 ,
, ,1,*, 张远东,2,*1
,1,*, 张远东,2,*1 2
Comparison of growth-climate relationship of Sabina przewalskii at different timberlines along a precipitation gradient in the northeast Qinghai-Xizang Plateau, China
SONG Wen-Qi1, ZHU Liang-Jun1, ZHANG Xu1, WANG Xiao-Chun,,1,*, ZHANG Yuan-Dong,2,*1 and 2
通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1528KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
宋文琦, 朱良军, 张旭, 王晓春, 张远东. 青藏高原东北部不同降水梯度下高山林线祁连圆柏径向生长与气候关系的比较. 植物生态学报[J], 2018, 42(1): 66-77 doi:10.17521/cjpe.2017.0251
SONG Wen-Qi.
气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(李博等, 2000; Allen et al., 2013)。探究植被对气候变化的响应是目前生态学研究的热点问题(Sutherland et al., 2013)。高山林线作为森林生态系统中垂直分布于森林上限与灌木林或苔原之间的一条生态过渡带(王庆锁等, 1997), 对气候变化的敏感程度远超过森林主体(Nemani et al., 2003; Holtmeier & Broll, 2005; Smith et al., 2009; Qi et al., 2015), 是研究全球气候变化对植被影响及其适应的理想区域(崔海亭等, 2005)。传统理论认为, 低温是高山林线树木生长的主要限制因子, 树木生长与温度变化显著正相关。随着全球气候变暖, 林线树木生长加快、林线上移, 并且高山林线分布的海拔上限从赤道到极地随纬度升高而逐渐降低(王晓春等, 2004; Harsch et al., 2009; Liang et al., 2014; Feurdean et al., 2016; Piper et al., 2016)。然而, 部分区域林线树木生长对温度敏感性下降或消失, 甚至由正相关转为负相关, 而对降水的敏感性却升高(Wilmking & Myers-Smith, 2008; Solberg et al., 2016), 降水及土壤水分可利用性也可能是限制林线树木生长的主要因子(Lloyd et al., 2002; Elliott & Baker, 2004; Peng et al., 2008)。Leuschner和Schulte (1991)提出低纬度干旱区的高山林线树木生长可能并非由温度控制, 而是由干旱胁迫控制(湿度限制假说), 在低纬度地区的林线研究中既存在水分限制树木生长的案例(Morales et al., 2004; Liang et al., 2014; Piper et al., 2016), 又存在低温限制树木生长的结果(Lü & Zhang, 2013)。因此, 水分在干旱区高山林线树木生长中的作用并不完全清楚。
青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要。青藏高原东北部近50年来年平均气温上升约1.3 ℃ (刘桂芳和卢鹤立, 2010), 平均年降水量也呈上升趋势(胡豪然和梁玲, 2013)。Yang等(2013)发现青藏高原东北部林线树木生长与生长季降水显著正相关, 与气温显著负相关, 并且随着气温升高树木生长对温度的敏感性下降(Gao et al., 2012; Liang et al., 2014)。也有研究表明, 林线树木生长受生长季气温限制作用明显强于降水(郑永宏等, 2008; Liang et al., 2010; 彭剑峰等, 2010; Zhang & Wilmking, 2010)。因此, 青藏高原东北部高山林线树木生长的气候限制因子目前尚不明确。在全球气候变化背景下, 理解并掌握该地区树木径向生长规律及其气候限制机制的空间(尤其是沿水分梯度下)变异性具有重要意义。不同树种间树木径向生长对气候响应可能存在差异(Zimmermann et al., 2015; Rollinson et al., 2016), 因此本研究所用树种仅选择在青藏高原东北部高山林线处广泛自然分布的祁连圆柏(Sabina przewalskii), 该树种对气候变化敏感, 适合进行年轮气候分析(Zhang et al., 2015a; Yang et al., 2017)。对于青藏高原东北部祁连圆柏径向生长与气候关系的研究较多(Zhu et al., 2008; 彭剑峰等, 2010; Yin et al., 2016), 但缺乏不同水分梯度下高山林线树木生长限制因子研究。因此, 本研究在青海东北部, 选择降水量依次增加的3个高山林线样点: 乌兰县哈里哈图国家森林公园(HL)、都兰县曲什岗(QS)、同德县河北林场(HB)。利用树木年轮学方法, 试图验证在干旱、半干旱区高山林线处水分是树木生长的主要限制因子假说, 同时分析水分梯度差异对树木生长与气候关系的影响。研究结果既有助于准确了解青藏高原高山林线树木生长的气候限制因子, 又能为预测全球气候变化情境下植被响应模式和未来高山林线动态提供理论依据。
1 材料和方法
1.1 研究区域概况
本研究区位于青藏高原东北部, 属高原干旱大陆性气候, 研究区内自西北向东南降水量逐渐增加(图1)。采样点位于青海省东部乌兰县哈里哈图国家森林公园(HL)、都兰县曲什岗地区(QS)和同德县河北林场(HB) (图1), 样品均取自各采样点森林分布海拔上限(约4 000 m) (表1)的天然纯林, 取样树种为各采样点内共有且唯一的优势树种——祁连圆柏。林线处森林郁闭度约为0.2, 稀疏生长有祁连圆柏, 无其他伴生树种。林下及林线以上植被均为以短花针茅(Stipa breviflora)、密生薹草(Carex crebra)等低矮草本为优势种的高山草甸。各采样点土壤有机质含量均较低, 不足5%。HL和QS采样点土壤为灰褐土, 土层厚度30-50 cm, 土体干燥至微润, 呈粒状结构且结构疏松, 石砾较多。HB采样点土壤以褐色针叶林土为主, 土层厚度较浅(20-40 cm), 腐殖质含量较低(中国森林编辑委员会, 1999)。图1
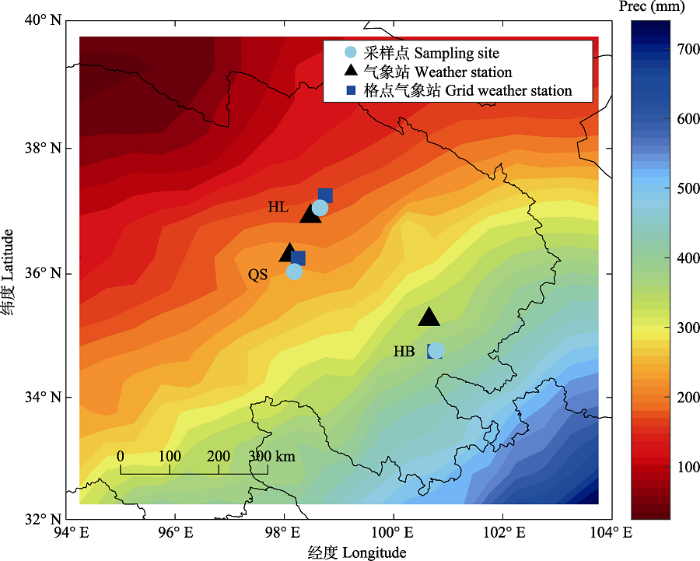 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1研究区域采样点、气象站及格点气象站分布位置。HB, 河北林场; HL, 哈里哈图国家森林公园; QS, 曲什岗地区。Prec, 降水量。
Fig. 1Location of the sampling sites, weather stations and grid weather stations. HB, Hebei Forest Farm; HL, Halihatu National Forest Park; QS, Qushigang; Prec, precipitation.
Table 1
表1
表1祁连圆柏标准年表采样点信息
Table 1
| 采样点 Sampling sites | 纬度 Latitude (N) | 经度 Longitude (E) | 海拔 Altitude (m) | 坡向 Aspect | 坡度 Slope (°) | 年降水量 Annual precipitation (mm) | 年平均气温 Annual mean air temperature (°C) |
|---|---|---|---|---|---|---|---|
| 哈里哈图 Halihatu (HL) | 37.05° | 98.67° | 3β972 | 西 West | 25 | 217 | -2.0 |
| 曲什岗 Qushi hillock (QS) | 36.03° | 98.19° | 4β024 | 东 East | 20 | 281 | -1.8 |
| 河北林场 Hebei forest farm (HB) | 34.76° | 100.79° | 3β791 | 西南 Southwest | 21 | 470 | -1.2 |
新窗口打开|下载CSV
各采样点降水量存在明显不同(图2), 从HL (217 mm)、QS (281 mm)到HB (470 mm)多年平均降水量依次升高, 降水多集中在夏季(6-8月), 分别为138 mm、163 mm和278 mm, 各采样点春季(3-5月)平均降水量与秋季(9-11月)相近。HL、QS和HB采样点气温差异不大, 年平均气温分别为-2.0、-1.8和-1.2 ℃, 7月平均气温分别为9.0、8.5和10.0 ℃, 1月平均气温分别为-16.2、-14.9和-12.4 ℃, 采样点年平均最低气温分别为-10.8、-7.5和-8.5 ℃ (图2)。
图2
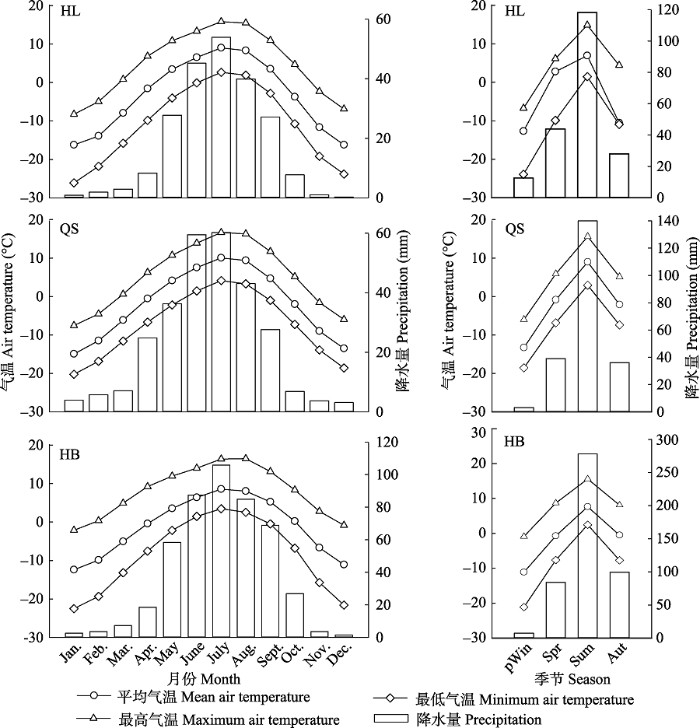 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2各采样点1955-2015年各月和各季节平均气温、最高气温、最低气温和降水量。HB, 河北林场; HL, 哈里哈图国家森林公园; QS, 曲什岗地区。pWin, 上年冬季; Spr, 当年春季; Sum, 当年夏季; Aut, 当年秋季。
Fig. 2Monthly and seasonally mean, maximum, and minimum air temperatures, and precipitation at each sampling site, 1955-2015. HB, Hebei Forest Farm; HL, Halihatu National Forest Park; QS, Qushigang. pWin, last winter; Spr, current spring; Sum, current summer; Aut, current autumn.
