 ,1,*1
,1,*1 2
Hierarchical responses of plant stoichiometry to phosphorus addition in an alpine meadow community
SUN Xiao-Mei1, CHEN Jing-Jing2, LI Jin-Xia1, LI Liang1, HAN Guo-Jun1, CHEN Nian-Lai,1,*1 and 2
通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1262KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孙小妹, 陈菁菁, 李金霞, 李良, 韩国君, 陈年来. 高寒草甸植物化学计量比对磷添加的分层响应. 植物生态学报[J], 2018, 42(1): 78-85 doi:10.17521/cjpe.2017.0253
SUN Xiao-Mei.
生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(Sterner & Elser, 2002), 利用种间植物组织中养分含量及其计量比特征的差异反映出土壤中养分条件、植物特征和生理机制的差异(Méndez & Karlsson, 2005; Ventura et al., 2008)。例如营养资源添加, 扰动土壤-植物的化学计量比过程, 植物中主要的生产组织会改变它们的营养元素含量与计量比来适应外界变化的养分生境。其中能够反映植物生存策略的C:N、C:P不仅反映植物的生长速率(Sun et al., 2016), 而且与植物的N、P利用策略有关(Han et al., 2014), 植物N:P在一定程度上能反映植物生长的养分限制元素(Güsewell, 2004)。
N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(Suding et al., 2005; Pe?uelas et al., 2013; Yan et al., 2015; Koerner et al., 2016)。增加的N、P沉降通过打破C、N和P在植物体内的平衡, 改变了植物的C:N:P计量比特征。先前的研究发现P的富集通常会增加植物体内的P含量, 相应降低C:P (Mayor et al., 2014; Hong et al., 2015; Mao et al., 2016); 然而P富集对植物体内N含量、C:N和N:P的影响效应通常跟植被限制元素类型相关。例如, 受P限制的生态系统, P添加使植物迅速增长, 由于稀释效应通常使植物N含量下降, 故而C:N增加而N:P下降(Feller et al., 2007; Yuan & Chen, 2015)。而在N限制的生态系统, 由于物种间不同的养分利用策略, N含量、C:N和N:P对P添加的响应无一致性规律(Yuan & Chen, 2015)。因此, 关于植物计量比特征对P添加的响应还有待进一步验证。
由于物种间对P富集的差异性响应会影响物种的生长速率而改变物种的竞争能力, 使群落的物种组成和构成发生变化。为了具体探究P富集导致哪些物种丧失, 基于功能群的物种多样性研究是综合评价物种和群落对环境波动响应的有效工具(Sun et al., 2016)。因为功能群是生态系统中物种的生长模式、特征和对环境因素响应特征相似的集合体, 例如在资源匮乏的生境中, 不同的功能群对限制因子采取不同的策略, 可能影响竞争层级而决定群落的构成和结构(Wang et al., 2013)。所以P富集不仅改变了物种水平的生态化学计量比过程, 而且调节了功能群水平的计量比特征。 而基于功能群的计量比特征更能准确反映物种间对于P富集响应的差异性, 从而对群落的构成和结构的改变做出解释。因此, 我们以已经探明受N限制的高寒草甸群落为研究对象(Sun et al., 2016), 分析P添加对物种水平和功能群水平C、N、P含量以及C:N、C:P和N:P的影响, 探究P添加对群落物种构成改变的内在机制, 为天然草原的保护和恢复提供参考资料。
1 材料和方法
1.1 研究区概况
本实验在甘肃省合作市(34.92° N, 102.88° E; 海拔3 000 m)进行。该地气候类型为寒温湿润的高原气候, 年平均气温2.4 ℃, 年降水量545 mm, 年蒸发量为1β200 mm。植被类型为多年生草本植物占优势的亚高寒草甸, 主要物种为禾本科的垂穗披碱草(Elymus nutans)和羊茅(Festuca ovina), 莎草科的矮嵩草(Kobresia humilis)和藨草(Scirpus triqueter), 豆科的黄花棘豆(Oxytropis ochrocephala)和黄花苜蓿(Medicago falcata), 以及杂类草川甘蒲公英(Taraxacum lugubre)、甘青蒿(Artemisia tangutica)、珠芽蓼(Polygonum viviparum)和甘青老鹳草(Geranium pylzowianum)等。1.2 试验设计
本试验采用完全随机设计, 于2009年在青藏高原高寒草甸群落设定不同浓度的P添加实验, 设定4个处理(C、P5、P10、P15), P添加量分别为0、5、10、15 g·m-2·a-1, 所施磷肥为磷酸二氢钠(NaH2PO4)。每个处理设3个重复, 共12个小区, 每小区面积为5 m × 5 m。各小区间有1 m的缓冲带, 缓冲带不施肥。每个小区内有4个亚样方(0.5 m × 0.5 m), 其中两个用于长期物种、多度与盖度的调查, 一个用于地上生物量的测定, 另外一个用于植株高度测定。为使肥料效应能充分体现出对群落结构的影响, 施肥于试验的前一年8月底至9月初进行。通常选择雨天添加营养元素, 使肥料颗粒溶解同时满足施肥后植物对水分的需求(Luo et al., 2006)。1.3 样品采集与室内测定
分别在施肥样地中选取群落共有物种作为研究对象, 包括: 垂穗披碱草(禾本科)、矮嵩草(莎草科)、黄花棘豆(豆科)、川甘蒲公英(莲座状杂类草)与甘青老鹳草(直立型杂类草)。地上净生物量产出测定于2013年8月中旬齐地面剪取每个处理地上部分装袋, 3次重复。单独封装5个典型种, 其他物种按照禾本科、莎草科、豆科、杂类草4类功能群单独装袋, 带回实验室60 ℃烘干48 h至恒质量并称量记录(精确到0.001 g), 然后用于C、N、P含量的测定。4类功能群C、N、P含量的测定中不包括5个典型种的植物样。植物样地上部分经研磨后用80 μm的筛子过筛。采用重铬酸钾容量法测定植物全C (外加热法); 采用HClO4-H2SO4消煮后的溶液经定容、沉淀和稀释后用SmartChem 200化学分析仪(WestCo Scientific Instrument, Brookfielg, USA)分析测定样品全N和全P (Sun et al., 2016)。1.4 数据分析
方差分析采用Duncan多重比较法比较不同处理间在物种水平上5个典型物种地上部分C、N、P含量及计量比特征和物种优势度的差异, 在功能群水平上4个功能群的C、N、P含量及计量比特征和生物量产出对P添加响应的差异, 以及同一处理不同功能群间的指标差异。物种优势度用相对生物量来表示, 也就是每个物种的地上生物量占群落的生物量的百分数。以上统计分析都是在SPSS 16.0中完成, 作图用GraphPad Prism。2 结果和分析
2.1 P添加对物种优势度和各功能群生物量产出的影响
比较4个功能群地上生物量平均值, 禾本科>杂类草>莎草>豆科的生物量积累(图1A)(p < 0.05)。禾本科地上生物量随P添加量的增加而显著上升(p < 0.05), 而杂类草的地上生物量显著下降(p < 0.01), P添加对莎草科和豆科物种的地上生物量影响不显著(图1A)。图1
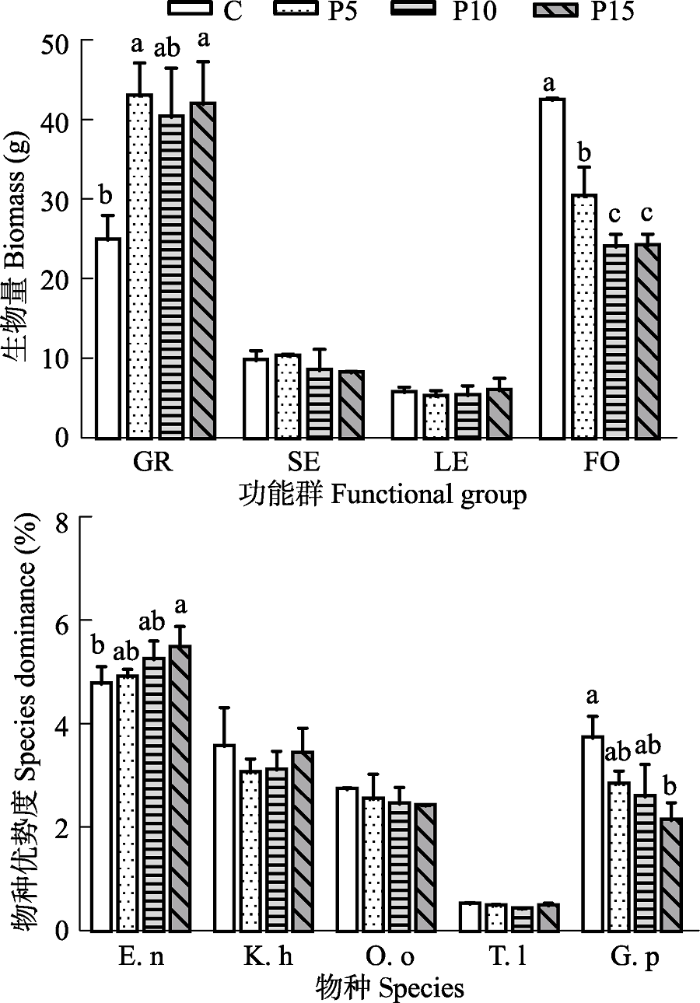 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1P添加对典型种物种优势度和各功能群生物量(平均值±标准误差, n = 3)的影响。C、P5、P10、P15分别指P添加量为0、5、10、15 g·m-2·a-1。GR, 禾本科; SE, 莎草科; LE, 豆科; FO, 杂类草; E.n, 垂穗披碱草; K.h, 矮嵩草; O.o, 黄花棘豆; T.l, 川甘蒲公英; G.p, 甘青老鹳草。不同小写字母表示处理间差异显著(p < 0.05)。
Fig. 1Effects of P addition on species dominance of exemplary species and biomass of each functional group (mean ± SE, n = 3). C, P5, P10, P15, nutrient addition 0, 5, 10, 15 g·m-2·a-1. GR, grasses; SE, sedges; LE, legumes; FO, forbs; E.n, Elymus nutans; K.h, Kobresia humili; O.o, Oxytropis ochoocephala; T.l, Taraxacum lugubre; G.p, Geranium pylzowianum. Different lowercase letters indicate significant differences between treatments (p < 0.05).
