Distribution and storage of soil organic carbon across the desert grasslands in the southeastern fringe of the Tengger Desert, China
YANG Hao-Tian, WANG Zeng-Ru,*, JIA Rong-Liang*Shapotou Desert Research and Experiment Station, Northwest Institute of Eco-environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
SupportedbytheStrategicPriorityResearchProgramoftheChineseAcademyofSciences(XDA05050406-1) the National Natural Science Foundation of China(41501110) the West Light Program for Talent Cultivation of Chinese Academy of Sciences.(41401112)
Abstract Aims The complexity of environments and high spatial heterogeneity of desert ecosystems are important factors contributing to the uncertainty in the estimation of soil organic carbon storage.Methods Ten types of desert grassland communities in the southeastern fringe of the Tengger Desert, China were investigated. The content and vertical distribution of soil organic carbon (SOC) content in seven soil depths (0-5, 5-10, 10-20, 20-30, 30-50, 50-70 and 70-100 cm) and the underlying drivers were examined. Soil organic carbon density (SOCD) of four soil profiles (0-5, 0-20, 0-50 and 0-100 cm) were quantified.Important findings We found significant differences in SOC content among the 10 vegetation communities, and the shrub community type was an important factor affecting SOC content. Two types of trends in SOC content changes with soil depth were observed: 1) monotonic decrease, 2) increase followed by decrease. The SOC content was significantly positively correlated with clay content, total N, total P and conductivity, but negatively correlated with sand content. There were significant differences in SOCD for soil profiles of 0-5, 0-20, 0-50 and 0-100 cm among different communities, of which the mean values of SOCD were 0.118, 0.478, 1.159 and 1.936 kg·m-2, respectively. Our results show that SOCD is far below the mean value of global or national grasslands. Using the average values of SOCD across either global or national grasslands (including the grassland in this study) to estimate the SOC storage of desert ecosystems may lead to the overestimation or underestimation. Using the SOCD of specific communities may greatly increase the accuracy of SOC storage estimation in desert grasslands. Keywords:the Tengger Desert;desert grassland;soil organic carbon content;soil organic carbon density
PDF (1003KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 杨昊天, 王增如, 贾荣亮. 腾格里沙漠东南缘荒漠草地不同群落类型土壤有机碳分布及储量特征. 植物生态学报, 2018, 42(3): 288-296 doi:10.17521/cjpe.2017.0068 YANG Hao-Tian, WANG Zeng-Ru, JIA Rong-Liang. Distribution and storage of soil organic carbon across the desert grasslands in the southeastern fringe of the Tengger Desert, China. Chinese Journal of Plant Ecology, 2018, 42(3): 288-296 doi:10.17521/cjpe.2017.0068
土壤有机碳(SOC)库是陆地生态系统中最大的有机碳库, 其微小的变化都会影响大气中CO2的浓度, 因而在全球碳循环过程和全球气候变化中起着重要的作用(Post et al., 1990; Davidson et al., 2000)。SOC储量是土壤碳输入和输出之间的净差值, 输入主要来自叶和根的凋落物, 输出主要为根呼吸及有机物分解释放的CO2 (Kirschbaum, 2000)。不同的植被类型伴随着不同的生境条件和不同的生理生态特征, 形成了不同的土壤有机质的输入和输出方式, 进而影响SOC密度分布和土壤碳储量的大小(丁越岿等, 2012)。目前植被类型与SOC密度分布特征和SOC储量关系的相关研究主要集中在森林、草地和湿地生态系统, 而荒漠植被类型因其具有较小的碳密度而受到较少的关注。
荒漠生态系统是陆地生态系统的重要组成部分, 面积占全球陆地面积的30%以上, 碳储量约为全球碳储量的8% (Janzen, 2004; Li et al., 2015), 主要以土壤碳的形式存在(Wohlfahrt et al., 2008; 赵有益等, 2008)。由于荒漠生态系统较为脆弱, 土壤碳储量容易受环境变化的影响, 在气候变化和人类活动加剧的背景下, 荒漠生态系统土壤碳在全球碳循环中可能起着更加重要的作用。有研究认为荒漠生态系统可能是全球碳循环长期寻找的“碳失汇”所在地, 近年来的研究表明荒漠生态系统具有很强的固碳潜力, 是未来固定CO2的重要区域(Stone, 2008; Rotenberg & Yakir, 2010; Evans et al., 2014), 但是, 目前还缺乏强有力的证据(Luyssaert et al., 2007; Wohlfahrt et al., 2008; Schlesinger et al., 2009)。环境的复杂性及高度的空间异质性给荒漠生态系统碳储量的精确评估带来较大的困难(Jasoni et al., 2005; Xie et al., 2009), 与其他生态系统相比, 荒漠生态系统土壤碳储量研究还很薄弱(Hastings et al., 2005; Gao et al., 2012)。近年来, 我国典型荒漠生态系统碳储量的研究逐渐增多, 如新疆温性荒漠碳储量(安尼瓦尔·买买提, 2006; 门学慧, 2013)、河西走廊荒漠草地碳储量(王敏, 2014)、内蒙古荒漠草原碳储量(马文红, 2006; 杨婷婷, 2013), 虽然这些研究极大地丰富和完善了我国荒漠草地碳储量研究, 但涉及荒漠植物群落与土壤碳储量关系的研究还较少, 还远不能代表我国丰富的荒漠植物群落的土壤碳储量特征。现有的研究结果显示, 荒漠生态系统土壤碳储量估算结果不确定性极为明显, 这已成为国家和区域尺度上碳储量评估以及国家政策制定的主要障碍。腾格里沙漠东南缘植被为典型的荒漠草原类型, 是草原化荒漠与荒漠化草原的过渡带, 地带性植被发育在波状起伏的沙质草原上, 母质是洪积冲积物, 该区灌木、半灌木群落类型丰富。由稀疏植被或荒漠植被构成干旱地区典型的生态系统(斑块植被结构或镶嵌植被结构), 这种植被类型基本是阿拉善高原沙质、沙砾质荒漠区的典型代表。其特殊的地理位置和气候条件, 使得该区植物类型及其生物适应性特征更具独特性(李新荣等, 2016)。研究不同类型荒漠植物群落SOC密度特征, 有助于我们在区域尺度上认识SOC的分布特征并在此基础上精确地估算荒漠生态系统SOC储量, 减少荒漠生态系统土壤碳储量估算的不确定性, 也对应对气候变化、制定提高土壤碳储量的管理措施具有重要意义。本研究选择腾格里沙漠东南缘现有的主要类型群落研究SOC含量、分布特点及其差异, 旨在全面地阐明在腾格里沙漠南缘这一特殊生境植被类型对SOC的影响。