1.2 样本采集与树轮年表建立
本研究野外调查及取样工作于2016年7-8月进行, 在HL、QS和HB 3个采样点高山林线处, 以生长于最高处的树木为海拔上限, 各设置3条垂直于林线的30 m × 100 m的矩形样带, 以样带下限左下角为原点, 记录样带内所有树木的坐标和胸径, 利用内径为5.15 mm的生长锥(HaGlof, Dalarna, Sweden)在胸高(约1.3 m)处进行树木年轮取样, 对胸径介于10-20 cm之间的树木一树一芯, 对胸径大于20 cm的树木一树两芯, 胸径低于10 cm的树木不取样芯, HL、QS和HB采样点, 调查树木773株, 共取得636根树木年轮样芯。将取得的样芯编号并置于塑料管中, 带回实验室后对样芯进行固定、风干、打磨等预处理(Stokes & Smiley, 1968)。预处理后的年轮样芯用骨架图定年法, 在双筒显微镜下目视交叉定年。在交叉定年过程中发现, 每个采样点3个样方的树轮(取自各样带)序列间存在高度的一致性, 所以将3个样方内所有树轮样本放到一起进行分析。由于HL、QS和HB采样点内树木年龄相差较大, 所以从HL和QS所有树木年轮样本中挑选出与HB树木年龄相近的样本, 以HB采样点内树龄最高的175年为标准, 定义树龄大于175年的树木为高龄, 低于175的树木为低龄。在HL和QS采样点, 低龄样芯数量分别为28根和16根, 占采样点调查树木总量的9%和6%, 高龄样芯树木数量分别为232根和231根, 占总量的72%和83%, 在HB采样点, 取样芯树木为139株, 占调查总量的81%。用树木年轮宽度量测仪(UniSlide A40Series, Velmex, New York, USA)分别对已完成交叉定年的高龄和低龄样芯进行逐年宽度测量, 测量精度为0.001 mm。为确保定年结果准确无误, 用COFECHA程序进行检验校准, 并在保证样本量的情况下剔除与主序列严重不符及在取样过程中有严重损坏的年轮样芯, 校准后HL采样点内剩余低龄树木样芯20根(占总数的71%), 高龄样芯95根(占43%); QS采样点内剩余低龄树木样芯7根(占44%), 高龄样芯83根(占36%); HB采样点内剩余树木年轮样芯90根(占65%)。为剔除年龄等非气候因素产生的径向生长趋势, 用ARSTAN程序去趋势和建立树轮年表, 去趋势方程采用负指数函数或线性函数拟合方法去趋势(Cook & Holmes, 1984), 本文采用去趋势后获得的标准年表进行随后的年轮-气候分析。1.3 气象数据及统计分析
气象数据从中国气象科学数据共享服务网(为弄清降水梯度下高山林线树木生长在不同气候特征年的气候限制因子是否一致, 用生长季前(1-4月)、生长季(5-8月)平均气温距平和SPEI将整个研究时间段分为生长季前低温年(PLT)、生长季前高温年(PHT)、生长季前干旱年(PD)、生长季前湿润年(PW)、生长季低温年(GLT)、生长季高温年(GHT)、生长季干旱年(GD)、生长季湿润年(GW)。定义平均气温距平小于0 ℃为低温年, 平均气温距平大于0 ℃为高温年, SPEI < 0为干旱年, SPEI > 0为湿润年, 前一年12月到当年2月为冬季, 3-5月为春季, 6-8月为夏季, 9-11月为秋季。年轮指数与气候因子的统计分析用SPSS 20.0完成。
2 结果
2.1 林线祁连圆柏树轮年表特征比较
HL和QS采样点内低龄年表与HB年表时间跨度相近, 高龄年表时间跨度较长(图3)。但是HL和QS采样点内高龄年表与低龄年表公共时间内具有明显的一致性, 变化趋势相近, 拟合度分别为: R2HL = 0.90, R2QS = 0.75 (p < 0.01), 说明标准年表中的年龄趋势去除较为成功。由于在实际取样过程中, HL和QS采样点内可用的低龄树木(树龄小于175年)样本极少, 分别为20个和7个(图3), 所以信噪比(SNR)和样本量的总体解释信号(EPS)数值较低(表2), 不宜用作随后年轮-气候关系的分析, 本文后续分析均采用高龄年表。其他年表SNR和EPS数值均较高, 说明均包含较多的气候信息, 并且年表足以代表采样点内所有树木生长变化的整体特征。HB采样点年表特征值中的MS和SD较HL和QS采样点数值大, 表明HB树木年轮在年际间具有更明显的宽窄变化。各年表的一阶自相关(AC)数值均较大, 说明树木当年的生长可能受前一年气候条件的影响。图3
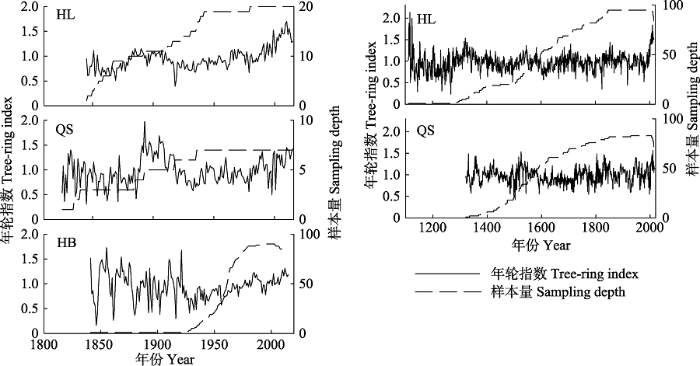 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3林线祁连圆柏树木年轮指数及各年表样本量。左图为低龄年表, 右图为高龄年表。HB, 河北林场; HL, 哈里哈图国家森林公园; QS, 曲什岗地区。
Fig. 3Tree-ring index of Sabina przewalskii in timberline and sampling depth of each chronology. The left and right figures indicate the tree-ring index of young- and old-growth S. przewalskii respectively. HB, Hebei Forest Farm; HL, Halihatu National Forest Park; QS, Qushigang.
Table 2
表2
表2林线祁连圆柏标准年表的主要统计特征
Table 2
| 采样点 Sampling site | 时间跨度 Time span | 标准年表特征值 Eigenvalue of standard chronology | 公共区间统计量 Common interval analysis | |||||
|---|---|---|---|---|---|---|---|---|
| MS | SD | MC | AC | SNR | EPS | VF | ||
| HL (高龄) HL (Old-growth) | 1β112-2β015 | 0.17 | 0.22 | 0.29 | 0.45 | 27.1 | 0.96 | 30.7 |
| HL (低龄) HL (Young-growth) | 1β845-2β015 | 0.13 | 0.22 | 0.51 | 0.76 | 19.6 | 0.95 | 56.6 |
| QS (高龄) QS (Old-growth) | 1β275-2β015 | 0.14 | 0.18 | 0.20 | 0.50 | 15.2 | 0.94 | 25.0 |
| QS (低龄) QS (Young-growth) | 1β825-2β015 | 0.23 | 0.29 | 0.26 | 0.52 | 1.70 | 0.63 | 51.7 |
| HB (低龄) HB (Young-growth) | 1β841-2β015 | 0.24 | 0.27 | 0.29 | 0.48 | 17.6 | 0.95 | 32.7 |
新窗口打开|下载CSV
2.2 不同降水条件下林线祁连圆柏年轮-气候关系的比较
随降水量的增加, 祁连圆柏径向生长与生长季前冬季最低气温由HL、QS的显著正相关(r = 0.29*, r = 0.36*, *和**代表p < 0.05和p < 0.01, 下同)变为HB的不显著负相关(r = -0.03)。径向生长与春季最低气温的关系, 由HL的显著正相关(r = 0.28*)变为QS的不显著正相关(r = 0.04), 及HB的显著负相关(r = -0.30*)。径向生长与夏季最低气温, 由HL的显著正相关(r = 0.49**)变为QS的不显著正相关(r = 0.05), 以及HB的不显著负相关(r = -0.15)。径向生长与秋季最低气温, 由HL的不显著负相关(r = -0.08)变为QS的不显著正相关(r = 0.09), 以及HB的显著负相关(r = -0.30*) (图4)。随着降水量增加, 生长季前及生长季气温对林线祁连圆柏径向生长正效应会逐渐衰弱, 并最终转变为负效应。图4
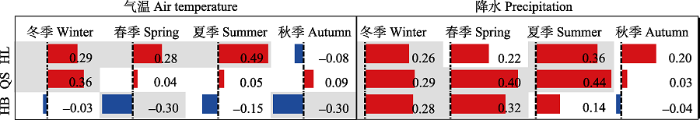 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同降水强度林线祁连圆柏径向生长与季节气候数据的相关性。红色表示正值; 蓝色表示负值; 底色阴影为显著相关(p < 0.05)。HB, 河北林场; HL, 哈里哈图国家森林公园; QS, 曲什岗地区。
Fig. 4Correlation coefficients between the radial growth of Qilian juniper and seasonal climatic data along the precipitation gradient. The red rectangles denote positive correlations. The blue rectangles denote negative correlations. The shaded background denotes significance at p < 0.05 level of Pearson correlation. HB, Hebei Forest Farm; HL, Halihatu National Forest Park; QS, Qushigang.
然而, 祁连圆柏径向生长与春、夏季降水的相关性却随降水变化呈现了不同的变化趋势。径向生长与冬季降水量的关系在3个样点全部显著正相关(HL: r = 0.26*, QS: r = 0.29*, HB: r = 0.28*)。径向生长与春季降水量的关系, 由HL的不显著正相关(r = 0.22)变为QS、HB的显著正相关(r = 0.40*, r = 0.32*)。径向生长与夏季降水量的关系, 由HL、QS的显著正相关(r = 0.36*, r = 0.44*)变为HB的不显著正相关(r = 0.14)。径向生长与秋季降水量的关系在3个样点全不显著(HL: r = 0.20; QS: r = 0.03; HB: r = -0.04) (图4)。
2.3 极端气温及降水年林线祁连圆柏径向生长与气候因子的关系比较
HL采样点, 在生长季前高温年、生长季低温年和高温年, 祁连圆柏径向生长与夏季最低气温均显著正相关(PHT: r = 0.58*, GLT: r = 0.57*, GHT: r = 0.53*)。在所有气候特征年, 祁连圆柏径向生长与夏季降水均显著正相关(图5)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5沿降水梯度祁连圆柏径向生长与不同气温、不同降水年季节气候因子的相关性。红色表示正值; 蓝色表示负值; 底色阴影为显著相关(p < 0.05); GHT, 生长季高温年; GLT, 生长季低温年; PHT, 生长季前高温年; PLT, 生长季前低温年。GD, 生长季干旱年; GW, 生长季湿润年; PD, 生长季前干旱年; PW, 生长季前湿润年。
Fig. 5Correlation coefficients between the radial growth of Qilian juniper and seasonal climate data in different temperature and precipitation years along the precipitation gradient. The red rectangles denote the positive value. The blue rectangles denote the negative value. The shaded background denotes significance at p < 0.05 level of Pearson correlation. GHT, year of growing-season high temperature; GLT, year of growing-season low temperature; PHT, year of pre-season high temperature; PLT, year of pre-season low temperature. GD, year of drought growing season; GW, year of wet growing season; PD, year of drought pre-season; PW, year of wet pre-season. HB, Hebei Forest Farm; HL, Halihatu National Forest Park; QS, Qushigang.
