Effects of fencing on ecosystem carbon exchange at meadow steppe in the northern slope of the Tianshan Mountains
HU Yi,1,2, ZHU Xin-Ping1,2, JIA Hong-Tao,1,2,*, HAN Dong-Liang1,2, HU Bao-An1,2, LI Dian-Peng11 College of Grassland and Environmental Sciences, Xinjiang Agricultural University, ürümqi 830052, China; 2 Xinjiang Key Laboratory of Soil and Plant Ecological Processes, ürümqi 830052, China;
SupportedbytheStrategicPriorityResearchProgramoftheChineseAcademyofSciences(XDA05050405) the National Natural Science Foundation of China(31560171) Innovation Plan of Postgraduate Education in Xinjiang Uygur Autonomous region of China.(XJGRI2014084)
Abstract Aims The carbon exchange between ecosystems and the atmosphere and its response to environmental factors is the focus of current research. The aim of this study was to examine the effects of fencing on ecosystem carbon exchange at meadow steppe in the northern slope of Tianshan Mountains.Methods The static box method with a LI-840 CO2/H2O infrared analyzer was used to evaluate daily and seasonal changes of ecosystem carbon exchange and their relationship with environmental factors in the inside fence and outside fence after 9 years fencing.Important findings We found the ecosystem carbon exchange inside the fence was significantly (p < 0.05) higher than that in outside the fence. The ecosystem carbon exchange had obvious daily and seasonal variation both in inside and outside the fence, which showed a unimodal curve during the plant growing season. The minimum net ecosystem CO2 exchange (NEE) in the inside and outside of the fences were -7.62 and -6.63 μmol·m-2·s-1, respectively; the maximum ecosystem respiration (ER) were 8.55 and 7.04 μmol·m-2·s-1, respectively; and the maximum gross ecosystem productivity (GEP) were -14.66 and -13.89 μmol·m-2·s-1, respectively. Due to the protection of fence, the vegetation in the fence was flourished with higher photosynthesis, and thus resulted in lower NEE. Meanwhile, organic carbon input enhanced ecosystem respiration. Besides, the ecosystem carbon exchange significantly correlated with the air temperature and soil temperature of 0 to 10 cm depth, and the correlation with the air temperature was higher than soil temperature of 0 to 10 cm depth. Also, the correlation in the inside of the fence was higher than that in the outside of the fence. Ecosystem carbon exchange had correlation with soil water content, but the correlation was slightly lower than that with soil temperature. Keywords:fencing;meadow steppe;ecosystem carbon exchange
PDF (1135KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 胡毅, 朱新萍, 贾宏涛, 韩东亮, 胡保安, 李典鹏. 围栏封育对天山北坡草甸草原生态系统碳交换的影响. 植物生态学报, 2018, 42(3): 372-381 doi:10.17521/cjpe.2016.0049 HU Yi, ZHU Xin-Ping, JIA Hong-Tao, HAN Dong-Liang, HU Bao-An, LI Dian-Peng. Effects of fencing on ecosystem carbon exchange at meadow steppe in the northern slope of the Tianshan Mountains. Chinese Journal of Plant Ecology, 2018, 42(3): 372-381 doi:10.17521/cjpe.2016.0049
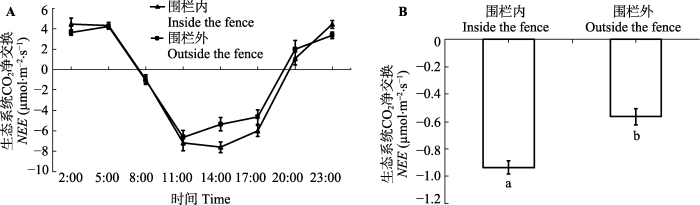
Fig. 1The daily dynamic (A) and daily average value (B) of the net ecosystem CO2 exchange (NEE) in inside and outside the fence (mean ± SE). The different lowercase letters represent significant differences at the 0.05 level.
Fig. 2The daily dynamic (A) and daily average value (B) of the ecosystem respiration (ER) in inside and outside the fence (mean ± SE). The same lowercase letters represent no significant differences at the 0.05 level.
Fig. 3The daily dynamics (A) and daily average value (B) of the gross ecosystem productivity (GEP) in inside and outside the fence, respectively (mean ± SE). The same lowercase letters represent no significant differences at the 0.05 level.
Fig. 4The seasonal dynamic (A) and average value (B) of the the net ecosystem CO2 exchange (NEE) in inside and outside the fence, respectively (mean ± SE). The different lowercase letters represent significant differences at the 0.05 level.
Fig. 5The seasonal dynamic (A) and month average value (B) of the ecosystem respiration (ER) in inside and outside the fence, respectively (mean ± SE). The same lowercase letters represent no significant differences at the 0.05 level.
Fig. 6The seasonal dynamic (A) and average value (B) of the GEP in inside and outside the fence, respectively (mean ± SE). The different lowercase letters represent significant differences at the 0.05 level.
Table 1 表1 表1围栏内外草甸草原生态系统CO2净交换速率(NEE)、生态系统呼吸(ER)、生态系统总初级生产力(GEP)与大气温度和0-10 cm土壤温度相关性 Table 1The inside and outside the fence correlation coefficients between the net ecosystem CO2 exchange (NEE), ecosystem respiration (ER), gross ecosystem the productivity (GEP) and atmospheric temperature and 0-10 cm soil temperature in meadow steppe
Table 2 表2 表2围栏内外草甸草原生态系统CO2净交换速率(NEE)、生态系统呼吸(ER)、生态系统总初级生产力(GEP)与温度和土壤含水量的回归分析 Table 2The regression analysis of the NEE (net ecosystem CO2 exchange), ER (ecosystem respiration), GEP (gross ecosystem the productivity) and temperature and soil water content in inside and outside the fence
BachmanS, Heisler-WhiteJL, PendallE, WilliamsD, MorganJA, NewcombJ ( 2010). Elevated carbon dioxide alters impacts of precipitation pulses on ecosystem photosynthesis and respiration in a semi-arid grassland Oecologia, 62, 791-802. DOI:10.1007/s00442-009-1511-xURLPMID:19943173 [本文引用: 1] Predicting net C balance under future global change scenarios requires a comprehensive understanding of how ecosystem photosynthesis (gross primary production; GPP) and respiration (Re) respond to elevated atmospheric [CO 2 ] and altered water availability. We measured net ecosystem exchange of CO 2 (NEE), GPP and Re under ambient and elevated [CO 2 ] in a northern mixed-grass prairie (Wyoming, USA) during dry intervals and in response to simulated precipitation pulse events. Elevated [CO 2 ] resulted in higher rates of both GPP and Re across the 2006 growing season, and the balance of these two fluxes (NEE) accounted for cumulative growing season C uptake (6114.402±028.302g02C02m 612 ). Despite lower GPP and Re, experimental plots under ambient [CO 2 ] had greater cumulative uptake (6136.202±028.202g02C02m 612 ) than plots under elevated [CO 2 ]. Non-irrigated control plots received 50% of average precipitation during the drought of 2006, and had near-zero NEE (1.902±026.402g02C02m 612 ) for the growing season. Elevated [CO 2 ] extended the magnitude and duration of pulse-related increases in GPP, resulting in a significant [CO 2 ] treatment by pulse day interaction, demonstrating the potential for elevated [CO 2 ] to increase the capacity of this ecosystem to respond to late-season precipitation. However, stimulation of Re throughout the growing season under elevated [CO 2 ] reduced net C uptake compared to plots under ambient [CO 2 ]. These results indicate that although elevated [CO 2 ] stimulates gross rates of ecosystem C fluxes, it does not necessarily enhance net C uptake, and that C cycle responses in semi-arid grasslands are likely to be more sensitive to changes in precipitation than atmospheric [CO 2 ].
