 ,
, ,1,2,*, 关正翾1, 盛建东1
,1,2,*, 关正翾1, 盛建东1Soil respiration features of mountain meadows under four typical land use types in Zhaosu Basin
WANG Xiang1, ZHU Ya-Qiong1, ZHENG Wei,,1,2,*, GUAN Zheng-Xuan1, SHENG Jian-Dong1通讯作者:
编委: 白永飞
责任编辑: 李敏
| 基金资助: |
Online:2018-03-20
| Supported by: | SupportedbytheStrategicPriorityResearchProgramofChineseAcademyofSciences( |

摘要
关键词:
Abstract
Keywords:
PDF (2022KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
王祥, 朱亚琼, 郑伟, 关正翾, 盛建东. 昭苏山地草甸4种典型土地利用方式下的土壤呼吸特征. 植物生态学报, 2018, 42(3): 382-396 doi:10.17521/cjpe.2017.0050
WANG Xiang, ZHU Ya-Qiong, ZHENG Wei, GUAN Zheng-Xuan, SHENG Jian-Dong.
土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(Willcock et al., 2016): 其通过改变土壤微环境、土壤物理化学过程和土壤微生物活动, 进而影响土壤CO2的产生与排放(Flechard et al., 2007; 刘慧峰等, 2014)。当前, 由人类活动引起的草地利用/土地利用方式的变化导致的陆地生态系统变化对全球碳循环有重要的影响。如Sampson等(1993)就认为, 草地开垦后, 虽然植被生产力有所提高, 但由于土壤呼吸增强, 土壤有机碳储量下降; 刘季骢等(2014)也认为, 无论草地开垦还是农田退耕还草, 均会增加土壤呼吸速率, 进而增加碳排放。因此, 准确地估计草地利用/土地利用方式变化对陆地生态系统碳循环和碳平衡的影响是当前全球气候变化和碳循环的研究重点(Sampson et al., 1993)。
目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(宁发等, 2008)。郭明英等(2012)对呼伦贝尔羊草草甸草原3种不同利用方式(围封、放牧和刈割)下的土壤呼吸和土壤微生物特性进行了研究, 结果表明不同利用方式下土壤呼吸速率表现为围封比放牧和刈割高, 其微生物生物量碳和过氧化氢酶活性也较高。王忆慧等(2015)则发现, 在干旱年份土壤呼吸最高的是放牧样地, 而湿润年份是刈割样地, 水分是影响温带草原碳循环的重要限制因素。不同草地土地利用方式一方面通过改变根系呼吸(Hanson et al., 2000), 扰动土壤(Lal et al., 1998), 导致土壤水分、温度(王小国等, 2007)和土壤有机碳(Lal, 2004)发生显著变化, 进而抑制或促进微生物活性, 降低或加快土壤呼吸; 另一方面也改变了土壤理化性质, 从而对土壤呼吸产生很大影响(Sparling et al., 1992)。草地开垦是引起草原土壤碳减少的最强烈的人为因素。草地开垦过程会破坏土壤中致密的根系, 使土壤深层的有机质暴露在空气中, 从而加速了其分解过程, 导致土壤呼吸作用增强(Anderson & Coleman, 1985)。张平良等(2007)研究发现在高寒地区种植的人工草地固碳效率是撂荒地的1-2倍, 从而提高了碳储量, 降低了土壤呼吸。但也有研究表明人工草地的建植也可能降低土壤有机碳含量, 或对土壤有机碳没有影响, 如加拿大开垦天然草原种植冰草(Agropyron cristatum) 13年后, 土壤碳储量和对照相比无显著变化(Wang, 2010)。
退化的生态系统恢复成草地往往是理想的生态选择, 能够增加水分利用效率, 增加碳储量, 减少温室气体排放(Zhang et al., 2015)。进一步讲, 利用豆禾牧草混播进行的草地补播/重建、间混作改良农田不仅有利于植被的恢复, 而且能有效地提高土地的固碳能力(张英俊等, 2013; Raiesi & Beheshti, 2014)。然而, 现有关于对退化草地进行人工补播改良或重建为人工草地的研究中, 多集中在牧草产量、品质和土壤养分利用改善方面, 对土地利用方式改变后土壤碳循环(包括土壤呼吸)的研究较少。本研究选取新疆伊犁昭苏马场退化山地草甸为研究对象, 将地势开阔、植被相对均一、 土壤类型一致的退化山地草甸翻耕建成的豆禾混播草地, 补播改良成的豆禾补播草地, 围封草地, 以及开垦成种植春小麦(Triticum aestivum)的农田作为此次试验的研究对象, 试从上述当地4种典型土地利用方式入手, 研究土地利用方式变化对新疆天山北坡西段山地草甸土壤呼吸特征的影响, 并探讨影响土壤呼吸的生物和非生物因素, 以期为天山北坡山地草甸不同草地土地利用方式下的草地碳汇管理机制提供依据, 并为深入理解我国草地生态系统碳循环过程和草地生态系统碳平衡估算提供依据。
1 材料和方法
1.1 研究区概况
试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏县的昭苏马场(81.05°-81.08° E, 40.92°-43.25° N), 属于大陆性温带山区半干旱半湿润冷凉气候。2014-2015年平均气温为4.1 ℃, ≥10 ℃年积温为1β416.8 ℃, 无霜期85-100天, 2014-2015年平均年降水量为490.1 mm, 降水集中于5-10月, 约占全年总降水量的74%, 平均年蒸发量1β261.6 mm, 约是降水量的2.5倍, 积雪期158天, 积雪厚度20-60 cm。试验小区位于天山北坡西段山地草甸, 未退化山地草甸植被主要成分有鸭茅(Dactylis glomerata)、无芒雀麦(Bromus inermis)、新疆鹅观草(Roegneria sinkiangensis)、猫尾草(Phleum pratense)、草原糙苏(Phlomis pratensis)、草地老鹳草(Geranium pratense)、大叶橐吾(Ligularia macrophylla)等。盖度85%-100%, 草层高45-85 cm, 鲜草产量4β300-9β050 kg·hm-2。土壤为黑钙土, 0-5 cm土壤有机碳含量为7.91%-8.64%, 全氮、全磷、全钾含量分别为9.53、2.16和12.20 g·kg-1, 碱解氮、有效磷、有效钾含量分别为459.20、144.43和473.58 mg·kg-1 (郑伟等, 2015)。退化山地草甸植被特征为: 植被主要成分有野苜蓿(Medicago falcata)、草地早熟禾(Poa pratensis)、拂子茅(Calamagrostis epigeios)、短柄薹草(Carex pediformis)、直立委陵菜(Potentilla recta)、野草莓(Fragaria vesca)、千叶蓍(Achillea millefolium)等, 盖度75%, 草层高度25 cm, 鲜草产量1β300 kg·hm-2。0-5 cm土壤有机碳下降为6.03%, 土壤其他理化性质变化较小(王雪娇, 2013)。1.2 样地选择
选择围封的围封草地(NG)、补播草地(RG)、翻耕重建的豆禾混播草地(LG)和农田(CR)为研究对象。其中, 天然草地为退化山地草甸进行围封(围封开始年为2011年, 2012年后开始割草利用); 补播草地为退化山地草甸上补播鸭茅、无芒雀麦、猫尾草、紫苜蓿(Medicago sativa)、驴食草(又名红豆草) (Onobrychis viciifolia)和红车轴草(Trifolium pratense), 豆禾比为5:5 (补播年份为2011年, 2012年后开始割草利用); 豆禾混播草地为退化山地草甸翻耕重建成鸭茅、无芒雀麦、猫尾草、紫苜蓿、驴食草和红车轴草的豆禾混播草地, 豆禾比为5:5 (建植年份为2011年, 2012年后开始割草利用); 2014年将退化山地草甸开垦成农田, 2015年和2016年均种植春小麦, 春小麦每年4月初至4月中旬播种, 9月初至9月中旬收割。1.3 研究方法
1.3.1 土壤呼吸日动态及水热的测定土壤呼吸的测定时间在2015年4月底至9月初和2016年4月底至9月初, 用LI-8100A (LI-COR, Lincoln NE, USA)对各样地进行测定, 在2015-5-29、2015-6-20、2015-7-6、2015-7-27、2015-8-17、2015-9-1; 2016-4-19、2016-5-14、2016-6-1、2016-6-29、2016-7-27、2016-8-13、2016-9-5测定各样地土壤呼吸日变化, 并根据天气预报尽量选择晴朗天气对各样地进行监测。测定前24 h将直径20 cm、高20 cm的PVC基座嵌入土壤中, 每样地设置3个相距10 m,露出地面5 cm的PVC基座; 同时, 将基座内的绿色植物齐地面剪掉, 但尽量不破坏土壤, 以减少土壤扰动及根系损伤对测量结果的影响。经24 h的平衡后, 在一天内9个时间点(北京时间6:00、8:00、10:00、12:00、14:00、16:00、18:00、20:00、22:00)测定PVC基座内的土壤呼吸速率。
同时, 用与预先埋下的针式地温计同步测定各样地5 cm地温。土壤含水量用烘干铝盒法测定, 取样方法为在测定土壤呼吸的同时, 取相应样地3处PVC基座附近0-10 cm土壤并混匀后装入铝盒带回实验室。
1.3.2 土壤微生物数量的测定
土壤微生物数量测定采用平板涂抹接种法, 以牛肉膏蛋白胨培养基培养细菌高氏1号培养基加重铬酸钾(3.30 mL·L-1)培养放线菌; 马丁氏培养基加链霉素(3.30 mL·L-1)培养真菌。培养3-6天观察(许光辉, 1986)。
1.3.3 土壤微生物生物量碳的测定
土壤采样于每次日变化测量结束后进行, 在各地段随机选取3个土壤采样点, 取样深度为0-5 cm, 从中取约100 g土壤进行预培养, 其中约30 g做熏蒸处理, 30 g做未熏蒸处理, 30 g测含水量。微生物生物量碳采用TOC分析仪(Elementar vario, Langenselbold, Germany)测定, 熏蒸试剂为氯仿。
1.3.4 土壤酶活性的测定
脲酶活性的测定采用奈氏比色法, 以1 g干土 24 h生成的NH3-N量为脲酶1个活性单位; 过氧化氢酶活性的测定采用KMnO4滴定法, 酶活性以1 g干土1 h内消耗的0.1 mol·L-1 KMnO4体积数(以mL计)表示; 转化酶活性的测定采用3,5-二硝基水杨酸比色法, 酶活性以24 h后1 g土壤葡萄糖的mg数表示; 蛋白酶活性的测定采用铜盐比色法, 以1 g土壤在 24 h内酶解蛋白质释放的质量代表蛋白酶的活性(关松荫, 1986)。
1.3.5 数据分析
采用指数模型分析土壤呼吸和土壤温度月变化的关系, 采用一元回归模型分析土壤呼吸和土壤含水量月变化关系, 方程如下:
式中, Rs为土壤呼吸速率(mg·m-2·h-1), a、b、c、d为拟合参数, Ts为5 cm深度的土壤温度(℃), Q10为土壤呼吸的温度敏感性系数, 指土壤温度每增加10 ℃土壤呼吸速率所增加的倍数, W10为10 cm深度的土壤体积含水量。
复合模型能较好地模拟土壤呼吸对温度和土壤含水量的响应, 土壤温度和土壤含水量对土壤呼吸速率的共同影响用多元非线性回归分析模型计算, 方程如下:
式中a、b、c、d、f为拟合参数。
利用Microsoft Excel 2010进行数据整理, 使用SPSS 20.0进行数据分析, 采用单因素方差分析方法分析比较4种处理样地不同时期的土壤呼吸速率。利用Origin Pro 8和Microsoft Excel 2010作图。
2 结果和分析
2.1 各样地不同时期土壤呼吸的变化特征
RG、LG、NG和CR土壤呼吸速率均表现出明显的季节变化, 且有着较为相似的变化规律, 即在生长季初期的4-5月最低, 在6-8月达到最高, 之后逐渐降低。但各处理间的季节变化特征又有差异。2015年各处理土壤呼吸速率季节变化表现为NG和CR均为双峰曲线, 在6月出现一个峰值, 然后下降, 这个下降的过程可能由于这个时期温度下降, 随之土壤呼吸速率下降, NG与CR的土壤呼吸对温度的响应的更加迅速, 因此变化更为明显。NG与CR处理在7月又上升并达最大值, NG出现当年监测土壤呼吸速率的最大值16.41 μmol·m-2·s-1, 随后下降; RG与LG土壤呼吸季节变化呈现单峰曲线, 最大值分别为13.63和11.37 μmol·m-2·s-1。2016年各处理土壤呼吸速率季节变化均呈现单峰曲线, 但各处理出现最大值的时间不同, RG和LG土壤呼吸速率在6月底达到最大值, 分别为16.22和17.97 μmol·m-2·s-1, 而NG与CR在7月底达到最大值, 分别为19.68和17.84 μmol·m-2·s-1; 2015年平均土壤呼吸速率由大到小依次为NG (10.44 μmol·m-2·s-1)、CR (9.61 μmol·m-2·s-1)、RG (8.65 μmol·m-2·s-1)、LG (8.49 μmol·m-2·s-1)。2016年平均土壤呼吸速率由大到小依次为NG (11.06 μmol·m-2·s-1)、RG (10.05 μmol·m-2·s-1)、LG (9.79 μmol·m-2·s-1)、CR (9.70 μmol·m-2·s-1); 2年监测的土壤呼吸速率平均值由大到小依次为: NG (10.83 μmol·m-2·s-1)、RG (9.53 μmol·m-2·s-1)、CR (9.46 μmol·m-2·s-1)、LG (9.19 μmol·m-2·s-1)。对不同时期4种样地的土壤呼吸速率变化进行方差分析显示, 在2015年4月CR与NG存在显著差异(p < 0.05), 与其他处理差异不显著(p > 0.05); 6月RG与NG、CR均存在显著差异(p < 0.05), 而CR与其他处理也存在显著差异(p < 0.05); 在7月初和7月底 均表现为, CR与其他3个处理存在显著差异 (p < 0.05), 而其他3个处理间差异不显著(p > 0.05); 在8月, NG与RG、LG存在显著差异(p < 0.05), 与CR差异不显著(p > 0.05), RG与LG差异不显著(p > 0.05); 9月各处理均无显著差异(p > 0.05)。在2016年4月, CR与其他各处理存在显著差异(p < 0.05), 而其他各处理无显著差异(p > 0.05); 而5月、6月初、6月底各处理均无显著差异(p > 0.05); 在7月底NG与RG、LG存在显著差异(p < 0.05)、与CR差异不显著(p > 0.05); 在8月, RG与NG存在显著差异(p < 0.05), 与其他处理差异不显著(p > 0.05), NG其他处理差异也不显著(p > 0.05); 在9月初各处理差异均不显著(图1)。
图1
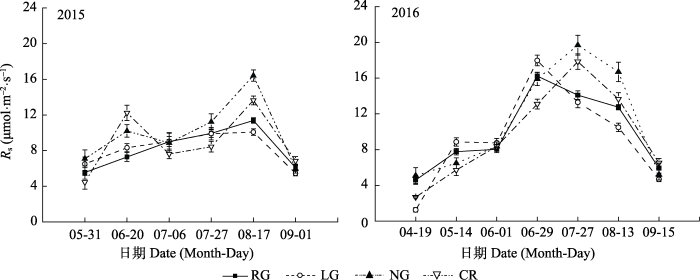 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同土地利用方式下土壤呼吸速率(Rs)季节变化(平均值±标准偏差)。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 1Seasonal variation of soil respiration rate (Rs) in plots with different land use types (mean ± SD). CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
2.2 土壤温度和水分对土壤呼吸速率的影响
不同利用方式下各样地土壤温度(图2)呈单峰曲线, 在7-8月达到最大值, 利用模型(1)对各样地0-5 cm处土壤温度(Ts)与土壤呼吸速率做相关性分析(图3), RG、LG、NG样地土壤呼吸速率均和土壤温度相关性达显著水平(p < 0.05), CR不显著(p > 0.05); 利用模型(2)计算温度敏感指数Q10, 各样地变化为NG > CR > RG > LG (图3)。各样地土壤含水量与相应土壤呼吸速率相关关系利用模型(3)进行模拟, 结果表明RG、LG、NG和CR土壤含水量与土壤呼吸速率均呈负相关关系(图4); 除CR的土壤含水量与土壤呼吸速率相关性不显著外, 其余各样地土壤呼吸速率均与土壤含水量显著相关, 且NG 达到极显著。决定系数(R2)的大小为NG > LG > RG > CR。图2
