Effect of sheep digestive tract on the recovery and germination of seeds of fifteen leguminous plants in the northern Xinjiang region, China
WANG Shu-Lin,, LU Wei-Hua,*, CHEN Yi-Shi*, JING Peng-Cheng*College of Animal Science and Technology, Shihezi University, Shihezi, Xinjiang 832003, China
通讯作者: 通讯作者Author for correspondence (E-mail: jshe@pku.edu.cn)
Abstract Aims The objective of this study was to learn the effect of sheep digestive tract on the recovery and germination of seeds of fifteen leguminous plants in the northern Xinjiang region.Methods The seeds were collected from fifteen leguminous species which were widely distributed in the natural mowing pasture of the northern Xinjiang region. After determining their morphological characteristics (seed length, width, thickness, single seed mass, and seed shape index), seeds were fed to sheep. Faeces collection was carried out at 6, 12, 24, 36, 48 and 72 h after seeds were eaten by sheep. Seed recovery percentage (SRP), mean retention time (MRT) and seed germinability before and after ingested were determined, and the relationships between seed size, seed shape index and SRP, MRT after ingested were also studied.Important findings Seed mass ranged from 1.50 to 37.68 mg, and seed shape index between 0.001 and 0.12, which indicated the seeds are all medium or large type sphericity (round) seeds. Seed excretion dynamic followed a Gaussian model: Y = 0.02 + 0.74e -0.5((X- 29.61)/9.41)2 (R2 = 0.62, p < 0.01), and excretion peak appeared 24-36 h after digestion. Seed recovery rate was ranged from 39.25% (Glycyrrhiza glabra) to 4.28% (Melilotus officinalis). The MRT ranged from 37.19 h (Melilotus officinalis) to 22.33 h (Oxytropis sinkiangensis). The relationship between SRP and seed size is expressed as the equation Y = 6.45 + 2.05X - 0.04X2 (R2 = 0.41, p < 0.05). The relationship between seed SRP and seed shape index is expressed as the equation Y = 2.59 + 36.97e -24.47X (R2 = 0.37, p < 0.05). The relationship between MRT and seed size is expressed as the equation Y = 12.48 + 37.44e -0.07X (R2 = 0.37, p < 0.05). The relationship between MRT and seed shape index is expressed as the equation Y = 3.93 + 2055.33X - 21757.99X2 (R2 = 0.42, p < 0.05). The results suggested that leguminous seeds in larger, or smaller, or irregular shape were more easily digested and excreted by sheep. After digested by sheep gut, germination percentages of recovered seeds of Caragana pleiophylla (35.17%) was significantly decreased (p < 0.05) compared with that of seeds before ingested (96.22%). Similarly, seed germination percentages of Trifolium lupinaster and Astragalus alopecurus seeds were decreased than those of no ingested seeds (not significantly, p > 0.05), but the germination percentages of other twelve kinds seeds were significantly increased (p < 0.05). Keywords:seed endozoochore dispersal;northern Xinjiang region;leguminous plant;seed mass;shape index;retention time;seed recovery percentage