QS采样点, 在生长季前和生长季高温年, 祁连圆柏径向生长与冬季最低气温显著正相关(PHT: r = 0.38*, GHT: r = 0.50*), 其余均相关不显著。在生长季前和生长季高温年、生长季低温年, 祁连圆柏径向生长与春季降水显著正相关(PHT: r = 0.40*, GLT: r = 0.45*, GHT: r = 0.39*)。与夏季降水的相关性同HL采样点一样, 在所有气候特征年, 祁连圆柏径向生长与夏季降水均显著正相关(图5)。
HB采样点, 在不同气温年祁连圆柏径向生长与气候关系的差异较大。在生长季前低温年, 除与冬(r = 0.54*)、春季(r = 0.37*)最低气温显著正相关外, 与其他季节气候因子均相关不显著。在生长季低温年, 祁连圆柏年径向生长与气温和降水的相关关系也不显著。但在生长季前和生长季高温年, 祁连圆柏径向生长与春季降水显著正相关(PHT: r = 0.39*, GHT: r = 0.57*), 而与春季最低气温显著负相关(PHT: r = -0.73*, GHT: r = -0.44*)。在生长季前高温年, 祁连圆柏径向生长与冬季最低气温显著负相关(r = -0.49*); 在生长季高温年, 祁连圆柏径向生长与秋季气温显著负相关(r = -0.42*)。可见在不同极端气温年, 高降水区域祁连圆柏径向生长与气候关系的差异相对于低降水区域更明显。因此极端气温变化可能对高降水区域祁连圆柏年轮-气候关系的影响更大(图5)。
HL采样点, 在生长季前干旱年、生长季干旱年、生长季湿润年, 祁连圆柏径向生长与夏季最低气温都显著正相关(PD: r = 0.64*, GD: r = 0.38*, GW: r = 0.79*)。在生长季前湿润年和生长季干旱年, 祁连圆柏径向生长与夏季降水显著正相关(图5, PW: r = 0.47*, GD: r = 0.35*)。
QS采样点, 在生长季前干旱年, 祁连圆柏径向生长与冬季最低气温(r = 0.44*)、夏季降水(r = 0.59*)显著正相关。在生长季前湿润年, 祁连圆柏径向生长与春季降水显著正相关(r = 0.43*)。在生长季干旱年, 祁连圆柏径向生长与气候因子均无显著相关关系。在生长季湿润年, 祁连圆柏径向生长与冬季(r = 0.42*)、夏季(r = 0.41*)最低气温和春季(r = 0.38*)、夏季(r = 0.51*)降水显著正相关(图5)。
HB采样点, 在生长季前湿润年, 祁连圆柏径向生长与冬季最低气温显著正相关(r = 0.40*)。在生长季前湿润年和生长季湿润年, 祁连圆柏径向生长与其他气候因子均无显著相关关系。在生长季前干旱年和生长季干旱年, 祁连圆柏径向生长与春季最低气温显著负相关(PD: r = -0.41*, GD: r = -0.50*), 祁连圆柏径向生长与春季降水量显著正相关(PD: r = 0.52*, GD: r = 0.71*)。在生长季前干旱年, 径向生长与夏季气温显著负相关(图5, r = -0.43*), 与夏季降水量显著正相关(r = 0.41*)。可见在不同降水年份, 高(HB)、中(QS)降水区域祁连圆柏径向生长与气候关系的差异相对于低降水区域(HL)更明显。因此, 水分对于低降水区域(HL)祁连圆柏年轮-气候关系影响较小, 而对中、高降水区域(QS和HB)祁连圆柏年轮-气候关系影响相对较大。
3 讨论
青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(Yang et al., 2013; Liang et al., 2014; Shi et al., 2014; Zhang et al., 2014, 2015a, 2015b, 2016; Wang et al., 2015; Yin et al., 2016), 本研究不支持干旱区林线树木生长主要受水分限制的假说, 但降水量变化会影响林线树木生长与气温的关系。在本研究中, 降水对不同水分梯度下林线树木生长的影响方式相同, 但气温主要以两种方式影响树木径向生长, 分别为: 低温限制作用(Shi et al., 2014; Zhang et al., 2014, 2015a; Wang et al., 2015; Yin et al., 2016)和高温胁迫作用(Yang et al., 2013; Zhang et al., 2015b, 2016), 这与本研究区附近其他研究结果相似。冬季低温和降雪少引起的土壤低温会损伤树木, 暖冬可提高土壤温度, 降雪增加雪被厚度, 保证树木根系不受低温伤害, 维持树木根系生长及各项生理功能正常发挥的能力(Grace & Norton, 1990; Toromani et al., 2011)。但是, 如果降水量低到不能为生长季前期光合作用提供充足的水分, 春季光合作用潜力的充分发挥就会受到严重影响(Wang et al., 2013)。春季树木休眠解除并进入生长季初期时, 光合作用微弱, 此时树木生长对于土壤可利用水分的敏感性较高(Liang et al., 2014)。生长季前低温导致树木根系生长受阻, 生理活动能力受限, 吸收土壤水分的能力较弱(Toromani et al., 2011), 也会使覆雪期和霜冻期延长, 推迟生长季开始的时间(Wang et al., 2013)。
气温过高会加剧呼吸作用对自身营养物质的消耗, 并且高温会加速土壤水分蒸散, 使得树木可利用水分降低; 气温过高将促进呼吸作用和蒸腾作用, 从而消耗树木所固定的有机物, 同时高温将加速土壤水分蒸发, 进而导致水分亏缺使生长受限(Lavergne et al., 2015)。夏季低温导致土壤和叶面温度降低, 树木光合作用减弱, 蒸腾作用受限, 根系吸水困难, 限制木质部的形成(Ren et al., 2015)。夏季降水增加, 有利于补充土壤可利用水分, 从而促进树木生长(潘瑞炽等, 2012)。
不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异。一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(Lloyd & Fastie, 2002; Lavergne et al., 2015)。Lavergne等(2015)发现随降水增加, 生长季前和生长季内降水对Nothofagus pumilio径向生长的限制作用减弱, 低温限制作用增加。本研究结果与之不同, 本研究生长季前降水的限制作用一直存在。Lavergne等(2015)的研究区域的平均年降水量跨度极大(从4β000 mm到500 mm), 本研究高降水区域(HB)的平均年降水量仅为470 mm, 且冬季和春季降水量非常少(仅约20 mm), 仍属半干旱区降水范围, 这些降水并不足以提供冬季雪被厚度和来年春季的水分需求。但本研究并未发现随降水量增加低温限制作用增加的现象, 却出现了随降水量增加, 冬季和夏季最低气温对祁连圆柏树木径向生长的低温限制减弱, 树木生长由春季低温限制变为高温胁迫的现象。其他祁连圆柏树木生长与气候关系的研究中也在采样点附近有类似发现(Yang et al., 2013; Zhang et al., 2014)。高降水区域(HB)冬季降雪增加, 不仅可以增加雪被厚度以维持地温, 使树木在冬季受到较少的低温损伤(Toromani et al., 2011), 而且可以为生长季初期提供充足的土壤水分。夏季树木生长旺盛, 较低的土壤和叶面温度会使树木光合作用受抑(潘瑞炽等, 2012), 蒸腾作用减弱, 根系吸水困难, 限制木质部的形成(Ren et al., 2015), 夏季降水增加则有利于补充土壤水分, 从而促进树木生长。春季树木光合作用较弱或者仍处于休眠期, 此时树木生长对于土壤水分的敏感性较高(Liang et al., 2014), 气温升高加快呼吸作用对树木自身营养物质的消耗(潘瑞炽等, 2012), 并且高温会加速土壤水分蒸发, 使得树木可利用水分降低。
同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(Macalady & Bugmann, 2014; Carnwath et al., 2016)类似。在低降水区域(HL) , 极端年份气候波动对祁连圆柏生长-气候关系影响不明显; 在中降水区域(QS), 极端年份水分变化对生长-气候关系影响显著。在生长季前干旱年, 树木生长主要受春夏季最低气温和夏季降水限制, 而在生长季(或季前)湿润年, 生长主要受春季降水和夏季气温限制(Carnwath et al., 2016)。在高降水区域(HB), 林线树木生长的限制因子在不同特征年中变化最大(图5), 在低温年(PLT和GLT)、湿润年(PW和GW)和生长季湿润年, 树木生长对气候变化不敏感(Macalady & Bugmann, 2014)。这可能是因为气温偏低, 水分相对充足, 使树木在生长季前和生长季内的水热条件适宜, 土壤和空气中可利用水分的蒸散减少, 从而不受气温和水分的限制(潘瑞炽等, 2012; Macalady & Bugmann, 2014)。在高温年(PHT和GHT)、干旱年(PD和GD), 树木生长主要受春季水分可利用性限制(Yang et al., 2013; Zhang et al., 2015b, 2016), 这表明虽然总体而言水分较充足, 但就不同季节而言, 气温升高也可能造成水分胁迫作用。
4 结论
青藏高原东北部高山林线祁连圆柏径向生长的气候限制作用在不同降水梯度下存在明显差异, 但本研究结果并不支持在干旱区水分是高山林线树木生长的主控因子这一假说。在低、中、高降水区域, 降水对径向生长的限制作用都存在, 但温度的限制作用在不同降水梯度下差异较大。随着降水量增加, 不同季节气温对祁连圆柏生长的限制作用明显出现差异, 水分的季节分配对生长更重要。在不同降水梯度下, 极端气候年也影响祁连圆柏径向生长与气候的关系, 这种影响在高降水区域最为明显, 在高温和干旱年祁连圆柏径向生长的气候敏感性明显高于低温和湿润年。
特邀编委: 张齐兵 责任编辑: 李 敏
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/S0925-7721(01)00003-7URL [本文引用: 1]

The Working Group 1 contribution to the IPCC Fifth Assessment Report (AR5) extends from observations and paleoclimate information regarding past changes in the climate system, a comprehensive evaluation of climate models, the detection and attribution of observed changes to natural or anthropogenic forcing, through to projected future changes on both near-term and long-term time scales. Human influence on the climate system is now detected with increased certainty, both globally and in most regions. Since the mid-20th century, the increase in anthropogenic greenhouse gas concentrations has led to surface warming over almost the entire globe, while at the same time, the ocean has continued to warm and store energy. Satellite based observations reveal with improved precision that summer sea ice extent is declining rapidly in the Arctic, glaciers are retreating world-wide, and global mean sea level continues to rise. Concurrent with a continued increase in atmospheric CO2 concentrations, the oceanic uptake of CO2 has resulted in decreasing pH of seawater since the beginning of the industrial era. Projections of future changes in the climate system to the end of the 21st century are based on a series of new climate models and new scenarios, but are broadly consistent with previous assessment findings, confirming widespread and significant changes across the climate system. Greater warming is projected to occur over land than ocean, with the most rapid warming in the Arctic region. Based on modeled changes in seasonal mean precipitation, the contrast between wet and dry regions, and wet and dry seasons is projected to increase as global temperatures rise. Confidence in projections of global mean sea level rise has increased since the previous IPCC assessment report, and projections now include future rapid ice-sheet dynamical changes. On long time scales, warming is dominated by total emissions of CO2, and many changes will persist for centuries even if CO2 emissions were stopped. In 2014, the Fifth Assessment cycle of the IPCC will be completed following the release of the reports of Working Groups II (impacts, adaptation and vulnerability) and Working Groups III (mitigation of climate change), and finally the Synthesis Report, the combined synthesis product based on all three underlying Working Group assessment reports.
DOI:10.1111/1365-2745.12604URL [本文引用: 2]
Summary To accurately predict how ecosystems will respond to climate change – and how management actions can influence such responses – scientists and managers need a better understanding of how and when biotic interactions modify climate–growth relationships. However, current research has largely ignored the role of competition in modulating climate–growth relationships of mature trees. In this study, we assessed the effect of competition on tree responses to drought and interannual climate variability as well as linkages between climate sensitivity and morphological characteristics of the stem wood. We sampled 10 sites in north-eastern Washington, USA, and used dendroecology to quantify responses of Douglas-fir ( Pseudotsuga menziesii ) to drought and climate variability. Tree-ring series were converted to basal area increment series, and the effects of competition on climate–growth relationships were assessed at the tree and site levels using a combination of correlation analyses and linear mixed-effects models. Competition did not affect tree responses to extreme drought. When soil moisture was below average, tree growth was tightly coupled to climate variability for all trees, regardless of their competitive status. However, in wet years, competition had a pronounced, positive effect on climate sensitivity of growth. Trees with more competition from neighbours exhibited a significantly higher proportion of sapwood area in latewood (a morphological trait associated with greater drought resistance). Synthesis . Our results suggest that a tree's ability to cope with environmental variability is driven not just by the proximate effects of neighbours on resource availability, but also by phenotypic plasticity and long-term adaptations to competitive stress (such as changes in carbon allocation). Findings have important implications for individual-tree and stand-level growth models and may help managers better understand how their activities will modify tree responses to climate change.
DOI:10.1111/gcb.13366URLPMID:27298138 [本文引用: 1]
Biogeographical and ecological theory suggests that species distributions should be driven to higher altitudes and latitudes as global temperatures rise. Such changes occur as growth improves at the poleward edge of a species distribution and declines at the range edge in the opposite or equatorial direction, mirrored by changes in the establishment of new individuals. A substantial body of evidence demonstrates that such processes are underway for a wide variety of species. Case studies from populations at the equatorial range edge of a variety of woody species have led us to understand that widespread growth decline and distributional shifts are underway. However, in apparent contrast, other studies report high productivity and reproduction in some range edge populations. We sought to assess temporal trends in the growth of the widespread European beech tree (Fagus sylvatica) across its latitudinal range. We explored the stability of populations to major drought events and the implications for predicted widespread growth decline at its equatorial range edge. In contrast to expectations, we found greatest sensitivity and low resistance to drought in the core of the species range, while dry range edge populations showed particularly high resistance to drought and little evidence of drought-linked growth decline. We hypothesise that this high range-edge resistance to drought is driven primarily by local environmental factors that allow relict populations to persist despite regionally unfavourable climate. The persistence of such populations demonstrates that range edge decline is not ubiquitous and is likely to be driven by declining population density at the landscape scale rather than sudden and widespread range retraction.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3354/cr008001URL [本文引用: 1]
Two methods are investigated for interpolating daily minimum and maximum air temperatures (Tmin and Tmax) at a 1 km spatial resolution over a large mountainous region (830 000 km2) in the U.S. Pacific Northwest. The methods were selected because of their ability to (1) account for the effect of elevation on temperature and (2) efficiently handle large volumes of data. The first method, the neutral stability algorithm (NSA), used the hydrostatic and potential temperature equations to convert measured temperatures and elevations to sea-level potential temperatures. The potential temperatures were spatially interpolated using an inverse-squared-distance algorithm and then mapped to the elevation surface of a digital elevation model (DEM). The second method, linear lapse rate adjustment (LLRA), involved the same basic procedure as the NSA, but used a constant linear lapse rate instead of the potential temperature equation. Cross-validation analyses were performed using the NSA and LLRA methods to interpolate Tmin and Tmax each day for the 1990 water year, and the methods were evaluated based on mean annual interpolation error (IE). The NSA method showed considerable bias for sites associated with vertical extrapolation. A correction based on climate station/grid cell elevation differences was developed and found to successfully remove the bias. The LLRA method was tested using 3 lapse rates, none of which produced a serious extrapolation bias. The bias-adjusted NSA and the 3 LLRA methods produced almost identical levels of accuracy (mean absolute errors between 1.2 and 1.3??C), and produced very similar temperature surfaces based on image difference statistics. In terms of accuracy, speed, and ease of implementation, LLRA was chosen as the best of the methods tested.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-2699.2004.01064.xURL [本文引用: 1]
Abstract During a repeat photography study quaking aspen ( Populus tremuloides Michx.) was observed invading conifer stands at treeline in the San Juan Mountains of south-western Colorado. Aspen tree core samples were collected from nine plots ranging in elevation from 3192 to 3547m, and estimated dates of establishment of aspen were grouped into 10-year intervals for analysis. Estimated periods of establishment were compared with century-long climate data records to derive any correlations with aspen invasion. Other disturbance agents, such as fire and livestock grazing were also considered. Quantitative analysis of climate variables suggests that decreased mean spring precipitation and increased mean summer maximum temperature provide optimal conditions for aspen establishment. Episodes of invasion were non-synchronous, but all occurred after 1900, and are likely from seed germination, considered unusual in aspen. Different climate variables explain stand initiation from seed and subsequent peak establishment from vegetative reproduction. Long-term climate records indicate a general warming since the beginning of the 20th century and explain the continued invasion and persistence of aspen at treeline, resulting from asexual reproduction. Short-term climate records identify anomalously cool, moist years that explain rarely observed sexual reproduction in aspen.