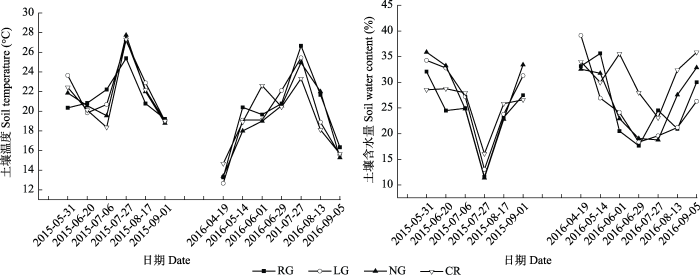 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同土地利用方式下各样地土壤温度和土壤含水量。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 2Soil temperature and soil water content in plots with different land use types. CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
图3
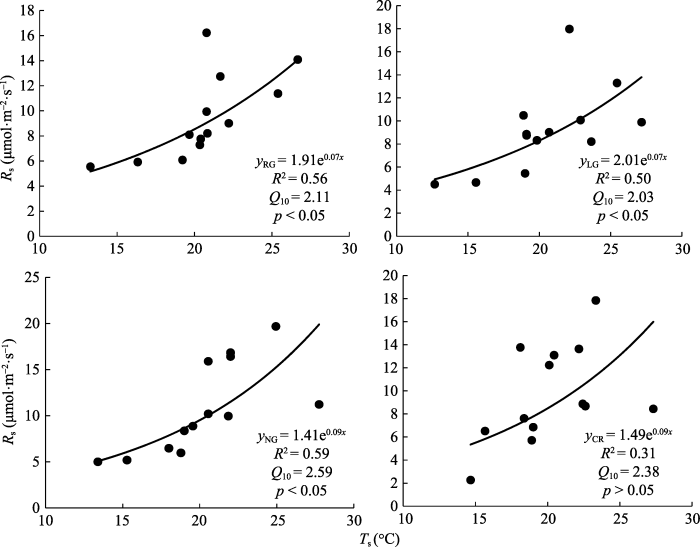 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同土地利用方式下土壤呼吸速率(Rs)与5 cm土壤温度的指数拟合关系。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 3The relationship between soil respiration rate (Rs) and soil temperature at 5 cm depth in plots with different land use types. CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
图4
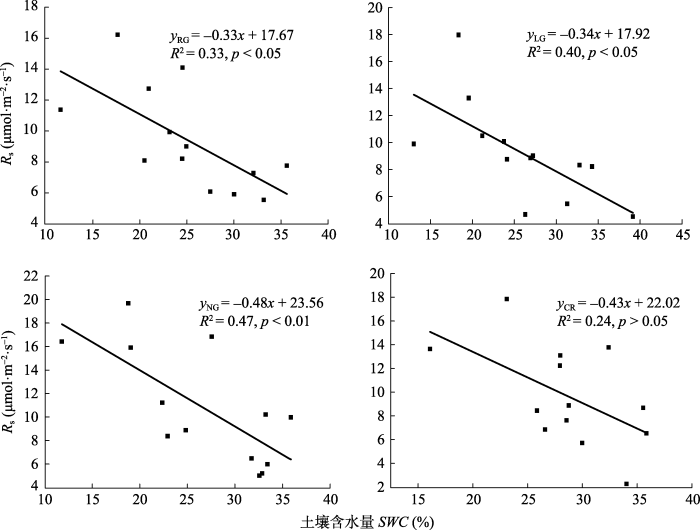 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同土地利用方式下土壤呼吸速率(Rs)与10 cm土壤含水量的线性拟合关系。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 4The relationship between soil respiration rate (Rs) and soil water content (SWC) at 10 cm depth in plots with different land use types. CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
复合模型能较好地模拟土壤呼吸对土壤温度和土壤含水量变化的协同响应, 因此采用复合模型(4)计算土壤温度和水分对土壤呼吸的协同影响(表1), 结果表明, R2较单因子模型均有不同程度的提高, 拟合效果更好, 土壤温度和水分共同解释土壤呼吸速率变异度的64%-89%。
Table 1
表1
表1土壤呼吸(Rs)与土壤温度(Ts)和土壤含水量(W10)的回归方程
Table 1
| 样地 Plot | 2015 | 2016 | ||||
|---|---|---|---|---|---|---|
| 拟合方程 Fitted equation | R2 | p | 拟合方程 Fitted equation | R2 | p | |
| RG | Rs = 0.63e0.10Ts +$0.47W_{10}^{2}$- 0.11W10 - 1.99 | 0.81 | < 0.001 | Rs = e-0.99Ts -$0.60W_{10}^{2}$+ 0.63W10 - 0.73 | 0.68 | < 0.01 |
| LG | Rs = 0.92e0.09Ts +$1.22W_{10}^{2}$- 0.02W10 - 12.45 | 0.67 | < 0.01 | Rs = e-0.99Ts +$0.45_{10}^{2}$- 0.24W10 - 0.97 | 0.89 | < 0.01 |
| NG | Rs = e-0.95Ts -$0.49W_{10}^{2}$+ 0.49W10 + 0.47 | 0.78 | < 0.001 | Rs = 0.46e0.14Ts -$0.02W_{10}^{2}$+ 0.64W10 - 1.73 | 0.88 | < 0.01 |
| CR | Rs = e-0.99Ts -$0.79W_{10}^{2}$+ 0.85W10 - 0.29 | 0.68 | < 0.01 | Rs = e-Ts +$0.86W_{10}^{2}$- 0.86W10 + 0.42 | 0.64 | < 0.01 |
新窗口打开|下载CSV
2.3 土壤微生物特征对土壤呼吸的影响
不同土地利用方式对土壤微生物数量的影响如表2所示, 从土壤微生物类群组成来看, 不同处理土壤微生物数量以细菌为主, 放线菌次之, 真菌数量最少, 细菌数量在土壤微生物组成中占绝对优势。各类群微生物数量占微生物总数比例由大到小依次为细菌>放线菌>真菌。Table 2
表2
表2不同土地利用方式下土壤微生物区系构成(cfu·g-1 dry soil)(平均值±标准偏差, n = 3)
Table 2
| 时期 Time | 样地 Plot | 总数量(105·g-1干土) Total abundance (105·g-1 dry soil) | 细菌 Bacteria | 放线菌 Actinomyces | 真菌 Fungi | |||
|---|---|---|---|---|---|---|---|---|
| 数量(105·g-1干土) Abundance (105·g-1 dry soil) | 比例 Proportion (%) | 数量(105·g-1干土) Abundance (105·g-1 dry soil) | 比例 Proportion (%) | 数量(105·g-1干土) Abundance (105·g-1 dry soil) | 比例 Proportion (%) | |||
| 4月 April | RG | 12.02 | 11.81 ± 0.58a | 98.22 | 1.75 ± 1.21b | 1.46 | 0.03 ± 2.91b | 0.32 |
| LG | 12.15 | 11.51 ± 1.73a | 94.67 | 1.99 ± 3.53b | 1.64 | 0.44 ± 1.76c | 3.69 | |
| NG | 19.39 | 19.19 ± 4.26c | 98.94 | 1.55 ± 1.15a | 0.80 | 0.04 ± 1.76b | 0.26 | |
| CR | 14.55 | 13.69 ± 1.73b | 94.24 | 8.26 ± 5.24c | 5.68 | 0.01 ± 0.33a | 0.08 | |
| 5月 May | RG | 16.46 | 16.08 ± 1.64b | 98.03 | 3.33 ± 1.53a | 1.66 | 0.04 ± 1.26a | 0.31 |
| LG | 13.36 | 12.69 ± 2.09a | 95.98 | 4.06 ± 3.98 ab | 1.83 | 0.03 ± 2.02b | 2.19 | |
| NG | 18.04 | 17.65 ± 4.51b | 98.65 | 3.21 ± 3.69a | 0.99 | 0.06 ± 2.69a | 0.36 | |
| CR | 14.53 | 13.58 ± 2.94a | 94.41 | 6.59 ± 3.02c | 5.19 | 0.03 ± 1.05b | 0.40 | |
| 6月 June | RG | 15.56 | 15.08 ± 3.07b | 96.42 | 4.79 ± 3.64b | 2.63 | 0.01 ± 1.34a | 0.95 |
| LG | 13.22 | 12.47 ± 1.33a | 95.21 | 2.51 ± 4.01a | 2.79 | 0.04 ± 3.17c | 2.00 | |
| NG | 16.98 | 16.43 ± 2.77b | 95.83 | 5.03 ± 2.79b | 2.40 | 0.02 ± 5.69b | 1.77 | |
| CR | 14.21 | 13.24 ± 2.50a | 93.29 | 4.91 ± 3.84b | 3.42 | 0.03 ± 3.89c | 3.29 | |
| 7月初 Early of July | RG | 16.86 | 15.59 ± 4.11c | 92.50 | 8.34 ± 1.31b | 4.93 | 0.04 ± 2.08a | 2.57 |
| LG | 10.44 | 9.43 ± 2.91a | 86.88 | 5.16 ± 4.53a | 7.12 | 0.05 ± 2.68a | 6.00 | |
| NG | 17.51 | 16.49 ± 3.19c | 93.92 | 5.99 ± 6.07a | 3.57 | 0.04 ± 4.16a | 2.51 | |
| CR | 13.83 | 12.91 ± 2.18b | 90.44 | 8.48 ± 2.71b | 6.18 | 0.05 ± 0.88a | 3.38 | |
| 7月末 End of July | RG | 12.88 | 11.84 ± 3.39b | 89.46 | 7.38 ± 3.62b | 7.75 | 0.03 ± 2.44a | 2.79 |
| LG | 5.18 | 5.41 ± 6.88a | 87.18 | 3.89 ± 2.56a | 6.25 | 0.04 ± 5,78a | 6.57 | |
| NG | 17.64 | 16.31 ± 2.87c | 92.47 | 7.22 ± 3.39b | 4.11 | 0.06 ± 0.88b | 3.42 | |
| CR | 12.66 | 11.49 ± 2.68b | 90.56 | 8.67 ± 1.81c | 6.98 | 0.03 ± 2.37a | 2.46 | |
| 8月 August | RG | 12.16 | 11.46 ± 11.55b | 93.87 | 5.79 ± 5.41b | 4.99 | 0.01 ± 1.73a | 1.14 |
| LG | 8.79 | 8.21 ± 8.13a | 90.53 | 4.71 ± 5.17b | 7.75 | 0.01 ± 2.57a | 1.72 | |
| NG | 11.96 | 11.58 ± 1.58b | 96.77 | 2.67 ± 4.01a | 2.27 | 0.01 ± 1.53a | 0.96 | |
| CR | 10.51 | 9.60 ± 0.70a | 89.78 | 7.91 ± 3.08c | 8.81 | 0.01 ± 2.35a | 1.41 | |
| 9月 September | RG | 15.61 | 15.14 ± 4.57b | 97.16 | 4.21 ± 2.97b | 2.56 | 0.04 ± 4.82a | 0.28 |
| LG | 12.92 | 12.71 ± 3.94b | 98.40 | 1.72 ± 2.54a | 1.33 | 0.04 ± 0.88a | 0.27 | |
| NG | 10.51 | 9.77 ± 7.97a | 86.48 | 6.98 ± 2.21c | 12.59 | 0.04 ± 1.47a | 0.93 | |
| CR | 7.74 | 7.15 ± 2.78a | 82.93 | 5.65 ± 1.81d | 16.53 | 0.03 ± 1.22a | 0.54 | |
新窗口打开|下载CSV
各样地总微生物数量存在一定的季节变化, 表现为RG随时间的变化数量不断上升并在7月初达到最大值(16.86 × 105·g-1干土), 而最小值出现在4月初, 温度较低可能是影响微生物生物量较低的原因; LG总微生物数量随时间的变化有先增加后降低再增加的趋势, 其最大值出现在5月(13.36 × 105·g-1干土), 而最小值出现在7月初(5.18 × 105·g-1干土); NG随时间变化出现先降低后增加再降低的趋势, 其最小值出现在9月(10.51 × 105·g-1干土); CR总微生物生物量从翻耕后随时间的变化始终呈明显的递减趋势, 最大值出现在4月(12.02 × 105·g-1干土), 最小值出现在9月(7.74 × 105·g-1干土)。微生物各类群受土地利用方式的影响有所不同(表3), 不同类群微生物随时间的变化而发生变化, 细菌的数量在各类群中占绝对多数, 其变化可以反映土地利用方式变化后, 土壤微生物生物量的变化, 本研究中RG和LG在生长季前期和后期均占90%以上, 而生长季旺期占80%, 其变化与土壤温度的变化相反, 与土壤含水量的变化趋势相似。NG和CR则表现为在生长季的前期和中期, 其细菌在总微生物群落中均占90%以上, 而到生长季后期则在80%以上90%以下, 其变化与土壤温度变化相似。为探明各区系对土壤呼吸的影响以及土壤呼吸变化与土壤微生物协同变化的趋势, 将单位体积土壤细菌、真菌和放线菌数量与相应样地土壤呼吸速率进行拟合分析, 如图5所示, RG土壤呼吸与放线菌呈显著的线性相关(p < 0.05)与细菌和真菌的相关性不显著(p > 0.05)。LG土壤呼吸与细菌和放线菌呈显著线性相关(p < 0.05)与真菌无线性相关; NG与CR的土壤呼吸与各微生物数量均无显著线性关系。
Table 3
表3
表3不同土地利用方式下各样地土壤微生物数量和微生物生物量碳比较(平均值±标准偏差, n = 3)
Table 3
| 指标 Index | LG | RG | NG | CR |
|---|---|---|---|---|
| 细菌 Bacteria (105·g-1 dry soil) | 14.17 ± 3.95bc | 10.31 ± 3.91a | 15.02 ± 4.93c | 11.25 ± 3.90ab |
| 放线菌 Actinomyces (105·g-1 dry soil) | 5.34 ± 24.42a | 3.55 ± 20.18a | 4.91 ± 30.91ab | 7.13 ± 19.87b |
| 真菌 Fungi (105·g-1 dry soil) | 0.02 ± 16.18a | 0.03 ± 18.42b | 0.02 ± 23.01b | 0.02 ± 19.45b |
| 微生物生物量碳 Microbial biomass carbon (mg·kg-1) | 82.15 ± 26.61a | 82.12 ± 29.26a | 97.41 ± 32.92b | 97.72 ± 35.79b |
新窗口打开|下载CSV
图5
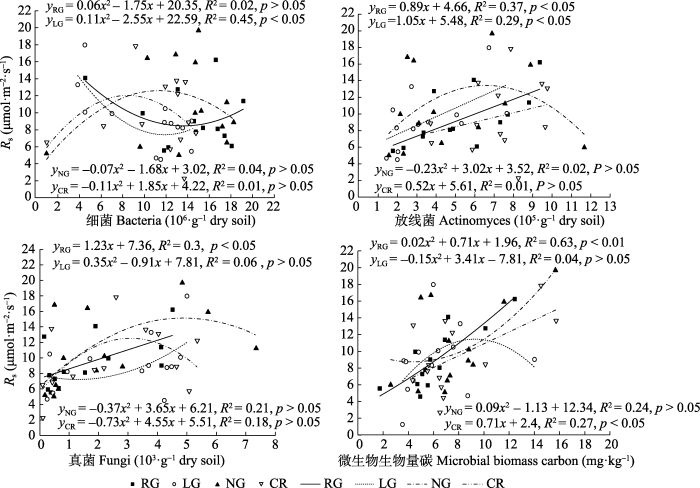 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同利用方式下土壤呼吸速率(Rs)与微生物区系、微生物生物量碳的相关关系(平均值±标准偏差)。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 5The relationship between soil respiration rate(Rs) and microflora, microbial biomass carbon in four plots with different land use types (mean ± SD). CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
不同利用方式下土壤微生物生物量碳如图6所示, 各处理随季节的温度和湿度的变化有较明显的变化, 但各处理微生物生物量碳最大值和最小值出现的时间有所不同, RG的最大值出现在2015年8月和2016年6月, 最小值出现在2016年6月初和2015年5月底。LG的最大值出现在2015年7月初和2016年9月, 最小值出现在2015年9月和2016年4月; NG的最大值出现在2015年6月和2016年7月底, 最小值出现在2015年8月和2016年9月; CR最小值出现在2015年7月初和2016年6月初, 最大值出现在2015年7月底和2016年8月。单因素方差分析的结果(表3)表明, LG与NG和CR存在显著差异, RG与NG和CR也存在显著差异, 其余差异不明显; 其平均值为CR > NG > LG > RG。