PDF (1034KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王树林, 鲁为华, 陈乙实, 景鹏成. 北疆地区15种豆科植物种子对绵羊消化道作用的响应. 植物生态学报, 2018, 42(2): 185-194 doi:10.17521/cjpe.2017.0118 WANG Shu-Lin, LU Wei-Hua, CHEN Yi-Shi, JING Peng-Cheng. Effect of sheep digestive tract on the recovery and germination of seeds of fifteen leguminous plants in the northern Xinjiang region, China. Chinese Journal of Plant Ecology, 2018, 42(2): 185-194 doi:10.17521/cjpe.2017.0118
Table 2 表2 表215种豆科种子经绵羊采食后的种子回收率及种子在绵羊体内的平均滞留时间(平均值±标准误差) Table 2Seed recovery percentages and mean retention time of 15 leguminous species after sheep ingestion (mean ± SE)
种名 Species
种子回收率 SRP (%)
平均滞留时间 MRT (h)
种名 Species
种子回收率 SRP (%)
平均滞留时间 MRT (h)
洋甘草 Glycyrrhiza glabra
39.25 ± 20.40a
31.62 ± 0.73bcd
骆驼刺 Alhagi sparsifolia
25.06 ± 9.25ab
28.27 ± 0.27bcd
草木犀 Melilotus officinalis
4.28 ± 1.76c
37.19 ± 2.12ab
山羊豆 Galega officinalis
38.02 ± 9.60a
27.55 ± 1.75bcd
苦马豆 Sphaerophysa salsula
38.60 ± 19.77a
32.48 ± 0.39abcd
铃铛刺 Halimodendron halodendron
34.56 ± 15.89a
36.51 ± 5.93ab
蒙古黄耆 Astragalus mongholicus
4.88 ± 1.02c
36.62 ± 0.54ab
野苜蓿 Medicago falcata
9.41 ± 1.26bc
24.58 ± 0.84cd
木黄耆 Astragalus arbuscula
5.02 ± 1.25c
32.51 ± 3.93abcd
细叶野豌豆 Vicia tenuifolia
34.90 ± 2.27a
32.78 ± 2.94abcd
新疆棘豆 Oxytropis sinkiangensis
16.04 ± 4.11bc
22.33 ± 0.77d
狐尾黄耆 Astragalus alopecurus
25.61 ± 11.75ab
34.60 ± 0.84abc
多叶锦鸡儿 Caragana pleiophylla
8.93 ± 4.24bc
33.76 ± 1.19abc
弯果胡卢巴 Trigonella arcuata
6.65 ± 1.44c
24.95 ± 1.34cd
野火球 Trifolium lupinaster
14.01 ± 7.18bc
30.04 ± 1.49bcd
SRP, seed recovery percentage; MRT, mean retention time. Different lowercase letters within the same column indicate significant difference (p < 0.05). 同列不同小写字母表示差异显著(p < 0.05)。
Table 3 表3 表3绵羊消化前后15种豆科种子的萌发率(平均值±标准误差) Table 3Germination percentages of seeds of 15 leguminous species before and after sheep ingestion (mean ± SE)
种名 Species
萌发率 Germination rates (%)
萌发率增值 Appreciation of germination rates (%)
对照 Control
消化后 After sheep ingested
洋甘草 Glycyrrhiza glabra
15.36 ± 5.55b
68.41 ± 13.86a
53.05
草木犀 Melilotus officinalis
6.67 ± 1.16b
23.68 ± 1.67a
17.01
苦马豆 Sphaerophysa salsula
15.00 ± 7.00b
71.01 ± 3.62a
56.01
蒙古黄耆 Astragalus mongholicus
14.33 ± 3.22b
25.00 ± 3.61a
10.67
木黄耆 Astragalus arbuscula
45.00 ± 2.00b
53.70 ± 1.57a
8.70
新疆棘豆 Oxytropis sinkiangensis
33.33 ± 1.54a
35.39 ± 3.08a
2.06
多叶锦鸡儿 Caragana pleiophylla
96.22 ± 6.33a
35.17 ± 3.32b
-61.05
野火球 Trifolium lupinaster
28.33 ± 3.06a
21.43 ± 3.11a
-6.90
骆驼刺 Alhagi sparsifolia
8.33 ± 3.51b
42.25 ± 2.63a
33.92
山羊豆 Galega officinalis
13.33 ± 1.53a
14.45 ± 1.27a
1.12
铃铛刺 Halimodendron halodendron
1.67 ± 0.58b
11.96 ± 2.00a
10.29
野苜蓿 Medicago falcata
7.82 ± 1.05b
62.36 ± 3.01a
54.54
细叶野豌豆 Vicia tenuifolia
8.33 ± 0.58b
28.52 ± 6.73a
20.19
狐尾黄耆 Astragalus alopecurus
6.30 ± 2.14a
5.83 ± 2.56a
-0.47
弯果胡卢巴 Trigonella arcuata
5.20 ± 1.31b
20.32 ± 5.51a
15.12
Negative value indicates that the germination rates decreased. Different lowercase letters within the same row indicate significant difference (p < 0.05). 负值表示萌发率降低。同行不同小写字母表示差异显著(p < 0.05)。
BaskinCC, BaskinJM ( 2014). Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. 2nd edn. Academic Press, New York.
