/17/9/008 URL 用样带法研究了草原群落植物多样性和初级生产力沿海拔和水分梯度的变化,结果表明:在样带梯度上,物种丰富度、多样性和群落初级生产力与海拔高度、年降水量和土壤有机C及全N含量呈正相关,而与年平均气温和干燥度呈负相关。随着海拔高度的降低,降水量的减少,热量和干燥度的增加,以及土壤有机C和全N含量的降低,草原群落的物种丰富度、多样性和初级生产力逐渐降低。典型相关分析的结果揭示出,土壤有机C含量和干燥度是对草
[2] Cai XB Zhang YQ Shao W 2007 ). Degradation and mechanism of grassland of North Tibet alpine prairieSoil, 39, 855 -858 . DOI:
10.3321/j.issn:0253-9829.2007.06.002 URL [本文引用: 2] 藏北高寒草原是西藏畜牧业发展的核心区域.近年来,日趋严重的草地退化已从整体上影响并制约 着区域经济的可持续发展.藏北高原自然环境严酷、生态系统脆弱,在全球变化的大背景下,随着人类干预强度和频度的不断提高,草地生态系统的退化正呈进一步 加重的趋势.在阐述藏北高原草地退化现状的基础上,就藏北高原草地生态系统的退化成因进行了深入分析.
[ 蔡晓布 , 张永青 , 邵伟 ( 2007 ). 藏北高寒草原草地退化及其驱动力分析土壤 , 39, 855 -858 .] DOI:
10.3321/j.issn:0253-9829.2007.06.002 URL [本文引用: 2] 藏北高寒草原是西藏畜牧业发展的核心区域.近年来,日趋严重的草地退化已从整体上影响并制约 着区域经济的可持续发展.藏北高原自然环境严酷、生态系统脆弱,在全球变化的大背景下,随着人类干预强度和频度的不断提高,草地生态系统的退化正呈进一步 加重的趋势.在阐述藏北高原草地退化现状的基础上,就藏北高原草地生态系统的退化成因进行了深入分析.
[3] Chen LL Shi JJ Wang YL Ma YS Dong QM Hou XK 2016 ). Study on different degraded degrees grassland community structure characteristics of the alpine areaActa Agrectir Sinica, 24, 210 -213 . DOI:
10.11733/j.issn.1007-0435.2016.01.029 URL [本文引用: 1] 在玉树市巴塘乡铁力角村冬春草地上对不同退化程度高寒草地植物群落、地上及地下生物量进行调查分析。结果表明,该地区草地植被由16科33属42种植物组成;随着草地退化程度的加剧,草地优势种由丝颖针茅(Stipa capillacea)向铁棒锤(Aconitum pendulum)过渡;草地总盖度、总生物量及优良牧草生物量均呈降低趋势;杂类草生物量显著增加(P〈0.05);草地质量和生产力明显降低。
[ 陈乐乐 , 施建军 , 王彦龙 , 马玉寿 , 董全民 , 侯宪宽 ( 2016 ). 高寒地区不同退化程度草地群落结构特征研究草地学报 , 24, 210 -213 .] DOI:
10.11733/j.issn.1007-0435.2016.01.029 URL [本文引用: 1] 在玉树市巴塘乡铁力角村冬春草地上对不同退化程度高寒草地植物群落、地上及地下生物量进行调查分析。结果表明,该地区草地植被由16科33属42种植物组成;随着草地退化程度的加剧,草地优势种由丝颖针茅(Stipa capillacea)向铁棒锤(Aconitum pendulum)过渡;草地总盖度、总生物量及优良牧草生物量均呈降低趋势;杂类草生物量显著增加(P〈0.05);草地质量和生产力明显降低。
[4] Dong XB Zhang XS 2005 ). View on the grassland in ChinaGreen Economy, ( 10 ), 70 -73 . [本文引用: 1] [ 董孝斌 , 张新时 ( 2005 ). 我国草地的发展观生态经济 , ( 10 ), 70 -73 .] [本文引用: 1] [5] Gao YZ Han XG Wang SP 2004 ). The effects of grazing on grassland soilsActa Ecologica Sinica, 24, 790 -797 . DOI:
10.3321/j.issn:1000-0933.2004.04.021 URL [本文引用: 1] 介绍了放牧对草原土壤物理性质(容重、渗透率)、化学性质(有机质、N素)和微生物的影响.由于草原土壤系统本身的复杂性、滞后性和弹性,放牧对土壤性质的影响不尽相同.一般而言,随放牧强度的增大,动物践踏作用的增强,土壤孔隙分布的空间格局发生变化,土壤的总孔隙减少,特别是大孔隙(>50μm)和较大中等孔隙(9~50μm)减少,使土壤容重增加,土壤的渗透阻力加大,土壤的保水和持水能力下降.但在有机质含量很低的沙质土壤中,超载过牧,造成有机质含量降低,土壤的团粒结构减少,稳定性团聚体减少,土壤结构遭到破坏,使得土壤容重反而降低.土壤有机质和放牧之间存在复杂的相互关系,土壤有机质对放牧的响应受多种因素的影响,这些因素包括植被和土壤的初始状况;环境因素,特别是水分和温度;放牧历史(强度、频率、持续时间和动物类型).同时,土壤有机质含量低的土壤比含量高的土壤更易受放牧的影响,而使有机质发生变化.土壤微生物量碳是最具活性的土壤碳库,对环境的变化敏感,能较早地指示生态系统功能的变化.当考虑时间尺度时,高强度放牧对土壤肥力有负面的影响,短期内,由于加速了养分的循环效率,产生有利的影响,但长期无管理的超载放牧必然造成系统物质(资源)输入和输出的不平衡,最终导致草原生态系统退化,特别是在相对脆弱的干旱和半干旱生态区.为了保护草原的土壤资源,应改变只注重草原的经济功能(获取各种畜产品)的观念,而更应注重发挥草原的生态功能和社会功能,使草原得到修养生息.
[ 高英志 , 韩兴国 , 汪诗平 ( 2004 ). 放牧对草原土壤的影响生态学报 , 24, 790 -797 .] DOI:
10.3321/j.issn:1000-0933.2004.04.021 URL [本文引用: 1] 介绍了放牧对草原土壤物理性质(容重、渗透率)、化学性质(有机质、N素)和微生物的影响.由于草原土壤系统本身的复杂性、滞后性和弹性,放牧对土壤性质的影响不尽相同.一般而言,随放牧强度的增大,动物践踏作用的增强,土壤孔隙分布的空间格局发生变化,土壤的总孔隙减少,特别是大孔隙(>50μm)和较大中等孔隙(9~50μm)减少,使土壤容重增加,土壤的渗透阻力加大,土壤的保水和持水能力下降.但在有机质含量很低的沙质土壤中,超载过牧,造成有机质含量降低,土壤的团粒结构减少,稳定性团聚体减少,土壤结构遭到破坏,使得土壤容重反而降低.土壤有机质和放牧之间存在复杂的相互关系,土壤有机质对放牧的响应受多种因素的影响,这些因素包括植被和土壤的初始状况;环境因素,特别是水分和温度;放牧历史(强度、频率、持续时间和动物类型).同时,土壤有机质含量低的土壤比含量高的土壤更易受放牧的影响,而使有机质发生变化.土壤微生物量碳是最具活性的土壤碳库,对环境的变化敏感,能较早地指示生态系统功能的变化.当考虑时间尺度时,高强度放牧对土壤肥力有负面的影响,短期内,由于加速了养分的循环效率,产生有利的影响,但长期无管理的超载放牧必然造成系统物质(资源)输入和输出的不平衡,最终导致草原生态系统退化,特别是在相对脆弱的干旱和半干旱生态区.为了保护草原的土壤资源,应改变只注重草原的经济功能(获取各种畜产品)的观念,而更应注重发挥草原的生态功能和社会功能,使草原得到修养生息.
[6] Gu YR Zhang GF Meng L 2005 ). Drought resistance of four forage species at seedling under water stressJournal of Sichuan Grassland, ( 4 ), 4 -7 . [本文引用: 1] [ 谷燕蓉 , 张国芳 , 孟林 ( 2005 ). 四种牧草幼苗对水分胁迫的相应及其抗旱性四川草原 , ( 4 ), 4 -7 .] [本文引用: 1] [7] He JS Fang JY Ma KP Huang JH 2003 ). Biodiversity and ecosystem productivity: Why is there a discrepancy in the relationship between experimental and natural ecosystems?Acta Phytoecologica Sinica, 27, 835 -843 . [本文引用: 1] [ 贺金生 , 方精云 , 马克平 , 黄建辉 ( 2003 ). 生物多样性与生态系统生产力: 为什么野外观测和受控实验结果不一致?植物生态学报 , 27, 835 -843 .] [本文引用: 1] [8] Hou JT Cheng ZJ 1999 ). Application and analytical strategies of structure equation modellingExploration of Psychology, 19, 54 -59 . DOI:
10.3969/j.issn.1003-5184.1999.01.010 URL [本文引用: 1] 差不多所有心理、教育、社会等概念,均难以直接准确测量,结构方程(SEM,Struc-tulalEquationModelling)提供一个处理测量误差的方法,采用多个指标去反映潜在变量,也令估计整个模型因子间关系,较传统回归方法更为准确合理。本文主要用一系列有关学习动机的虚拟例子,指出每个问题的主要分析策略,以展示SEM在教育及心理学可以应用的研究范畴。文内探讨的方法包括:验证性因素、高阶因子、路径及因果分析、多时段(multiwave)设计、单形模型(SimpleModel)、及多组比较等。
[ 侯杰泰 , 成子娟 ( 1999 ). 结构方程模型的应用及分析策略心理学探新 , 19, 54 -59 .] DOI:
10.3969/j.issn.1003-5184.1999.01.010 URL [本文引用: 1] 差不多所有心理、教育、社会等概念,均难以直接准确测量,结构方程(SEM,Struc-tulalEquationModelling)提供一个处理测量误差的方法,采用多个指标去反映潜在变量,也令估计整个模型因子间关系,较传统回归方法更为准确合理。本文主要用一系列有关学习动机的虚拟例子,指出每个问题的主要分析策略,以展示SEM在教育及心理学可以应用的研究范畴。文内探讨的方法包括:验证性因素、高阶因子、路径及因果分析、多时段(multiwave)设计、单形模型(SimpleModel)、及多组比较等。
[9] Kong FZ Yu RC Xu ZJ Zhou MJ 2012 ). Application of excel in calculation of biodiversity indicesMarine Sciences, 36(4 ), 57 -62 . Magsci 生物多样性指数是生物群落研究中的重要参数, 广泛应用于海洋生态系统生物多样性的评价和研究。作者利用微软的电子表格软件Excel 中的数组函数、条件函数及其他函数, 提供了一种快速、简便、可靠的同时计算Shannon、Simpson、Berger-Parker、Margalef 和Pielou 均匀度指数等8 种常用生物多样性指数的方法, 应用开发的Excel 文件——Biodixcel.xlsx, 使用者只需在此文件列出的表格中一次性录入所要计算的原始数据即可在此表格中同时得到上述几个指数的计算结果, 本方法为群落生物多样性的计算提供了一种简单、实用的工具。
[ 孔凡洲 , 于仁成 , 徐子钧 , 周名江 ( 2012 ). 应用Excel软件计算生物多样性指数海洋科学 , 36(4 ), 57 -62 .] Magsci 生物多样性指数是生物群落研究中的重要参数, 广泛应用于海洋生态系统生物多样性的评价和研究。作者利用微软的电子表格软件Excel 中的数组函数、条件函数及其他函数, 提供了一种快速、简便、可靠的同时计算Shannon、Simpson、Berger-Parker、Margalef 和Pielou 均匀度指数等8 种常用生物多样性指数的方法, 应用开发的Excel 文件——Biodixcel.xlsx, 使用者只需在此文件列出的表格中一次性录入所要计算的原始数据即可在此表格中同时得到上述几个指数的计算结果, 本方法为群落生物多样性的计算提供了一种简单、实用的工具。
[10] Li L Liu W 2011 ). Relationship between plant functional groups, species richness and community’s productivity in degraded grasslandActa Agrectir Sinica, 19, 917 -921 . DOI:
10.11733/j.issn.1007-0435.2011.06.005 URL [本文引用: 2] 为了探讨草地退化后功能群的变化情况,采用群落调查和空间代替时 间的方法,在青海省达日地区对不同退化程度的矮嵩草(Kobresia humilis)草甸物种丰富度和生产力之间的关系进行了分析.结果表明:物种丰富度和物种多样性指数在中度退化草地最高,轻度退化草地次之,重度退化草 地最低;群落生产力则在轻度退化草地最高,中度退化草地次之,重度退化草地最低,且与物种总数和多样性指数无显著相关性.不同植物功能群的变化不尽相同, 随着退化程度的加剧杂类草生物量增加、豆科和莎草科生物量减少、禾本科生物量呈现“低-高-低”的变化趋势.相关分析结果表明,功能群生物量与功能群内植 物种数不存在明显的相关关系.
[ 李里 , 刘伟 ( 2011 ). 退化草地植物功能群和物种丰富度与群落生产力关系的研究草地学报 , 19, 917 -921 .] DOI:
10.11733/j.issn.1007-0435.2011.06.005 URL [本文引用: 2] 为了探讨草地退化后功能群的变化情况,采用群落调查和空间代替时 间的方法,在青海省达日地区对不同退化程度的矮嵩草(Kobresia humilis)草甸物种丰富度和生产力之间的关系进行了分析.结果表明:物种丰富度和物种多样性指数在中度退化草地最高,轻度退化草地次之,重度退化草 地最低;群落生产力则在轻度退化草地最高,中度退化草地次之,重度退化草地最低,且与物种总数和多样性指数无显著相关性.不同植物功能群的变化不尽相同, 随着退化程度的加剧杂类草生物量增加、豆科和莎草科生物量减少、禾本科生物量呈现“低-高-低”的变化趋势.相关分析结果表明,功能群生物量与功能群内植 物种数不存在明显的相关关系.