1.2.2.1 灌木和草本调查 灌木调查: 灌木样方大小为10 m × 10 m, 调查样方内所有灌木的总盖度, 分物种调查所有灌木的株高及冠幅。草本调查: 草本样方大小为1 m × 1 m, 调查样方内所有草本植物的总盖度、群落高度。并分物种调查高度、株(丛)数量。用收获法获得样方内所有草本地上部分活体生物量、凋落物生物量和半分解层生物量。
SOCD =$\sum\limits_{i=1}^{j}{{{C}_{i}}}$× Ti × ρi × (1 - τi)/10 (2)
数据统计分析在Excel和SPSS 19.0中进行。
2 结果和分析
2.1 不同植被类型的土壤有机碳含量特征
在自然条件下, 植被是SOC的重要来源, 植被类型会影响SOC的输入和输出方式, 进而影响SOC含量。本研究显示, 在腾格里沙漠南缘的荒漠化草地, 灌木群落类型是SOC含量的重要影响因素。对于7个土层(0-5、5-10、10-20、20-30、30-50、50-70和70-100 cm土层)的SOC含量, 10个以灌木为优势种的典型荒漠植物群落之间均存在显著差异(表1), 0-5 cm土层SOC含量, F = 26.549, p ? 0.05; 5-10 cm土层SOC含量, F = 8.651, p ? 0.05; 10-20 cm土层SOC含量, F = 21.810, p ? 0.05; 20-30 cm土层SOC含量, F = 15.652, p ? 0.05; 30- 50 cm土层SOC含量, F = 7.488, p ? 0.05; 50-70 cm土层SOC含量, F = 23.374, p ? 0.05; 70-100 cm土层SOC含量, F = 6.992, p ? 0.05 (表1)。
Table 1 表1 表1不同类型群落土壤有机碳含量(g·kg-1) (平均值±标准偏差) Table 1Soil organic carbon content (g·kg-1) of different communities (mean ± SD)
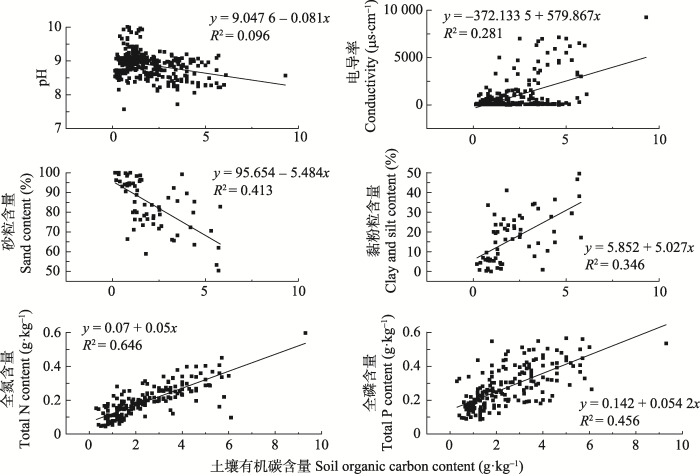
Fig. 1The relationships between soil organic carbon content and pH value, conductivity, sand content, clay and silt content, total nitrogen (N) and total phosphorus (P) content.
Fig. 2Soil organic carbon density in different depth for different community types (mean ± SD). The abbreviations of community types are the same as in Table 1. Different superscript letters indicate significant difference of soil organic carbon density between different communities (p < 0.05).
Table 2 表2 表2不同土层土壤有机碳密度占0-100 cm土层有机碳密度的比率 Table 2The ratio of soil organic carbon density of 0-5, 0-20, and 0-50 cm soil profile to soil organic carbon density of 0-100 cm soil profile
群落类型 Community type
土层 Soil layer
0-5
0-20
0-50
AM
0.071
0.256
0.562
CLA
0.104
0.306
0.622
KF
0.036
0.197
0.535
NT
0.063
0.248
0.541
OAL
0.062
0.174
0.500
PM
0.046
0.284
0.674
RS
0.018
0.151
0.520
RSK
0.113
0.348
0.715
SP
0.089
0.299
0.682
ZX
0.033
0.193
0.543
平均值 Mean
0.061
0.247
0.599
The abbreviations of community types are the same as in Table 1. 群落类型代码见表1。
BatjesNH ( 1996). Total carbon and nitrogen in the soils of the world European Journal of Soil Science, 47, 151-163. DOI:10.1111/ejs.1996.47.issue-2URL
DavidsonEA, TrumboreSE, AmundsonR ( 2000). Biogeochemistry: Soil warming and organic carbon content Nature, 408, 789-790. DOI:10.1038/35048672URLPMID:11130707 [本文引用: 1] Soils store two or three times more carbon than exists in the atmosphere as CO, and it is thought that the temperature sensitivity of decomposing organic matter in soil partly determines how much carbon will be transferred to the atmosphere as a result of global warming. Giardina and Ryan have questioned whether turnover times of soil carbon depend on temperature, however, on the basis of experiments involving isotope analysis and laboratory incubation of soils. We believe that their conclusions are undermined by methodological factors and also by their turnover times being estimated on the assumption that soil carbon exists as a single homogeneous pool, which can mask the dynamics of a smaller, temperature-dependent soil-carbon fraction. The real issue about release of carbon from soils to the atmosphere, however, is how temperature, soil water content and other factors interact to influence decomposition of soil organic matter. And, contrary to one interpretation of Giardina and Ryan's results, we believe that positive feedback to global warming is still a concern.