BlackshawRE, RodeLM ( 1991). Effect of ensiling and rumen digestion by cattle on weed seed viability Weed Science, 39, 104-108. DOI:10.1007/BF00224983URL [本文引用: 1] Studies were conducted to determine the effect of ensiling and/or rumen digestion by cattle on the germination and viability of several common weed species. Seed survival of grass species subjected to ensiling and/or rumen digestion tended to be less than that of broadleaf species. Downy brome, foxtail barley, and barnyardgrass were nonviable after either ensiling for 8 weeks or rumen digestion for 24 h. Some green foxtail (17%) and wild oats (0 to 88%) seeds survived digestion in the rumen but were killed by the ensiling process. Varying percentages of seeds of kochia, redroot pigweed, common lambsquarters, wild buckwheat, round-leaved mallow, and field pennycress remained viable after ensiling (3 to 30%), rumen digestion (15 to 98%), and ensiling plus rumen digestion (2 to 19%). A time course study of rumen digestion indicated that loss of seed viability often was not a gradual process. With some species, there was an initial lag phase while degradation of the protective seed coat likely occurred, followed by a rapid decline in embryo viability. The diet fed to livestock appeared to affect viability losses caused by rumen digestion. Estimates of seed survival with varying rates of passage through the rumen due to differing ratios of grain to forage in the diet are presented.
BrochetAL, GuillemainM, Gauthier-ClercM, FritzH, GreenAJ ( 2010). Endozoochory of Mediterranean aquatic plant seeds by teal after a period of desiccation: Determinants of seed survival and influence of retention time on germinability and viability Aquatic Botany, 93, 99-106. DOI:10.1016/j.aquabot.2010.04.001URL [本文引用: 1] The capacity of eight wetland seeds to be internally dispersed by common teal ( Anas crecca L.) after a dry period typical of natural Mediterranean wetlands was experimentally quantified. These plant taxa ( Chara spp., Echinochloa crus-galli (L.) P. Beauv., Eleocharis palustris (L.) Roem. & Schult, Polygonum lapathifolium L., Potamogeton nodosus Poir., Potamogeton pusillus L., Schoenoplectus mucronatus (L.) Palla, Scirpus maritimus L.) were frequently ingested by teal in the field. Captive teal were fed with known quantities of dry seeds and faecal samples were collected at intervals of 1–2 h for 48 h. The proportion of seeds retrieved after gut passage that germinated varied between taxa from 3% to 83%. From 2% to 63% of the seeds ingested were recovered intact, and 18-95% of these were still viable. Therefore, an overall 32% of the seeds ingested were evacuated in a viable condition, ranging from 0.2% for Chara spp. to 54% for S. mucronatus. Mean seed retention time was 3–5 h. Overall, 94% of seeds were evacuated within 12 h, but 2% were recovered after more than 24 h. Seed viability was reduced at longer retention times, although viable seeds of all taxa except Chara spp. were recovered 44–48 h after the experiment began. Germinability was increased by gut passage, although for some species this effect was only significant after stratification, indicating that they exhibit both mechanical and physical dormancy. Seed germination was accelerated by gut passage, especially at longer retention times. Gut passage increased the proportion of viable seeds, suggesting selective digestion of non-viable ones. Endozoochorous transport by teal appears to be a widespread dispersal mechanism among wetland plants.