DOI:10.1016/j.quascirev.2015.12.020URL [本文引用: 1]
61The timberline had exceeded 154002m by 10,20002cal.02yr BP when summers were warmer.61The treeline reached its maximum after 850002cal.02yr BP under milder winter conditions.61Cool summers led to a lowering of the timberline and treeline from 490002cal.02yr BP.61Since the Bronze Age (350002cal.02yr BP) human impact has accentuated treeline depression.61Future upslope tree movements will favour abandoned agricultural areas.
DOI:10.1007/s00468-012-0776-6URL [本文引用: 1]
In the context of global warming, it is of high importance to assess the influence of climatic change and geographic factors on the radial growth of high-elevation trees. Using tree-ring data collected from four stands of Qilian juniper ( Juniperus przewalskii Kom. ) across an altitudinal gradient in the central Qilian Mountains, northwest China, we compared the radial growth characteristics and climate–growth relationships at different elevations. Results indicated that there was little difference in the tree-ring parameters of the four chronologies. Correlation analyses both for unfiltered and 10-year high-passed data of monthly climatic variables and chronologies were presented to investigate the climatic forcing on tree growth, and results revealed that the correlation patterns were consistent among the four sites, especially for high-passed data. We employed the principal components analysis method to obtain the first principal component (PC1) of the four chronologies and computed the correlations between PC1 and climate factors. The PC1 correlated significantly with winter (November–January) temperature, prior August and current May temperature, and precipitation in the previous September and current January and April, indicating that tree growth in this region was mainly limited by cold winter temperature and drought in early growing season and prior growing season (prior August and September). However, the climate–growth relationships were unstable; with an increase in temperature, the sensitivity of tree growth to temperature had decreased over the past few decades. Considering the instability of the climate–growth relationships, climate reconstructions based on tree rings in the study area should be approached with more caution.
DOI:10.2307/2260887URL [本文引用: 1]
(1) The influence of climatic factors on the growth of Pinus sylvestris was evaluated by comparing annual ring-widths from seven sites along an altitudinal gradient in Scotland with monthly climatic data. (2) Temperature was found to be more important than rainfall in influencing growth. Both late-winter (January-February) and summer (July-August) temperatures were significantly and positively correlated with ring-width. Winter browning or frost drought is a possible cause of the winter temperature correlation. (3) Krummholz trees, at the tree-line, were found to grow more, and their growth was less strongly correlated with climatic factors, than trees at lower elevations. It is suggested that this is due to their short stature and resultant higher tissue temperatures.
DOI:10.1111/j.1461-0248.2009.01355.xURLPMID:19682007 [本文引用: 1]
Treelines are temperature sensitive transition zones that are expected to respond to climate warming by advancing beyond their current position. Response to climate warming over the last century, however, has been mixed, with some treelines showing evidence of recruitment at higher altitudes and/or latitudes (advance) whereas others reveal no marked change in the upper limit of tree establishment. To explore this variation, we analysed a global dataset of 166 sites for which treeline dynamics had been recorded since 1900 AD. Advance was recorded at 52% of sites with only 1% reporting treeline recession. Treelines that experienced strong winter warming were more likely to have advanced, and treelines with a diffuse form were more likely to have advanced than those with an abrupt or krummholz form. Diffuse treelines may be more responsive to warming because they are more strongly growth limited, whereas other treeline forms may be subject to additional constraints.
DOI:10.1111/j.1466-822X.2005.00168.xURL [本文引用: 1]
ABSTRACT Top of page ABSTRACT INTRODUCTION SENSITIVITY AND RESPONSE IN DIFFERENT TREELINE TYPES CONCLUSIONS ACKNOWLEDGEMENTS REFERENCES The sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change are increasingly discussed in terms of climate change, often forgetting that climate is only one aspect of environmental variation. As treeline heterogeneity increases from global to regional and smaller scales, assessment of treeline sensitivity at the landscape and local scales requires a more complex approach than at the global scale. The time scale (short-, medium-, long-term) also plays an important role when considering treeline sensitivity. The sensitivity of the treeline to a changing environment varies among different types of treeline. Treelines controlled mainly by orographic influences are not very susceptible to the effects of warming climates. Greatest sensitivity can be expected in anthropogenic treelines after the cessation of human activity. However, tree invasion into former forested areas above the anthropogenic forest limit is controlled by site conditions, and in particular, by microclimates and soils. Apart from changes in tree physiognomy, the spontaneous advance of young growth of forest-forming tree species into present treeless areas within the treeline ecotone and beyond the tree limit is considered to be the best indicator of treeline sensitivity to environmental change. The sensitivity of climatic treelines to climate warming varies both in the local and regional topographical conditions. Furthermore, treeline history and its after-effects also play an important role. The sensitivity of treelines to changes in given factors (e.g. winter snow pack, soil moisture, temperature, evaporation, etc.) may vary among areas with differing climatic characteristics. In general, forest will not advance in a closed front but will follow sites that became more favourable to tree establishment under the changed climatic conditions.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.dendro.2015.09.001URL [本文引用: 2]
Over the last decades, gradual changes in summer climate in the Southern Hemisphere have affected forest growth in contrasting ways in moist and dry regions. Here, we use correlation analysis and a forward process-based model (Vaganov hashkin-Lite) to investigate changes in climate limitation of the interannual tree-ring growth ofNothofagus pumilioat the upper treeline along a precipitation gradient in northern Patagonia. Patterns of climate limitation vary consistently along the gradient. At mesic and humid treelines, tree-ring growth is positively related to growing season temperature and negatively to precipitation. At xeric treelines, the opposite is observed. Moreover, the climate-growth relations are not stationary. In particular, according to the model, the step decrease in precipitation in 1952 induced an increase of the moisture limitation at the dry edge of the gradient. Correlation analyses evidence that the dependence of growth on moisture after 1952 has enhanced since 1976. While the model consistently reproduces tree-ring width variations over the 1931 1975 period, it does not capture the growth patterns in the following years. Some environmental parameters (cloudiness, snowpack, atmospheric CO2) affecting moisture, radiation and stomatal aperture may have reached thresholds beyond which the effect on tree-growth has become sizable.
URL
Micrometeorological measurements were made in the lower alpinezone of Mt. Haleakala on Maui, Hawaiian Islands, in March 1988 to characterizeecologically significant climatological parameters. Daily courses ofphotosynthetically active and total net radiation; temperatures of air, soil, andplant canopy; wind speed; air humidity; leaf wetness; and precipitation wererecorded at an elevation of 2100 m in alpine scrub slightly above the timberline.A battery-powered data-logging system was used, which gave high temporalresolution. Influence of variable cloud cover on microclimate of the study sitewas evaluated on five selected days with highly differing weather conditions.Based on comparison with data from other high mountains of the humid tropicalzone, it is concluded that the alpine timberline on Maui is caused by a complexof factors. Plant water availability is probably the dominating one; temperatureseems to be of lesser importance. The possible role of other important factors isdiscussed. The extraordinary microclimatological conditions of the alpine zoneof Maui are examined in the context of the atmospheric circulation system inthe region of the Hawaiian archipelago.
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/13-1904.1URL [本文引用: 5]
Abstract Birch ( Betula ) trees and forests are found across much of the temperate and boreal zones of the Northern Hemisphere. Yet, despite being an ecologically significant genus, it is not well studied compared to other genera like Pinus , Picea , Larix , Juniperus , Quercus , or Fagus . In the Himalayas, Himalayan birch ( Betula utilis ) is a widespread broadleaf timberline species that survives in mountain rain shadows via access to water from snowmelt. Because precipitation in the Nepalese Himalayas decreases with increasing elevation, we hypothesized that the growth of birch at the upper timberlines between 3900 and 4150 m above sea level is primarily limited by moisture availability rather than by low temperature. To examine this assumption, a total of 292 increment cores from 211 birch trees at nine timberline sites were taken for dendroecological analysis. The synchronous occurrence of narrow rings and the high interseries correlations within and among sites evidenced a reliable cross-dating and a common climatic signal in the tree-ring width variations. From March to May, all nine tree-ring-width site chronologies showed a strong positive response to total precipitation and a less-strong negative response to temperature. During the instrumental meteorological record (from 1960 to the present), years with a high percentage of locally missing rings coincided with dry and warm pre-monsoon seasons. Moreover, periods of below-average growth are in phase with well-known drought events all over monsoon Asia, showing additional evidence that Himalayan birch growth at the upper timberlines is persistently limited by moisture availability. Our study describes the rare case of a drought-induced alpine timberline that is comprised of a broadleaf tree species.
[本文引用: 1]
DOI:10.11821/yj2010120017URL [本文引用: 1]
基于69个气象台站的气象数据,对青藏高原地区1961~2005年来的主要气候因子特征进行了分析。结果表明:1961~2005年的45年间,青藏高原地区年平均温度呈上升趋势,其倾向率为0.265℃/10a,其中青藏高原地区冬季气温变暖趋势明显,春季变暖趋势不明显;20世纪80年代以来青藏高原地区的温度升高有加速的趋势。近45年来青藏高原地区年降水量呈现微弱增加趋势,其倾向率为8.21 mm/10a。青藏高原地区春季和冬季降水量都以增加趋势为主,但春季增加趋势远远大于冬季。青藏高原地区降水存在一定的周期性,32个站表现出短周期特性,为2~4年左右;11个站表现出中周期特性,为5~8年;6个站表现出长周期特性,均大于10年。1961~2005年间,青藏高原地区整体气候变化以暖湿化趋势为主,暖湿化站点占总数的67%。
DOI:10.11821/yj2010120017URL [本文引用: 1]
基于69个气象台站的气象数据,对青藏高原地区1961~2005年来的主要气候因子特征进行了分析。结果表明:1961~2005年的45年间,青藏高原地区年平均温度呈上升趋势,其倾向率为0.265℃/10a,其中青藏高原地区冬季气温变暖趋势明显,春季变暖趋势不明显;20世纪80年代以来青藏高原地区的温度升高有加速的趋势。近45年来青藏高原地区年降水量呈现微弱增加趋势,其倾向率为8.21 mm/10a。青藏高原地区春季和冬季降水量都以增加趋势为主,但春季增加趋势远远大于冬季。青藏高原地区降水存在一定的周期性,32个站表现出短周期特性,为2~4年左右;11个站表现出中周期特性,为5~8年;6个站表现出长周期特性,均大于10年。1961~2005年间,青藏高原地区整体气候变化以暖湿化趋势为主,暖湿化站点占总数的67%。
DOI:10.1023/A:1014278819094URL [本文引用: 2]
In this study, we investigated the response of trees growing at the cold margins of the boreal forest to climate variation in the 20th century. Working at eight sites at and near alpine and arctic treeline in three regions in Alaska, we compared tree growth (from measured tree ring-widths) to historical climate data to document how growth has responded to climate variation in the 20th century. We found that there was substantial regional variability in response to climate variation. Contrary to our expectations, we found that after 1950 warmer temperatures were associated with decreased tree growth in all but the wettest region, the Alaska Range. Although tree growth increased from 1900 1950 at almost all sites, significant declines in tree growth were common after 1950 in all but the Alaska Range sites. We also found that there was substantial variability in response to climate variation according to distance to treeline. Inverse growth responses to temperature were more common at sites below the forest margin than at sites at the forest margin. Together, these results suggest that inverse responses to temperature are widespread, affecting even the coldest parts of the boreal forest. Even in such close proximity to treeline, warm temperatures after 1950 have been associated with reduced tree growth. Growth declines were most common in the warmer and drier sites, and thus support the hypothesis that drought-stress may accompany increased warming in the boreal forest.
DOI:10.1029/2001JD000852URL
[1] Boreal tree species are expected to invade tundra ecosystems as climate warms. Because forested ecosystems differ from tundra ecosystems in a number of climatically relevant characteristics, this advance of the altitudinal and latitudinal tree limit may ultimately feedback on regional climate. We used tree rings to reconstruct the response of the treeline ecotone on the Seward Peninsula in northwestern Alaska to 20th century warming. We further explored the sensitivity of the treeline ecotone to climate change using the spatially explicit, frame-based model ALFRESCO. Our reconstructions of forest response to past warming indicate that in upland tundra spruce have successfully established progressively farther from the forest limit since the 1880s. Shrub tundra has thus been converted to low-density forest tundra within a band extending approximately 10 km from the forest limit. In lowland sites, where soil thermal and hydrologic properties are the primary constraint on tree distribution, trees began establishing in tundra after 1920, and establishment was restricted to areas that had experienced thermokarst activity in the past. Modeling experiments suggested that changes in disturbance regime and spruce growth response to climate may introduce strongly nonlinear responses to climate change at treeline. Field data and model experiments thus both indicate that large and nearly instantaneous responses to warming are likely at the treeline ecotone but that sensitivity to warming is likely to vary substantially over space and time.