图6
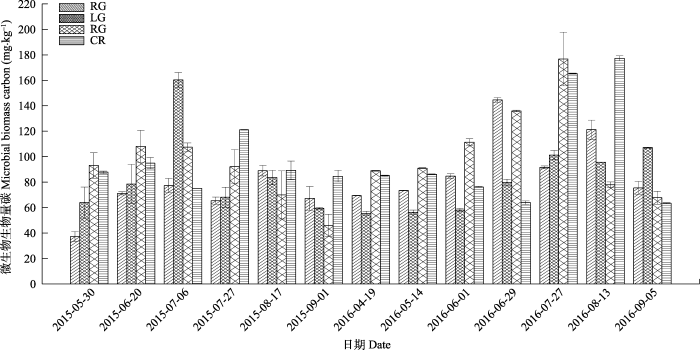 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同利用方式下土壤微生物生物量碳(平均值±标准偏差)。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 6Soil microbial biomass carbon in four plots with different land use types (mean ± SD). CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
微生物生物量碳表征微生物的总量, 各样地植被结构不同, 土壤表面覆盖度不同, 使土壤温度和湿度存在差异, 从而影响微生物碳的变化。各样地土壤呼吸速率与微生物生物量碳拟合分析(图5)显示, RG与CR的土壤呼吸速率与微生物生物量碳显著线性相关(p < 0.05), 其中CR达到极显著(p < 0.01), LG与NG相关性不明显。
2.4 土壤酶活性变化对土壤呼吸的影响
不同利用方式下土壤各时期酶活性如图7所示, 各样地各酶活性随季节的变化呈现一定的变化; 土壤过氧化氢酶在2015年, RG、LG、NG、CR均呈现先增加后降低的趋势, 在2016年, LG与NG呈现由低到高再到低的过程, RG表现为先增加后降低的趋势, NG翻耕后过氧化氢酶出现明显降低, 各样地两年监测期过氧化氢酶平均值大小为CR > NG > RG > LG; 脲酶活性RG、LG与NG在两年生长季内均先增加后降低, CR则在翻耕后酶活性降低, 在生长季旺期出现极低值, 两年脲酶活性平均值由大到小为CR > NG > LG > RG; 蔗糖酶呈现明显的季节变化, 均在生长季旺期达到最大值, 其大小为CR > NG > RG > LG。蛋白酶生长季初期和生长季末期变化都较小, 在生长季旺期变化较为剧烈, 其大小为CR > NG > LG > RG (表4)。图7
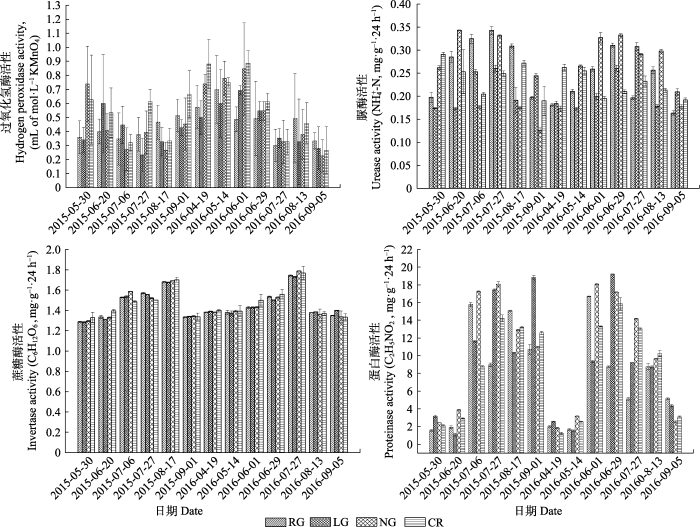 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同利用方式草地土壤酶活性的变化(平均值±标准偏差)。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 7Soil enzyme activities in four plots with different land use types (mean ± SD). CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
Table 4
表4
表4不同土地利用方式下土壤酶活性各样地酶活性比较(平均值±标准偏差, n = 3)
Table 4
| 样地 Plot | 过氧化氢酶 Hydrogen peroxidase (mL of mol·L-1 KMnO4) | 脲酶 Urease (NH4+-N, mg·g-1·24 h-1) | 蔗糖酶 Invertase (C6H12O6, mg·g-1·24 h-1) | 蛋白酶 Proteinase (C2H5NO2, mg·g-1·24 h-1) |
|---|---|---|---|---|
| RG | 0.45 ± 0.11a | 0.22 ± 0.06a | 1.45 ± 0.14a | 7.86 ± 5.53a |
| LG | 0.44 ± 0.14a | 0.23 ± 0.04a | 1.46 ± 0.13a | 8.71 ± 6.43a |
| NG | 0.49 ± 0.22ab | 0.25 ± 0.07a | 1.46 ± 0.15a | 9.04 ± 6.61a |
| CR | 0.64 ± 0.29b | 0.25 ± 0.03a | 1.47 ± 0.14a | 10.17 ± 5.48a |
新窗口打开|下载CSV
将各样地土壤呼吸与土壤酶活性进行拟合相关分析(图8), 土壤过氧化氢酶和脲酶与土壤呼吸速率拟合显示, 各样地拟合参数均较低, 且相关性均未达到显著水平(p > 0.05); 将蛋白酶与土壤呼吸速率线性拟合显示, 除NG相关性达到显著水平(p < 0.05), 其余各样地均未达到显著; 将蔗糖酶与土壤呼吸速率拟合显示, 除LG未达到显著水平(p > 0.05), 其余各样地均达到显著水平。
图8
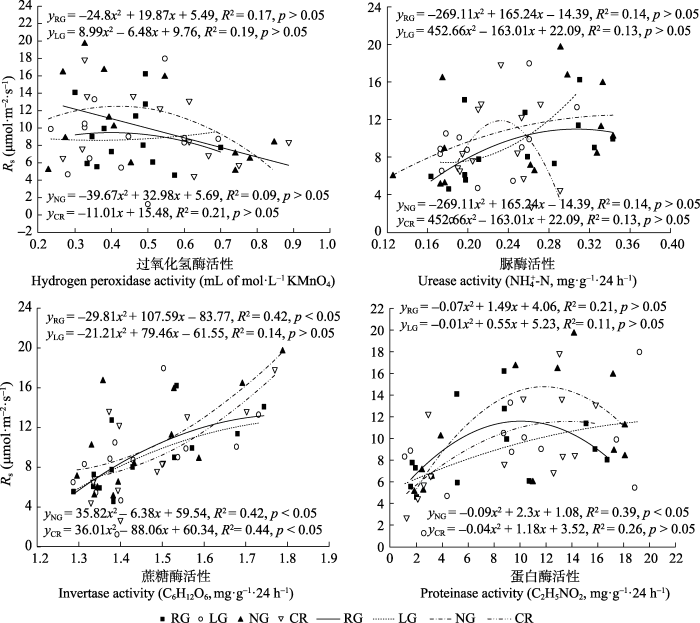 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同土地利用方式下各样地土壤呼吸速率(Rs)与土壤酶活性的相关关系。CR, 农田; LG, 豆禾混播草地; NG, 围封草地; RG, 补播草地。
Fig. 8The relationship between soil respiration rate(Rs) and enzymatic activity in four plots with different land use types. CR, cropland; LG, legume-grass mixture; NG, natural grassland; RG, reseeding grassland.
3 讨论和结论
3.1 不同土地利用方式下土壤呼吸特征及其对水热因子的响应
土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(刘伟等, 2011)。本研究表明, 在土地利用方式变化后, 土壤温度变化表现为LG > RG > NG > CR, 土壤含水量的变化为CR > NG > RG > LG, 土壤呼吸速率年平均值为: NG > RG > CR > LG; 较高的温度可能抑制豆禾混播草地土壤呼吸, 使其土壤呼吸速率最低; 农田由于灌溉的缘故, 保持较低的温度和较高的土壤含水量, 因此抑制土壤呼吸CO2排放; 补播草地的土壤温度低于混播草地、土壤含水量高于混播草地, 平均土壤呼吸速率也高于豆禾混播草地; 农田的土壤呼吸速率低于围封草地。这与郭慧敏等(2014)的研究结果不同, 其原因是本试验样地土壤温度和土壤含水量有关: 本区域常年雨水充足, 土壤水分含量较高, 当土壤含水量饱和后, 就会抑制土壤呼吸。李文等(2015)也报道了高寒草甸草原土壤质量含水量对土壤呼吸速率的影响存在一个临界值, 30%左右为临界值, 在此之前二者呈正相关关系, 此后呈负相关关系。另外, 各处理样地的土壤呼吸速率的季节变化表现为: 在生长季初期和生长季末期均较低, 而在生长旺季土壤呼吸速率达到一年中的最大值。这与温度的季节变化相一致, 因此温度的变化仍然是土壤呼吸变化的重要决定因素。影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(亓伟伟等, 2012)。本研究发现, 各处理5 cm土壤温度与土壤呼吸指数拟合除农田不显著外, 其余均达到显著水平; 就拟合效果而言, 围封草地土壤呼吸速率与温度的拟合效果最好(R2 = 0.59), 其次为补播草地(R2 = 0.56)和豆禾混播草地(R2 = 0.50), 而农田温度与土壤呼吸的拟合效果最差, 这在一定程度上表明, 土壤利用方式变化后, 补播草地和豆禾混播草地对温度变化较为敏感, 因为土壤呼吸通常对最限制它的因子产生响应, 即当土壤水分含量较低时, 土壤呼吸对温度的反应不敏感; 当温度变低时, 土壤呼吸对土壤含水量的响应不敏感(杨阳等, 2012)。农田的土壤呼吸速率与土壤含水量和温度相关性均不明显, 说明含水量和温度均不是其限制因子; 从土壤呼吸速率与土壤温度和土壤湿度的拟合关系可以发现, 土地利用方式变化后, 围封草地土壤呼吸速率与水热因子的拟合效果均高于豆禾混播草地和补播草地。由此可以说明, 由于补播和人工重建使草地对水热因子的响应相对围封草地均有所降低, 从而降低了草地对热量和水分的依赖。有研究表明土壤水分与土壤呼吸呈线性或二次正相关关系(李志刚和侯扶江, 2010; 王铭等, 2014), 但本研究发现土壤水分对土壤呼吸的影响呈显著的线性负相关关系, 这与本区域降水充足, 土壤含水量较高有关; 在较为干旱的地区, 降水导致的土壤含水量升高可能会提高土壤呼吸速率(Zhang et al., 2015), 但本区域土壤水分长期充足, 水分减少反而能增加土壤呼吸速率, 因此土壤呼吸速率与土壤温度表现为负相关。复合模型能很好地解释水热对土壤呼吸速率的协同影响, 因此本研究通过建立土壤呼吸与土壤温度和土壤水分间的复合模型, 拟合关系结果表明, 土壤温度和水分共同解释了土壤呼吸变异度的64%-89%, 土壤温度和土壤含水量是土壤呼吸的主要影响因素。
总体而言, Q10与温度呈负相关关系, 即在温度上升相同幅度下低温区比高温区有更大的Q10 (陈骥等, 2014), 同时, 土壤干旱也会降低Q10值, 一定范围内随着土壤含水量的增加, 土壤呼吸对温度的敏感性也会增加(Kirschbaum, 2006)。土地利用方式的变化引起上层植被和植被盖度的变化, 从而改变土壤温度, 使Q10随之发生变化, Q10变化则促使土壤呼吸对温度的响应更加迅速; 本研究中土地利用方式变化后, 围封草地的Q10数值最高, 其次是农田, 豆禾混播草地最低。农田和围封草地Q10较高, 也表明在土地利用方式变化后其对温度变化的响应较为敏感, 而温度对补播草地和豆禾混播草地的影响较小。
3.2 不同土地利用方式下土壤微生物特征及其与土壤呼吸速率的关系
土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(Su et al., 2005)。在本研究中, 各样地土壤微生物数量以细菌占绝对优势, 放线菌次之, 真菌数量最少, 这与大多数****的研究结果(张文婷等, 2008; 张成霞和南志标, 2010)一致。总微生物生物量则表现为NG > RG > CR > LG, 这与两年监测期平均土壤呼吸排放速率大小完全一致。本研究中细菌在微生物群落中占80%-90%, 因此各土地利用方式对细菌数量的影响也最大, 而细菌的变化一定程度上也是土壤呼吸变化的主要原因。本研究中各处理微生物生物量碳的大小为CR > NG > LG > RG, 微生物生物量碳含量的提高, 标志着土壤微生物活性得到一定程度的改善, 可以促进土壤团聚体的形成, 加快动植物残体的分解速度和养分的转化进程, 微生物生物量的大小主要取决于输入土壤的物质和质量(蒋德明等, 2006); 本研究结果表明, 补播草地和农田的土壤呼吸速率与土壤微生物生物量碳有密切关系, 说明微生物对这两种土地利用方式的土壤呼吸速率有一定的影响。农田微生物生物量碳的含量较高是由于农田翻耕和施肥, 且历年收割后秸秆还田导致微生物生物量碳增加。3.3 不同土地利用方式下土壤酶活性特征及其与土壤呼吸的关系
土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(陈书涛等, 2016)。本研究各样地4种土壤酶活性的高低均表现为农田最大、围封草地次之, 豆禾混播草地和补播草地最小, 农田和围封草地土壤酶活性的较高也在一定程度上解释了其土壤呼吸速率较高的原因, 而补播草地和豆禾混播草地较低的土壤酶活性也是其土壤呼吸速率较低的一个重要原因。农田酶活性较高可能是由于本研究样地采用秸秆还田的方式, 在一定程度上保持了土壤肥力, 为土壤微生物提供了营养, 间接促进了土壤酶活性; 研究表明增温、秸秆施用均会提高土壤微生物的数量和活性(Rinnan et al., 2009; 孙建等, 2009; 王学娟等, 2014)。土壤呼吸与酶活性的线性相关分析表明: 蛋白酶与围封草地的相关性达到显著水平, 蔗糖酶与农田、围封草地和补播草地的土壤呼吸均有显著相关性, 而豆禾混播草地与酶活性均无显著相关性。土壤蔗糖酶能催化糖的水解过程, 与土壤碳循环过程有关, 以往也有研究验证了土壤微生物呼吸与蔗糖酶具有相关关系(吴静等, 2015)。蛋白酶是参与土壤氮素循环的主要酶, 其活性大小直接影响土壤养分, 从而影响土壤微生物呼吸速率的大小。因此, 4种土地利用方式影响了天山北坡西段山地草甸的水热和生物因素, 从而导致土壤呼吸速率的改变。其中, 水热因素和温度敏感性(Q10)对围封草地和农田土壤呼吸速率影响较大, 而对豆禾混播草地和补播草地影响较小; 围封草地和农田也具有较高的微生物生物量和土壤酶活性。故而, 退化的山地草甸在不同土地利用方式下土壤呼吸速率表现为围封草地>补播草地>农田>豆禾混播草地, 即豆禾混播草地和补播草地相对于围封草地和农田, 显著降低了土壤呼吸速率, 提高了草地土壤的固碳能力。因此在昭苏地区, 对退化山地草甸的合理利用和生态保护措施中, 建植人工草地和补播草地是较好的选择。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.2307/3899755URL [本文引用: 1]

Abstract Research on organic matter in semiarid and grassland soils is reviewed in terms of carbon additions, decomposition rates, organic matter quality, the effects of cultivation, translocation of humic materials, humification and the role of organic matter in carbon cycles. Farming systems which minimise tillage, maintain soil cover and use soil moisture efficiently benefit both these soils and the producer. -R.H.Johnson
DOI:10.11686/cyxb20140610URLMagsci [本文引用: 1]
非生长季土壤呼吸是生态系统碳循环的重要组成部分,显著地影响着碳收支。本研究利用Li-8100开路式碳通量测定系统,研究了青海湖北岸高寒草甸草原非生长季土壤呼吸对短期围栏封育(自由放牧,3年围栏封育和5年围栏封育样地)的响应,并讨论了温度和湿度对非生长季土壤呼吸的影响。结果表明,1)自由放牧、3年围栏封育和5年围栏封育样地非生长季土壤呼吸平均速率分别是全年土壤呼吸平均速率的0.21,0.22和0.19倍;2)自由放牧、3年围栏封育和5年围栏封育样地非生长季土壤呼吸所排放的碳量分别为117.2,109.2和100.7 g C/m<sup>2</sup>,占全年土壤呼吸所排放的碳量的21.2%,22.3%和23.2%;3)相比于生长季和全年土壤呼吸的温度敏感性,非生长季具有更低的土壤呼吸温度敏感性;4)非生长季土壤温度和土壤湿度对土壤呼吸的解释率相当,一定的土壤湿度是保证土壤呼吸对温度响应的必要条件。因此,当考虑到碳收支和生态系统碳循环时不能忽略掉非生长季土壤呼吸的作用,而且水分在调节非生长季土壤呼吸中起着重要的作用。
DOI:10.11686/cyxb20140610URLMagsci [本文引用: 1]
非生长季土壤呼吸是生态系统碳循环的重要组成部分,显著地影响着碳收支。本研究利用Li-8100开路式碳通量测定系统,研究了青海湖北岸高寒草甸草原非生长季土壤呼吸对短期围栏封育(自由放牧,3年围栏封育和5年围栏封育样地)的响应,并讨论了温度和湿度对非生长季土壤呼吸的影响。结果表明,1)自由放牧、3年围栏封育和5年围栏封育样地非生长季土壤呼吸平均速率分别是全年土壤呼吸平均速率的0.21,0.22和0.19倍;2)自由放牧、3年围栏封育和5年围栏封育样地非生长季土壤呼吸所排放的碳量分别为117.2,109.2和100.7 g C/m<sup>2</sup>,占全年土壤呼吸所排放的碳量的21.2%,22.3%和23.2%;3)相比于生长季和全年土壤呼吸的温度敏感性,非生长季具有更低的土壤呼吸温度敏感性;4)非生长季土壤温度和土壤湿度对土壤呼吸的解释率相当,一定的土壤湿度是保证土壤呼吸对温度响应的必要条件。因此,当考虑到碳收支和生态系统碳循环时不能忽略掉非生长季土壤呼吸的作用,而且水分在调节非生长季土壤呼吸中起着重要的作用。