Calvi?o-CancelaM, Martín-HerreroJ ( 2009). Effectiveness of a varied assemblage of seed dispersers of a fleshy-fruited plant Ecology, 90, 3503-3515. DOI:10.1890/08-1629.1URLPMID:20120817 [本文引用: 1] Disperser effectiveness is the number of new plants resulting from the activity of one disperser relative to other dispersers or to nondispersed seeds. Effectiveness remains largely uninvestigated due to the complexity of its measurement. We measured the effectiveness of seed dispersers (Larus michahellis, Turdus merula, and Oryctolagus cuniculus) of the shrub Corema album (Empetraceae) using a simulation model of the recruitment process that was parameterized with field data of seed dispersal, predation, and seedling emergence and validated with independent data on seedling density. The model allows tracking the fate of seeds dispersed by each animal and estimating, for the first time, disperser effectiveness as seedlings per square meter contributed by each disperser. It also allows quantifying the relative importance of different recruitment processes in determining the quantity and spatial distribution of recruitment. Larus michahellis was the most effective disperser in two of the three habitats studied, contributing 3-125 times more than the other two species, whose lower effectiveness depended mostly on deposition patterns (T. merula) or deleterious effects on seedling emergence (O. cuniculus). The dependence of the plant on each disperser differed between habitats and was the greatest in sparse scrub, where recruitment depended almost exclusively on gulls (90%). Quantity and quality of dispersal were not correlated; quality was a better predictor of disperser effectiveness. Seedling emergence was the most crucial process in determining both the spatial pattern of recruitment among microhabitats (99.8% of variance explained) and the quantity of recruitment within microhabitats (43-83%). A sensitivity analysis showed that increasing seed dispersal improved the recruitment for all dispersers when there was no competition for fruits. However, with limited fruits, increased dispersal of lower quality dispersers reduced overall recruitment. Our results show important differences in effectiveness among dispersers and illustrate the different influences of the components of effectiveness, which varied depending not only on the disperser but also on the circumstances (e.g., type of habitat).
CosynsE, DelporteA, LensL, HoffmannM ( 2005). Germination success of temperate grassland species after passage through ungulate and rabbit guts Journal of Ecology, 93, 353-361. DOI:10.1111/j.0022-0477.2005.00982.xURL [本文引用: 1] Summary 1 Dispersal of endozoochorous seed involves uptake by a herbivore and exposure to different kinds of digestive fluids during passage through the gastrointestinal tract. Assessment of the ecological significance of endozoochory therefore requires examination of the survival rate of seeds during this phase. 2 A feeding experiment was conducted with seeds of 19 plant species that are important constituents of temperate semi-natural grasslands and five animal species (two ruminants, two colon fermenters and a caecum fermenter). Mean retention time of germinable seeds was determined and seed characteristics that might affect germination success were examined. 3 Gut-passed seeds had a much lower germination success (0–26%) than non-gut-passed seeds either sown directly on dung (2–79%) or bare soil (7–89%). 4 Relative germination success differed considerably between both plant and animal species. This may result from complex, herbivore-specific interactions between animal behaviour (chewing, digestion) and seed characteristics. 5 Germination success was positively related to seed longevity and, remarkably, also to seed mass and seed shape. Retention time of germinable seeds varied from c . 12 hours (rabbit) to 72 hours (ungulates), potentially allowing long-distance seed dispersal. This study highlights both the complex interaction between animal species and seed characteristics and the considerable differences in germination success of gut-passed seeds, which exist between plant species. The loss of seed germinability after gut passage calls into question the ecological significance of endozoochory, although the costs of other dispersal mechanisms remain to be tested.
