 ,*, ��־ǿ*, ������,*���ɹ�ũҵ��ѧ��ԭ����Դ����ѧԺ, ���ͺ��� 010019
,*, ��־ǿ*, ������,*���ɹ�ũҵ��ѧ��ԭ����Դ����ѧԺ, ���ͺ��� 010019Net ecosystem carbon exchange characteristics in Stipa breviflora desert steppe with different stocking rates
JIN Yu-Xi, LIU Fang, ZHANG Jun, HAN Meng-Qi, WANG Zhong-Wu,*, QU Zhi-Qiang*, HAN Guo-Dong,*College of Grassland, Resources and Environment, Inner Mongolia Agricultural University, Hohhot 010019, ChinaͨѶ����:
��ί: �ƽ���
���α༭: ����
| ��������: |
Online:2018-03-20
| Supported by: | SupportedbytheNationalNaturalScienceFoundationofChina( |

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (1613KB)ժҪҳ����ά�����������������EndNote|Ris|Bibtex�ղر���
����
������, ����, �ž�, ������, ������, ��־ǿ, ������. ��ͬ�����ʴ����¶̻���é��Į��ԭ��̬ϵͳ��̼��������. ֲ����̬ѧ��, 2018, 42(3): 361-371 doi:10.17521/cjpe.2017.0066
JIN Yu-Xi, LIU Fang, ZHANG Jun, HAN Meng-Qi, WANG Zhong-Wu, QU Zhi-Qiang, HAN Guo-Dong.
�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ�⡣��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ����ʡ���̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��)������Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(Xu et al., 2004; Ma et al., 2007; Zhou et al., 2007)���ҹ�������̬ϵͳ̼�������о�����ظ�ԭ(�����, 2014; �����, 2016)���ݵ��ԭ(����, 2011; �����, 2018)�����Ͳ�ԭ(�����ҵ�, 2006; �콨����, 2016)�ͻ�Į��ԭ(���½�, 2015; ��ٻ��, 2016)���зֲ��������Dzݵص���Ҫ���÷�ʽ, ��������ͨ����ʳ����̤����йӰ��Ⱥ����������ֲ���ṹ�Լ������������ʺ�����(An et al., 2015), ����Ӱ����̬ϵͳ̼ͨ������֡������о�����, ������������̬ϵͳ��̼����, ���ȷ��������CO2���ͷ�((Frank et al., 2000; Kuzyakov & Gavrichkova, 2010)����ͬʱ���о������ʶȷ����ٽ��ݵ���̬ϵͳCO2����DZ��(Kang et al., 2013; ������, 2016), �����о����ַ����Բݵ�CO2ͨ��û������Ӱ��(Risch & Frank, 2006; ���IJ���, 2015)���ݵ�̼Դ��̼��״̬�Է�������Ӧ���ܴ��ڿռ�߶ȵIJ����ԡ��̻���é(Stipa breviflora)��ԭռ���Ի�Į��ԭ���������11.2% (��������, 2007), �Dz�ԭ�����Į�����ɵ���̬ϵͳ����, �Ի�Į��ԭ��̬ϵͳ�����幦��Ӱ��ܴ��о����������ض��˻��ݵ�ȫ��ʵʩΧ�����, ��̼DZ��ÿ���12.01 Tg C (Niu et al., 2011)������, �ݵصĹ�̼�����Բ�ͬ�ݵ����͡���ͬ�������÷�ʽ����Ӧ��ͬ(����������ȫ, 2015), ��˻���һ�������ȶ������в�ͬ�����ʵ�ʵ��ƽ̨���������о���ʮ�ֱ�Ҫ�ġ�����̽�������ù̶������ʷ���10�����̬ϵͳ̼��������, ּ�ڽ�ʾ��̬ϵͳ̼�����������ʵ���Ӧ������ˮ�����صĹ�ϵ�������˽�ݵ�̼Դ��̼���ת��״̬, ½����̬ϵͳ̼ѭ������������Ҫ���塣
1 ���Ϻͷ���
1.1 �о����ſ�
�����λ�����ɹ������������첼�����ɹ�ũ��ҵ��ѧԺ�����������(41.47�� N, 111.53�� E), ���θ߶�Ϊ1 450 m������������Ϊ��������, �л�̼ƽ������Ϊ1.3%, �������Ϊȱ�������ס���������ص㡣��������Ϊ���´���½�Լ�������, ������������, �ļ��̴���ˬ, ��ˮ�ٶ�����, ����ͬ��; �о�����ƽ������3.4 ��, 6-8���������, ��10 ��������Ϊ2 200-2 500 ��, ��˪��90-120�졣�꽵ˮ��288 mm, ��Ҫ�����ڲݵط�������ʱ���(5-9��), ���ڼ�Ľ�ˮ��ռȫ���80%���ϡ��õ���ֲ��������ɽ�ƶ��, Ⱥ��Ͱ�ϡ��, ��Ⱥ��Ϊ�̻���é, ������Ϊ����(Artemisia frigida)����â���Ӳ�(Cleistogenes songorica), ���ڵ��͵Ļ�Į��ԭ��̬ϵͳ��1.2 �������
��������ʼ��2004��, �ڶ̻���é��Į��ԭѡ��������ƽ̹��һ�µĵط�Χ��Լ50 hm2��Ȼ�ݵء�������ȫ��������������, ��Χ������Ϊ3������, ��3���ظ�, ��ÿ����������4������, ��4����ͬ��������ˮƽ, �ֱ�Ϊ������(CK)����ȷ���(LG)���жȷ���(MG)���ضȷ���(HG), 4��������ȫ������С���������С������������, Ϊ4.4 hm2���������Ǿ���(2000)�����ɹŻ�Į��ԭ�����ʵ��о����, �趨��ͬ��������ֵ�ֱ�Ϊÿ����ÿ����0 (CK)��0.91 (LG)��1.82 (MG)��2.71 (HG)��λ, �������������ֱ�Ϊ0��4��8��12ֻ, ÿ��6�³�ѡȡ2������ɹ�������ָ����С�������ɲ�ʳ, ÿ�����ʱ��Ϊ6:00-18:00, ������11�½��������о�ѡȡ�˲���������ȷ������ضȷ���3��������ˮƽ�����о���1.3 ���ݲɼ�
1.3.1 ��̬ϵͳ̼����2014-2016��������(5-10��)�ñ�Яʽ�����LI-6400 (LI-COR, Lincoln, USA)���ܱ�ʽ�䷨�ⶨ��̬ϵͳ��̼������ÿ��������鿪ʼǰ, ��ÿ������С���������3�����Ͻ�ˮ�ۿ�(50 �� 50 cm2), ��������8 cm�������, �������������ϲ�����ƽ�ȡ��ⶨǰ�Ƚ����ƴ�Ҷ�Һ�LI-6400����, �ⶨʱ��һ��50 �� 50 �� 50 cm3�����л�������(��Ҷ��)�����ڽ�������, ʹ��Ӵ����ܲ���֤�ܱա�Ϊʹ���������Ͼ���, �������ڲ����߶��Ǵ�װ������С���ȡ��ⶨʱ��ѡ���������Ƶ�����, ��8:00-12:00���вⶨ, ÿ�²ⶨ2��(����������ԭ����Щ�·�ֻ��һ��)��ÿ120 s�ⶨһ������, CO2Ũ�Ⱥ�ˮ��ͨ����ֵÿ10 s�Զ���¼һ��, ����ʱ�����С�CO2Ũ�Ⱥ�ˮ��ͨ��ֵ�仯���Լ������̬ϵͳ��CO2����(NEE)��NEE��ֵ��ʾ��CO2����, ��ֵ��ʾ��CO2�ͷš���̬ϵͳ�ܳ���������(GEP)����NEE��ER����ó���
1.3.2 �����¶���ʪ��
��LI-6400�ⶨ��ͬʱ, ��2��TP3001���¼Ʋⶨ����10 cm�¶�, �ȶ����¼���ݡ�ͬʱ, �ڲⶨCO2ͨ����ÿ�����㸽��, ʹ��ֱ��2.5 cm������ȡ��0-10 cm������Ʒ, װ��������, ��ʱ������¼ʪ����, ��105 ���º��24 h��Ƹ�����, ������������ˮ����
1.4 ���ݷ���
�������ڶ�βⶨ��ָ��, ʹ��SPSS 20.0�е��ظ��ⶨ������������鲻ͬ�����ʶ���̬ϵͳ��CO2�����������¶Ⱥ�ʪ�ȵ�Ӱ��, ����R���Է��������¶ȡ�����ʪ�ȶ�NEE�Ĺ����ʼ������Ҫ�ԡ���Sigmaplot 10.0������ͼ��2 ����ͷ���
2.1 ���ºͽ�ˮ
2014-2016����ƽ�����ºͽ�ˮ�ļ��ڶ�̬������ͼ1��ʾ, ��ƽ�����³ʵ���������, �����ƽ�����¼�����7��, �ֱ�Ϊ21.12��20.12��19.60 �档������ƽ�����·ֱ�Ϊ14.39��14.54��13.87 �档2014��2015�꽵ˮ��������, ��˫�������ߡ�6�·ݽ�ˮ���, 2014��2015��ֱ�Ϊ36.1��44.2 mm, 9��ĩ10�³��ٴγ��ַ�ֵ���������������ۻ���ˮ���ֱ�Ϊ195.6��120.7 mm��2016�꽵ˮ������, ����Ϊ�ǶԳ��Եĵ�������, ��Ҫ������6-7��, 7�µĽ�ˮ���ﵽ131.2 mm, �����������ۻ���ˮ��Ϊ332.9 mm��
ͼ1
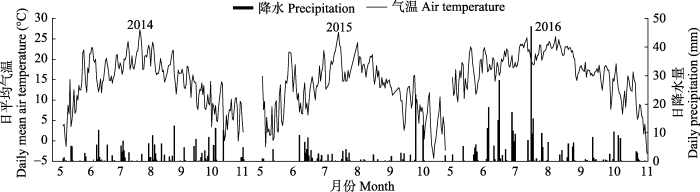 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ12014-2016����������ƽ�����º��ս�ˮ����ÿһ�����ݵ�Ϊ���չ۲�ֵ��
Fig. 1Daily mean air temperature and precipitation in the growing seasons from 2014 to 2016. Each data point is the observation value of the day.
2.2 �����¶���ʪ��
2014-2016���������ڼ�10 cm����������¶���ʪ�ȵĶ�̬�仯�ʵ�����(ͼ2), ��仯����������¶Ⱥͽ�ˮһ�¡������ʶ������¶�û������Ӱ��, ����ͬ�����ʴ����������¶ȵı仯����һ��, �����ֳ�������<��ȷ�����<�ضȷ��������ܽ�ˮӰ������, 2016������ʪ����������2014��2015��(p < 0.000 1, ��1)��2014��2015��10��������ˮ���ﵽ�˷�ֵ, �ֱ���9.54%��9.94%, 2016��7��������ˮ�����Ϊ13.86%�������ʶ�����ʪ��û������Ӱ��, ��ͬ�����ʴ�����, ����ʪ�ȱ���Ϊ������>��ȷ�����>�ضȷ�������ͼ2
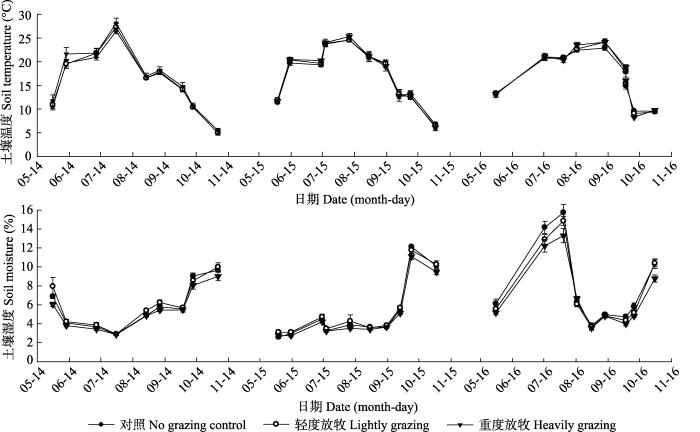 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ22014-2016����������ͬ�����ʴ�����10 cm���������¶Ⱥ�����ʪ�ȡ�ÿһ�����ݵ�Ϊ���ոô�����, �����ظ��۲��ƽ��ֵ��
Fig. 2During the growing season 2014-2016, 10 cm surface soil temperature and moisture under different stocking rates. Each data point is the average of all repeated observations for that day under that treatment.
Table 1
��1
��1�����ʡ���ݼ��佻�����ö���̬ϵͳ̼������Ӱ��
Table 1
| ��̬ϵͳ��̼���� Net ecosystem carbon exchange | ��̬ϵͳ���� Ecosystem respiration | ��̬ϵͳ�ܳ��������� Gross ecosystem productivity | �����¶� Soil temperature | ����ʪ�� Soil moisture | |
|---|---|---|---|---|---|
| ������ Stocking rate | <0.001** | <0.000 1*** | <0.000 1*** | 0.740 | 0.085 |
| �� Year | <0.000 1*** | <0.000 1*** | <0.000 1*** | 0.080 | <0.000 1*** |
| �����ʡ��� Stocking rate �� Year | 0.287 | 0.022* | 0.071 | 0.984 | 0.808 |
�´��ڴ�|����CSV
2.3 ����������̬ϵͳ̼����
�Ӷ����ط����������, �����ʺ����������Ӱ������̬ϵͳ̼����, ���ߵĽ������ý���ER������Ӱ��(p < 0.05, ��1)��2014-2016�������������ڼ�, �̻���é��Į��ԭ��̬ϵͳ���崦�ھ�̼���յ�״̬, 2016����̬ϵͳ��̼������������2014��2015��(p < 0.000 1)����̬ϵͳ��̼�����������ʵ���Ӧ�������, 2014���2016��������������ضȷ���, ��ȷ���û����������; 2015��, ���պ���ȷ������������ضȷ���(ͼ3)����������, �ضȷ���������������������NEE (48.6%)��ER (35.3%)��GEP (40.4%)��NEE�ļ��ڱ仯�����ºͽ�ˮ��Ӱ��, 2014��NEE�ķ�ֵ������8��ĩ9�³�; 2015��NEEΪ˫��������, ��6��ĩ7�³��ﵽ��ֵ, 9��ĩ���ڽ�ˮ�������ٴγ��ַ�ֵ; 2016��NEE���ֳ����Եĵ���������, 7��ĩ8�³��ﵽ��ֵ(ͼ4)��ͼ3
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ32014-2016�겻ͬ����������̬ϵͳ̼�����ı仯(ƽ��ֵ�������)��CK, ����; LG, ��ȷ���; HG, �ضȷ�������д��ĸ��ʾͬ���ڲ�ͬ�����ʲ�������(p < 0.05), Сд��ĸ��ʾͬһ�������ڲ�ͬ��ݼ��������(p < 0.05)����ͬ��ĸ��ʾ����������(p > 0.05)��
Fig. 3Variations of ecosystem carbon exchange in plots with different stocking rates from 2014 to 2016 (mean �� SE). CK, no grazing control; LG, lightly grazing; HG, heavily grazing. Capital letters indicate significant differences (p < 0.05) in sites with different stocking rates in the same year. Lowercase letters indicate significant differences (p < 0.05) in different years at sites with the same rate of stocking. The same letters mean no significant difference (p > 0.05).
ͼ4
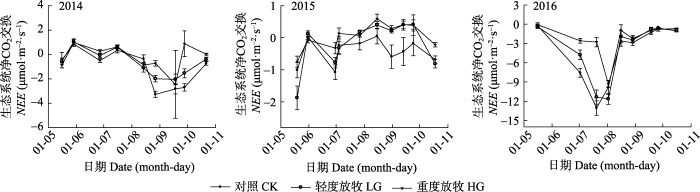 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ42014-2016����̬ϵͳ��̼�����¶�̬(ƽ��ֵ�������)��NEE, ��̬ϵͳ��̼������ÿһ�����ݵ�Ϊ���չ۲��ÿ�������ʴ����Ĺ۲�ֵ��
Fig. 4The monthly dynamic of net ecosystem carbon exchange from 2014 to 2016 (mean �� SE). NEE, net ecosystem carbon exchange. Each data point is the observed value of each stocking rate treatment observed on the day.