BaldocchiD ( 1997). Measuring and modeling carbon dioxide and water vapor exchange over a temperate broad-leafed forest during the 1985 summer drought Plant, Cell & Environment, 20, 1108-1122. DOI:10.1046/j.1365-3040.1997.d01-147.xURL [本文引用: 1] ABSTRACT Forests in the south-eastern United States experienced a prolonged dry spell and above-normal temperatures during the 1995 growing season. During this episode, nearly continuous, eddy covariance measurements of carbon dioxide and water vapour fluxes were acquired over a temperate, hardwood forest. These data are used to examine how environmental factors and accumulating soil moisture deficits affected the diurnal pattern and magnitude of canopy-scale carbon dioxide and water vapour fluxes. The field data are also used to test an integrative leaf-to-canopy scaling model (CANOAK), which uses micrometeorological and physiological theory, to calculate mass and energy fluxes. When soil moisture was ample in the spring, peak rates of net ecosystem CO 2 exchange ( N F ) occurred around midday and exceeded 20 μmol m 612 s 611 . Rates of N K were near optimal when air temperature ranged between 22 and 25°C. The accumulation of soil moisture deficits and a co-occurrence of high temperatures caused peak rates of daytime carbon dioxide uptake to occur earlier in the morning. High air temperatures and soil moisture deficits were also correlated with a dramatic reduction in the magnitude of N E . On average, the magnitude of N E decreased from 20 to 7 μmol m 612 s 611 as air temperature increased from 24 to 30°C and the soil dried. The CANAOK model yielded accurate estimates of canopy-scale carbon dioxide and water vapour fluxes when the forest had an ample supply of soil moisture. During the drought and heat spell, a cumulative drought index was needed to adjust the proportionality constant of the stomatal conductance model to yield accurate estimates of canopy CO 2 exchange. The adoption of the drought index also enabled the CANOAK model to give improved estimates of evaporation until midday. On the other hand, the scheme failed to yield accurate estimates of evaporation during the afternoon.
BlackTA, HartogGD, NeumannHH, BlankenPD, YangPC, RussellC, NesicZ, LeeX, ChenSG, StaeblerR, NovakMD ( 1996). Annual cycles of water vapour and carbon dioxide fluxes in and above a boreal aspen forest Global Change Biology, 2, 219-229. DOI:10.1111/j.1365-2486.1996.tb00074.xURL [本文引用: 1] Abstract Water vapour and CO 2 fluxes were measured using the eddy correlation method above and below the overstorey of a 21-m tall aspen stand in the boreal forest of central Saskatchewan as part of the Boreal Ecosystem-Atmosphere Study (BOREAS). Measurements were made at the 39.5-m and 4-m heights using 3-dimensional sonic anemometers (Kaijo-Denki and Solent, respectively) and closed-path gas analysers (LI-COR 6262) with 6-m and 4.7-m long heated sampling tubing, respectively. Continuous measurements were made from early October to mid-November 1993 and from early February to late-September 1994. Soil CO 2 flux (respiration) was measured using a LI-COR 6000-09 soil chamber and soil evaporation was measured using Iysimetry. The leaf area index of the aspen and hazelnut understorey reached 1.8 and 3.3, respectively. The maximum daily evapotranspiration ( E ) rate was 5–6 mm d 611 . Following leaf-out the hazelnut and soil accounted for 22% of the forest E . The estimated total E was 403 mm for 1994. About 88% of the precipitation in 1994 was lost as evapotranspiration. During the growing season, the magnitude of half-hourly eddy fluxes of CO 2 from the atmosphere into the forest reached 1.2 mg CO 2 m 612 s 611 (33 μmol C m 612 s 611 ) during the daytime. Downward eddy fluxes at the 4-m height were observed when the hazelnut was growing rapidly in June and July. Under well-ventilated night-time conditions, the eddy fluxes of CO 2 above the aspen and hazelnut, corrected for canopy storage, increased exponentially with soil temperature at the 2-cm depth. Estimates of daytime respiration rates using these relationships agreed well with soil chamber measurements. During the 1994 growing season, the cumulative net ecosystem exchange (NEE) was -3.5 t C ha 611 y 611 (a net gain by the system). For 1994, cumulative NEE, ecosystem respiration (R) and gross ecosystem photosynthesis ( GEP = R - NEE ) were estimated to be -1.3, 8.9 and 10.2 t C ha 611 y 611 respectively. Gross photosynthesis of the hazelnut was 32% of GEP.
ChenJ, ShiWY, CaoJJ ( 2014). Effects of grazing on ecosystem CO2 exchange in a meadow grassland on the Tibetan Plateau during the growing season Environmental Management, 55, 347-359. DOI:10.1007/s00267-014-0390-zURLPMID:25355630 [本文引用: 1] Abstract Effects of human activity on ecosystem carbon fluxes (e.g., net ecosystem exchange (NEE), ecosystem respiration (R(eco)), and gross ecosystem exchange (GEE)) are crucial for projecting future uptake of CO2 in terrestrial ecosystems. However, how ecosystem that carbon fluxes respond to grazing exclusion is still under debate. In this study, a field experiment was conducted to study the effects of grazing exclusion on R(eco), NEE, and GEE with three treatments (free-range grazing (FG) and grazing exclusion for 3 and 5 years (GE3 and GE5, respectively)) in a meadow grassland on the Tibetan Plateau. Our results show that grazing exclusion significantly increased NEE by 47.37 and 15.84%, and R eco by 33.14 and 4.29% under GE3 and GE5 plots, respectively, although carbon sinks occurred in all plots during the growing season, with values of 192.11, 283.12, and 222.54 g C m(-2) for FG, GE3, and GE5, respectively. Interestingly, grazing exclusion increased temperature sensitivity (Q10) of R eco with larger increases at the beginning and end of growing season (i.e., May and October, respectively). Soil temperature and soil moisture were key factors on controlling the diurnal and seasonal variations of R(eco), NEE, and GEE, with soil temperature having a stronger influence. Therefore, the combined effects of grazing and temperature suggest that grazing should be taken into consideration in assessing global warming effects on grassland ecosystem CO2 exchange.
DongG ( 2011). Carbon and Water Fluxes and Water Use Efficiency of the Songnen Meadow Steppe in North East China. PhD dissertation, Northeast Normal University, Changchun. URL [本文引用: 1] 陆地生态系统与大气间的碳水交换是物质和能量循环的关键过程,开展植被与大气间碳、水通量的长期观测对人们深入研究和理解全球变化与陆地生态系统碳水循环的相互关系及其控制机理具有重要的意义。本研究以中国东北松嫩草甸草原为研究对象,以涡度相关方法为主要技术手段,以长岭野外实验站四年的碳、水通量及微气象数据为依托,客观评价了涡度相关技术在草甸草原碳、水循环研究中的适用性,完成了通量数据的质量评价和控制,集中探讨了该草地生态系统净碳交换和水分蒸散在不同时间尺度上的变化特征及其驱动机制,对比分析了能量和水量平衡的年际收支差... 董刚 ( 2011). 中国东北松嫩草甸草原碳水通量及水分利用效率研究 博士学位论文, 东北师范大学, 长春. URL [本文引用: 1] 陆地生态系统与大气间的碳水交换是物质和能量循环的关键过程,开展植被与大气间碳、水通量的长期观测对人们深入研究和理解全球变化与陆地生态系统碳水循环的相互关系及其控制机理具有重要的意义。本研究以中国东北松嫩草甸草原为研究对象,以涡度相关方法为主要技术手段,以长岭野外实验站四年的碳、水通量及微气象数据为依托,客观评价了涡度相关技术在草甸草原碳、水循环研究中的适用性,完成了通量数据的质量评价和控制,集中探讨了该草地生态系统净碳交换和水分蒸散在不同时间尺度上的变化特征及其驱动机制,对比分析了能量和水量平衡的年际收支差...