[11] Li MS 2000 ). Countermeasures for environmental protection in the Qinghai-Tibet PlateauResources Science, 22(4 ), 78 -82 . DOI:
10.3321/j.issn:1007-7588.2000.04.012 URL Magsci [本文引用: 1] 青藏高原地域辽阔,地势高亢,生态环境独特而又脆弱。严酷的生存环境使其人口稀少,经济活动对环境的影响也较弱,故高原原始自然面貌基本完好。但是最近数十年间,局部地区的水、土、生物与矿产等资源的过度开发与不合理利用,不仅使部分资源质量下降,也导致局部地区环境趋于恶化,诸如草场退化、森林减少、荒漠化土地增大以及一些主要城乡的三废污染日益严重等。所以,适当控制人口、适度开发与合理利用资源、加强环境整治与自然保护区建设等乃是协调人地关系,保护高原资源与环境,促使高原地区经济可持续发展的基本对策。
[ 李明森 ( 2000 ). 青藏高原环境保护对策资源科学 , 22(4 ), 78 -82 .] DOI:
10.3321/j.issn:1007-7588.2000.04.012 URL Magsci [本文引用: 1] 青藏高原地域辽阔,地势高亢,生态环境独特而又脆弱。严酷的生存环境使其人口稀少,经济活动对环境的影响也较弱,故高原原始自然面貌基本完好。但是最近数十年间,局部地区的水、土、生物与矿产等资源的过度开发与不合理利用,不仅使部分资源质量下降,也导致局部地区环境趋于恶化,诸如草场退化、森林减少、荒漠化土地增大以及一些主要城乡的三废污染日益严重等。所以,适当控制人口、适度开发与合理利用资源、加强环境整治与自然保护区建设等乃是协调人地关系,保护高原资源与环境,促使高原地区经济可持续发展的基本对策。
[12] Lin HL Hou FJ Ren JZ 2008 ). Evaluation of indicators of grazing trampling intensityActa Prataculturae Sinica, 17(1 ), 85 -92 . DOI:
10.3321/j.issn:1004-5759.2008.01.014 URL [本文引用: 1] 基于对践踏强度评定中存在问题的分析,提出践踏强度是在特定的放牧方式(自由放牧或轮牧) 下,单一种或组合种畜群单位时间内(通常指1个放牧季内)对单位草地面积所施加的践踏重力或践踏面积,它应有3种表示方法:重力表示法、面积表示法和频率 表示法。首次提出了践踏单位和践踏当量概念,以明确践踏强度的计算标准和折算标准。具体是指在放牧季中,一头体重40k的滩羊每蹄践踏1次所施加于草地的 践踏称为1个践踏单位——羊践踏,促进了践踏强度单位表示方法的统一化。引入的践踏当量概念,使得不同类别、不同年龄的家畜个体对草地的践踏量,均可通过 践踏当量换算为统一单位——羊践踏,是制定出合理而简明的践踏强度指标计算公式的依据,有助于使当前不统一的践踏强度表示方法规范化。从纷繁复杂的影响践 踏强度的众多因素中提炼出践踏次数或行走步数作为变量指标,把家畜的放牧行为及其时间分配模式归结到践踏次数中去。为计算单一种群或混合群践踏强度指标奠 定了计算基础。在相同的放牧强度下,通过对虚拟的3个单一种群和1个混合畜群践踏强度的计算比较表明,如果忽略畜种在践踏次数上的差异,牛群和马群的践踏 强度分别是羊群的5.6和4.7倍,混合群的践踏强度是羊群的4.8倍。而在此种情况下,用放牧强度或用单位面积草地上家畜体重与放牧时间的乘积表示的践 踏强度均相等,因此有理由认为用放牧强度或用单位面积草地上家畜体重与放牧时间的乘积表示单一种群或混合畜群的践踏强度均是不准确的。
[ 林慧龙 , 侯扶江 , 任继周 ( 2008 ). 放牧家畜的践踏强度指标探讨草业学报 , 17(1 ), 85 -92 .] DOI:
10.3321/j.issn:1004-5759.2008.01.014 URL [本文引用: 1] 基于对践踏强度评定中存在问题的分析,提出践踏强度是在特定的放牧方式(自由放牧或轮牧) 下,单一种或组合种畜群单位时间内(通常指1个放牧季内)对单位草地面积所施加的践踏重力或践踏面积,它应有3种表示方法:重力表示法、面积表示法和频率 表示法。首次提出了践踏单位和践踏当量概念,以明确践踏强度的计算标准和折算标准。具体是指在放牧季中,一头体重40k的滩羊每蹄践踏1次所施加于草地的 践踏称为1个践踏单位——羊践踏,促进了践踏强度单位表示方法的统一化。引入的践踏当量概念,使得不同类别、不同年龄的家畜个体对草地的践踏量,均可通过 践踏当量换算为统一单位——羊践踏,是制定出合理而简明的践踏强度指标计算公式的依据,有助于使当前不统一的践踏强度表示方法规范化。从纷繁复杂的影响践 踏强度的众多因素中提炼出践踏次数或行走步数作为变量指标,把家畜的放牧行为及其时间分配模式归结到践踏次数中去。为计算单一种群或混合群践踏强度指标奠 定了计算基础。在相同的放牧强度下,通过对虚拟的3个单一种群和1个混合畜群践踏强度的计算比较表明,如果忽略畜种在践踏次数上的差异,牛群和马群的践踏 强度分别是羊群的5.6和4.7倍,混合群的践踏强度是羊群的4.8倍。而在此种情况下,用放牧强度或用单位面积草地上家畜体重与放牧时间的乘积表示的践 踏强度均相等,因此有理由认为用放牧强度或用单位面积草地上家畜体重与放牧时间的乘积表示单一种群或混合畜群的践踏强度均是不准确的。
[13] Liu W Wang QJ Wang X Zhou L Li YF Li FJ 1999 ). Ecological process of forming “black soil-type” degraded grasslandActa Agrestia Sinica, 7, 301 -305 . URL [本文引用: 1] 研究分析退化草地的植被组成、地上生物量、优良牧草比例、高原鼠免密度以及土经性质等特征,探讨“黑土型”退化草地的成因及生态过程。结果透明,中度退化草地生物量最高950.3g/0.25m^2)。原生植被优良牧草比例最高(78.40%)。重度退化草地最低(4.49%),轻度和中度退化草地居中(57.2%和16.42%)。中度退化草地高原鼠兔平均密度较高(148只/hm^2),二者之间呈显著的正相关(r=
[ 刘伟 , 王启基 , 王溪 , 周立 , 李有福 , 李发吉 ( 1999 ). 高寒草甸“黑土型”退化草地的成因及生态过程草地学报 , 7, 301 -305 .] URL [本文引用: 1] 研究分析退化草地的植被组成、地上生物量、优良牧草比例、高原鼠免密度以及土经性质等特征,探讨“黑土型”退化草地的成因及生态过程。结果透明,中度退化草地生物量最高950.3g/0.25m^2)。原生植被优良牧草比例最高(78.40%)。重度退化草地最低(4.49%),轻度和中度退化草地居中(57.2%和16.42%)。中度退化草地高原鼠兔平均密度较高(148只/hm^2),二者之间呈显著的正相关(r=
[14] Liu XN Sun JL Zhang DG Pu XP Xu GP 2008 ). A study on the community structure and plant diversity of alpine meadow under different degrees of degradation in the Eastern Qilian MountainsActa Prataculturae Sinica, 17(4 ), 1 -11 . [本文引用: 3] [ 柳小妮 , 孙九林 , 张德罡 , 蒲小鹏 , 徐广平 ( 2008 ). 东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究,草业学报 , 17(4 ), 1 -11 .] [本文引用: 3] [15] Liu XQ Wu QH Li HQ Mao SJ Li YN 2013 ). A comparison of the vegetation/soil carbon density and net ecosystem CO2 exchange of alpine meadow with different enclosure durationsJournal of Glaciology and Geocryology, 35, 848 -856 . DOI:
10.7522/j.issn.1000-0240.2013.0096 URL [本文引用: 1] 封育是推广范围最广的草地恢复措施之一.为研究不同封育年限高寒 草甸植被、土壤碳密度变化,对1 a、6 a和16a不同封育年限样地监测结果进行分析.结果表明:不同封育年限高寒草甸植被现存碳密度表现出封育16 a>封育1 a>封育6 a,分别为1 522.57 gC·m_2、1 323.12 gC·m-2和1 148.17 gC·m_2,但不同封育年限之间植被现存碳密度差异不显著(P>0.05).土壤碳密度垂直分布明显,0~5 cm和5~10 cm土层有机碳密度较高,随土层深度增加土壤有机碳密度明显下降,土壤容重上升;不同封育年限之间0~40 cm层次土壤碳密度和土壤容重差异性均不显著,但仍可表现出土壤碳密度封育1 a>封育6 a>封育16a,分别为28 636.32 gC·m-2、26 570.92 gC·m-2和26 060.71gC· m-2;同时,土壤容重随封育时间延长而下降.对7月下旬到10月上旬净生态系统CO2交换率(NEE)监测来看,封育1 a植被土壤碳吸收速率显著高于封育16 a(P<0.05);而排放率与封育16a样地接近,差异不显著(P>0.05).
[ 刘晓琴 , 吴启华 , 李红琴 , 毛绍娟 , 李英年 ( 2013 ). 不同封育年限高寒草甸植被/土壤碳密度及净生态系统CO2 交换量的比较冰川冻土 , 35, 848 -856 .] DOI:
10.7522/j.issn.1000-0240.2013.0096 URL [本文引用: 1] 封育是推广范围最广的草地恢复措施之一.为研究不同封育年限高寒 草甸植被、土壤碳密度变化,对1 a、6 a和16a不同封育年限样地监测结果进行分析.结果表明:不同封育年限高寒草甸植被现存碳密度表现出封育16 a>封育1 a>封育6 a,分别为1 522.57 gC·m_2、1 323.12 gC·m-2和1 148.17 gC·m_2,但不同封育年限之间植被现存碳密度差异不显著(P>0.05).土壤碳密度垂直分布明显,0~5 cm和5~10 cm土层有机碳密度较高,随土层深度增加土壤有机碳密度明显下降,土壤容重上升;不同封育年限之间0~40 cm层次土壤碳密度和土壤容重差异性均不显著,但仍可表现出土壤碳密度封育1 a>封育6 a>封育16a,分别为28 636.32 gC·m-2、26 570.92 gC·m-2和26 060.71gC· m-2;同时,土壤容重随封育时间延长而下降.对7月下旬到10月上旬净生态系统CO2交换率(NEE)监测来看,封育1 a植被土壤碳吸收速率显著高于封育16 a(P<0.05);而排放率与封育16a样地接近,差异不显著(P>0.05).
[16] Long Y Meng FD Wang CS Bai L Zhong Y Wang SP 2015 ). Above- and below-ground biomass distribution of main alpine meadow plants and impact of degradation on root/shoot ratio and root areaGuihaia, 35, 532 -538 . DOI:
10.11931/guihaia.gxzw201406032 URL [本文引用: 2] 研究高寒草甸主要植物地上地下生物量的分布及其对退化的响应有利于了解高寒草甸的退化过程。该研究首先在西藏那曲生态环境综合观测研究站小嵩草围栏内(2009年围封)选择原生植被较好的地点随机选择小嵩草(Kobresia pygmaea)、矮嵩草(K.humilis)、紫花针茅(Stipa purpurea)、二裂委陵菜(Potentila bifurca)和青藏苔草(Carex moorcroftii)等5种植物斑块,选择退化斑块上(与原生植被相比)的二裂委陵菜和青藏苔草;然后用烘箱烘至恒重并称重,用扫描仪对根系进行扫描用于估算根系表面积;最后利用2因子方差分析检验不同物种个体、不同取样层次对地上和地下生物量的影响,利用物种和退化状态2因子方差分析检验对地上生物量的影响,以及利用物种、取样层次和退化状态3因子方差分析检验对二裂委陵菜和青藏苔草地下生物量、根冠比和根系表面积的影响。结果表明:在未退化条件下,小嵩草、矮嵩草和紫花针茅0~10cml地下生物量占0~30cm地下生物量的70%以上,0~30cm地下生物量占其地上地下总生物量的96%以上;二裂委陵菜(Potentilla bifurca)和青藏苔草(Carex moorcroftii)0~10cm地下生物量占0~30cml地下生物量的50%以上,其中二裂委陵菜0~30cm地下生物量占其地上地下总生物量的57%,青藏苔草0~30cm地下生物量占其地上地下总生物量的87%;对于退化草甸的主要植物,退化显著降低了二裂委陵菜的地上生物量、地下生物量和根冠比,对其根系表面积影响不大,但显著增加了青藏苔草的地上生物量,降低了其根冠比,对其地下生物量和根系表面积影响不大。
[ 龙毅 , 孟凡栋 , 王常顺 , 白玲 , 钟扬 , 汪诗平 ( 2015 ). 高寒草甸主要植物地上地下生物量分布及退化对根冠比和根系表面积的影响广西植物 , 35, 532 -538 .] DOI:
10.11931/guihaia.gxzw201406032 URL [本文引用: 2] 研究高寒草甸主要植物地上地下生物量的分布及其对退化的响应有利于了解高寒草甸的退化过程。该研究首先在西藏那曲生态环境综合观测研究站小嵩草围栏内(2009年围封)选择原生植被较好的地点随机选择小嵩草(Kobresia pygmaea)、矮嵩草(K.humilis)、紫花针茅(Stipa purpurea)、二裂委陵菜(Potentila bifurca)和青藏苔草(Carex moorcroftii)等5种植物斑块,选择退化斑块上(与原生植被相比)的二裂委陵菜和青藏苔草;然后用烘箱烘至恒重并称重,用扫描仪对根系进行扫描用于估算根系表面积;最后利用2因子方差分析检验不同物种个体、不同取样层次对地上和地下生物量的影响,利用物种和退化状态2因子方差分析检验对地上生物量的影响,以及利用物种、取样层次和退化状态3因子方差分析检验对二裂委陵菜和青藏苔草地下生物量、根冠比和根系表面积的影响。结果表明:在未退化条件下,小嵩草、矮嵩草和紫花针茅0~10cml地下生物量占0~30cm地下生物量的70%以上,0~30cm地下生物量占其地上地下总生物量的96%以上;二裂委陵菜(Potentilla bifurca)和青藏苔草(Carex moorcroftii)0~10cm地下生物量占0~30cml地下生物量的50%以上,其中二裂委陵菜0~30cm地下生物量占其地上地下总生物量的57%,青藏苔草0~30cm地下生物量占其地上地下总生物量的87%;对于退化草甸的主要植物,退化显著降低了二裂委陵菜的地上生物量、地下生物量和根冠比,对其根系表面积影响不大,但显著增加了青藏苔草的地上生物量,降低了其根冠比,对其地下生物量和根系表面积影响不大。
[17] Luo YY Meng QT Zhang JH Zhao XY Qin Y 2014 ). Species diversity and biomass in relation to soil properties of alpine meadows in the eastern Tibetan Plateau in different degradation stagesJournal of Glaciology and Geocryology, 36, 1298 -1305 . [本文引用: 12] [ 罗亚勇 , 孟庆涛 , 张静辉 , 赵学勇 , 秦彧 ( 2014 ). 青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系冰川冻土 , 36, 1298 -1305 .] [本文引用: 12] [18] Luo YY Zhang Y Zhang JH Ka ZJ Shang LY Wang SY 2012 ). Soil stiochiometry characteristics among alpine meadows of different degradation stages in Maqu CountyChinese Journal of Ecology, 31, 254 -260 . URL [本文引用: 1] 为了阐明不同退化阶段高寒草甸土壤的化学计量特征,沿着高寒草甸 退化的梯度选取了原生嵩草草甸、轻度退化草甸和严重沙化草甸,测定了高寒草甸退化过程中不同深度土壤的有机碳、全氮、全磷和全钾含量.结果表明:随着高寒 草甸的退化,0~100 cm土壤的有机碳、全氮、全磷和全钾含量以及碳氮比、碳磷比、碳钾比、氮磷比、氮钾比和磷钾比均呈降低趋势,且土壤有机碳对高寒草甸退化的敏感性最高,全 氮、全磷和全钾的敏感性依次降低,表层20 cm的土壤有机碳和金氮可作为表征高寒草甸退化程度最敏感的土壤养分指标.另外,随着草甸的退化,土壤的有机碳、全氮、全磷和全钾含量及其化学计量比的垂 直分布明显不同:随着土壤深度的增加,原生嵩草草甸和轻度退化草甸的土壤有机碳、全氮和全磷含量以及碳氮比、碳磷比、碳钾比、氮磷比、氮钾比和磷钾比在0 ~ 40 cm范围内锐减,在40cm以下缓慢降低并趋于稳定;而沙化草甸土壤的有机碳、全氮、全磷和全钾及其化学计量比随着土壤深度的增加保持不变.