DingYK, YangJ, SongBY, HuGJLT, ZhangL ( 2012). Effect of different vegetation types on soil organic carbon in Mu Us Desert Acta Prataculturae Sinica, 21(2), 18-25. DOI:10.11686/cyxb20120203URLMagsci [本文引用: 1] 通过对比分析毛乌素沙地9种主要植被类型样地中的土壤有机碳含量和密度及其在土壤剖面上的分布差异,研究了不同植被类型对毛乌素沙地土壤有机碳的影响。结果表明,0~80cm 深度的土壤有机碳密度与平均土壤有机碳含量的大小关系均为:马蔺群落>芨芨草群落>旱柳群落>固定沙地油蒿群落>沙柳群落>半固定沙地油蒿群落>农田>中间锦鸡儿群落>流动沙地。随着土层加深,固定沙地油蒿群落、中间锦鸡儿群落、旱柳群落、芨芨草群落、马蔺群落和农田的有机碳含量均显著增加,而半固定沙地油蒿群落、流动沙地、沙柳群落的土壤有机碳含量随土层加深逐渐减少。各植被类型土壤有机碳密度在土壤剖面上的垂直分布格局和有机碳含量相似,但二者变化幅度不同。在流动沙地上进行植被建设有助于土壤固碳,且油蒿群落和沙柳群落的固碳效果较好。应尽量减少和避免强烈的人为干扰,以防止浅层土壤碳发生变化,维持土壤碳库稳定。 丁越岿, 杨劼, 宋炳煜, 呼格吉勒图, 张琳 ( 2012). 不同植被类型对毛乌素沙地土壤有机碳的影响 草业学报, 21(2), 18-25. DOI:10.11686/cyxb20120203URLMagsci [本文引用: 1] 通过对比分析毛乌素沙地9种主要植被类型样地中的土壤有机碳含量和密度及其在土壤剖面上的分布差异,研究了不同植被类型对毛乌素沙地土壤有机碳的影响。结果表明,0~80cm 深度的土壤有机碳密度与平均土壤有机碳含量的大小关系均为:马蔺群落>芨芨草群落>旱柳群落>固定沙地油蒿群落>沙柳群落>半固定沙地油蒿群落>农田>中间锦鸡儿群落>流动沙地。随着土层加深,固定沙地油蒿群落、中间锦鸡儿群落、旱柳群落、芨芨草群落、马蔺群落和农田的有机碳含量均显著增加,而半固定沙地油蒿群落、流动沙地、沙柳群落的土壤有机碳含量随土层加深逐渐减少。各植被类型土壤有机碳密度在土壤剖面上的垂直分布格局和有机碳含量相似,但二者变化幅度不同。在流动沙地上进行植被建设有助于土壤固碳,且油蒿群落和沙柳群落的固碳效果较好。应尽量减少和避免强烈的人为干扰,以防止浅层土壤碳发生变化,维持土壤碳库稳定。
EvansRD, KoyamaA, SondereggerDL, ChenX, MaisupovaB, MadaminovAA, HanQ, DjenbaevBM ( 2014). Greater ecosystem carbon in the Mojave Desert after ten years exposure to elevated CO2 Nature Climate Change, 4, 394-397. DOI:10.1038/NCLIMATE2184URL [本文引用: 2] Carbon dioxide is the main greenhouse gas inducing climate change. Increased global CO2 emissions, estimated at 8.4 Pg C yr611 at present, have accelerated from 1% yr611 during 1990–1999 to 2.5% yr611 during 2000–2009 (ref. 1). The carbon balance of terrestrial ecosystems is the greatest unknown in the global C budget because the actual magnitude, location and causes of terrestrial sinks are uncertain2; estimates of terrestrial C uptake, therefore, are often based on the residuals between direct measurements of the atmospheric sink and well-constrained models of ocean uptake of CO2...
FangJY, YangYH, MaWH, MohhamotA, ShenHH ( 2010). Ecosystem carbon stocks and their changes in China’s grasslands Scientia Sinica Vitae, 40, 566-576. [本文引用: 2]
GaoYH, LiXR, LiuLC, JiaRL, YangHT, LiG, WeiY ( 2012). Seasonal variation of carbon exchange from a revegetation area in a Chinese desert Agricultural and Forest Meteorology, 156, 134-142. DOI:10.1016/j.agrformet.2012.01.007URL [本文引用: 1] Revegetation is a common method to combat desertification and to reduce soil carbon loss in arid and semi-arid areas worldwide. Revegetated areas in the temperate arid Shapotou area of Northern China were established to stabilize sand dunes in an area subject to wind erosion. The objective of this study was to characterize the carbon flux of a 20-year old revegetated area. During the measurement period from May 2008 to December 2010 carbon flux was measured by the eddy covariance technique, and rainfall, temperature, and soil water content were monitored simultaneously with the aim of understanding the relationship between carbon flux and meteorological factors. We found that the revegetated ecosystem is a carbon sink during the growing season when most of the annual precipitation falls. The revegetated area changed from being a carbon source to a sink in the 1 or 2 days following an effective rainfall event. In the non-growing season the ecosystem was a carbon source. The soil water content (SWC) affected the relationship between ecosystem respiration and temperature. The nighttime respiration rate increased with soil temperature. When the SWC was 3% the correlation between respiration rate and soil temperature was stronger (R2=0.23) than when SWC was <3% (R2=0.08). Although the net ecosystem carbon flux (NEE) in temperate arid desert areas is relatively small compared to that of humid areas, it represents huge carbon fixation potential because of the very large area of desert in northern China. Further research is required to clarify the effects of climatic factors on the various components of temperate arid ecosystems, especially the roles of cryptogam and biological soil crusts in the carbon exchange processes.
HastingsSJ, OechelWC, Muhlia-MeloA ( 2005). Diurnal, seasonal and annual variation in the net ecosystem CO2 exchange of a desert shrub community (Sarcocaulescent) in Baja California, Mexico Global Change Biology, 11, 927-939. DOI:10.1111/j.1365-2486.2005.00951.xURL [本文引用: 1] Abstract Estimates of net ecosystem exchange (NEE) of CO 2 have been measured on a variety of ecosystems world wide including grasslands, savannahs, boreal, pine, deciduous, Mediterranean and tropical rain forests as well as arctic tundra. While there have been numerous comparisons between net primary productivity of arid and semiarid grasslands and shrublands, notably lacking are estimates of NEE with a few exceptions. The objective of this study was to characterize the seasonal and annual carbon flux of a desert shrub ecosystem using the eddy covariance technique to determine the sensitivity of the system to the timing and varying amounts of precipitation. Measurements began in July of 2001, a year with 339mm of rainfall, considerably above the long-term average of 174mm and preceded by 2 years of below average rainfall (50–62mm). Over the 2 complete years of measurements, precipitation was 147 and 197mm in 2002 and 2003, respectively. In all years, the majority of the precipitation fell between August and September. The site was a sink of 6139gCm 612 yr 611 in 2002 with a relatively strong uptake in the early part of the year and reduced uptake after the suboptimal rainfall in September. This contrasts with 2003 when the ecosystem took up 6152gCm 612 yr 611 concentrated in the fall after significant rain in August and September. Likely, extremely low rainfall years would result in a carbon loss while a strengthening of the typical winter secondary peak in precipitation (notably absent in the 2 years of measurements) may extend uptake into the spring resulting in more carbon accumulation. The system appears to be buffered against variations in annual rainfall attributed to water storage in the stems and roots.