DuanXN, WangXK, FengZZ, OuyangZY ( 2005). Study of net ecosystem exchange for seedling stage of spring wheat ecosystem in Hetao irrigation district, Inner Mongolia Journal of Environmental Science, 25, 166-171. [本文引用: 1]
FanLL, XieJP, MaJ, WangZY, ZhaoHM ( 2013). Variation characteristics of herb layer carbon flux and soil respiration in southern edge of Jungggar Basin, Northwest China Chinese Journal of Ecology, 32, 2567-2573. URL [本文引用: 2] 利用LI-840以及LI-8100土壤呼吸测量仪,对准噶尔盆地南缘草本层片碳通量以及沙丘不同部位的土壤呼吸速率进行了测量,探讨了草本层片碳通量季节动态以及日过程的变化特征,沙丘不同部位土壤呼吸速率的年变化特征以及环境因子对草本层片碳通量以及土壤呼吸速率的影响。结果表明:草本层片碳通量不仅在日尺度上呈现单峰的变化趋势,在季节尺度上也呈现相同的变化趋势。生长旺盛季草本层碳通量日过程变化幅度较大,草本层片净碳交换(NEE)从-2.5~0.02μmol·m~(~(-2))·s~(-1),草本层总生产力(GEP)则从-4.4~-1.1μmol·m~(-2)·s~(-1),而9月份以后则基本维持在0附近。通过分析碳通量与环境因子的关系,草本层片碳通量与土壤温度呈正相关关系,草本层土壤呼吸(ER)与光合有效辐射呈正相关关系,而草本层GEP与光合有效辐射则呈负相关关系。沙丘不同部位(沙丘顶部、中部、底部)土壤呼吸速率年动态变化为双峰型,且不同坡位之间土壤呼吸速率差异显著。 范连连, 谢继萍, 马健, 王忠媛, 赵红梅 ( 2013). 准噶尔盆地南缘草本层碳通量及土壤呼吸的变化特征 生态学杂志, 32, 2567-2573. URL [本文引用: 2] 利用LI-840以及LI-8100土壤呼吸测量仪,对准噶尔盆地南缘草本层片碳通量以及沙丘不同部位的土壤呼吸速率进行了测量,探讨了草本层片碳通量季节动态以及日过程的变化特征,沙丘不同部位土壤呼吸速率的年变化特征以及环境因子对草本层片碳通量以及土壤呼吸速率的影响。结果表明:草本层片碳通量不仅在日尺度上呈现单峰的变化趋势,在季节尺度上也呈现相同的变化趋势。生长旺盛季草本层碳通量日过程变化幅度较大,草本层片净碳交换(NEE)从-2.5~0.02μmol·m~(~(-2))·s~(-1),草本层总生产力(GEP)则从-4.4~-1.1μmol·m~(-2)·s~(-1),而9月份以后则基本维持在0附近。通过分析碳通量与环境因子的关系,草本层片碳通量与土壤温度呈正相关关系,草本层土壤呼吸(ER)与光合有效辐射呈正相关关系,而草本层GEP与光合有效辐射则呈负相关关系。沙丘不同部位(沙丘顶部、中部、底部)土壤呼吸速率年动态变化为双峰型,且不同坡位之间土壤呼吸速率差异显著。
FlanaganLB, JohnsonBG ( 2005). Interacting effects of temperature, soil moisture and plant biomass production on ecosystem respiration in a northern temperate grassland Agricultural and Forest Meteorology, 130, 237-253. DOI:10.1016/j.agrformet.2005.04.002URL [本文引用: 1] Chamber measurements of total ecosystem respiration (TER) in a native Canadian grassland ecosystem were made during two study years with different precipitation. The growing season (April–September) precipitation during 2001 was less than one-half of the 30-year mean (1971–2000), while 2002 received almost double the normal growing season precipitation. As a consequence soil moisture remained higher in 2002 than 2001 during most of the growing season and peak aboveground biomass production (253.9 g m 612) in 2002 was 60% higher than in 2001. Maximum respiration rates were approximately 9 μmol m 612 s 611 in 2002 while only approximately 5 μmol m 612 s 611 in 2001. Large diurnal variation in TER, which occurred during times of peak biomass and adequate soil moisture, was primarily controlled by changes in temperature. The temperature sensitivity coefficient ( Q 10) for ecosystem respiration was on average 1.83 ± 0.08, and it declined in association with reductions in soil moisture. Approximately 94% of the seasonal and interannual variation in R 10 (standardized rate of respiration at 10 °C) data was explained by the interaction of changes in soil moisture and aboveground biomass, which suggested that plant aboveground biomass was good proxy for accounting for variations in both autotrophic and heterotrophic capacity for respiration. Soil moisture was the dominant environmental factor that controlled seasonal and interannual variation in TER in this grassland, when variation in temperature was held constant. We compared respiration rates measured with chambers and that determined from nighttime eddy covariance (EC) measurements. Respiration rates measured by both techniques showed very similar seasonal patterns of variation in both years. When TER was integrated over the entire growing season period, the chamber method produced slightly higher values than the EC method by approximately 4.5% and 13.6% during 2001 and 2002, respectively, much less than the estimated uncertainty for both measurement techniques. The two methods for calculating respiration had only minor effects on the seasonal-integrated estimates of net ecosystem CO 2 exchange and ecosystem gross photosynthesis.