DOI:10.13227/j.hjkx.2016.02.040URL [本文引用: 1]
为研究增温及秸秆施用对冬小麦田土壤呼吸和酶活性的影响,于2014-11~2015-05进行田间试验.设置对照、增温、秸秆施用、增温及秸秆施用这4个处理,观测了不同处理下土壤呼吸、土壤温度、土壤湿度(体积含水量)的季节动态,并在拔节期、孕穗期、扬花期观测了不同处理下的土壤脲酶、转化酶、过氧化氢酶活性.结果表明,对照、增温、秸秆施用、增温及秸秆施用这4个处理的季节平均土壤呼吸速率分别为1.46、1.96、1.92、2.45μmol?(m2?s)-1.方差分析表明,增温处理与对照、秸秆施用处理与对照、增温及秸秆施用处理与对照3对处理之间的季节平均土壤呼吸速率均存在显著差异( P <0.05).不同处理下土壤呼吸与土壤温度的关系均可用指数回归方程拟合,指数回归方程可分别解释对照、增温、秸秆施用、增温及秸秆施用这4个处理土壤呼吸34.3% 、28.1% 、24.6%、32.0%的变异.增温、秸秆施用比对照处理显著(P <0.05)提高了脲酶、转化酶、过氧化氢酶活性.土壤呼吸与脲酶活性存在线性回归关系,其 P 值为0.061,接近显著水平;土壤呼吸与转化酶(P =0.013)、过氧化氢酶活性(P =0.002)均存在极显著的线性回归关系.
DOI:10.13227/j.hjkx.2016.02.040URL [本文引用: 1]
为研究增温及秸秆施用对冬小麦田土壤呼吸和酶活性的影响,于2014-11~2015-05进行田间试验.设置对照、增温、秸秆施用、增温及秸秆施用这4个处理,观测了不同处理下土壤呼吸、土壤温度、土壤湿度(体积含水量)的季节动态,并在拔节期、孕穗期、扬花期观测了不同处理下的土壤脲酶、转化酶、过氧化氢酶活性.结果表明,对照、增温、秸秆施用、增温及秸秆施用这4个处理的季节平均土壤呼吸速率分别为1.46、1.96、1.92、2.45μmol?(m2?s)-1.方差分析表明,增温处理与对照、秸秆施用处理与对照、增温及秸秆施用处理与对照3对处理之间的季节平均土壤呼吸速率均存在显著差异( P <0.05).不同处理下土壤呼吸与土壤温度的关系均可用指数回归方程拟合,指数回归方程可分别解释对照、增温、秸秆施用、增温及秸秆施用这4个处理土壤呼吸34.3% 、28.1% 、24.6%、32.0%的变异.增温、秸秆施用比对照处理显著(P <0.05)提高了脲酶、转化酶、过氧化氢酶活性.土壤呼吸与脲酶活性存在线性回归关系,其 P 值为0.061,接近显著水平;土壤呼吸与转化酶(P =0.013)、过氧化氢酶活性(P =0.002)均存在极显著的线性回归关系.
DOI:10.1016/j.agee.2006.12.024URL [本文引用: 1]
Soil/atmosphere exchange fluxes of nitrous oxide were monitored for a 3-year period at 10 grassland sites in eight European countries (Denmark, France, Hungary, Ireland, Italy, The Netherlands, Switzerland and United Kingdom), spanning a wide range of climatic, environmental and soil conditions. Most study sites investigated the influence of one or several management practices on N 2O exchange, such as nitrogen fertilization and grazing intensity. Fluxes were measured using non-steady state chambers at most sites, and alternative measurement techniques such as eddy covariance and fast-box using tunable diode laser spectroscopy were implemented at some sites. The overall uncertainty in annual flux estimates derived from chamber measurements may be as high as 50% due to the temporal and spatial variability in fluxes, which warrants the future use of continuous measurements, if possible at the field scale. Annual emission rates were higher from intensive than from extensive grasslands, by a factor 4 if grazed (1.77 versus 0.48 kg N 2O-N ha 611 year 611) and by a factor 3 if ungrazed (0.95 versus 0.32 kg N 2O-N ha 611 year 611). Annual emission factors for fertilized systems were highly variable, ranging from 0.01% to 3.56%, but the mean emission factor across all sites (0.75%) was substantially lower than the IPCC default value of 1.25%. Emission factors for individual fertilization events increased with soil temperature and were generally higher for water-filled pore space values in the range 60–90%, though precipitation onto dry soils was also shown to lead to high losses of N 2O-N from applied fertilizer. An empirical, multiple regression model to predict N 2O emission factors on the basis of soil temperature, moisture and rainfall is developed, explaining half of the variability in observed emission factors.
[本文引用: 1]
[本文引用: 1]
DOI:10.11733/j.issn.1007-0435.2012.01.007URL [本文引用: 1]
通过对呼伦贝尔羊草(Leymus chinensis)草甸草原土壤呼吸特性及土壤微生物的测定,研究了3种不同利用方式(围封、放牧和刈割)对天然草地土壤呼吸特性及土壤微生物的影响,为草甸草原土壤呼吸研究提供基础理论依据。结果表明:不同利用方式下土壤呼吸速率、土壤微生物量碳含量与土壤酶活性(除脲酶)均表现为草地围封比放牧和刈割呼吸速率高(含量高、活性强);土壤微生物量氮含量、脲酶活性和微生物数量表现为草地放牧和刈割比围封的含量高(活性强、数量多);土壤微生物量碳、氮含量、土壤酶活性(除脲酶)和土壤微生物数量均以表层(0~10cm)最高(活性最强、数量最多),随着土层深度增加而降低(活性下降、数量减少)。土壤呼吸速率与微生物量碳、土壤酶活性(除过氧化氢酶)成正相关,与微生物量氮、过氧化氢酶和微生物数量成负相关,其中只与微生物量氮相关性显著,而其余之间均不显著。
DOI:10.11733/j.issn.1007-0435.2012.01.007URL [本文引用: 1]
通过对呼伦贝尔羊草(Leymus chinensis)草甸草原土壤呼吸特性及土壤微生物的测定,研究了3种不同利用方式(围封、放牧和刈割)对天然草地土壤呼吸特性及土壤微生物的影响,为草甸草原土壤呼吸研究提供基础理论依据。结果表明:不同利用方式下土壤呼吸速率、土壤微生物量碳含量与土壤酶活性(除脲酶)均表现为草地围封比放牧和刈割呼吸速率高(含量高、活性强);土壤微生物量氮含量、脲酶活性和微生物数量表现为草地放牧和刈割比围封的含量高(活性强、数量多);土壤微生物量碳、氮含量、土壤酶活性(除脲酶)和土壤微生物数量均以表层(0~10cm)最高(活性最强、数量最多),随着土层深度增加而降低(活性下降、数量减少)。土壤呼吸速率与微生物量碳、土壤酶活性(除过氧化氢酶)成正相关,与微生物量氮、过氧化氢酶和微生物数量成负相关,其中只与微生物量氮相关性显著,而其余之间均不显著。
[本文引用: 2]
DOI:10.3321/j.issn:1673-5021.2006.04.004URL [本文引用: 1]
对科尔沁地区碱化草甸草地翻耙补播沙打旺后的土壤理化性质和土壤生物活性的变化进行了研究. 结果表明,翻耙补播改良后草地综合生产性能得到明显改善,土壤容重、pH、含盐量和碱化度降低,土壤含水量和孔隙度增加,土壤有机质和N、P、K等养分含 量增加;同时,土壤微生物C、N的含量以及土壤酶的活性也得到较大幅度提高.
DOI:10.3321/j.issn:1673-5021.2006.04.004URL [本文引用: 1]
对科尔沁地区碱化草甸草地翻耙补播沙打旺后的土壤理化性质和土壤生物活性的变化进行了研究. 结果表明,翻耙补播改良后草地综合生产性能得到明显改善,土壤容重、pH、含盐量和碱化度降低,土壤含水量和孔隙度增加,土壤有机质和N、P、K等养分含 量增加;同时,土壤微生物C、N的含量以及土壤酶的活性也得到较大幅度提高.
DOI:10.1016/j.soilbio.2006.01.030URL [本文引用: 1]
The temperature dependence of organic matter decomposition is of considerable ecosphysiological importance, especially in the context of possible climate-change feedback effects. It effectively controls whether, or how much, carbon will be released with global warming, and to what extent that release of carbon constitutes a dangerous positive feedback effect that leads to further warming. The present paper is an invited contribution in a series of Citation Classics based on a review paper of the temperature dependence of organic matter decomposition that was published in 1995. It discusses the context and main findings of the 1995 study, the progress has been made since then and what issues still remain unresolved. Despite the continuation of much further experimental work and repeated publication of summary articles, there is still no scientific consensus on the temperature dependence of organic matter decomposition. It is likely that this lack of consensus is largely due to different studies referring to different experimental conditions where confounding factors play a greater or lesser role. Substrate availability is particularly important. If it changes during the course of measurements, it can greatly confound the derived apparent temperature dependence. This confounding effect is illustrated through simulations and examples of experimental work drawn from the literature. The paper speculates that much of the current disagreement between studies might disappear if different studies would ensure that they are all studying the same system attributes, and if confounding factors were always considered and, if possible, eliminated.
DOI:10.1126/science.1097396URLPMID:15192216 [本文引用: 1]
Abstract The carbon sink capacity of the world's agricultural and degraded soils is 50 to 66% of the historic carbon loss of 42 to 78 gigatons of carbon. The rate of soil organic carbon sequestration with adoption of recommended technologies depends on soil texture and structure, rainfall, temperature, farming system, and soil management. Strategies to increase the soil carbon pool include soil restoration and woodland regeneration, no-till farming, cover crops, nutrient management, manuring and sludge application, improved grazing, water conservation and harvesting, efficient irrigation, agroforestry practices, and growing energy crops on spare lands. An increase of 1 ton of soil carbon pool of degraded cropland soils may increase crop yield by 20 to 40 kilograms per hectare (kg/ha) for wheat, 10 to 20 kg/ha for maize, and 0.5 to 1 kg/ha for cowpeas. As well as enhancing food security, carbon sequestration has the potential to offset fossil fuel emissions by 0.4 to 1.2 gigatons of carbon per year, or 5 to 15% of the global fossil-fuel emissions.