HouXY ( 1982). Vegetation Map of the People’s Republic of China and Its Illustration. China Cartographic Publishing House, Beijing. [本文引用: 1]
JanzenHH ( 2004). Carbon cycling in earth systems—A soil science perspective Agriculture Ecosystems & Environment, 104, 399-417. DOI:10.1016/j.agee.2004.01.040URL [本文引用: 1] The changing carbon cycle poses new questions for scientists. Now we will be asked, not how things are, but how they will be. For example: How will changes in CO 2 alter flows of carbon through biological carbon stocks? Can we manage ecosystems to hold more carbon? Are current carbon stores vulnerable should the earth warm, or water cycles shift, or nitrogen flows be altered? What will the C cycle look like a century from now; and will it then still provide all that we expect from it? These and other new questions may elicit from us fresh insights and approaches. We may learn to look more broadly at the C cycle, seeing all the ‘ecosystem services’ (not just C sequestration). We may insist on studies yielding deeper understanding of the C cycle, relevant beyond current issues. We may further emphasize ‘time’ in our studies, looking more at flows and changes than at describing what is—and looking long enough to see even subtle shifts. We may learn to follow C beyond the usual boundaries set by arbitrary disciplines. And we may come to see, more than before, how the carbon cycle weaves through our fields and skies and forests—and find new ways to reveal its grandeur to those who have not yet seen it. And then, it may happen that our successors, a century from now, will look back, almost in envy, at the urgent, enticing questions we were given to solve.
JasoniRL, SmithSD, ArnoneJA ( 2005). Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2 Global Change Biology, 11, 749-756. DOI:10.1111/j.1365-2486.2005.00948.xURL [本文引用: 1] Abstract Arid ecosystems, which occupy about 35% of the Earth's terrestrial surface area, are believed to be among the most responsive to elevated [CO 2 ]. Net ecosystem CO 2 exchange (NEE) was measured in the eighth year of CO 2 enrichment at the Nevada Desert Free-Air CO 2 Enrichment (FACE) Facility between the months of December 2003–December 2004. On most dates mean daily NEE (24h) (μmolCO 2 m 612 s 611 ) of ecosystems exposed to elevated atmospheric CO 2 were similar to those maintained at current ambient CO 2 levels. However, on sampling dates following rains, mean daily NEEs of ecosystems exposed to elevated [CO 2 ] averaged 23 to 56% lower than mean daily NEEs of ecosystems maintained at ambient [CO 2 ]. Mean daily NEE varied seasonally across both CO 2 treatments, increasing from about 0.1μmolCO 2 m 612 s 611 in December to a maximum of 0.5–0.6μmolCO 2 m 612 s 611 in early spring. Maximum NEE in ecosystems exposed to elevated CO 2 occurred 1 month earlier than it did in ecosystems exposed to ambient CO 2 , with declines in both treatments to lowest seasonal levels by early October (0.09±0.03μmolCO 2 m 612 s 611 ), but then increasing to near peak levels in late October (0.36±0.08μmolCO 2 m 612 s 611 ), November (0.28±0.03μmolCO 2 m 612 s 611 ), and December (0.54±0.06μmolCO 2 m 612 s 611 ). Seasonal patterns of mean daily NEE primarily resulted from larger seasonal fluctuations in rates of daytime net ecosystem CO 2 uptake which were closely tied to plant community phenology and precipitation. Photosynthesis in the autotrophic crust community (lichens, mosses, and free-living cyanobacteria) following rains were probably responsible for the high NEEs observed in January, February, and late October 2004 when vascular plant photosynthesis was low. Both CO 2 treatments were net CO 2 sinks in 2004, but exposure to elevated CO 2 reduced CO 2 sink strength by 30% (positive net ecosystem productivity=127±17gCm 612 yr 611 ambient CO 2 and 90±11gCm 612 yr 611 elevated CO 2 , P =0.011). This level of net C uptake rivals or exceeds levels observed in some forested and grassland ecosystems. Thus, the decrease in C sequestration seen in our study under elevated CO 2 – along with the extensive coverage of arid and semi-arid ecosystems globally – points to a significant drop in global C sequestration potential in the next several decades because of responses of heretofore overlooked dryland ecosystems.
JobbágyEG, SalaOE ( 2000). Controls of grass and shrub aboveground production in the Patagonian steppe Ecological Applications, 10, 541-549. DOI:10.2307/2641113URL [本文引用: 2] We studied the temporal dynamics of aboveground net primary production (ANPP) and senescence in a semiarid steppe representative of the Occidental District of Patagonia. We focused on the association of ANPP and senescence with climatic fluctuations at annual and seasonal scales. We estimated annual ANPP based on biomass harvests of grasses and shrubs (the dominant functional groups) at the peak of the growing season (January). The ANPP and senescence of grasses alone were also estimated for several periods within each year. The mean annual ANPP of the studied community for 10 years was 56 g m-2 yr-1, and the coefficient of variation was 26%. Grass and shrub mean annual ANPP had similar levels. Annual shrub production was associated with cumulative precipitation for the whole year, whereas annual grass production was not associated with precipitation. The average seasonal dynamics of grass ANPP showed its peak during spring, and there was no grass production during late summer and early fall. The peak green biomass of grass was achieved in early summer. Average senescence rates were more constant throughout the year, except for winter, when senescence was almost interrupted. Winter grass ANPP was positively associated with fall temperature, spring grass ANPP was positively related to winter precipitation, and summer grass ANPP was positively related to spring precipitation. Grass senescence during spring was negatively related to precipitation in the same season. Our results highlight the importance of separating functional types in productivity studies. The differential effect of precipitation on grass and shrub ANPP masked the association when these functional groups were pooled. ANPP association with climatic variables in the Occidental District of the Patagonian steppe shifted from precipitation alone to precipitation and temperature when the temporal scale of analysis changed from annual to seasonal.
KirschbaumMUF ( 2000). Will changes in soil organic carbon act as a positive or negative feedback on global warming? Biogeochemistry, 48, 21-51. DOI:10.1023/A:1006238902976URL [本文引用: 1] The world's soils contain about 1500 Gt of organic carbon to a depth of 1m and a further 900 Gt from 1--2m. A change of total soil organic carbon by just 10% would thus be equivalent to all the anthropogenic CO 2 emitted over 30 years. Warming is likely to increase both the rate of decomposition and net primary production (NPP), with a fraction of NPP forming new organic carbon. Evidence from various sources can be used to assess whether NPP or the rate of decomposition has the greater temperature sensitivity, and, hence, whether warming is likely to lead to an increase or decrease in soil organic carbon.Evidence is reviewed from laboratory-based incubations, field measurements of organic carbon storage, carbon isotope ratios and soil respiration with either naturally varying temperatures or after experimentally increasing soil temperatures. Estimates of terrestrial carbon stored at the Last Glacial Maximum are also reviewed. The review concludes that the temperature dependence of organic matter decomposition can be best described as: d(T) = exp[3.36 (T 61 40)/(T + 31.79)] where d(T) is the normalised decomposition rate at temperature T (in °C). In this equation, decomposition rate is normalised to ‘1’ at 40 °C.The review concludes by simulating the likely changes in soil organic carbon with warming. In summary, it appears likely that warming will have the effect of reducing soil organic carbon by stimulating decomposition rates more than NPP. However, increasing CO 2 is likely to simultaneously have the effect of increasing soil organic carbon through increases in NPP. Any changes are also likely to be very slow. The net effect of changes in soil organic carbon on atmospheric CO 2 loading over the next decades to centuries is, therefore, likely to be small.