[ 罗亚勇 , 张宇 , 张静辉 , 卡召加 , 尚伦宇 , 王少影 ( 2012 ). 不同退化阶段高寒草甸土壤化学计量特征生态学杂志 , 31, 254 -260 .] URL [本文引用: 1] 为了阐明不同退化阶段高寒草甸土壤的化学计量特征,沿着高寒草甸 退化的梯度选取了原生嵩草草甸、轻度退化草甸和严重沙化草甸,测定了高寒草甸退化过程中不同深度土壤的有机碳、全氮、全磷和全钾含量.结果表明:随着高寒 草甸的退化,0~100 cm土壤的有机碳、全氮、全磷和全钾含量以及碳氮比、碳磷比、碳钾比、氮磷比、氮钾比和磷钾比均呈降低趋势,且土壤有机碳对高寒草甸退化的敏感性最高,全 氮、全磷和全钾的敏感性依次降低,表层20 cm的土壤有机碳和金氮可作为表征高寒草甸退化程度最敏感的土壤养分指标.另外,随着草甸的退化,土壤的有机碳、全氮、全磷和全钾含量及其化学计量比的垂 直分布明显不同:随着土壤深度的增加,原生嵩草草甸和轻度退化草甸的土壤有机碳、全氮和全磷含量以及碳氮比、碳磷比、碳钾比、氮磷比、氮钾比和磷钾比在0 ~ 40 cm范围内锐减,在40cm以下缓慢降低并趋于稳定;而沙化草甸土壤的有机碳、全氮、全磷和全钾及其化学计量比随着土壤深度的增加保持不变.
[19] Mao F Zhang YH Hou YY Tang SH Lu ZG Zhang JH 2008 ). Dynamic assessment of grassland degradation in Naqu of northern TibetChinese Journal of Applied Ecology, 19, 278 -284 . [本文引用: 3] [ 毛飞 , 张艳红 , 侯英雨 , 唐世浩 , 卢志光 , 张佳华 ( 2008 ). 藏北那曲地区草地退化动态评价应用生态学报 , 19, 278 -284 .] [本文引用: 3] [20] Niu KC Luo YJ Choler P Du GZ 2008 ). The role of biomass allocation strategy in diversity loss due to fertilizationBasic and Applied Ecology, 9, 485 -493 . DOI:
10.1016/j.baae.2007.06.015 URL [本文引用: 1] Vorgeschlagene Mechanismen, die den Biodiversit01tsverlust erkl01ren, schlie08en interspezifische Konkurrenz und eine Ausdünnung der Vergesellschaftungsebenen ein. Die interspezifische Konkurrenz Hypothese geht von einer Verbindung zwischen den Populations01nderungen und der Konkurrenzf01higkeit von Arten aus, die mit funktionellen Eigenschaften wie z. B. der Biomassenallokation zusammenh01ngt. Auf der Basis eines zweij01hrigen Düngeexperimentes auf einer alpinen Wiese des tibetischen Plateaus versuchten wir die Beziehungen zwischen inviduellen und Populationsreaktionen herauszufinden. Die individuellen Reaktionen wurden durch die Ver01nderungen in der Biomasse und der Biomassenallokation gemessen, und die Populationsreaktion wurde durch Ver01nderungen in den Artenabundanzen abgesch01tzt. Die Ergebnisse lassen erkennen, dass als Folge der Düngung (1) sich die Ver01nderungen der individuellen Biomasse zwischen Arten und funktionellen Gruppen unterscheiden, (2) die Reproduktionsallokation bei allen Arten zu einer Abnahme tendiert, w01hrend die Blattallokation für Gr01ser zunimmt und für Stauden abnimmt, (3) eine starke positive Korrelation zwischen der relativen Ver01nderung der Artenabundanz und der individuellen Biomassenreaktion existiert und (4) die relative Ver01nderung der Artenabundanz eine positive Korrelation zur Blattallokationsreaktion, eine negative zur Stammallokationsreaktion und keine signifikante Korrelation zur Reproduktionsallokationsreaktion hat. Wir schlie08en daraus, dass die individuellen Biomassenreaktionen und die Biomassenallokations-Strategien den Diversit01tsverlust aufgrund von Düngung zum Teil erkl01ren kann. Dies ist ein Ergebnis, das mit der interspezifischen Konkurrenz Hypothese übereinstimmt.
[21] Qi B Zhang DG Ding LL Zhang L Wang X Yang YH 2005 ). Properties of plant community diversity of degraded alpine arid grasslandsJournal of Gansu Agricultural University, 40, 626 -631 . DOI:
10.3969/j.issn.1003-4315.2005.05.012 URL [本文引用: 1] 对青海省三角城种羊场的高寒干旱草地植物群落多样性特征进行了调查研究,结果表明,随退化程度的加剧,放牧草地的群落物种组成、结构和植被群落多样性发生了重大改变.在长期的放牧利用过程中,草地群落的优势物种由原始的赖草、早熟禾逐渐演变为紫花针茅,其伴生种也发生了明显的相互更替.随着草地退化程度的加重,群落的植物多样性和物种丰富度及均匀度都明显的随之降低,而群落的优势度逐步增大.而围栏封育的草地物种多样性、丰富度和均匀度均显著低于适度放牧利用的草地.因而,对于高寒干旱草地,适度的放牧利用有利于群落物种丰富度和多样性的提高,过度放牧势必导致植被群落的逆向演替,长期的围栏封育并不利于物种多样性和丰富度的提高.
[ 祁彪 , 张德罡 , 丁玲玲 , 张力 , 汪玺 , 杨予海 ( 2005 ). 退化高寒干旱草地植物群落多样性特征甘肃农业大学学报 , 40, 626 -631 .] DOI:
10.3969/j.issn.1003-4315.2005.05.012 URL [本文引用: 1] 对青海省三角城种羊场的高寒干旱草地植物群落多样性特征进行了调查研究,结果表明,随退化程度的加剧,放牧草地的群落物种组成、结构和植被群落多样性发生了重大改变.在长期的放牧利用过程中,草地群落的优势物种由原始的赖草、早熟禾逐渐演变为紫花针茅,其伴生种也发生了明显的相互更替.随着草地退化程度的加重,群落的植物多样性和物种丰富度及均匀度都明显的随之降低,而群落的优势度逐步增大.而围栏封育的草地物种多样性、丰富度和均匀度均显著低于适度放牧利用的草地.因而,对于高寒干旱草地,适度的放牧利用有利于群落物种丰富度和多样性的提高,过度放牧势必导致植被群落的逆向演替,长期的围栏封育并不利于物种多样性和丰富度的提高.
[22] Ren QJ Cui XL Zhao BB 2008 ). Effects of grazing impact on community structure and productivity in an alpine meadowActa Prataculturae Sinica, 17(6 ), 134 -140 . DOI:
10.3321/j.issn:1004-5759.2008.06.019 URL Magsci [本文引用: 6] <FONT face=Verdana>草地的退化是当前畜牧生产面临的一个主要问题,本试验选取青藏高原高寒草地自然群落中等放牧强度和不放牧2个梯度进行对比研究,探讨当前的普遍放牧强度对群落生产力、不同功能群产量分配和群落结构的影响。结果表明,3年中放牧使群落总体生产力下降,其中不同功能群所占比例也有所变化,禾草和莎草比例由平均76.6%下降到57.4%,杂草、豆科和毒草的比例上升;群落高度降低,平均最大高度由46.8cm 下降到22.4cm;物种多样性和多度增加,多样性由单位样方平均26.5种增加到30.9种,多度由单位样方215.7株增加到304.6株;总盖度并没有明显差异,禾草和莎草盖度减小,杂草盖度增加,群落结构由垂直分层为主演化为水平生态位分化为主。3年中放牧的影响没有显示出草地的进一步退化趋势。</FONT>
[ 仁青吉 , 崔现亮 , 赵彬彬 ( 2008 ). 放牧对高寒草甸植物群落结构及生产力的影响草业学报 , 17(6 ), 134 -140 .] DOI:
10.3321/j.issn:1004-5759.2008.06.019 URL Magsci [本文引用: 6] <FONT face=Verdana>草地的退化是当前畜牧生产面临的一个主要问题,本试验选取青藏高原高寒草地自然群落中等放牧强度和不放牧2个梯度进行对比研究,探讨当前的普遍放牧强度对群落生产力、不同功能群产量分配和群落结构的影响。结果表明,3年中放牧使群落总体生产力下降,其中不同功能群所占比例也有所变化,禾草和莎草比例由平均76.6%下降到57.4%,杂草、豆科和毒草的比例上升;群落高度降低,平均最大高度由46.8cm 下降到22.4cm;物种多样性和多度增加,多样性由单位样方平均26.5种增加到30.9种,多度由单位样方215.7株增加到304.6株;总盖度并没有明显差异,禾草和莎草盖度减小,杂草盖度增加,群落结构由垂直分层为主演化为水平生态位分化为主。3年中放牧的影响没有显示出草地的进一步退化趋势。</FONT>
[23] Sala OE Oesterheld M León RJC Soriano A 1986 ). Grazing effects upon plant community structure in subhumid grasslands of ArgentinaVegetatio, 67, 27 -32 . DOI:
10.1007/BF00040315 URL [本文引用: 1] Changes in plant community structure are identified as a result of grazing in grasslands of the flooding pampa which evolved under supposedly light grazing conditions. The effect of excluding grazing upon total leaf area index was an increase of 30%. The largest response was observed in the distribution of leaves in the canopy. In the grazed areas, most of the green material was concentrated in the 0–5 cm layer while in the ungrazed treatments the largest portion of the leaf area was in the 10–30 cm layer. Grazing exclusion resulted in a small change in total basal area but a larger change in its distribution, from many small tussocks to less numerous large ones. The effect of grazing upon leaf area and basal area was accounted for by changes in vigor as well as by changes in species composition. The major effect of excluding grazing upon species composition was the disappearance of some native planophile species and most of the exotics. The species composition of grazed areas of both communities was very similar while there were large differences between the ungrazed areas and between the grazed and ungrazed areas of the same community. It is suggested that there is a group of species which responds to the coarse-grained ‘signal’ of grazing and its presence can cause dissimilar communities to converge under grazing conditions. The other group of species responded to the fine-grained ‘signal’ of the environmental conditions associated with topography.
[24] Sheng L Wang YL 2010 ). Characteristics variation of soil nutrition in different degraded degree alpine meadow in Dawu region of Guoluo prefectureChinese Qinghai Journal of Animal and Veterinary Sciences, 40(4 ), 4 -6 . DOI:
10.3969/j.issn.1003-7950.2010.04.003 URL [本文引用: 1] 对果洛大武地区不同退化梯度下高寒草甸土壤养分特征变化进行了研 究,结果表明:各退化梯度群落内土壤有机质,全氮,速效氮损失较大,平均流失率为46.7%,44.1%以及49.8%;全磷流失相对较少;速效磷含量的 变化没有规律性,在轻度和中度退化草地中有较大幅度的损失,在重度退化草地中则明显升高.