DOI:10.11686/cyxb2015152Magsci [本文引用: 1]
为探讨不同放牧管理模式对青藏高原东缘高寒草甸草原土壤呼吸速率的影响,于2014年5-10月用土壤呼吸测量仪(LI-8100A,LI-COR,Lincoln,USA)对禁牧(NG)、全生长季休牧(RG)、传统夏季休牧(TG)和全年连续放牧(CG)4种不同放牧管理模式高寒草甸草原土壤呼吸速率进行测定,并分析了土壤呼吸速率与其影响因子间的关系。结果表明:1)不同放牧管理模式高寒草甸草原土壤呼吸速率月动态均表现出先增加后降低的变化态势,峰值出现在8月,除CG外,8月显著高于其余月份;2)在整个生长季,NG, RG和TG平均土壤呼吸速率显著高于CG,其中,NG和RG间无显著差异,但均显著高于TG;3)除CG外,NG, RG和TG土壤呼吸速率与土壤温度呈显著正相关,<em>Q</em><sub>10</sub>值排序为:NG>RG>TG>CG;4)土壤质量含水量对土壤呼吸速率的影响存在一个临界值,30%左右为临界值,在此之前二者呈正相关,此后呈负相关;5)地上、地下生物量均与土壤呼吸速率呈显著正相关。在保障牧民收入稳定的前提下,为实现草地资源的可持续利用与牧草的更新,相对于全年禁牧,全生长季休牧既可充分利用牧草资源,也能有效保护草地系统稳定与生态恢复,是青藏高原高寒草甸草原类草地放牧管理的理想选择。
DOI:10.11686/cyxb2015152Magsci [本文引用: 1]
为探讨不同放牧管理模式对青藏高原东缘高寒草甸草原土壤呼吸速率的影响,于2014年5-10月用土壤呼吸测量仪(LI-8100A,LI-COR,Lincoln,USA)对禁牧(NG)、全生长季休牧(RG)、传统夏季休牧(TG)和全年连续放牧(CG)4种不同放牧管理模式高寒草甸草原土壤呼吸速率进行测定,并分析了土壤呼吸速率与其影响因子间的关系。结果表明:1)不同放牧管理模式高寒草甸草原土壤呼吸速率月动态均表现出先增加后降低的变化态势,峰值出现在8月,除CG外,8月显著高于其余月份;2)在整个生长季,NG, RG和TG平均土壤呼吸速率显著高于CG,其中,NG和RG间无显著差异,但均显著高于TG;3)除CG外,NG, RG和TG土壤呼吸速率与土壤温度呈显著正相关,<em>Q</em><sub>10</sub>值排序为:NG>RG>TG>CG;4)土壤质量含水量对土壤呼吸速率的影响存在一个临界值,30%左右为临界值,在此之前二者呈正相关,此后呈负相关;5)地上、地下生物量均与土壤呼吸速率呈显著正相关。在保障牧民收入稳定的前提下,为实现草地资源的可持续利用与牧草的更新,相对于全年禁牧,全生长季休牧既可充分利用牧草资源,也能有效保护草地系统稳定与生态恢复,是青藏高原高寒草甸草原类草地放牧管理的理想选择。
DOI:10.11686/cyxb20100107Magsci [本文引用: 2]
<p><font face="Verdana">采用动态密闭气室红外CO<sub>2</sub> 分析法(IRGA)对黄土高原丘陵沟壑区不同地形的封育草地土壤呼吸速率日动态进行了测定,并分析了土壤呼吸与土壤温度、土壤水分、地下生物量和地形间的关系。研究结果显示,草地的土壤呼吸日动态呈单峰曲线,最小值为0.393~1.103μmol/(m<sup>2</sup>·s),出现在凌晨;最大值为1.034~1.405μmol/(m<sup>2</sup>·s),出现在14时左右,可以用正弦曲线拟合;而且不同样地间土壤呼吸速率与土壤温度间的关系均可用指数函数描述。土壤日均呼吸速率在不同地形的草地间存在差异,平坦草地为1.229μmol/(m<sup>2</sup>·s),东北坡15°,30°和45°草地分别为1.138,1.033 和0.890μmol/(m<sup>2</sup>·s);西南坡15°,30° 和45° 草地分别为1.036,0.859 和0.379μmol/(m<sup>2</sup>·s);土壤呼吸速率随坡度的增大而减小,东北坡土壤呼吸速率小于西南坡,与不同样地间土壤水分和地下生物量的变化趋势一致,而且研究结果表明土壤呼吸的这种差异主要是受地形影响的土壤水分和地下生物量空间分布差异性决定的,而不同样地间土壤平均温度差异不显著(<em>P</em>>0.05)。</br></font></p>
DOI:10.11686/cyxb20100107Magsci [本文引用: 2]
<p><font face="Verdana">采用动态密闭气室红外CO<sub>2</sub> 分析法(IRGA)对黄土高原丘陵沟壑区不同地形的封育草地土壤呼吸速率日动态进行了测定,并分析了土壤呼吸与土壤温度、土壤水分、地下生物量和地形间的关系。研究结果显示,草地的土壤呼吸日动态呈单峰曲线,最小值为0.393~1.103μmol/(m<sup>2</sup>·s),出现在凌晨;最大值为1.034~1.405μmol/(m<sup>2</sup>·s),出现在14时左右,可以用正弦曲线拟合;而且不同样地间土壤呼吸速率与土壤温度间的关系均可用指数函数描述。土壤日均呼吸速率在不同地形的草地间存在差异,平坦草地为1.229μmol/(m<sup>2</sup>·s),东北坡15°,30°和45°草地分别为1.138,1.033 和0.890μmol/(m<sup>2</sup>·s);西南坡15°,30° 和45° 草地分别为1.036,0.859 和0.379μmol/(m<sup>2</sup>·s);土壤呼吸速率随坡度的增大而减小,东北坡土壤呼吸速率小于西南坡,与不同样地间土壤水分和地下生物量的变化趋势一致,而且研究结果表明土壤呼吸的这种差异主要是受地形影响的土壤水分和地下生物量空间分布差异性决定的,而不同样地间土壤平均温度差异不显著(<em>P</em>>0.05)。</br></font></p>
URL [本文引用: 1]
土壤是大气中主要温室气体(如CO2、CH4和N2O)重要的源或汇,土地利用方式的改变将会导致土壤相关微环境及其生理生化过程发生改变,从而显著影响土壤中温室气体的产生与排放。在全球变化和土地利用大幅度改变的背景下,国际上已逐步开展了关于土地利用变化对土壤温室气体通量的研究。本文在简要介绍土地利用变化与土壤温室气体通量研究的基础上,重点论述了农田、草地和森林互换、湿地向农田转变、不同土地利用类型(森林、草地、湿地和农田)内部变化对3种土壤温室气体排放的影响,并从3种土壤温室气体产生的关键过程简单阐述其主要影响机理。根据目前研究中存在的不足,提出了今后需要加强的领域,以期更好地揭示土地利用变化对土壤温室气体通量的影响及作用机理,为今后深入开展相关研究提供参考。
URL [本文引用: 1]
土壤是大气中主要温室气体(如CO2、CH4和N2O)重要的源或汇,土地利用方式的改变将会导致土壤相关微环境及其生理生化过程发生改变,从而显著影响土壤中温室气体的产生与排放。在全球变化和土地利用大幅度改变的背景下,国际上已逐步开展了关于土地利用变化对土壤温室气体通量的研究。本文在简要介绍土地利用变化与土壤温室气体通量研究的基础上,重点论述了农田、草地和森林互换、湿地向农田转变、不同土地利用类型(森林、草地、湿地和农田)内部变化对3种土壤温室气体排放的影响,并从3种土壤温室气体产生的关键过程简单阐述其主要影响机理。根据目前研究中存在的不足,提出了今后需要加强的领域,以期更好地揭示土地利用变化对土壤温室气体通量的影响及作用机理,为今后深入开展相关研究提供参考。
DOI:10.3969/j.issn.1671-0815.2014.03.018URL
农牧交错带地区土地利用方式多变,人为的改变土地利用方式会影响 土壤释放CO2的速率.分别对新开垦农田、多年耕种农田、退耕还草草地、多年生草地4种利用类型的土壤温度、土壤有机碳含量、土壤呼吸速率进行监测和研 究,结果表明:4种土地利用类型土壤呼吸速率的日变化图像呈单峰性曲线,其中草地平均呼吸速率最大,为0.85g·m-2·d-1.4种土地利用类型土壤 温度与土壤呼吸速率呈正相关关系,其中新开垦农田变化明显,K值最高,达到0.045.从整个生长季来看,4种土地利用类型平均呼吸速率表现为新开农田> 草地>还草草地>农田,分别为0.97g·m-2·d-1、0.85g·m-2·d-1、0.77g·m-2·d-1和0.56g·m-2·d- 1.0-10cm和10-20cm土层土壤有机碳含量大小关系与土壤呼吸速率基本吻合.试验表明不论对草地进行开垦还是对农田进行退耕还草都会增加土壤 CO2的释放.
DOI:10.3969/j.issn.1671-0815.2014.03.018URL
农牧交错带地区土地利用方式多变,人为的改变土地利用方式会影响 土壤释放CO2的速率.分别对新开垦农田、多年耕种农田、退耕还草草地、多年生草地4种利用类型的土壤温度、土壤有机碳含量、土壤呼吸速率进行监测和研 究,结果表明:4种土地利用类型土壤呼吸速率的日变化图像呈单峰性曲线,其中草地平均呼吸速率最大,为0.85g·m-2·d-1.4种土地利用类型土壤 温度与土壤呼吸速率呈正相关关系,其中新开垦农田变化明显,K值最高,达到0.045.从整个生长季来看,4种土地利用类型平均呼吸速率表现为新开农田> 草地>还草草地>农田,分别为0.97g·m-2·d-1、0.85g·m-2·d-1、0.77g·m-2·d-1和0.56g·m-2·d- 1.0-10cm和10-20cm土层土壤有机碳含量大小关系与土壤呼吸速率基本吻合.试验表明不论对草地进行开垦还是对农田进行退耕还草都会增加土壤 CO2的释放.
DOI:10.3724/SP.J.1258.2011.00275URL [本文引用: 2]
甲烷(CH4)是仅次于CO2的重要温室气体。内蒙古草原是欧亚温带草原的重要类型,具有典 型的生态地域代表性。该文以内蒙古温带典型草原为研究对象,通过人工剔除植物种的方法来确定群落中的植物功能型,并应用静态箱技术,观测土壤CH4的吸 收,以理解植物功能型对土壤CH4吸收的影响。结果表明:1)土壤CH4的吸收受温度和水分变化的影响,具有明显的季节差异,且与温度显著相关。2)在 2008年和2009年所测的大部分月份中,植物功能型的土壤CH4吸收量之间没有显著差异;然而在植物生长旺季(8月),不同植物功能型的土壤CH4吸 收量之间存在显著差异,多年生丛生禾草的土壤CH4吸收量最小。3)处理中一、二年生植物、多年生杂类草的存在能够增加土壤CH4的吸收量,而处理中多年 生根茎类禾草、多年生丛生禾草的存在对土壤CH4吸收的影响不大。这可能是因为,植物功能型影响土壤的微生物代谢和环境因子,进而影响土壤CH4吸收量。 该试验说明,在痕量气体层面上,植物功能型组成在生态系统功能中具有重要作用,特别是群落中的亚优势种和伴生种(一、二年生植物、多年生杂类草),通过调 控土壤微生物和环境因子,对地-气的CH4交换产生重要影响。
DOI:10.3724/SP.J.1258.2011.00275URL [本文引用: 2]
甲烷(CH4)是仅次于CO2的重要温室气体。内蒙古草原是欧亚温带草原的重要类型,具有典 型的生态地域代表性。该文以内蒙古温带典型草原为研究对象,通过人工剔除植物种的方法来确定群落中的植物功能型,并应用静态箱技术,观测土壤CH4的吸 收,以理解植物功能型对土壤CH4吸收的影响。结果表明:1)土壤CH4的吸收受温度和水分变化的影响,具有明显的季节差异,且与温度显著相关。2)在 2008年和2009年所测的大部分月份中,植物功能型的土壤CH4吸收量之间没有显著差异;然而在植物生长旺季(8月),不同植物功能型的土壤CH4吸 收量之间存在显著差异,多年生丛生禾草的土壤CH4吸收量最小。3)处理中一、二年生植物、多年生杂类草的存在能够增加土壤CH4的吸收量,而处理中多年 生根茎类禾草、多年生丛生禾草的存在对土壤CH4吸收的影响不大。这可能是因为,植物功能型影响土壤的微生物代谢和环境因子,进而影响土壤CH4吸收量。 该试验说明,在痕量气体层面上,植物功能型组成在生态系统功能中具有重要作用,特别是群落中的亚优势种和伴生种(一、二年生植物、多年生杂类草),通过调 控土壤微生物和环境因子,对地-气的CH4交换产生重要影响。
URL [本文引用: 1]
在内蒙古太仆寺旗典型草原开展不同干扰方式对土壤理化性质影响的比较研究,结果表明:放牧地与季节性封育草地土壤的容重显著高于割草地(P0.05),而含水量显著低于割草地(P0.05),分析原因为放牧对草地土壤具有明显的压实作用,从而增加土壤的容重,降低土壤对水分的渗透与蓄积能力;季节性封育草地土壤的有机质、全N、全P、全K、速效P、速效K含量显著高于放牧地与割草地,说明目前普遍采用的长期重度放牧利用及连年割草利用都不利于土壤肥力的蓄积及草地资源的可持续利用,而季节性围封禁牧的管理方式能促进家畜粪便的归还及地表凋落物的分解,有利于土壤肥力的增加,但围封时间段的选择有待进一步研究。
URL [本文引用: 1]
在内蒙古太仆寺旗典型草原开展不同干扰方式对土壤理化性质影响的比较研究,结果表明:放牧地与季节性封育草地土壤的容重显著高于割草地(P0.05),而含水量显著低于割草地(P0.05),分析原因为放牧对草地土壤具有明显的压实作用,从而增加土壤的容重,降低土壤对水分的渗透与蓄积能力;季节性封育草地土壤的有机质、全N、全P、全K、速效P、速效K含量显著高于放牧地与割草地,说明目前普遍采用的长期重度放牧利用及连年割草利用都不利于土壤肥力的蓄积及草地资源的可持续利用,而季节性围封禁牧的管理方式能促进家畜粪便的归还及地表凋落物的分解,有利于土壤肥力的增加,但围封时间段的选择有待进一步研究。
DOI:10.5846/stxb201102250215URL [本文引用: 1]
低温被广泛认为是高寒草甸生态系统首要限制性因子,因此增温可能会在某种程度上促进初级生产力,但是也可能由于土壤水分、N素营养状况的改变形成新胁迫而抑制生产力提高。此外,生态系统呼吸由于增温而提高的幅度也可能高于初级生产力提高的幅度,造成总碳库平衡的改变。利用青藏高原海北高寒草甸实测数据对生态系统过程模型Biome-BGC(V.4.2)进行了参数化,并利用研究区实测土壤水分(0-40 cm)和其它观测数据对模型进行了检验,证明模型模拟结果较为可靠。模型使用2005-2008年的海北气象站实测气象数据包括气温、降水等作为驱动数据,模拟了增温1.2-1.7℃下青藏高原海北定位站高寒草甸生态系统碳通量的变化,并整合分析增温试验平台上已发表的试验,与模拟结果进行对比,探讨增温对海北高寒草甸生态系统碳收支的可能影响。结果表明:2005-2008年青藏高原高寒草甸生态系统为弱的碳汇,短期增温导致系统净碳固定增加。增温直接影响系统碳通量,也通过土壤水分和土壤矿化氮变化间接影响碳通量,相比土壤水分和氮素,增温对影响碳通量变化过程中的效应更大;研究也揭示,在增温条件下,植物对土壤矿化氮的吸收量小于有机质分解产生的土壤矿化氮量,土壤矿化氮含量增加。
DOI:10.5846/stxb201102250215URL [本文引用: 1]
低温被广泛认为是高寒草甸生态系统首要限制性因子,因此增温可能会在某种程度上促进初级生产力,但是也可能由于土壤水分、N素营养状况的改变形成新胁迫而抑制生产力提高。此外,生态系统呼吸由于增温而提高的幅度也可能高于初级生产力提高的幅度,造成总碳库平衡的改变。利用青藏高原海北高寒草甸实测数据对生态系统过程模型Biome-BGC(V.4.2)进行了参数化,并利用研究区实测土壤水分(0-40 cm)和其它观测数据对模型进行了检验,证明模型模拟结果较为可靠。模型使用2005-2008年的海北气象站实测气象数据包括气温、降水等作为驱动数据,模拟了增温1.2-1.7℃下青藏高原海北定位站高寒草甸生态系统碳通量的变化,并整合分析增温试验平台上已发表的试验,与模拟结果进行对比,探讨增温对海北高寒草甸生态系统碳收支的可能影响。结果表明:2005-2008年青藏高原高寒草甸生态系统为弱的碳汇,短期增温导致系统净碳固定增加。增温直接影响系统碳通量,也通过土壤水分和土壤矿化氮变化间接影响碳通量,相比土壤水分和氮素,增温对影响碳通量变化过程中的效应更大;研究也揭示,在增温条件下,植物对土壤矿化氮的吸收量小于有机质分解产生的土壤矿化氮量,土壤矿化氮含量增加。
DOI:10.1007/s12665-014-3376-5URL [本文引用: 1]
Conversion from rangeland to cropland potentially influences soil organic matter (SOM) dynamics and biochemical properties. The purpose of the current study was to assess changes in soil properties and investigate the interaction between SOM and enzyme activities following cultivation of native rangelands for more than 4002years. The quantities of soil organic carbon (C) and microbial biomass C (MBC), aggregate stability, microbial respiration rate (MRR), potential C mineralization (PCM) and the activities of soil enzymes involved in C, N, P and S cycling at the 0–20 and 20–4002cm depths were evaluated in never-cultivated natural rangelands and their corresponding farmlands with similar soil type and climatic conditions at Dehnow located in Central west, Iran. Cultivation in native rangelands resulted in a significant decline of most soil properties in farmlands, depending on soil sampling depths. Conversion of rangelands to their farmland counterparts decreased soil C (50–6602%), MBC (48–6102%), MRR (20–4402%), PCM (45–5902%), the activities of urease (5–1702%), invertase (7–2802%) and arylsulfatase (17–2002%) per soil mass, while increased C turnover rate (61–6302%), microbial metabolic quotient ( q CO 2 ) values (41–5402%) and enzyme activities per unit C and MBC (61–19002%) without affecting microbial quotient (MBC/C ratio) values and the activities of soil phosphatases per soil mass. The decreases in soil microbial and biochemical properties were largely the result of a substantial decline in soil C contents and aggregate stability by tillage activities in farmlands. Although the absolute activities of soil enzymes (i.e., activity per dry soil mass) showed inconsistent responses to land use changes and were even less responsive than soil organic C and MBC contents to changing land uses, the specific enzyme activities expressed either per unit of C or MBC tended to increase considerably and consistently with this conversion and were more sensitive than soil C and MBC contents. Soil C turnover rates and metabolic quotients were found to correlate positively with the specific enzyme activities, indicating an interaction between SOM and enzyme activities. The specific enzymatic activity could be an appropriate and integrative simple index to (1) assess soil quality changes, (2) detect changes in soil microbial community; and even (3) associate the losses of SOM with the presence of metabolically more active decomposer biota in rangelands cultivated for the long term.