比较物种优势度发现, 垂穗披碱草的物种优势度最高, 川甘蒲公英的最低(图1B)。物种优势度与各功能群地上生物量产出对P添加的响应规律基本一致。禾本科物种垂穗披碱草的物种优势度随P添加显著增加(p < 0.05); 莎草科的矮嵩草和豆科黄花棘豆的物种优势度对P添加响应不显著; 杂类草中莲座状杂类草川甘蒲公英对P添加响应不显著, 而直立型杂类草甘青老鹳草的物种优势度随P添加显著下降, 与杂类草对P添加的响应规律一致(图1B)。
2.2 物种水平和功能群水平C、N、P含量对P添加的响应
除豆科(不含典型物种)外, C含量在其他3个功能群(不含典型物种)和物种水平对P添加的响应规律一致, P添加对C含量的影响不显著(图2A、2B)。禾本科和莎草科的N含量对P添加在物种水平和功能群水平(不含典型物种)上的响应规律一致, 均表现出显著降低(p < 0.05); 而豆科在物种水平上显著增加, 但在功能群水平上(不含典型物种)无显著差异; P添加对杂类草功能群水平上(不含典型物种)的N含量无显著影响, 但是物种水平上莲座状物种川甘蒲公英的N含量随P添加显著下降, 而直立型杂类草甘青老鹳草则显著上升(p < 0.05)。在物种水平和功能群水平上(不含典型物种), P添加后, 均是豆科系列的植物N含量显著高于其他物种和功能群(p < 0.05)(图2C、2D)。
图2
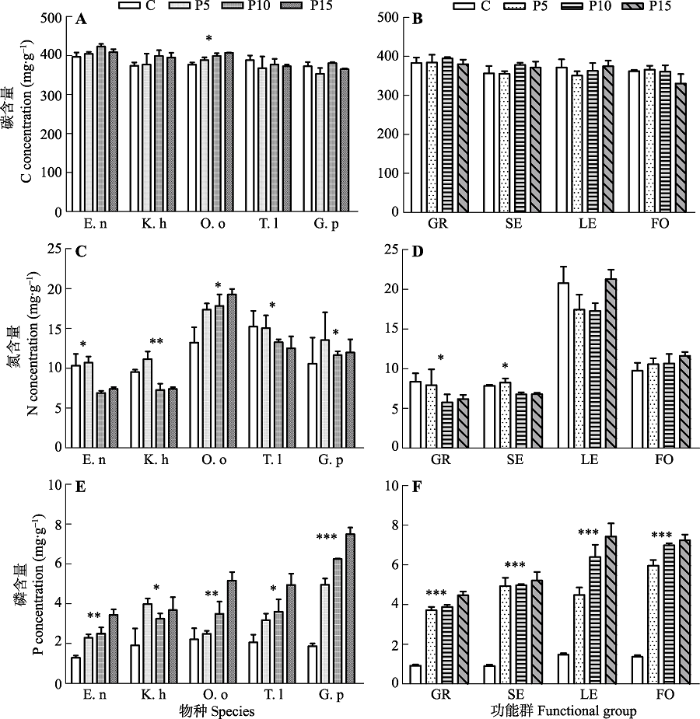 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2P添加对物种水平和功能群水平C、N、P含量的影响(平均值±标准误差, n = 3)。Duncan’s多重比较: ***, p < 0.001; **, p < 0.01; *, p < 0.05。C、P5、P10、P15分别指P添加量为0、5、10、15 g·m-2·a-1。GR, 禾本科; SE, 莎草科; LE, 豆科; FO, 杂类草; E.n, 垂穗披碱草; K.h, 矮嵩草; O.o, 黄花棘豆; T.l, 川甘蒲公英; G.p, 甘青老鹳草。
Fig. 2Effects of P addition on C, N and P concentration at exemplar species and functional group levels (mean ± SE, n = 3). ***, p < 0.001; **, p < 0.01; *, p < 0.05 among treatments following Duncan’s multiple range tests. C, P5, P10, P15, nutrient addition 0, 5, 10, 15 g·m-2·a-1. GR, grasses; SE, sedges; LE, legumes; FO, forbs; E.n, Elymus nutans; K.h, Kobresia humili; O.o, Oxytropis ochoocephala; T.l, Taraxacum lugubre; G.p, Geranium pylzowianum.