[ 盛丽 , 王彦龙 ( 2010 ). 果洛大武地区不同退化梯度下高寒草甸土壤养分特征变化青海畜牧兽医杂志 , 40(4 ), 4 -6 .] DOI:
10.3969/j.issn.1003-7950.2010.04.003 URL [本文引用: 1] 对果洛大武地区不同退化梯度下高寒草甸土壤养分特征变化进行了研 究,结果表明:各退化梯度群落内土壤有机质,全氮,速效氮损失较大,平均流失率为46.7%,44.1%以及49.8%;全磷流失相对较少;速效磷含量的 变化没有规律性,在轻度和中度退化草地中有较大幅度的损失,在重度退化草地中则明显升高.
[25] Studies on the Development of Agriculture and Animal Husbandry Industry with Characteristics of Tibet Autonomous Region Group ( 2003 ). Studies on the development of agriculture and animal husbandry industry with characteristics of Tibet Autonomous RegionJournal of China Agricultural Resources and Regional Planning, 24(6 ), 8 -12 . [本文引用: 1] [ 西藏自治区农牧业特色产业发展研究课题组 ( 2003 ). 西藏自治区农牧业特色产业发展研究中国农业资源与区划 , 24(6 ), 8 -12 .] [本文引用: 1] [26] Wang CT Long RJ Wang QL Cao GM Shi JJ Du YG 2008 ). Response of plant diversity and productivity to soil response changing under grazing disturbance on an alpine meadowActa Ecologica Sinica, 28, 4144 -4152 . DOI:
10.3321/j.issn:1000-0933.2008.09.011 URL [本文引用: 1] 为了揭示高寒小嵩草草甸群落在放牧扰动下,探讨土壤养分供给水平的变化对生态系统初级生产力和多样性影响,为高寒草地的退化演替机理研究提供依据,以野外样地调查和室内分析法研究了放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应。结果表明,放牧干扰不仅改变了高寒小蒿草草甸群落土壤根系和蕴育土壤根系的"载体"量及根土比例,改变了植物群落的结构和功能,而且使土壤的物理和化学特性发生了明显的改变。随着放牧强度的增加,蕴育土壤根系的基质量逐渐减少,根土比特别是0~10 cm土层的根土比例增加;"载体"量减少导致大部分地下根系由于营养供给水平的降低而死亡,归还土壤中有机质的数量逐渐减少,加之地上部分持续利用,土壤养分也在不断消耗,土壤基质量的减少和土壤资源持续供给能力的下降,草地发生逆向演替(退化),表现在:物种数减少、多样性下降、能量的分配转向地下等;土壤性状上的某些改变(土壤容重、土壤湿度等),也会引起植被组成、物种多样性变化;放牧主要通过影响土壤环境及其养分含量来改变草地群落生物量(地上、地下);土壤表面的适度干扰和原有植物的适度破坏为新成员提供了小生境,从而允许新的植物侵入群落,并提高了植物的丰富度。但是,在受到强度干扰时,草地植物群落的主要物种的优势地位发生明显的替代变化。
[ 王长庭 , 龙瑞军 , 王启兰 , 曹广民 , 施建军 , 杜岩功 ( 2008 ). 放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应生态学报 , 28, 4144 -4152 .] DOI:
10.3321/j.issn:1000-0933.2008.09.011 URL [本文引用: 1] 为了揭示高寒小嵩草草甸群落在放牧扰动下,探讨土壤养分供给水平的变化对生态系统初级生产力和多样性影响,为高寒草地的退化演替机理研究提供依据,以野外样地调查和室内分析法研究了放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应。结果表明,放牧干扰不仅改变了高寒小蒿草草甸群落土壤根系和蕴育土壤根系的"载体"量及根土比例,改变了植物群落的结构和功能,而且使土壤的物理和化学特性发生了明显的改变。随着放牧强度的增加,蕴育土壤根系的基质量逐渐减少,根土比特别是0~10 cm土层的根土比例增加;"载体"量减少导致大部分地下根系由于营养供给水平的降低而死亡,归还土壤中有机质的数量逐渐减少,加之地上部分持续利用,土壤养分也在不断消耗,土壤基质量的减少和土壤资源持续供给能力的下降,草地发生逆向演替(退化),表现在:物种数减少、多样性下降、能量的分配转向地下等;土壤性状上的某些改变(土壤容重、土壤湿度等),也会引起植被组成、物种多样性变化;放牧主要通过影响土壤环境及其养分含量来改变草地群落生物量(地上、地下);土壤表面的适度干扰和原有植物的适度破坏为新成员提供了小生境,从而允许新的植物侵入群落,并提高了植物的丰富度。但是,在受到强度干扰时,草地植物群落的主要物种的优势地位发生明显的替代变化。
[27] Wang DL Lü XL Luo WD 1996 ). Analysis to effects of different grazing density on characteristics of rangeland vegetationActa Prataculturae Sinica, 5(3 ), 28 -33 . [本文引用: 1] [ 王德利 , 吕新龙 , 罗卫东 ( 1996 ). 不同放牧密度对草原植被特征的影响分析草业学报 , 5(3 ), 28 -33 .] [本文引用: 1] [28] Wang GX Shen YP Cheng GD 2000 ). Eco-environmental changes and causal analysis in the source regions of the Yellow RiverJournal of Glaciology and Geocryology, 22, 200 -205 . [本文引用: 1] [ 王根绪 , 沈永平 , 程国栋 ( 2000 ). 黄河源区生态环境变化与成因分析冰川冻土 , 22, 200 -205 .] [本文引用: 1] [29] Wang QL Wang CT Du YG Cao GM 2008 ). Grazing impact on soil microbial biomass carbon and relationships with soil environment in alpine Kobresia meadowActa Prataculturae Sinica, 2 ), 39 -46 . DOI:
10.3321/j.issn:1004-5759.2008.02.006 URL [本文引用: 1] 通过对高寒嵩草草甸不同放牧压力下土壤微生物量碳、土壤物理性状和速效养分的变化特征及其相互关系的研究,表明随着放牧压力的增大,土壤微生物量碳、有机质及速效磷显著降低;土壤pH值、容重及根土比显著升高;速效氮和速效钾变化不一致。相关性分析表明,土壤有机质、根土比、有效磷、土壤容重与微生物量碳具有较大的关联度;土壤有机质、有效磷、土壤容重和根土比对微生物量碳的形成有较强的直接作用,有效氮表现出较强的间接效应;根土比-有效磷、有机质-土壤容重、有效氮-土壤容重之间互相调节,互相制约,间接调控着微生物量碳形成。过度放牧使土壤系统的微环境恶化,速效养分的供给能力及有机物质的转化能力降低,从而导致了植被和土壤系统的双重退化和微生物量碳的低下。
[ 王启兰 , 王长庭 , 杜岩功 , 曹广民 ( 2008 ). 放牧对高寒嵩草草甸土壤微生物量碳的影响及其与土壤环境的关系草业学报 , 17(2 ), 39 -46 .] DOI:
10.3321/j.issn:1004-5759.2008.02.006 URL [本文引用: 1] 通过对高寒嵩草草甸不同放牧压力下土壤微生物量碳、土壤物理性状和速效养分的变化特征及其相互关系的研究,表明随着放牧压力的增大,土壤微生物量碳、有机质及速效磷显著降低;土壤pH值、容重及根土比显著升高;速效氮和速效钾变化不一致。相关性分析表明,土壤有机质、根土比、有效磷、土壤容重与微生物量碳具有较大的关联度;土壤有机质、有效磷、土壤容重和根土比对微生物量碳的形成有较强的直接作用,有效氮表现出较强的间接效应;根土比-有效磷、有机质-土壤容重、有效氮-土壤容重之间互相调节,互相制约,间接调控着微生物量碳形成。过度放牧使土壤系统的微环境恶化,速效养分的供给能力及有机物质的转化能力降低,从而导致了植被和土壤系统的双重退化和微生物量碳的低下。
[30] Wang W Liu ZL Hao DY Liang CZ 1996 ). Research on the restoring succession of the degenerated grassland in Inner Mongolia II. Analysis of the restoring processActa Phytoecologica Sinica, 20, 460 -471 . URL [本文引用: 2] 根据内蒙古典型草原地带的羊草+大针茅草原退化变型-冷蒿群落封育12年(1983~1994)的动态监测数据进行分析,对群落恢复演替轨迹取得以下认识:1.依据群落优势种的更替及主分量分析结果可将恢复演替过程划分为冷蒿优势阶段、冷蒿+冰草阶段、冰草优势阶段、羊草优势阶段。2.退化草原群落在恢复演替过程中,群落生产力的变化表现出阶梯式跃变和亚稳态阶面相间的特点。第一次跃变发生在1984年,上升到第二个阶面,第二次跃变发生在1990年,进入了第三个阶面,已接近于原生群落的生产力。3.群落生产力与水资源量的关系因恢复演替阶段不同而异。第一亚稳态时期,群落地上现存生物量大体处于166g·m ̄-2的水平上,生长季降水量达176mm以上时,增加降水对群落生产力的提高不发生显著影响。第二亚稳态时期,群落生物量与降水量之间的相关性显著。可推算出群落于物质生产用水量介于1.1~1.6mm·g ̄-1之间。此值在1.1mm·g ̄-1时,群落对水资源的利用效率最高,而在1.6mm·g ̄-1时群落生物量达到最大值。4.在恢复演替进程中,群落密度的位点常数约为271.5株·m ̄-2,循此常数上下波动,表现出拥挤与稀疏交替发生的过程,构成了恢
[ 王炜 , 刘钟龄 , 郝敦元 , 梁存柱 ( 1996 ). 内蒙古草原退化群落恢复演替的研究: II.恢复演替时间进程的分析植物生态学报 , 20, 460 -471 .] URL [本文引用: 2] 根据内蒙古典型草原地带的羊草+大针茅草原退化变型-冷蒿群落封育12年(1983~1994)的动态监测数据进行分析,对群落恢复演替轨迹取得以下认识:1.依据群落优势种的更替及主分量分析结果可将恢复演替过程划分为冷蒿优势阶段、冷蒿+冰草阶段、冰草优势阶段、羊草优势阶段。2.退化草原群落在恢复演替过程中,群落生产力的变化表现出阶梯式跃变和亚稳态阶面相间的特点。第一次跃变发生在1984年,上升到第二个阶面,第二次跃变发生在1990年,进入了第三个阶面,已接近于原生群落的生产力。3.群落生产力与水资源量的关系因恢复演替阶段不同而异。第一亚稳态时期,群落地上现存生物量大体处于166g·m ̄-2的水平上,生长季降水量达176mm以上时,增加降水对群落生产力的提高不发生显著影响。第二亚稳态时期,群落生物量与降水量之间的相关性显著。可推算出群落于物质生产用水量介于1.1~1.6mm·g ̄-1之间。此值在1.1mm·g ̄-1时,群落对水资源的利用效率最高,而在1.6mm·g ̄-1时群落生物量达到最大值。4.在恢复演替进程中,群落密度的位点常数约为271.5株·m ̄-2,循此常数上下波动,表现出拥挤与稀疏交替发生的过程,构成了恢
[31] Wang YX Cao JM 2007 ). How to solve the overgrazing problem in Inner MongoliaGreen Economy, ( 7 ), 58 -64 . DOI:
10.3969/j.issn.1671-4407.2007.07.011 URL [本文引用: 2] 内蒙古草地资源丰富,但面临着逐年退化、面积日益缩小的危机,其中草原过度放牧是重要原因之一。本文基于内蒙古草原面积过百万(公顷)的31个旗县的数据测算了全区草原过牧的程度,结果表明,内蒙古草原过牧率高居70%左右,而且过牧程度由轻度转向中度。最后本文对内蒙古草原过牧的原因进行了深入分析,并提出了相应的对策建议。
[ 王云霞 , 曹建民 ( 2007 ). 内蒙古草原过度放牧的解决途径生态经济 , ( 7 ), 58 -64 .] DOI:
10.3969/j.issn.1671-4407.2007.07.011 URL [本文引用: 2] 内蒙古草地资源丰富,但面临着逐年退化、面积日益缩小的危机,其中草原过度放牧是重要原因之一。本文基于内蒙古草原面积过百万(公顷)的31个旗县的数据测算了全区草原过牧的程度,结果表明,内蒙古草原过牧率高居70%左右,而且过牧程度由轻度转向中度。最后本文对内蒙古草原过牧的原因进行了深入分析,并提出了相应的对策建议。
[32] Wu ML 2010 ). Structural Equation Modeling: Operation and Response of AMOS . The University of Chongqing Press, Chongqing. [本文引用: 1] (in Chinese) [ 吴明隆 ( 2010 ).结构方程模型: AMOS的操作与应用 . 重庆大学出版.] [本文引用: 1] [33] Yang FY Zhang YW Miao YJ Wei XH 2003 ). Main limiting factors for deteriorated grasslands vegetation restoration of Northern Tibet PlateauBulletin of Soil and Water Conservation, 23(4 ), 17 -20 . DOI:
10.3969/j.issn.1000-288X.2003.04.005 URL [本文引用: 1] 在介绍藏北高寒草原现状的基础上,从自然和人为因素两方面对藏北高寒退化草地植被恢复与重建过程中的障碍因子进行了初步探讨;提出退化草地植被恢复的一些措施,包括天然草场合理利用、土壤培育、南草北调和加强科学研究等对策.
[ 杨富裕 , 张蕴微 , 苗彦军 , 魏学红 ( 2003 ). 藏北高寒退化草地植被恢复过程的障碍因子初探水土保持通报 , 23(4 ), 17 -20 .] DOI:
10.3969/j.issn.1000-288X.2003.04.005 URL [本文引用: 1] 在介绍藏北高寒草原现状的基础上,从自然和人为因素两方面对藏北高寒退化草地植被恢复与重建过程中的障碍因子进行了初步探讨;提出退化草地植被恢复的一些措施,包括天然草场合理利用、土壤培育、南草北调和加强科学研究等对策.