2.4 ��ͬ�����ʴ�����GEP��ER��NEE�Ĺ���
3����������, ��ͬ�����ʴ�����, NEE����GEP�����ӳ�������������������(p < 0.000 1)������, GEP��NEE�Ľ��ͳ̶�ΪLG > CK > HG��NEE��ER�Ĺ�ϵ������졣2014��2015�겻ͬ�����ʴ�����, NEE��ERû����ع�ϵ, ֻ����2016�����������������ع�ϵ, ����ER�����Ӿ�̼������������(p < 0.000 1)����ͬ�����ʴ�����, ER��NEE�Ľ��ͳ̶�ΪLG > CK > HG��NEE-GEP֮�������Ը���NEE-ER��NEE��GEP��б������: 2014��, CK 0.688, LG 0.772, HG 0.788; 2015��, CK 0.603, LG 0.684, HG 0.692; 2016��, CK 0.494, LG 0.552, HG 0.531��2014��2015��NEE��GEP��б�ʸ���2016��, ���������ʵ�����, б��������, ��2016������LG > HG > CK��2.5 ˮ�����ض���̬ϵͳ̼������Ӱ��
ˮ������������NEE����Ҫ���������ء�����2014-2016��3���е�ˮ���������ܴ�, ����������̼ͨ���������¶ȡ�����ʪ�ȵĻع�, ���������¶ȡ�����ʪ����CO2ͨ��֮��Ĺ�ϵ��3���������в�ͬ��2014��, NEE�������¶ȳ������Ķ��κ�����ϵ(p < 0.000 1), ���������¶����߾�̼����������, �������¶ȳ���20 ��ʱ, ��̼�������Լ���, �����־�̼�ͷš�GEP�ı仯������NEEһ��, ���������¶ȵ�����, GEP�����Ӻ���١�ER�������¶ȳ������Ķ��κ�����ϵ(p < 0.000 1), ���������¶ȵ�����, ER�����Ӻ���١�����ʪ����CO2ͨ��֮��Ĺ�ϵ�������¶�����, NEE��GEP������ʪ�ȳ������Ķ��κ�����ϵ(p < 0.000 1), ��������ʪ�ȵ�����, ��̼���պ�GEP�����Ӻ���١�ER������ʪ�ȳ������Ķ��κ�����ϵ (p = 0.008), ��������ʪ�ȵ�����, ER�����Ӻ����; 2015��, NEE�������¶ȳ����������Թ�ϵ(p < 0.000 1), ���������¶�����, ��̼���ճ������������ơ�GEP�ı仯������NEEһ��, GEP�������¶ȵ����߶����ӡ�ER�������¶ȳ������Ķ��κ�����ϵ(p = 0.0012), ���������¶ȵ�����, ER�����Ӻ���١�Ȼ��2015�������ʪ����CO2ͨ��֮��û�б��ֳ���������ع�ϵ; 2016��, CO2ͨ���������¶Ⱥ�ʪ�ȵ����ӱ��ֳ���ͬ�ı仯���ơ�NEE��GEP�������¶ȡ�����ʪ�ȳ����������Թ�ϵ(p < 0.000 1), ���������¶Ⱥ�ʪ�ȵ�����, CO2���ճ������������ơ�ER�������¶ȡ�����ʪ�ȳ����������Թ�ϵ(p < 0.000 1), ���������¶Ⱥ�ʪ�ȵ�����, ER�������ӡ��ڼ��ڳ߶���, �����¶ȶ���̬ϵͳ̼ͨ����ֵĽ��ͳ̶�(R2 = 0.02-0.54)��������ʪ��(R2 = 0-0.28)����ͬ��������¶ȡ�ʪ�ȶ�NEE�������Ҫ�Խ������2��ʾ, 2014�������¶Ⱥ�ʪ�ȿ��Խ���NEE�����7.9%, ���������¶ȶ�NEE����Թ���Ϊ5.7%, ����ʪ�ȶ�NEE����Թ���Ϊ2.2%, ���Ƿֱ�ռ���������72.28%��27.72%��2015�������¶Ⱥ�ʪ�ȿ��Խ���NEE�����18.4%, �����¶ȶ�NEE����Թ���Ϊ13.7%, ����ʪ�ȶ�NEE����Թ���Ϊ4.6%, �ֱ�ռ���������74.71%��25.29%��2016�������¶Ⱥ�ʪ�ȿ��Խ���NEE�����38.2%, �����¶ȶ�NEE����Թ���Ϊ20.2%, ����ʪ�ȶ�NEE����Թ���Ϊ18%, �ֱ�ռ���������52.88%��47.12%��3����, ����ƽˮ��(2014��)��Ƿˮ��(2015��)���Ƿ�ˮ��(2016��), �����¶ȶ�NEE�Ĺ����ʾ���������ʪ�ȡ�
Table 2
��2
��2�����¶ȡ�����ʪ�ȶ���̬ϵͳ��̼��������Թ���
Table 2
| ��ģ�� Sub-model | 2014����ⷽ��� Extra variance contribution in 2014 | 2015����ⷽ��� Extra variance contribution in 2015 | 2016����ⷽ��� Extra variance contribution in 2016 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| R2 | ST | SM | R2 | ST | SM | R2 | ST | SM | |
| K = 0 | 0 | 0.075 | 0.041 | 0 | 0.094 | 0.003 | 0 | 0.228 | 0.21 |
| ST | 0.075 | 0.004 | 0.094 | 0.090 | 0.228 | 0.154 | |||
| SM | 0.041 | 0.039 | 0.003 | 0.181 | 0.206 | 0.176 | |||
| K = 1 | 0.039 | 0.004 | 0.181 | 0.090 | 0.176 | 0.154 | |||
| ST, SM | 0.079 | 0.184 | 0.382 | ||||||
| ��ƽ������ Total average contribution | 0.057 | 0.022 | 0.137 | 0.046 | 0.202 | 0.180 | |||
| �ڷ����еı��� Ratio in variance | 72.28% | 27.72% | 74.71% | 25.29% | 52.88% | 47.12% | |||
�´��ڴ�|����CSV
ͼ5
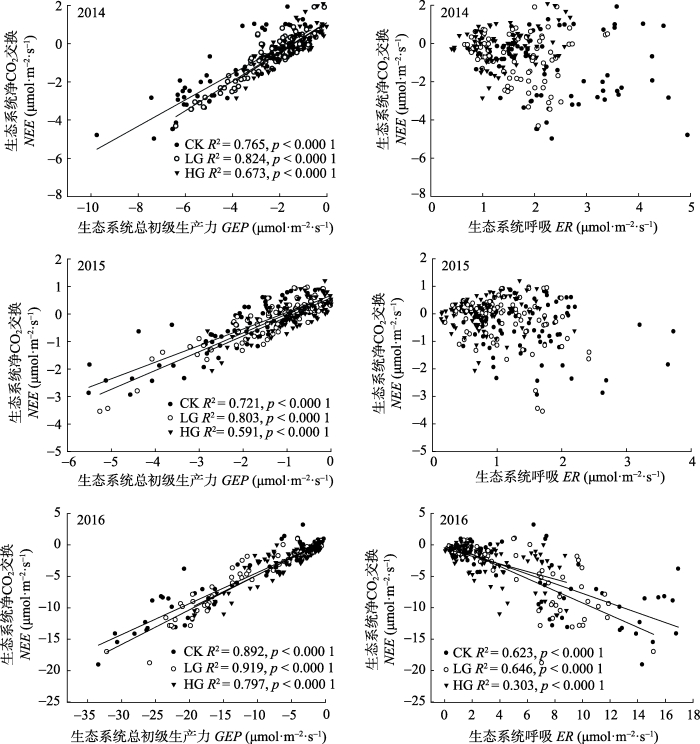 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ52014-2016�겻ͬ�����ʴ�������̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)����̬ϵͳ��̼����(NEE)�Ĺ��ס�CK, ����; LG, ��ȷ���; HG, �ضȷ�����ͼ��ÿһ�����ݵ�Ϊ���չ۲��ÿ�������ʴ�����ÿ���ظ���ֵ��
Fig. 5Relationship between gross primary productivity (GEP) and net ecosystem carbon exchange (NEE) (left panel) and ecosystem respiration (ER) and net ecosystem carbon exchange (NEE) (right panel) in plots with different stocking rates from 2014 to 2016. CK, no grazing control; LG, lightly grazing; HG, heavily grazing. Each data point in the graph is the value of each replicate to each stocking rate observed during the day.
3 ����
3.1 ��̬ϵͳ��̼�����ļ��ڶ�̬
5�·ݶ̻���é��ԭֲ���ոտ�ʼ����, ��ˮ��, ��̬ϵͳ̼���ս���, NEE��Ҫ��ER�����������ۻ��ķ���ЧӦʹ����ȷ������ضȷ�����������Խ�ʵ, ͨ�Խ���, ���³������������ڶ���, ��֮�������ڵĵ�������۽϶�, Ӱ����ֲ���ȶ�, ����Ⱥ��Ĺ�����ýϵ�, ��˳���������������̼�����������ڶ������������6�·ݽ�ˮ������, �ٽ���ֲ��������, ��̬ϵͳ̼����������ǿ��������������ضȷ������������NEE��7��������������ƽ���¶���ߵ��·�, ˮ�ֳ�Ϊ����ֲ��������GEP����Ҫ������, 2014��7�����д���������Ϊ̼Դ, ��2015��7���������Ϊ����̼�㡣2016��7�½�ˮƵ������������, ��ӽ����������¶�, ������ˮ��ԼΪ2014�ꡢ2015���4��, NEE�ﵽȫ�����(-7.03 mol��m-2��s-1)��8�����������ĸ߷���, 2014��8�¾�̼�����ӽ���ֵ, 2015����һ���ɺ������, 8�½�ˮΪ�������������, ����ֲ������������Ч��ˮ��, �ڷ���������, ֲ��ϡ�貢����ݻ�, ����ʪ�ȵ�, ֲ�������֮���̼��������, ������������Ϊ̼Դ��2016��8�²�ͬ�����ʴ�����NEE����Ϊ��̼����, �����岻��7�¡�����������ʢ�ڵ���, ���������¸�����������, �Һ������ʼӿ�, ����ϵ�����������������߶����, �ϸߵĸ�ϵ��������ٽ�CO2���ͷ�(���½ܵ�, 2015)��9���ӽ���������ĩ��, 2014��9�½�ˮ����, ��̼���մﵽ��ֵ, 10��̼���������½�, �ضȷ���������̼�ͷš�2015��9����Ȼ�н�ˮ, ������8�·ݵĸɺ�����ֲ��ϡ������ݻ�, ��˳�������, ��������������Ϊ̼�ͷš�����10��������ˮ���ﵽ��ֵ, ��̼������С���ȵ����ӡ�������������, ��ˮ���ͽ�ˮ�ļ��ڷֲ����꾻̼������Ӱ��ܴ�(Polky et al., 2010; �����, 2018)��ͼ6
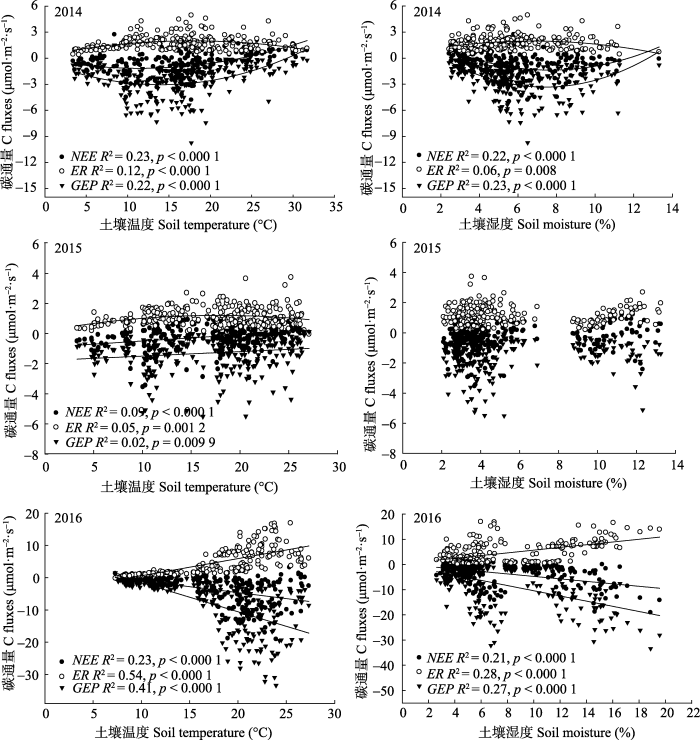 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ62014-2016����̬ϵͳ̼�����������¶ȡ�����ʪ�ȵĹ�ϵ��NEE, ��̬ϵͳ��̼����; ER, ��̬ϵͳ����; GEP, ��̬ϵͳ�ܳ�����������2014�����ָ�����̶�NEE��ER��GEP�������¶ȡ�����ʪ�ȼ�Ĺ�ϵ��ϡ�2015�����ָ�����̶�ER�������¶ȼ��ϵ���, �������Է��̶�NEE��GEP�������¶ȼ��ϵ��ϡ�2016��������Է��̶������¶ȡ�����ʪ�ȼ��ϵ��ϡ�ÿһ�����ݵ�Ϊ���չ۲��ʵ��ֵ��
Fig. 6The relationship between ecosystem carbon exchange and soil temperature and soil moisture from 2014 to 2016. NEE, net ecosystem carbon exchange; ER, ecosystem respiration; GEP, gross primary productivity. In 2014, the exponential model was used to fit the relationship between carbon flux and soil temperature and soil moisture, In 2015, the exponential model was used to fit the relationship between ER and soil temperature, and the linear model was used to fit the relationship between NEE, GEP and soil temperature. In 2016, the linear model was used to fit the relationship between carbon flux and soil temperature and soil moisture. Each data point is the observed value of the day.
�о������ڸɺ��ش�, �ݵص�̼��֧�ܽ�ˮ�������Լ(Kang et al., 2013)��ͨ��3��IJⶨ����, ��ˮ����ʼ���̬ϵͳ̼�������¶�̬��Ӱ��ʮ�����ԡ�2016��CO2ͨ����ֵ��������2014��2015�ꡣ��ˮ������, ������ˮ������, Ⱥ��Ĺ��������ǿ, GEP����, GEP���ӿ���Ϊ��ϵ���������������������ṩ����ĵ���, �ٽ�ER������GEP�漰�Ĺ������ֻ��ֲ���й�, ��ER������ֲ���йػ��ܵ�����������������������Ӱ��, ���, GEP�Խ�ˮ�����жȴ���ER, ��NEE�ı仯���������á� ��ˮ�ǵ��ڶ̻���é��Į��ԭCO2ͨ���仯����Ҫ����, ���Ž�ˮ����, �̻���é��Į��ԭ̼��������ǿ���˽������´���ԭ�Ĺ۲���(Niu et al., 2008; Li et al., 2017)һ��, ��ݵ��ԭ���о����(���S��, 2016)�෴��������Dz�ͬ�IJݵ������������Ӳ�ͬ, GEP��ER������仯����Ӧ��С�ͷ���ͬ���µġ�
��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼�������Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ�����ԡ��о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(Jia et al., 2013; ������, 2016)��2016��, �����¶Ⱥ�ʪ�ȹ�ͬ������NEE�����38.24%, ����2014��2015�ꡣ3������ݾ�����Ϊ�����¶ȶ�NEE�Ľ��ͳ̶ȸ�������ʪ��, �����¶ȶ�NEE�Ĺ����ʸ�������ʪ�ȡ������о����������¶�����ԼNEE����Ҫ����(����Ƶ�, 2000; ��ȫʤ��, 2003; ������, 2014)�����о��Ľ���֧����һ�۵㡣������(2011)�ڿ�����é��ԭ���о�������ֵ������¶ȳ���20 �� ʱ, ������GEP�����ö��ٽ�ER�����á����о���Ϊ, �����¶�����, ֲ����ϵ�ĺ����Լ�����Ļ����, �ٽ���CO2���ͷš��������¶ȹ��ߵ�������ʪ�Ƚ���, ֲ���ܵ�ˮ��в�Ȼ��ȡҶƬ���ر�(��ռ�غ����ڹ�, 1990), ������������, ���ˮ������Ч������Ӧ�ɺ�, Ȼ�������е�ˮ����Ϊ������õ�ԭ��, ˮ�ֿ�ȱֱ��Ӱ��ֲ��Ĺ������, �Ӷ�Ӱ����̬ϵͳ�ľ�̼���ա���Ȼ�����¶Ȼ���Լ��̬ϵͳ��̼����, ���ǵ���������ˮ����ʱ, ֲ������������ˮ�ָ�, �������ͨ�����¶ȵ����߶���ǿ(��������, 2010), ��̬ϵͳ̼�������������¶ȵ����ӳ������������������ơ����, �ڶ̻���é��Į��ԭ, ����Ľ�ˮ������ͻ�������¶ȶ���̬ϵͳ̼��������Լ���á�
3.2 �����ʶ���̬ϵͳ̼������Ӱ��
��ˮ����������ʼ�̻���é��Į��ԭ��̬ϵͳCO2ͨ��������������, �̻���é��Į��ԭ������Ϊ��̼����, ���������ʵ�����, NEE��ER��GEP�������͡��ضȷ������������˲ݵع�̼����, ����ȷ���û������Ӱ�졣�ڼ��ڳ߶���, �����¶���Ӱ����̬ϵͳ��̼������������Ҫ�������ӡ���ˮ�ٽ��������¶ȡ�����ʪ����̼ͨ����ֵ�����ԡ���л ���ɹ�ũҵ��ѧ���������ѧ����(2014XYQ-7)�������о�, �ش���л��

�ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
DOI:10.1007/s40333-014-0049-xURL [��������: 1]

Grazing can modulate the feedback between vegetation and soil nutrient dynamics(carbon and nitrogen),altering the cycles of these elements in grassland ecosystems.For clarifying the impact of grazing on the C and N in plants and soils in the desert grassland of Ningxia,China,we examined the plant biomass,SOC(soil organic carbon),total soil N and stable isotope signatures of plants and soils from both the grazed and ungrazed sites.Significantly lower aboveground biomass,root biomass,litter biomass and vegetation coverage were found in the grazed site compared to the ungrazed site,with decreases of 42.0%,16.2%,59.4% and 30.0%,respectively.The effects of grazing on plant carbon,nitrogen,?15N and ?13C values were uniform among species.The levels of plant carbon and nitrogen in grasses were greater than those in the forbs(except for the carbon of Cynanchum komarovii and Euphorbia esula).Root 15 N and 13 C values increased with grazing,while the responses of root carbon and nitrogen to grazing showed no consistent patterns.Root 15 N and 13 C were increased by 79.0% and 22.4% in the grazed site compared to the ungrazed site,respectively.The values of SOC and total N were significantly lower in the grazed than in the ungrazed sites for all sampling depths(0�C10 and 10�C20 cm),and values of SOC and total N at the surface(0�C10 cm) were lower than those in the deeper soils(10�C20 cm).Soil ?15N values were not affected by grazing at any sampling depth,whereas soil ?13C values were significantly affected by grazing and increased by 19.3% and 8.6% in the soils at 0�C10 and 10�C20 cm,respectively.The soil ?13C values(�C8.3�� to �C6.7��) were higher than those for roots(�C20.2�� to �C15.6��) and plant tissues(�C27.9�� to �C13.3��).Our study suggests that grazing could greatly affect soil organic carbon and nitrogen in contrast to ungrazed grassland and that grazing appears to exert a negative effect on soil carbon and nitrogen in desert grassland.
DOI:10.1046/j.1365-2435.2001.00517.xURL
Abstract 1. Increases in photosynthesis of leaves remaining after defoliation may result from shifts in leaf photosynthetic characteristics or from an improved light penetration in the canopy. However, few studies have tried to estimate the relative contribution of these factors to mitigating the negative effects of defoliation. We present a quantitative framework for such an analysis. 2. In a field and greenhouse experiment, plants of the rain forest understorey palm Chamaedorea elegans were subjected to three levels of defoliation (0, 50 or 66% of leaves removed) and, in the greenhouse, grown at two irradiances (5 or 16% of daylight, 'low light' and 'high light' hereafter). For each plant, leaf photosynthesis and light penetration in the canopy were measured and these data were used to calculate whole-canopy carbon gain. 3. Defoliation significantly increased the light available to the remaining leaves. In the field and in the 'high-light' greenhouse plants, defoliation also resulted in an increase in the light-saturated photosynthesis per unit leaf area ( P max ), although in the 'low-light' plants this was not the case. 4. A sensitivity analysis revealed that in the 'low-light' plants, a proportional increase in leaf P max did not result in increased canopy-level carbon gain, while in the 'high-light' and field plants, it did. This suggests that the lack of plasticity in P max in response to defoliation, at low light, may be the appropriate pattern to maximize carbon gain. 5. Defoliated plants had 10 18% more average photosynthesis per unit leaf area ( P area ) than the control plants, and this increase was mostly the result of an improved light penetration in the canopy. 6. A relatively small fraction (5 30%) of the total estimated loss in canopy carbon gain caused by the removal of leaves was compensated for by an increase in P area . This suggests that in rain forest understorey plants, an increase in diurnal photosynthesis of remaining leaves has limited potential for mitigating the negative effects of defoliation.
DOI:10.17521/cjpe.2003.0031URL [��������: 1]
���ü�Һ���շ������ֺ������´����Ͳ�ԭһ�˻�Ⱥ����������������˲ⶨ ,���������¶Ⱥ�ˮ�ֶ�����������Ӱ�� ,������� :1 )�������������������ļ��� ,�������ڵ� ,�����ڶ�̬���ֲ����ɵIJ������� ;2 )���¡��ر��¶��Լ� 5cm��1 0cm��1 5cm�� 2 5cm�������¶Ⱦ��������������ʳ�������ָ����ϵ ,�¶ȶ�����������Ӱ���ڵ���ʱ�ȸ���ʱ������ ;3) 0�� 1 0cm�� 1 0�� 2 0cm�����������ˮ�����������������ʳ����������Թ�ϵ ,�������µ�Ӱ�����ʸ�Ϊ�����ij��ݹ�ϵ ;4)���ݱ�����p =0 .0 5ˮƽ�ϵĶ�Ԫ�ع��������õ������������������º� 1 0�� 2 0cm������ˮ���Ĺ�ϵģ�� :y =5 91 1 .648��e0 .0 42 1 6Ta��M20 .90 758(R2 =0 .8584,p 0 .0 0 0 1 ) ,��һģ�ͱȵ�����ģ���ܸ��õؽ������������ı仯��� ;5)ʵ���ڼ�����������ƽ������Ϊ 661 .35mgC��m-2 ��d-1 ,�����¡��ر��¶��Լ� 5cm��1 0cm��1 5cm�� 2 5cm�������¶�Ϊ���ݵõ���Q1 0 ֵ����Ϊ 1 .63��1 .47��1 .52��1 .70��1 .90��1 .97��
DOI:10.17521/cjpe.2003.0031URL [��������: 1]
���ü�Һ���շ������ֺ������´����Ͳ�ԭһ�˻�Ⱥ����������������˲ⶨ ,���������¶Ⱥ�ˮ�ֶ�����������Ӱ�� ,������� :1 )�������������������ļ��� ,�������ڵ� ,�����ڶ�̬���ֲ����ɵIJ������� ;2 )���¡��ر��¶��Լ� 5cm��1 0cm��1 5cm�� 2 5cm�������¶Ⱦ��������������ʳ�������ָ����ϵ ,�¶ȶ�����������Ӱ���ڵ���ʱ�ȸ���ʱ������ ;3) 0�� 1 0cm�� 1 0�� 2 0cm�����������ˮ�����������������ʳ����������Թ�ϵ ,�������µ�Ӱ�����ʸ�Ϊ�����ij��ݹ�ϵ ;4)���ݱ�����p =0 .0 5ˮƽ�ϵĶ�Ԫ�ع��������õ������������������º� 1 0�� 2 0cm������ˮ���Ĺ�ϵģ�� :y =5 91 1 .648��e0 .0 42 1 6Ta��M20 .90 758(R2 =0 .8584,p 0 .0 0 0 1 ) ,��һģ�ͱȵ�����ģ���ܸ��õؽ������������ı仯��� ;5)ʵ���ڼ�����������ƽ������Ϊ 661 .35mgC��m-2 ��d-1 ,�����¡��ر��¶��Լ� 5cm��1 0cm��1 5cm�� 2 5cm�������¶�Ϊ���ݵõ���Q1 0 ֵ����Ϊ 1 .63��1 .47��1 .52��1 .70��1 .90��1 .97��
URL [��������: 1]
½����̬ϵͳ��������̼ˮ���������ʺ�����ѭ���Ĺؼ����̣���չֲ���������̼��ˮͨ���ij��ڹ۲�����������о�������ȫ��仯��½����̬ϵͳ̼ˮѭ�������ϵ������ƻ���������Ҫ�����塣���о����й��������۲ݵ��ԭΪ�о��������ж���ط���Ϊ��Ҫ�����ֶΣ��Գ���Ұ��ʵ��վ�����̼��ˮͨ������������Ϊ���У����������ж���ؼ����ڲݵ��ԭ̼��ˮѭ���о��е������ԣ������ͨ�����ݵ��������ۺͿ��ƣ�����̽���˸òݵ���̬ϵͳ��̼������ˮ����ɢ�ڲ�ͬʱ��߶��ϵı仯���������������ƣ��Աȷ�����������ˮ��ƽ��������֧��...