DOI:10.1111/j.1365-2745.2009.01506.xURL [本文引用: 1]
Summary Top of page Summary Introduction Methods Results Discussion Conclusions Acknowledgements References 1 Climate warming increases the cover of deciduous shrubs in arctic ecosystems and herbivory is also known to have a strong influence on the biomass and composition of vegetation. However, research combining herbivory with warming is largely lacking. Our study describes how warming and simulated herbivory affect vegetation, soil nutrient concentrations and soil microbial communities after 10 13 years of exposure. 2 We established a factorial warming and herbivory-simulation experiment at a subarctic tundra heath in Kilpisj盲rvi, Finland, in 1994. Warming was carried out using the open-top chamber setup of the International Tundra Experiment (ITEX). Wounding of the dominant deciduous dwarf shrub Vaccinium myrtillus L. to simulate herbivory was carried out annually. We measured vegetation cover in 2003 and 2007, soil nutrient concentrations in 2003 and 2006, soil microbial respiration in 2003, and composition and function of soil microbial communities in 2006. 3 Warming increased the cover of V. myrtillus , whereas other plant groups did not show any response. Simulated herbivory of V. myrtillus cancelled out the impact of warming on the species cover, and increased the cover of other dwarf shrubs. 4 The concentrations of , and microbial biomass C and N in the soil were significantly reduced by warming after 10 treatment years but not after 13 treatment years. The reduction in by warming was significant only without simultaneous herbivory treatment, which indicates that simulated herbivory reduced N uptake by vegetation. 5 Soil microbial community composition, based on phospholipid fatty acid (PLFA) analysis, was slightly altered by warming. The activity of cultivable bacterial and fungal communities was significantly increased by warming and the substrate utilization patterns were influenced by warming and herbivory. 6 Synthesis. Our results show that warming increases the cover of V. myrtillus , which seems to enhance the nutrient sink strength of vegetation in the studied ecosystem. However, herbivory partially negates the effect of warming on plant N uptake and interacts with the effect of warming on microbial N immobilization. Our study demonstrates that effects of warming on soil microorganisms are likely to differ in the presence and absence of herbivores.
DOI:10.1007/BF01104985URL [本文引用: 1]
Understanding the role of terrestrial ecosystems in the global carbon (C) cycle has become increasingly important as policymakers consider options to address the issues associated with global change, particularly climate change. Sound scientific theories are critical in predicting how these systems may respond in the future, both to climate change and human actions.In March 1993, 60 scientists from 13 nations gathered in Bad Harzburg, Germany, to develop a state-of-the-science assessment of the present and likely future C fluxes associated with the major components of the earth's terrestrial biosphere. In the process, particular emphasis was placed on the potential for improving C sinks and managing long-term C sequestration.The majority of the week's work was conducted in eight working groups which independently considered a particular biome or subject area. The working groups considered: the Global Carbon Cycle; Boreal Forests and Tundra; Temperate Forests; Tropical Forests; Grasslands, Savannas and Deserts; Land and Water Interface Zones; Agroecosystems; and Biomass Management. This paper presents a brief overview of their major conclusions and findings. In addition, Table 1 brings together the best estimates from each group as to the current magnitude and estimated future direction of changes in the terrestrial C fluxes.
DOI:10.1016/0167-1987(92)90089-TURL [本文引用: 2]
ABSTRACT Soil compaction can be a major limitation to continuous maize production on the heavier-textured soils of the Manawatu region, New Zealand. A survey was undertaken of commercially-operated farms that were producing continuous maize using conventional mouldboard ploughing and cultivation methods. The effects of cultivation on the proportions of water-stable macroaggregates, total organic C content, microbial biomass C and soil respiration were measured in the predominant soil type, a poorly-drained Kairanga silty clay loam (Typic Haplaquept). Three other regional soil types, a Manawatu silt loam (well drained), Kairanga silt loam (poorly drained) and Moutoa humic clay (very poorly drained) were incluced for comparison. The effects of cropping with wheat or barley, and the recovery of the soils after restoration to pasture, were also measured.Continuous cultivation with maize on the Kairanga silty clay loam for up to 11 years decreased the total C content in the top 20 cm of soil by 21%, microbial C by 49% and water-stable aggregates (greater than 2 mm) by 54% compared with the levels under long-term permanent pasture. Losses on cultivation of pastures were greater on Manawatu silt loam with decreases of 49% in organic C, 60% in microbial C and 98% of the greater than 2 mm aggregates. Changes were much less pronounced on the Moutoa soil with a greater initial organic matter content, and the soil rapidly established new, only slightly lower, equilibria. Cultivation for barley or wheat had a less detrimental effect than cultivation for maize on the total organic and microbial C contents and aggregate stability of Kairanga silty clay loam. There was a strong linear relationship between the decline in the proportion of stable aggregates and the loss of microbial C and organic C in the top 10 cm of cultivated Kairanga soil. The relationship was stronger between microbial C and aggregate stability (R2 = 77.8%) than total organic C and aggregate stability (R2 = 64%), but was only valid when soil organic C was declining, and was not significant in the soil at the 10鈥20 cm depth.Re-establishment of pasture caused a more rapid recovery in microbial biomass C than total C, and increased the proportion of organic C comprised of microbial C. However, recovery of the total organic and microbial C pools and aggregate stability was very variable, and after 4 years of pasture none of the sites had re-established the levels found under permanent pasture. The previous cropping history of these soils before being returned to pasture, rather than the organic or microbial C content, appeared to be of greater importance in controlling the aggregate stability characteristics.
DOI:10.1016/j.catena.2004.09.001URL [本文引用: 1]
Overgrazing is one of the main causes of desertification in the semiarid Horqin sandy grassland of northern China. Excluding grazing livestock is considered as an alternative to restore vegetation in degraded sandy grassland in this region. However, few data are available concerning the impacts of continuous grazing and livestock exclusion on soil properties. In this paper, characteristics of vegetation and soil properties under continuous grazing and exclusion of livestock for 5 and 10 years were examined in representative degraded sandy grassland. Continuous grazing resulted in a considerable decrease in ground cover, which accelerates soil erosion by wind, leading to a further coarseness in surface soil, loss of soil organic C and N, and a decrease in soil biological properties. The grassland under continuous grazing is in the stage of very strong degradation. Excluding livestock grazing enhances vegetation recovery, litter accumulation, and development of annual and perennial grasses. Soil organic C and total N concentrations, soil biological properties including some enzyme activities and basal soil respiration improved following 10-year exclusion of livestock, suggesting that degradation of the grassland is being reversed. The results suggest that excluding grazing livestock on the desertified sandy grassland in the erosion-prone Horqin region has a great potential to restore soil fertility, sequester soil organic carbon and improve biological activity. Soil restoration is a slow process although the vegetation can recover rapidly after removal of livestock. A viable option for sandy grassland management should be to adopt proper exclosure in a rotation grazing system in the initial stage of grassland degradation.
DOI:10.3321/j.issn:1000-0933.2009.10.040URL [本文引用: 1]
2005~2008年在内蒙古呼和浩特市清水河县进行定位试验,设免耕留 低茬(NL)、免耕留高茬覆盖(NHS)和传统耕翻(T)3种耕作处理方式.结果表明:(1)免耕留高茬覆盖及免耕留低茬长期实施,能显著提高表层土壤有 机质、全氮、全钾、碱解氮、速效磷和速效钾含量,且免耕留高茬覆盖处理比传统耕翻分别提高了11%、41%、22%、15%、29%、27%、13%;在 测定各个时期内,土壤各营养指标含量整体趋势为NHS>NL>T.(2)免耕留高茬覆盖及免耕留低茬耕作方式有利于提高土壤微生物量C、N含量,在各测定 时期均以免耕留高茬覆盖处理的土壤微生物量C、N含量最高,传统耕翻最低.与传统耕翻相比,免耕留高茬覆盖处理土壤微生物量C、N含量分别平均提高了 69%、43%;测定各个时期,不同处理土壤生物量C、N含量均以7月份含量最高、5月份次之、10月份最低.(3)免耕留高茬覆盖及免耕留低茬处理土壤 碱性磷酸酶、蔗糖酶、过氧化氢酶活性和脲酶活性高于传统耕翻,整个测定期内免耕留高茬覆盖处理4种酶平均活性,分别较传统耕翻增加了57%、82%、 93%和25%;春季土壤酶活性开始增强,在7月份蔗糖酶、过氧化氢酶和脲酶活性达到最大值,而碱性磷酸酶的峰值出现在6月份.土壤微生物量C、N及土壤 酶活性是评价土壤质量的重要因子.
DOI:10.3321/j.issn:1000-0933.2009.10.040URL [本文引用: 1]
2005~2008年在内蒙古呼和浩特市清水河县进行定位试验,设免耕留 低茬(NL)、免耕留高茬覆盖(NHS)和传统耕翻(T)3种耕作处理方式.结果表明:(1)免耕留高茬覆盖及免耕留低茬长期实施,能显著提高表层土壤有 机质、全氮、全钾、碱解氮、速效磷和速效钾含量,且免耕留高茬覆盖处理比传统耕翻分别提高了11%、41%、22%、15%、29%、27%、13%;在 测定各个时期内,土壤各营养指标含量整体趋势为NHS>NL>T.(2)免耕留高茬覆盖及免耕留低茬耕作方式有利于提高土壤微生物量C、N含量,在各测定 时期均以免耕留高茬覆盖处理的土壤微生物量C、N含量最高,传统耕翻最低.与传统耕翻相比,免耕留高茬覆盖处理土壤微生物量C、N含量分别平均提高了 69%、43%;测定各个时期,不同处理土壤生物量C、N含量均以7月份含量最高、5月份次之、10月份最低.(3)免耕留高茬覆盖及免耕留低茬处理土壤 碱性磷酸酶、蔗糖酶、过氧化氢酶活性和脲酶活性高于传统耕翻,整个测定期内免耕留高茬覆盖处理4种酶平均活性,分别较传统耕翻增加了57%、82%、 93%和25%;春季土壤酶活性开始增强,在7月份蔗糖酶、过氧化氢酶和脲酶活性达到最大值,而碱性磷酸酶的峰值出现在6月份.土壤微生物量C、N及土壤 酶活性是评价土壤质量的重要因子.
URL [本文引用: 1]
以松嫩平原西部草甸草原中典型植物虎尾草、碱茅、芦苇和羊草群落 为对象,分析了4种植被群落土壤呼吸速率日动态和季节动态及其影响因素,以及土壤盐碱度与土壤呼吸碳排放量的关系.结果表明:4种植物群落的土壤呼吸速率 日变化均呈明显的单峰曲线,峰值出现在11:00-15:00,而谷值大多出现在21:00-1:00或3:00-5:00;4种植被群落土壤呼吸速率的 季节变化趋势一致,7、8月的土壤呼吸速率(3.21~4.84 μmol CO2·m-2·s-1)最高,10月最低(0.46~1.51 μmol CO2·m-2·s-1);各群落土壤呼吸速率与土壤和近地表大气温度之间呈极显著相关关系,其中,虎尾草群落的土壤呼吸速率与土壤表层含水量极显著相 关,芦苇和羊草群落土壤呼吸速率与近地表的相对湿度显著相关.土壤盐分含量明显抑制了土壤CO2排放量,土壤pH、电导率和土壤交换性钠可以解释该草甸草 原土壤呼吸空间变异的87% ~91%.
URL [本文引用: 1]
以松嫩平原西部草甸草原中典型植物虎尾草、碱茅、芦苇和羊草群落 为对象,分析了4种植被群落土壤呼吸速率日动态和季节动态及其影响因素,以及土壤盐碱度与土壤呼吸碳排放量的关系.结果表明:4种植物群落的土壤呼吸速率 日变化均呈明显的单峰曲线,峰值出现在11:00-15:00,而谷值大多出现在21:00-1:00或3:00-5:00;4种植被群落土壤呼吸速率的 季节变化趋势一致,7、8月的土壤呼吸速率(3.21~4.84 μmol CO2·m-2·s-1)最高,10月最低(0.46~1.51 μmol CO2·m-2·s-1);各群落土壤呼吸速率与土壤和近地表大气温度之间呈极显著相关关系,其中,虎尾草群落的土壤呼吸速率与土壤表层含水量极显著相 关,芦苇和羊草群落土壤呼吸速率与近地表的相对湿度显著相关.土壤盐分含量明显抑制了土壤CO2排放量,土壤pH、电导率和土壤交换性钠可以解释该草甸草 原土壤呼吸空间变异的87% ~91%.
DOI:10.3321/j.issn:1000-0933.2007.05.036URL [本文引用: 1]
采用静态箱-气相色谱法对四川盆地中部紫色土丘陵区3种土地利用方式(林地、草地和轮作旱地)土壤呼吸进行测定,结果表明,林地、草地和旱地土壤呼吸速率变化范围分别为78.63~577.97、39.28~584.18和34.48~484.65mgCO2·m^-2·h^-1,年平均土壤呼吸速率分别为264.68、242.91、182.21mgCO2·m^-2h^-1。3种土地利用方式的土壤呼吸速率季节变化趋势均呈单峰曲线,林地和草地土壤呼吸速率最大值均出现在夏末(7月底与8月初之间),旱地土壤呼吸速率最大值出现的时间比林地和草地要早,在6月底与7月初之间;最小值均出现在12月底与翌年1月初之间。土壤温度和土壤湿度是影响本地区土壤呼吸的主要因子,双因素关系模型(R=αe^bTw^c)较好地拟合了土壤温度和土壤湿度对土壤呼吸的影响,二者共同解释了土壤呼吸变化的64%~90%。土壤呼吸的温度敏感性指数Q10值受土壤(5cm处)温度和土壤(0~10cm)湿度的影响。分析表明3种土地利用土壤的Q10值与土壤温度呈显著负相关关系,而与土壤湿度呈显著正相关关系。
DOI:10.3321/j.issn:1000-0933.2007.05.036URL [本文引用: 1]
采用静态箱-气相色谱法对四川盆地中部紫色土丘陵区3种土地利用方式(林地、草地和轮作旱地)土壤呼吸进行测定,结果表明,林地、草地和旱地土壤呼吸速率变化范围分别为78.63~577.97、39.28~584.18和34.48~484.65mgCO2·m^-2·h^-1,年平均土壤呼吸速率分别为264.68、242.91、182.21mgCO2·m^-2h^-1。3种土地利用方式的土壤呼吸速率季节变化趋势均呈单峰曲线,林地和草地土壤呼吸速率最大值均出现在夏末(7月底与8月初之间),旱地土壤呼吸速率最大值出现的时间比林地和草地要早,在6月底与7月初之间;最小值均出现在12月底与翌年1月初之间。土壤温度和土壤湿度是影响本地区土壤呼吸的主要因子,双因素关系模型(R=αe^bTw^c)较好地拟合了土壤温度和土壤湿度对土壤呼吸的影响,二者共同解释了土壤呼吸变化的64%~90%。土壤呼吸的温度敏感性指数Q10值受土壤(5cm处)温度和土壤(0~10cm)湿度的影响。分析表明3种土地利用土壤的Q10值与土壤温度呈显著负相关关系,而与土壤湿度呈显著正相关关系。
URL [本文引用: 1]
本研究以伊犁昭苏马场严重退化草地为对象,以彻底翻耕建植放牧型 草地为目的,探讨其重建及合理利用技术。通过重建前后对比,分析群落特征变化,筛选适宜的重建组合,明确重建技术;进而对不同放牧利用强度下重建草地进行 群落数量及质量动态测定,揭示其变化规律,综合评价其利用效果,确定重建草地合理的放牧利用方式。主要研究内容包括: ⑴秋季对重 建草地群落特征进行连续2年测定结果表明各重建处理除盖度外,其余指标第二年高于第一年,且基本优于未重建草地;重建处理群落特征指标排序存在一定的波动 性;群落盖度、高度、密度、产量和豆科牧草产量比值...