FuYL, YuGR, WangYF, LiZQ, HaoYB ( 2006). Effect of water stress on ecosystem photosynthesis and respiration of a Leymus chinensis steppe in Inner Mongolia. Science in China: Series D, Earth Sciences, 36(Suppl. I), 183-193. URL [本文引用: 1] 在全球气候变化条件下,全球陆地区域的降水量及其空间/时间分布模式将发生明显改变,这种变 化将对中国的陆地生态系统,尤其是干旱地区的温带草地生态系统碳收支产生重要影响.作为中国陆地生态系统通量观测研究网络(ChinaFLUX)的一部 分,本研究利用2003至2004年在内蒙古羊草草原的涡度相关通量观测数据,初步探讨了水分胁迫对羊草草原生态系统光合和呼吸作用的影响.研究发现温度 和水分是影响该生态系统在生长季(5~9月)的光合和呼吸作用的主要因子.在土壤水分适宜条件下,生态系统呼吸对温度变化的敏感性较大 (Q10=2.0),而当土壤含水量降低时生态系统呼吸对温度的敏感性明显降低(Q10=1.6).高温和干旱会显著降低生态系统的光合生产力.生长季的 降水量及其季节分配模式的不同对草地生态系统的生物物候有明显的影响,在2003年6月初生态系统就开始净吸收CO2、在7月初出现最大净生态系统CO2 吸收量,而因随后发生的干旱和高温胁迫使其在8月就提早进入休眠期;2004年春季的严重干旱导致生态系在7月初才开始净吸收CO2,并在降水丰沛气温适 宜的8月出现最大CO2吸收量,水分胁迫导致该草地植物生长发育比2003年推迟1个多月.观测结果显示该草地生态系统在2004年5~9月比2003年 同期多吸收30gCO2·m^-2.我国温带草原植被类型复杂多样,要准确估算我国草地生态系统碳收支还需要开展更多的长期联合观带研究. 伏玉玲, 于贵瑞, 王艳芬, 李正泉, 郝彦宾 ( 2006). 水分胁迫对内蒙古羊草草原生态系统光合和呼吸作用的影响 中国科学: D辑地球科学, 36(增刊I), 183-193. URL [本文引用: 1] 在全球气候变化条件下,全球陆地区域的降水量及其空间/时间分布模式将发生明显改变,这种变 化将对中国的陆地生态系统,尤其是干旱地区的温带草地生态系统碳收支产生重要影响.作为中国陆地生态系统通量观测研究网络(ChinaFLUX)的一部 分,本研究利用2003至2004年在内蒙古羊草草原的涡度相关通量观测数据,初步探讨了水分胁迫对羊草草原生态系统光合和呼吸作用的影响.研究发现温度 和水分是影响该生态系统在生长季(5~9月)的光合和呼吸作用的主要因子.在土壤水分适宜条件下,生态系统呼吸对温度变化的敏感性较大 (Q10=2.0),而当土壤含水量降低时生态系统呼吸对温度的敏感性明显降低(Q10=1.6).高温和干旱会显著降低生态系统的光合生产力.生长季的 降水量及其季节分配模式的不同对草地生态系统的生物物候有明显的影响,在2003年6月初生态系统就开始净吸收CO2、在7月初出现最大净生态系统CO2 吸收量,而因随后发生的干旱和高温胁迫使其在8月就提早进入休眠期;2004年春季的严重干旱导致生态系在7月初才开始净吸收CO2,并在降水丰沛气温适 宜的8月出现最大CO2吸收量,水分胁迫导致该草地植物生长发育比2003年推迟1个多月.观测结果显示该草地生态系统在2004年5~9月比2003年 同期多吸收30gCO2·m^-2.我国温带草原植被类型复杂多样,要准确估算我国草地生态系统碳收支还需要开展更多的长期联合观带研究.
GaoCD, SunXY, ZhangL, LiZG, AL ( 2009). Changing characteristics of CO2 flux from the interface of soil-atmosphere at arid area in north temperate Journal of Beijing Forestry University, 31(6), 32-38. URL [本文引用: 1] 采用箱法对栗钙土、灰钙土、粗骨土和山地灰褐土4种有代表性的干旱土壤表面CO2通量进行观测和研究。结果表明:森林土壤(粗骨土和山地灰褐土)的通量显著大于草原土壤(栗钙土和灰钙土)。干旱区土壤表面CO2通量的平均值为230.05μmol/(m2·h),变化范围为-147.27~2319.55μmol/(m2·h)。不同土壤类型之间存在差异,粗骨土(351.82μmol/(m2·h))〉山地灰褐土(347.33μmol/(m2·h))〉栗钙土(193.36μmol/(m2·h))〉灰钙土(162.37μmol/(m2·h))。土壤表面CO2通量存在季节变化,趋势呈“S”形。9月份最高(516.79μmol/(m2·h)),以土壤向大气释放为主;1月份最低(-7.09μmol/(m2·h)),以大气进入土壤为主;具有春夏秋冬交替规律,与气候变化趋势基本一致,土壤表面CO2通量稍有后滞。全天候土壤表面CO2通量呈“山峰”形变化,04:00最小(154.13μmol/(m2·h)),12:00最大(349.65μmol/(m2·h)),具有昼夜交替规律,比气候日变化稍有滞后。影响土壤表面CO2通量的环境因子有地表空气温度、土壤温度(0~10cm、10~20cm和20~30cm)、土壤含水量(0~10cm、10~20cm和20~30cm);其中,地表空气温度、土壤温度(0~10cm、10~20cm和20~30cm)和土壤含水量(0~10cm)分别与土壤表面CO2通量呈正相关关系,而10~20cm和20~30cm深度的土壤含水量与土壤表面CO2通量呈负相关关系,地表空气相对湿度与土壤表面CO2通量的关系不显著。大气与土壤之间的CO2存在双向转移机制,CO2不仅从土壤向大气转移,而且也从大气向土壤转移,热量在地球表面的差异性分布,导致温带和寒带地区的土壤具有平衡大气CO2浓度的功能,是温带、寒带地区的显著特征。 高程达, 孙向阳, 张林, 李志刚, 阿拉塔 ( 2009). 北温带干旱地区土壤-大气界面CO2通量的变化特征 北京林业大学学报, 31(6), 32-38. URL [本文引用: 1] 采用箱法对栗钙土、灰钙土、粗骨土和山地灰褐土4种有代表性的干旱土壤表面CO2通量进行观测和研究。结果表明:森林土壤(粗骨土和山地灰褐土)的通量显著大于草原土壤(栗钙土和灰钙土)。干旱区土壤表面CO2通量的平均值为230.05μmol/(m2·h),变化范围为-147.27~2319.55μmol/(m2·h)。不同土壤类型之间存在差异,粗骨土(351.82μmol/(m2·h))〉山地灰褐土(347.33μmol/(m2·h))〉栗钙土(193.36μmol/(m2·h))〉灰钙土(162.37μmol/(m2·h))。土壤表面CO2通量存在季节变化,趋势呈“S”形。9月份最高(516.79μmol/(m2·h)),以土壤向大气释放为主;1月份最低(-7.09μmol/(m2·h)),以大气进入土壤为主;具有春夏秋冬交替规律,与气候变化趋势基本一致,土壤表面CO2通量稍有后滞。全天候土壤表面CO2通量呈“山峰”形变化,04:00最小(154.13μmol/(m2·h)),12:00最大(349.65μmol/(m2·h)),具有昼夜交替规律,比气候日变化稍有滞后。影响土壤表面CO2通量的环境因子有地表空气温度、土壤温度(0~10cm、10~20cm和20~30cm)、土壤含水量(0~10cm、10~20cm和20~30cm);其中,地表空气温度、土壤温度(0~10cm、10~20cm和20~30cm)和土壤含水量(0~10cm)分别与土壤表面CO2通量呈正相关关系,而10~20cm和20~30cm深度的土壤含水量与土壤表面CO2通量呈负相关关系,地表空气相对湿度与土壤表面CO2通量的关系不显著。大气与土壤之间的CO2存在双向转移机制,CO2不仅从土壤向大气转移,而且也从大气向土壤转移,热量在地球表面的差异性分布,导致温带和寒带地区的土壤具有平衡大气CO2浓度的功能,是温带、寒带地区的显著特征。
GuoMY, ChaoKL, YouJC, XuLJ, WangLJ, JiaSJ, XinXP ( 2012). Soil microbial characteristic and soil respiration in grassland under different use patterns Acta Agrestia Sinica, 20, 42-48. DOI:10.11733/j.issn.1007-0435.2012.01.007URL [本文引用: 2] 通过对呼伦贝尔羊草(Leymus chinensis)草甸草原土壤呼吸特性及土壤微生物的测定,研究了3种不同利用方式(围封、放牧和刈割)对天然草地土壤呼吸特性及土壤微生物的影响,为草甸草原土壤呼吸研究提供基础理论依据。结果表明:不同利用方式下土壤呼吸速率、土壤微生物量碳含量与土壤酶活性(除脲酶)均表现为草地围封比放牧和刈割呼吸速率高(含量高、活性强);土壤微生物量氮含量、脲酶活性和微生物数量表现为草地放牧和刈割比围封的含量高(活性强、数量多);土壤微生物量碳、氮含量、土壤酶活性(除脲酶)和土壤微生物数量均以表层(0~10cm)最高(活性最强、数量最多),随着土层深度增加而降低(活性下降、数量减少)。土壤呼吸速率与微生物量碳、土壤酶活性(除过氧化氢酶)成正相关,与微生物量氮、过氧化氢酶和微生物数量成负相关,其中只与微生物量氮相关性显著,而其余之间均不显著。 郭明英, 朝克图, 尤金成, 徐丽君, 王丽娟, 贾淑杰, 辛晓平 ( 2012). 不同利用方式下草地土壤微生物及土壤呼吸特性 草地学报, 20, 42-48. DOI:10.11733/j.issn.1007-0435.2012.01.007URL [本文引用: 2] 通过对呼伦贝尔羊草(Leymus chinensis)草甸草原土壤呼吸特性及土壤微生物的测定,研究了3种不同利用方式(围封、放牧和刈割)对天然草地土壤呼吸特性及土壤微生物的影响,为草甸草原土壤呼吸研究提供基础理论依据。结果表明:不同利用方式下土壤呼吸速率、土壤微生物量碳含量与土壤酶活性(除脲酶)均表现为草地围封比放牧和刈割呼吸速率高(含量高、活性强);土壤微生物量氮含量、脲酶活性和微生物数量表现为草地放牧和刈割比围封的含量高(活性强、数量多);土壤微生物量碳、氮含量、土壤酶活性(除脲酶)和土壤微生物数量均以表层(0~10cm)最高(活性最强、数量最多),随着土层深度增加而降低(活性下降、数量减少)。土壤呼吸速率与微生物量碳、土壤酶活性(除过氧化氢酶)成正相关,与微生物量氮、过氧化氢酶和微生物数量成负相关,其中只与微生物量氮相关性显著,而其余之间均不显著。
HoughtonRA ( 2003). Revised estimates of the annual net flux of carbon to the atmosphere from changes in land use and land management 1850-2000 Tellus Series B—Chemical and Physical Meteorology, 55, 378-390. [本文引用: 1]