URL [��������: 1]
½����̬ϵͳ��������̼ˮ���������ʺ�����ѭ���Ĺؼ����̣���չֲ���������̼��ˮͨ���ij��ڹ۲�����������о�������ȫ��仯��½����̬ϵͳ̼ˮѭ�������ϵ������ƻ���������Ҫ�����塣���о����й��������۲ݵ��ԭΪ�о��������ж���ط���Ϊ��Ҫ�����ֶΣ��Գ���Ұ��ʵ��վ�����̼��ˮͨ������������Ϊ���У����������ж���ؼ����ڲݵ��ԭ̼��ˮѭ���о��е������ԣ������ͨ�����ݵ��������ۺͿ��ƣ�����̽���˸òݵ���̬ϵͳ��̼������ˮ����ɢ�ڲ�ͬʱ��߶��ϵı仯���������������ƣ��Աȷ�����������ˮ��ƽ��������֧��...
[��������: 1]
[��������: 1]
URL [��������: 1]
Terrestrial grassland ecosystems are an important component of the global carbon (C) cycle. We used the Bowen ratio/energy balance technique to measure CO2 fluxes and evapotranspiration (ET) over a mid-grass prairie at Mandan, ND; a mixed-grass prairie at Woodward, OK; and a tallgrass prairie at Temple, TX representing three diverse grassland ecosystems along a latitudinal gradient of 31N to 46N in the Great Plains of the USA. Measurements from early spring through late autumn 1995 through 1997 were used to estimate daily, seasonal, and annual net CO2 uptake. Annual net CO2 uptake, calculated using estimates of CO2 efflux during winter, averaged 348, 336, and 1822 g/m2/y at Mandan, Woodward, and Temple, respectively. These fluxes are equivalent to 946, 914, and 4956 kg C/ha/y being stored annually at Mandan, Woodward, and Temple, respectively. The greater net CO2 fixation at Temple resulted from effects of burning the esite in February 1995, a longer season of active CO2 fixation, and greater precipitation. Carbon dioxide fluxes were frequently reduced by periods of below normal precipitation, but all sites sequestered C. Peak above ground and root biomass were 1028 and 15630 kg/ha for Mandan, 3234 and 16463 kg/ ha for Woodward, and 3834 and 19352 kg/ha for Temple. Average peak LAI was 0.5, 1.2, and 2.7 for Mandan, Woodward, and Temple, respectively. Daily ET rates during the period of measurement averaged 2.6, 2.3, and 3.1 mm/d at Mandan, Woodward, and Temple, respectively. These results suggest that these grasslands, which are typical of grassland ecosystems that cover millions of hectares in the Great Plains, are potential sinks for CO2.
DOI:10.3321/j.issn:1000-0933.2007.01.021URL [��������: 1]
���ò�ͬ�����ʵ�Χ����������,�о������ɹŸ�ԭ��Į��ԭ�Ǵ��̻���é(Stipa breviflora Griseb.)��ԭȺ���ڲ�ͬ������ˮƽֲ������Ա仯���ɺͶԲݵ���������Ӱ��.�о��������:��2a�ķ���������,��ͬ������ֲ�������ָ���ľ�ֵ ����ȵ������н��͵�����,����ȼ����δ�ﵽ����ˮƽ;��ͬ�����, ֲ�������ָ������������1.027 ֻ��/(hm2��a)�������ַ�ֵ;��������Ϊ1.027ֻ��/(hm2��a)ʱֲ�ﲹ�����������,���������������ˮƽ.�о����֧��Ⱥ�����ֶ����� ����������ظ���еĸ����κ�����ϵ�ĵ�����ģ��,���е�������ˮƽ���ֶ��������.
DOI:10.3321/j.issn:1000-0933.2007.01.021URL [��������: 1]
���ò�ͬ�����ʵ�Χ����������,�о������ɹŸ�ԭ��Į��ԭ�Ǵ��̻���é(Stipa breviflora Griseb.)��ԭȺ���ڲ�ͬ������ˮƽֲ������Ա仯���ɺͶԲݵ���������Ӱ��.�о��������:��2a�ķ���������,��ͬ������ֲ�������ָ���ľ�ֵ ����ȵ������н��͵�����,����ȼ����δ�ﵽ����ˮƽ;��ͬ�����, ֲ�������ָ������������1.027 ֻ��/(hm2��a)�������ַ�ֵ;��������Ϊ1.027ֻ��/(hm2��a)ʱֲ�ﲹ�����������,���������������ˮƽ.�о����֧��Ⱥ�����ֶ����� ����������ظ���еĸ����κ�����ϵ�ĵ�����ģ��,���е�������ˮƽ���ֶ��������.
DOI:10.5846/stxb201603110430URL
��ȷ�����½��ݵ���̬ϵͳ̼Դ/��ЧӦ��������߶�̼ѭ���о�������Ҫ���塣�������½��ݵ���̬ϵͳ����Ҫ���������������Բݵ�̼ƽ����̼��̬��Ӱ�컹���кܴ�IJ�ȷ���ԡ�������̬ϵͳ����ģ��Biome-BGC grazing��ͨ���龰ģ���ۺ������½��ݵ���̬ϵͳ̼Դ/��Ķ�̬�����������1�� 1979-2007���½��ݵ���̬ϵͳ��̼Դ����Ϊ0.38PgC�������ɷ������µ�̼�ͷ�Ϊ0.37PgC��2�� ��ƽ��������С��0.24ͷ����/hm^2ʱ�������ܹ��ٽ��ݵ�̼�̶����о�ʵ����Biome-BGC grazingģ��������߶ȵ�Ӧ�ã��о��������������������仯�������Ըɺ����ݵ���̬ϵͳ̼��̬�仯��������������̽���ɺ�����ԭ��̬ϵͳ��Դ/������������Ҫ���塣
DOI:10.5846/stxb201603110430URL
��ȷ�����½��ݵ���̬ϵͳ̼Դ/��ЧӦ��������߶�̼ѭ���о�������Ҫ���塣�������½��ݵ���̬ϵͳ����Ҫ���������������Բݵ�̼ƽ����̼��̬��Ӱ�컹���кܴ�IJ�ȷ���ԡ�������̬ϵͳ����ģ��Biome-BGC grazing��ͨ���龰ģ���ۺ������½��ݵ���̬ϵͳ̼Դ/��Ķ�̬�����������1�� 1979-2007���½��ݵ���̬ϵͳ��̼Դ����Ϊ0.38PgC�������ɷ������µ�̼�ͷ�Ϊ0.37PgC��2�� ��ƽ��������С��0.24ͷ����/hm^2ʱ�������ܹ��ٽ��ݵ�̼�̶����о�ʵ����Biome-BGC grazingģ��������߶ȵ�Ӧ�ã��о��������������������仯�������Ըɺ����ݵ���̬ϵͳ̼��̬�仯��������������̽���ɺ�����ԭ��̬ϵͳ��Դ/������������Ҫ���塣
URL [��������: 2]
�ݱ���-������CO2��������Ի������ӵ���Ӧ��Ŀǰ�о����ȵ㡣���о�ͨ����̬�䷨������Li-840 CO2/H2O��������ǣ����½���ɽ���²ݵ��ԭΧ��9������غ�Χ�������̬ϵͳ̼�������м�⣬����Χ��������̬ϵͳ̼�����IJ����ԡ��ձ仯�����ڱ仯�����뻷�����ӵĹ�ϵ�������������̬ϵͳ̼����Χ���ڸ���Χ���⣬���ֳ����ԵIJ����ԣ�P<0.05����Χ��������̬ϵͳ̼�������������Ե��ձ仯�ͼ��ڱ仯���ɣ��ʵ������ߣ�����ֲ�����������αȽ����ԡ�����������ڼ䣬Χ��������̬ϵͳ��̼������net ecosystem CO2 exchange, NEE�����ֵ�ֱ�Ϊ-7.62 ��mol?m-2?s-1��-6.63 ��mol?m-2?s-1����̬ϵͳ������ecosystem respiration , ER�����ֵ�ֱ�Ϊ8.55 ��mol?m-2?s-1��7.04��mol?m-2?s-1����̬ϵͳ�ܳ�����������gross ecosystem productivity, GEP�����ֵ�ֱ�Ϊ-14.66 ��mol?m-2?s-1��-13.89 ��mol?m-2?s-1����Χ����ֲ���õ�������ֲ������ïʢ���������ǿ��NEE��С��ͬʱ�л�̼���������ǿ����̬ϵͳ�������������֣���̬ϵͳ̼���������º�10 cm���³�������أ������µ�����Ը���10 cm���£���Χ��������Ժ���Χ���⣻��ˮ������̬ϵͳ̼��������һ��������ԣ���������ʪ���������С�������¶ȡ�
URL [��������: 2]
�ݱ���-������CO2��������Ի������ӵ���Ӧ��Ŀǰ�о����ȵ㡣���о�ͨ����̬�䷨������Li-840 CO2/H2O��������ǣ����½���ɽ���²ݵ��ԭΧ��9������غ�Χ�������̬ϵͳ̼�������м�⣬����Χ��������̬ϵͳ̼�����IJ����ԡ��ձ仯�����ڱ仯�����뻷�����ӵĹ�ϵ�������������̬ϵͳ̼����Χ���ڸ���Χ���⣬���ֳ����ԵIJ����ԣ�P<0.05����Χ��������̬ϵͳ̼�������������Ե��ձ仯�ͼ��ڱ仯���ɣ��ʵ������ߣ�����ֲ�����������αȽ����ԡ�����������ڼ䣬Χ��������̬ϵͳ��̼������net ecosystem CO2 exchange, NEE�����ֵ�ֱ�Ϊ-7.62 ��mol?m-2?s-1��-6.63 ��mol?m-2?s-1����̬ϵͳ������ecosystem respiration , ER�����ֵ�ֱ�Ϊ8.55 ��mol?m-2?s-1��7.04��mol?m-2?s-1����̬ϵͳ�ܳ�����������gross ecosystem productivity, GEP�����ֵ�ֱ�Ϊ-14.66 ��mol?m-2?s-1��-13.89 ��mol?m-2?s-1����Χ����ֲ���õ�������ֲ������ïʢ���������ǿ��NEE��С��ͬʱ�л�̼���������ǿ����̬ϵͳ�������������֣���̬ϵͳ̼���������º�10 cm���³�������أ������µ�����Ը���10 cm���£���Χ��������Ժ���Χ���⣻��ˮ������̬ϵͳ̼��������һ��������ԣ���������ʪ���������С�������¶ȡ�
DOI:10.17521/cjpe.2006.0113URL [��������: 1]
�����ж���ط���2005�����������ɹ����ֺ��������(Leymus chinensis)��ԭ����̬ϵͳ����(Net ecosystem exchange, NEE)�����˹۲�.�۲�������:��Ϊ����������������126 mm�ĸɺ���,���ֺ�������ݲ�ԭ��̬ϵͳ�ܵ�ǿ�ҵĸɺ�в��,�侻��̬ϵͳ̼�������ն�̬����Ϊ�����������ո߷�,�����շ�ֵ������8��00�� 18��00����.����CO2������Ϊ-0.38 mg CO2��m-2��s-1,������6�µ�,���ˮ�������̬ϵͳ���CO2�������½���1��.����������������,�����ǰ��컹������2005�궼����Ϊ�� CO2�ŷ�,����������CO2���ŷ���Ϊ372.56 g CO2��m-2,��һ�����Ե�CO2Դ.������ˮ���������¶ȿ�������̬ϵͳCO2ͨ���Ĵ�С,�������ڰ���,CO2ͨ����������ˮ���ı仯���ֳ������� ����ع�ϵ,�������¶ȱ���Ϊ����ع�ϵ.
DOI:10.17521/cjpe.2006.0113URL [��������: 1]
�����ж���ط���2005�����������ɹ����ֺ��������(Leymus chinensis)��ԭ����̬ϵͳ����(Net ecosystem exchange, NEE)�����˹۲�.�۲�������:��Ϊ����������������126 mm�ĸɺ���,���ֺ�������ݲ�ԭ��̬ϵͳ�ܵ�ǿ�ҵĸɺ�в��,�侻��̬ϵͳ̼�������ն�̬����Ϊ�����������ո߷�,�����շ�ֵ������8��00�� 18��00����.����CO2������Ϊ-0.38 mg CO2��m-2��s-1,������6�µ�,���ˮ�������̬ϵͳ���CO2�������½���1��.����������������,�����ǰ��컹������2005�궼����Ϊ�� CO2�ŷ�,����������CO2���ŷ���Ϊ372.56 g CO2��m-2,��һ�����Ե�CO2Դ.������ˮ���������¶ȿ�������̬ϵͳCO2ͨ���Ĵ�С,�������ڰ���,CO2ͨ����������ˮ���ı仯���ֳ������� ����ع�ϵ,�������¶ȱ���Ϊ����ع�ϵ.
DOI:10.1371/journal. pone. 0057858URLPMID:358512851472782026862081134 [��������: 1]
Abstract Although the temperature response of soil respiration (Rs ) has been studied extensively, several issues remain unresolved, including hysteresis in the Rs -temperature relationship and differences in the long- vs. short-term Rs sensitivity to temperature. Progress on these issues will contribute to reduced uncertainties in carbon cycle modeling. We monitored soil CO2 efflux with an automated chamber system in a Pinus tabulaeformis plantation near Beijing throughout 2011. Soil temperature at 10-cm depth (Ts ) exerted a strong control over Rs , with the annual temperature sensitivity (Q10) and basal rate at 10 C (Rs10) being 2.76 and 1.40 mol m(-2) s(-1), respectively. Both Rs and short-term (i.e., daily) estimates of Rs10 showed pronounced seasonal hysteresis with respect to Ts , with the efflux in the second half of the year being larger than that early in the season for a given temperature. The hysteresis may be associated with the confounding effects of microbial population dynamics and/or litter input. As a result, all of the applied regression models failed to yield unbiased estimates of Rs over the entire annual cycle. Lags between Rs and Ts were observed at the diel scale in the early and late growing season, but not in summer. The seasonality in these lags may be due to the use of a single Ts measurement depth, which failed to represent seasonal changes in the depth of CO2 production. Daily estimates of Q10 averaged 2.04, smaller than the value obtained from the seasonal relationship. In addition, daily Q10 decreased with increasing Ts , which may contribute feedback to the climate system under global warming scenarios. The use of a fixed, universal Q10 is considered adequate when modeling annual carbon budgets across large spatial extents. In contrast, a seasonally-varying, environmentally-controlled Q10 should be used when short-term accuracy is required.
DOI:10.1007/s11368-013-0675-5URL [��������: 2]
AbstractPurposeCarbon (C) dynamics in grassland ecosystem contributes to regional and global fluxes in carbon dioxide (CO) concentrations. Grazing is one of the main structuring factors in grassland, but the impact of grazing on the C budget is still under debate. In this study, in situ net ecosystem CO exchange (NEE) observations by the eddy covariance technique were integrated with a modified process-oriented biogeochemistry model (denitrification�Cdecomposition) to investigate the impacts of grazing on the long-term C budget of semiarid grasslands.Materials and methodsNEE measurements were conducted in two adjacent grassland sites, non-grazing (NG) and moderate grazing (MG), during 2006�C2007. We then used daily weather data for 1978�C2007 in conjunction with soil properties and grazing scenarios as model inputs to simulate grassland productivity and C dynamics. The observed and simulated CO fluxes under moderate grazing intensity were compared with those without grazing.Results and discussionNEE data from 2-year observations showed that moderate grazing significantly decreased grassland ecosystem CO release and shifted the ecosystem from a negative CO balance (releasing 34.0002g C65m) at the NG site to a positive CO balance (absorbing 6143.0202g C65m) at the MG site. Supporting our experimental findings, the 30-year simulation also showed that moderate grazing significantly enhances the CO uptake potential of the targeted grassland, shifting the ecosystem from a negative CO balance (57.0865��6516.4502g C65m65year) without grazing to a positive CO balance (6128.5865��6514.6002g C65m65year) under moderate grazing. The positive effects of grazing on CO balance could primarily be attributed to an increase in productivity combined with a significant decrease of soil heterotrophic respiration and total ecosystem respiration.ConclusionsWe conclude that moderate grazing prevails over no-management practices in maintaining CO balance in semiarid grasslands, moderating and mitigating the negative effects of global climate change on the CO balance in grassland ecosystems.
DOI:10.1111/j.1365-2486.2010.02179.xURL [��������: 1]
Abstract CO 2 efflux from soil depends on the availability of organic substances respired by roots and microorganisms. Therefore, photosynthetic activity supplying carbohydrates from leaves to roots and rhizosphere is a key driver of soil CO 2 . This fact has been overlooked in most soil CO 2 studies because temperature variations are highly correlated with solar radiation and mask the direct effect of photosynthesis on substrate availability in soil. This review highlights the importance of photosynthesis for rhizosphere processes and evaluates the time lag between carbon (C) assimilation and CO 2 release from soil. Mechanisms and processes contributing to the lag were evaluated. We compared the advantages and shortcomings of four main approaches used to estimate this time lag: (1) interruption of assimilate flow from leaves into the roots and rhizosphere, and analysis of the decrease of CO 2 efflux from soil, (2) time series analysis (TSA) of CO 2 fluxes from soil and photosynthesis proxies, (3) analysis of natural �� 13 C variation in CO 2 with photosynthesis-related parameters or �� 13 C in the phloem and leaves, and (4) pulse labeling of plants in artificial 14 CO 2 or 13 CO 2 atmosphere with subsequent tracing of 14 C or 13 C in CO 2 efflux from soil. We concluded that pulse labeling is the most advantageous approach. It allows clear evaluation not only of the time lag, but also of the label dynamics in soil CO 2 , and helps estimate the mean residence time of recently assimilated C in various above- and belowground C pools. The impossibility of tracing the phloem pressure�Cconcentration waves by labeling approach may be overcome by its combination with approaches based on TSA of CO 2 fluxes and its �� 13 C with photosynthesis proxies. Numerous studies showed that the time lag for grasses is about 12.5��7.5 (SD)h. The time lag for mature trees was much longer (654�C5 days). Tree height slightly affected the lag, with increasing delay of 0.1daym 611 . By evaluating bottle-neck processes responsible for the time lag, we conclude that, for trees, the transport of assimilates in phloem is the rate-limiting step. However, it was not possible to predict the lag based on the phloem transport rates reported in the literature. We conclude that studies of CO 2 fluxes from soil, especially in ecosystems with a high contribution of root-derived CO 2 , should consider photosynthesis as one of the main drivers of C fluxes. This calls for incorporating photosynthesis in soil C turnover models.