[34] Zhang GS Li L 1999 ). Effects of climatic changes of South Qinghai Plateau on the alpine meadowActa Prataculturae Sinica, 8(3 ), 1 -10 . DOI:
10.1088/0256-307X/16/12/025 URL [本文引用: 1] 通过对1961 年以来青南高原气候变化的分析,发现该地区年平均降水量及秋季降水量无明显的变化趋势,且其年际间的波动趋于缓和;冬季(12~2 月)和春季(3~5 月)降水量的变化趋势呈现出逐年增多的趋势,气候倾向率分别为2.6 m m /10 a 和3.6 m m /10 a;夏季(6~8 月)降水量变化却表现出显著的减少趋势,其倾向率为- 6.5 m m /10 a。青南牧区牧草返青期气温回升速度在逐年减缓,而牧草枯黄期气温降低速度在逐年增大。该区的气候变化使青南高原主要优势牧草——嵩草的生长高度由80 年代末期的6~8 cm 下降到现在的3~5cm ,牧草开花期和籽粒成熟期的发育百分率普遍下降25% ~50% ,发育程度一般达不到50% ,天然草地产鲜草量和干草产量减少约为70% ~80% 。
[ 张国胜 , 李林 ( 1999 ). 青南高原气候变化及其对高寒草甸牧草生长影响的研究草业学报 , 8(3 ), 1 -10 .] DOI:
10.1088/0256-307X/16/12/025 URL [本文引用: 1] 通过对1961 年以来青南高原气候变化的分析,发现该地区年平均降水量及秋季降水量无明显的变化趋势,且其年际间的波动趋于缓和;冬季(12~2 月)和春季(3~5 月)降水量的变化趋势呈现出逐年增多的趋势,气候倾向率分别为2.6 m m /10 a 和3.6 m m /10 a;夏季(6~8 月)降水量变化却表现出显著的减少趋势,其倾向率为- 6.5 m m /10 a。青南牧区牧草返青期气温回升速度在逐年减缓,而牧草枯黄期气温降低速度在逐年增大。该区的气候变化使青南高原主要优势牧草——嵩草的生长高度由80 年代末期的6~8 cm 下降到现在的3~5cm ,牧草开花期和籽粒成熟期的发育百分率普遍下降25% ~50% ,发育程度一般达不到50% ,天然草地产鲜草量和干草产量减少约为70% ~80% 。
[35] Zhang J Zhao H Zhang T Zhao X Drake S 2005 ). Community succession along a chronosequence of vegetation restoration on sand dunes in Horqin Sandy LandJournal of Arid Environments, 62, 555 -566 . DOI:
10.1016/j.jaridenv.2005.01.016 URL [本文引用: 1] Species diversity influences plant community structure and function. This paper examines the patterns and dynamics of species diversity along a chronosequence of vegetation recovery on sand dunes in a semi-arid region to assess the probability of vegetation recovery via succession, and provides some implications for revegetation practices in this region. Species richness and diversity indices gradually increased with succession, except for a decline in the community of 18 years, which is attributed to the strong dominance of Artemisia halodendron. In each stage of the restoration process, there was a dominant species with particular life history traits which contribute to the dominance of this species. Species replacement and habitat changes were the main drivers of succession, while plant species and community succession drove the process of vegetation recovery. Results showed that restoration via succession holds promise for vegetation recovery and desertification control within protected, fenced enclosures.
[36] Zhang JP Chen XH Zou XY Ji HL Du J Cao ZJ Fu XF 2001 ). The eco-environmental problems and its countermeasures in TibetJournal of Mountain Science, 19, 81 -86 . DOI:
10.3969/j.issn.1008-2786.2001.01.016 URL [本文引用: 1] 以青藏高原生态环境构成和社会经济发展。分析了西藏自治区存在的生态环境问题及生态环境发展趋势,提出生态环境建设分区及区域生态环境建设对策。
[ 张建平 , 陈学华 , 邹学勇 , 靳鹤龄 , 杜杰 , 曹宗杰 , 付雪峰 ( 2001 ). 西藏自治区生态环境问题及对策山地学报 , 19, 81 -86 .] DOI:
10.3969/j.issn.1008-2786.2001.01.016 URL [本文引用: 1] 以青藏高原生态环境构成和社会经济发展。分析了西藏自治区存在的生态环境问题及生态环境发展趋势,提出生态环境建设分区及区域生态环境建设对策。
[37] Zhao CZ Long RJ Ma YH Ji SZ 2005 ). The impact of the institution of grassland property right on overgrazing—A case from Hongshiwo countryside, Sunan County, GansuActa Prataculturae Sinica, 14(1 ), 1 -5 . [本文引用: 2] [ 赵成章 , 龙瑞军 , 马永欢 , 吉生柱 ( 2005 ). 草地产权制度对草地超载过牧的影响——以肃南县红石窝乡的调查为例草业学报 , 14(1 ), 1 -5 .] [本文引用: 2] [38] Zheng W Dong QM Li SX Li HT Liu Y Yang SH 2012 ). Impact of grazing intensities on community biodiversity and productivity of alpine grassland in Qinghai Lake RegionActa Agrestia Sinica, 20, 1033 -1038 . [本文引用: 1] [ 郑伟 , 董全民 , 李世雄 , 李红涛 , 刘玉 , 杨时海 ( 2012 ). 放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响草地学报 , 20, 1033 -1038 .] [本文引用: 1] [39] Zhong XH 2005 ). Pay close attention to an issue at home and abroad academic circles all along: Research of the Qinghai-Tibet PlateauJournal of Mountain Science, 23, 257 -259 . [本文引用: 1] [ 钟祥浩 ( 2005 ). 国内外学术界一直关注的问题: 青藏高原研究山地学报 , 23, 257 -259 .] [本文引用: 1] [40] Zhou HK Zhao XQ Zhou Li Liu W Li YN Tang YH 2005 ). A study on correlations between vegetation degradation and soil degradation in the alpine meadow of the Qinghai-Tibetan PlateauActa Prataculturae Sinica, 14(3 ), 31 -40 . [本文引用: 8] [ 周华坤 , 赵新全 , 周立 , 刘伟 , 李英年 , 唐艳鸿 ( 2005 ). 青藏高原高寒草甸的植被退化与土壤退化特征研究草业学报 , 14(3 ), 31 -40 .] [本文引用: 8] [41] Zhu JT 2016 ). Effects of experimental warming on plant phenology in Xizang alpine meadowChinese Journal of Plant Ecology, 40, 1028 -1036 . [本文引用: 2] [ 朱军涛 ( 2016 ). 实验增温对藏北高寒草甸植物物候的影响植物生态学报 , 40, 1028 -1036 .] [本文引用: 2] [42] Zong N Shi PL Niu B Jiang J Song MH Zhang XZ He YT 2010 ). Effects of nitrogen and phosphorous fertilization on community structure and productivity of degraded alpine meadows in northern Tibet, ChinaChinese Journal of Applied Ecology, 25, 3458 -3468 . URL [本文引用: 1] 施肥是退化草地初期恢复常用的管理措施,被用于改良草地群落结构和提高牧草生产力.本文以藏北轻度和重度 退化高寒草甸为研究对象,设置低氮(50kgN·hm-2 ·a-1,LN)、高氮(100 kg N· hm-2·a-1,HN)施肥水平,及其与磷配施组合试验(50 kg N·hm-2·a-1+50kgP·hm-2·a-1,LN+P; 100kgN ·hm-2·a-1+50kgP ·hm-2·a-1,HN+P),研究施肥梯度与施肥类型对两种退化程度高寒草甸群落结构、物种丰富度与多样性、群落生物量及其分配的影响.结果表明:单 独施氮对轻度和重度退化高寒草甸群落盖度和生物量均无显著影响,而氮磷配施显著提高群落盖度及地上、地下生物量,其中LN+P处理的促进效果最显著.这表 明本研究区地上生产力受到氮和磷的共同限制.施肥处理对重度退化样地总生物量无显著影响,但显著降低了根冠比.HN与HN+P处理均显著降低轻度退化群落 的物种丰富度和均匀度,说明HN处理不利于轻度退化草地物种多样性和稳定性的维持.在轻度退化样地,施氮(尤其是HN处理)主要提高了禾草植物的重要值和 生物量,而氮磷配施则有利于莎草类植物的生长.施肥对重度退化群落植物的重要值影响较小,主要提高了杂草生物量.说明轻度退化高寒草甸可选择低氮配施磷肥 的措施,而对重度退化草地可能需要结合围栏和补播牧草等其他管理措施进行改良.
[ 宗宁 , 石培礼 , 牛犇 , 蒋婧 , 宋明华 , 张宪洲 , 何永涛 ( 2010 ). 氮磷配施对藏北退化高寒草甸群落结构和生产力的影响应用生态学报 , 25, 3458 -3468 .] URL [本文引用: 1] 施肥是退化草地初期恢复常用的管理措施,被用于改良草地群落结构和提高牧草生产力.本文以藏北轻度和重度 退化高寒草甸为研究对象,设置低氮(50kgN·hm-2 ·a-1,LN)、高氮(100 kg N· hm-2·a-1,HN)施肥水平,及其与磷配施组合试验(50 kg N·hm-2·a-1+50kgP·hm-2·a-1,LN+P; 100kgN ·hm-2·a-1+50kgP ·hm-2·a-1,HN+P),研究施肥梯度与施肥类型对两种退化程度高寒草甸群落结构、物种丰富度与多样性、群落生物量及其分配的影响.结果表明:单 独施氮对轻度和重度退化高寒草甸群落盖度和生物量均无显著影响,而氮磷配施显著提高群落盖度及地上、地下生物量,其中LN+P处理的促进效果最显著.这表 明本研究区地上生产力受到氮和磷的共同限制.施肥处理对重度退化样地总生物量无显著影响,但显著降低了根冠比.HN与HN+P处理均显著降低轻度退化群落 的物种丰富度和均匀度,说明HN处理不利于轻度退化草地物种多样性和稳定性的维持.在轻度退化样地,施氮(尤其是HN处理)主要提高了禾草植物的重要值和 生物量,而氮磷配施则有利于莎草类植物的生长.施肥对重度退化群落植物的重要值影响较小,主要提高了杂草生物量.说明轻度退化高寒草甸可选择低氮配施磷肥 的措施,而对重度退化草地可能需要结合围栏和补播牧草等其他管理措施进行改良.
[43] Zou XY Dong GR Li S Dong YX Yang P Ji HL Zhang CL Cheng H 2003 ). Desertification and its prevention and control strategy in TibetJournal of Natural Disasters, 17-24. DOI:
10.3969/j.issn.1004-4574.2003.01.003 URL [本文引用: 1] 西藏自治区荒漠化研究始于20世纪90年代初,但近10年来在探 索荒漠化面积、类型、分布、危害、动态监测和防治等方面都取得了重大进展.全区荒漠化土地面积20474114hm2,潜在荒漠化土地面积 1365180hm2.从影响荒漠化进程的多种因素的发展趋势来看,目前荒漠化仍处于发展阶段.为了有效地促进西藏自治区的经济发展,改善生态环境,西藏 自治区政府于1997年制定了详细的防治荒漠化计划,制定了"保护优先、积极治理、适度开发"的防治战略.按该计划,在2020年前共治理荒漠化土地面积 5243870hm2.考虑到西藏国民经济发展战略的总体布局,防治荒漠化在布局上将以"一江两河"中部流域为重点,兼顾其他地区.达到上述目标,将能初 步实现西藏社会经济与自然环境的协调发展.
[ 邹学勇 , 董光荣 , 李森 , 董玉祥 , 杨萍 , 靳鹤龄 , 张春来 , 程宏 ( 2003 ). 西藏荒漠化及其防治战略自然灾害学报 , 12, 17 -24 .] DOI:
10.3969/j.issn.1004-4574.2003.01.003 URL [本文引用: 1] 西藏自治区荒漠化研究始于20世纪90年代初,但近10年来在探 索荒漠化面积、类型、分布、危害、动态监测和防治等方面都取得了重大进展.全区荒漠化土地面积20474114hm2,潜在荒漠化土地面积 1365180hm2.从影响荒漠化进程的多种因素的发展趋势来看,目前荒漠化仍处于发展阶段.为了有效地促进西藏自治区的经济发展,改善生态环境,西藏 自治区政府于1997年制定了详细的防治荒漠化计划,制定了"保护优先、积极治理、适度开发"的防治战略.按该计划,在2020年前共治理荒漠化土地面积 5243870hm2.考虑到西藏国民经济发展战略的总体布局,防治荒漠化在布局上将以"一江两河"中部流域为重点,兼顾其他地区.达到上述目标,将能初 步实现西藏社会经济与自然环境的协调发展.
[44] Zuo XA Zhao XY Zhao HL Li YQ Guo ZR Zhao YP 2007 ). Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin Sand LandEnvironmental Science, 28, 945 -951 . DOI:
10.3321/j.issn:0250-3301.2007.05.003 URL [本文引用: 3] 研究了科尔沁沙地6个典型生境类型的沙质草地群落物种多样性与生产力的变化,分析了植物群落格局、物种多样性、生产力与土壤特性的关系.结果表明,从湿草甸向干草甸、固定沙丘、半固定沙丘、半流动沙丘和流动沙丘退化过程中,群落生产力逐渐下降;群落物种多样性先增加后减小,表现出由湿生化和土壤贫瘠化生境向中生、中旱生生境逐渐增加的趋势;土壤极细沙和粉粒含量逐渐递减,土壤有机碳和全氮含量、电导率逐渐递减.典范对应分析(CCA)表明,土壤有机碳、全氮、有效氮、有效钾、土壤含水量、酸碱度和盐分含量的变化共同影响植物群落分布格局,其解释总方差为40%,其中土壤养分梯度是沙质草地群落分布格局的主要土壤限制因子.沙质草地植物群落的生态优势度、物种多样性指数分别与土壤养分梯度和水盐及酸碱因子二元指标之间存在显著的二元线性关系.沙质草地群落物种多样性变化受土壤养分、水盐及酸碱度因子的共同影响.多元回归模型分析表明,土壤养分对生物量的贡献率为86.73%,明显大于水盐及酸碱度对群落生产力的影响.