DOI:10.3354/cr01144URL [本文引用: 1]
Information about historical summer temperature variability is rare on the Qinghai-Tibetan Plateau (QTP). Here we report a 290 yr summer (June to August) mean minimum temperature (SMMT) record reconstructed from tree-ring widths of alpine juniper Sabina squamata in Yadong County at the southern edge of QTP, China. The reconstruction accounts for 51.2% of the SMMT variance in the instrumental period from 1956 to 2002. Five cold intervals in 1725-1734, 1745-1789, 1817-1825, 1860-1869 and 1967-1978, and 4 warm intervals in 17351744, 1790-1816, 1870-1879 and 1990-2002 were identified. Wavelet analysis of the reconstructed temperatures suggested the existence of 2 to 7 yr cycles in discontinuous periods and 20 to 25 yr cycles in the 1850s to 1880s. The reconstructed summer temperature variations were consistent with other temperature records from tree rings, ice cores and glacier activities on the QTP, which demonstrate the fidelity of the reconstruction and suggested that the climate in the study region was part of a large-scale climate system. The 1980s to 2000s were the warmest period in the last 3 centuries in our tree-ring records, but the amplitude of the warming was still within the natural variation.
DOI:10.1371/journal.pone.? 0092770URLPMID:4008371 [本文引用: 3]
Abstract The processes leading to drought-associated tree mortality are poorly understood, particularly long-term predisposing factors, memory effects, and variability in mortality processes and thresholds in space and time. We use tree rings from four sites to investigate Pinus edulis mortality during two drought periods in the southwestern USA. We draw on recent sampling and archived collections to (1) analyze P. edulis growth patterns and mortality during the 1950s and 2000s droughts; (2) determine the influence of climate and competition on growth in trees that died and survived; and (3) derive regression models of growth-mortality risk and evaluate their performance across space and time. Recent growth was 53% higher in surviving vs. dying trees, with some sites exhibiting decades-long growth divergences associated with previous drought. Differential growth response to climate partly explained growth differences between live and dead trees, with responses wet/cool conditions most influencing eventual tree status. Competition constrained tree growth, and reduced trees' ability to respond to favorable climate. The best predictors in growth-mortality models included long-term (15-30 year) average growth rate combined with a metric of growth variability and the number of abrupt growth increases over 15 and 10 years, respectively. The most parsimonious models had high discriminatory power (ROC>0.84) and correctly classified 70% of trees, suggesting that aspects of tree growth, especially over decades, can be powerful predictors of widespread drought-associated die-off. However, model discrimination varied across sites and drought events. Weaker growth-mortality relationships and higher growth at lower survival probabilities for some sites during the 2000s event suggest a shift in mortality processes from longer-term growth-related constraints to shorter-term processes, such as rapid metabolic decline even in vigorous trees due to acute drought stress, and/or increases in the attack rate of both chronically stressed and more vigorous trees by bark beetles.
DOI:10.1029/2009JD013493URL [本文引用: 1]
[1] The typically sparse distribution of weather stations in mountainous terrain inadequately resolves temperature variability. Accordingly, high-resolution gridding of climate data (for applications such as hydrological modeling) often relies on assumptions such as a constant surface temperature lapse rate (i.e., decrease of surface temperature with altitude) of 6.500°C km0908081. Using an example of the Cascade Mountains, we describe the temporal and spatial variability of the surface temperature lapse rate, combining data from: (1) COOP stations, (2) nearby radiosonde launches, (3) a temporary dense network of sensors, (4) forecasts from the MM5 regional model, and (5) PRISM geo-statistical analyses. On the windward side of the range, the various data sources reveal annual mean lapse rates of 3.90900095.200°C km0908081, substantially smaller than the often-assumed 6.500°C km0908081. The data sets show similar seasonal and diurnal variability, with lapse rates smallest (2.50900093.500°C km0908081) in late-summer minimum temperatures, and largest (6.50900097.500°C km0908081) in spring maximum temperatures. Geographic (windward versus lee side) differences in lapse rates are found to be substantial. Using a simple runoff model, we show the appreciable implications of these results for hydrological modeling.
DOI:10.1890/03-0269URL [本文引用: 1]
We present a framework for fitting multiple random walks to animal movement paths consisting of ordered sets of step lengths and turning angles. Each step and turn is assigned to one of a number of random walks, each characteristic of a different behavioral state. Behavioral state assignments may be inferred purely from movement data or may include the habitat type in which the animals are located. Switching between different behavioral states may be modeled explicitly using a state transition matrix estimated directly from data, or switching probabilities may take into account the proximity of animals to landscape features. Model fitting is undertaken within a Bayesian framework using the WinBUGS software. These methods allow for identification of different movement states using several properties of observed paths and lead naturally to the formulation of movement models. Analysis of relocation data from elk released in east-central Ontario, Canada, suggests a biphasic movement behavior: elk are either in an "encamped" state in which step lengths are small and turning angles are high, or in an "exploratory" state, in which daily step lengths are several kilometers and turning angles are small. Animals encamp in open habitat (agricultural fields and opened forest), but the exploratory state is not associated with any particular habitat type.
DOI:10.1126/science.1082750URLPMID:12791990 [本文引用: 1]
Recent climatic changes have enhanced plant growth in northern mid-latitudes and high latitudes. However, a comprehensive analysis of the impact of global climatic changes on vegetation productivity has not before been expressed in the context of variable limiting factors to plant growth. We present a global investigation of vegetation responses to climatic changes by analyzing 18 years (1982 to 1999) of both climatic data and satellite observations of vegetation activity. Our results indicate that global changes in climate have eased several critical climatic constraints to plant growth, such that net primary production increased 6% (3.4 petagrams of carbon over 18 years) globally. The largest increase was in tropical ecosystems. Amazon rain forests accounted for 42% of the global increase in net primary production, owing mainly to decreased cloud cover and the resulting increase in solar radiation.
[本文引用: 4]
[本文引用: 4]
DOI:10.3773/j.issn.1005-264x.2010.05.005URL [本文引用: 2]
选择青海省同德县南部河北林场的一个连续坡面,根据不同海拔和坡 向设置4个采样点,采集祁连圆柏(Sabina przewalskii)树轮数据,分析不同海拔和坡向对树木生长的影响.结果表明:坡面上部3个采样点的树轮年表特征值均呈一定的变化规律--平均敏感 值(MS)和标准差(SD)随海拔升高而增大,一阶自相关(AC)随海拔升高而递减,下限年表特征值均表现出与其他3点的不同,都是最值(MS和SD均最 大,AC最小);年表间相关和主成分分析结果都显示出海拔梯度上的变化规律,但下限差异显著;树轮指数与当年6-18月平均气温的相关系数呈增强趋势,森 林上限受当年7、8月平均气温影响较大,下限树轮指数不仅与当年6月和前一年11月的气温显著负相关,而且受前一年8月和当年5月的月降水量影响显著.与 通常情况"下限树木生长受降水制约"比较,这里的温度作用增强而降水限制减弱.显然,坡向扭转是海拔梯度上影响祁连圆柏生长变化的重要因子.
DOI:10.3773/j.issn.1005-264x.2010.05.005URL [本文引用: 2]
选择青海省同德县南部河北林场的一个连续坡面,根据不同海拔和坡 向设置4个采样点,采集祁连圆柏(Sabina przewalskii)树轮数据,分析不同海拔和坡向对树木生长的影响.结果表明:坡面上部3个采样点的树轮年表特征值均呈一定的变化规律--平均敏感 值(MS)和标准差(SD)随海拔升高而增大,一阶自相关(AC)随海拔升高而递减,下限年表特征值均表现出与其他3点的不同,都是最值(MS和SD均最 大,AC最小);年表间相关和主成分分析结果都显示出海拔梯度上的变化规律,但下限差异显著;树轮指数与当年6-18月平均气温的相关系数呈增强趋势,森 林上限受当年7、8月平均气温影响较大,下限树轮指数不仅与当年6月和前一年11月的气温显著负相关,而且受前一年8月和当年5月的月降水量影响显著.与 通常情况"下限树木生长受降水制约"比较,这里的温度作用增强而降水限制减弱.显然,坡向扭转是海拔梯度上影响祁连圆柏生长变化的重要因子.
DOI:10.1111/j.1744-7909.2008.00688.xURLPMID:18713348 [本文引用: 1]
Three ring-width chronologies were developed from Qilian Juniper (Sabina przewalskii Kom.) at the upper treeline along a west-east gradient in the Anyemaqen Mountains.Most chronological statistics,except for mean sensitivity (MS),decreased from west to east.The first principal component (PC1) Ioadings indicated that stands in a similar climate condition were most important to the variability of radial growth.PC2 Ioadings decreased from west to east,suggesting the difference of tree-growth between eastern and western Anyemaqen Mountains.Correlations between standard chronologies and climatic factors revealed different climatic influences on radial growth along a west-east gradient in the study area.Temperature of warm season (July-August) was important to the radial growth at the upper treeline in the whole study area.Precipitation of current May was an important limiting factor of tree growth only in the western (drier) upper treeline,whereas precipitation of current September limited tree growth in the eastern (wetter) upper treeline.Response function analysis results showed that there were regional differences between tree growth and climatic factors in various sampling sites of the whole study area.Temperature and precipitation were the important factors influencing tree growth in western (drier) upper treeline.However,tree growth was greatly limited by temperature at the upper treeline in the middle area,and was more limited by precipitation than temperature in the eastern (wetter) upper treeline.
DOI:10.1111/1365-2745.12555URL [本文引用: 1]
Summary The growth limitation hypothesis (GLH) is the most accepted explanation for treeline formation, but it has been scarcely examined in Mediterranean regions, where treelines are located at lower elevations than in temperate regions. The GLH states that low temperature is the ultimate environmental driver for treeline formation, constraining C-sinks (i.e. tissue formation) more than C-sources. The GLH predicts similar or increasing (but not decreasing) non-structural carbohydrate (NSC) concentrations with elevation throughout the course of the growing season. We hypothesized that elevational trends in growth and NSC in Mediterranean regions are not determined by low temperature alone. We tested the GLH by examining the mean annual basal area and NSC concentrations in developing (new twigs) and ripened tissues (branch, stem) of trees growing at three elevations in three Mediterranean and three temperate treelines in the Chilean southern Andes (33°02S, Kageneckia angustifolia ; 36, 40 and 46°02S, Nothofagus pumilio ) and in Spain (36°02N, Pinus sylvestris ; 42°02N, Pinus uncinata ). Samples for NSC were taken at the onset of summer and autumn, which represent periods of contrasting drought intensities in Mediterranean regions. Tree growth decreased significantly with elevation in temperate treelines but not in Mediterranean treelines. In Mediterranean treelines, new twig NSC concentrations increased significantly with elevation in the early summer but not in the early autumn. In temperate treelines, in contrast, no elevational or seasonal variation (or interaction between them) was observed in new twig NSC concentrations. The NSC concentrations of the branches and stems from both climates showed no elevational trends. The soluble sugars' NSC fraction increased over the season in Mediterranean treelines and decreased in temperate treelines. Synthesis . Although we found support for the growth limitation hypothesis (GLH) in temperate and Mediterranean treelines, our study shows that the tree growth and C balance in Mediterranean treelines are not controlled by low temperature alone. We suggest that environmental factors other than temperature explain the lower global elevation of Mediterranean treelines when compared to temperate treelines.
DOI:10.1111/gcb.12703URLPMID:25099555 [本文引用: 1]
Forest growth is sensitive to interannual climatic change in the alpine treeline ecotone (ATE). Whether the alpine treeline ecotone shares a similar pattern of forest growth with lower elevational closed forest belt (CFB) under changing climate remains unclear. Here, we reported an unprecedented acceleration of Picea schrenkiana forest growth since 1960s in the ATE of Tianshan Mountains, northwestern China by a stand-total sampling along six altitudinal transects with three plots in each transect: one from the ATE between the treeline and the forest line, and the other two from the CFB. All the sampled P. schrenkiana forest patches show a higher growth speed after 1960 and, comparatively, forest growth in the CFB has sped up much slower than that in the ATE. The speedup of forest growth at the ATE is mainly accounted for by climate factors, with increasing temperature suggested to be the primary driver. Stronger water deficit as well as more competition within the CFB might have restricted forest growth there more than that within the ATE, implying biotic factors were also significant for the accelerated forest growth in the ATE, which should be excluded from simulations and predictions of warming-induced treeline dynamics.