DOI:10.11829\j.issn.1001-0629.2013-0528URL [��������: 1]
��������ظ�ԭ��Ҫ���������÷�ʽ֮һ������ǿ�Ȳ�ͬ,����ظ�ԭ�ߺ��ݵ���̬ϵͳNEE��Ӱ��Ҳ��ͬ��Ϊ��ʾ��ͬ����ǿ���¸ߺ��ݵ���̬ϵͳ���彻���ı仯����,ͬʱΪ��̬ϵͳ̼��֧�����ṩ��������,���о���2012��5-10�²���Li-6400��Яʽ����Ǻ��ܱ�ʽ�䷨,����ظ�ԭ�ߺ��ݵ���̬ϵͳ��ͬ����ǿ���µ����彻�����з����ⶨ���������,1)����̬ϵͳ������(NEE)����̬ϵͳ������(Reco)����̬ϵͳ����������(GPP)�����ֳ����Եļ��ڱ仯,NEE�������������ı仯���Ƴ�"U"��,Reco��GPPΪ�����ͱ仯����;2)��5����Ѯ��9����Ѯ,NEEΪ��ֵ,�����ߺ��ݵ���̬ϵͳΪCO2������;3)�ڷ�����Ϊ50%���жȷ���ǿ����,�ߺ��ݵ��NEE��GPP�������ֵ,�����жȷ�����̬ϵͳ���нϸߵ�̼��ˮƽ��
DOI:10.11829\j.issn.1001-0629.2013-0528URL [��������: 1]
��������ظ�ԭ��Ҫ���������÷�ʽ֮һ������ǿ�Ȳ�ͬ,����ظ�ԭ�ߺ��ݵ���̬ϵͳNEE��Ӱ��Ҳ��ͬ��Ϊ��ʾ��ͬ����ǿ���¸ߺ��ݵ���̬ϵͳ���彻���ı仯����,ͬʱΪ��̬ϵͳ̼��֧�����ṩ��������,���о���2012��5-10�²���Li-6400��Яʽ����Ǻ��ܱ�ʽ�䷨,����ظ�ԭ�ߺ��ݵ���̬ϵͳ��ͬ����ǿ���µ����彻�����з����ⶨ���������,1)����̬ϵͳ������(NEE)����̬ϵͳ������(Reco)����̬ϵͳ����������(GPP)�����ֳ����Եļ��ڱ仯,NEE�������������ı仯���Ƴ�"U"��,Reco��GPPΪ�����ͱ仯����;2)��5����Ѯ��9����Ѯ,NEEΪ��ֵ,�����ߺ��ݵ���̬ϵͳΪCO2������;3)�ڷ�����Ϊ50%���жȷ���ǿ����,�ߺ��ݵ��NEE��GPP�������ֵ,�����жȷ�����̬ϵͳ���нϸߵ�̼��ˮƽ��
DOI:10.1016/j.agrformet.2016.09.004URL [��������: 1]
The balance between ecosystem carbon dioxide (CO 2 ) uptake and release determines the level of carbon (C) sequestration in terrestrial ecosystems and its potential impact on CO 2 concentration in the atmosphere. However, how changes in temperature and precipitation will affect the relationships of net ecosystem productivity (NEP) with gross primary productivity (GPP) and ecosystem respiration (ER) remains unclear. In this study, a nine-year field manipulative experiment was conducted with elevated temperature and increased precipitation in a semiarid steppe of Inner Mongolia, China. Experimental warming reduced GPP and ER by almost the same amount, leading to a slight change in NEP (610.1602��mol02m 612 02s 611 ), whereas increased precipitation stimulated GPP more than ER during the growing seasons, resulting in an enhanced NEP (+0.6302��mol02m 612 02s 611 ). In addition, seasonal patterns of ecosystem C fluxes and the NEP-GPP or NEP-ER relationships were not altered by experimental warming. However, increased precipitation delayed the peak of GPP during the growing seasons and enhanced the correlation between NEP and GPP in the steppe ecosystem. The enhanced control of GPP over NEP under the increased precipitation suggests that ecosystem C sequestration is attributed more to C uptake than C release when water availability is improved in the semiarid grassland. Our findings provide an insight into the response mechanism of ecosystem C flux to warming and precipitation change in semiarid grasslands, and facilitate the projection of terrestrial ecosystem C dynamics and climate feedbacks in the future.
DOI:10.1088/0256-307X/17/9/008URL [��������: 1]
���ü�Һ���շ������ɹ����ֺ�������� (L eymus chinensis)��ԭȺ�������������ʽ��������� 2���Ұ��ⶨ ,��ͨ����Ԫ�ع��ֶζ���Ӱ�����ӽ����˷������������ :1) 1998�� 1999���������������Ⱥ�������������ʵļ��ڶ�̬��Ϊ���������� ,���ֵ������ 7����Ѯ ;2 ) CO2 �ͷ����ʡ����º�������ˮ��֮�������¹�ϵ :L n Res=5 .85 96 + 0 .0 12 5 M+ 0 .0 394T+ 0 .0 0 49M�� T,R2 =0 .70 96 ,����Լ�� 70 %�������������仯�������º�������ˮ����ͬ������ ,�������¶�Ϊ�� (Res=2 5 6 .75 T0 .6 5 5 7,R2 =0 .5 779) ,Q1 0 �� 2 .0�� 3.0֮�� ;3)�������Ⱥ������������������������ 2 49.4�� 32 0 .7g C.m- 2 .a- 1֮�� ,����Ⱥ���� 2 37.0�� 30 5 .6 g C.m- 2 .a- 1֮�� ,��������������Ӱ�첻��
DOI:10.1088/0256-307X/17/9/008URL [��������: 1]
���ü�Һ���շ������ɹ����ֺ�������� (L eymus chinensis)��ԭȺ�������������ʽ��������� 2���Ұ��ⶨ ,��ͨ����Ԫ�ع��ֶζ���Ӱ�����ӽ����˷������������ :1) 1998�� 1999���������������Ⱥ�������������ʵļ��ڶ�̬��Ϊ���������� ,���ֵ������ 7����Ѯ ;2 ) CO2 �ͷ����ʡ����º�������ˮ��֮�������¹�ϵ :L n Res=5 .85 96 + 0 .0 12 5 M+ 0 .0 394T+ 0 .0 0 49M�� T,R2 =0 .70 96 ,����Լ�� 70 %�������������仯�������º�������ˮ����ͬ������ ,�������¶�Ϊ�� (Res=2 5 6 .75 T0 .6 5 5 7,R2 =0 .5 779) ,Q1 0 �� 2 .0�� 3.0֮�� ;3)�������Ⱥ������������������������ 2 49.4�� 32 0 .7g C.m- 2 .a- 1֮�� ,����Ⱥ���� 2 37.0�� 30 5 .6 g C.m- 2 .a- 1֮�� ,��������������Ӱ�첻��
[��������: 1]
[��������: 1]
DOI:10.1016/j.agrformet.2007.07.008URL [��������: 1]
To understand the dynamics of ecosystem carbon cycling, CO 2 fluxes were measured over and under an oak�Cgrass savanna and over a proximate grassland in California. The measurements were made from 2000 to 2006 using the eddy covariance technique. Annual net carbon exchange (NEE) ranged from 61155 to 615602gC02m 612 02year 611 and from 6188 to 14102gC02m 612 02year 611 at the savanna and nearby grassland, respectively. Inter-annual variability in NEE was significantly related to length of growing season for the savanna, grassland, and tree canopy. We partitioned the NEE into two separate terms��primary productivity (GPP) and respiration (ecosystem respiration, R eco )��and found that the GPP and R eco of the savanna and grassland depended primarily on the amount of seasonal precipitation that occurred while grass and tree canopies were simultaneously active rather than depending on annual precipitation. We also found that NEE was greatly constrained by both R eco and GPP and R eco was significantly constrained by GPP. R eco increased by 7902gC02m 612 02year 611 with each 10002gC02m 612 02year 611 increase in GPP. In addition, measuring CO 2 exchange separately in the overstory and understory of the savanna over multiple years enabled us to partition R eco into heterotrophic and autotrophic respiration terms in a new and direct way. The sensitivity of R eco to GPP (d R eco /dGPP) and the baseline of respiration terms both provide useful tools for understanding the dynamics of ecosystem CO 2 uptake under current conditions of climate and ecosystem succession stage.
DOI:10.1016/j.agee.2011.06.002URL [��������: 1]
Grasslands cover about 40% of China's land area. This paper synthesizes 133 papers from China on the impacts of land use conversion and improved management practices on soil organic carbon (SOC) in China's grasslands. The synthesis finds that overgrazing and conversion of freely grazed grassland to cropland lead to an annual average decline of 2.3�C2.8% in SOC, and have caused a loss of 30�C35% of total grassland SOC in China. Improved management practices may reverse the loss of SOC. Exclosure of degraded grassland from grazing and conversion of cropland to abandoned fields (i.e. natural restoration) increased carbon content by 34% and 62% on average. Carbon sequestration rates were greatest during the first 30 yr after treatments began and tended to be greatest in the top 10 cm of soil. Carbon sequestration potential was negatively related to initial carbon and nitrogen concentrations in soils. Exclosure from grazing and the conversion of cropland to abandoned fields resulted in average carbon sequestration rates of 130.4 g C m 612 yr 611 for 0�C40 cm soil and 128.0 g C m 612 yr 611 for 0�C30 cm soil, representing annual average increases of 5.4�C6.3%. Based on our results, achievement of the national objective to exclude grazing livestock from 150 million ha of China's grasslands and to establish 30 million ha of cultivated pasture by 2020 would sequester over 0.24 Pg C yr 611, which is equivalent to about 16% of fossil fuel CO 2 emissions in China in 2006.
DOI:10.1111/j.1469-8137.2007.02237.xURLPMID:17944829 [��������: 1]
090004 Global warming and a changing precipitation regime could have a profound impact on ecosystem carbon fluxes, especially in arid and semiarid grasslands where water is limited. A field experiment manipulating temperature and precipitation has been conducted in a temperate steppe in northern China since 2005. 090004 A paired, nested experimental design was used, with increased precipitation as the primary factor and warming simulated by infrared radiators as the secondary factor. 090004 The results for the first 2 yr showed that gross ecosystem productivity (GEP) was higher than ecosystem respiration, leading to net C sink (measured by net ecosystem CO 2 exchange, NEE) over the growing season in the study site. The interannual variation of NEE resulted from the difference in mean annual precipitation. Experimental warming reduced GEP and NEE, whereas increased precipitation stimulated ecosystem C and water fluxes in both years. Increased precipitation also alleviated the negative effect of experimental warming on NEE. 090004 The results demonstrate that water availability plays a dominant role in regulating ecosystem C and water fluxes and their responses to climatic change in the temperate steppe of northern China.
DOI:10.1016/j.jaridenv.2007.01.002URL
In order to explore the response patterns of gas exchange to the increased grazing intensity and to test whether or not plants have compensatory growth under animal grazing disturbance, we have conducted this experiment in Hunshandak Sandland of Northern China. Changes in net photosynthetic rate (Pn), transpiration rate (), and water use efficiency (WUE) of four plant species (Cleistogenes squarrosa, Agropyron cristatum, Artemisia frigida, Potentilla acaulis) in four grazing intensities, non-, light-, moderate- and high-grazed plots were investigated. Pn of all four species increased from non-grazed plots to moderate-grazed plots then decreased under high-grazed conditions. However, P. acaulis increased its Pn from moderately grazed plots to high-grazed plots, suggesting a strong compensatory photosynthetic growth. Transpirations of the four species elevated considerably with the increase of grazing intensity. Except for Artemisia frigida, WUE of others increased significantly from non-grazed plots to moderately grazed plots then decreased in high-grazed plots. Our results indicate that, while a sharp decrease in leaf areas and aboveground biomass, photosynthetic growth and WUE were enhanced under moderate level of grazing. With a level of grazing increased, the plant functions were declined as a result. Therefore, grassland in Hunshandak Sandland fails to support the present stocking rate.
DOI:10.2111/REM-D-09-00015.1URL [��������: 1]
Los pastizales ocupan 50% de la superficie del planeta y por lo tanto son importantes en el ciclo de Carbono (C) terrestre. Para los pastizales y otros ecosistemas terrestres, el balance entre el consumo fotosint��tico del di��xido de carbono (CO 2 ) y la p��rdida de CO 2 debido a la respiraci��n var��a de un a09o a otro en respuesta a la variaci��n del medio ambiente en el transcurso del a09o. La variabilidad en el intercambio del CO 2 resulta de las diferencias que se dan durante el a09o en 1) variables ambientales en un momento dado en el ciclo anual (efectos directos del medio ambiente) y en 2) la respuesta de los flujos hacia un determinado cambio en el medioambiente debido a los cambios en el transcurso del a09o en los factores biol��gicos que regulan la fotos��ntesis y la respiraci��n (cambio funcional). El cambio funcional se calcula como la contribuci��n de las diferencias entre los a09os en la pendiente de las relaciones del flujo y el medio ambiente a la varianza total en los flujos explicada por el medio ambiente. El cambio funcional complica las predicciones basadas en el medio ambiente del intercambio del CO 2 . Sin embargo, sus causas y la contribuci��n a la variabilidad de los flujos no se han definido satisfactoriamente. Determinamos las contribuciones del cambio funcional y los efectos directos del medio ambiente a la variaci��n entre a09os en el intercambio neto del ecosistema de CO 2 (NEE) de ocho ecosistemas de pastizales en la parte oeste del los Estados Unidos (58-sitios-a09os de datos). Pronosticamos que 1) el cambio funcional esta correlacionado con los cambios entre a09os de la precipitaci��n en cada tipo de pastizal y 2) la contribuci��n del cambio funcional a la variaci��n en NEE aumenta entre los pastizales a medida que la precipitaci��n se incrementa. El cambio funcional explic�� 10�C40% de la variaci��n en NEE y representaron m��s del doble de la variaci��n en los flujos de los efectos directos de la variabilidad ambiental para seis de ocho ecosistemas. Los cambios funcionales estuvieron asociados con la variaci��n entre a09os en la precipitaci��n de la mayor��a del los pastizales, pero contrariamente a la predicci��n contribuy�� proporcionalmente m��s a variaci��n en NEE en ecosistemas ��ridos que los m��sicos. Estos resultados indican que debemos de tomar en cuenta la influencia de la precipitaci��n en la relaci��n del flujo y medio ambiente para poder distinguir los efectos que ejerce el medio ambiente de los que tiene el manejo del balance de C en los pastizales.