URL [本文引用: 1]
本研究以伊犁昭苏马场严重退化草地为对象,以彻底翻耕建植放牧型 草地为目的,探讨其重建及合理利用技术。通过重建前后对比,分析群落特征变化,筛选适宜的重建组合,明确重建技术;进而对不同放牧利用强度下重建草地进行 群落数量及质量动态测定,揭示其变化规律,综合评价其利用效果,确定重建草地合理的放牧利用方式。主要研究内容包括: ⑴秋季对重 建草地群落特征进行连续2年测定结果表明各重建处理除盖度外,其余指标第二年高于第一年,且基本优于未重建草地;重建处理群落特征指标排序存在一定的波动 性;群落盖度、高度、密度、产量和豆科牧草产量比值...
URL [本文引用: 1]
采用开顶箱 (open-top chamber,OTC) 增温方法 ( 1.1-1.9 ℃),研究了长白山苔原生态系统土壤酶活性、土壤微生物生物量、土壤微生物群落结构及土壤微生物呼吸对温度升高的响应。结果表明,连续三个生长季 (6-9月) 增温,没有明显地改变土壤蔗糖酶 (58.1和45.9 mg g^-1 24 h^-1) 和纤维素酶 (0.34 和0.26 mg g^-1 72 h^-1) 的活性,但土壤脲酶活性升高80.1% (0.82和 0.46 mg g^-1 24 h^-1),过氧化氢酶活性也升高10.1% (1.18 和 1.07 ml KMnO4 g^-1 h^-1)。增温与对照条件下土壤微生物生物量碳含量 (0.85 和0.75 mg g^-1)、氮 (0.07 和0.06 mg g^-1) 、磷 (0.013 和0.011 mg g^-1) 和土壤微生物呼吸 (6.1 和 6.3 μmol m^-2 s-1) 无明显差异。相关分析表明,土壤微生物生物量月际间明显的变化与土壤含水量及土壤有机质的相对变化有关。增温改变了土壤微生物的群落结构。增温并未引起与碳循环相关的酶活性、土壤微生物生物量和土壤微生物呼吸发生明显变化,可能是短期增温及增温幅度不足以使土壤微生物活性产生明显的改变。
URL [本文引用: 1]
采用开顶箱 (open-top chamber,OTC) 增温方法 ( 1.1-1.9 ℃),研究了长白山苔原生态系统土壤酶活性、土壤微生物生物量、土壤微生物群落结构及土壤微生物呼吸对温度升高的响应。结果表明,连续三个生长季 (6-9月) 增温,没有明显地改变土壤蔗糖酶 (58.1和45.9 mg g^-1 24 h^-1) 和纤维素酶 (0.34 和0.26 mg g^-1 72 h^-1) 的活性,但土壤脲酶活性升高80.1% (0.82和 0.46 mg g^-1 24 h^-1),过氧化氢酶活性也升高10.1% (1.18 和 1.07 ml KMnO4 g^-1 h^-1)。增温与对照条件下土壤微生物生物量碳含量 (0.85 和0.75 mg g^-1)、氮 (0.07 和0.06 mg g^-1) 、磷 (0.013 和0.011 mg g^-1) 和土壤微生物呼吸 (6.1 和 6.3 μmol m^-2 s-1) 无明显差异。相关分析表明,土壤微生物生物量月际间明显的变化与土壤含水量及土壤有机质的相对变化有关。增温改变了土壤微生物的群落结构。增温并未引起与碳循环相关的酶活性、土壤微生物生物量和土壤微生物呼吸发生明显变化,可能是短期增温及增温幅度不足以使土壤微生物活性产生明显的改变。
DOI:10.2136/sssaj2009.0366URL [本文引用: 1]
Abstract This study investigated how disturbances from cultivation and haying altered the soil organic C (SOC) at two sites: one (Lethbridge) in a Stipa–Agropyron–Bouteloua community (49°701 N, 112°5701 W) and a second (Onefour) in a more xeric Stipa–Bouteloua community (49°301 N, 110°2701 W). Seven disturbance treatments were established on previously uncultivated native grassland to assess their effect on SOC 12 or 13 yr after establishment. A randomized complete block design with four replicates was used. Soils were sampled in the fall of 1995, 1997, and 2006 at two increments (0–15 and 15–30 cm) and a third (30–90 cm) was sampled in 2006. The soils were all analyzed for their SOC and, in 2006, also for their light fraction organic C (LF OC) and mineralizable C. Breaking and immediate establishment of perennial grass monocultures had no effect on SOC, LF OC, or mineralizable C. Wheat (Triticum aestivum L.) cropping (either fallow or continuous) resulted in a 19% loss of SOC after 13 yr. The rate of loss varied from an average of about 1.7 Mg ha-1 yr-1 in the first 4 yr to about 0.32 Mg ha-1 yr-1 in the subsequent 9 yr. These results are consistent with a more rapid loss of SOC in the first decade of cropping virgin soils, a slowing rate of loss, and a new steady state within a few decades.
DOI:10.1111/gcb.13218URLPMID:26748590 [本文引用: 2]
Abstract Agricultural expansion has resulted in both land use and land cover change (LULCC) across the tropics. However, the spatial and temporal patterns of such change and their resulting impacts are poorly understood, particularly for the presatellite era. Here, we quantify the LULCC history across the 33.902million ha watershed of Tanzania's Eastern Arc Mountains, using geo-referenced and digitized historical land cover maps (dated 1908, 1923, 1949 and 2000). Our time series from this biodiversity hotspot shows that forest and savanna area both declined, by 74% (2.802million ha) and 10% (2.902million ha), respectively, between 1908 and 2000. This vegetation was replaced by a fivefold increase in cropland, from 1.202million ha to 6.702million ha. This LULCC implies a committed release of 0.902Pg02C (95% CI: 0.4–1.5) across the watershed for the same period, equivalent to 0.302Mg02C02ha61102yr611. This is at least threefold higher than previous estimates from global models for the same study area. We then used the LULCC data from before and after protected area creation, as well as from areas where no protection was established, to analyse the effectiveness of legal protection on land cover change despite the underlying spatial variation in protected areas. We found that, between 1949 and 2000, forest expanded within legally protected areas, resulting in carbon uptake of 4.8 (3.8–5.7) Mg02C02ha611, compared to a committed loss of 11.9 (7.2–16.6) Mg02C02ha611 within areas lacking such protection. Furthermore, for nine protected areas where LULCC data are available prior to and following establishment, we show that protection reduces deforestation rates by 150% relative to unprotected portions of the watershed. Our results highlight that considerable LULCC occurred prior to the satellite era, thus other data sources are required to better understand long-term land cover trends in the tropics.
DOI:10.13227/j.hjkx.2015.04.050URL [本文引用: 1]
为研究不同温度下的土壤微生物呼吸及其与水溶性有机碳(DOC)和转化酶的关系,设置了室内培养实验.采集南京市周边老山、紫金山、宝华山的土壤,研究不同土壤的微生物呼吸对温度升高的响应规律,并分析土壤DOC含量及转化酶活性.结果表明,不同土壤的累积微生物呼吸与土壤温度之间的关系均可用指数方程描述,其P值均达到极显著水平(P0.001),不同地点土壤的微生物呼吸温度敏感系数(Q10值)在1.762~1.895之间变异.累积土壤微生物呼吸的Q10值随着土壤温度升高表现出降低的趋势.培养后27 d土壤微生物呼吸的Q10值与培养后1 d的Q10值无显著差异(P0.05),这表明难分解有机质的温度敏感性与易分解有机质的温度敏感性一致.对于所有土壤而言,累积土壤微生物呼吸与DOC含量之间存在极显著(P=0.003)的线性回归关系,DOC可以解释累积土壤微生物呼吸31.6%的变异性.无论是单独分析不同土壤还是综合所有土壤的测定结果,累积微生物呼吸与土壤转化酶活性均存在极显著(P0.01)的一元线性回归关系,由此说明转化酶活性是衡量土壤微生物呼吸大小的一个较好的指标.
DOI:10.13227/j.hjkx.2015.04.050URL [本文引用: 1]
为研究不同温度下的土壤微生物呼吸及其与水溶性有机碳(DOC)和转化酶的关系,设置了室内培养实验.采集南京市周边老山、紫金山、宝华山的土壤,研究不同土壤的微生物呼吸对温度升高的响应规律,并分析土壤DOC含量及转化酶活性.结果表明,不同土壤的累积微生物呼吸与土壤温度之间的关系均可用指数方程描述,其P值均达到极显著水平(P0.001),不同地点土壤的微生物呼吸温度敏感系数(Q10值)在1.762~1.895之间变异.累积土壤微生物呼吸的Q10值随着土壤温度升高表现出降低的趋势.培养后27 d土壤微生物呼吸的Q10值与培养后1 d的Q10值无显著差异(P0.05),这表明难分解有机质的温度敏感性与易分解有机质的温度敏感性一致.对于所有土壤而言,累积土壤微生物呼吸与DOC含量之间存在极显著(P=0.003)的线性回归关系,DOC可以解释累积土壤微生物呼吸31.6%的变异性.无论是单独分析不同土壤还是综合所有土壤的测定结果,累积微生物呼吸与土壤转化酶活性均存在极显著(P0.01)的一元线性回归关系,由此说明转化酶活性是衡量土壤微生物呼吸大小的一个较好的指标.
[本文引用: 2]
[本文引用: 2]
URLMagsci [本文引用: 1]
<p>张成霞,南志标综述了我国不同放牧强度对草地土壤微生物区系组成、微生物数量及土壤微生物的时空分布变化研究,草地的不同利用方式对土壤微生物的影响。草地土壤微生物类群主要由真菌、细菌和放线菌等组成,它们也是草地土壤微生物中数量最大的3个类群,其中,细菌数量最多,放线菌次之,真菌最少。在空间上,草地土壤微生物数量表现为随土层的加深逐渐减少,季节分布表现为春季随温度的上升逐渐增高,最高峰出现在8月中旬,冬季降到最低。因此微生物在草地生态系统的能量流动和物质转化中起着重要的作用。</p>
URLMagsci [本文引用: 1]
<p>张成霞,南志标综述了我国不同放牧强度对草地土壤微生物区系组成、微生物数量及土壤微生物的时空分布变化研究,草地的不同利用方式对土壤微生物的影响。草地土壤微生物类群主要由真菌、细菌和放线菌等组成,它们也是草地土壤微生物中数量最大的3个类群,其中,细菌数量最多,放线菌次之,真菌最少。在空间上,草地土壤微生物数量表现为随土层的加深逐渐减少,季节分布表现为春季随温度的上升逐渐增高,最高峰出现在8月中旬,冬季降到最低。因此微生物在草地生态系统的能量流动和物质转化中起着重要的作用。</p>
DOI:10.3969/j.issn.1003-4315.2007.02.022URL
以耕地为对照,研究了高寒农牧交错带种植老芒麦草和撂荒2种植被恢复措施对土壤有机碳和全氮含量的影响.结果表明,与耕地相比较种植老芒麦草和撂荒使0-30 cm土层土壤总有机碳含量分别增加了19.7%-22.4%和7.3%-10.6%,颗粒有机碳(0.05-2.00 mm)含量分别增加了43.0%-52.3%和28.7%-59.8%,土壤全氮含量分别增加了12.0%-20.6%和5.4%-13.7%.这一结果说明,在高寒农牧交错带通过种植老芒麦草或者撂荒植被恢复措施,对于提高土壤有机质含量和全氮含量、恢复土壤质量具有非常显著的效果.
DOI:10.3969/j.issn.1003-4315.2007.02.022URL
以耕地为对照,研究了高寒农牧交错带种植老芒麦草和撂荒2种植被恢复措施对土壤有机碳和全氮含量的影响.结果表明,与耕地相比较种植老芒麦草和撂荒使0-30 cm土层土壤总有机碳含量分别增加了19.7%-22.4%和7.3%-10.6%,颗粒有机碳(0.05-2.00 mm)含量分别增加了43.0%-52.3%和28.7%-59.8%,土壤全氮含量分别增加了12.0%-20.6%和5.4%-13.7%.这一结果说明,在高寒农牧交错带通过种植老芒麦草或者撂荒植被恢复措施,对于提高土壤有机质含量和全氮含量、恢复土壤质量具有非常显著的效果.
[本文引用: 2]
[本文引用: 2]
DOI:10.1016/j.ecoleng.2014.10.003URL [本文引用: 2]
A better understanding of the response of soil respiration to land-use conversion has important practical implications for ecological restoration in degraded regions. In this study, in situ soil respiration was monitored in a typical land-use sequence on a ridge slope in Wangdonggou watershed of the Loess Plateau, China, during a three-year period from 2011 to 2013. The land-use conversion sequences included cropland (control), apple orchard, grassland, and woodland. The results clearly showed that soil respiration and temperature sensitivity ( Q 10 ) varied significantly with land-use conversion. Soil respiration was decreased by 10% after conversion of cropland to orchard, and increased by 7 46% after conversion of cropland to grassland and woodland. Q 10 was increased by 19% after conversion of cropland to woodland, and decreased by 9 26% after conversion of cropland to grassland and orchard. Soil respiration increased linearly with soil organic carbon (SOC) storage and fine root biomass (<2 mm). The results indicated that root biomass and SOC storage were the major factors influencing Q 10 after conversion of cropland to non-natural ecosystem, and substrate quality or root system adaptability may be the real reason for the difference in Q 10 after conversion of cropland to natural grassland ecosystem. Although soil temperature and moisture significantly influenced soil respiration among the four typical land-use types, their difference derived from land-use conversions could not well explain the difference in soil respiration among land-use conversions. In conclusion, the increases in SOC storage and fine root biomass were the major factors influencing soil respiration among land-use conversions. Thus, conversion of cropland to natural grassland seemed to be the most effective integrated small watershed management to increase soil carbon storage and decrease CO 2 concentration in the loess regions of China.