DOI:10.1093/aob/mcu259URLPMID:25725006 [本文引用: 2]
BACKGROUND AND AIMS: A series of studies have shown that temperature triggers the onset of xylogenesis of trees after winter dormancy. However, little is known about whether and how moisture availability influences xylogenesis in spring in drought-prone areas.METHODS: Xylogenesis was monitored in five mature Qilian junipers (Juniperus przewalskii) by microcore sampling from 2009 to 2011 in a semi-arid area of the north-eastern Tibetan Plateau. A simple physical model of xylem cell production was developed and its sensitivity was analysed. The relationship between climate and growth was then evaluated, using weekly wood production data and climatic data from the study site.KEY RESULTS: Delayed onset of xylogenesis in 2010 corresponded to a negative standardized precipitation evapotranspiration index (SPEI) value and a continuous period without rainfall in early May. The main period of wood formation was in June and July, and drier conditions from May to July led to a smaller number of xylem cells. Dry conditions in July could cause early cessation of xylem differentiation. The final number of xylem cells was mainly determined by the average production rate rather than the duration of new cell production. Xylem growth showed a positive and significant response to precipitation, but not to temperature.CONCLUSIONS: Precipitation in late spring and summer can play a critical role in the onset of xylogenesis and xylem cell production. The delay in the initiation of xylogenesis under extremely dry conditions seems to be a stress-avoidance strategy against hydraulic failure. These findings could thus demonstrate an evolutionary adaptation of Qilian juniper to the extremely dry conditions of the north-eastern Tibetan Plateau.
DOI:10.1890/15-1549URLPMID:27220216 [本文引用: 1]
Abstract Climate and competition are often presented from two opposing views of the dominant driver of individual tree growth and species distribution in temperate forests, such as those in the eastern United States. Previous studies have provided abundant evidence indicating that both factors influence tree growth, and we argue that these effects are not independent of one another and rather that interactions between climate, competition, and size best describe tree growth. To illustrate this point, we describe the growth responses of five common eastern tree species to interacting effects of temperature, precipitation, competition, and individual size using maximum likelihood estimation. Models that explicitly include interactions among these four factors explained over half of the variance in annual growth for four out of five species using annual climate. Expanding temperature and precipitation analyses to include seasonal interactions resulted in slightly improved models with a mean R2 of 0.61 (SD 0.10). Growth responses to individual factors as well their interactions varied greatly among species. For example, growth sensitivity to temperature for Quercus rubra increased with maximum annual precipitation, but other species showed no change in sensitivity or slightly reduced annual growth. Our results also indicate that three-way interactions among individual stem size, competition, and temperature may determine which of the five co-occurring species in our study could have the highest growth rate in a given year. Continued consideration and quantification of interactions among climate, competition, and individual-based characteristics are likely to increase understanding of key biological processes such as tree growth. Greater parameterization of interactions between traditionally segregated factors such as climate and competition may also help build a framework to reconcile drivers of individual-based processes such as growth with larger-scale patterns of species distribution.
DOI:10.1007/s00382-014-2386-zURL [本文引用: 2]
Despite instrumental records showing recent large temperature rises on the Tibetan Plateau (TP), only a few tree-ring temperature reconstructions do capture this warming trend. Here, we sampled 260 trees from seven Alpine treeline locations across the southeast TP. Standardized tree-ring width chronologies of Abies squamata and Sabina squamat were produced following Regional Curve Standardization detrending. The leading principal component of these records is well correlated with the regional summer (JJA) minimum temperature (MinT) (R 2 02=020.47, P 02<020.001, 1953–2009). Hence we produce a regional summer MinT reconstruction spanning the last 21202years. This reconstruction reveals a long-term persistent warming trend, starting in the 1820s, at a rate of 0.4502±020.0902°C/century (1820–2009). This trend is also detected since the 1820s in the Asian summer MinT reconstruction produced by the PAGES 2K project, with a very close warming rate (0.4302±020.0802°C/century, 1820–1989). Our record also displays an enhanced multi-decadal variability since the mid-twentieth century. The 1990s–2000s are the warmest of our whole record, due to the superposition of the gradual warming trend and decadal variability during this interval. The strongest decadal cooling occurs during the 1950s and the largest warming trend during the 1970s. The magnitude of warming from 1973 to 2003 was larger than the total warming trend from 1820s to 2009. Extreme events are also more frequent since 1950. The pattern of multi-decadal variability has similarities with the Atlantic multi-decadal oscillation, suggesting common causality. CMIP5 historical simulations fail to capture both the magnitude and timing of this multi-decadal variability. The ensemble CMIP5 average produces a steady warming trend starting in the 1970s, which only accounts for about 6002% of the observed warming trend during this period. We conclude that TP summer temperature could reflect a climate response to increased greenhouse gas concentrations, however modulated by multi-decadal variations common with the Atlantic sector.
DOI:10.1007/s12229-009-9030-3URL [本文引用: 1]
Because of the characteristically low temperatures and ambient CO 2 concentrations associated with greater altitudes, mountain forests may be particularly sensitive to global warming and increased atmospheric CO 2 . Moreover, the upper treeline is probably the most stressful location within these forests, possibly providing an early bellwether of forest response. Most treeline studies of the past century, as well as recently, have correlated temperatures with the altitudinal limits observed for treelines. In contrast, investigations on pre-establishment seedlings, the most vulnerable life stage of most tree species, are rare. There appears to be specific microclimatic factors dictated by wind and sky exposure that limit seedling survival, and also generate the distorted tree forms commonly observed at treeline. Seedling survival appears critical for creating the biological facilitation of microclimate at the community level which is necessary for the growth of seedlings to normal tree stature, forming new subalpine forest at a higher altitude.
DOI:10.1080/11956860.2002.11682693URL [本文引用: 1]
Climate-growth relationships since the end of the 19th century were analysed by using regional climate data and North Atlantic Oscillation index data, together with spruce (Picea abies (L) Karst.) tree-ring data from an altitudinal gradient close to the coast in central Norway (64 掳N). Correlation and response function analyses were used to decipher both spatial and temporal diameter growth responses. A positive response for May and June temperatures dominated along the entire gradient, but the importance of individual months shifted through time. In periods when May and June temperatures were above their long-term means, the importance of other climate factors increased. Winter climate was significantly related to radial tree growth mainly for periods up to the 1940s, but not during the second half of the 20th century. This pattern was principally caused by changes in the response to winter precipitation. The summer climate acquired increased importance in periods with warm and moist winters. The climate-growth relationship in this study changed continuously and dynamically during the studied period; this might be a general phenomenon that should be carefully considered in dendroclimatological studies. In scenarios for forest development, it is of vital importance to build on detailed knowledge of growth responses to multiple climate variables for all seasons, particularly where the oceanic influence is predicted to be spatially extended.
[本文引用: 1]
DOI:10.1111/1365-2745.12025URL [本文引用: 1]
Fundamental ecological research is both intrinsically interesting and provides the basic knowledge required to answer applied questions of importance to the management of the natural world. The 100th anniversary of the British Ecological Society in 2013 is an opportune moment to reflect on the current status of ecology as a science and look forward to high-light priorities for future work.To do this, we identified 100 important questions of fundamental importance in pure ecology. We elicited questions from ecologists working across a wide range of systems and disciplines. The 754 questions submitted (listed in the online appendix) from 388 participants were narrowed down to the final 100 through a process of discussion, rewording and repeated rounds of voting. This was done during a two-day workshop and thereafter.The questions reflect many of the important current conceptual and technical pre-occupations of ecology. For example, many questions concerned the dynamics of environmental change and complex ecosystem interactions, as well as the interaction between ecology and evolution.The questions reveal a dynamic science with novel subfields emerging. For example, a group of questions was dedicated to disease and micro-organisms and another on human impacts and global change reflecting the emergence of new subdisciplines that would not have been foreseen a few decades ago.The list also contained a number of questions that have perplexed ecologists for decades and are still seen as crucial to answer, such as the link between population dynamics and life-history evolution.Synthesis. These 100 questions identified reflect the state of ecology today. Using them as an agenda for further research would lead to a substantial enhancement in understanding of the discipline, with practical relevance for the conservation of biodiversity and ecosystem function.
DOI:10.1046/j.1440-1703.2003.00577.xURL [本文引用: 1]
DOI:10.1139/x11-096URL [本文引用: 3]
This paper deals with the response of silver fir (Mill.) growth to climate, mainly drought, presenting data from southern Kosovo, for which only little information is available. Two cores from 55 dominant trees were extracted at two sites representing the limits of the natural distribution of silver fir. We built 80- and 112-year-long chronologies for each site. Comparison of both ring width chronologies showed that they were diverse. Pointer year analysis indicated that high spring temperatures and ample rainfall (108202mm) influenced positively silver fir growth (wide ring), while low spring temperatures and scarce precipitation had an inverse effect. Bootstrapped response functions confirmed that April and May temperatures of the current growth year influenced positively silver fir growth at the low-elevation site, while the July precipitation of the previous growth year had a negative influence. At the high-elevation site, warm previous December temperatures had a positive influence on silver fir growth. Drought indices indicated two periods (1953–1961 and 1994–2001) with moderate drought. Correlation analysis between silver fir radial growth and drought indices revealed that the impact of drought on radial growth was significant at the time scale of 1.502months during the summer months.
DOI:10.1175/2012EI000434.1URL [本文引用: 1]
In this study, the authors provide a global assessment of the performance of different drought indices for monitoring drought impacts on several hydrological, agricultural, and ecological response variables. For this purpose, they compare the performance of several drought indices [the standardized precipitation index (SPI); four versions of the Palmer drought severity index (PDSI); and the standardized precipitation evapotranspiration index (SPEI)] to predict changes in streamflow, soil moisture, forest growth, and crop yield. The authors found a superior capability of the SPEI and the SPI drought indices, which are calculated on different time scales than the Palmer indices to capture the drought impacts on the aforementioned hydrological, agricultural, and ecological variables. They detected small differences in the comparative performance of the SPI and the SPEI indices, but the SPEI was the drought index that best captured the responses of the assessed variables to drought in summer, the season in which more drought-related impacts are recorded and in which drought monitoring is critical. Hence, the SPEI shows improved capability to identify drought impacts as compared with the SPI. In conclusion, it seems reasonable to recommend the use of the SPEI if the responses of the variables of interest to drought are not known a priori.
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1306157110URLPMID:23739768 [本文引用: 2]
Zhang et al. (1) report that the Tibetan Plateau experienced a continuous advancing start of green-up date (SOS) from 1982 to 2011 based on the merged Global Inventory Modeling and Mapping Studies (GIMMS)-based with Moderate Resolution Imaging Spectroradiometer (MODIS) [Système Pour l’Observation de la Terre vegetation (SPOT-VGT)]-based SOSs. It challenged...
DOI:10.1007/s10584-015-1467-2URL [本文引用: 2]
中国科学院机构知识库(中国科学院机构知识库网格(CAS IR GRID))以发展机构知识能力和知识管理能力为目标,快速实现对本机构知识资产的收集、长期保存、合理传播利用,积极建设对知识内容进行捕获、转化、传播、利用和审计的能力,逐步建设包括知识内容分析、关系分析和能力审计在内的知识服务能力,开展综合知识管理。
DOI:10.3321/j.issn:1000-0933.2004.11.010URL [本文引用: 1]
运用样带样方法和年轮气候学方法对大海林地区的气候因子和样地数据进行了分析。结果表明 ,近 30 a来老秃顶子地区气候变暖明显 ,尤其是冬季增温最明显 ,月份增温中以 2月份最大 ;寒冷时期 (12月份、翌年 1和 2月份 )和温暖时期 (6~ 9月份 )的温度都有增加 ,但是寒冷时期温度的增加幅度较大 ;冬季与夏季温差稍有减少 ,但积温有所增加 ,整年的热量正在增加。全球变暖导致的大海林地区增温对老秃顶子林线结构特征产生了很大的影响 ,由样地调查和分析可知 ,全球变暖导致林线中上部幼苗、幼树的更新和存活增多 ,森林密度加大 ,树木平均年龄降低 ,年龄结构呈倒 J字型 ,并且多呈聚集分布 ;而在林线的下部 ,幼苗更新很少 ,主要以中龄林存在 ,并且多呈零散分布形式。通过年轮分析得出 ,气候变暖导致林线树木径生长和高生长增加 ,而且增加的趋势和近 30 a来温度的变化基本一致。通过对年轮指数与气候因子的相关性分析 ,表明林线树木年轮指数与温度的相关性较强 ,而与降水的相关性较弱 ,并且年轮指数与温暖时期温度和积温呈正相关 ,而与寒冷时期温度和年平均温度呈负相关 ,表明温暖时期温度和积温控制着林线的海拔高度 ,而寒冷时期的温度和年平均温度主要对林线树种类型起着决定性的作用。从敏感度分析看出 ,林线
DOI:10.3321/j.issn:1000-0933.2004.11.010URL [本文引用: 1]
运用样带样方法和年轮气候学方法对大海林地区的气候因子和样地数据进行了分析。结果表明 ,近 30 a来老秃顶子地区气候变暖明显 ,尤其是冬季增温最明显 ,月份增温中以 2月份最大 ;寒冷时期 (12月份、翌年 1和 2月份 )和温暖时期 (6~ 9月份 )的温度都有增加 ,但是寒冷时期温度的增加幅度较大 ;冬季与夏季温差稍有减少 ,但积温有所增加 ,整年的热量正在增加。全球变暖导致的大海林地区增温对老秃顶子林线结构特征产生了很大的影响 ,由样地调查和分析可知 ,全球变暖导致林线中上部幼苗、幼树的更新和存活增多 ,森林密度加大 ,树木平均年龄降低 ,年龄结构呈倒 J字型 ,并且多呈聚集分布 ;而在林线的下部 ,幼苗更新很少 ,主要以中龄林存在 ,并且多呈零散分布形式。通过年轮分析得出 ,气候变暖导致林线树木径生长和高生长增加 ,而且增加的趋势和近 30 a来温度的变化基本一致。通过对年轮指数与气候因子的相关性分析 ,表明林线树木年轮指数与温度的相关性较强 ,而与降水的相关性较弱 ,并且年轮指数与温暖时期温度和积温呈正相关 ,而与寒冷时期温度和年平均温度呈负相关 ,表明温暖时期温度和积温控制着林线的海拔高度 ,而寒冷时期的温度和年平均温度主要对林线树种类型起着决定性的作用。从敏感度分析看出 ,林线
DOI:10.1016/j.dendro.2007.04.003URL [本文引用: 1]
Dendroclimatological research is often based on the assumption that the relationship between tree growth and climate is not variable over time. Here we test this assumption by exploring if climate sensitivity of Picea mariana (Mill.) trees growing in open-stand lowland forest and on top of a neighboring peatland in Interior Alaska is stable or changing over time. Climate–growth correlations at the study sites are strongly dependent on microtopography and vary substantially over time. Trees growing in the open forest site generally display stronger climate–growth correlations, especially significantly negative correlations with late summer temperatures (July, August) starting in the period 1920–1970. Trees growing on the peatland site are less climate sensitive, but display positive correlations between annual growth and temperature of October and December in the early 20th century, while in the late 20th century, significant negative correlations exist with January and February temperatures. This study, thus, demonstrates a transient climate–growth response for P. mariana (Mill.) on two sites typical for lowland Interior Alaska. However, due to multiple possible explanations (e.g. changing climate, coupled with aging trees and a growing peatland surface) it is not possible at this time to pinpoint the exact cause for these changes in the climate–growth relationships.