植物P含量在4个功能群水平(不含典型物种)和相应物种水平对P素添加的响应规律一致, 均随P素添加量增加显著增加(p < 0.05)(图2E、2F)。与对照相比, 杂类草的P含量相较于其他3个物种和功能群的增幅要高。
2.3 物种水平和功能群水平计量比特征对P添加的响应
比较物种间和功能群间(不含典型物种)的C:N大小, 垂穗披碱草和矮嵩草的C:N平均值显著高于其他3个物种, 功能群禾本科的C:N平均值要显著高于其他3个功能群(图3A、3B)。5个代表种的C:N对P添加的响应规律不同, 其中垂穗披碱草和矮嵩草的C:N随P添加量增加显著增加(p < 0.05); 而黄花棘豆和甘青老鹳草的C:N在对照处理显著大于P添加处理, 而P5、P10和P15处理间差异不显著; P添加对川甘蒲公英的C:N影响不显著(图3A)。功能群水平上(不含典型物种)禾本科和莎草科的C:N对P添加的响应规律分别与物种水平的响应规律一致(p < 0.05); P添加对功能群豆科(不含典型物种)的C:N影响不显著, 而显著降低功能群杂类草(不含典型物种)的C:N, 这与直立型杂类草甘青老鹳草的响应规律一致(图3B)。图3
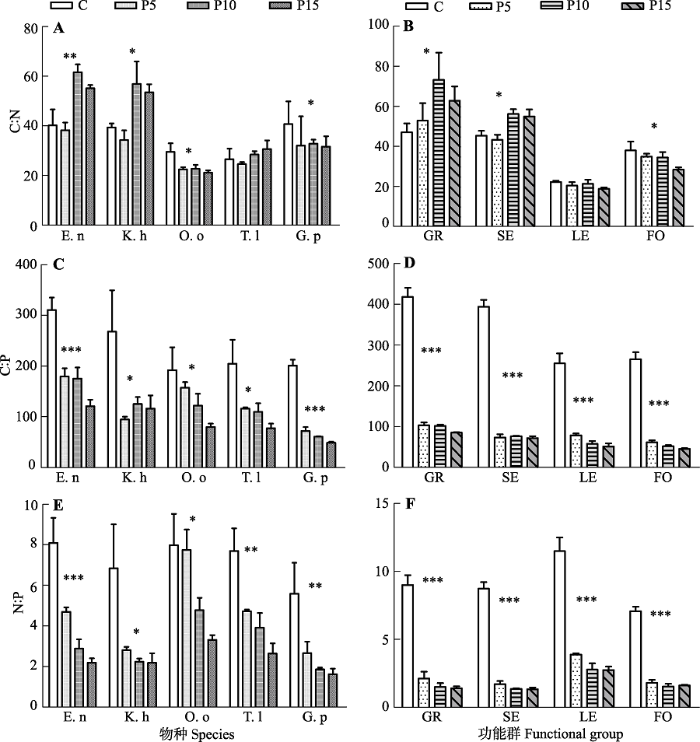 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3P添加对物种水平和功能群水平计量比特征的影响(平均值±标准误差, n = 3)。Duncan’s多重比较: ***, p < 0.001; **, p < 0.01; *, p < 0.05。C、P5、P10、P15分别指P添加量为0、5、10、15 g·m-2·a-1。GR, 禾本科; SE, 莎草科; LE, 豆科; FO, 杂类草; E.n, 垂穗披碱草; K.h, 矮嵩草; O.o, 黄花棘豆; T.l, 川甘蒲公英; G.p, 甘青老鹳草。
Fig. 3Effects of P addition on C:N:P stoichiometric characters at exemplar species and functional groups-levels (mean ± SE, n = 3). ***, p < 0.001; **, p < 0.01; *, p < 0.05 among treatments following Duncan’s multiple range tests. C, P5, P10, P15, nutrient addition 0, 5, 10, 15 g·m-2·a-1. GR, grasses; SE, sedges; LE, legumes; FO, forbs; E.n, Elymus nutans; K.h, Kobresia humili; O.o, Oxytropis ochoocephala; T.l, Taraxacum lugubre; G.p, Geranium pylzowianum.
比较物种间的C:P值大小, 垂穗披碱草的C:P显著高于其他4个物种(图3C)。随P添加量的增加, 5个代表种的C:P显著下降(p < 0.05)。4个功能群水平(不含典型物种)上C:P的变化规律跟相应物种水平上的变化规律一致, 均随土壤P添加量的增加而显著下降(p < 0.05)(图3D)。
物种水平上N:P随P添加的变化规律与C:P相似, 随P添加量的增加N:P显著下降(p < 0.05); 且在任何处理下, 5个代表种的N:P < 10 (图3E)。在功能群水平上, 4个功能群(不含典型物种)的N:P与相应物种水平上的变化规律一致, 随P添加量的增加N:P显著下降(p < 0.001); 且在P添加处理下, 4个功能群的N:P < 13 (图3F)。
3 结论和讨论
我们发现在青藏高原高寒草甸连续5年添加P显著改变了植物的营养物质含量, 故而植物的生态化学计量比也相应发生了变化, 并且物种水平和功能群水平的响应规律基本一致。在P添加处理下, 除豆科外, C含量在其他3个功能群和相应物种水平对P添加的响应规律一致: P添加对C含量的影响不显著(图2A、2B)。Mao等(2016)在我国东北的温带湿地添加不同梯度的P, 结果发现P添加对优势种小叶章(Deyeuxia angustifolia)与狭叶甜茅(Glyceria spiculosa)的整株有机碳影响也不显著。由于组成植物体组织的结构性物质(C)较功能性和贮藏性物质(N、P)受环境的影响较小而相对稳定(Sterner & Elser, 2002), 这是植物组织C含量不随土壤P添加量的增加而增大的原因。P添加显著增加了物种水平和功能群水平植物组织中的P含量(图2E、2F), 相应降低了C:P (图3C、3D), 这与之前的研究结果相一致(Mayor et al., 2014; Hong et al., 2015; Yuan & Chen, 2015; Chen et al., 2016; Li et al., 2017)。土壤P含量与土壤母质的理化性质相关, 土壤中的P主要源于岩石的风化和淋洗。因此, 土壤P在土壤发育时间较长、温暖地区被高度淋溶的土壤中是植物生长主要的限制因子; 而岩石中N的风化过程是一个温度敏感的生物过程(Heerwaarden et al., 2003; Reich & Oleksyn, 2004)。青藏高原高寒草甸由于低温限制微生物的活性和抑制土壤中有机质的分解和矿化, 土壤N是主要的限制因子。我们之前的研究也表明, 在青藏高原高寒草甸N是提高地上净生物量产出的关键限制因子(Sun et al., 2016)。因此, 本研究中物种水平和功能群水平植物组织中的C:P对P添加的负响应归因于物种对非限制营养元素P的奢侈利用。
随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(图2C、2D; 图3A、3B)。先前的研究发现P添加对植物N含量的影响有正效应(Iversen et al., 2010; Mao et al., 2016)、负效应(Feller et al., 2007; Ostertag, 2010)或者无显著影响(Mayor et al., 2014; Yan et al., 2015)。这些结果的不一致是由于植物物种养分利用策略的差异性以及生境中可利用养分的不同造成的(Danger et al., 2008; Townsend & Asner, 2013)。P添加使豆科植物的N含量和C:N在物种水平上分别显著增加和降低, 但是功能群水平上无显著影响(图2C、2D; 图3A、3B)。由于豆科植物能与固氮菌结合形成互惠共生体, 通过生物固氮作用一般能满足豆科植物对氮的需求, 因此相比于其他功能群, 豆科植物更倾向于P限制(Dybzinski et al., 2008; Zhang et al., 2011)。P对豆科作物的生长和结瘤固氮均有重要的作用, 苗淑杰等(2006)的研究表明随着P添加浓度的增加, 大豆(Glycine max)的根瘤数、根瘤干质量和植株N含量均显著增加。然而陈凌云等(2010)发现, 氮磷复合肥添加对豆科植物花苜蓿(Medicago ruthenica)、异叶米口袋(Gueldenstaedtia diversifolia)和甘肃棘豆(Oxytropis kansuensis)叶片的N含量无显著影响, 主要增加了豆科植物对P的养分吸收。植物对N、P的吸收能力具有物种特异性(Güsewell, 2004)可能是造成P添加使豆科和杂类草物种水平和功能群水平N含量响应规律不一致的原因。随P添加量增加, 禾本科在功能群和物种水平上N含量显著降低, 而C:N显著增加。由于禾草具有庞大的根系系统和更快的生长速率, 能够迅速吸收与利用土壤中的可溶性N、P等速效养分, 使叶片积累大量的N以保证高效的光合效率, 随着光合产物的积累, 固定的碳水化合物无法及时地输出至其他器官, 造成了叶片中N的稀释(Evans, 1989)。故而稀释效应可能是造成P添加使禾本科物种水平和功能群水平N含量显著下降, C:N增加的原因。
P添加显著增加了物种水平和功能群水平的P含量, 尽管N含量有增加有降低, 但是参试物种和功能群水平的N:P均显著下降(图3E、3F)。这与其他受N限制的森林(Ostertag, 2010)、草地(Li et al., 2011)和亚北极苔原(de Long et al., 2016)生态系统的研究结果一致。本研究中发现P处理使物种水平的N:P < 10, 而功能群水平的N:P < 13 (图3E、3F)。考虑到植物的N:P比可作为判断N限制或P限制的一个重要指标, N:P < 13和> 16分别表示植被受N限制和P限制(Güsewell, 2004), 我们的结果表明P的添加潜在加剧了高寒草甸群落的N限制。
叶片C:N和C:P能够反映出植物对N和P利用效率的高低(Vitousek, 1982)。禾本科物种垂穗披碱草的C:N和C:P平均值要高于其他4个物种, 除豆科外杂类草的C:N和C:P平均值则低于其他物种。比较代表种的物种优势度发现, 禾本科物种垂穗披碱草的物种优势度随P添加显著提高, 而直立型杂类草甘青老鹳草的物种优势度显著下降; 相应地在功能群水平上禾本科的生物量显著增加, 而杂类草的生物量显著下降。表明物种间由于养分利用效率的差异改变了物种的竞争能力, 禾本科物种具有逐渐增高的C:N和C:P而在群落中占据优势, 相应杂类草由于逐渐降低的N和P利用效率而在群落中的竞争能力下降, 但是由于杂类草物种数目最大, 仍在决定群落结构方面起重要作用。
综合分析表明, 在青藏高原高寒草甸连续5年添加P显著改变了植物的C、N、P含量以及C:N:P计量比特征, 并且物种水平和功能群水平的响应规律基本一致。除豆科系列外, C含量在其他3个功能群和相应物种水平上P添加对C含量的影响不显著; P添加显著增加了物种水平和功能群水平植物中的P含量, 降低了C:P和N:P。物种水平和功能群水平的N含量和C:N对P添加的响应规律不一致。在N限制的生境中添加P, 禾本科物种具有逐渐增高的C:N和C:P而在群落中具有强的竞争能力, 相应禾本科功能群水平的生物量显著增加; 而杂类草由于逐渐降低的N和P利用效率而使其在群落中生物量所占的比重有所下降。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1371/journal.pone.0161492URLPMID:27560190 [本文引用: 1]

Atmospheric depositions pose significant threats to biodiversity and ecosystem function. However, the underlying physiological mechanisms are not well understood, and few studies have considered the combined effects and interactions of multiple pollutants. Thisin situstudy explored the physiological responses of two epiphytic bryophytes to combined addition of nitrogen, phosphorus and sulfur. We investigated the electrical conductivity (EC), total chlorophyll concentration (Chl), nutrient stoichiometry and chlorophyll fluorescence signals in a subtropical montane cloud forest in south-west China. The results showed that enhanced fertilizer additions imposed detrimental effects on bryophytes, and the combined enrichment of simulated fertilization exerted limited synergistic effects in their natural environments. On the whole, EC, Chl, the effective quantum yield of photosystem II (PSII) and photochemical quenching (qP) were the more reliable indicators of increased artificial fertilization. However, conclusions on nutrient stoichiometry should be drawn cautiously concerning the saturation uptake and nutrient interactions in bryophytes. Finally, we discuss the limitations of prevailing fertilization experiments and emphasize the importance of long-term data available for future investigations.