LiC, ZhangC, LuoG, ChenX, MaisupovaB, MadaminovAA, HanQ, DjenbaevBM ( 2015). Carbon stock and its responses to climate change in Central Asia Global Change Biology, 21, 1951-1967. DOI:10.1111/gcb.12846URLPMID:25626071 [本文引用: 1] Abstract Central Asia has a land area of 5.602×0210602km2 and contains 80–90% of the world's temperate deserts. Yet it is one of the least characterized areas in the estimation of the global carbon (C) stock/balance. This study assessed the sizes and spatiotemporal patterns of C pools in Central Asia using both inventory (based on 353 biomass and 284 soil samples) and process-based modeling approaches. The results showed that the C stock in Central Asia was 31.34–34.1602Pg in the top 1-m soil with another 10.42–11.4302Pg stored in deep soil (1–302m) of the temperate deserts. They amounted to 18–24% of the global C stock in deserts and dry shrublands. The C stock was comparable to that of the neighboring regions in Eurasia or major drylands around the world (e.g. Australia). However, 90% of Central Asia C pool was stored in soil, and the fraction was much higher than in other regions. Compared to hot deserts of the world, the temperate deserts in Central Asia had relatively high soil organic carbon density. The C stock in Central Asia is under threat from dramatic climate change. During a decadal drought between 1998 and 2008, which was possibly related to protracted La Ni09a episodes, the dryland lost approximately 0.4602Pg C from 1979 to 2011. The largest C losses were found in northern Kazakhstan, where annual precipitation declined at a rate of 9002mm02decade611. The regional C dynamics were mainly determined by changes in the vegetation C pool, and the SOC pool was stable due to the balance between reduced plant-derived C influx and inhibited respiration.
LiD, HuangY, WuQ, MingZ, JinDY ( 2010). Modeling dynamics of soil organic carbon in an alpine meadow ecosystem on Qinghai-Tibetan Plateau using the Century model Acta Prataculturae Sinica, 19(2), 160-168. DOI:10.11686/cyxb20100223Magsci [本文引用: 1] <FONT face=Verdana>利用高寒草甸区植被、土壤和气候等资料,借助Century模型研究了青藏高原高寒草甸生态系统土壤有机碳的动态。以2003年7月-2005年7月逐月气象观测资料为输入变量,模拟了自然条件下高寒矮嵩草草甸0~20cm 土壤微生物呼吸CO2-C 通量季节变化。模拟值与观测值进行的回归分析显示,二者具有较好的一致性(<EM>R</EM><SUP>2</SUP>=0.89,<EM>P</EM><0.05)。土壤有机碳动态模拟表明:1)自然条件下高寒草甸土壤有机碳在经历了一个快速积累过程后,积累速率逐渐趋于缓和,最终达到并接近稳定状态。稳定状态下0~20cm 土壤总有机碳库约7597.50~7694.10gC/m<SUP>2</SUP>。其中活性、缓性和惰性土壤有机碳组分占土壤总有机碳储量的2.80%,58.50% 和38.70%。2)过去45年(1960-2005)高寒草甸土壤有机碳呈振幅较为稳定的波动变化,但这种变化主要是土壤有机碳各组分波动变化的结果。气候波动对土壤有机碳影响主要与温度变化引起的惰性土壤有机碳库的变化有关,二者呈显著负相关(<EM>r</EM>=-0.548,<EM>P</EM><0.01)。降水量对土壤有机碳及有机碳各组分的影响不显著。</FONT> 李东, 黄耀, 吴琴, 明珠, 靳代樱 ( 2010). 青藏高原高寒草甸生态系统土壤有机碳动态模拟研究 草业学报, 19(2), 160-168. DOI:10.11686/cyxb20100223Magsci [本文引用: 1] <FONT face=Verdana>利用高寒草甸区植被、土壤和气候等资料,借助Century模型研究了青藏高原高寒草甸生态系统土壤有机碳的动态。以2003年7月-2005年7月逐月气象观测资料为输入变量,模拟了自然条件下高寒矮嵩草草甸0~20cm 土壤微生物呼吸CO2-C 通量季节变化。模拟值与观测值进行的回归分析显示,二者具有较好的一致性(<EM>R</EM><SUP>2</SUP>=0.89,<EM>P</EM><0.05)。土壤有机碳动态模拟表明:1)自然条件下高寒草甸土壤有机碳在经历了一个快速积累过程后,积累速率逐渐趋于缓和,最终达到并接近稳定状态。稳定状态下0~20cm 土壤总有机碳库约7597.50~7694.10gC/m<SUP>2</SUP>。其中活性、缓性和惰性土壤有机碳组分占土壤总有机碳储量的2.80%,58.50% 和38.70%。2)过去45年(1960-2005)高寒草甸土壤有机碳呈振幅较为稳定的波动变化,但这种变化主要是土壤有机碳各组分波动变化的结果。气候波动对土壤有机碳影响主要与温度变化引起的惰性土壤有机碳库的变化有关,二者呈显著负相关(<EM>r</EM>=-0.548,<EM>P</EM><0.01)。降水量对土壤有机碳及有机碳各组分的影响不显著。</FONT>
LiXR ( 2012). Eco-hydrology of Biological Soil Crusts in Desert Regions of China. Higher Education Press, Beijing. [本文引用: 1]
LiXR, HeMZ, DuanZH, XiaoHL, JiaXH ( 2007a ). Recovery of topsoil physicochemical properties in revegetated sites in the sand-burial ecosystems of the Tengger Desert, northern China Geomorphology, 88, 254-265. DOI:10.1016/j.geomorph.2006.11.009URL [本文引用: 1] Recovery in soil properties and processes after sand burial in the Tengger Desert, northern China, was documented at five different-aged revegetated sites (1956, 1964, 1973, 1982, and 1991) and at a reference site with native vegetation, which had never been damaged by sand burial and was enclosed for grazing. The proportions of silt and clay, depth of topsoil and biological soil crusts, and concentrations of soil organic C, K, total N and total P increased with years since revegetation. Most characteristics of topsoil (0–5cm) characteristics had recovered to 60% of those measured at the reference site by 50years after sand-binding vegetation had been established. Exceptions were electrical conductivity and contents of sand, silt, CaCO 3 and organic C, which recovered to 20–40% of the values at the reference site. The difference in annual recovery rates of soil properties between the two most recently revegetated sites (0–14years) was greater than the difference between the two oldest revegetated sites (43–50years). Best-fit asymptote models showed that the estimated times for the soil properties in the 50-year-old site to reach the same levels as in the reference site (i.e. an undisturbed, native steppified desert ecosystem) would be between 23 and 245years, but for some properties even maximum recovery after >0250years still fell significantly short of the level at the reference site. These results suggest that soil recovery is a slow process in an extremely arid desert environment, and therefore the conservation of soil habitat is a crucial issue for land managers.