DOI:10.1371/journal.pone.0069065URLPMID:3708892 [本文引用: 4]
Abstract It is generally hypothesized that tree growth at the upper treeline is normally controlled by temperature while that at the lower treeline is precipitation limited. However, uniform patterns of inter-annual ring-width variations along altitudinal gradients are also observed in some situations. How changing elevation influences tree growth in the cold and arid Qilian Mountains, on the northeastern Tibetan Plateau, is of considerable interest because of the sensitivity of the region's local climate to different atmospheric circulation patterns. Here, a network of four Qilian juniper (Sabina przewalskii Kom.) ring-width chronologies was developed from trees distributed on a typical mountain slope at elevations ranging from 3000 to 3520 m above sea level (a.s.l.). The statistical characteristics of the four tree-ring chronologies show no significant correlation with increasing elevation. All the sampled tree growth was controlled by a common climatic signal (local precipitation) across the investigated altitudinal gradient (520 m). During the common reliable period, covering the past 450 years, the four chronologies have exhibited coherent growth patterns in both the high- and low-frequency domains. These results contradict the notion of contrasting climate growth controls at higher and lower elevations, and specifically the assumption that inter-annual tree-growth variability is controlled by temperature at the upper treeline. It should be stressed that these results relate to the relatively arid conditions at the sampling sites in the Qilian Mountains.
DOI:10.1073/pnas.1616608114URLPMID:28630302 [本文引用: 1]
Phenological responses of vegetation to climate, in particular to the ongoing warming trend, have received much attention. However, divergent results from the analyses of remote sensing data have been obtained for the Tibetan Plateau (TP), the world’s largest high-elevation region. This study provides a perspective on vegetation phenology shifts during...
DOI:10.1007/s00468-015-1348-3URL [本文引用: 3]
Key message Both temperature and precipitation are strong factors of radial tree growth at all elevations in the semi-arid study area, except at the upper
DOI:10.1007/s11442-015-1226-3URL [本文引用: 1]
There are differing views regarding the climatic factors that limit radial growth of Qilian juniper ( Juniperus przewalskii Kom.) at the upper treelines on the northeastern Tibetan Plateau. In this study, trees from an upper treeline site in the Anyemaqen Mountains of the northeastern Tibetan Plateau were selected to present new evidence that low temperature limits the radial growth of Qilian juniper. Using a signal-free regional curve standardization (SF-RCS) method, a ring-width chronology for Qilian juniper was developed extending from AD 1082 to 2010. The results of correlation analysis between tree-ring index and instrumental climatic variables showed that both winter (December in the previous year and January in the current year) and summer (July and August in the current year) temperatures were significantly and positively correlated with the tree-ring index. Precipitation in June was also significantly and positively correlated with the tree-ring index, but was not as important as low temperature in controlling tree growth. To test the key limiting climatic factor for tree radial growth at different altitudes, an indicator termed the “relative distance to upper treeline” (RDUT) was developed to quantify the representativeness of collected samples for the forest’s upper treeline. The RDUT showed that the upper 20% of the forest belt may be an important boundary in terms of capturing the temperature signal from tree-ring width at the upper treelines on the northeastern Tibetan Plateau. Our findings enhance the existing understanding that temperature is the limiting factor at upper treelines on the northeastern Tibetan Plateau, and will be useful in the reconstruction of past temperature in future studies.
[本文引用: 4]
DOI:10.5194/cpd-10-341-2014URL [本文引用: 3]
A 1342 yr-long tree-ring chronology was developed from Qilian junipers inthe central Qilian Mountains of the north-eastern Tibetan Plateau, China.The climatic implications of this chronology were investigated using simplecorrelation, partial correlation and response function analyses. Thechronology was significantly positively correlated with temperaturevariables during the pre- and current growing seasons, especially withminimum temperature. The variability of the mean minimum temperature fromJanuary to August since 670 AD was then reconstructed based on the tree-ringchronology. The reconstruction explained 58.5% of the variance in theinstrumental temperature records during the calibration period (1960–2011)and captured the variation patterns in minimum temperature at the annual tocentennial time scales over the past millennium. The most recent 50 yrwere the warmest period, while 1690–1880 was the coldest period since670 AD. Comparisons with other temperature series from neighbouring regions andfor the Northern Hemisphere as a whole supported the validity of ourreconstruction and suggested that it provided a good regional representationof temperature change in the north-eastern Tibetan Plateau. The results ofmulti-taper spectral analysis showed the occurrence of significantquasi-periodic behaviour at a number of periods (2–3, 28.8–66.2,113.6–169.5, and 500 yr), which were consistent with those associatedwith El Ni09o-Southern Oscillation (ENSO), Pacific Decadal Oscillation(PDO) and solar activity. Some reconstructed cold events may have closerelationship with the volcanic eruptions.
DOI:10.1007/s10584-016-1695-0URL [本文引用: 3]
Abstract An annually resolved and absolutely dated ring-width chronology spanning 657 yrs. is constructed with Whitebark pine (Pinus bungeana Zucc.) samples from the southern Taihang Mountains, Eastern China. On the basis of a significant correlation between the tree-ring width index and observed instrumental data, precipitation in current May is reconstructed for the region since AD 1510, with predictor variables accounting for 37.9 % of the variance in precipitation data. In agreement with other drought reconstructions, notable dry spells occur in the 1630s–1650s, 1680s–1700s, and 1770s–1800s, whereas wet periods prevail in the 1530s–1570s, 1840s–1870s, and 1950s-present. Wavelet analysis reveals clear 2–8, 20–40, and 80–130 yrs cycles at the 95 % confidence level for the reconstructed series over the past 500 yrs, suggesting possible linkages with the El Ni09o-Southern Oscillation (ENSO) and the Pacific Decadal Oscillation (PDO). Correlation analysis between the tree-ring series, ENSO, and PDO index further demonstrates that precipitation is negatively correlated with PDO and ENSO in the long term.
DOI:10.1016/j.foreco.2010.06.034URL [本文引用: 1]
Divergent responses between tree growth and climate factors have been widely reported at high latitudes in the northern hemisphere. Here we show variable climate-growth relationships and divergent growth responses of Qinghai spruce ( Picea crassfolia) along an elevation gradient at a mid-latitude site at the northeastern Tibetan Plateau, China. Trees from higher elevations, limited mainly by temperature, show divergent growth trends over time and two responses to climate. Some trees show increasing positive and some increasing negative responses to growing season temperature during the last decades. Trees from lower treeline show a strengthening drought stress signal over time and no divergent growth trends within sites. Our results indicate that single tree analysis might be a worthwhile tool to (1) uncover spatial emporal changes in climate-growth relationships of trees, (2) better understand future growth performance and (3) help overcome current limitations of tree ring based climatic reconstructions.
DOI:10.3321/j.issn:1000-1522.2008.03.002URL [本文引用: 1]
通过在青海都兰曲什岗地区建立不同生境的祁连圆柏树轮宽度年表及分析,揭示了不同微环境下祁连圆柏的径向生长对气候变化的响应。从标准轮宽年表的统计特征来看,森林中部年表的平均敏感度、标准差以及序列间的相关系数等统计量均高于其他地点,而凹地年表的上述统计量都较低;各序列之间的相关分析表明,无论在高频变化还是在低频变化上,森林上限年表与其他各采样点的年表均存在较大差异,这种差异在低频变化上尤为明显;采样点轮宽年表与气候资料的相关分析表明,森林中部、下限及水分条件相对较好的凹地,树木径向生长主要与当年6月份降水量之间存在显著的正相关关系,而与同期月平均最高温度之间存在显著的负相关关系。值得提出的是,森林上限祁连圆柏的径向生长与当年7月份温度之间存在极显著的正相关关系。该研究所揭示的不同生境树木生长特性及其对气候响应的差异,一方面对于模拟不同微环境下树木生长对全球变化的响应具有重要意义,另一方面对于利用同一地区的树轮资料重建不同气候要素提供了依据。
DOI:10.3321/j.issn:1000-1522.2008.03.002URL [本文引用: 1]
通过在青海都兰曲什岗地区建立不同生境的祁连圆柏树轮宽度年表及分析,揭示了不同微环境下祁连圆柏的径向生长对气候变化的响应。从标准轮宽年表的统计特征来看,森林中部年表的平均敏感度、标准差以及序列间的相关系数等统计量均高于其他地点,而凹地年表的上述统计量都较低;各序列之间的相关分析表明,无论在高频变化还是在低频变化上,森林上限年表与其他各采样点的年表均存在较大差异,这种差异在低频变化上尤为明显;采样点轮宽年表与气候资料的相关分析表明,森林中部、下限及水分条件相对较好的凹地,树木径向生长主要与当年6月份降水量之间存在显著的正相关关系,而与同期月平均最高温度之间存在显著的负相关关系。值得提出的是,森林上限祁连圆柏的径向生长与当年7月份温度之间存在极显著的正相关关系。该研究所揭示的不同生境树木生长特性及其对气候响应的差异,一方面对于模拟不同微环境下树木生长对全球变化的响应具有重要意义,另一方面对于利用同一地区的树轮资料重建不同气候要素提供了依据。
DOI:10.1007/s11434-008-0400-8URL [本文引用: 1]
The climate of the past 1000 years is an important context for evaluating the recent climate warming.However,there are few 1000-year-long climate reconstructions with annual resolution in the Qing-hai-Tibet Plateau.In this paper,a dendroclimatic analysis was conducted for the radial growth of Qilian juniper from the upper forest limit in Wulan,Qinghai Province.The results of correlation analysis between the tree-ring widths and the climate variables indicate that the growth of junipers at the upper forest limit is mainly limited by low temperatures of September,November and February of the pregrowth season,and July of the current growth season.There is no significant correlation between the tree-ring widths and precipitation.A mean temperature from the previous year's September to the current year's April was reconstructed for the Wulan area since A.D.1000.The reconstruction can explain 40.8% of the instrumental variance in the calibration period(1856―2002).The reconstruction shows that the 20th century is the warmest 100 years,and the 1990s is the warmest decade during the past 1000 years,while the coldest 100 years and decade occur at 1600―1699 and 1642―1651,respectively.The variations are verified well by the temperature reconstruction of the middle Qilian Mountain(QL) and the total organic carbon(TOC) in the Qinghai Lake sediments.The comparison of our reconstruction with the annual temperature reconstruction of extra-tropical Northern Hemisphere suggests that the climate of Wulan during the Medieval Warm Period is of obvious regional specialty,but there was a good response to the climate of hemispheric scales during the recent 400 years.
DOI:10.1007/s10021-015-9849-xURL [本文引用: 1]
Climate warming is predicted to extend the duration and enhance the severity of summer droughts in Central Europe, which may pose a serious risk to forest productivity and forest health. Fagus sylvatica (European beech), the most abundant tree species of Central Europe natural forest vegetation and one of the key species in forestry, is thought to be particularly vulnerable to drought. Here, we present a dendrochronological analysis in three mixed temperate broad-leaved forests along a precipitation gradient with the aim of comparing the climatic response of radial growth of F. sylvatica with the performance of four co-existing species ( Acer pseudoplatanus , A. platanoides , Quercus petraea , Fraxinus excelsior ). We hypothesized that Fagus is the most drought sensitive of the five species, which implies that it could lose its competitive advantage at drier sites in the course of climate warming. In support of this hypothesis, we found that F. sylvatica in all stands exhibited an increase in the number of negative pointer years and a decrease in radial increment in the driest stand since about 1980, in parallel to increasing summer temperatures and drought intensity. Such a response was missing in the other four species and may point to shifts in the competitive hierarchy in these mixed forests under a future warmer climate. We conclude that Central Europe forestry sector should consider carefully the risk of failure of beech in regions with relatively low and decreasing summer precipitation.