DOI:10.1111/j.1600-0706.2008.16793.xURL [本文引用: 1]
Abstract Liebig's law of the minimum, which states that only one element limits the growth of organisms at any given time, is widely used in ecology. This principle is routinely applied to organisms, populations and communities, but can it really be applied indistinguishably across these different scales? Here we show, by prediction of a resource ratio conceptual model and with an experimental test carried out in microcosms with bacteria that, unlike single species, communities are likely to adjust their stoichiometry to that of their resources. This adjustment results from competitive exclusion and coexistence mechanisms, and is sensitive to the overall diversity of species in the community. It guaranties co-limitation, i.e. simultaneous limitation by multiple resources, at the community scale and optimal use of resources and maximization of community biomass for wide ranges of resource ratios. These results question the applicability of the Liebig's law of the minimum at the community level, and the relevance of ecosystem models relying on this principle.
DOI:10.1111/1365-2435.12493URL [本文引用: 1]
Summary Plant chemical and structural defence compounds are well known to impact upon herbivory of fresh leaves and influence decomposition rates after leaf senescence. A number of theories predict that alleviating nutrient limitation and reducing other environmental stressors will result in decreased production of plant chemical defences. In this study, we measured plant defence properties [total polyphenols (TP), condensed tannins (CT) and lignin concentrations, and protein complexation capacity (PCC)] in both fresh and senesced plant leaves in a fully factorial N and P fertilization experiment set-up at each of three elevations along an elevational gradient in Swedish subarctic tundra heath vegetation. Further, we performed a decomposition of variance analysis on community-weighted averages (CWAs) of plant defence properties to determine the relative contributions of interspecific and intraspecific variation to the total variation observed in response to elevation and nutrient addition. We hypothesized that N fertilization would reduce plant defence properties and that this reduction would be greater at higher elevations, while the effects of P fertilization would have no effect at any elevation. At the community level, N addition reduced CT and PCC in both fresh and senesced leaves and TP in senesced leaves, while P addition had few effects, broadly in line with our hypothesis. The effects of N addition frequently varied with elevation, but in contrast to our hypothesis, the said effects were strongest at the lowest elevations. The effects of N addition and the interactive effect of N with elevation were primarily driven by intraspecific, rather than interspecific, variation. Our findings suggest that as temperatures warm and N availability increases due to global climate change, secondary metabolites in subarctic heath vegetation will decline particularly within species. Our results highlight the need to consider the effects of both nutrient availability and temperature, and their interaction, in driving subarctic plant defence.
DOI:10.1007/s00442-008-1123-xURL [本文引用: 1]
DOI:10.1007/BF00377192URLPMID:28311896 [本文引用: 1]
The photosynthetic capacity of leaves is related to the nitrogen content primarily bacause the proteins of the Calvin cycle and thylakoids represent the majority of leaf nitrogen. To a first approximation, thylakoid nitrogen is proportional to the chlorophyll content (50 mol thylakoid N mol -1 Chl). Within species there are strong linear relationships between nitrogen and both RuBP carboxylase and chlorophyll. With increasing nitrogen per unit leaf area, the proportion of total leaf nitrogen in the thylakoids remains the same while the proportion in soluble protein increases. In many species, growth under lower irradiance greatly increases the partitioning of nitrogen into chlorophyll and the thylakoids, while the electron transport capacity per unit of chlorophyll declines. If growth irradiance influences the relationship between photosynthetic capacity and nitrogen content, predicting nitrogen distribution between leaves in a canopy becomes more complicated. When both photosynthetic capacity and leaf nitrogen content are expressed on the basis of leaf area, considerable variation in the photosynthetic capacity for a given leaf nitrogen content is found between species. The variation reflects different strategies of nitrogen partitioning, the electron transport capacity per unit of chlorophyll and the specific activity of RuBP carboxylase. Survival in certain environments clearly does not require maximising photosynthetic capacity for a given leaf nitrogen content. Species that flourish in the shade partition relatively more nitrogen into the thylakoids, although this is associated with lower photosynthetic capacity per unit of nitrogen.
[本文引用: 2]
DOI:10.1111/j.1469-8137.2004.01192.xURL [本文引用: 3]
Abstract Contents Summary I Introduction II Variability of N : P ratios in response to nutrient supply III Critical N : P ratios as indicators of nutrient limitation IV Interspecific variation in N : P ratios V Vegetation properties in relation to N : P ratios VI Implications of N : P ratios for human impacts on ecosystems VII Conclusions Acknowledgements References Summary Nitrogen (N) and phosphorus (P) availability limit plant growth in most terrestrial ecosystems. This review examines how variation in the relative availability of N and P, as reflected by N : P ratios of plant biomass, influences vegetation composition and functioning. Plastic responses of plants to N and P supply cause up to 50-fold variation in biomass N : P ratios, associated with differences in root allocation, nutrient uptake, biomass turnover and reproductive output. Optimal N : P ratios those of plants whose growth is equally limited by N and P depend on species, growth rate, plant age and plant parts. At vegetation level, N : P ratios 20 often (not always) correspond to N- and P-limited biomass production, as shown by short-term fertilization experiments; however long-term effects of fertilization or effects on individual species can be different. N : P ratios are on average higher in graminoids than in forbs, and in stress-tolerant species compared with ruderals; they correlate negatively with the maximal relative growth rates of species and with their N-indicator values. At vegetation level, N : P ratios often correlate negatively with biomass production; high N : P ratios promote graminoids and stress tolerators relative to other species, whereas relationships with species richness are not consistent. N : P ratios are influenced by global change, increased atmospheric N deposition, and conservation managment.
DOI:10.1007/s11104-014-2154-1URL [本文引用: 1]
Background and aims Stoichiometric relations drive powerful constraints on many ecosystem processes. However, our understanding of the hierarchical responses of plant C:N:P stoichiometry at different...
DOI:10.1046/j.1365-2745.2003.00828.xURL [本文引用: 1]
1 Plant growth at high-latitude sites is usually strongly nutrient-limited. The increased nutrient availability predicted in response to global warming may affect internal plant nutrient cycling, including nutrient resorption from senescing leaves.