CouvreurM, CosynsE, HermyM, HoffmannM ( 2005). Complementarity of epi- and endozoochory of plant seeds by free ranging donkeys Ecography, 28, 37-48. DOI:10.1111/j.0906-7590.2005.04159.xURL [本文引用: 1] Epizoochory and endozoochory are well-recognized long-distance seed dispersal mechanisms, yet their relative importance has hardly been studied before. Here, epi- and endozoochory were compared on donkeys foraging in a species-rich 100 ha coastal dune ecosystem, through in vitro germination of zoochorous material obtained by fur brushing and dung collection. We identified 6675 seedlings of 66 plant species, covering 20% of the species recorded in the study area. Of the 66 species, only 16 occurred in both epi- and endozoochory samples, demonstrating the complementarity of both dispersal mechanisms. The species composition in the zoochory samples reflected a strong seasonality, and seedling numbers were partly correlated with species abundance in the study area. The non-zoochorously dispersed species in the study area differed from the zoochorous species in seed size and weight, plant height, life span, dispersal strategy and seedbank persistence. Dispersal-relevant plant traits were used to derive dispersal-functional plant types for all species in the study area. Epizoochory showed to be more specific than endozoochory and was associated with a more narrow range of dispersal-functional plant types.
D’HondtB, HoffmannM ( 2011). A reassessment of the role of simple seed traits in mortality following herbivore ingestion Plant Biology, 13, 118-124. DOI:10.1111/j.1438-8677.2010.00335.xURLPMID:21134095 [本文引用: 2] Abstract Grazing mammals are regarded as major vectors in seed dispersal of grassland plants, through seed ingestion and subsequent excretion (endozoochory). The (evolutionary) ecology underlying this dispersal mode is relatively poorly understood because there are limited data, among others, on how seed attributes perform in this process (and could thus be selected for). For seed mortality following ingestion, contrasting patterns described for the role of simple seed traits seem partly due to inadequate comparative analytical methods. We conducted a feeding experiment in which controlled seed quantities from 48 grassland herb and grass species were fed to cattle. Seed mass, length and shape measurements were related to seed mortality rate using phylogenetically independent contrasts, which account for taxonomic interdependence. The proportion of seeds surviving the digestive tract was generally low, but it reached 100%, with increased germination for some species. Neither seed size nor shape correlated significantly with mortality. Structural traits are likely to overrule these simple seed traits, as illustrated by high survival of species having water-impermeable seeds. As this trait has interesting properties with respect to intraspecific variation and genetics, it could provide opportunities for inference on evolutionary consequences for this type of dispersal.
G?kbulakF ( 2002). Effect of American bison ( Bison bison L.) on the recovery and germinability of seeds of range forage species. Grass & Forage Science, 57, 395-400. [本文引用: 1]
G?kbulakF ( 2006). Recovery and germination of grass seeds ingested by cattle The Journal of Biological Sciences, 6, 23-27. DOI:10.3844/ojbsci.2006.23.27URL [本文引用: 1] ABSTRACT Seeds of bluebunch wheatgrass [Psuedoroegneria spicata (Pursh) A. Love] and Sandberg bluegrass (Poa secunda Presl.) were fed to Holstein heifers in different amounts to investigate the effects of seed feeding level and seed size on the recovery and germinability of passed seed. Animals were fed 60,000, 30,000,15,000, and 7,500 seeds of each species. Passed seeds were recovered from dungcollected daily over a 4-day period and tested for germinability. In general,recovery of the largerseeded bluebunch wheatgrass and the smaller-seeded Sandberg bluegrass declined as seed feeding levels decreased from 60,000 to 7,500 seeds per animal, and as time after seed ingestion increased from 1 to 4 days. Total seed recovery over the 4-day period was greater for bluebunch wheatgrass at the 60,000 seed feeding level, similar for both species at the 30,000 seed feeding level, and greater for Sandberg bluegrass at the 15,000 and 7,500 seed feeding levels.Germinability of bluebunch wheatgrass seeds decreased with each additional day in the digestive tract, while germinability of Sandberg bluegrass seeds remained constant or increased with time. Germinability of both species tended to increase as seed feeding levels decreased from 60,000 to 15,000 seeds per animal. Sandberg bluegrass seeds had greater germinability than bluebunch wheatgrass at all seed feeding levels and collection dates. Recovered seeds had significantly lower germinability than noningested seeds for both species.Results showed that livestock have the potential for dispersing enough germinable seeds on degraded rangelands.