HuY, ZhuXP, HanDL, JiaHT, HuBA, LiDP ( 2016). Effects of fencing on soil respiration rates in the meadow steppes in the northern slope of the Tianshan Mountains Acta Ecologica Sinica, 36, 6379-6386. [本文引用: 3]
JiaBR, ZhouGS, WangFY, WangYH, WengEH ( 2007). Effects of grazing on soil respiration of Leymus chinensis steppe. Climatic Change, 82, 211-223. [本文引用: 1]
KatoT, TangYH, GuS, HirotaM, DuMY, LiYN, ZhaoXQ ( 2006). Temperature and biomass influences on interannual changes in CO2 exchange in an alpine meadow on the Qinghai-Tibetan Plateau Global Change Biology, 12, 1285-1298. DOI:10.1137/040614359URL [本文引用: 1] Three years of eddy covariance measurements were used to characterize the seasonal and interannual variability of the CO2 fluxes above an alpine meadow (3250 m a.s.l.) on the Qinghai-Tibetan Plateau, China. This alpine meadow was a weak sink for atmospheric CO2, with a net ecosystem production (NEP) of 78.5, 91.7, and 192.5 g C m(-2) yr(-1) in 2002, 2003, and 2004, respectively. The prominent, high NEP in 2004 resulted from the combination of high gross primary production (GPP) and low ecosystem respiration (R-e) during the growing season. The period of net absorption of CO2 in 2004, 179 days, was 10 days longer than that in 2002 and 5 days longer than that in 2003. Moreover, the date on which the mean air temperature first exceeded 5.0 degrees C was 10 days earlier in 2004 (DOY110) than in 2002 or 2003. This date agrees well with that on which the green aboveground biomass (Green AGB) started to increase. The relationship between light-use efficiency and Green AGB was similar among the three years. In 2002, however, earlier senescence possibly caused low autumn GPP, and thus the annual NEP, to be lower. The low summertime R-e in 2004 was apparently caused by lower soil temperatures and the relatively lower temperature dependence of R-e in comparison with the other years. These results suggest that (1) the Qinghai-Tibetan Plateau plays a potentially significant role in global carbon sequestration, because alpine meadow covers about one-third of this vast plateau, and (2) the annual NEP in the alpine meadow was comprehensively controlled by the temperature environment, including its effect on biomass growth.
LawBE, FalgeE, GuL, BaldocchiDD, BakwinP, BerbigierP, DavisK, DolmanAJ, FalkM, FuentesJD, GoldsteinA, GranierA, GrelleA, HollingerD, JanssensIA, JarvisP, JensenNO, KatulG, MahliY, MatteucciG, MeyersT, MonsonR, MungerW, OechelW, OlsonR, PilegaardK, PawKT, ThorgeirssonH, ValentiniR, VermaS, VesalaT, WilsonK, WofsyS ( 2002). Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation Agricultural and Forest Meteorology, 113, 97-120. DOI:10.1016/S0168-1923(02)00104-1URL [本文引用: 1] The objective of this research was to compare seasonal and annual estimates of CO 2 and water vapor exchange across sites in forests, grasslands, crops, and tundra that are part of an international network called FLUXNET, and to investigating the responses of vegetation to environmental variables. FLUXNETs goals are to understand the mechanisms controlling the exchanges of CO 2, water vapor and energy across a spectrum of time and space scales, and to provide information for modeling of carbon and water cycling across regions and the globe. At a subset of sites, net carbon uptake (net ecosystem exchange, the net of photosynthesis and respiration) was greater under diffuse than under direct radiation conditions, perhaps because of a more efficient distribution of non-saturating light conditions for photosynthesis, lower vapor pressure deficit limitation to photosynthesis, and lower respiration associated with reduced temperature. The slope of the relation between monthly gross ecosystem production and evapotranspiration was similar between biomes, except for tundra vegetation, showing a strong linkage between carbon gain and water loss integrated over the year (slopes=3.4 g CO 2/kg H 2O for grasslands, 3.2 for deciduous broadleaf forests, 3.1 for crops, 2.4 for evergreen conifers, and 1.5 for tundra vegetation). The ratio of annual ecosystem respiration to gross photosynthesis averaged 0.83, with lower values for grasslands, presumably because of less investment in respiring plant tissue compared with forests. Ecosystem respiration was weakly correlated with mean annual temperature across biomes, in spite of within site sensitivity over shorter temporal scales. Mean annual temperature and site water balance explained much of the variation in gross photosynthesis. Water availability limits leaf area index over the long-term, and inter-annual climate variability can limit carbon uptake below the potential of the leaf area present.