DOI:10.1007/s11104-015-2416-6URL [本文引用: 2]
Background and aims Plant nutrient concentrations and their correlations with soil nutrient conditions are regarded as effective tools for exploring plant adaptation and resource utilisation...
DOI:10.1890/09-0064.1URLPMID:20426329 [本文引用: 1]
Abstract Nitrogen (N) is the primary growth-limiting nutrient in many terrestrial ecosystems, and therefore plant production per unit N taken up (i.e., N use efficiency, NUE) is a fundamentally important component of ecosystem function. Nitrogen use efficiency comprises two components: N productivity (A(N), plant production per peak biomass N content) and the mean residence time of N in plant biomass (MRT(N)). We utilized a five-year fertilization experiment to examine the manner in which increases in N and phosphorus (P) availability affected plant NUE at multiple biological scales (i.e., from leaf to community level). We fertilized a natural gradient of nutrient-limited peatland ecosystems in the Upper Peninsula of Michigan, USA, with 6 g N x m(-2) x yr(-1), 2 g P x m(-2) x yr(-1), or a combination of N and P. Our objectives were to determine how changes in carbon and N allocation within a plant to leaf and woody tissue and changes in species composition within a community, both above- and belowground, would affect (1) NUE; (2) the adaptive trade-off between the components of NUE; (3) the efficiency with which plants acquired N from the soil (N uptake efficiency); and (4) plant community production per unit soil N availability (N response efficiency, NRE). As expected, N and P addition generally increased aboveground production and N uptake. In particular, P availability strongly affected the way in which plants took up and used N. Nitrogen use efficiency response to nutrient addition was not straightforward. Nitrogen use efficiency differed between leaf and woody tissue, among species, and across the ombrotrophic-minerotrophic gradient because plants and communities were adapted to maximize either A(N) or MRT(N), but not both concurrently. Increased N availability strongly decreased plant and community N uptake efficiency, while increased P availability increased N uptake efficiency, particularly in a nitrogen-fixing shrub. Nitrogen uptake efficiency was more important in controlling overall plant community response to soil N availability than was NUE, and above- and belowground community N uptake efficiencies responded to nutrient addition in a similar manner. Our results demonstrate that plants respond to nutrient availability at multiple biological scales, and we suggest that N uptake efficiency may be a more representative measurement of plant responses to nutrient availability gradients than plant NUE.
DOI:10.1111/1365-2745.12610URL [本文引用: 1]
Summary Nitrogen (N) deposition and phosphorus (P) deposition due to pollution and land-use change are dramatically altering biogeochemical cycles. These altered nutrient inputs affect plant communities by generally increasing dominance and reducing diversity, as well as altering community variability (heterogeneity). Less well studied are the effects of changes in community variability on ecosystem functions, such as productivity, or the stability of those functions. Here, we use a twelve-year nutrient addition experiment in tallgrass prairie to determine the variability in community responses to N and P additions and link these responses to ecosystem productivity and stability. We added two levels of N and four levels of P in a fully factorial design to 25-m2 plots in native tallgrass prairie in north-eastern Kansas, USA. Each year percentage cover of each species was measured in June and August in a 1-m2 subplot of each plot, and annual net primary productivity was measured in two 0.1-m2 subplots in each plot at the end of each growing season. The addition of N and P together increased plant community variability across space (i.e. the replicates were significantly more different from each other in the N + P treatments than they were in the control treatment). We also found that the variability of the plant community within a single plot through time increased with the addition of N alone and N and P together. The highest level of both spatial and temporal variability occurred in plots with the highest level of nutrient addition (10gm2 of both N and P). While we found no linkage between spatial variability of community composition and the spatial stability of productivity, the temporal stability of productivity decreased with increasing temporal plant community variability. Additionally, the ability to predict the productivity response to growing season precipitation, a key environmental variable, also decreased under higher temporal community variability. Synthesis . Using a 12-year nutrient addition experiment, we found that nutrient addition leads to both spatial and temporal community variability in mesic tallgrass prairie. The changes in community variability through time were directly related to ecosystem stability. While overall shifts in community structure in response to nutrient additions are important, the change in variability of local communities has significant implications for our ability to predict how patterns of biodiversity and ecosystem function will respond to a rapidly changing world.
DOI:10.1007/s10265-017-0930-8URLPMID:28364378 [本文引用: 1]
Nitrogen (N) input by atmospheric deposition and human activity enhances the availability of N in various ecosystems, which may further affect N and phosphorus (P) cycling and use by plants. However, the internal use of N, P, and N:P stoichiometry by plants in response to N supply, particularly for grass species in a desert steppe ecosystem, remains unclear. In this work, a field experiment was conducted at an infertile area in a desert steppe to investigate the effects of N fertilizer addition rates on the stoichiometry of N and P in a dominant grass species, Seriphidium korovinii . Results showed that for both aboveground and fine roots of S. korovinii , N inputs exponentially increased the N concentration and N:P ratios while P concentrations decreased. Meanwhile, the relationships between N and P concentrations for both aboveground and fine roots were significantly negative. Furthermore, while the N concentrations in the plants were relatively low, P concentrations were higher than the global means, resulting in a relatively low N:P ratio. These results suggest that the stoichiometric characteristics of N were different from that of P for this desert plant species. Results also show that the intraspecific variations in the main element traits (N, P, and N:P ratios) were consistent at the whole-plant level. Our results also suggest that N should be part of any short-term fertilization plan that is part of a management strategy designed to restore degraded desert grassland. These findings highlight that nutrient addition by atmospheric N deposition and human activity can have significant effects on the internal use of N and P by plants. Therefore, establishing a nutrient-conservation strategy for desert grasslands is important.
DOI:10.1111/j.1365-2494.2011.00781.xURL [本文引用: 1]
To examine whether the critical leaf N/P ratios (of 14, 16) are valid to test nutrient limitation in the context of semi-arid sandy grasslands, an experiment was conducted on a Keerqin sandy grassland in North-east China to investigate the responses of plant biomass and nutrient concentrations to fertilization. Plant biomass production and leaf nutrient concentrations were measured after five consecutive years of fertilization with N (20 g N m612 year611) and/or P (10 g P2O5 m612 year611). Nitrogen fertilization increased the shoot biomass by twofold and consequently the shoot/root ratio, whereas P fertilization had little effect on either shoot biomass or shoot/root ratio. Leaf N/P ratio varied among species with an average of 5·6 in the control, while the mean leaf N/P ratio (7·5) under the N fertilization treatment remained below the threshold of 14. Our results suggest that the critical N/P ratio (14, 16) is not applicable as a test for nutrient limitations in the context of semi-arid, sandy grassland.
DOI:10.1111/j.1654-1103.2006.tb02462.xURL [本文引用: 1]
Question: Which fraction of the decrease in species richness under fertilization can be explained by assemblage-level thinning? Location: An alpine meadow on the eastern Tibet plateau. Methods: 60-m2 plots were randomly assigned to a control or one of four levels of ammonium phosphate fertilizer. Treatments were repeated for three years. The effect of assemblage-level thinning was decided based on similarity in quadrats within and between fertilizing levels, bootstrap simulation based on random thinning of the high density (low production, low fertility) quadrats and correlation of species' biomass in low fertility and high fertility. Results: Fertilization increased production, reduced species richness and reduced density of individuals. Heavily fertilized quadrats are more similar in species composition in 2000 but less similar in 2001 and 2002. Rarefaction showed that a decrease in density can account for 32.3-42.9% decrease of species richness, but the simulated species richness is always significantly higher than the observed one. When production and species richness are similar at two levels of fertilization, species biomass in the higher fertility treatment is positively correlated with biomass at lower fertility. When the two fertilizer levels differed in production and species richness, there was no evidence of correlation in species biomass, suggesting that assemblage level thinning cannot explain all the loss of species. Conclusion: Although a decrease in density could explain much of the decrease (up to 42.9%) in species richness when this alpine meadow was fertilized, other important mechanisms such as interspecific competition cannot be ignored. Future studies should investigate the effect of assemblage level thinning on species diversity, and search for mechanisms responsible for a decrease in diversity..