GrandeD, MartinvicenteA ( 2013). Endozoochorus seed dispersal by goats: Recovery, germinability and emergence of five Mediterranean shrub species Spanish Journal of Agricultural Research, 11, 347-355. DOI:10.5424/sjar/2013112-3673URL [本文引用: 1] Herbivores can play an important role as seed dispersal vectors, ungulates constituting potential seed dispersal agents of Mediterranean grasses and shrubs. We evaluated the role of domestic goats as seed dispersers of five representative Mediterranean forage shrub species [Cistus albidus L., Phillyrea angustifolia L., Calicotome villosa (Poir.) Link., Rhamnus lycioides L. and Atriplex halimus ...
HeidrunW, OliverT ( 2008). A mechanistic simulation model of seed dispersal by animals Journal of Ecology, 96, 1011-1022. [本文引用: 1]
JanzenDH ( 1977). Variation in seed size within a crop of a costa rican Mucuna andreana( Leguminosae). American Journal of Botany, 64, 347-349.
JanzenDH ( 1984). Dispersal of small seeds by big herbivores: Foliage is the fruit The American Naturalist, 123, 338-353. [本文引用: 2]
JaroszewiczB ( 2013). Endozoochory by European bison influences the build-up of the soil seed bank in subcontinental coniferous forest European Journal of Forest Research, 132, 445-452. [本文引用: 1]
JuradoE, EstradaE, MolesA ( 2001). Characterizing plant attributes with particular emphasis on seeds in Tamaulipan thornscrub in semi-arid Mexico Journal of Arid Environments, 48, 309-321.
KleinJ ( 1981). La Mesta. 2nd edn. Alianza Editorial, Madrid Spain. [本文引用: 1]
LuWH, WanJJ, YangJJ, RenAT, YuL ( 2013). Review of endozoochory of plant seeds by herbivores Acta Prataculturae Sinica, 22, 306-313. [本文引用: 2]
MaSB, JiangHQ ( 1999). Study on the seed weight and seed seize variation pattern and their biological significance in Podophylloideae (Berberjdaceae) Acta Botanica Boreali-?Occidentalia Sinica, 19, 715-724. [本文引用: 1]
Mancilla-LeytónJM, Fernández-AlésR, VicenteAM ( 2012). Plant-ungulate interaction: Goat gut passage effect on survival and germination of Mediterranean shrub seeds Journal of Vegetation Science, 22, 1031-1037. [本文引用: 1]
ManzanoP, MaloJE, PecoB ( 2005). Sheep gut passage and survival of Mediterranean shrub seeds Seed Science Research, 15, 21-28. [本文引用: 5]
Miloti?T, HoffmannM ( 2016). How does gut passage impact endozoochorous seed dispersal success? Evidence from a gut environment simulation experiment Basic and Applied Ecology, 17, 165-176. [本文引用: 2]
MouXM, YuYW, ZhangHM, SunH, WangHC, XuCL, HuaLM ( 2013). Effects of yakdung deposition on community characteristics and niche parameters in alpine meadow Pratacultural Science, 30, 1594-1601. [本文引用: 1]
MouissieAM, VeenCEJVD, VeenGF, DiggelenRV ( 2005 a). Ecological correlates of seed survival after ingestion by fallow deer Functional Ecology, 19, 284-290. [本文引用: 2]
MouissieAM, VosP, VerhagenHMC, BakkerJP ( 2005 b). Endozoochory by free-ranging, large herbivores: Ecological correlates and perspectives for restoration Basic and Applied Ecology, 6, 547-558. [本文引用: 1]
MyersJA, VellendM, GardescuS, MarksPL ( 2004). Seed dispersal by white-tailed deer: Implications for long-????distance dispersal, invasion, and migration of plants in eastern North America Oecologia, 139, 35-44. [本文引用: 2]