URL [��������: 1]
���������ɹŲ�ԭ����Ҫ�����÷�ʽ����Щ����,���ڳ��ڸ�ǿ�ȵķ������²ݵ��������ֺ������͡���ˮ�����½�,�����˲ݵص��˻��ٶ�,�Ӷ�ʹ�ݵ���̬ϵͳ��̼��DZ�����͡�ˮ����Ϊ�����ɹŵ��Ͳ�ԭ��̬ϵͳ̼ѭ�����̵�������Ҫ�������ӡ�̼��Ԫ����Ϊ½����̬ϵͳ���������������ԴҪ��,���ߵ�ѭ����������,����Ӱ�졣������ֲ���л����ں������еı�����ϵ�ںܴ�̶��Ͽ�������̬ϵͳ̼ѭ����ƽ��,�����Ųݵ���̬ϵͳ̼Դ����ת������������̬ϵͳ̼���������е�ЧӦͨ����������ˮ�����������,��Ӱ��ݵ�̼�������̵ĸ�������,ˮ�������Ӱ�첻�ݺ��ӡ� ���о������ɵ��Ͳ�ԭ���ڷ���ʵ����,�����ˮ������ʵ�顣����̽�ֲ�ͬ����ǿ�ȶ��ҹ��´����Ͳ�ԭ̼������Ӱ�졣�����о�ˮ�����ӶԳ��ڷ��������µIJ�ԭ��̬ϵͳ̼������Ӱ�졣�ر��Dz�ͬ�����ݶ���,���Ͳ�ԭ��̬ϵͳ̼������ˮ�����ӵ���Ӧ��ȡ��������Ҫ����� (1)���ڵķ������ŵ�����̬ϵͳ��̼����(Net Ecosystem CO2Exchanges,NEE)����̬ϵͳ�ܳ���������(Gross Ecosystem Productivity, GEP)����̬ϵͳ����(Ecosystem Respiration, ER)������,���������ݶȼ����������졣��������û�иı���̬ϵͳ̼Դ��̼��Ĺ�ϵ�� (2)�����Ե��Ͳ�ԭȺ���������������ݺʹ���é�ı����仯������Ӱ�졣���ŷ���ǿ�ȵ����Ӵ���é��Ⱥ���еı�����������,����ݵı��������½��� (3)�µ���̬ϵͳ��̼����(NEE)��������ƽ��,������(ƽ�غ��µ�)����̬ϵͳ����������(GEP)û������Ӱ�졣 (4)ˮ�����Ӷ���̬ϵͳ̼�����дٽ�����,ˮ������ЧӦ��Ϊ������ˮ��ͬʱ����ʹƽ��ϵͳ��NEE, ER��GEP�ֱ�������28%,9%��18%,�µطֱ�������2%,14%��21%�� (5)�ݵ���̬ϵͳ�ľ�̼���������������ⶨ��ݾ�����Ϊ�������ߡ����������ڽϵ�,�������ݵ�����������,̼���յķ�ֵ��������8�³�,֮������,��������ĩ������ת��Ϊ̼Դ����̬ϵͳ̼�����ļ��ڶ�̬�뽵�������¶ȵļ��ڱ仯ģ�����Ǻϡ� (6)����������ˮ��ͬʱ���Ӷ�����������Ⱥ��ĵ���������,����Ⱥ����������ݺʹ���é����Ⱥ���б����仯û������Ӱ�졣 (7)��̬ϵͳ̼����ָ��NEE��ER��GEP���������¶ȳ���������ع�ϵ,����������ˮ�ֹ�ϵ�������� ������ͬ����ǿ�ȸ�����,�ݵ���̬ϵͳ̼������ˮ�����ӵ���Ӧ,�Լ���ȫ��C02Ũ�����ߵĴ���,ͨ��ˮ�����Ӷ��˻���ԭ�ָ����о�,Ϊ����ˮ��������������ŶԲݵ�̼����������Ӱ�������Ҫ����������,���ڶ����ɹŵ��Ͳ�ԭ�ƶ������ķ����ƶȺ��˻���̬ϵͳ���ܵĻָ��ṩһ�����������ݡ�
URL [��������: 1]
���������ɹŲ�ԭ����Ҫ�����÷�ʽ����Щ����,���ڳ��ڸ�ǿ�ȵķ������²ݵ��������ֺ������͡���ˮ�����½�,�����˲ݵص��˻��ٶ�,�Ӷ�ʹ�ݵ���̬ϵͳ��̼��DZ�����͡�ˮ����Ϊ�����ɹŵ��Ͳ�ԭ��̬ϵͳ̼ѭ�����̵�������Ҫ�������ӡ�̼��Ԫ����Ϊ½����̬ϵͳ���������������ԴҪ��,���ߵ�ѭ����������,����Ӱ�졣������ֲ���л����ں������еı�����ϵ�ںܴ�̶��Ͽ�������̬ϵͳ̼ѭ����ƽ��,�����Ųݵ���̬ϵͳ̼Դ����ת������������̬ϵͳ̼���������е�ЧӦͨ����������ˮ�����������,��Ӱ��ݵ�̼�������̵ĸ�������,ˮ�������Ӱ�첻�ݺ��ӡ� ���о������ɵ��Ͳ�ԭ���ڷ���ʵ����,�����ˮ������ʵ�顣����̽�ֲ�ͬ����ǿ�ȶ��ҹ��´����Ͳ�ԭ̼������Ӱ�졣�����о�ˮ�����ӶԳ��ڷ��������µIJ�ԭ��̬ϵͳ̼������Ӱ�졣�ر��Dz�ͬ�����ݶ���,���Ͳ�ԭ��̬ϵͳ̼������ˮ�����ӵ���Ӧ��ȡ��������Ҫ����� (1)���ڵķ������ŵ�����̬ϵͳ��̼����(Net Ecosystem CO2Exchanges,NEE)����̬ϵͳ�ܳ���������(Gross Ecosystem Productivity, GEP)����̬ϵͳ����(Ecosystem Respiration, ER)������,���������ݶȼ����������졣��������û�иı���̬ϵͳ̼Դ��̼��Ĺ�ϵ�� (2)�����Ե��Ͳ�ԭȺ���������������ݺʹ���é�ı����仯������Ӱ�졣���ŷ���ǿ�ȵ����Ӵ���é��Ⱥ���еı�����������,����ݵı��������½��� (3)�µ���̬ϵͳ��̼����(NEE)��������ƽ��,������(ƽ�غ��µ�)����̬ϵͳ����������(GEP)û������Ӱ�졣 (4)ˮ�����Ӷ���̬ϵͳ̼�����дٽ�����,ˮ������ЧӦ��Ϊ������ˮ��ͬʱ����ʹƽ��ϵͳ��NEE, ER��GEP�ֱ�������28%,9%��18%,�µطֱ�������2%,14%��21%�� (5)�ݵ���̬ϵͳ�ľ�̼���������������ⶨ��ݾ�����Ϊ�������ߡ����������ڽϵ�,�������ݵ�����������,̼���յķ�ֵ��������8�³�,֮������,��������ĩ������ת��Ϊ̼Դ����̬ϵͳ̼�����ļ��ڶ�̬�뽵�������¶ȵļ��ڱ仯ģ�����Ǻϡ� (6)����������ˮ��ͬʱ���Ӷ�����������Ⱥ��ĵ���������,����Ⱥ����������ݺʹ���é����Ⱥ���б����仯û������Ӱ�졣 (7)��̬ϵͳ̼����ָ��NEE��ER��GEP���������¶ȳ���������ع�ϵ,����������ˮ�ֹ�ϵ�������� ������ͬ����ǿ�ȸ�����,�ݵ���̬ϵͳ̼������ˮ�����ӵ���Ӧ,�Լ���ȫ��C02Ũ�����ߵĴ���,ͨ��ˮ�����Ӷ��˻���ԭ�ָ����о�,Ϊ����ˮ��������������ŶԲݵ�̼����������Ӱ�������Ҫ����������,���ڶ����ɹŵ��Ͳ�ԭ�ƶ������ķ����ƶȺ��˻���̬ϵͳ���ܵĻָ��ṩһ�����������ݡ�
DOI:10.1007/s00442-005-0261-7URLPMID:16205950 [��������: 1]
Abstract Landscape position, grazing, and seasonal variation in precipitation and temperature create spatial and temporal variability in soil processes, and plant biomass and composition in grasslands. However, it is unclear how this variation in plant and soil properties affects carbon dioxide (CO2) fluxes. The aim of this study is to explore the effect of grazing, topographic position, and seasonal variation in soil moisture and temperature on plant assimilation, shoot and soil respiration, and net ecosystem CO2 exchange (NEE). Carbon dioxide fluxes, vegetation, and environmental variables were measured once a month inside and outside long-term ungulate exclosures in hilltop (dry) to slope bottom (mesic) grassland throughout the 2004 growing season in Yellowstone National Park. There was no difference in vegetation properties and CO2 fluxes between the grazed and the ungrazed sites. The spatial and temporal variability in CO2 fluxes were related to differences in aboveground biomass and total shoot nitrogen content, which were both related to variability in soil moisture. All sites were CO2 sinks (NEE>0) for all our measurements taken throughout the growing season; but CO2 fluxes were four- to fivefold higher at sites supporting the most aboveground biomass located at slope bottoms, compared to the sites with low biomass located at hilltops or slopes. The dry sites assimilated more CO2 per gram aboveground biomass and stored proportionally more of the gross-assimilated CO2 in the soil, compared to wet sites. These results indicate large spatio-temporal variability of CO2 fluxes and suggest factors that control the variability in Yellowstone National Park.
URL [��������: 1]
����仯��������������ó̶ȼӾ罫�����ı���̬ϵͳ�Ľṹ����,��ͨ��Ӱ��̼ѭ����������ȫ������仯��½����̬ϵͳ̼ѭ����ȫ��̼ѭ������Ҫ��ɲ��֡��ݵ�Լռ½�������20-25%,��������̼����ȫ��̼ƽ������з�������Ҫ�ĵ��ڹ��ܡ�ͬʱ,�ݵ������������ϵ͡���̬ϵͳ�ṹ��ԭ�����ܵ��ؼ�������Դ�������Ըı���ȷ�����Ӱ��,������Ѹ�ٻָ������,��֪�ݵ���̬ϵͳ̼�����Ի����仯����Ӧ���ɼ���DZ�ڵ��ػ���,����ȷ��������仯����Ϊ���żӾ�״����ȫ��̼��֧��ֲ��ɻ�ȱ�Ĺؼ����ڡ����о����ҹ�������Ȼ�ݵؽ��С��Է���ʵ��ƽ̨Ϊ����,��չ�˷����������ʩ��������Ӱ������̬ϵͳ̼�����ı�ij���ϵͳ�о�,�Լ������ʩ��������,ģ��ͻ�����˸ɺ��¼��Ծ���̬ϵͳCO2����(NEE)Ӱ��Ķ����о���ͨ���Ƚϲ�ͬ������,��̬ϵͳ̼�������ͨ���;�ͨ���IJ���,�������뻷������������֮�����ϵ,��ʾ���۲ݵ���̬ϵͳ̼ͨ����δ����ˮ��ָı䡢����ǿ�Ⱥ͵��������ӵ���Ӧ����,̽����Ӱ�����Ҫ;��,��ȷ���еĹؼ��Ե������Ӻ���Ӧ���ơ�ͨ��3��ķ�����ʩ��(ģ��س���)������(ģ�⽵ˮ����)����Ұ�����ʵ��,�Լ�2014�����ģ�⼫�˸ɺ�ʵ��,�����������Ҫ�о����:(1)�������Ӽ���ش̼��˵�����������NEE,�������۲ݵ��������صĿ���������̬ϵͳ̼ѭ������Ҫ���������ӡ���ʵ���ڼ�,���ӽ�ˮ�������������γɺ���������Ӱ�첻�������ݵ����������ڽ�ˮ����������ƽ��NEE��Ӱ��ҪԶ�����������ܽ�ˮ�������ۺϿ�����Ϊ��ˮ����Ȼ��ˮ�Խ�ˮ��ֵ�Ӱ�����,NEE�뽵��Ƶ�ʵ�����Ը��ڽ���������Щ���ͻ����ʾ���ڽ�Ϊʪ������۲ݵ���̬ϵͳ��,��ˮʱ��ֲ��ڵ���NEE�����о�����Ҫ���á�(2)ģ�⼫�˸ɺ��¼������ؽ����˲ݵ���̬ϵͳCO2����,����������Ӱ���С��ͬʱ,�о�������������ˮ�ֹ�����NEE�Ļָ����ʳ�����ع�ϵ,˵��ˮ��ȱ���Ἣ�������ֲ��Ĺ������,�ݵ�ֲ���������ά������ˮ�ֵ��ȶ������ٵĻ��Զ���̼�������Ļ��,�Ի�ȡ���ź���ߵĻָ�����������ʩ�������괦���ᵼ��NEE�Ըɺ����г̶�(SNEE)�����ӡ��ۺϷ����������ص�������ݺ���,ֲ��ĸ��ڱȺ�����������SNEE�ĸı��м��ߵ����Ͻ��Ͷȡ���Щ�������,������Դ�ḻ������ı�ֲ���������������,����������̬ϵͳˮ�ֹ���ƽ���ˮ��в�ȵ����ܺ�Ӧ������,���Բݵ���δ���ɺ�Ƶ�����ӵĴ���ά�ֻ������̼�㹦�ܲ���������Ӱ�졣(3)�����������������۲ݵ�Ⱥ��ֲ�������,�ܹ���һ���̶��ϻ�����ʩ�������괦�����µ�ֲ������Խ������⡣��ֲ���������,��ʳ������������ֵĴ�����ʳ,�Լ���Դ�ḻ���������������ʿ��Ե�����Ƿ�������ֲ����������ӵ���Ҫԭ��ͬʱ,�����������д�������nee�����������Ĵٽ�����,������ʩ�������ͷ��������괦������������Ľ�������,��nee��Ӱ����Ϊ���ԡ����,�������ܻ��Ƴ�����ֲ������������,���Բݵػ�����Դ���ú�ת��Ч�ʵĸ���,ʹ�侻����Ч�������ڴٽ���̬ϵͳ̼�Ĺ̶���(4)���۲ݵ�������������(srh)������ˮ�ֺ͵��صĸı伫Ϊ����,�������ܺ���(srtot)����������(sra)��ʩ�����������Ӧ����к�ǿ�����������ԡ�ʩ�������괦��,���ڸɺ�����ж�srtot��sra������Ӱ�졣��Щ�������,���۲ݵ���̬ϵͳsrhͨ����Ҫ�ܵ��������ص�����,��sraͨ������Ҫ�ܵ�ֲ���������ơ���������������,�ر���srh��srtot���ڼ�ǿ����������,������Դ���Ӷ�ֲ�������Ĵ̼�������һ���̶��ϻ��������sra�����ơ��ṹ����ģ�ͷ�������,������ʩ�������괦��ͨ��Ӱ����ڱȡ��������������������������زݵ���̬ϵͳsra��srh����һ����ǿ���˺���̼����Ĺ���,�Լ���������Ժ���������������������۲ݵ���������������������Ҫ�����á��ܶ���֮,���۲ݵ���δ��ȫ������仯�ͷ�������Ӱ����,���������ر���srh�����������ڽ�ˮ�͵����������ӡ���������Ϊ������������,����ǿ������,���۲ݵؿ��������Ÿ�Ϊ�Ͼ�������Ӫ��Ԫ�غͺ���̼����黹������ʧ��ʹ�λ,�Ӷ�����δ���ݵ�������������̬ϵͳ̼���ͷŹ�������ռ�ı������ۺ������о����,��δ��ȫ������仯Ӱ����,���۲ݵ������������������ڵ����������ӡ������ڼ�����������,��ˮ�������,Ƶ���½�,���۲ݵؿ��������Ÿ�Ϊ�Ͼ���ˮ�ֹ���ʧ��ʹ�λ,�Ӷ�����δ���ݵ���̬ϵͳ̼�㹦��;������ʩ��������������۲ݵ���̬ϵͳȺ����ɺ�ֲ���Ͼ���������Ӱ�졣������ȫ��仯���Ӵ��ڽ�������,������ά�ֲݵ�Ⱥ��ṹ�ȶ���,���ܷŴ�ȫ��仯���Ӷ����۲ݵ���̬ϵͳco2��������������;ͬʱ,���ڷ�����ֲ�������������ڱ����ߵ���������,��δ����ˮ�͵��������ӱ�����,�ݵؼ�������ܹ�������̬ϵͳ̼�����Լ��˸ɺ��¼���������,��������������ʧ�����о�����̬ϵͳCO2���������������ĽǶ�,��һ��֤ʵ��ȫ������仯����Ϊ�������ö����۲ݵ���̬ϵͳ̼ѭ����̼�㹦�ܵ�����Ӱ��,����ЩӰ��DZ�ڻ����IJ��;�����Ҫ�����ۼ�ֵ������,���ڷ����ڲݵ���Դ�������䷽���ͻ������,��Щ����ҲΪ�ݵع����߶Բݵ���̬��ֵ�����ü�ֵ�ı����������ṩ������֧�ֺ�ָ����
URL [��������: 1]
����仯��������������ó̶ȼӾ罫�����ı���̬ϵͳ�Ľṹ����,��ͨ��Ӱ��̼ѭ����������ȫ������仯��½����̬ϵͳ̼ѭ����ȫ��̼ѭ������Ҫ��ɲ��֡��ݵ�Լռ½�������20-25%,��������̼����ȫ��̼ƽ������з�������Ҫ�ĵ��ڹ��ܡ�ͬʱ,�ݵ������������ϵ͡���̬ϵͳ�ṹ��ԭ�����ܵ��ؼ�������Դ�������Ըı���ȷ�����Ӱ��,������Ѹ�ٻָ������,��֪�ݵ���̬ϵͳ̼�����Ի����仯����Ӧ���ɼ���DZ�ڵ��ػ���,����ȷ��������仯����Ϊ���żӾ�״����ȫ��̼��֧��ֲ��ɻ�ȱ�Ĺؼ����ڡ����о����ҹ�������Ȼ�ݵؽ��С��Է���ʵ��ƽ̨Ϊ����,��չ�˷����������ʩ��������Ӱ������̬ϵͳ̼�����ı�ij���ϵͳ�о�,�Լ������ʩ��������,ģ��ͻ�����˸ɺ��¼��Ծ���̬ϵͳCO2����(NEE)Ӱ��Ķ����о���ͨ���Ƚϲ�ͬ������,��̬ϵͳ̼�������ͨ���;�ͨ���IJ���,�������뻷������������֮�����ϵ,��ʾ���۲ݵ���̬ϵͳ̼ͨ����δ����ˮ��ָı䡢����ǿ�Ⱥ͵��������ӵ���Ӧ����,̽����Ӱ�����Ҫ;��,��ȷ���еĹؼ��Ե������Ӻ���Ӧ���ơ�ͨ��3��ķ�����ʩ��(ģ��س���)������(ģ�⽵ˮ����)����Ұ�����ʵ��,�Լ�2014�����ģ�⼫�˸ɺ�ʵ��,�����������Ҫ�о����:(1)�������Ӽ���ش̼��˵�����������NEE,�������۲ݵ��������صĿ���������̬ϵͳ̼ѭ������Ҫ���������ӡ���ʵ���ڼ�,���ӽ�ˮ�������������γɺ���������Ӱ�첻�������ݵ����������ڽ�ˮ����������ƽ��NEE��Ӱ��ҪԶ�����������ܽ�ˮ�������ۺϿ�����Ϊ��ˮ����Ȼ��ˮ�Խ�ˮ��ֵ�Ӱ�����,NEE�뽵��Ƶ�ʵ�����Ը��ڽ���������Щ���ͻ����ʾ���ڽ�Ϊʪ������۲ݵ���̬ϵͳ��,��ˮʱ��ֲ��ڵ���NEE�����о�����Ҫ���á�(2)ģ�⼫�˸ɺ��¼������ؽ����˲ݵ���̬ϵͳCO2����,����������Ӱ���С��ͬʱ,�о�������������ˮ�ֹ�����NEE�Ļָ����ʳ�����ع�ϵ,˵��ˮ��ȱ���Ἣ�������ֲ��Ĺ������,�ݵ�ֲ���������ά������ˮ�ֵ��ȶ������ٵĻ��Զ���̼�������Ļ��,�Ի�ȡ���ź���ߵĻָ�����������ʩ�������괦���ᵼ��NEE�Ըɺ����г̶�(SNEE)�����ӡ��ۺϷ����������ص�������ݺ���,ֲ��ĸ��ڱȺ�����������SNEE�ĸı��м��ߵ����Ͻ��Ͷȡ���Щ�������,������Դ�ḻ������ı�ֲ���������������,����������̬ϵͳˮ�ֹ���ƽ���ˮ��в�ȵ����ܺ�Ӧ������,���Բݵ���δ���ɺ�Ƶ�����ӵĴ���ά�ֻ������̼�㹦�ܲ���������Ӱ�졣(3)�����������������۲ݵ�Ⱥ��ֲ�������,�ܹ���һ���̶��ϻ�����ʩ�������괦�����µ�ֲ������Խ������⡣��ֲ���������,��ʳ������������ֵĴ�����ʳ,�Լ���Դ�ḻ���������������ʿ��Ե�����Ƿ�������ֲ����������ӵ���Ҫԭ��ͬʱ,�����������д�������nee�����������Ĵٽ�����,������ʩ�������ͷ��������괦������������Ľ�������,��nee��Ӱ����Ϊ���ԡ����,�������ܻ��Ƴ�����ֲ������������,���Բݵػ�����Դ���ú�ת��Ч�ʵĸ���,ʹ�侻����Ч�������ڴٽ���̬ϵͳ̼�Ĺ̶���(4)���۲ݵ�������������(srh)������ˮ�ֺ͵��صĸı伫Ϊ����,�������ܺ���(srtot)����������(sra)��ʩ�����������Ӧ����к�ǿ�����������ԡ�ʩ�������괦��,���ڸɺ�����ж�srtot��sra������Ӱ�졣��Щ�������,���۲ݵ���̬ϵͳsrhͨ����Ҫ�ܵ��������ص�����,��sraͨ������Ҫ�ܵ�ֲ���������ơ���������������,�ر���srh��srtot���ڼ�ǿ����������,������Դ���Ӷ�ֲ�������Ĵ̼�������һ���̶��ϻ��������sra�����ơ��ṹ����ģ�ͷ�������,������ʩ�������괦��ͨ��Ӱ����ڱȡ��������������������������زݵ���̬ϵͳsra��srh����һ����ǿ���˺���̼����Ĺ���,�Լ���������Ժ���������������������۲ݵ���������������������Ҫ�����á��ܶ���֮,���۲ݵ���δ��ȫ������仯�ͷ�������Ӱ����,���������ر���srh�����������ڽ�ˮ�͵����������ӡ���������Ϊ������������,����ǿ������,���۲ݵؿ��������Ÿ�Ϊ�Ͼ�������Ӫ��Ԫ�غͺ���̼����黹������ʧ��ʹ�λ,�Ӷ�����δ���ݵ�������������̬ϵͳ̼���ͷŹ�������ռ�ı������ۺ������о����,��δ��ȫ������仯Ӱ����,���۲ݵ������������������ڵ����������ӡ������ڼ�����������,��ˮ�������,Ƶ���½�,���۲ݵؿ��������Ÿ�Ϊ�Ͼ���ˮ�ֹ���ʧ��ʹ�λ,�Ӷ�����δ���ݵ���̬ϵͳ̼�㹦��;������ʩ��������������۲ݵ���̬ϵͳȺ����ɺ�ֲ���Ͼ���������Ӱ�졣������ȫ��仯���Ӵ��ڽ�������,������ά�ֲݵ�Ⱥ��ṹ�ȶ���,���ܷŴ�ȫ��仯���Ӷ����۲ݵ���̬ϵͳco2��������������;ͬʱ,���ڷ�����ֲ�������������ڱ����ߵ���������,��δ����ˮ�͵��������ӱ�����,�ݵؼ�������ܹ�������̬ϵͳ̼�����Լ��˸ɺ��¼���������,��������������ʧ�����о�����̬ϵͳCO2���������������ĽǶ�,��һ��֤ʵ��ȫ������仯����Ϊ�������ö����۲ݵ���̬ϵͳ̼ѭ����̼�㹦�ܵ�����Ӱ��,����ЩӰ��DZ�ڻ����IJ��;�����Ҫ�����ۼ�ֵ������,���ڷ����ڲݵ���Դ�������䷽���ͻ������,��Щ����ҲΪ�ݵع����߶Բݵ���̬��ֵ�����ü�ֵ�ı����������ṩ������֧�ֺ�ָ����
URL [��������: 2]
�ݵط����Ϳ������й�����ũ������������Ҫ�������÷�ʽ,�о���ͬ��������������̬ϵͳ��̼ͨ���仯���̼�Ӱ������,�Ժ�������ũ��������������������,������̬ϵͳ̼�̳�����������Ҫ���塣������ũ���������ĺӱ����ϵ��Ͳ�ԭΪ�о�����,���þ�̬����䷨�о�������(������UG,�ʶȷ�����MG,�ضȷ�����HG)��ũ��(CL)�Ͷ������˹���ݵ�(AG)�Ȳ�ͬ��������������̬ϵͳCO2������(NEE)���������̬ϵͳ����(Re)����̬ϵͳCO2�̶ܹ���(GEP)��Ӱ��,�ۺϷ�����ͨ�����ӱ仯����Ҫ��������,��Ҫ�о�������£�(1)���������������ǿ�����Ӷ���С,�ʶȷ��������ڵ���0-12cm��ϵ���������ۻ���HG���ص�����������UG��������58%,��ϵ��������������12%(p0.05)��MG���ص�����������UG��������23%,��ϵ��������������10%(p0.05)��CL��AG�ĵ�����������������UG(p0.05),����CL���ظ�ϵ��������������UG��AG(p0.05)���ʶȷ����������Ӳݵ����������л�̼��ȫ������(p0.05)��(2)�����غ�ũ���NEE�������(GEP��Re)�������Եļ��ڱ仯,��ʱ����������Ķ��η����Իع��ϵ(p0.05),����AG���ؽ�GEP��ʱ��ķ����Իع���ϴﵽ����ˮƽ(p0.05)��UG��MG��HG���ص�NEE��ʱ����ϵ����(2012-2014��)����ϵ��R2�ֱ�Ϊ0.64,0.79��0.50,������NEE��ʱ���ƽ������ϵ��(0.63)Re(0.70)GEP(0.77),˵���ڷ����ݵ���̬ϵͳ,�����NEE��Re,GEP�����ܼ��ڱ仯Ӱ�졣�����ص�NEE��GEP��ʱ仯��������(p0.05),2013���NEE�ֱ��2012��2014����������33%��34%(p0.05),GEP�ֱ���������24%��26%(p0.05)������Re��ʲ��첻����(p0.05)��(3)�ۺ�������������������ع�������ַ������¾�NEE��AGB(����������)��VWC(������ˮ��)������������ع�ϵ(NEE=-0.05AGB-0.09VWC, R2=0.95, p0.0001), AGB�ɽ���NEE�仯��92%,���¾�NEE�仯����Ҫ��������(p0.0001)���¾�GEP����Ҫ��������Ҳ��AGB,ͬ������GEP�仯��92%(GEP=-0.068AGB-0.202Ta-0.102Ts, R2=0.97, p0.0001);�¾�Re����Ҫ���������Ǵ����¶�(Ta),�ɽ���Re�仯��93%(Re=0.23Ta-0.057VWC+0.017AGB+0.038Ts, R2=0.97, p0.0001)�� CL����,�¾�NEE (NEE=-0.026AGB, R2=0.81, p0.0001)��GEP (GEP=-0.04AGB-1.39VWC+0.98Ts, R2=0.95, p0.0001)����Ҫ�������Ӿ���AGB, AGB������NEE��GEP�仯��81%(p0.0001)��VWC���¾�Re�仯����Ҫ��������(Re=0.51VWC-0.11Ta+0.013AGB, R2=0.97, p0.0001),����Re�仯��90%��AG����,�¾�NEE (NEE=-1.29Ta+0.57Ts+0.46VWC, R2=0.98, p0.0001)��GEP (GEP=-0.71Ta+0.32Ts -0.01AGB, R2=0.99, p0.0001)����Ҫ�������Ӿ���Ta,�ֱ����NEE��GEP�仯��95%��98%��Re�仯����Ҫ���������������¶�(Ts),����Re�仯��96%(Re=0.22Ts+0.01AGB, R2=0.98, p0.0001)��(4)ʵ������������������������������(2012-2014��)������̼����״̬,��HG���ص�NEE��UG��MG�ֱ���������36%��51%(p0.05)��MG����NEE��UG��HG�ֱ���������31%��106%(p0.05)���ɼ��ضȷ����ή�Ͳݵ���̬ϵͳCO2����DZ�����ʶȷ�����ٽ�̼���ա�UG��CL��AG���ص�NEE��GEP���첻����(p0.05),��CL����Re��������UG(p0.05)��UG��MG��HG������������(2012-2014��)������ۻ���̼���ֱ�Ϊ:771g C m-2,1005 gCm-2��491gCm-2, HGUGMG, HG���������˲ݵصĹ�̼����(p0.05)��2013��,UG��MG��HG��AG��CL������ص��������ۻ���̼���ֱ�Ϊ1008 gCm-2,1240g Cm-2,555 gCm-2,1240 g Cm-2��926 g Cm-2, AG, MGUG��CLHG����ʱ仯��Ӱ������ݵ��ۻ���̼��,2012-2014��ķ����ص�����������ۻ���̼���ֱ�Ϊ643gCm-2,934gC m2��690 g Cm-2,201320142012�ꡣ
URL [��������: 2]
�ݵط����Ϳ������й�����ũ������������Ҫ�������÷�ʽ,�о���ͬ��������������̬ϵͳ��̼ͨ���仯���̼�Ӱ������,�Ժ�������ũ��������������������,������̬ϵͳ̼�̳�����������Ҫ���塣������ũ���������ĺӱ����ϵ��Ͳ�ԭΪ�о�����,���þ�̬����䷨�о�������(������UG,�ʶȷ�����MG,�ضȷ�����HG)��ũ��(CL)�Ͷ������˹���ݵ�(AG)�Ȳ�ͬ��������������̬ϵͳCO2������(NEE)���������̬ϵͳ����(Re)����̬ϵͳCO2�̶ܹ���(GEP)��Ӱ��,�ۺϷ�����ͨ�����ӱ仯����Ҫ��������,��Ҫ�о�������£�(1)���������������ǿ�����Ӷ���С,�ʶȷ��������ڵ���0-12cm��ϵ���������ۻ���HG���ص�����������UG��������58%,��ϵ��������������12%(p0.05)��MG���ص�����������UG��������23%,��ϵ��������������10%(p0.05)��CL��AG�ĵ�����������������UG(p0.05),����CL���ظ�ϵ��������������UG��AG(p0.05)���ʶȷ����������Ӳݵ����������л�̼��ȫ������(p0.05)��(2)�����غ�ũ���NEE�������(GEP��Re)�������Եļ��ڱ仯,��ʱ����������Ķ��η����Իع��ϵ(p0.05),����AG���ؽ�GEP��ʱ��ķ����Իع���ϴﵽ����ˮƽ(p0.05)��UG��MG��HG���ص�NEE��ʱ����ϵ����(2012-2014��)����ϵ��R2�ֱ�Ϊ0.64,0.79��0.50,������NEE��ʱ���ƽ������ϵ��(0.63)Re(0.70)GEP(0.77),˵���ڷ����ݵ���̬ϵͳ,�����NEE��Re,GEP�����ܼ��ڱ仯Ӱ�졣�����ص�NEE��GEP��ʱ仯��������(p0.05),2013���NEE�ֱ��2012��2014����������33%��34%(p0.05),GEP�ֱ���������24%��26%(p0.05)������Re��ʲ��첻����(p0.05)��(3)�ۺ�������������������ع�������ַ������¾�NEE��AGB(����������)��VWC(������ˮ��)������������ع�ϵ(NEE=-0.05AGB-0.09VWC, R2=0.95, p0.0001), AGB�ɽ���NEE�仯��92%,���¾�NEE�仯����Ҫ��������(p0.0001)���¾�GEP����Ҫ��������Ҳ��AGB,ͬ������GEP�仯��92%(GEP=-0.068AGB-0.202Ta-0.102Ts, R2=0.97, p0.0001);�¾�Re����Ҫ���������Ǵ����¶�(Ta),�ɽ���Re�仯��93%(Re=0.23Ta-0.057VWC+0.017AGB+0.038Ts, R2=0.97, p0.0001)�� CL����,�¾�NEE (NEE=-0.026AGB, R2=0.81, p0.0001)��GEP (GEP=-0.04AGB-1.39VWC+0.98Ts, R2=0.95, p0.0001)����Ҫ�������Ӿ���AGB, AGB������NEE��GEP�仯��81%(p0.0001)��VWC���¾�Re�仯����Ҫ��������(Re=0.51VWC-0.11Ta+0.013AGB, R2=0.97, p0.0001),����Re�仯��90%��AG����,�¾�NEE (NEE=-1.29Ta+0.57Ts+0.46VWC, R2=0.98, p0.0001)��GEP (GEP=-0.71Ta+0.32Ts -0.01AGB, R2=0.99, p0.0001)����Ҫ�������Ӿ���Ta,�ֱ����NEE��GEP�仯��95%��98%��Re�仯����Ҫ���������������¶�(Ts),����Re�仯��96%(Re=0.22Ts+0.01AGB, R2=0.98, p0.0001)��(4)ʵ������������������������������(2012-2014��)������̼����״̬,��HG���ص�NEE��UG��MG�ֱ���������36%��51%(p0.05)��MG����NEE��UG��HG�ֱ���������31%��106%(p0.05)���ɼ��ضȷ����ή�Ͳݵ���̬ϵͳCO2����DZ�����ʶȷ�����ٽ�̼���ա�UG��CL��AG���ص�NEE��GEP���첻����(p0.05),��CL����Re��������UG(p0.05)��UG��MG��HG������������(2012-2014��)������ۻ���̼���ֱ�Ϊ:771g C m-2,1005 gCm-2��491gCm-2, HGUGMG, HG���������˲ݵصĹ�̼����(p0.05)��2013��,UG��MG��HG��AG��CL������ص��������ۻ���̼���ֱ�Ϊ1008 gCm-2,1240g Cm-2,555 gCm-2,1240 g Cm-2��926 g Cm-2, AG, MGUG��CLHG����ʱ仯��Ӱ������ݵ��ۻ���̼��,2012-2014��ķ����ص�����������ۻ���̼���ֱ�Ϊ643gCm-2,934gC m2��690 g Cm-2,201320142012�ꡣ
DOI:10.3321/j.issn:1673-5021.2000.06.001URL
�����ɹŶ̻���é��Į��ԭ���� 5��������ˮƽ����������� ,�о�ֲ��Ⱥ��������ʵ���Ӧ���о�������� ,��Ҫֲ��Ƕȡ��߶Ⱥ��ܶ��ڵ������ʺ��е��������±��ֽϸ�ˮƽ ;�е������������ڶ̻���é���� ;���������ʵ����� ,ֲ��Ⱥ���ִ����½� ;�̻���é��ԭ���˵�������Ϊ 1.7�� 2ֻ�� /hm2 /���ꡣ
DOI:10.3321/j.issn:1673-5021.2000.06.001URL
�����ɹŶ̻���é��Į��ԭ���� 5��������ˮƽ����������� ,�о�ֲ��Ⱥ��������ʵ���Ӧ���о�������� ,��Ҫֲ��Ƕȡ��߶Ⱥ��ܶ��ڵ������ʺ��е��������±��ֽϸ�ˮƽ ;�е������������ڶ̻���é���� ;���������ʵ����� ,ֲ��Ⱥ���ִ����½� ;�̻���é��ԭ���˵�������Ϊ 1.7�� 2ֻ�� /hm2 /���ꡣ
DOI:10.3773/j.issn.1005-264x.2010.07.002URL [��������: 1]
Ϊ�˽�ʾ����Դ�����������(Elymus nutans)�˹��ݵ���̬ϵͳ(100��26'-100��41'E,34��17'-34��25'N,����3 980 m)�ľ���̬ϵͳCO2����(NEE),���о�����2006���ж����ϵͳ�۲�����ݷ����˸��˹��ݵص�NEE,�ܳ���������(GPP)����̬ϵͳ���� (Reco)�Լ�Reco/GPP�ı仯��������Ӱ������.CO2���������ֵΪ6.56 g CO2��m-2��d-1,����ŷ�ֵΪ4.87 gCO2��m-2��d-1.GPP������Ϊ1 761 g CO2��m-2,����Լ90%���ϱ���̬ϵͳ����������,CO2����������Ϊ111 gCO2��m-2.5�µ�Reco/GPP�Ը����������������·�,Ϊ90%;6��Reco/GPP��ֵ���,Ϊ79%.��̬ϵͳ�ĺ�����(Q10)Ϊ 4.81,��������������̬ϵͳ.���о�����:��������NEE��Ҫ�ܹ�����ͨ���ܶ�(PPFD)���¶Ⱥͱ���ˮ��ѹ��(VPD)��Ӱ��,��̬ϵͳ�������� Ҫ�������¶ȵĿ���.
DOI:10.3773/j.issn.1005-264x.2010.07.002URL [��������: 1]
Ϊ�˽�ʾ����Դ�����������(Elymus nutans)�˹��ݵ���̬ϵͳ(100��26'-100��41'E,34��17'-34��25'N,����3 980 m)�ľ���̬ϵͳCO2����(NEE),���о�����2006���ж����ϵͳ�۲�����ݷ����˸��˹��ݵص�NEE,�ܳ���������(GPP)����̬ϵͳ���� (Reco)�Լ�Reco/GPP�ı仯��������Ӱ������.CO2���������ֵΪ6.56 g CO2��m-2��d-1,����ŷ�ֵΪ4.87 gCO2��m-2��d-1.GPP������Ϊ1 761 g CO2��m-2,����Լ90%���ϱ���̬ϵͳ����������,CO2����������Ϊ111 gCO2��m-2.5�µ�Reco/GPP�Ը����������������·�,Ϊ90%;6��Reco/GPP��ֵ���,Ϊ79%.��̬ϵͳ�ĺ�����(Q10)Ϊ 4.81,��������������̬ϵͳ.���о�����:��������NEE��Ҫ�ܹ�����ͨ���ܶ�(PPFD)���¶Ⱥͱ���ˮ��ѹ��(VPD)��Ӱ��,��̬ϵͳ�������� Ҫ�������¶ȵĿ���.
DOI:10.13292/j.1000-4890.201606.023URL [��������: 1]
�����ů�ʹ�����������ȫ��仯����Ҫ�������ӡ��ڲݵ���̬ϵͳ��,�������ߺʹ�������������ı�ݵع�̼��C��״��,Ȼ���¶����Ӻʹ��������������Ӱ����̬ϵͳ̼����Ŀǰ������ȷ�����о�ּ���о����º͵������ӶԻ�Į��ԭ̼������Ӱ�졣�ڶ̻���é��Į��ԭ�ϲ���2��2������ȫ���������������,ʹ�ú����������ģ�������ů����ʹ�����ӵ��ʵķ�����ģ�����������,�ڲ�ͬ���������²ⶨ��̬ϵͳ��̼������NEE������̬ϵͳ������ER��������̬ϵͳ��������GEP��,������2013��2014��Ӱ��̻���é��Į��ԭ��̬ϵͳC����������,������£���1������ʹ�����¶�����������0.70�棨P��0.001��,����ʪ������������7.58%��P��0.001������2�����¡��������Ӽ��佻����������������GEP��ER��P��0.05��,������NEEû������Ӱ�죨P��0.05������3��2013��GEP��8�³��ﵽ��ֵ,ER��8��ĩ9�³��ﵽ��ֵ,NEE����GEP��ER�ı仯������;2014��GEP��ER��NEE����8��ĩ9�³��ﵽ��ֵ����4��ER��GEP���Ŵ����¶����ߺͽ�ˮ���Ӷ�����,�����¶Ⱥ�����ʪ��Ҳ��Ӱ����̬ϵͳC��������Ҫ���ء�
DOI:10.13292/j.1000-4890.201606.023URL [��������: 1]
�����ů�ʹ�����������ȫ��仯����Ҫ�������ӡ��ڲݵ���̬ϵͳ��,�������ߺʹ�������������ı�ݵع�̼��C��״��,Ȼ���¶����Ӻʹ��������������Ӱ����̬ϵͳ̼����Ŀǰ������ȷ�����о�ּ���о����º͵������ӶԻ�Į��ԭ̼������Ӱ�졣�ڶ̻���é��Į��ԭ�ϲ���2��2������ȫ���������������,ʹ�ú����������ģ�������ů����ʹ�����ӵ��ʵķ�����ģ�����������,�ڲ�ͬ���������²ⶨ��̬ϵͳ��̼������NEE������̬ϵͳ������ER��������̬ϵͳ��������GEP��,������2013��2014��Ӱ��̻���é��Į��ԭ��̬ϵͳC����������,������£���1������ʹ�����¶�����������0.70�棨P��0.001��,����ʪ������������7.58%��P��0.001������2�����¡��������Ӽ��佻����������������GEP��ER��P��0.05��,������NEEû������Ӱ�죨P��0.05������3��2013��GEP��8�³��ﵽ��ֵ,ER��8��ĩ9�³��ﵽ��ֵ,NEE����GEP��ER�ı仯������;2014��GEP��ER��NEE����8��ĩ9�³��ﵽ��ֵ����4��ER��GEP���Ŵ����¶����ߺͽ�ˮ���Ӷ�����,�����¶Ⱥ�����ʪ��Ҳ��Ӱ����̬ϵͳC��������Ҫ���ء�
DOI:10.17521/cjpe.2004.0044URL [��������: 1]
������������Ƶķ����о������(Leymus chinensis)����ҶƬ��ϲ�����5���¶Ⱥ�5��ˮ���ݶȵ���Ӧ����Ӧ.�������:��ȡ��ж������ɺ���û���������ҶƬ������,�����彻��������������Ӱ��,��ӳ��������������ˮ��в�ȵĽϸ�����.ҶƬ��������26 ��ʱ���,��������Ϊ23 �桢20 �桢29 ���32 ��.�¶�����ʹ�����Ⱥ�������������, ȴʹ������ʺ�ˮ������Ч�ʽ���.ˮ�ֺ��¶ȶ�ҶƬ��������������ʡ������Ⱥ��������ʴ��������Ľ�������,�������¼�ǿ�˸ɺ���ҶƬ���������彻����Ӱ��, ��������ݶ������ɺ�����Ӧ����.���ºɺ��Ľ������ý����������ҹ���ɺ�������ԭ�����������.
DOI:10.17521/cjpe.2004.0044URL [��������: 1]
������������Ƶķ����о������(Leymus chinensis)����ҶƬ��ϲ�����5���¶Ⱥ�5��ˮ���ݶȵ���Ӧ����Ӧ.�������:��ȡ��ж������ɺ���û���������ҶƬ������,�����彻��������������Ӱ��,��ӳ��������������ˮ��в�ȵĽϸ�����.ҶƬ��������26 ��ʱ���,��������Ϊ23 �桢20 �桢29 ���32 ��.�¶�����ʹ�����Ⱥ�������������, ȴʹ������ʺ�ˮ������Ч�ʽ���.ˮ�ֺ��¶ȶ�ҶƬ��������������ʡ������Ⱥ��������ʴ��������Ľ�������,�������¼�ǿ�˸ɺ���ҶƬ���������彻����Ӱ��, ��������ݶ������ɺ�����Ӧ����.���ºɺ��Ľ������ý����������ҹ���ɺ�������ԭ�����������.