LiXR, MaFY, XiaoHL, WangXP, KimKC ( 2004). Long term effects of revegetation on soil water content of sand dunes in arid region of Northern China Journal of Arid Environments, 57, 1-16. DOI:10.1016/S0140-1963(03)00089-2URL [本文引用: 1] Desert is a challenging environment for organisms largely controlled by extreme abiotic factors including soil water content. The objectives of this study were to: (1) determine the effect of vegetative water utilization on soil water content; and (2) monitor the long-term vegetative response to the change of soil water content after establishing the revegetation of sand dunes of the Shapotou Region in the Tengger Desert, Northern China. The permanent revegetation plots were established in different sites in 1956, 1964, 1976 and 1982. Soil water content was monitored at 2002cm intervals of 10–30002cm in depth. Water content in sand dunes, was measured gravimetrically after 24-h desiccation at 105°C during the entire period of revegetation through the year to 2000. Our data show that soil water contents rapidly decreased at 9–10 years after revegetation, and similarly the vertical profile of soil water contents also decreased by depth. Annual precipitation is positively correlated with the water contents at depths 0–4002cm, although no clear-cut relationship was shown between precipitation and soil water at the deeper soil layers (from 40 to 30002cm). It is therefore concluded that water utilization of different plant species: shrub with deep-rooting system and annual plants with shallow-rooting system influences the spatial distribution of water inside the soil profile. Temporal and spatial variances of soil water also affected the changes in composition, coverage and biomass of vegetation. Vegetation composition of dominant species has successively changed from all shrub species, such as Caragana korshinskii and Hedysarum scoparaium to annual plants and shrub with shallow-rooting system ( Artemisia ordosica ). The shrub biomass also changed along with soil water content, although the biomass of herb species increased over time. After 45 years, the shrub coverage decreased from the highest average 30.21% to the current level of 6%, whereas herb coverage increased over time. The soil water content was, however, kept relatively constant at lower level (1.23%) despite shrub coverage was reduced to about 6%. These results provide important baseline information for establishing vegetation and utilization of limited water resource in arid desert regions.
LuoCY, BaoXY, WangSP, ZhuXX, CuiSJ, ZhangZH, XuBB, NiuHS, ZhaoL, ZhaoXQ ( 2015). Impacts of seasonal grazing on net ecosystem carbon exchange in alpine meadow on the Tibetan Plateau Plant and Soil, 396, 381-395. DOI:10.1007/s11104-015-2602-6URL [本文引用: 1] BACKGROUND AND AIMS: Understanding the effect of grazing season on net ecosystem CO60 exchange (NEE) and its components is crucial to predict the feedback of grazing management to climate change. METHODS: We estimated NEE, gross primary productivity (GPP) and ecosystem respiration (Re) under different seasonal grazing practices (i.e. no-grazing (NG), warm season grazing (WG) and cold season grazing (CG)) by sheep during the growing seasons from 2008 to 2012 on the Tibetan Plateau. RESULTS: Our results show that the impacts of seasonal grazing on daily GPP, Re and NEE in the alpine meadow ecosystem varied with sampling date and year. Compared with NG and CG, WG significantly reduced average seasonal NEE by 22.702%, because grazing impact was exacerbated by drought in July in 2010. Soil temperature only explained 19–3102% of the variation in daily GPP, Re and NEE for all grazing treatments. The interannual variabilities of GPP, Re and NEE were mainly determined by root biomass and/or average soil temperature during the growing season. CONCLUSIONS: Our results suggest that although WG may decrease sequestration of CO60 under continuous drought conditions after grazing, it would have little impact on CO60 sequestration during the growing season under conditions of future warming with greater rainfall in alpine meadows on the Tibetan Plateau.
MaJ, TangHP ( 2011). Variations of soil respiration rate and its temperature sensitivity among different land use types in the agro-pastoral ecotone of Inner Mongolia Chinese Journal of Plant Ecology, 35, 167-175. DOI:10.3724/SP.J.1258.2011.00167URL [本文引用: 1] 2009年8–10月,采用动态气室法观测了内蒙古农牧交错区多伦县农田、弃耕和围封3种土地利用方式下,土壤呼吸速率从6:00到18:00的变化规律,分析了不同深度的土壤温度与土壤含水量对土壤呼吸速率的控制作用。结果表明,空间尺度上,不同土地利用方式的土壤呼吸速率由高到低依次为:农田〉弃耕〉围封;时间尺度上,土壤呼吸速率在6:00–18:00的变化趋势为单峰曲线,在12:00–15:00达到峰值,随后降低,在18:00基本恢复到6:00左右的呼吸水平,同时,土壤呼吸速率在9、10月显著降低。利用Van’tHoff指数模型研究不同深度土壤温度对土壤呼吸速率的影响发现,10–15cm深度的土壤温度对土壤呼吸速率的影响最为显著,其中,土壤呼吸温度敏感性由高到低分别为:农田〉围封〉弃耕。相反,由于8–10月土壤含水量变化较小,故土壤含水量与土壤呼吸速率间的相关性不显著,土壤含水量不能解释该时段土壤呼吸速率的变化。 马骏, 唐海萍 ( 2011). 内蒙古农牧交错区不同土地利用方式下土壤呼吸速率及其温度敏感性变化 植物生态学报, 35, 167-175. DOI:10.3724/SP.J.1258.2011.00167URL [本文引用: 1] 2009年8–10月,采用动态气室法观测了内蒙古农牧交错区多伦县农田、弃耕和围封3种土地利用方式下,土壤呼吸速率从6:00到18:00的变化规律,分析了不同深度的土壤温度与土壤含水量对土壤呼吸速率的控制作用。结果表明,空间尺度上,不同土地利用方式的土壤呼吸速率由高到低依次为:农田〉弃耕〉围封;时间尺度上,土壤呼吸速率在6:00–18:00的变化趋势为单峰曲线,在12:00–15:00达到峰值,随后降低,在18:00基本恢复到6:00左右的呼吸水平,同时,土壤呼吸速率在9、10月显著降低。利用Van’tHoff指数模型研究不同深度土壤温度对土壤呼吸速率的影响发现,10–15cm深度的土壤温度对土壤呼吸速率的影响最为显著,其中,土壤呼吸温度敏感性由高到低分别为:农田〉围封〉弃耕。相反,由于8–10月土壤含水量变化较小,故土壤含水量与土壤呼吸速率间的相关性不显著,土壤含水量不能解释该时段土壤呼吸速率的变化。
McfaddenJP, EugsterW, ChapinFS ( 2003). A regional study of the controls on water vapor and CO2 exchange in arctic tundra Ecology, 84, 2762-2776. DOI:10.1890/01-0444URL [本文引用: 1] Water vapor and CO2exchanges were measured by the eddy covariance method in 24 ecosystems along a transect from the Arctic coast to the latitudinal tree line in northern Alaska during three growing seasons. Variations in net ecosystem exchange across the region were controlled by differences in the net uptake of CO2due to photosynthesis, rather than by differences in ecosystem respiration. Daytime CO2uptake was related mainly to site differences in leaf area index, whereas nighttime CO2efflux was related to leaf area index and soil moisture. Temperature had no effect on regional patterns of ecosystem respiration during the growing season. Regional variations in surface conductance were largely explained by the water content of the moss-soil layer. Water vapor and CO2fluxes were poorly coupled because water vapor exchange was determined largely by evaporation from mosses, whereas CO2exchange was controlled by vascular plant activity. This is in contrast to the close correlation between maximum rates of surface conductance and CO2assimilation that is observed at the global scale. The results also suggest that the relationship between the maximum rates of surface conductance and stomatal conductance in arctic tundra departs markedly from the global pattern, because the moss layer dried more readily than a moist soil. These spatially distributed eddy covariance measurements revealed new functional relationships among water vapor and CO2exchanges and their environmental controls in arctic tundra; however, there remains a need for winter flux observations and long-term measurements to understand the net effect of these processes on the annual carbon balance of the region.