[ 左小安 , 赵学勇 , 赵哈林 , 李玉强 , 郭轶瑞 , 赵玉萍 ( 2007 ). 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系环境科学 , 28, 945 -951 .] DOI:
10.3321/j.issn:0250-3301.2007.05.003 URL [本文引用: 3] 研究了科尔沁沙地6个典型生境类型的沙质草地群落物种多样性与生产力的变化,分析了植物群落格局、物种多样性、生产力与土壤特性的关系.结果表明,从湿草甸向干草甸、固定沙丘、半固定沙丘、半流动沙丘和流动沙丘退化过程中,群落生产力逐渐下降;群落物种多样性先增加后减小,表现出由湿生化和土壤贫瘠化生境向中生、中旱生生境逐渐增加的趋势;土壤极细沙和粉粒含量逐渐递减,土壤有机碳和全氮含量、电导率逐渐递减.典范对应分析(CCA)表明,土壤有机碳、全氮、有效氮、有效钾、土壤含水量、酸碱度和盐分含量的变化共同影响植物群落分布格局,其解释总方差为40%,其中土壤养分梯度是沙质草地群落分布格局的主要土壤限制因子.沙质草地植物群落的生态优势度、物种多样性指数分别与土壤养分梯度和水盐及酸碱因子二元指标之间存在显著的二元线性关系.沙质草地群落物种多样性变化受土壤养分、水盐及酸碱度因子的共同影响.多元回归模型分析表明,土壤养分对生物量的贡献率为86.73%,明显大于水盐及酸碱度对群落生产力的影响.
锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
2000
锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
2000
藏北高寒草原草地退化及其驱动力分析
2
2007
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
... )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
藏北高寒草原草地退化及其驱动力分析
2
2007
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
... )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
高寒地区不同退化程度草地群落结构特征研究
1
2016
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
高寒地区不同退化程度草地群落结构特征研究
1
2016
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
我国草地的发展观
1
2005
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
我国草地的发展观
1
2005
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
放牧对草原土壤的影响
1
2004
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
放牧对草原土壤的影响
1
2004
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
四种牧草幼苗对水分胁迫的相应及其抗旱性
1
2005
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
四种牧草幼苗对水分胁迫的相应及其抗旱性
1
2005
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
生物多样性与生态系统生产力: 为什么野外观测和受控实验结果不一致?
1
2003
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
生物多样性与生态系统生产力: 为什么野外观测和受控实验结果不一致?
1
2003
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
结构方程模型的应用及分析策略
1
1999
... id="C35">结构等式模型假定一组隐变量之间存在相关关系, 隐变量是几个显变量的线性组合(
侯杰泰和成子娟, 1999 ).通过检验显变量之间的相关系数或协方差, 进而估计模型中的通径系数, 因此可以在统计上检验所假设的模型对所研究的过程是否合适. ...
结构方程模型的应用及分析策略
1
1999
... id="C35">结构等式模型假定一组隐变量之间存在相关关系, 隐变量是几个显变量的线性组合(
侯杰泰和成子娟, 1999 ).通过检验显变量之间的相关系数或协方差, 进而估计模型中的通径系数, 因此可以在统计上检验所假设的模型对所研究的过程是否合适. ...
应用Excel软件计算生物多样性指数
2012
应用Excel软件计算生物多样性指数
2012
退化草地植物功能群和物种丰富度与群落生产力关系的研究
2
2011
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
退化草地植物功能群和物种丰富度与群落生产力关系的研究
2
2011
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
青藏高原环境保护对策
1
2000
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
青藏高原环境保护对策
1
2000
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
放牧家畜的践踏强度指标探讨
1
2008
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
放牧家畜的践踏强度指标探讨
1
2008
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
高寒草甸“黑土型”退化草地的成因及生态过程
1
1999
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
高寒草甸“黑土型”退化草地的成因及生态过程
1
1999
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究,
3
2008
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C68">物种多样性不仅能够度量群落的组成结构和功能的复杂性, 而且也能指示环境状况, 例如土壤养分等(
罗亚勇等, 2014 ).研究结果表明, 在高寒草甸退化过程中群落多样性呈现出非线性增加趋势, 该结果与仁青吉等(2008)的研究结果一致.研究进一步发现, 均匀度指数和多样性指数在重度退化梯度上达到最大, 物种丰富度指数在极度退化梯度上达到最大, 与之前的研究结果(
柳小妮等, 2008 )相似.造成这种差异的主要原因是群落种类数目和种类之间个体分配的均匀性, 过度放牧抑制了优势物种的竞争能力, 导致处于竞争弱势物种的入侵和定居成为可能, 在重度、极度退化阶段物种数增幅较大, 因此物种丰富度显著增加.同时在重度和极度退化阶段, 优势种对其他物种的抑制作用明显减弱, 表现为优势种高山嵩草盖度逐渐降低, 矮火绒草盖度逐渐增加, 因此在重度退化梯度上, 高山嵩草、委陵菜属和矮火绒草在群落中达到相对平衡, 导致均匀度在重度退化阶段最高. ...
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究,
3
2008
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C68">物种多样性不仅能够度量群落的组成结构和功能的复杂性, 而且也能指示环境状况, 例如土壤养分等(
罗亚勇等, 2014 ).研究结果表明, 在高寒草甸退化过程中群落多样性呈现出非线性增加趋势, 该结果与仁青吉等(2008)的研究结果一致.研究进一步发现, 均匀度指数和多样性指数在重度退化梯度上达到最大, 物种丰富度指数在极度退化梯度上达到最大, 与之前的研究结果(
柳小妮等, 2008 )相似.造成这种差异的主要原因是群落种类数目和种类之间个体分配的均匀性, 过度放牧抑制了优势物种的竞争能力, 导致处于竞争弱势物种的入侵和定居成为可能, 在重度、极度退化阶段物种数增幅较大, 因此物种丰富度显著增加.同时在重度和极度退化阶段, 优势种对其他物种的抑制作用明显减弱, 表现为优势种高山嵩草盖度逐渐降低, 矮火绒草盖度逐渐增加, 因此在重度退化梯度上, 高山嵩草、委陵菜属和矮火绒草在群落中达到相对平衡, 导致均匀度在重度退化阶段最高. ...
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
不同封育年限高寒草甸植被/土壤碳密度及净生态系统CO
2 交换量的比较
1
2013
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
不同封育年限高寒草甸植被/土壤碳密度及净生态系统CO
2 交换量的比较
1
2013
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
高寒草甸主要植物地上地下生物量分布及退化对根冠比和根系表面积的影响
2
2015
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
高寒草甸主要植物地上地下生物量分布及退化对根冠比和根系表面积的影响
2
2015
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系
12
2014
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
... ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C68">物种多样性不仅能够度量群落的组成结构和功能的复杂性, 而且也能指示环境状况, 例如土壤养分等(
罗亚勇等, 2014 ).研究结果表明, 在高寒草甸退化过程中群落多样性呈现出非线性增加趋势, 该结果与仁青吉等(2008)的研究结果一致.研究进一步发现, 均匀度指数和多样性指数在重度退化梯度上达到最大, 物种丰富度指数在极度退化梯度上达到最大, 与之前的研究结果(
柳小妮等, 2008 )相似.造成这种差异的主要原因是群落种类数目和种类之间个体分配的均匀性, 过度放牧抑制了优势物种的竞争能力, 导致处于竞争弱势物种的入侵和定居成为可能, 在重度、极度退化阶段物种数增幅较大, 因此物种丰富度显著增加.同时在重度和极度退化阶段, 优势种对其他物种的抑制作用明显减弱, 表现为优势种高山嵩草盖度逐渐降低, 矮火绒草盖度逐渐增加, 因此在重度退化梯度上, 高山嵩草、委陵菜属和矮火绒草在群落中达到相对平衡, 导致均匀度在重度退化阶段最高. ...
... id="C78">熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法.值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003).本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降. ...
青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系
12
2014
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
... ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C68">物种多样性不仅能够度量群落的组成结构和功能的复杂性, 而且也能指示环境状况, 例如土壤养分等(
罗亚勇等, 2014 ).研究结果表明, 在高寒草甸退化过程中群落多样性呈现出非线性增加趋势, 该结果与仁青吉等(2008)的研究结果一致.研究进一步发现, 均匀度指数和多样性指数在重度退化梯度上达到最大, 物种丰富度指数在极度退化梯度上达到最大, 与之前的研究结果(
柳小妮等, 2008 )相似.造成这种差异的主要原因是群落种类数目和种类之间个体分配的均匀性, 过度放牧抑制了优势物种的竞争能力, 导致处于竞争弱势物种的入侵和定居成为可能, 在重度、极度退化阶段物种数增幅较大, 因此物种丰富度显著增加.同时在重度和极度退化阶段, 优势种对其他物种的抑制作用明显减弱, 表现为优势种高山嵩草盖度逐渐降低, 矮火绒草盖度逐渐增加, 因此在重度退化梯度上, 高山嵩草、委陵菜属和矮火绒草在群落中达到相对平衡, 导致均匀度在重度退化阶段最高. ...
... id="C78">熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法.值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003).本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降. ...
不同退化阶段高寒草甸土壤化学计量特征
1
2012
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
不同退化阶段高寒草甸土壤化学计量特征
1
2012
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
藏北那曲地区草地退化动态评价
3
2008
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
... id="C75">其中频次分析结果表明: 应用频次最高的4个指标分别为植被盖度、有机质含量、经济收入水平及坡度, 频次分别为116、48、43和42 (附件
表2 ; 邱秋兰, 2012).在以上分析基础上, 参考以往相关研究(
周华坤等, 2003 ;
毛飞等, 2008 ), 并结合本研究的数据获取情况, 本文在评价藏北高寒草甸退化过程中选取的评价指标为: 群落总盖度、高山嵩草盖度、土壤碳氮比、地上生物量、地下生物量及土壤砾石比等6个指标. ...
... id="C78">熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法.值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003).本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降. ...
藏北那曲地区草地退化动态评价
3
2008
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
... id="C75">其中频次分析结果表明: 应用频次最高的4个指标分别为植被盖度、有机质含量、经济收入水平及坡度, 频次分别为116、48、43和42 (附件
表2 ; 邱秋兰, 2012).在以上分析基础上, 参考以往相关研究(
周华坤等, 2003 ;
毛飞等, 2008 ), 并结合本研究的数据获取情况, 本文在评价藏北高寒草甸退化过程中选取的评价指标为: 群落总盖度、高山嵩草盖度、土壤碳氮比、地上生物量、地下生物量及土壤砾石比等6个指标. ...
... id="C78">熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法.值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003).本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降. ...
The role of biomass allocation strategy in diversity loss due to fertilization
1
2008
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
退化高寒干旱草地植物群落多样性特征
1
2005
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
退化高寒干旱草地植物群落多样性特征
1
2005
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
放牧对高寒草甸植物群落结构及生产力的影响
6
2008
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
... ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
放牧对高寒草甸植物群落结构及生产力的影响
6
2008
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
... ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
Grazing effects upon plant community structure in subhumid grasslands of Argentina
1
1986
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
果洛大武地区不同退化梯度下高寒草甸土壤养分特征变化
1
2010
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
果洛大武地区不同退化梯度下高寒草甸土壤养分特征变化
1
2010
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
西藏自治区农牧业特色产业发展研究
1
2003
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
西藏自治区农牧业特色产业发展研究
1
2003
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应
1
2008
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应
1
2008
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
不同放牧密度对草原植被特征的影响分析
1
1996
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
不同放牧密度对草原植被特征的影响分析
1
1996
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
黄河源区生态环境变化与成因分析
1
2000
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
黄河源区生态环境变化与成因分析
1
2000
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
放牧对高寒嵩草草甸土壤微生物量碳的影响及其与土壤环境的关系
1
2008
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
放牧对高寒嵩草草甸土壤微生物量碳的影响及其与土壤环境的关系
1
2008
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
内蒙古草原退化群落恢复演替的研究: II.恢复演替时间进程的分析
2
1996
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
内蒙古草原退化群落恢复演替的研究: II.恢复演替时间进程的分析
2
1996
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C69">随着退化程度的加剧, 群落多样性指数非线性增加, 群落总盖度及高山嵩草盖度非线性降低, 矮火绒草盖度非线性增加, 委陵菜属则没有显著变化.植物群落的结构特征通常以优势种和其他主要物种组成为特征, 优势种的更替可成为植物群落演替的重要表现形式 (
Sala et al ., 1986 ;
王炜等, 1996 ), 在退化过程中, 高寒草甸植被群落逆向演替表现为原优势种高山嵩草在群落竞争中逐步丧失优势地位, 成为群落的伴生种, 群落向杂类草草地过渡, 最终形成毒杂草占优势的极度退化草地植被类型.这主要是因为: 1)过度放牧对高寒草甸退化的影响是一个综合因素, 其中包括动物的采食和践踏(
林慧龙等, 2008 ), 以及由此引起的土壤理化状况的改变(
高英志等, 2004 ;
王启兰等, 2008 ), 受到干扰后的土壤表面和原有植物的适度破坏为先锋种提供了小生境, 从而使得新的植物侵入群落, 提高了植物的丰富度(
王长庭等, 2008 ); 2)高寒草甸植物的竞争主要是光竞争和营养竞争, 其中竞争力变化较大的是物种对光资源的获取能力(
仁青吉等, 2008 ), 由于放牧(尤其是过度放牧)使得高山嵩草被优先采食, 导致高山嵩草的盖度降低, 进而降低整个群落的光照闭合度, 使光资源不再是影响物种生长和生存的限制因子, 光资源的释放使弱势物种在对光资源的竞争过程中受益, 被排斥的物种又重新占据原来的生态位, 使得物种丰富度和多度增加, 主要表现为杂类草的增加(
仁青吉等, 2008 ), 例如, 本研究中, 随着退化程度不断加剧, 高山嵩草盖度不断降低, 矮火绒草盖度不断增加, 到极度退化, 矮火绒草取代高山嵩草占领其生态位; 3)由于放牧强度的增大导致高寒草场储存在土壤库中的氮磷元素不断输出,降低土壤肥力和生产能力, 进而加速了植物群落的演替(
柳小妮等, 2008 ).我们的研究结果表明, 相对于土壤物理性质, 土壤营养状况是影响高寒草甸退化过程中物种多样性的重要因素, 该结论与罗亚勇等(2014)和白永飞等(2000)的研究结果较一致.由于高寒草甸退化导致土壤C、N含量非线性降低, 而我们的结果发现土壤营养状况是影响群落多样性的重要因素, 因此高寒草甸退化过程中土壤养分的非线性响应可能导致群落多样性的非线性响应. ...