URL [��������: 2]
����C02Ũ����������������ů������㷺��ע,�ݵ���������������,���кܴ��̼��DZ���������Dzݵص���Ҫ���÷�ʽ,��������ͨ����ʳ����̤,���زݵ���������Ⱥ��ṹ��������������,��Ӹı�ݵ���̬ϵͳ��̼ѭ����ͬʱ,ˮ������Ҳ����Լ�ݵ�̼��֧�Ĺؼ����ء����о�ѡ��̻���é��Į��ԭ��Ϊ�о�����,�������ڲ�ͬ����������ʵ��ƽ̨,���ա��¡���߶Ȳⶨ�ݵ���̬ϵͳ�����CO2��������,���۲�ֲ��Ⱥ�䡢����Ⱥ�������ֵ����ִ�����ֲ���Ƕȡ�������������������ʪ�ȡ������л�̼�����������¶ȡ�����ı仯����,Ŀ����̽���̻���é��Į��ԭ��̬ϵͳ̼�������ʶԳ��ڲ�ͬ�����ʵ���Ӧ,��ʾֲ�����������������Ӷ�̼������Ӱ��������о�������¡�1.���ڹ��ȷ������¶����������̲��ִ�����������,����ݡ���ľ�����ľ���ִ��������½�,C4ֲ���ִ����������ӡ�2.��̬ϵͳ̼�������������뽵�����Ĺ�ͬӰ��,�������ܹ�������̬ϵͳ̼�����������ʵ���Ӧ�̶ȡ�3.�����������ۻ�����ϵ���������������л�̼����������������������ʡ�4.����ʪ�ȡ������¶ȡ������¶ȶ���̬ϵͳ̼��������Ӱ�졣
URL [��������: 2]
����C02Ũ����������������ů������㷺��ע,�ݵ���������������,���кܴ��̼��DZ���������Dzݵص���Ҫ���÷�ʽ,��������ͨ����ʳ����̤,���زݵ���������Ⱥ��ṹ��������������,��Ӹı�ݵ���̬ϵͳ��̼ѭ����ͬʱ,ˮ������Ҳ����Լ�ݵ�̼��֧�Ĺؼ����ء����о�ѡ��̻���é��Į��ԭ��Ϊ�о�����,�������ڲ�ͬ����������ʵ��ƽ̨,���ա��¡���߶Ȳⶨ�ݵ���̬ϵͳ�����CO2��������,���۲�ֲ��Ⱥ�䡢����Ⱥ�������ֵ����ִ�����ֲ���Ƕȡ�������������������ʪ�ȡ������л�̼�����������¶ȡ�����ı仯����,Ŀ����̽���̻���é��Į��ԭ��̬ϵͳ̼�������ʶԳ��ڲ�ͬ�����ʵ���Ӧ,��ʾֲ�����������������Ӷ�̼������Ӱ��������о�������¡�1.���ڹ��ȷ������¶����������̲��ִ�����������,����ݡ���ľ�����ľ���ִ��������½�,C4ֲ���ִ����������ӡ�2.��̬ϵͳ̼�������������뽵�����Ĺ�ͬӰ��,�������ܹ�������̬ϵͳ̼�����������ʵ���Ӧ�̶ȡ�3.�����������ۻ�����ϵ���������������л�̼����������������������ʡ�4.����ʪ�ȡ������¶ȡ������¶ȶ���̬ϵͳ̼��������Ӱ�졣
DOI:10.11733/j.issn.1007-0435.2015.03.008URL
�������ÿ�·ʽ����̼ͨ������ ϵͳ-LI-8100�ڶ̻���é(Stipa breviflora Griseb)��Į��ԭ�ⶨ4�鲻ͬ�����ʵ������������ʶ�̬,�������귨�ⶨ��ֲ������ۻ�������,��������ⶨ�˸�ϵ����������Ŀ���DZȽϲ�ͬ������ ������������ֲ������ۻ���������ϵ����������Ӱ��,��̽������������������8�·�����������ֲ������ۻ�����������ϵ������������Ӧ���������:���� ����������,ֲ������ۻ�����������,��ϵ��������,������������������Ӱ��;����������������ֲ������ۻ������������������,8�·���������ֵ���ϵ ���������������������������ֲ�����������Ӱ������,����������,�¸��տ�ʼ����,���������������ϸ�,������������,�¸�����������,�������� ���ʼӿ�,�¸�����������������
DOI:10.11733/j.issn.1007-0435.2015.03.008URL
�������ÿ�·ʽ����̼ͨ������ ϵͳ-LI-8100�ڶ̻���é(Stipa breviflora Griseb)��Į��ԭ�ⶨ4�鲻ͬ�����ʵ������������ʶ�̬,�������귨�ⶨ��ֲ������ۻ�������,��������ⶨ�˸�ϵ����������Ŀ���DZȽϲ�ͬ������ ������������ֲ������ۻ���������ϵ����������Ӱ��,��̽������������������8�·�����������ֲ������ۻ�����������ϵ������������Ӧ���������:���� ����������,ֲ������ۻ�����������,��ϵ��������,������������������Ӱ��;����������������ֲ������ۻ������������������,8�·���������ֵ���ϵ ���������������������������ֲ�����������Ӱ������,����������,�¸��տ�ʼ����,���������������ϸ�,������������,�¸�����������,�������� ���ʼӿ�,�¸�����������������
[��������: 1]
[��������: 1]
DOI:10.1111/j.1365-2486.2007.01333.xURL [��������: 1]
Partitioning soil CO 2 efflux into autotrophic ( R A ) and heterotrophic ( R H ) components is crucial for understanding their differential responses to climate change. We conducted a long-term experiment (2000�C2005) to investigate effects of warming 2��C and yearly clipping on soil CO 2 efflux and its components (i.e. R A and R H ) in a tallgrass prairie ecosystem. Interannual variability of these fluxes was also examined. Deep collars (70 cm) were inserted into soil to measure R H . R A was quantified as the difference between soil CO 2 efflux and R H . Warming treatment significantly stimulated soil CO 2 efflux and its components (i.e. R A and R H ) in most years. In contrast, yearly clipping significantly reduced soil CO 2 efflux only in the last 2 years, although it decreased R H in every year of the study. Temperature sensitivity (i.e. apparent Q 10 values) of soil CO 2 efflux was slightly lower under warming ( P >0.05) and reduced considerably by clipping ( P <0.05) compared with that in the control. On average over the 4 years, R H accounted for approximately 65% of soil CO 2 efflux with a range from 58% to 73% in the four treatments. Over seasons, the contribution of R H to soil CO 2 efflux reached a maximum in winter (6590%) and a minimum in summer (6535%). Annual soil CO 2 efflux did not vary substantially among years as precipitation did. The interannual variability of soil CO 2 efflux may be mainly caused by precipitation distribution and summer severe drought. Our results suggest that the effects of warming and yearly clipping on soil CO 2 efflux and its components did not result in significant changes in R H or R A contribution, and rainfall timing may be more important in determining interannual variability of soil CO 2 efflux than the amount of annual precipitation.
URL [��������: 1]
��Щ����,��������Ũ�����������ܵ�Խ��Խ�㷺�Ĺ�ע,�������dz����ӹ������������ŷ��Լ���������ļ��Ź�����Ϊ��Ӧ��ȫ��仯,�ƶ���ѧ�����ķ�չ����,�ҹ���������硶�й�����������Ƶ�������Բߡ������й���ȫ������仯���ҶԲ��о����Լ�������仯�����о�������������ǰ,�ҹ�ͨ���˸�����,ֲ������,��̬�ָ���һϵ�д�ʩ,������������ź�ά�����������������Լ��Ĺ��ס���ظ�ԭ���ҹ���Ҫ������ҵ����,����ҵ�����ǵ��ص�֧���Բ�ҵ�����ǽ����������˿ڵ�����,������ҵ��Ʒ������������ʹ��������ֳ��ģ���������ȷ����������²ݳ������˻���������ɳ����ˮλ�½�����������Խ��͵�һϵ��Σ��������ƶ�����������ҵ����,ʹ������ҵ�����ɳ����ķ�չ,���ҹ�����ؽ����������⡣����ͨ�������Բݵؽ��������Ǹߺ��ݵ�����Ҫ�����÷�ʽ֮һ�����������еIJ�ʳ����̤����й���ȹ��̻��̼�ŷŲ�����ҪӰ�졣�ڷ���������������ʪ�ȡ�ֲ��Ⱥ����ɡ������л�̼�������������ֵ�ָ����ܻᷢ���仯,��Щ���ض���̬ϵͳ����������ҪӰ�졣������������̬ϵͳ����CO_2�Ĺ��̼��临��,����ǿ�Ⱥͷ�����ʷ�ۺ����ö���̬ϵͳ����CO_2���Ӱ�졣����֮����ܻ������ڷ����������͡����ڷ������̶Ըߺ��ݵ�CO_2�ŷŵ�Ӱ�켫�临��,��Ŀǰ,����������̬ϵͳ������������δ���һ�¹�ʶ����Ȼ���кܶ����Ƶ��о�,�����ò�ͬ����ǿ���ҳ�ʱ�������۲�ߺ��ݵ�CO_2�ŷŵ��о������б����������������ظ�ԭ��Ե�ĸߺ��ݵ鿪չ�˳�ʱ��ij����۲�,Ϊ������ظ�ԭ��Ե�ߺ��ݵ�̼�������Լ��ƶ������ķ��������ṩ�������ݡ�����ظ�ԭ��Ե�Ĵ�ʡ���Ӳ���Ǽ�������ݺ�ԭ�ؾ��ڿ�չ��Ϊ������ķ���ʵ�顣���о�����������ƽ��ʵ��,����ţ����ʵ��Ͳ��������ʵ�顣������ʵ���зֱ������˲�ͬ�ķ���ǿ��,���þ�̬��-����ɫ���ⶨCO_2Ũ��,��ȫ��̽����ظ�ԭ������Ե�ߺ��ݵ��ڲ�ͬ����ǿ���µ�CO_2�ŷ�ͨ��,��ͬ����ǿ�ȶ�CO_2�ŷ�ͨ���������Լ�����������ߺ��ݵ���̬ϵͳCO_2�ŷ�ͨ���Ĺ�ϵ���õ�����Ҫ����ͽ�������:(1)����ţ����ʵ����,���ֲ�ͬ����ǿ��,��:����������ȷ������жȷ������ضȷ�����ƽ�����ۻ��ŷ����ֱ�Ϊ:470.31,482.71,462.16��431.44 g C m-2 y-1,���ܵ�ƽ��ֵΪ461.66g C m-2 y-1;�ڲ��������ʵ����,���ֲ�ͬ����ǿ��,��:��ȷ������жȷ������ضȷ�����ƽ�����ۻ��ŷ����ֱ�Ϊ:487.66,430.11��434.78g C m-2y-1,���ܵ�ƽ��ֵΪ450.85g C m-2y-1��(2)������ʵ���о������������е�CO_2�ŷ�ͨ��ҪԶԶ���ڷ�������,���ֳ������Եļ�������������ţ����ʵ����,�������е�CO_2�ŷ�ͨ��ռ���ۻ�CO_2�ŷ�ͨ����68.02%~74.36%;�ڲ��������ʵ����,�������е�CO_2�ŷ�ͨ��ռ���ۻ�CO_2�ŷ�ͨ����69.34%~77.21%��(3)�ڱ��о��������ʵ�������õķ���ǿ���µĽ������:��ͬ����ǿ���¸ߺ��ݵ�CO_2�ŷ�ͨ�����������졣(4)������ʵ����,���Ǿ�����:�¶���CO_2�ŷ�ͨ������ع�ϵ,�������ۻ���ˮ����CO_2�ŷ�ͨ��������Ը���һЩ��
URL [��������: 1]
��Щ����,��������Ũ�����������ܵ�Խ��Խ�㷺�Ĺ�ע,�������dz����ӹ������������ŷ��Լ���������ļ��Ź�����Ϊ��Ӧ��ȫ��仯,�ƶ���ѧ�����ķ�չ����,�ҹ���������硶�й�����������Ƶ�������Բߡ������й���ȫ������仯���ҶԲ��о����Լ�������仯�����о�������������ǰ,�ҹ�ͨ���˸�����,ֲ������,��̬�ָ���һϵ�д�ʩ,������������ź�ά�����������������Լ��Ĺ��ס���ظ�ԭ���ҹ���Ҫ������ҵ����,����ҵ�����ǵ��ص�֧���Բ�ҵ�����ǽ����������˿ڵ�����,������ҵ��Ʒ������������ʹ��������ֳ��ģ���������ȷ����������²ݳ������˻���������ɳ����ˮλ�½�����������Խ��͵�һϵ��Σ��������ƶ�����������ҵ����,ʹ������ҵ�����ɳ����ķ�չ,���ҹ�����ؽ����������⡣����ͨ�������Բݵؽ��������Ǹߺ��ݵ�����Ҫ�����÷�ʽ֮һ�����������еIJ�ʳ����̤����й���ȹ��̻��̼�ŷŲ�����ҪӰ�졣�ڷ���������������ʪ�ȡ�ֲ��Ⱥ����ɡ������л�̼�������������ֵ�ָ����ܻᷢ���仯,��Щ���ض���̬ϵͳ����������ҪӰ�졣������������̬ϵͳ����CO_2�Ĺ��̼��临��,����ǿ�Ⱥͷ�����ʷ�ۺ����ö���̬ϵͳ����CO_2���Ӱ�졣����֮����ܻ������ڷ����������͡����ڷ������̶Ըߺ��ݵ�CO_2�ŷŵ�Ӱ�켫�临��,��Ŀǰ,����������̬ϵͳ������������δ���һ�¹�ʶ����Ȼ���кܶ����Ƶ��о�,�����ò�ͬ����ǿ���ҳ�ʱ�������۲�ߺ��ݵ�CO_2�ŷŵ��о������б����������������ظ�ԭ��Ե�ĸߺ��ݵ鿪չ�˳�ʱ��ij����۲�,Ϊ������ظ�ԭ��Ե�ߺ��ݵ�̼�������Լ��ƶ������ķ��������ṩ�������ݡ�����ظ�ԭ��Ե�Ĵ�ʡ���Ӳ���Ǽ�������ݺ�ԭ�ؾ��ڿ�չ��Ϊ������ķ���ʵ�顣���о�����������ƽ��ʵ��,����ţ����ʵ��Ͳ��������ʵ�顣������ʵ���зֱ������˲�ͬ�ķ���ǿ��,���þ�̬��-����ɫ���ⶨCO_2Ũ��,��ȫ��̽����ظ�ԭ������Ե�ߺ��ݵ��ڲ�ͬ����ǿ���µ�CO_2�ŷ�ͨ��,��ͬ����ǿ�ȶ�CO_2�ŷ�ͨ���������Լ�����������ߺ��ݵ���̬ϵͳCO_2�ŷ�ͨ���Ĺ�ϵ���õ�����Ҫ����ͽ�������:(1)����ţ����ʵ����,���ֲ�ͬ����ǿ��,��:����������ȷ������жȷ������ضȷ�����ƽ�����ۻ��ŷ����ֱ�Ϊ:470.31,482.71,462.16��431.44 g C m-2 y-1,���ܵ�ƽ��ֵΪ461.66g C m-2 y-1;�ڲ��������ʵ����,���ֲ�ͬ����ǿ��,��:��ȷ������жȷ������ضȷ�����ƽ�����ۻ��ŷ����ֱ�Ϊ:487.66,430.11��434.78g C m-2y-1,���ܵ�ƽ��ֵΪ450.85g C m-2y-1��(2)������ʵ���о������������е�CO_2�ŷ�ͨ��ҪԶԶ���ڷ�������,���ֳ������Եļ�������������ţ����ʵ����,�������е�CO_2�ŷ�ͨ��ռ���ۻ�CO_2�ŷ�ͨ����68.02%~74.36%;�ڲ��������ʵ����,�������е�CO_2�ŷ�ͨ��ռ���ۻ�CO_2�ŷ�ͨ����69.34%~77.21%��(3)�ڱ��о��������ʵ�������õķ���ǿ���µĽ������:��ͬ����ǿ���¸ߺ��ݵ�CO_2�ŷ�ͨ�����������졣(4)������ʵ����,���Ǿ�����:�¶���CO_2�ŷ�ͨ������ع�ϵ,�������ۻ���ˮ����CO_2�ŷ�ͨ��������Ը���һЩ��
DOI:10.11920/xnmdzk.2016.05.009URL [��������: 1]
��ȫ��仯�ı�����,���ɹŲ�ԭ��̬ϵͳ�Ľ�ˮ���������ӻ�����.����,��ˮ���ĸı�Ը�����ݵ���̬ϵͳ̼������Ӱ�����ǻ�֪֮����.Ϊ��,���о�ͨ����ȫ��ˮ������,��չ�˽�ˮ�ݶ�ʵ��,̽������̬ϵͳ̼����(ecosystem carbon exchange)�潵ˮ���仯����Ӧ�켣.�������:������������ˮ����100mm���ӵ�500mm,����̬ϵͳ̼����(net ecosystem exchange,NEE)����̬ϵͳ����(ecosystem respiration,ER)����̬ϵͳ��������(gross ecosystem productivity,GEP)���������,��Щ���̾�����Ϊ���Եķ�������Ӧ.NEE��ER��GEP�ı��͵��Ӧ�Ľ�ˮ���ֱ�Ϊ350mm,200mm��275mm.����Ӧ�ʵı仯б�ʿ�,��ˮ������(��275 mm�Ĵ���)��ЧӦ����ǿ�ڽ�ˮ������(�� 275 mm�Ĵ���)��ЧӦ.������Դ����˵��,���Ž�ˮ���ĸı�,Ⱥ���������Dz�ԭ��̬ϵͳ̼��������ҪӰ������,���Խ���NEE��ER��GEP�����30.89����41.90����40.60��,��������ˮ���������¶�ֻ�ܽ���NEE��ER��GEP�����11.51����7.78����9.28��.�������Ϊ���������Ԥ��δ����ˮ�仯�����ɹ��´����Ͳ�ԭ��̬ϵͳ̼������Ӱ���ṩ������.
DOI:10.11920/xnmdzk.2016.05.009URL [��������: 1]
��ȫ��仯�ı�����,���ɹŲ�ԭ��̬ϵͳ�Ľ�ˮ���������ӻ�����.����,��ˮ���ĸı�Ը�����ݵ���̬ϵͳ̼������Ӱ�����ǻ�֪֮����.Ϊ��,���о�ͨ����ȫ��ˮ������,��չ�˽�ˮ�ݶ�ʵ��,̽������̬ϵͳ̼����(ecosystem carbon exchange)�潵ˮ���仯����Ӧ�켣.�������:������������ˮ����100mm���ӵ�500mm,����̬ϵͳ̼����(net ecosystem exchange,NEE)����̬ϵͳ����(ecosystem respiration,ER)����̬ϵͳ��������(gross ecosystem productivity,GEP)���������,��Щ���̾�����Ϊ���Եķ�������Ӧ.NEE��ER��GEP�ı��͵��Ӧ�Ľ�ˮ���ֱ�Ϊ350mm,200mm��275mm.����Ӧ�ʵı仯б�ʿ�,��ˮ������(��275 mm�Ĵ���)��ЧӦ����ǿ�ڽ�ˮ������(�� 275 mm�Ĵ���)��ЧӦ.������Դ����˵��,���Ž�ˮ���ĸı�,Ⱥ���������Dz�ԭ��̬ϵͳ̼��������ҪӰ������,���Խ���NEE��ER��GEP�����30.89����41.90����40.60��,��������ˮ���������¶�ֻ�ܽ���NEE��ER��GEP�����11.51����7.78����9.28��.�������Ϊ���������Ԥ��δ����ˮ�仯�����ɹ��´����Ͳ�ԭ��̬ϵͳ̼������Ӱ���ṩ������.