LiXR, KongDS, TanHJ, WangXP ( 2007b ). Changes in soil and vegetation following stabilisation of dunes in the southeastern fringe of the Tengger Desert, China Plant and Soil, 300, 221-231. DOI:10.1007/s11104-007-9407-1URL [本文引用: 1]
LiXR, ZhangZS, LiuYB, LiXJ, YangHT ( 2016). Fundamental Ecohydrology of Ecological Restoration and Recovery in Sandy Desert Regions of China. Science Press, Beijing. [本文引用: 2]
LuyssaertS, IngliinaI, JungM, RichardsonA, ReichsteinM, PapaleD, PiaoS, SchulzeED, WingateL, MatteucciG ( 2007). CO2 balance of boreal, temperate, and tropical forests derived from a global database Global Change Biology, 13, 2509-2537. DOI:10.1111/gcb.2007.13.issue-12URL [本文引用: 1]
MaWH ( 2006). Carbon Storage of Grasslands in Inner Mongolia. PhD dissertation, Peking University, Beijing. [本文引用: 2]
MohhamotA ( 2006). Carbon and Nitrogen Storage of Grassland Ecosystem in Xinjiang. PhD dissertation, Peking University Beijing. URL [本文引用: 2] 草地生态系统占陆地生态系统总面积的25﹪,是地球生物圈的重要组成部分,在全球碳循环和气候变化中发挥着重要作用。新疆地处中亚、西伯利亚、蒙古、西藏的交会处,地形复杂,草地类型丰富,在我国草地碳库中占据重要地位。研究新疆草地植被、土壤碳、氮库大小及其分布机制和决定因素,对评价新疆干旱荒漠区温带草地生物地球化学循环对我国乃至全球变化的响应和贡献具有重要的意义。 本文基于在新疆天山、阿尔泰山、准噶尔西部山地以及帕米尔高原等地区实地调查获取的460个生物量样方和276个土壤剖面数据,结合新疆土壤普查的部分数据,研究了新疆主要草地植... 安尼瓦尔·买买提 ( 2006). 新疆草地生态系统碳、氮储量的研究 博士学位论文, 北京大学, 北京.] URL [本文引用: 2] 草地生态系统占陆地生态系统总面积的25﹪,是地球生物圈的重要组成部分,在全球碳循环和气候变化中发挥着重要作用。新疆地处中亚、西伯利亚、蒙古、西藏的交会处,地形复杂,草地类型丰富,在我国草地碳库中占据重要地位。研究新疆草地植被、土壤碳、氮库大小及其分布机制和决定因素,对评价新疆干旱荒漠区温带草地生物地球化学循环对我国乃至全球变化的响应和贡献具有重要的意义。 本文基于在新疆天山、阿尔泰山、准噶尔西部山地以及帕米尔高原等地区实地调查获取的460个生物量样方和276个土壤剖面数据,结合新疆土壤普查的部分数据,研究了新疆主要草地植...
PostWM, EmanuelWR, ZinkePJ, StangenbergerAG ( 1982). Soil carbon pools and world life zones Nature, 298, 156-159. DOI:10.1038/298156a0URL [本文引用: 1]
PostWM, PengTH, EmanuelWR, KingAW, DaleVH, DeAngelisDL ( 1990). The global carbon cycle American Scientist, 78, 310-326. [本文引用: 1]
RotenbergE, YakirD ( 2010). Contribution of semi-arid forests to the climate system Science, 327, 451-454. DOI:10.1126/science.1179998URLPMID:20093470 [本文引用: 2] Abstract Forests both take up CO2 and enhance absorption of solar radiation, with contrasting effects on global temperature. Based on a 9-year study in the forests' dry timberline, we show that substantial carbon sequestration (cooling effect) is maintained in the large dry transition zone (precipitation from 200 to 600 millimeters) by shifts in peak photosynthetic activities from summer to early spring, and this is counteracted by longwave radiation (L) suppression (warming effect), doubling the forestation shortwave (S) albedo effect. Several decades of carbon accumulation are required to balance the twofold S + L effect. Desertification over the past several decades, however, contributed negative forcing at Earth's surface equivalent to approximately 20% of the global anthropogenic CO2 effect over the same period, moderating warming trends.
SchlesingerWH, BelnapJ, MarionG ( 2009). On carbon sequestration in desert ecosystems Global Change Biology, 15, 1488-1490. DOI:10.1111/j.1365-2486.2008.01763.xURL [本文引用: 1] Recent reports of net ecosysytem production >100 gCm 612 yr 611 in deserts are incompatible with existing measurements of net primary production and carbon pools in deserts. The comparisions suggest that gas exchange measurements should be used with caution and better validation if they are expected to indicate the magnitude of carbon sink in these ecosysytems.
StoneR ( 2008). Have desert researchers discovered a hidden loop in the carbon cycle? Science, 320, 1409-1410. DOI:10.1126/science.320.5882.1409URLPMID:18556524 [本文引用: 2] Findings in two deserts on opposite sides of the world suggest that deserts are a larger sink for carbon dioxide than scientists had assumed.
WangM ( 2014). Vegetation Biomass and Soil Organic Carbon Storage in Desert Grasslands of Hexi Corridor. PhD dissertation, University of Chinese Academy of Sciences Beijing. URL [本文引用: 1] 草地是陆地生态系统的重要组成部分,是地球表面分布最广的植被类型之一,在全球生态系统碳循环中发挥着极为重要的作用。准确估算草地生物量的大小,揭示其控制因素,阐明生物量的分配模式,对于认识草地生态系统土壤中的碳输入和整个生态系统的碳循环具有重要的意义。准确估算土壤有机碳的储量,揭示其分布格局和控制因素将有助于评价土壤和草地生态系统在陆地生态系统碳循环中的重要作用。但是,目前关于草地生态系统生物量和碳储量大小、分配格局及其控制因素等方面的研究主要集中在温带草地,重点关注干旱荒漠草地生态系统的相关研究较少。河西走廊荒... 王敏 ( 2014). 河西走廊荒漠草地生物量和土壤有机碳储量 博士学位论文, 中国科学院大学, 北京. URL [本文引用: 1] 草地是陆地生态系统的重要组成部分,是地球表面分布最广的植被类型之一,在全球生态系统碳循环中发挥着极为重要的作用。准确估算草地生物量的大小,揭示其控制因素,阐明生物量的分配模式,对于认识草地生态系统土壤中的碳输入和整个生态系统的碳循环具有重要的意义。准确估算土壤有机碳的储量,揭示其分布格局和控制因素将有助于评价土壤和草地生态系统在陆地生态系统碳循环中的重要作用。但是,目前关于草地生态系统生物量和碳储量大小、分配格局及其控制因素等方面的研究主要集中在温带草地,重点关注干旱荒漠草地生态系统的相关研究较少。河西走廊荒...