Climate change 2013: The physical science basis. An overview of the Working Group I contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC)
1
2013
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
The effect of competition on responses to drought and interannual climate variability of a dominant conifer tree of western North America
2
2016
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
... )类似.在低降水区域(HL) , 极端年份气候波动对祁连圆柏生长-气候关系影响不明显; 在中降水区域(QS), 极端年份水分变化对生长-气候关系影响显著.在生长季前干旱年, 树木生长主要受春夏季最低气温和夏季降水限制, 而在生长季(或季前)湿润年, 生长主要受春季降水和夏季气温限制(
Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge
1
2017
... id="C11">气象数据从中国气象科学数据共享服务网(
Program ARSTAN User Manual
1
1984
... id="C10">本研究野外调查及取样工作于2016年7-8月进行, 在HL、QS和HB 3个采样点高山林线处, 以生长于最高处的树木为海拔上限, 各设置3条垂直于林线的30 m × 100 m的矩形样带, 以样带下限左下角为原点, 记录样带内所有树木的坐标和胸径, 利用内径为5.15 mm的生长锥(HaGlof, Dalarna, Sweden)在胸高(约1.3 m)处进行树木年轮取样, 对胸径介于10-20 cm之间的树木一树一芯, 对胸径大于20 cm的树木一树两芯, 胸径低于10 cm的树木不取样芯, HL、QS和HB采样点, 调查树木773株, 共取得636根树木年轮样芯.将取得的样芯编号并置于塑料管中, 带回实验室后对样芯进行固定、风干、打磨等预处理(
1
2005
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
1
2005
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Daily air temperature interpolated at high spatial resolution over a large mountainous region
1
1997
... id="C11">气象数据从中国气象科学数据共享服务网(
1
1999
... id="C6">本研究区位于青藏高原东北部, 属高原干旱大陆性气候, 研究区内自西北向东南降水量逐渐增加(
1
1999
... id="C6">本研究区位于青藏高原东北部, 属高原干旱大陆性气候, 研究区内自西北向东南降水量逐渐增加(
Quaking aspen (Populus tremuloides Michx.) at treeline: A century of change in the San Juan Mountains, Colorado, USA
1
2004
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Tree and timberline shifts in the northern Romanian Carpathians during the Holocene and the responses to environmental changes
1
2016
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Climate-growth analysis of Qilian juniper across an altitudinal gradient in the central Qilian Mountains, northwest China
1
2012
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Climate and growth of Pinus sylvestris at its upper altitudinal limit in Scotland: Evidence from tree growth-rings
1
1990
... id="C27">冬季低温和降雪少引起的土壤低温会损伤树木, 暖冬可提高土壤温度, 降雪增加雪被厚度, 保证树木根系不受低温伤害, 维持树木根系生长及各项生理功能正常发挥的能力(
Are treelines advancing? A global meta-analysis of treeline response to climate warming
1
2009
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales
1
2005
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
近50年青藏高原东部降水的时空变化特征
1
2013
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
近50年青藏高原东部降水的时空变化特征
1
2013
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Temporal changes in climatic limitation of tree-growth at upper treeline forests: Contrasted responses along the west-to- east humidity gradient in Northern Patagonia
2
2015
... id="C28">气温过高会加剧呼吸作用对自身营养物质的消耗, 并且高温会加速土壤水分蒸散, 使得树木可利用水分降低; 气温过高将促进呼吸作用和蒸腾作用, 从而消耗树木所固定的有机物, 同时高温将加速土壤水分蒸发, 进而导致水分亏缺使生长受限(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
Microclimatological investigations in the tropical alpine scrub of Maui, Hawaii: Evidence for a drought-induced alpine timberline
1991
1
2000
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
1
2000
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature?
5
2014
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... id="C27">冬季低温和降雪少引起的土壤低温会损伤树木, 暖冬可提高土壤温度, 降雪增加雪被厚度, 保证树木根系不受低温伤害, 维持树木根系生长及各项生理功能正常发挥的能力(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan Plateau
1
2010
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
1961-2005年来青藏高原主要气候因子的基本特征
1
2010
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
1961-2005年来青藏高原主要气候因子的基本特征
1
2010
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Spatial and temporal variability in the growth and climate response of treeline trees in Alaska
2
2002
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
Patterns and dynamics of treeline advance on the Seward Peninsula, Alaska
2002
Tree-ring based summer minimum temperature reconstruction for the southern edge of the Qinghai-Tibetan Plateau, China
1
2013
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Growth-mortality relationships in pinon pine (Pinus edulis) during severe droughts of the past century: Shifting processes in space and time
3
2014
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
... ), 在低温年(PLT和GLT)、湿润年(PW和GW)和生长季湿润年, 树木生长对气候变化不敏感(
... ;
Surface temperature lapse rates over complex terrain: Lessons from the Cascade Mountains
1
2010
... id="C11">气象数据从中国气象科学数据共享服务网(
Extracting more out of relocation ata: Building movement models as mixtures of random walks
1
2004
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Climate- driven increases in global terrestrial net primary production from 1982 to 1999
1
2003
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
4
2012
... id="C28">气温过高会加剧呼吸作用对自身营养物质的消耗, 并且高温会加速土壤水分蒸散, 使得树木可利用水分降低; 气温过高将促进呼吸作用和蒸腾作用, 从而消耗树木所固定的有机物, 同时高温将加速土壤水分蒸发, 进而导致水分亏缺使生长受限(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
... ), 气温升高加快呼吸作用对树木自身营养物质的消耗(
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
4
2012
... id="C28">气温过高会加剧呼吸作用对自身营养物质的消耗, 并且高温会加速土壤水分蒸散, 使得树木可利用水分降低; 气温过高将促进呼吸作用和蒸腾作用, 从而消耗树木所固定的有机物, 同时高温将加速土壤水分蒸发, 进而导致水分亏缺使生长受限(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
... ), 气温升高加快呼吸作用对树木自身营养物质的消耗(
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
坡向对海拔梯度上祁连圆柏树木生长的影响
2
2010
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
... ;
坡向对海拔梯度上祁连圆柏树木生长的影响
2
2010
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
... ;
Difference in tree growth responses to climate at the upper treeline: Qilian juniper in the Anyemaqen Mountains
1
2008
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Mediterranean and temperate treelines are controlled by different environmental drivers
1
2016
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Climate-driven speedup of alpine treeline forest growth in the Tianshan Mountains, Northwestern China
1
2015
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Is precipitation a trigger for the onset of xylogenesis in Juniperus przewalskii on the north-eastern Tibetan Plateau?
2
2015
... id="C28">气温过高会加剧呼吸作用对自身营养物质的消耗, 并且高温会加速土壤水分蒸散, 使得树木可利用水分降低; 气温过高将促进呼吸作用和蒸腾作用, 从而消耗树木所固定的有机物, 同时高温将加速土壤水分蒸发, 进而导致水分亏缺使生长受限(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
Interspecific variation in growth responses to climate and competition of five eastern tree species
1
2016
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Unprecedented recent warming rate and temperature variability over the east Tibetan Plateau inferred from alpine treeline dendrochronology
2
2014
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... ), 本研究不支持干旱区林线树木生长主要受水分限制的假说, 但降水量变化会影响林线树木生长与气温的关系.在本研究中, 降水对不同水分梯度下林线树木生长的影响方式相同, 但气温主要以两种方式影响树木径向生长, 分别为: 低温限制作用(
The altitude of alpine treeline: A bellwether of climate change effects
1
2009
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Shifts in radial growth responses of coastal Picea abies induced by climatic change during the 20th Century, central Norway
1
2016
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
1
1968
... id="C10">本研究野外调查及取样工作于2016年7-8月进行, 在HL、QS和HB 3个采样点高山林线处, 以生长于最高处的树木为海拔上限, 各设置3条垂直于林线的30 m × 100 m的矩形样带, 以样带下限左下角为原点, 记录样带内所有树木的坐标和胸径, 利用内径为5.15 mm的生长锥(HaGlof, Dalarna, Sweden)在胸高(约1.3 m)处进行树木年轮取样, 对胸径介于10-20 cm之间的树木一树一芯, 对胸径大于20 cm的树木一树两芯, 胸径低于10 cm的树木不取样芯, HL、QS和HB采样点, 调查树木773株, 共取得636根树木年轮样芯.将取得的样芯编号并置于塑料管中, 带回实验室后对样芯进行固定、风干、打磨等预处理(
Identification of 100 fundamental ecological questions
1
2013
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Effects of climate on the radial growth of tree species in the upper and lower distribution limits of an altitudinal ecotone on Mount Norikura, central Japan
1
2003
... id="C11">气象数据从中国气象科学数据共享服务网(
Growth responses to climate and drought in silver fir (Abies alba) along an altitudinal gradient in southern Kosovo
3
2011
... id="C27">冬季低温和降雪少引起的土壤低温会损伤树木, 暖冬可提高土壤温度, 降雪增加雪被厚度, 保证树木根系不受低温伤害, 维持树木根系生长及各项生理功能正常发挥的能力(
... ).生长季前低温导致树木根系生长受阻, 生理活动能力受限, 吸收土壤水分的能力较弱(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
Performance of drought indices for ecological, agricultural, and hydrological applications
1
2012
... id="C11">气象数据从中国气象科学数据共享服务网(
生态交错带与生物多样性
1
1997
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
生态交错带与生物多样性
1
1997
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Declining snow cover may affect spring phenological trend on the Tibetan Plateau
2
2013
... id="C27">冬季低温和降雪少引起的土壤低温会损伤树木, 暖冬可提高土壤温度, 降雪增加雪被厚度, 保证树木根系不受低温伤害, 维持树木根系生长及各项生理功能正常发挥的能力(
... ), 也会使覆雪期和霜冻期延长, 推迟生长季开始的时间(
Differential response of Qilian juniper radial growth to climate variations in the middle of Qilian Mountains and the northeastern Qaidam Basin
2
2015
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... , 2015a;
气候变暖对老秃顶子林线结构特征的影响
1
2004
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
气候变暖对老秃顶子林线结构特征的影响
1
2004
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Changing climate sensitivity of black spruce (Picea mariana Mill.) in a peatlandforest landscape in Interior Alaska
1
2008
... id="C4">气候变化对地球植被的生长、生存及分布等有巨大影响, 甚至会加速物种消亡(
Climate control on tree growth at the upper and lower treelines: A case study in the Qilian Mountains, Tibetan Plateau
4
2013
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... )和高温胁迫作用(
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data
1
2017
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Growth climate relationships along an elevation gradient on a southeast- facing mountain slope in the semi-arid eastern Qaidam Basin, northeastern Tibetan Plateau
3
2016
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... ;
a). Which climatic factors limit radial growth of Qilian juniper at the upper treeline on the northeastern Tibetan Plateau?
1
2015
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
b). Forward modeling analyses of Qilian Juniper (Sabina przewalskii) growth in response to climate factors in different regions of the Qilian Mountains, northwestern China
4
2015
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... ;
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
Millennial minimum temperature variations in the Qilian Mountains, China: Evidence from tree rings
3
2014
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... ;
... id="C29">不同降水梯度下祁连圆柏树木径向生长的气候限制因子存在明显差异.一般认为随降水增加, 降水对树木生长的限制作用减弱, 温度限制作用增加(
500-yr. precipitation variability in Southern Taihang Mountains, China, and its linkages to ENSO and PDO
3
2016
... id="C26">青藏高原及其周边地区高山树线处, 树木生长既存在温度限制也存在水分限制(
... ,
... id="C30">同一采样点, 在不同气候特征年(低温、高温、干旱、湿润), 高山林线祁连圆柏径向生长的限制因子也存在差异, 并且这种差异在不同降水梯度下也有所不同, 这与其他研究结果(
Divergent growth responses and increasing temperature limitation of Qinghai spruce growth along an elevation gradient at the northeast Tibet Plateau
1
2010
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
不同生境祁连圆柏径向生长对气候变化的响应
1
2008
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
不同生境祁连圆柏径向生长对气候变化的响应
1
2008
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Millennial temperature reconstruction based on tree-ring widths of Qilian juniper from Wulan, Qinghai Province, China
1
2008
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (
Climate warming-related growth decline affects Fagus sylvatica, but not other broad-leaved tree species in central European mixed forests.
1
2015
... id="C5">青藏高原东北部分布着大面积的高山林线, 该区自然环境条件严酷, 其生态系统十分脆弱且对气候变化响应极其敏感, 生态环境和森林资源逐渐恶化, 了解该区气候变化对林线树木的影响及气候限制机制尤为重要.青藏高原东北部近50年来年平均气温上升约1.3 ℃ (

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}