DOI:10.1016/j.scitotenv.2016.03.158URLPMID:27045368 [本文引用: 2]
Phosphorus (P) enrichment induced by anthropogenic activities results in modified plant nutrient status, which potentially alters the stoichiometry of carbon (C), nitrogen (N), and P in plants. However, how increased P availability changes plant C:N:P stoichiometry at different hierarchical scales is unclear in N-limited ecosystems. In this study, we conducted a four-level P addition experiment (0, 1.2, 4.8, and 9.602g02P02m 61 2 02year 61021 ) to elucidate the effect of P enrichment on plant C:N:P stoichiometric ratios at both the species and community levels in a freshwater wetland in the Sanjiang Plain, Northeast China. We found that species- and community-level plant C:N:P stoichiometry responded consistently to six years of P addition, although there was a shift in species dominance. Phosphorus addition increased plant N and P concentrations and thus decreased C:N, C:P, and N:P ratios irrespective of the P addition levels. These similar change trends at different scales resulted from the identical responses of plant N and P concentrations in different species to P addition. Moreover, plant N concentration exhibited an increasing trend with increasing P addition levels, whereas plant C:N ratio showed a declining trend. At the community level, P addition at the rates of 1.2, 4.8, and 9.602g02P02m 61022 02year 61021 decreased the C:N ratio by 24%, 27%, and 34%; decreased the C:P ratio by 33%, 35%, and 38%; and decreased the N:P ratio by 12%, 10%, and 6%, respectively. Our results indicate that the stoichiometric responses to P addition are scale-independent, and suggest that altered plant C:N:P stoichiometry induced by P enrichment would stimulate organic matter decomposition and accelerate nutrient cycles in N -limited temperate freshwater wetlands.
DOI:10.1111/1365-2745.12190URL [本文引用: 3]
The concentration, stoichiometry and resorption of nitrogen (N) and phosphorus (P) in plant leaves are often used as proxies of the availability of these growth-limiting nutrients, but the responses of these metrics to changes in nutrient availability remain largely untested for tropical forest trees.We evaluated changes in N and P concentrations, N/P ratios and resorption for four common tree species after 13 years of factorial N and P additions in a lowland tropical forest in Panama.Chronic P addition increased foliar P concentrations, decreased P resorption proficiency and decreased N/P ratios in three locally common eudicot tree species (Alseis blackiana, Heisteria concinna, Tetragastris panamensis). The increase in foliar P involved similar proportional increases in organic and inorganic P in two species and a disproportionately large increase in inorganic P in A. blackiana.Nitrogen addition did not alter foliar N concentrations in any species, but did decrease N resorption proficiency in H. concinna.A fourth species, the palm Oenocarpus mapora, demonstrated remarkably static foliar nutrient concentrations, responding only with a marginal decrease in P resorption proficiency under N plus P co-addition.Synthesis. Collectively, these results suggest that adjustment of N/P ratios can be expected in eudicots exposed to elevated P, but foliar N appears to already be at optimal levels in these lowland rain forest tree species. The complexity of species-specific responses to altered nutrient availability highlights the difficulty in predicting future responses of tropical forest trees to a changing world.
DOI:10.1890/04-0354URL [本文引用: 1]
Current understanding of the extent, causes for, and consequences of variation in nutrient composition in plants is limited. Important questions to be addressed include to what extent nutrients covary, how flexible nutrient ratios are within a population or species, how reproduction influences nutrient ratios, and how much the ratios of nutrients to mass and nutrients to each other change through ontogeny. This information is needed to assess the physiological and ecological consequences of plant nutrient composition and to what extent plants function as balanced systems in acquisition and allocation of resources. We studied the variation in nutrient stoichiometry (i.e., the ratio between contents of different nutrients within a plant) in relation to three factors: (1) environmental availability of nitrogen, (2) plant size, and (3) reproductive status. We investigated these questions in 11 populations of the carnivorous plant Pinguicula vulgaris in northern Scandinavia. Dry mass and N and P content were measured for reproductive and vegetative portions of flowering individuals and for winter buds corresponding to four reproductive states: control reproductive individuals, experimentally vegetative individuals (from which flower buds were removed), adult vegetative individuals, and individuals below the threshold size for reproduction. [N], [P], and to a lesser extent, N and P content were positively related to soil N, but not to prey capture. Nutrient stoichiometry was also size dependent; in general, small plants were relatively enriched in N and relatively depleted in P compared to larger plants. Reproductive status affected not only size, but also nutrient stoichiometry of the resulting winter bud. Winter buds derived from reproductive individuals had a higher [N] and lower [P] than those of the different types of nonproductive individuals. Our findings indicate that studies of nutrient stoichiometry in plants must go beyond links between environmental and plant nutrient concentrations to consider internal processes such as growth and reproduction.
DOI:10.3969/j.issn.1001-0068.2006.04.010URL
在温室条件下,利用水培技术,研究了磷对大豆生长、根生物量和根瘤固氮的影响.结果表明,大豆在生长初期表现出缺氮症状,随着磷浓度的增加,根瘤数、根瘤干重、氮和磷累积量均显著增加.在移栽后0d~14d,磷对大豆根瘤的生长影响不大,而在21d后,磷对根瘤生长的影响逐渐增大,移栽后35d,高磷处理比低磷增长了138%.植株不同部位磷含量大小顺序是:根瘤>根>地上部分.图5,表1,参6.
DOI:10.3969/j.issn.1001-0068.2006.04.010URL
在温室条件下,利用水培技术,研究了磷对大豆生长、根生物量和根瘤固氮的影响.结果表明,大豆在生长初期表现出缺氮症状,随着磷浓度的增加,根瘤数、根瘤干重、氮和磷累积量均显著增加.在移栽后0d~14d,磷对大豆根瘤的生长影响不大,而在21d后,磷对根瘤生长的影响逐渐增大,移栽后35d,高磷处理比低磷增长了138%.植株不同部位磷含量大小顺序是:根瘤>根>地上部分.图5,表1,参6.
DOI:10.1007/s11104-010-0281-xURL [本文引用: 2]
How plants respond to long-term nutrient enrichment can provide insights into physiological and evolutionary constraints in various ecosystems. The present study examined foliar concentrations after fertilization o determine if nutrient accumulation responses of the most abundant species in a plant community reflect differences in N and P uptake and storage. Using a chronosequence in the Hawaiian Islands that differs in N and P availability, it was shown that after fertilization, plants increase foliar P to a much greater degree than foliar N, as indicated by response ratios. In addition, foliar P responses after fertilization were more variable and largely driving the observed changes in N:P values. Across species, both inorganic and organic P increased but neither form of N increased significantly. This pattern of P accumulation was consistent across 13 species of varying life forms and occurred at both the N-limited and P-limited site, although its magnitude was larger at the P-limited site. Foliar P accumulation after nutrient enrichment may indicate nutrient storage and may have evolved to be a general strategy to deal with uncertainties in P availability. Storage of P complicates interpretations of N:P values and the determination of nutrient limitation.
DOI:10.1038/ncomms3934URLPMID:24343268 [本文引用: 1]
Abstract The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen:phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
DOI:10.1073/pnas.0403588101URLPMID:15213326 [本文引用: 1]
A global data set including 5,087 observations of leaf nitrogen (N) and phosphorus (P) for 1,280 plant species at 452 sites and of associated mean climate indices demonstrates broad biogeographic patterns. In general, leaf N and P decline and the N/P ratio increases toward the equator as average temperature and growing season length increase. These patterns are similar for five dominant plant groups, coniferous trees and four angiosperm groups (grasses, herbs, shrubs, and trees). These results support the hypotheses that (i) leaf N and P increase from the tropics to the cooler and drier midlatitudes because of temperature-related plant physiological stoichiometry and biogeographical gradients in soil substrate age and then plateau or decrease at high latitudes because of cold temperature effects on biogeochemistry and (ii) the N/P ratio increases with mean temperature and toward the equator, because P is a major limiting nutrient in older tropical soils and N is the major limiting nutrient in younger temperate and high-latitude soils.
[本文引用: 2]
DOI:10.1073/pnas.0408648102URLPMID:15755810 [本文引用: 1]
Human activities have increased N availability dramatically in terrestrial and aquatic ecosystems. Extensive research demonstrates that local plant species diversity generally declines in response to nutrient enrichment, yet the mechanisms for this decline remain unclear. Based on an analysis of >900 species responses from 34 N-fertilization experiments across nine terrestrial ecosystems in North America, we show that both trait-neutral and trait-based mechanisms operate simultaneously to influence diversity loss as production increases. Rare species were often lost because of soil fertilization, randomly with respect to traits. The risk of species loss due to fertilization ranged from >60% for the rarest species to 10% for the most abundant species. Perennials, species with N-fixing symbionts, and those of native origin also experienced increased risk of local extinction after fertilization, regardless of their initial abundance. Whereas abundance was consistently important across all systems, functional mechanisms were often system-dependent. As N availability continues to increase globally, management that focuses on locally susceptible functional groups and generally susceptible rare species will be essential to maintain biodiversity.