内蒙古草原过度放牧的解决途径
2
2007
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
内蒙古草原过度放牧的解决途径
2
2007
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
1
2010
... id="C42">本文选取
χ 2 、绝对适配度指数(
RMSEA, <0.05)、增值适配度指数(
NFI, >0.9)和最小信息准则(
AIC )(理论模型的
AIC 值要小于独立模型和饱和模型的
AIC 值)衡量SEM的拟合优度(
吴明隆, 2010 ). ...
1
2010
... id="C42">本文选取
χ 2 、绝对适配度指数(
RMSEA, <0.05)、增值适配度指数(
NFI, >0.9)和最小信息准则(
AIC )(理论模型的
AIC 值要小于独立模型和饱和模型的
AIC 值)衡量SEM的拟合优度(
吴明隆, 2010 ). ...
藏北高寒退化草地植被恢复过程的障碍因子初探
1
2003
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
藏北高寒退化草地植被恢复过程的障碍因子初探
1
2003
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
青南高原气候变化及其对高寒草甸牧草生长影响的研究
1
1999
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
青南高原气候变化及其对高寒草甸牧草生长影响的研究
1
1999
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
Community succession along a chronosequence of vegetation restoration on sand dunes in Horqin Sandy Land
1
2005
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
西藏自治区生态环境问题及对策
1
2001
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
西藏自治区生态环境问题及对策
1
2001
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
草地产权制度对草地超载过牧的影响——以肃南县红石窝乡的调查为例
2
2005
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
草地产权制度对草地超载过牧的影响——以肃南县红石窝乡的调查为例
2
2005
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
... id="C40">由于过度放牧使家畜对草地土壤的践踏作用加剧, 造成土壤紧实度增加, 容重上升, 砂粒含量逐渐增加, 因此导致土壤持水力降低, 土壤水分含量减少(
赵成章等, 2005 ;
王云霞和曹建民, 2007 ); 在此过程中土壤pH值升高, 土壤养分逐渐降低(
罗亚勇等, 2014 ), 而土壤理化性质的改变将直接影响植被的生长、发育及演替过程(
贺金生等, 2003 ), 并且土壤理化性质的改变及其相互关系共同决定着高寒草甸生产力的变化.基于以上理论支持, 本文利用结构等式模型(SEM)解析土壤C含量, 碎石质量比, 体积含水量的改变对群落中高山嵩草盖度、矮火绒草盖度和总盖度及地上、地下生物量的直接和间接影响. ...
放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响
1
2012
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响
1
2012
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
国内外学术界一直关注的问题: 青藏高原研究
1
2005
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
国内外学术界一直关注的问题: 青藏高原研究
1
2005
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
青藏高原高寒草甸的植被退化与土壤退化特征研究
8
2005
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C75">其中频次分析结果表明: 应用频次最高的4个指标分别为植被盖度、有机质含量、经济收入水平及坡度, 频次分别为116、48、43和42 (附件
表2 ; 邱秋兰, 2012).在以上分析基础上, 参考以往相关研究(
周华坤等, 2003 ;
毛飞等, 2008 ), 并结合本研究的数据获取情况, 本文在评价藏北高寒草甸退化过程中选取的评价指标为: 群落总盖度、高山嵩草盖度、土壤碳氮比、地上生物量、地下生物量及土壤砾石比等6个指标. ...
... id="C78">熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法.值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003).本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降. ...
青藏高原高寒草甸的植被退化与土壤退化特征研究
8
2005
... id="C4">高寒草甸是青藏高原广泛分布的主要植被类型, 也是我国草地生态系统的重要组成部分.据粗略测算, 青藏高原的高寒草甸面积为70 × 10
4 km
2 , 约占青藏高原可利用草场面积的50%.特殊的地理特征和恶劣的环境条件, 以及草甸资源存在粗放经营、不合理利用和管理混乱等问题, 导致草甸退化现象尤为突出(
王根绪等, 2000 ;
周华坤等, 2005 ;
罗亚勇等, 2012 ;
刘晓琴等, 2013 ).自20世纪80年代以来, 覆盖青藏高原面积37%的高寒草甸原生植被严重退化(
罗亚勇等, 2014 ).以那曲地区为例, 该地区草地退化面积为1.81 × 10
7 hm
2 , 占该地区土地总面积的43.1%, 其中轻度退化面积占退化草地总面积的57.3%, 中度退化面积占38.3%, 重度退化面积占4.4% (
毛飞等, 2008 ). ...
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
... id="C7">群落生产力是衡量群落生态功能的最重要指标, 能够反映群落结构及其功能的变化.在高寒草甸退化过程中, 土壤质地、养分和水分等复杂变化及其相互关系共同调控物种多样性和群落生产力的变化(
罗亚勇等, 2014 ).随着退化程度的加剧, 禾本科及莎草科等优良牧草的地上生物量下降, 杂类草生物量明显升高, 在群落中逐渐占据优势地位, 使草地总生物量不断减少, 草地生产力降低(
仁青吉等, 2008 ;
李里和刘伟, 2011 ;
陈乐乐等, 2016 ), 但是与多样性对退化的响应趋势类似, 罗亚勇等(2014)研究发现, 随着高寒草甸退化程度的加剧, 植被群落地上生物量呈先稳定后降低的趋势, 地上生物量在轻度退化阶段或者中度退化阶段最高, 在极度退化阶段最低(
刘伟等, 1999 ;
周华坤等, 2005 ).然而, 在物种多样性和群落生产力对退化的响应过程中, 多数研究定量比较不同退化阶段的群落生物量之间的差异, 可能不能充分揭示生产力对退化的具体响应模式. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C67">与地上生物量对退化的响应不同, 随着退化程度的不断加剧, 地下生物量非线性降低, 与之前研究(
周华坤等, 2005 ;
罗亚勇等, 2014 )结论一致.在水分胁迫的情况下, 植物的叶片含水量降低得越多, 表明该物种受干旱胁迫就越强, 抗旱能力越弱(
谷燕蓉等, 2005 ), 研究发现在受到干旱胁迫时, 高山嵩草的叶片相对含水量较低导致其抗旱能力较弱, 因此在高寒草甸退化过程中, 土壤含水量不断降低, 致使高山嵩草盖度逐渐降低.与此同时, 在退化过程中土壤pH值不断增加, 土壤养分含量明显降低(
罗亚勇等, 2014 ), 因此随着高寒草甸退化的不断加剧, 土壤C含量的显著降低同样会导致高山嵩草盖度不断下降.研究发现高寒草甸优势物种高山嵩草属于密丛须根型多年生植物, 可以形成庞大的地下生物量(
柳小妮等, 2008 ), 其根冠比接近70 (
龙毅等, 2015 ), 地下生物量约为委陵菜地下生物量的60倍(
龙毅等, 2015 ), 因此高山嵩草对高寒草甸退化的响应模式在一定程度上决定地下生物量的变化趋势.随着退化程度不断加剧, 群落中高山嵩草的比例逐渐下降, 地下草毡层遭到破坏, 地下生物量迅速降低, 即使矮火绒草的盖度不断增加, 但其属于轴根型植物, 其数量的增加对地下生物量的贡献较低(
王炜等, 1996 ), 并且在此过程中伴随着土壤砾石含量增加、土壤持水力降低, 导致土壤含水量降低, 进一步加剧地下生物量的降低, 最终这些直接和间接因素导致在退化过程中地下生物量的非线性降低.由于高寒草甸总生物量中, 地下生物量占据主导地位, 因此总生物量的变化主要受地下生物量变异的影响. ...
... id="C75">其中频次分析结果表明: 应用频次最高的4个指标分别为植被盖度、有机质含量、经济收入水平及坡度, 频次分别为116、48、43和42 (附件
表2 ; 邱秋兰, 2012).在以上分析基础上, 参考以往相关研究(
周华坤等, 2003 ;
毛飞等, 2008 ), 并结合本研究的数据获取情况, 本文在评价藏北高寒草甸退化过程中选取的评价指标为: 群落总盖度、高山嵩草盖度、土壤碳氮比、地上生物量、地下生物量及土壤砾石比等6个指标. ...
... id="C78">熵权法是一种客观赋权法, 它充分利用了原始数据提供的信息, 在实践应用中是一种简单可行的科学方法.值得注意的是, 由于计算中要用到对数函数, 对数据有一定的限制, 要求各项数据大于0 (肖香玉, 2003).本文在借鉴以往研究成果基础上将高寒草甸退化程度分为轻度退化、中度退化、重度退化、极度退化(
周华坤等, 2005 ; 沈渭寿等, 2006;
毛飞等, 2008 ;
罗亚勇等, 2014 ), 结合藏北高寒草甸退化的分级及综合评价的指标选取情况, 依据熵权法综合评价得出不同退化程度的得分, 由对照处理到极度退化处理评价得分分别是70.52、62.14、45.24、29.4和22.55,可以发现随着退化程度的不断加深综合评价得分持续下降. ...
实验增温对藏北高寒草甸植物物候的影响
2
2016
... id="C9">研究区域位于藏北高原核心地带, 那曲地区那曲县那曲镇曲果仁毛村境内的藏北高原草地生态系统研究站(那曲站)(31.64° N, 92.02° E, 海拔4 585 m), 隶属于中国科学院地理科学与资源研究所.那曲县属于高原亚寒带季风半湿润气候区.全年大风日100天左右, 平均风速2.7 m·s
-1 , 最大风速可达26.3 m·s
-1 ; 年平均气温-1.16 ℃, 全年日照时间2 788 h; 年降水量430 mm, 集中在6-9月, 占全年降水量的85% (数据来源于中国气象科学数据共享服务网那曲站点).全年无绝对无霜期, 每年10月至次年5月为风雪期和土壤冻结期(
朱军涛, 2016 ).植被类型为典型的高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 常见伴生种有钉柱委陵菜(
Potentilla saundersiana )、无茎黄鹌菜(
Youngia simulatrix )、藏西风毛菊(
Saussurea stoliczkai )、矮羊茅(
Festuca coelestis )、楔叶委陵菜(
Potentilla cuneata )、矮火绒草(
Leontopodium nanum )等.土壤类型为高寒草甸土(
朱军涛, 2016 ). ...
... )等.土壤类型为高寒草甸土(
朱军涛, 2016 ). ...
实验增温对藏北高寒草甸植物物候的影响
2
2016
... id="C9">研究区域位于藏北高原核心地带, 那曲地区那曲县那曲镇曲果仁毛村境内的藏北高原草地生态系统研究站(那曲站)(31.64° N, 92.02° E, 海拔4 585 m), 隶属于中国科学院地理科学与资源研究所.那曲县属于高原亚寒带季风半湿润气候区.全年大风日100天左右, 平均风速2.7 m·s
-1 , 最大风速可达26.3 m·s
-1 ; 年平均气温-1.16 ℃, 全年日照时间2 788 h; 年降水量430 mm, 集中在6-9月, 占全年降水量的85% (数据来源于中国气象科学数据共享服务网那曲站点).全年无绝对无霜期, 每年10月至次年5月为风雪期和土壤冻结期(
朱军涛, 2016 ).植被类型为典型的高寒草甸, 优势种为高山嵩草(
Kobresia pygmaea ), 常见伴生种有钉柱委陵菜(
Potentilla saundersiana )、无茎黄鹌菜(
Youngia simulatrix )、藏西风毛菊(
Saussurea stoliczkai )、矮羊茅(
Festuca coelestis )、楔叶委陵菜(
Potentilla cuneata )、矮火绒草(
Leontopodium nanum )等.土壤类型为高寒草甸土(
朱军涛, 2016 ). ...
... )等.土壤类型为高寒草甸土(
朱军涛, 2016 ). ...
氮磷配施对藏北退化高寒草甸群落结构和生产力的影响
1
2010
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
氮磷配施对藏北退化高寒草甸群落结构和生产力的影响
1
2010
... id="C6">草地退化是一个复杂的生态过程, 虽然关于退化的原因和机理在不同区域和不同草地类型中存在一定争议(
王德利等, 1996 ;
张国胜和李林, 1999 ), 但是有关退化引起物种组成及生产力发生变化的结论是一致的(
李里和刘伟, 2011 ).草地退化程度越严重, 其群落结构越简单, 原群落中的优势物种逐渐在竞争中被适口性较差的一些物种取代, 导致优良牧草数量减少, 盖度降低, 而杂类草等适口性差的物种数量逐渐增加, 群落的多样性、丰富度和均匀度逐渐降低(
祁彪等, 2005 ;
周华坤等, 2005 ;
宗宁等, 2010 ;
罗亚勇等, 2014 ), 但随着退化程度加剧, 物种多样性不一定总是降低, 也可能增加(
仁青吉等, 2008 ).即使不考虑物种多样性对退化的正负响应, 也可能存在其他响应模式, 例如研究表明, 中度退化草地的物种多样性最高(
周华坤等, 2005 ;
郑伟等, 2012 ), 物种多样性对退化的响应呈现先稳定后降低的趋势(
罗亚勇等, 2014 ), 因此在物种多样性对退化的响应过程中可能存在非线性的响应模式. ...