[本文引用: 1]
[本文引用: 1]
DOI:10.11686/cyxb20150316URLMagsci [本文引用: 1]
以红豆草、紫花苜蓿、红三叶、鸭茅、无芒雀麦和猫尾草6种豆禾牧草在混播种类为3、4、5、6与豆禾比5∶5、4∶6和3∶7条件下建立混播草地。依据2008-2012年各混播组合的群落组分稳定性、功能稳定性和可入侵性,比较了21个豆禾混播组合的群落稳定性。从组分稳定性来看,含红豆草的组合豆科植物稳定性较差;而不含红豆草组合豆科植物稳定性较高;从功能稳定性来看,随着混播种类数量的增加,群落稳定性呈增加趋势,豆禾比4∶6的群落稳定性低于豆禾比5∶5和3∶7。从可入侵性来看,混播种类对其影响较小,豆禾比5∶5具有较高的稳定性。通过模糊综合评价,包含组分稳定性、功能稳定性和可入侵性3个方面的群落稳定性以豆禾比为4∶6较低,而混播种类对群落稳定性影响较小。因此,混播种类与比例仅能影响到群落稳定性的某一方面,还需探寻适宜的时空尺度与评价“标尺”下影响群落稳定性的其他因素。
DOI:10.11686/cyxb20150316URLMagsci [本文引用: 1]
以红豆草、紫花苜蓿、红三叶、鸭茅、无芒雀麦和猫尾草6种豆禾牧草在混播种类为3、4、5、6与豆禾比5∶5、4∶6和3∶7条件下建立混播草地。依据2008-2012年各混播组合的群落组分稳定性、功能稳定性和可入侵性,比较了21个豆禾混播组合的群落稳定性。从组分稳定性来看,含红豆草的组合豆科植物稳定性较差;而不含红豆草组合豆科植物稳定性较高;从功能稳定性来看,随着混播种类数量的增加,群落稳定性呈增加趋势,豆禾比4∶6的群落稳定性低于豆禾比5∶5和3∶7。从可入侵性来看,混播种类对其影响较小,豆禾比5∶5具有较高的稳定性。通过模糊综合评价,包含组分稳定性、功能稳定性和可入侵性3个方面的群落稳定性以豆禾比为4∶6较低,而混播种类对群落稳定性影响较小。因此,混播种类与比例仅能影响到群落稳定性的某一方面,还需探寻适宜的时空尺度与评价“标尺”下影响群落稳定性的其他因素。
The dynamics of organic matter in grassland soils
1
1985
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
青海湖北岸高寒草甸草原非生长季土壤呼吸对温度和湿度的响应
1
2014
... id="C54">总体而言, Q10与温度呈负相关关系, 即在温度上升相同幅度下低温区比高温区有更大的Q10 (
青海湖北岸高寒草甸草原非生长季土壤呼吸对温度和湿度的响应
1
2014
... id="C54">总体而言, Q10与温度呈负相关关系, 即在温度上升相同幅度下低温区比高温区有更大的Q10 (
增温及秸秆施用对冬小麦田土壤呼吸和酶活性的影响
1
2016
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
增温及秸秆施用对冬小麦田土壤呼吸和酶活性的影响
1
2016
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
Effects of climate and management intensity on nitrous oxide emissions in grassland systems across Europe
1
2007
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
1986
1986
黄土高原半干旱区土壤呼吸对土地利用变化的响应
1
2014
... id="C19">脲酶活性的测定采用奈氏比色法, 以1 g干土 24 h生成的NH3-N量为脲酶1个活性单位; 过氧化氢酶活性的测定采用KMnO4滴定法, 酶活性以1 g干土1 h内消耗的0.1 mol·L-1 KMnO4体积数(以mL计)表示; 转化酶活性的测定采用3,5-二硝基水杨酸比色法, 酶活性以24 h后1 g土壤葡萄糖的mg数表示; 蛋白酶活性的测定采用铜盐比色法, 以1 g土壤在 24 h内酶解蛋白质释放的质量代表蛋白酶的活性(
黄土高原半干旱区土壤呼吸对土地利用变化的响应
1
2014
... id="C19">脲酶活性的测定采用奈氏比色法, 以1 g干土 24 h生成的NH3-N量为脲酶1个活性单位; 过氧化氢酶活性的测定采用KMnO4滴定法, 酶活性以1 g干土1 h内消耗的0.1 mol·L-1 KMnO4体积数(以mL计)表示; 转化酶活性的测定采用3,5-二硝基水杨酸比色法, 酶活性以24 h后1 g土壤葡萄糖的mg数表示; 蛋白酶活性的测定采用铜盐比色法, 以1 g土壤在 24 h内酶解蛋白质释放的质量代表蛋白酶的活性(
不同利用方式下草地土壤微生物及土壤呼吸特性. 草地学报
1
2012
... id="C52">土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(
不同利用方式下草地土壤微生物及土壤呼吸特性. 草地学报
1
2012
... id="C52">土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(
Separating root and soil microbial contributions to soil respiration: A review of methods and observations
2
2000
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
... 则发现, 在干旱年份土壤呼吸最高的是放牧样地, 而湿润年份是刈割样地, 水分是影响温带草原碳循环的重要限制因素.不同草地土地利用方式一方面通过改变根系呼吸(
翻耙补播对科尔沁碱化草地土壤理化性质和生物活性的影响
1
2006
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
翻耙补播对科尔沁碱化草地土壤理化性质和生物活性的影响
1
2006
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
The temperature dependence of organic- matter decomposition-still a topic of debate. S
1
2006
... id="C54">总体而言, Q10与温度呈负相关关系, 即在温度上升相同幅度下低温区比高温区有更大的Q10 (
Soil carbon sequestration impacts on global climate change and food security
1
2004
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
不同放牧管理模式对高寒草甸草原土壤呼吸特征的影响
1
2015
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
不同放牧管理模式对高寒草甸草原土壤呼吸特征的影响
1
2015
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
黄土高原不同地形封育草地的土壤呼吸日动态与影响因子分析
2
2010
... id="C52">土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
黄土高原不同地形封育草地的土壤呼吸日动态与影响因子分析
2
2010
... id="C52">土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
土地利用变化对土壤温室气体排放通量影响研究进展
1
2014
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
土地利用变化对土壤温室气体排放通量影响研究进展
1
2014
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
北方农牧交错带不同土地利用方式下土壤呼吸速率探究
2014
北方农牧交错带不同土地利用方式下土壤呼吸速率探究
2014
内蒙古典型草原植物功能型对土壤甲烷吸收的影响
2
2011
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
... id="C52">土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(
内蒙古典型草原植物功能型对土壤甲烷吸收的影响
2
2011
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
... id="C52">土壤呼吸受自然环境和人为活动干扰的共同影响, 不同气候环境、土壤环境和地上植被都会影响土壤呼吸CO2的产生量(
干扰方式对典型草原土壤理化性质的影响
1
2008
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
干扰方式对典型草原土壤理化性质的影响
1
2008
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
增温对青藏高原高寒草甸生态系统固碳通量影响的模拟研究
1
2012
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
增温对青藏高原高寒草甸生态系统固碳通量影响的模拟研究
1
2012
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
Soil C turnover, microbial biomass and respiration, and enzymatic activities following range and conversion to wheat-alfalfa cropping in a semi-arid climate
1
2014
... id="C8">退化的生态系统恢复成草地往往是理想的生态选择, 能够增加水分利用效率, 增加碳储量, 减少温室气体排放(
Responses of vegetation and soil microbial communities to warming and simulated herbivory in a subarctic heath
1
2009
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
Terrestrial biosphere carbon fluxes quantification of sinks and sources of CO2
1
1993
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
Changes in soil organic C, microbial C and aggregate stability under continuous maize and cereal cropping, and after restoration to pasture in soils from the Manawatu Region, New Zealand
2
1992
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
Influences of continuous grazing and livestock exclusion on soil properties in a degraded early grassland, Inner Mongolia, northern China
1
2005
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
免耕与留茬对土壤微生物生物量C、N及酶活性的影响
1
2009
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
免耕与留茬对土壤微生物生物量C、N及酶活性的影响
1
2009
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
松嫩平原西部草甸草原典型植物群落土壤呼吸动态及影响因素
1
2014
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
松嫩平原西部草甸草原典型植物群落土壤呼吸动态及影响因素
1
2014
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
不同土地利用方式下土壤呼吸及其温度敏感性
1
2007
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
不同土地利用方式下土壤呼吸及其温度敏感性
1
2007
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
放牧型草地重建与利用研究
1
2013
... id="C9">试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏县的昭苏马场(81.05°-81.08° E, 40.92°-43.25° N), 属于大陆性温带山区半干旱半湿润冷凉气候.2014-2015年平均气温为4.1 ℃, ≥10 ℃年积温为1β416.8 ℃, 无霜期85-100天, 2014-2015年平均年降水量为490.1 mm, 降水集中于5-10月, 约占全年总降水量的74%, 平均年蒸发量1β261.6 mm, 约是降水量的2.5倍, 积雪期158天, 积雪厚度20-60 cm.试验小区位于天山北坡西段山地草甸, 未退化山地草甸植被主要成分有鸭茅(Dactylis glomerata)、无芒雀麦(Bromus inermis)、新疆鹅观草(Roegneria sinkiangensis)、猫尾草(Phleum pratense)、草原糙苏(Phlomis pratensis)、草地老鹳草(Geranium pratense)、大叶橐吾(Ligularia macrophylla)等.盖度85%-100%, 草层高45-85 cm, 鲜草产量4β300-9β050 kg·hm-2.土壤为黑钙土, 0-5 cm土壤有机碳含量为7.91%-8.64%, 全氮、全磷、全钾含量分别为9.53、2.16和12.20 g·kg-1, 碱解氮、有效磷、有效钾含量分别为459.20、144.43和473.58 mg·kg-1 (
放牧型草地重建与利用研究
1
2013
... id="C9">试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏县的昭苏马场(81.05°-81.08° E, 40.92°-43.25° N), 属于大陆性温带山区半干旱半湿润冷凉气候.2014-2015年平均气温为4.1 ℃, ≥10 ℃年积温为1β416.8 ℃, 无霜期85-100天, 2014-2015年平均年降水量为490.1 mm, 降水集中于5-10月, 约占全年总降水量的74%, 平均年蒸发量1β261.6 mm, 约是降水量的2.5倍, 积雪期158天, 积雪厚度20-60 cm.试验小区位于天山北坡西段山地草甸, 未退化山地草甸植被主要成分有鸭茅(Dactylis glomerata)、无芒雀麦(Bromus inermis)、新疆鹅观草(Roegneria sinkiangensis)、猫尾草(Phleum pratense)、草原糙苏(Phlomis pratensis)、草地老鹳草(Geranium pratense)、大叶橐吾(Ligularia macrophylla)等.盖度85%-100%, 草层高45-85 cm, 鲜草产量4β300-9β050 kg·hm-2.土壤为黑钙土, 0-5 cm土壤有机碳含量为7.91%-8.64%, 全氮、全磷、全钾含量分别为9.53、2.16和12.20 g·kg-1, 碱解氮、有效磷、有效钾含量分别为459.20、144.43和473.58 mg·kg-1 (
长白山苔原生态系统土壤酶活性及微生物生物量对增温的响应
1
2014
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
长白山苔原生态系统土壤酶活性及微生物生物量对增温的响应
1
2014
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
Cultivation and reseeding effects on soil organic matter in the mixed prairie
1
2010
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
草地利用方式对土壤呼吸和凋落物分解的影响
2015
草地利用方式对土壤呼吸和凋落物分解的影响
2015
Land cover change and carbon emissions over 100 years in an African biodiversity hotspot
2
2016
... id="C6">土地利用方式的变化及其导致的植被变化是影响陆地生态系统碳循环的一个关键因素(
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
不同温度下的土壤微生物呼吸及其与水溶性有机碳和转化酶的关系
1
2015
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
不同温度下的土壤微生物呼吸及其与水溶性有机碳和转化酶的关系
1
2015
... id="C56">土壤酶活性的大小影响土壤中的生物学过程, 而土壤呼吸的大小反映了土壤中的生物学过程的强弱(
1986
1986
内蒙古不同草原类型土壤呼吸对放牧强度及水热因子的响应
2
2012
... id="C15">土壤微生物数量测定采用平板涂抹接种法, 以牛肉膏蛋白胨培养基培养细菌高氏1号培养基加重铬酸钾(3.30 mL·L-1)培养放线菌; 马丁氏培养基加链霉素(3.30 mL·L-1)培养真菌.培养3-6天观察(
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
内蒙古不同草原类型土壤呼吸对放牧强度及水热因子的响应
2
2012
... id="C15">土壤微生物数量测定采用平板涂抹接种法, 以牛肉膏蛋白胨培养基培养细菌高氏1号培养基加重铬酸钾(3.30 mL·L-1)培养放线菌; 马丁氏培养基加链霉素(3.30 mL·L-1)培养真菌.培养3-6天观察(
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
放牧对草地土壤微生物影响的研究述评
1
2010
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
放牧对草地土壤微生物影响的研究述评
1
2010
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
高寒农牧交错带植被恢复对土壤有机碳、全氮含量的影响
2007
高寒农牧交错带植被恢复对土壤有机碳、全氮含量的影响
2007
黄土高原不同植被坡地土壤微生物区系特征
2
2008
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
黄土高原不同植被坡地土壤微生物区系特征
2
2008
... id="C7">目前, 对天然草地的利用方式中, 放牧、割草和围封是最主要的方式(
... id="C55">土壤微生物是草地生态系统的重要组成部分, 其活动能力的强弱与植被类型、干扰强度、土壤肥力等诸多因素有密切的关系(
Responses of soil respiration to land use conversions in degraded ecosystem of the semi-arid Loess Plateau
2
2015
... id="C8">退化的生态系统恢复成草地往往是理想的生态选择, 能够增加水分利用效率, 增加碳储量, 减少温室气体排放(
... id="C53">影响土壤呼吸的主要因子是所在区域的环境因子, 特别是温度和水分(
草原碳汇管理对策
1
2013
... id="C8">退化的生态系统恢复成草地往往是理想的生态选择, 能够增加水分利用效率, 增加碳储量, 减少温室气体排放(
草原碳汇管理对策
1
2013
... id="C8">退化的生态系统恢复成草地往往是理想的生态选择, 能够增加水分利用效率, 增加碳储量, 减少温室气体排放(
不同混播方式下豆禾混播草地群落稳定性的测度与比较
1
2015
... id="C9">试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏县的昭苏马场(81.05°-81.08° E, 40.92°-43.25° N), 属于大陆性温带山区半干旱半湿润冷凉气候.2014-2015年平均气温为4.1 ℃, ≥10 ℃年积温为1β416.8 ℃, 无霜期85-100天, 2014-2015年平均年降水量为490.1 mm, 降水集中于5-10月, 约占全年总降水量的74%, 平均年蒸发量1β261.6 mm, 约是降水量的2.5倍, 积雪期158天, 积雪厚度20-60 cm.试验小区位于天山北坡西段山地草甸, 未退化山地草甸植被主要成分有鸭茅(Dactylis glomerata)、无芒雀麦(Bromus inermis)、新疆鹅观草(Roegneria sinkiangensis)、猫尾草(Phleum pratense)、草原糙苏(Phlomis pratensis)、草地老鹳草(Geranium pratense)、大叶橐吾(Ligularia macrophylla)等.盖度85%-100%, 草层高45-85 cm, 鲜草产量4β300-9β050 kg·hm-2.土壤为黑钙土, 0-5 cm土壤有机碳含量为7.91%-8.64%, 全氮、全磷、全钾含量分别为9.53、2.16和12.20 g·kg-1, 碱解氮、有效磷、有效钾含量分别为459.20、144.43和473.58 mg·kg-1 (
不同混播方式下豆禾混播草地群落稳定性的测度与比较
1
2015
... id="C9">试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏县的昭苏马场(81.05°-81.08° E, 40.92°-43.25° N), 属于大陆性温带山区半干旱半湿润冷凉气候.2014-2015年平均气温为4.1 ℃, ≥10 ℃年积温为1β416.8 ℃, 无霜期85-100天, 2014-2015年平均年降水量为490.1 mm, 降水集中于5-10月, 约占全年总降水量的74%, 平均年蒸发量1β261.6 mm, 约是降水量的2.5倍, 积雪期158天, 积雪厚度20-60 cm.试验小区位于天山北坡西段山地草甸, 未退化山地草甸植被主要成分有鸭茅(Dactylis glomerata)、无芒雀麦(Bromus inermis)、新疆鹅观草(Roegneria sinkiangensis)、猫尾草(Phleum pratense)、草原糙苏(Phlomis pratensis)、草地老鹳草(Geranium pratense)、大叶橐吾(Ligularia macrophylla)等.盖度85%-100%, 草层高45-85 cm, 鲜草产量4β300-9β050 kg·hm-2.土壤为黑钙土, 0-5 cm土壤有机碳含量为7.91%-8.64%, 全氮、全磷、全钾含量分别为9.53、2.16和12.20 g·kg-1, 碱解氮、有效磷、有效钾含量分别为459.20、144.43和473.58 mg·kg-1 (
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}