OberbauerSF, TweedieCE, Welker JM FahnestockJT, HenryGHR, WebberPJ, HollisterRD, WaikerMD, KuchyA, ElmoreE, StarrG ( 2007). Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients Ecological Monographs, 77, 221-238. DOI:10.1890/06-0649URL [本文引用: 1] Climate warming is expected to differentially affect CO2 exchange of the diverse ecosystems in the Arctic. Quantifying responses of M exchange to warming in these ecosystems will require coordinated experimentation using standard temperature manipulations and measurements. Here, we used the International Tundra Experiment (ITEX) standard warming treatment to determine CO2 flux responses to growing-season warming for ecosystems spanning natural temperature and moisture ranges across the Arctic biome. We used the four North American Arctic ITEX sites (Toolik Lake, Atqasuk, and Barrow [USA] and Alexandra Fiord [Canada]) that span 10 degrees of latitude. At each site, we investigated the CO2 responses to warming in both dry and wet or moist ecosystems. Net ecosystem CO2 exchange (NEE), ecosystem respiration (ER), and gross ecosystem photosynthesis (GEP) were assessed using chamber techniques conducted over 24-h periods sampled regularly throughout the summers of two years at all sites. At Toolik Lake, warming increased net CO2 losses in both moist and dry ecosystems. In contrast, at Atqasuk and Barrow, warming increased net CO2 uptake in wet ecosystems but increased losses from dry ecosystems. At Alexandra Fiord, warming improved net carbon uptake in the moist ecosystem in both years, but in the wet and dry ecosystems uptake increased in one year and decreased the other. Warming generally increased ER, with the largest increases in dry ecosystems. In wet ecosystems, high soil moisture limited increases in respiration relative to increases in photosynthesis. Warming generally increased GEP, with the notable exception of the Toolik Lake moist ecosystem, where warming unexpectedly decreased GEP >25%. Overall, the respiration response determined the effect of warming on ecosystem CO2 balance. Our results provide the first multiple-site comparison of arctic tundra CO2 flux responses to standard warming treatments across a large climate gradient. These results indicate tha
SharkhuuA, PlanteAF, EnkhmandalO, GonneauC, CasperBB, BoldgivB, PetraitisPS ( 2016). Soil and ecosystem respiration responses to grazing, watering and experimental warming chamber treatments across topographical gradients in northern Mongolia Geoderma, 269, 91-98. DOI:10.1016/j.geoderma.2016.01.041URLPMID:28239190 [本文引用: 1] Abstract Globally, soil respiration is one of the largest fluxes of carbon to the atmosphere and is known to be sensitive to climate change, representing a potential positive feedback. We conducted a number of field experiments to study independent and combined impacts of topography, watering, grazing and climate manipulations on bare soil and vegetated soil (i.e., ecosystem) respiration in northern Mongolia, an area known to be highly vulnerable to climate change and overgrazing. Our results indicated that soil moisture is the most important driving factor for carbon fluxes in this semi-arid ecosystem, based on smaller carbon fluxes under drier conditions. Warmer conditions did not result in increased respiration. Although the system has local topographical gradients in terms of nutrient, moisture availability and plant species, soil respiration responses to OTC treatments were similar on the upper and lower slopes, implying that local heterogeneity may not be important for scaling up the results. In contrast, ecosystem respiration responses to OTCs differed between the upper and the lower slopes, implying that the response of vegetation to climate change may override microbial responses. Our results also showed that light grazing may actually enhance soil respiration while decreasing ecosystem respiration, and grazing impact may not depend on climate change. Overall, our results indicate that soil and ecosystem respiration in this semi-arid steppe are more sensitive to precipitation fluctuation and grazing pressure than to temperature change.
SunDC, LiYL, ZhaoXY, ZuoXA, MaoW ( 2015). Effects of enclosure and grazing on carbon and water fluxes of sandy grassland Chinese Journal of Plant Ecology, 39, 565-576. [本文引用: 2]
SuykerAE, VermanSB, BurbaGG ( 2003). Interannual variability in net CO2 exchange of a native tall grass prairie Global Change Biology, 9, 255-265. DOI:10.1046/j.1365-2486.2003.00567.xURL [本文引用: 1] Abstract Year-round eddy covariance flux measurements were made in a native tallgrass prairie in north-central Oklahoma, USA during 1997鈥2000 to quantify carbon exchange and its interannual variability. This prairie is dominated by warm season C 4 grasses. The soil is a relatively shallow silty clay loam underlined with a heavy clay layer and a limestone bedrock. During the study period, the prairie was burned in the spring of each year, and was not grazed. In 1997 there was adequate soil moisture through the growing season, but 1998 had two extended periods of substantially low soil moisture (with concurrent high air temperatures and vapor pressure deficits), one early and one later in the growing season. There was also moisture stress in 1999, but it was less severe and occurred later in the season. The annual net ecosystem CO 2 exchange, NEE (before including carbon loss during the burn) was 274, 46 and 124gCm 2 yr 1 in 1997, 1998, and 1999, respectively (flux toward the surface is positive), and the associated variation seemed to mirror the severity of moisture stress. We also examined integrated values of NEE during different periods (e.g. day/night; growing season/senescence). Annually integrated carbon dioxide uptake during the daytime showed the greatest variability from year to year, and was primarily linked to the severity of moisture stress. Carbon loss during nighttime was a significant part of the annual daytime NEE, and was fairly stable from year to year. When carbon loss during the burn (estimated from pre- and post-burn biomass samples) was incorporated in the annual NEE, the prairie was found to be approximately carbon neutral (i.e. net carbon uptake/release was near zero) in years with no moisture stress (1997) or with some stress late in the season (1999). During a year with severe moisture stress early in the season (1998), the prairie was a net source of carbon. It appears that moisture stress (severity as well as timing of occurrence) was a dominating factor regulating the annual carbon exchange of the prairie.
WuLB, GuS, ZhaoL, XuSX, ZhouHK, FengC, XuWX, LiYN, ZhaoXQ, TangYH ( 2010). Variation in net CO2 exchange, gross primary production and its affecting factors in the planted pasture ecosystem in Sanjiangyuan Region of the Qinghai-Tibetan Plateau of China Chinese Journal of Plant Ecology, 34, 770-780. DOI:10.3773/j.issn.1005-264x.2010.07.002URL [本文引用: 1] 为了揭示三江源区垂穗披碱草(Elymus nutans)人工草地生态系统(100°26'-100°41'E,34°17'-34°25'N,海拔3 980 m)的净生态系统CO2交换(NEE),该研究利用2006年涡度相关系统观测的数据分析了该人工草地的NEE,总初级生产力(GPP)、生态系统呼吸 (Reco)以及Reco/GPP的变化特征及其影响因子.CO2日最大吸收值为6.56 g CO2·m-2·d-1,最大排放值为4.87 gCO2·m-2·d-1.GPP年总量为1 761 g CO2·m-2,其中约90%以上被生态系统呼吸所消耗,CO2的年吸收量为111 gCO2·m-2.5月的Reco/GPP略高于生长季的其他月份,为90%;6月Reco/GPP比值最低,为79%.生态系统的呼吸商(Q10)为 4.81,显著高于其他生态系统.该研究表明:生长季的NEE主要受光量子通量密度(PPFD)、温度和饱和水汽压差(VPD)的影响,生态系统呼吸则主 要受土壤温度的控制. 吴力博, 古松, 赵亮, 徐世晓, 周华坤, 冯超, 徐维新, 李英年, 赵新全, 唐艳鸿 ( 2010). 三江源地区人工草地的生态系统CO2净交换、总初级生产力及其影响因子 植物生态学报, 34, 770-780. DOI:10.3773/j.issn.1005-264x.2010.07.002URL [本文引用: 1] 为了揭示三江源区垂穗披碱草(Elymus nutans)人工草地生态系统(100°26'-100°41'E,34°17'-34°25'N,海拔3 980 m)的净生态系统CO2交换(NEE),该研究利用2006年涡度相关系统观测的数据分析了该人工草地的NEE,总初级生产力(GPP)、生态系统呼吸 (Reco)以及Reco/GPP的变化特征及其影响因子.CO2日最大吸收值为6.56 g CO2·m-2·d-1,最大排放值为4.87 gCO2·m-2·d-1.GPP年总量为1 761 g CO2·m-2,其中约90%以上被生态系统呼吸所消耗,CO2的年吸收量为111 gCO2·m-2.5月的Reco/GPP略高于生长季的其他月份,为90%;6月Reco/GPP比值最低,为79%.生态系统的呼吸商(Q10)为 4.81,显著高于其他生态系统.该研究表明:生长季的NEE主要受光量子通量密度(PPFD)、温度和饱和水汽压差(VPD)的影响,生态系统呼吸则主 要受土壤温度的控制.