WohlfahrtQ, FenstermakerLF, ArnoneJA ( 2008). Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem Global Change Biology, 14, 1475-1487. DOI:10.1111/j.1365-2486.2008.01593.xURL [本文引用: 2] Abstract The net ecosystem CO 2 exchange (NEE) between a Mojave Desert ecosystem and the atmosphere was measured over the course of 2 years at the Mojave Global Change Facility (MGCF, Nevada, USA) using the eddy covariance method. The investigated desert ecosystem was a sink for CO 2 , taking up 102±67 and 110±70gCm 612 during 2005 and 2006, respectively. A comprehensive uncertainty analysis showed that most of the uncertainty of the inferred sink strength was due to the need to account for the effects of air density fluctuations on CO 2 densities measured with an open-path infrared gas analyser. In order to keep this uncertainty within acceptable bounds, highest standards with regard to maintenance of instrumentation and flux measurement postprocessing have to be met. Most of the variability in half-hourly NEE was explained by the amount of incident photosynthetically active radiation (PAR). On a seasonal scale, PAR and soil water content were the most important determinants of NEE. Precipitation events resulted in an initial pulse of CO 2 to the atmosphere, temporarily reducing NEE or even causing it to switch sign. During summer, when soil moisture was low, a lag of 3–4 days was observed before the correlation between NEE and precipitation switched from positive to negative, as opposed to conditions of high soil water availability in spring, when this transition occurred within the same day the rain took place. Our results indicate that desert ecosystem CO 2 exchange may be playing a much larger role in global carbon cycling and in modulating atmospheric CO 2 levels than previously assumed – especially since arid and semiarid biomes make up >30% of Earth's land surface.
XieJ, LiY, ZhaiC, LiC, LanZ ( 2009). CO2 absorption by alkaline soils and its implication to the global carbon cycle Environmental Geology, 56, 953-961. DOI:10.1007/s00254-008-1197-0URL [本文引用: 1] Motivated by the rapid increase in atmospheric CO2 due to human activities since the Industrial Revolution, and the climate changes it produced, the world0964s concerned scientific community has made a huge effort to investigate the global carbon cycle. However, the results reveal that the global CO2 budget cannot be balanced, unless a 0904missing sink0909 is invoked. Although numerous studies claimed to find the 0904missing sink0909, none of those claims has been widely accepted. This current study showed that alkaline soil on land are absorbing CO2 at a rate of 0.309“3.0 0204mol m0903’2 s0903’1 with an inorganic, non-biological process. The intensity of this CO2 absorption is determined by the salinity, alkalinity, temperature and water content of the saline/alkaline soils, which are widely distributed on land. Further studies revealed that high salinity or alkalinity positively affected the CO2 absorbing intensity, while high temperature and water content had a negative effect on the CO2 absorbing intensity of these soils. This inorganic, non-biological process of CO2 absorption by alkaline soils might have significant implications to the global carbon budget accounting.
YangHT, LiXR, WangZR, JiaRL, LiuLC, ChenYL, WeiYP, GaoYH, LiG ( 2014). Carbon sequestration capacity of shifting sand dune after establishing new vegetation in the Tengger Desert, northern China Science of the Total Environment, 478, 1-11. DOI:10.1016/j.scitotenv.2014.01.063URLPMID:24530579 [本文引用: 1] Abstract Reconstructing vegetation in arid and semiarid areas has become an increasingly important management strategy to realize habitat recovery, mitigate desertification and global climate change. To assess the carbon sequestration potential in areas where sand-binding vegetation has been established on shifting sand dunes by planting xeric shrubs located near the southeastern edge of the Tengger Desert in northern China, we conducted a field investigation of restored dune regions that were established at different times (20, 30, 47, and 55 years ago) in the same area. We quantified the total organic carbon (TOC) in each ecosystem by summing the individual carbon contributions from the soil (soil organic carbon; SOC), shrubs, and grasses in each system. We found that the TOC, as well as the amount of organic carbon in the soil, shrubs, and grasses, significantly increased over time in the restored areas. The average annual rate of carbon sequestration was highest in the first 20 years after restoration (3.26 0103 10(-2)kg00·m(-2) 00·year(-1)), and reached a stable rate (2.14 0103 10(-2) kg00·m(-2) 00·year(-1)) after 47 years. Organic carbon storage in soil represented the largest carbon pool for both restored systems and a system containing native vegetation, accounting for 67.6%-85.0% of the TOC. Carbon in grass root biomass, aboveground grass biomass, litter, aboveground shrub biomass, and shrub root biomass account for 10.0%-21.0%, 0.2%-0.6%, 0.1%-0.2%, 1.7%-12.1% and 0.9%-6.2% of the TOC, respectively. Furthermore, we found that the 55-year-old restored system has the capacity to accumulate more TOC (1.02 kg00·m(-2) more) to reach the TOC level found in the natural vegetation system. These results suggest that restoring desert ecosystems may be a cost-effective and environmentally friendly way to sequester CO2 from the atmosphere and mitigate the effects of global climate change. Copyright 0008 2014 Elsevier B.V. All rights reserved.