DOI:10.1093/jpe/rtv037URL [本文引用: 5]
DOI:10.1073/pnas.1301606110URL [本文引用: 1]
The microbial production of ethanol from lignocellulosic biomass is a multi-component process that involves biomass hydrolysis, carbohydrate transport and utilization, and finally, the production of ethanol. Strains of the genus Thermoanaerobacter have been studied for decades due to their innate abilities to produce comparatively high ethanol yields from hemicellulose constituent sugars. However, their inability to hydrolyze cellulose, limits their usefulness in lignocellulosic biofuel production. As such, co-culturing Thermoanaerobacter spp. with cellulolytic organisms is a plausible approach to improving lignocellulose conversion efficiencies and yields of biofuels. To evaluate native lignocellulosic ethanol production capacities relative to competing fermentative end-products, comparative genomic analysis of 11 sequenced Thermoanaerobacter strains, including a de novo genome, Thermoanaerobacter thermohydrosulfuricus WC1, was conducted. Analysis was specifically focused on the genomic potential for each strain to address all aspects of ethanol production mentioned through a consolidated bioprocessing approach. Whole genome functional annotation analysis identified three distinct clades within the genus. The genomes of Clade 1 strains encode the fewest extracellular carbohydrate active enzymes and also show the least diversity in terms of lignocellulose relevant carbohydrate utilization pathways. However, these same strains reportedly are capable of directing a higher proportion of their total carbon flux towards ethanol, rather than non-biofuel end-products, than other Thermoanaerobacter strains. Strains in Clade 2 show the greatest diversity in terms of lignocellulose hydrolysis and utilization, but proportionately produce more non-ethanol end-products than Clade 1 strains. Strains in Clade 3, in which T. thermohydrosulfuricus WC1 is included, show mid-range potential for lignocellulose hydrolysis and utilization, but also exhibit extensive divergence from both Clade 1 and Clade 2 strains in terms of cellular energetics. The potential implications regarding strain selection and suitability for industrial ethanol production through a consolidated bioprocessing co-culturing approach are examined throughout the manuscript.
DOI:10.1159/000090648URL [本文引用: 1]
1. We studied the effects of increased water temperatures (0-4.5 pC) and nutrient enrichment on the stoichiometric composition of different primary producers (macrophytes, epiphytes, seston and sediment biofilm) and invertebrate consumers in 24 mesocosm ecosystems created to mimic shallow pond environments. The nutrient ratios of primary producers were used as indicative of relative nitrogen (N) or phosphorus (P) limitation. We further used carbon stable isotopic composition (tp#pdC) of the different primary producers to elucidate differences in the degree of CO limitation. 2. Epiphytes were the only primary producer with significantly higher tp#pdC in the enriched mesocosms. No temperature effects were observed in tp#pdC composition of any primary producer. Independently of the treatment effects, the four primary producers had different tp#pdC signatures indicative of differences in CO limitation. Seston had signatures indicating negligible or low CO limitation, followed by epiphytes and sediment biofilm, with moderate CO limitation, while macrophytes showed the strongest CO limitation. CO together with biomass of epiphytes were the key variables explaining between 50 and 70% of the variability in tp#pdC of the different primary producers, suggesting that epiphytes play an important role in carbon flow of temperate shallow lakes. 3. The ratio of carbon to chlorophyll a decreased with increasing temperature and enrichment in both epiphytes and seston. The effects of temperature were mainly attributed to changes in algal Chl a content, while the decrease with enrichment was probably a result of a higher proportion of algae in the seston and epiphytes. 4. Macrophytes, epiphytes and seston decreased their C : N with enrichment, probably as an adaptation to the different N availability levels. The C : N of epiphytes and Elodea canadensis decreased with increasing temperature in the control mesocosms. Sediment biofilm was the only primary producer with lower C : P and N
DOI:10.1086/283931URL [本文引用: 1]
DOI:10.1111/j.1438-8677.2012.00605.xURLPMID:22672784 [本文引用: 1]
Variations in foliar stable carbon isotope signatures (020713C) of different plant functional groups (PFGs) and their relationships with environmental factors in China were investigated in this meta-analysis. There were some significant, but small differences in 020713C among PFGs categorised by life form (<10900°). Trees (09080826.780900°) and shrubs (09080826.890900°) had similar mean 020713C that were significantly higher than those of herbs (09080827.490900°). Evergreen shrubs (09080825.820900°) had significantly higher mean 020713C than deciduous shrubs (09080826.920900°). Perennial herbs (09080826.830900°) had significantly higher mean 020713C than annual herbs (09080827.100900°). Grasses (09080826.460900°) had significantly higher mean 020713C than forbs (09080826.960900°). For pooled data, 020713C was significantly and negatively correlated with mean annual precipitation (MAP) and mean annual temperature (MAT), while it was significantly and positively correlated with latitude and altitude. There was a threshold value of MAP along the gradients, and 020713C did not change significantly with higher rainfall. The 020713C of PFGs changed with altitude, suggesting that increases in 020713C with altitude cannot be generalised. Differences in 020713C between PFGs were generally much <10900° and therefore insignificant. In contrast, MAP and MAT had relatively large effects on 020713C (more than 40900° between extremes). The 020713C of some PFGs responded to environmental gradients in the same manner, while their 090004rates090005 of change were significantly different in some cases. This information could help predict potential changes in the distribution of PFGs in response to future climate change.
DOI:10.1007/s11104-014-2316-1URL [本文引用: 2]
Background Nitrogen (N) and phosphorus (P) availabilities affect plant growth and the balance of elements, yet how these plant traits respond to N and P supply remains unclear.
DOI:10.1890/14-0140.1URLPMID:26240859 [本文引用: 3]
Abstract Plants in infertile habitats are thought to have a high rate of nutrient resorption to enable them reuse nutrients more efficiently than those in fertile habitats. However, there is still much debate on how plant nutrient resorption responds to nutrient availability. Here we used a meta-analysis from a global data set of 9703 observations at 306 sites from 508 published articles to examine the effects of nitrogen (N) and phosphorus (P) fertilization on plant foliar N and P concentrations and resorption efficiency. We found that N fertilization enhanced N concentration in green leaves by 27% and P fertilization enhanced green-leaf P by 73% on average. The N and P concentrations in senesced leaves also increased with respective nutrient fertilization. Resorption efficiencies (percentage of nutrient recovered from senescing leaves) of both N and P declined in response to respective nutrient fertilization. Combined N and P fertilization also had negative effects on both N and P resorption efficiencies. Whether nutrient resorption efficiency differs among plant growth types and among ecosystems, however, remains uncertain due to the limited sample sizes when analyzed by plant growth types or ecosystem types. Our analysis indicates that fertilization decreases plant nutrient resorption and the view that nutrient resorption is a critical nutrient conservation strategy for plants in nutrient-poor environments cannot be abandoned. The response values to fertilization presented in our analysis can help improve biogeochemical models.
DOI:10.3402/polar.v30i0.15942URL [本文引用: 1]
It is known that the dominance of graminoid species is promoted by N addition; however, there has been relatively little effort to examine the pattern induced by natural N fertility.Since nutrie nt use e fficiency (NUE) is an important trait determining plant competitive ability, we expected that the species guild with higher NUE (lower nutrient content) may be more competitive on infertile soils. We explo red t he relationships between relative forbs biomass share, soil N and productivity by the linear regressi on analysis on a natural alpine meadow in northeast of Qinghai-Tibetan Plateau (3600 m a.s.l.). To test the variety of leaf N:P stoichiometry, paired t test and general linear model multivariate (GLM) analysis were also used. We f ound that the leaf N:P ratios of the whole community were below 13 in studied sites, which may be consistent with the N limitation on the veget ation. Graminoids re tained lower concentrations of leaf N and P than forbs in community on the Nlimited grassland. Consistent with our prediction, we found that the biomass fraction of graminoids declined with soil N content and aboveground production on the grassland. Different from the pattern along fertility gradients induced by N fertilization, our results showed that gr aminoids with lower internal nutrient content w ere able to resist low levels of nutrient availability on the natural alpine grassland when compared to forbs.