西藏荒漠化及其防治战略
1
2003
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
西藏荒漠化及其防治战略
1
2003
... id="C5">近些年在全球气候变化和人类活动加剧的背景下, 青藏高原高寒草甸面临着严峻的退化问题(盛丽和王彦龙2010).与全球气候变干、变暖的趋势相同, 青藏高原气候面临着干暖趋势(
杨富裕等, 2003 ), 由此引发的负面影响更为突出(
蔡晓布等, 2007 ), 例如气温增幅大、降水量持续降低(
邹学勇等, 2003 )、冻土层变薄(
钟祥浩, 2005 )等.气候变化引起的不利因素与大风等因素相结合加剧了土壤水蒸发, 尤其在地表物质疏松、土壤团粒结构和有机质含量低的条件下, 更易引发土壤风蚀, 加剧土地沙漠化、盐碱化, 进而严重抑制草地植被的生长(
蔡晓布等, 2007 ), 并且高寒草原植被相对比较稀疏、根系较浅, 导致这种状况进一步恶化(
盛丽和王彦龙, 2010 ).但是相对于气候变化对青藏高原草地退化的影响, 人类活动的影响更加严重(
李明森, 2000 ).过度放牧被认为是导致草地退化最重要的人为因素, 畜牧业是藏北高原支柱型产业, 近些年发展速度不断加快, 规模不断扩大, 已远远超过天然草地的承载能力(
张建平等, 2001 ).过度放牧与草地不合理利用、鼠害等因素共同导致土壤退化、沙化, 而且由于草地土壤养分大量输出, 土壤容重、肥力、植被的种类和数目发生明显变化(
董孝斌和张新时, 2005 ;
赵成章等, 2005 ;
王云霞和曹建民, 2007 ).研究发现畜种、畜群的构成中存在严重的不合理性, 饲草在季节间存在不平衡, 这些因素将进一步加剧过度放牧对草地生态系统的负面影响(
西藏自治区农牧业特色产业发展研究课题组, 2003 ), 导致人为因素对草地退化的影响更大. ...
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
3
2007
... id="C21">通过选用物种丰富度指数(
C )、辛普森指数(
D )、香农多样性指数(
H )和均匀度指数(
J ) 4类多样性指数来计算群落的物种多样性(
左小安等, 2007 ), 上述4种指数的计算均使用孔凡洲等(2012)的计算方法. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
3
2007
... id="C21">通过选用物种丰富度指数(
C )、辛普森指数(
D )、香农多样性指数(
H )和均匀度指数(
J ) 4类多样性指数来计算群落的物种多样性(
左小安等, 2007 ), 上述4种指数的计算均使用孔凡洲等(2012)的计算方法. ...
... id="C65">本研究利用空间上的多退化梯度, 探究群落生产力和物种多样性在退化过程中的动态响应.研究结果表明, 群落生产力与物种多样性在高寒草甸退化过程中存在非线性响应模式, 这个重要的变化模式在以往研究中少有报道(
周华坤等, 2005 ;
左小安等, 2007 ;
仁青吉等, 2008 ;
罗亚勇等, 2014 ). ...
... id="C66">我们的研究结果表明, 随着退化程度的不断加剧, 地上总生物量非线性增加, 该结果与之前的研究结果(
仁青吉等, 2008 ;
罗亚勇等, 2014 )相悖.由于群落中主要物种对干扰环境变化的响应对策存在差异, 引起群落组成发生变化(
Niu et al ., 2008 ), 进而影响植被过程和生态系统功能.在高寒草甸退化过程中, 草地生境的改变引起杂类草和毒草的入侵(
Zhang et al ., 2005 ), 导致草地植物功能群发生替代, 优势物种逐渐减少, 杂类草和毒草不断增加, 并且毒草和杂类草具有较高的地上生物量(
罗亚勇等, 2014 ).我们的研究结果显示, 矮火绒草生物量非线性增加, 高山嵩草地上生物量非线性降低, 委陵菜属地上生物量没有显著变化, 在重度退化到极度退化阶段, 矮火绒草地上生物量占地上总生物量的比重更高, 导致地上生物量非线性增加.但是, 我们的结果未发现土壤理化性质对地上生物量有直接或者间接的影响, 一方面可能是由于高寒草甸在退化过程中改变群落中物种组成, 杂类草和毒草比重不断增加, 而杂类草和毒草资源比率的大小和空间异质性的高低, 严重制约着物种之间竞争的激烈程度, 从而影响群落生产力的变化(
左小安等, 2007 ); 另一方面, 可能不同物种对土壤养分和土壤水分的响应不同, 导致物种间的此消彼长, 最终导致地上生物量对土壤理化性质变化不敏感. ...
 ,
, ,1,*, 李军祥5, 刘瑶杰1,2, 俎佳星1,2, 丛楠1, 黄珂1, 王荔51
,1,*, 李军祥5, 刘瑶杰1,2, 俎佳星1,2, 丛楠1, 黄珂1, 王荔51 
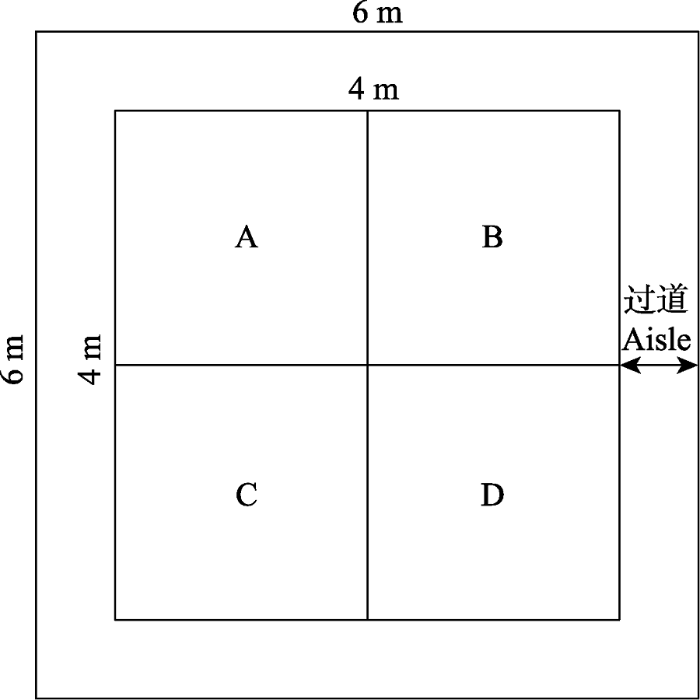 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

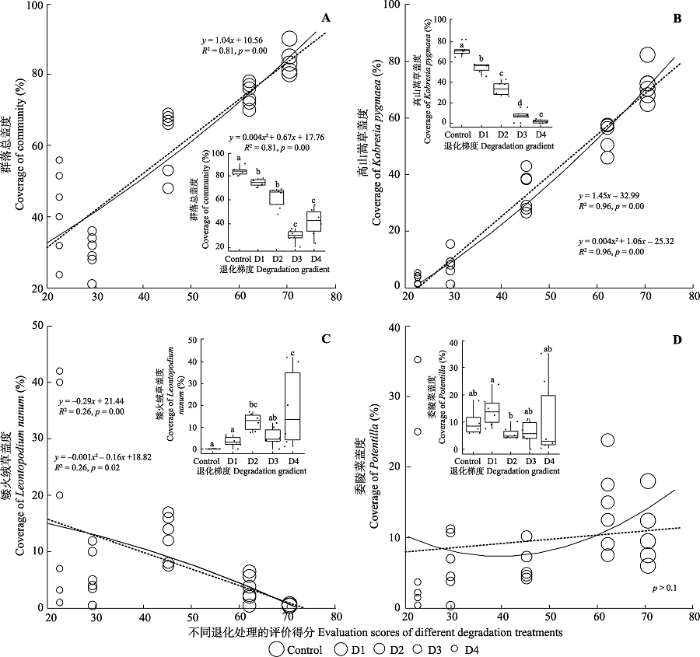 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT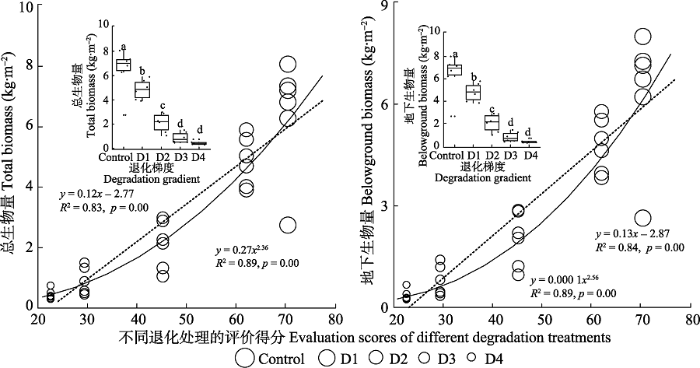 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT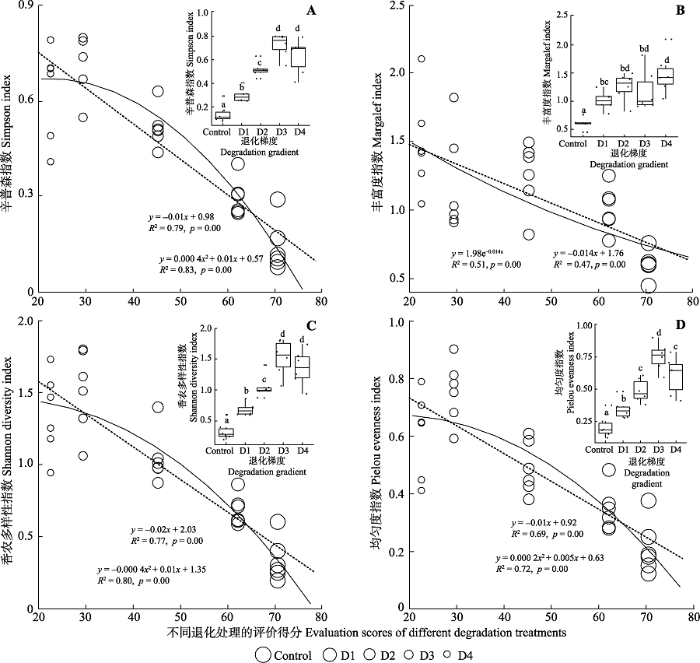 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT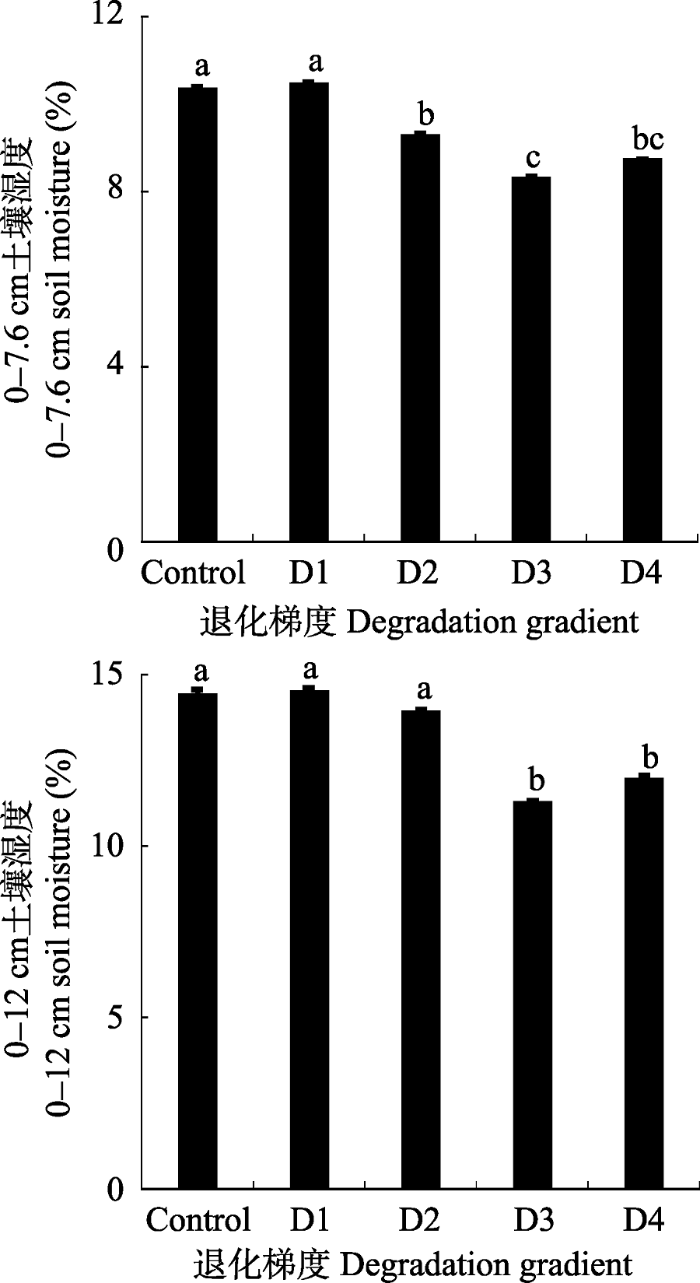 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT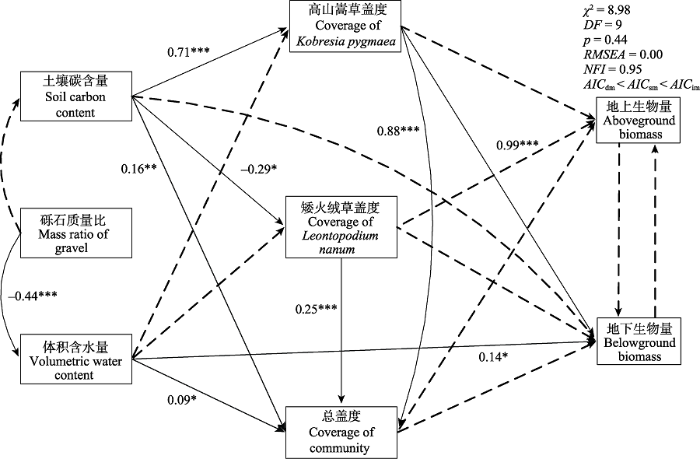 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}