DOI:10.11829\j.issn.1001-0629.2015-0096URL [��������: 1]
�ݵ���̬ϵͳ��̼������ȫ��̼�����Ĵ�С����ҪӰ�졣������Ϊ�ݵ���㷺�����÷�ʽ֮һӰ���Ųݵص�̼����,���ȺͲ������ķ�����ʽ��ɲݵ��˻�,����ݵ�̼����ʧ��Χ������������Բݵ���̬ϵͳ��̼������Ӱ����Ҫ������ֲ��������������̼������Ӱ��,���Ķ��йصı��������������������ʾ,������ֲ�������Ⱥ��ṹ������ȶ����ص�Ӱ��,Χ�������Բݵ�ֲ������������������ЧӦ;������ֲ����������Ӱ��Ҳû��һ�µĽ��,����,������ֲ����������Ӱ�컹�������ǿ���йء�Χ���������������ͨ������;���Բݵ�����̼����������������Ӱ�졣��֮,Χ������������Բݵ���̬ϵͳ��̼������Ӱ���Ǹ��Ӷ�����,��Ҫ�Բݵ�̼�������й���,������صĻ��ƽ����о�,���������ڲݵع����������������ľ��ߡ�
DOI:10.11829\j.issn.1001-0629.2015-0096URL [��������: 1]
�ݵ���̬ϵͳ��̼������ȫ��̼�����Ĵ�С����ҪӰ�졣������Ϊ�ݵ���㷺�����÷�ʽ֮һӰ���Ųݵص�̼����,���ȺͲ������ķ�����ʽ��ɲݵ��˻�,����ݵ�̼����ʧ��Χ������������Բݵ���̬ϵͳ��̼������Ӱ����Ҫ������ֲ��������������̼������Ӱ��,���Ķ��йصı��������������������ʾ,������ֲ�������Ⱥ��ṹ������ȶ����ص�Ӱ��,Χ�������Բݵ�ֲ������������������ЧӦ;������ֲ����������Ӱ��Ҳû��һ�µĽ��,����,������ֲ����������Ӱ�컹�������ǿ���йء�Χ���������������ͨ������;���Բݵ�����̼����������������Ӱ�졣��֮,Χ������������Բݵ���̬ϵͳ��̼������Ӱ���Ǹ��Ӷ�����,��Ҫ�Բݵ�̼�������й���,������صĻ��ƽ����о�,���������ڲݵع����������������ľ��ߡ�
Effects of grazing on carbon and nitrogen in plants and soils in a semiarid desert grassland, China
1
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Canopy-level photosynthetic compensation after defoliation in a tropical understory palm
2001
ˮ�����������ֺ�������Ͳ�ԭ�˻�Ⱥ������������Ӱ��
1
2003
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
ˮ�����������ֺ�������Ͳ�ԭ�˻�Ⱥ������������Ӱ��
1
2003
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
�й��������۲ݵ��ԭ̼ˮͨ����ˮ������Ч���о�
1
2011
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
�й��������۲ݵ��ԭ̼ˮͨ����ˮ������Ч���о�
1
2011
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
��ݺʹ���é���������併������̬���ӹ�ϵ���о�
1
1900
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
��ݺʹ���é���������併������̬���ӹ�ϵ���о�
1
1900
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
Carbon dioxide fluxes over three great plains grasslands
1
2000
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
�̻���é��ԭ��ͬ�����ʶ�ֲ������ԺͲݵ���������Ӱ��
1
2007
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
�̻���é��ԭ��ͬ�����ʶ�ֲ������ԺͲݵ���������Ӱ��
1
2007
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
�������½��ݵ���̬ϵͳ̼Դ/���Ӱ��ģ���о�
2017
�������½��ݵ���̬ϵͳ̼Դ/���Ӱ��ģ���о�
2017
Χ����������ɽ���²ݵ��ԭ��̬ϵͳ̼������Ӱ��
2
2018
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C28">5�·ݶ̻���é��ԭֲ���ոտ�ʼ����, ��ˮ��, ��̬ϵͳ̼���ս���, NEE��Ҫ��ER����.�����ۻ��ķ���ЧӦʹ����ȷ������ضȷ�����������Խ�ʵ, ͨ�Խ���, ���³������������ڶ���, ��֮�������ڵĵ�������۽϶�, Ӱ����ֲ���ȶ�, ����Ⱥ��Ĺ�����ýϵ�, ��˳���������������̼�����������ڶ����������.6�·ݽ�ˮ������, �ٽ���ֲ��������, ��̬ϵͳ̼����������ǿ.������������ضȷ������������NEE.7��������������ƽ���¶���ߵ��·�, ˮ�ֳ�Ϊ����ֲ��������GEP����Ҫ������, 2014��7�����д���������Ϊ̼Դ, ��2015��7���������Ϊ����̼��.2016��7�½�ˮƵ������������, ��ӽ����������¶�, ������ˮ��ԼΪ2014�ꡢ2015���4��, NEE�ﵽȫ�����(-7.03 mol��m-2��s-1).8�����������ĸ߷���, 2014��8�¾�̼�����ӽ���ֵ, 2015����һ���ɺ������, 8�½�ˮΪ�������������, ����ֲ������������Ч��ˮ��, �ڷ���������, ֲ��ϡ�貢����ݻ�, ����ʪ�ȵ�, ֲ�������֮���̼��������, ������������Ϊ̼Դ.2016��8�²�ͬ�����ʴ�����NEE����Ϊ��̼����, �����岻��7��.����������ʢ�ڵ���, ���������¸�����������, �Һ������ʼӿ�, ����ϵ�����������������߶����, �ϸߵĸ�ϵ��������ٽ�CO2���ͷ�(
Χ����������ɽ���²ݵ��ԭ��̬ϵͳ̼������Ӱ��
2
2018
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C28">5�·ݶ̻���é��ԭֲ���ոտ�ʼ����, ��ˮ��, ��̬ϵͳ̼���ս���, NEE��Ҫ��ER����.�����ۻ��ķ���ЧӦʹ����ȷ������ضȷ�����������Խ�ʵ, ͨ�Խ���, ���³������������ڶ���, ��֮�������ڵĵ�������۽϶�, Ӱ����ֲ���ȶ�, ����Ⱥ��Ĺ�����ýϵ�, ��˳���������������̼�����������ڶ����������.6�·ݽ�ˮ������, �ٽ���ֲ��������, ��̬ϵͳ̼����������ǿ.������������ضȷ������������NEE.7��������������ƽ���¶���ߵ��·�, ˮ�ֳ�Ϊ����ֲ��������GEP����Ҫ������, 2014��7�����д���������Ϊ̼Դ, ��2015��7���������Ϊ����̼��.2016��7�½�ˮƵ������������, ��ӽ����������¶�, ������ˮ��ԼΪ2014�ꡢ2015���4��, NEE�ﵽȫ�����(-7.03 mol��m-2��s-1).8�����������ĸ߷���, 2014��8�¾�̼�����ӽ���ֵ, 2015����һ���ɺ������, 8�½�ˮΪ�������������, ����ֲ������������Ч��ˮ��, �ڷ���������, ֲ��ϡ�貢����ݻ�, ����ʪ�ȵ�, ֲ�������֮���̼��������, ������������Ϊ̼Դ.2016��8�²�ͬ�����ʴ�����NEE����Ϊ��̼����, �����岻��7��.����������ʢ�ڵ���, ���������¸�����������, �Һ������ʼӿ�, ����ϵ�����������������߶����, �ϸߵĸ�ϵ��������ٽ�CO2���ͷ�(
���˸ɺ����������ֺ�������ݲ�ԭ����̬ϵͳ̼��������
1
2006
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
���˸ɺ����������ֺ�������ݲ�ԭ����̬ϵͳ̼��������
1
2006
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Temperature response of soil respiration in a Chinese pine plantation: Hysteresis and seasonal vs. Diel Q10
1
2013
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
Effects of grazing on CO2 balance in a semiarid steppe: Field observations and modeling
2
2013
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C29">�о������ڸɺ��ش�, �ݵص�̼��֧�ܽ�ˮ�������Լ(
Review: Time lag between photosynthesis and carbon dioxide efflux from soil: A review of mechanisms and controls
1
2010
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
����ǿ�ȶ���ظ�ԭ�ߺ��ݵ龻��̬ϵͳ��������Ӱ��
1
2014
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
����ǿ�ȶ���ظ�ԭ�ߺ��ݵ龻��̬ϵͳ��������Ӱ��
1
2014
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Effects of warming and increased precipitation on net ecosystem productivity: A long-term manipulative experiment in a semiarid grassland
1
2017
... id="C29">�о������ڸɺ��ش�, �ݵص�̼��֧�ܽ�ˮ�������Լ(
���ֺ�������ݲ�ԭȺ��������������Ӱ�����ӵ��о�
1
2000
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
���ֺ�������ݲ�ԭȺ��������������Ӱ�����ӵ��о�
1
2000
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
�����¶Ⱥ�ˮ�ֶԿ�����é��ԭ��̬ϵͳ̼ͨ����Ӱ���̽
1
2011
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
�����¶Ⱥ�ˮ�ֶԿ�����é��ԭ��̬ϵͳ̼ͨ����Ӱ���̽
1
2011
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
Inter-annual variability in carbon dioxide exchange of an oak/grass savanna and open grassland in California
1
2007
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Management and land use change effects on soil carbon in northern China��s grasslands: A synthesis
1
2011
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe
1
2008
... id="C29">�о������ڸɺ��ش�, �ݵص�̼��֧�ܽ�ˮ�������Լ(
Compensatory growth of the African dwarf shrub
2000
Photosynthesis, transpiration and water use efficiency of four plant species with grazing intensities in Hunshandake Sandland, China
2007
Precipitation regulates the response of net ecosystem CO2 exchange to environmental variation on United States rangelands
1
2010
... id="C28">5�·ݶ̻���é��ԭֲ���ոտ�ʼ����, ��ˮ��, ��̬ϵͳ̼���ս���, NEE��Ҫ��ER����.�����ۻ��ķ���ЧӦʹ����ȷ������ضȷ�����������Խ�ʵ, ͨ�Խ���, ���³������������ڶ���, ��֮�������ڵĵ�������۽϶�, Ӱ����ֲ���ȶ�, ����Ⱥ��Ĺ�����ýϵ�, ��˳���������������̼�����������ڶ����������.6�·ݽ�ˮ������, �ٽ���ֲ��������, ��̬ϵͳ̼����������ǿ.������������ضȷ������������NEE.7��������������ƽ���¶���ߵ��·�, ˮ�ֳ�Ϊ����ֲ��������GEP����Ҫ������, 2014��7�����д���������Ϊ̼Դ, ��2015��7���������Ϊ����̼��.2016��7�½�ˮƵ������������, ��ӽ����������¶�, ������ˮ��ԼΪ2014�ꡢ2015���4��, NEE�ﵽȫ�����(-7.03 mol��m-2��s-1).8�����������ĸ߷���, 2014��8�¾�̼�����ӽ���ֵ, 2015����һ���ɺ������, 8�½�ˮΪ�������������, ����ֲ������������Ч��ˮ��, �ڷ���������, ֲ��ϡ�貢����ݻ�, ����ʪ�ȵ�, ֲ�������֮���̼��������, ������������Ϊ̼Դ.2016��8�²�ͬ�����ʴ�����NEE����Ϊ��̼����, �����岻��7��.����������ʢ�ڵ���, ���������¸�����������, �Һ������ʼӿ�, ����ϵ�����������������߶����, �ϸߵĸ�ϵ��������ٽ�CO2���ͷ�(
ˮ�����Ӷ����ɹŵ��Ͳ�ԭ������̬ϵͳ̼������Ӱ��
1
2014
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
ˮ�����Ӷ����ɹŵ��Ͳ�ԭ������̬ϵͳ̼������Ӱ��
1
2014
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
Carbon dioxide fluxes in a spatially and temporally heterogeneous temperate grassland
1
2006
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
���۲ݵ���̬ϵͳCO2�����Ե���������ˮ���Ӽ���������Ӧ����
1
2016
... id="C29">�о������ڸɺ��ش�, �ݵص�̼��֧�ܽ�ˮ�������Լ(
���۲ݵ���̬ϵͳCO2�����Ե���������ˮ���Ӽ���������Ӧ����
1
2016
... id="C29">�о������ڸɺ��ش�, �ݵص�̼��֧�ܽ�ˮ�������Լ(
�̻���é(Stipa breviflora)ҶƬ�� 13C�������ʵ���Ӧ
2017
�̻���é(Stipa breviflora)ҶƬ�� 13C�������ʵ���Ӧ
2017
�����Ϳ��Ѷ��´����Ͳ�ԭCO2ͨ����Ӱ��
2
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
�����Ϳ��Ѷ��´����Ͳ�ԭCO2ͨ����Ӱ��
2
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
�̻���é��Į��ԭֲ��Ⱥ�������Բ�ͬ������ˮƽ����Ӧ
2000
�̻���é��Į��ԭֲ��Ⱥ�������Բ�ͬ������ˮƽ����Ӧ
2000
����Դ�����˹��ݵص���̬ϵͳCO2���������ܳ�������������Ӱ������
1
2010
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
����Դ�����˹��ݵص���̬ϵͳCO2���������ܳ�������������Ӱ������
1
2010
... id="C30">��ˮͨ��Ӱ�������¶ȡ�������ˮ����Ӱ����̬ϵͳ̼����.�Ƚ�3����NEE��GEP�Ĺ�ϵ���Կ���, ��ˮ���ӿ�����ǿNEE�������¶ȡ�ʪ�ȵ������.�о�����ֲ��������ٶȡ���������Լ������������¶Ⱥ�ˮ�������е�, ���, �����¶Ⱥ�����ʪ�ȹ�ͬ������̬ϵͳCO2ͨ����ֵı���(
ģ�����º͵������ӶԻ�Į��ԭ��̬ϵͳ̼������Ӱ��
1
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
ģ�����º͵������ӶԻ�Į��ԭ��̬ϵͳ̼������Ӱ��
1
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
���ҶƬ���彻���������¶Ⱥ�����ˮ�ֵ���Ӧ
1
2004
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
���ҶƬ���彻���������¶Ⱥ�����ˮ�ֵ���Ӧ
1
2004
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
�̻���é��Į��ԭ��̬ϵͳ̼�����Բ�ͬ�����ʵ���Ӧ
2
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C28">5�·ݶ̻���é��ԭֲ���ոտ�ʼ����, ��ˮ��, ��̬ϵͳ̼���ս���, NEE��Ҫ��ER����.�����ۻ��ķ���ЧӦʹ����ȷ������ضȷ�����������Խ�ʵ, ͨ�Խ���, ���³������������ڶ���, ��֮�������ڵĵ�������۽϶�, Ӱ����ֲ���ȶ�, ����Ⱥ��Ĺ�����ýϵ�, ��˳���������������̼�����������ڶ����������.6�·ݽ�ˮ������, �ٽ���ֲ��������, ��̬ϵͳ̼����������ǿ.������������ضȷ������������NEE.7��������������ƽ���¶���ߵ��·�, ˮ�ֳ�Ϊ����ֲ��������GEP����Ҫ������, 2014��7�����д���������Ϊ̼Դ, ��2015��7���������Ϊ����̼��.2016��7�½�ˮƵ������������, ��ӽ����������¶�, ������ˮ��ԼΪ2014�ꡢ2015���4��, NEE�ﵽȫ�����(-7.03 mol��m-2��s-1).8�����������ĸ߷���, 2014��8�¾�̼�����ӽ���ֵ, 2015����һ���ɺ������, 8�½�ˮΪ�������������, ����ֲ������������Ч��ˮ��, �ڷ���������, ֲ��ϡ�貢����ݻ�, ����ʪ�ȵ�, ֲ�������֮���̼��������, ������������Ϊ̼Դ.2016��8�²�ͬ�����ʴ�����NEE����Ϊ��̼����, �����岻��7��.����������ʢ�ڵ���, ���������¸�����������, �Һ������ʼӿ�, ����ϵ�����������������߶����, �ϸߵĸ�ϵ��������ٽ�CO2���ͷ�(
�̻���é��Į��ԭ��̬ϵͳ̼�����Բ�ͬ�����ʵ���Ӧ
2
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
... id="C28">5�·ݶ̻���é��ԭֲ���ոտ�ʼ����, ��ˮ��, ��̬ϵͳ̼���ս���, NEE��Ҫ��ER����.�����ۻ��ķ���ЧӦʹ����ȷ������ضȷ�����������Խ�ʵ, ͨ�Խ���, ���³������������ڶ���, ��֮�������ڵĵ�������۽϶�, Ӱ����ֲ���ȶ�, ����Ⱥ��Ĺ�����ýϵ�, ��˳���������������̼�����������ڶ����������.6�·ݽ�ˮ������, �ٽ���ֲ��������, ��̬ϵͳ̼����������ǿ.������������ضȷ������������NEE.7��������������ƽ���¶���ߵ��·�, ˮ�ֳ�Ϊ����ֲ��������GEP����Ҫ������, 2014��7�����д���������Ϊ̼Դ, ��2015��7���������Ϊ����̼��.2016��7�½�ˮƵ������������, ��ӽ����������¶�, ������ˮ��ԼΪ2014�ꡢ2015���4��, NEE�ﵽȫ�����(-7.03 mol��m-2��s-1).8�����������ĸ߷���, 2014��8�¾�̼�����ӽ���ֵ, 2015����һ���ɺ������, 8�½�ˮΪ�������������, ����ֲ������������Ч��ˮ��, �ڷ���������, ֲ��ϡ�貢����ݻ�, ����ʪ�ȵ�, ֲ�������֮���̼��������, ������������Ϊ̼Դ.2016��8�²�ͬ�����ʴ�����NEE����Ϊ��̼����, �����岻��7��.����������ʢ�ڵ���, ���������¸�����������, �Һ������ʼӿ�, ����ϵ�����������������߶����, �ϸߵĸ�ϵ��������ٽ�CO2���ͷ�(
�̻���é��Į��ԭ��ͬ�����ʵ�����������ֲ������������Ĺ�ϵ
2015
�̻���é��Į��ԭ��ͬ�����ʵ�����������ֲ������������Ĺ�ϵ
2015
Χ������������������Ǹ�ԭ��̿��CO2��CH4�ŷŵ�Ӱ��
1
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Χ������������������Ǹ�ԭ��̿��CO2��CH4�ŷŵ�Ӱ��
1
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem
1
2007
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
��ͬ����ǿ�ȶԸߺ��ݵ�CO2�ŷ�ͨ����Ӱ��
1
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
��ͬ����ǿ�ȶԸߺ��ݵ�CO2�ŷ�ͨ����Ӱ��
1
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
��ˮ���仯�����ɹ��´����Ͳ�ԭ��̬ϵͳ̼������Ӱ��
1
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
��ˮ���仯�����ɹ��´����Ͳ�ԭ��̬ϵͳ̼������Ӱ��
1
2016
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Χ������������Բݵ���̬ϵͳ��̼������Ӱ��
1
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Χ������������Բݵ���̬ϵͳ��̼������Ӱ��
1
2015
... id="C6">�ݵ���̬ϵͳ��̼��֧ƽ����Ҫ����̬ϵͳ����������������֮���ƽ��.��̬ϵͳ�ܳ���������(GEP)����̬ϵͳ����(ER)��������̬ϵͳ��CO2���ջ��ͷŵ�����.��̬ϵͳ��̼����(NEE)Ϊ��ֵ������̬ϵͳ��̼�ͷ�(��̼Դ), NEEΪ��ֵ������̬ϵͳ��̼����(��̼��).����Χ����̬ϵͳ̼�������о���Ҫ��չ�ڵ��к���ԭ���߲ݲ�ԭ���ȴ�ϡ����ԭ(
Copyright © 2021 ��Ȩ���� ��ֲ����̬ѧ�����༭��
��ַ: ������ɽ������20��, �ʱ�: 100093
Tel.: 010-62836134, 62836138;��Fax: 010-82599431;��E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
������: ��ICP��16067583��-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}