氮磷复合肥添加对高寒草甸金露梅灌丛中豆科植物的影响
2010
氮磷复合肥添加对高寒草甸金露梅灌丛中豆科植物的影响
2010
Physiological Responses of two epiphytic bryophytes to nitrogen, phosphorus and sulfur addition in a subtropical montane cloud forest
1
2016
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
Does Liebig’s law of the minimum scale up from species to communities?
1
2008
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Effects of elevation and nitrogen and phosphorus fertilization on plant defence compounds in subarctic tundra heath vegetation
1
2016
... id="C22">P添加显著增加了物种水平和功能群水平的P含量, 尽管N含量有增加有降低, 但是参试物种和功能群水平的N:P均显著下降(
Soil fertility increases with plant species diversity in a long-term biodiversity experiment
1
2008
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Photosynthesis and nitrogen relationships in leaves of C3 plants
1
1989
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Nutrient addition differentially affects ecological processes of Avicennia germinans in nitrogen versus phosphorus limited mangrove ecosystems
2
2007
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
N:P ratios in terrestrial plants: Variation and functional significance
3
2004
... id="C4">生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
... id="C22">P添加显著增加了物种水平和功能群水平的P含量, 尽管N含量有增加有降低, 但是参试物种和功能群水平的N:P均显著下降(
Hierarchical responses of plant stoichiometry to nitrogen deposition and mowing in a temperate steppe
1
2014
... id="C4">生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(
Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization
1
2003
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
Effects of soil fertility on the N:P stoichiometry of herbaceous plants on a nutrient-?limited alpine steppe on the northern Tibetan Plateau
2
2015
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
Scaling plant nitrogen use and uptake efficiencies in response to nutrient addition in peatlands
1
2010
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Nutrient additions cause divergence of tallgrass prairie plant communities resulting in loss of ecosystem stability
1
2016
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
Stoichiometry in aboveground and fine roots of Seriphidium korovinii in desert grassland in response to artificial nitrogen addition
1
2017
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
Foliar N/P ratio and nutrient limitation to vegetation growth on Keerqin sandy grassland of Northeast China
1
2011
... id="C22">P添加显著增加了物种水平和功能群水平的P含量, 尽管N含量有增加有降低, 但是参试物种和功能群水平的N:P均显著下降(
Importance of assemblage-level thinning: A field experiment in an alpine meadow on the Tibet Plateau
1
2006
... id="C8">本试验采用完全随机设计, 于2009年在青藏高原高寒草甸群落设定不同浓度的P添加实验, 设定4个处理(C、P5、P10、P15), P添加量分别为0、5、10、15 g·m-2·a-1, 所施磷肥为磷酸二氢钠(NaH2PO4).每个处理设3个重复, 共12个小区, 每小区面积为5 m × 5 m.各小区间有1 m的缓冲带, 缓冲带不施肥.每个小区内有4个亚样方(0.5 m × 0.5 m), 其中两个用于长期物种、多度与盖度的调查, 一个用于地上生物量的测定, 另外一个用于植株高度测定.为使肥料效应能充分体现出对群落结构的影响, 施肥于试验的前一年8月底至9月初进行.通常选择雨天添加营养元素, 使肥料颗粒溶解同时满足施肥后植物对水分的需求(
Effects of P addition on plant C:N:P stoichiometry in an N-limited temperate wetland of Northeast China
2
2016
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest
3
2014
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Nutrient stoichiometry in Pinguicula vulgaris: Nutrient availability, plant size, and reproductive status
1
2005
... id="C4">生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(
大豆结瘤固氮对磷素的需求
2006
大豆结瘤固氮对磷素的需求
2006
Foliar nitrogen and phosphorus accumulation responses after fertilization: An example from nutrient-??limited Hawaiian forests
2
2010
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
... id="C22">P添加显著增加了物种水平和功能群水平的P含量, 尽管N含量有增加有降低, 但是参试物种和功能群水平的N:P均显著下降(
Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe
1
2013
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
Global patterns of plant leaf N and P in relation to temperature and latitude
1
2004
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
2
2002
... id="C4">生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(
... id="C19">我们发现在青藏高原高寒草甸连续5年添加P显著改变了植物的营养物质含量, 故而植物的生态化学计量比也相应发生了变化, 并且物种水平和功能群水平的响应规律基本一致.在P添加处理下, 除豆科外, C含量在其他3个功能群和相应物种水平对P添加的响应规律一致: P添加对C含量的影响不显著(
Functional- and abundance-based mechanisms explain diversity loss due to N fertilization
1
2005
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
Species richness loss after nutrient addition as affected by N:C ratios and phytohormone GA3 contents in an alpine meadow community
5
2016
... id="C4">生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(
... id="C6">由于物种间对P富集的差异性响应会影响物种的生长速率而改变物种的竞争能力, 使群落的物种组成和构成发生变化.为了具体探究P富集导致哪些物种丧失, 基于功能群的物种多样性研究是综合评价物种和群落对环境波动响应的有效工具(
... ).所以P富集不仅改变了物种水平的生态化学计量比过程, 而且调节了功能群水平的计量比特征. 而基于功能群的计量比特征更能准确反映物种间对于P富集响应的差异性, 从而对群落的构成和结构的改变做出解释.因此, 我们以已经探明受N限制的高寒草甸群落为研究对象(
... id="C9">分别在施肥样地中选取群落共有物种作为研究对象, 包括: 垂穗披碱草(禾本科)、矮嵩草(莎草科)、黄花棘豆(豆科)、川甘蒲公英(莲座状杂类草)与甘青老鹳草(直立型杂类草).地上净生物量产出测定于2013年8月中旬齐地面剪取每个处理地上部分装袋, 3次重复.单独封装5个典型种, 其他物种按照禾本科、莎草科、豆科、杂类草4类功能群单独装袋, 带回实验室60 ℃烘干48 h至恒质量并称量记录(精确到0.001 g), 然后用于C、N、P含量的测定.4类功能群C、N、P含量的测定中不包括5个典型种的植物样.植物样地上部分经研磨后用80 μm的筛子过筛.采用重铬酸钾容量法测定植物全C (外加热法); 采用HClO4-H2SO4消煮后的溶液经定容、沉淀和稀释后用SmartChem 200化学分析仪(WestCo Scientific Instrument, Brookfielg, USA)分析测定样品全N和全P (
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
Multiple dimensions of resource limitation in tropical forests
1
2013
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Effects of increased temperature and nutrient enrichment on the stoichiometry of primary producers and consumers in temperate shallow lakes
1
2008
... id="C4">生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳(C)、氮(N)、磷(P))平衡的科学(
Nutrient cycling and nutrient use efficiency
1
1982
... id="C23">叶片C:N和C:P能够反映出植物对N和P利用效率的高低(
Variations in foliar stable carbon isotopes among functional groups and along environmental gradients in China—A meta-analysis
1
2013
... id="C6">由于物种间对P富集的差异性响应会影响物种的生长速率而改变物种的竞争能力, 使群落的物种组成和构成发生变化.为了具体探究P富集导致哪些物种丧失, 基于功能群的物种多样性研究是综合评价物种和群落对环境波动响应的有效工具(
Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana
2
2015
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(
Negative effects of fertilization on plant nutrient resorption
3
2015
... id="C5">N和P是植物生长所需的基本元素, 近几十年来, 由于人类大力发展农业和工业, N、P排放量增加不仅改变了区域或全球的N、P循环模式, 而且对自然群落的结构和构成、生态系统的生产力和功能稳定性等构成威胁(
... ).而在N限制的生态系统, 由于物种间不同的养分利用策略, N含量、C:N和N:P对P添加的响应无一致性规律(
... id="C20">P添加显著增加了物种水平和功能群水平植物组织中的P含量(
Biomass fraction of graminoids and forbs in N-limited alpine grassland: N:P stoichiometry
1
2011
... id="C21">随着P添加量增加, 物种间地上部分的N含量和C:N响应规律不一致, 且豆科和杂类草系列在物种水平和功能群水平的响应规律也不一致(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}