Seasonal variation of carbon exchange from a revegetation area in a Chinese desert 1 2012
... id="C7">荒漠生态系统是陆地生态系统的重要组成部分, 面积占全球陆地面积的30%以上, 碳储量约为全球碳储量的8% (Janzen, 2004; Li et al., 2015), 主要以土壤碳的形式存在(Wohlfahrt et al., 2008; 赵有益等, 2008).由于荒漠生态系统较为脆弱, 土壤碳储量容易受环境变化的影响, 在气候变化和人类活动加剧的背景下, 荒漠生态系统土壤碳在全球碳循环中可能起着更加重要的作用.有研究认为荒漠生态系统可能是全球碳循环长期寻找的“碳失汇”所在地, 近年来的研究表明荒漠生态系统具有很强的固碳潜力, 是未来固定CO2的重要区域(Stone, 2008; Rotenberg & Yakir, 2010; Evans et al., 2014), 但是, 目前还缺乏强有力的证据(Luyssaert et al., 2007; Wohlfahrt et al., 2008; Schlesinger et al., 2009).环境的复杂性及高度的空间异质性给荒漠生态系统碳储量的精确评估带来较大的困难(Jasoni et al., 2005; Xie et al., 2009), 与其他生态系统相比, 荒漠生态系统土壤碳储量研究还很薄弱(Hastings et al., 2005; Gao et al., 2012).近年来, 我国典型荒漠生态系统碳储量的研究逐渐增多, 如新疆温性荒漠碳储量(安尼瓦尔·买买提, 2006; 门学慧, 2013)、河西走廊荒漠草地碳储量(王敏, 2014)、内蒙古荒漠草原碳储量(马文红, 2006; 杨婷婷, 2013), 虽然这些研究极大地丰富和完善了我国荒漠草地碳储量研究, 但涉及荒漠植物群落与土壤碳储量关系的研究还较少, 还远不能代表我国丰富的荒漠植物群落的土壤碳储量特征.现有的研究结果显示, 荒漠生态系统土壤碳储量估算结果不确定性极为明显, 这已成为国家和区域尺度上碳储量评估以及国家政策制定的主要障碍.腾格里沙漠东南缘植被为典型的荒漠草原类型, 是草原化荒漠与荒漠化草原的过渡带, 地带性植被发育在波状起伏的沙质草原上, 母质是洪积冲积物, 该区灌木、半灌木群落类型丰富.由稀疏植被或荒漠植被构成干旱地区典型的生态系统(斑块植被结构或镶嵌植被结构), 这种植被类型基本是阿拉善高原沙质、沙砾质荒漠区的典型代表.其特殊的地理位置和气候条件, 使得该区植物类型及其生物适应性特征更具独特性(李新荣等, 2016).研究不同类型荒漠植物群落SOC密度特征, 有助于我们在区域尺度上认识SOC的分布特征并在此基础上精确地估算荒漠生态系统SOC储量, 减少荒漠生态系统土壤碳储量估算的不确定性, 也对应对气候变化、制定提高土壤碳储量的管理措施具有重要意义.本研究选择腾格里沙漠东南缘现有的主要类型群落研究SOC含量、分布特点及其差异, 旨在全面地阐明在腾格里沙漠南缘这一特殊生境植被类型对SOC的影响. ...
Diurnal, seasonal and annual variation in the net ecosystem CO2 exchange of a desert shrub community (Sarcocaulescent) in Baja California, Mexico 1 2005
... id="C7">荒漠生态系统是陆地生态系统的重要组成部分, 面积占全球陆地面积的30%以上, 碳储量约为全球碳储量的8% (Janzen, 2004; Li et al., 2015), 主要以土壤碳的形式存在(Wohlfahrt et al., 2008; 赵有益等, 2008).由于荒漠生态系统较为脆弱, 土壤碳储量容易受环境变化的影响, 在气候变化和人类活动加剧的背景下, 荒漠生态系统土壤碳在全球碳循环中可能起着更加重要的作用.有研究认为荒漠生态系统可能是全球碳循环长期寻找的“碳失汇”所在地, 近年来的研究表明荒漠生态系统具有很强的固碳潜力, 是未来固定CO2的重要区域(Stone, 2008; Rotenberg & Yakir, 2010; Evans et al., 2014), 但是, 目前还缺乏强有力的证据(Luyssaert et al., 2007; Wohlfahrt et al., 2008; Schlesinger et al., 2009).环境的复杂性及高度的空间异质性给荒漠生态系统碳储量的精确评估带来较大的困难(Jasoni et al., 2005; Xie et al., 2009), 与其他生态系统相比, 荒漠生态系统土壤碳储量研究还很薄弱(Hastings et al., 2005; Gao et al., 2012).近年来, 我国典型荒漠生态系统碳储量的研究逐渐增多, 如新疆温性荒漠碳储量(安尼瓦尔·买买提, 2006; 门学慧, 2013)、河西走廊荒漠草地碳储量(王敏, 2014)、内蒙古荒漠草原碳储量(马文红, 2006; 杨婷婷, 2013), 虽然这些研究极大地丰富和完善了我国荒漠草地碳储量研究, 但涉及荒漠植物群落与土壤碳储量关系的研究还较少, 还远不能代表我国丰富的荒漠植物群落的土壤碳储量特征.现有的研究结果显示, 荒漠生态系统土壤碳储量估算结果不确定性极为明显, 这已成为国家和区域尺度上碳储量评估以及国家政策制定的主要障碍.腾格里沙漠东南缘植被为典型的荒漠草原类型, 是草原化荒漠与荒漠化草原的过渡带, 地带性植被发育在波状起伏的沙质草原上, 母质是洪积冲积物, 该区灌木、半灌木群落类型丰富.由稀疏植被或荒漠植被构成干旱地区典型的生态系统(斑块植被结构或镶嵌植被结构), 这种植被类型基本是阿拉善高原沙质、沙砾质荒漠区的典型代表.其特殊的地理位置和气候条件, 使得该区植物类型及其生物适应性特征更具独特性(李新荣等, 2016).研究不同类型荒漠植物群落SOC密度特征, 有助于我们在区域尺度上认识SOC的分布特征并在此基础上精确地估算荒漠生态系统SOC储量, 减少荒漠生态系统土壤碳储量估算的不确定性, 也对应对气候变化、制定提高土壤碳储量的管理措施具有重要意义.本研究选择腾格里沙漠东南缘现有的主要类型群落研究SOC含量、分布特点及其差异, 旨在全面地阐明在腾格里沙漠南缘这一特殊生境植被类型对SOC的影响. ...
 ,*, 贾荣亮*中国科学院西北生态环境资源研究院, 沙坡头沙漠研究试验站, 兰州 730000
,*, 贾荣亮*中国科学院西北生态环境资源研究院, 沙坡头沙漠研究试验站, 兰州 730000
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT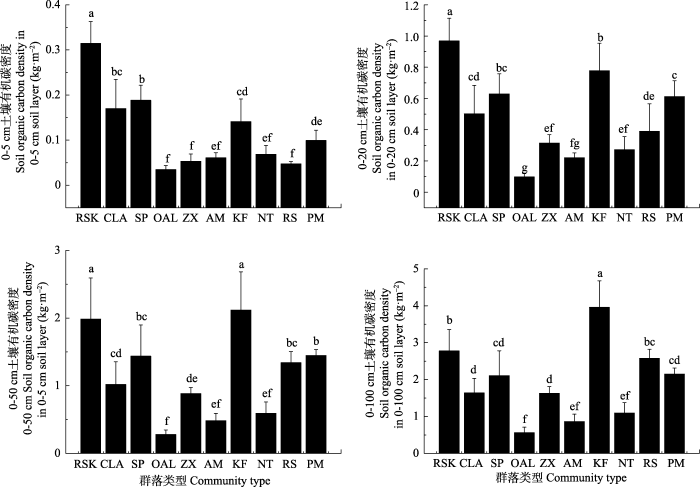 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}