XuP ( 1993). Grassland Resources and Its Utilization in Xinjiang. Science and Technology and Health Press, ürümqi. [本文引用: 1]
YangLL, GongJR, WangYH, LiuM, LuoQP, XuS, PanY, ZhaiZW ( 2016). Effects of grazing intensity and grazing exclusion on litter decomposition in the temperate steppe of Nei Mongol, China Chinese Journal of Plant Ecology, 40, 748-759. [本文引用: 1]
YangY, HanGD, LiYH, ChenZF, WangCJ ( 2012). Response of soil respiration to grazing intensity, water contents, and temperature of soil indifferent grasslands of Inner Mongolia Acta Prataculturae Sinica, 21(6), 8-14. [本文引用: 2]
ZhangWH, GuanSY, LiYJ ( 2000). Effect of grazing capacity on water content, nutrient and biomass of steppe soil Journal of Arid Land Resources and Environment, 14(4), 62-65. [本文引用: 1]
Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients 1 2007
... id="C6">生态系统CO2净交换速率(NEE)代表生态系统吸收或释放CO2的能力, 它由生态系统总初级生产力(GEP)与生态系统呼吸(ER)共同决定(Oberbauer et al., 2007).随着全球气候变化, CO2浓度升高, 草本层和大气间的碳交换及其对环境因子的响应机制成为目前研究的热点问题.许多****通过不同尺度的试验观测试图掌握在不同时空上的生态系统碳交换现状、潜力、速率和机制(Baldocchi, 1997; Suyker et al., 2003).国外在这方面已经做了大量研究(Black et al., 1996, Flanagan & Johnson, 2005).相比之下, 我国碳通量的观测与研究启动较晚, 而且多集中于农田和森林生态系统(张永强等, 2002; 段晓男等, 2005), 对草地生态系统碳收支的研究大多集中在荒漠草原、典型草原、高寒草原等(Jia et al., 2007; Chen et al., 2014; Luo et al., 2015; Sharkhuu et al., 2016), 虽然对草甸草原的研究有较多报道, 但较少涉及天山山脉山地草甸草原的研究报道. ...
Soil and ecosystem respiration responses to grazing, watering and experimental warming chamber treatments across topographical gradients in northern Mongolia 1 2016
... id="C6">生态系统CO2净交换速率(NEE)代表生态系统吸收或释放CO2的能力, 它由生态系统总初级生产力(GEP)与生态系统呼吸(ER)共同决定(Oberbauer et al., 2007).随着全球气候变化, CO2浓度升高, 草本层和大气间的碳交换及其对环境因子的响应机制成为目前研究的热点问题.许多****通过不同尺度的试验观测试图掌握在不同时空上的生态系统碳交换现状、潜力、速率和机制(Baldocchi, 1997; Suyker et al., 2003).国外在这方面已经做了大量研究(Black et al., 1996, Flanagan & Johnson, 2005).相比之下, 我国碳通量的观测与研究启动较晚, 而且多集中于农田和森林生态系统(张永强等, 2002; 段晓男等, 2005), 对草地生态系统碳收支的研究大多集中在荒漠草原、典型草原、高寒草原等(Jia et al., 2007; Chen et al., 2014; Luo et al., 2015; Sharkhuu et al., 2016), 虽然对草甸草原的研究有较多报道, 但较少涉及天山山脉山地草甸草原的研究报道. ...
 ,1,2, 朱新萍1,2, 贾宏涛
,1,2, 朱新萍1,2, 贾宏涛 ,1,2,*, 韩东亮1,2, 胡保安1,2, 李典鹏1
,1,2,*, 韩东亮1,2, 胡保安1,2, 李典鹏1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT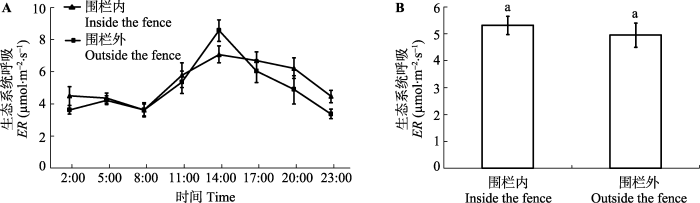 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT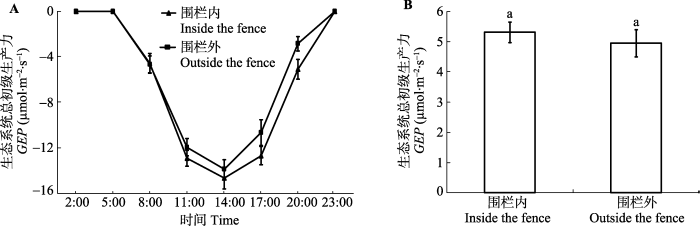 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT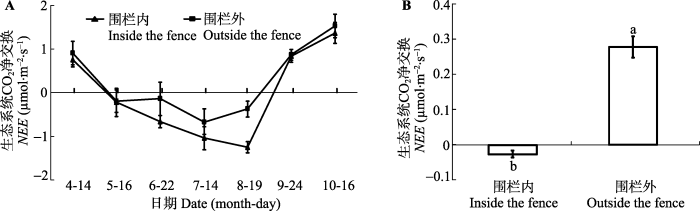 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT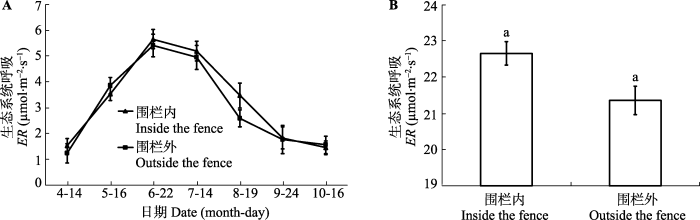 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT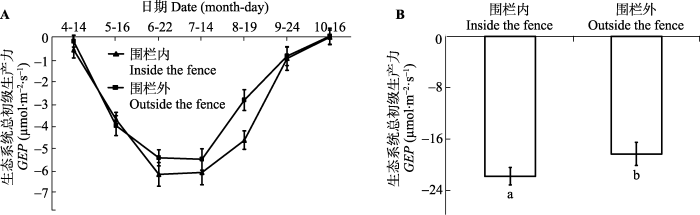 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}