PlumptreAJ ( 2003). Seed germination and early seedling establishment of some elephant-dispersed species in Banyang-?mbo wildlife sanctuary, South-western Cameroon Journal of Tropical Ecology, 19, 229-237. [本文引用: 2]
RamosME, RoblesAB, CastroJ ( 2006). Efficiency of endozoochorous seed dispersal in six dry-fruited species (Cistaceae): From seed ingestion to early seedling establishment Plant Ecology, 185, 97-106. [本文引用: 1]
RussiL, CocksPS, RobertsEH ( 1992). The fate of Legume seeds eaten by sheep from a Mediterranean grassland Journal of Applied Ecology, 29, 772-778. [本文引用: 1]
SoonsMB, van der VlugtC, Van LithB, HeilGW, KlaassenM ( 2008). Small seed size increases the potential for dispersal of wetland plants by ducks Journal of Ecology, 96, 619-627. [本文引用: 1]
SpiegelO, NathanR ( 2007). Incorporating dispersal distance into the disperser effectiveness framework: Frugivorous birds provide complementary dispersal to plants in a patchy environment Ecology Letters, 10, 718-728. [本文引用: 1]
ThompsonK, BandSR, HodgsonJG ( 1993). Seed size and shape predict persistance in soil Functional Ecology, 7, 236-241. [本文引用: 2]
ThomsonFJ, MolesAT, AuldTD, KingsfordRT ( 2011). Seed dispersal distance is more strongly correlated with plant height than with seed mass Journal of Ecology, 99, 1299-1307. [本文引用: 1]
WaliN, WangSL, ZhaoG, YangJJ, LüYP, LuWH ( 2016). Effect of sheep digestion on morphology, recovery and germinability of four Leguminous plants seed Pratacultural Science, 33, 1566-1573. [本文引用: 4]
WangBC, SmithTB ( 2002). Closing the seed dispersal loop Trends in Ecology and Evolution, 17, 379-386. [本文引用: 1]
WangCJ, WangWQ, LuWH, WenCL, YinXJ, ZhaoQZ ( 2016). Feed intake distribution model for herd based on grazing spatio-temporal trajectory data Transactions of the Chinese Society of Agricultural Engineering, 32(13), 125-130. [本文引用: 1]
WangSL, LuWH, WaliN, MaCH, ZhangQB, WangCJ ( 2017). Recovery and germination of seeds after passage through the gut of Kazakh sheep on the north slope of the Tianshan Mountains Seed Science Research, 27, 43-49. [本文引用: 1]
WangSL, WaliN, LuWH, WangWQ, WangCJ ( 2017). Morphological characteristics of six kinds of mucilaginous seeds and their response to digestion in rumen of sheep Acta Prataculturae Sinica, 26(4), 89-98. [本文引用: 1]
WillmsWD, AcharyaSN, RodeLM ( 1995). Feasibility of using cattle to disperse cicer milkvetch ( Astragalus cicer L.) seed in pastures. Canadian Veterinary Journal, 75, 173-175. [本文引用: 1]
YanXH, AnSZ, YuX, JiaSB, PanXD, ChengFX, LiH ( 2009). Observation on grazing behavior of Kazak sheep in summer Journal of Xinjiang Agricultural University, 32(4), 38-40. [本文引用: 1]
YuL, LuWH, YanP, LuoLJ, ZhangQB ( 2014). Resources and assessment of natural grassland in Shaertao Mountains, Zhaosu County in Xinjiang Chinese Journal of Grassland, 36(6), 4-11. [本文引用: 2]
YuXJ, XuCL, WangF, ShangZH, LongRJ ( 2012). Recovery and germinability of seeds ingested by yaks and Tibetan sheep could have important effects on the population dynamics of alpine meadow plants on the Qinghai-Tibetan Plateau Rangeland Journal, 34, 249-255. [本文引用: 1]
ZhaoLL, WangZL, DuJC, LiQF, SongCL ( 2007). The effect of sulfuric acid treatment on hard seeds of Melimotoides ruthenica strains. Chinese Journal of Grassland, 29(3), 73-77. [本文引用: 1]
Effect of ensiling and rumen digestion by cattle on weed seed viability 1 1991
... id="C5">种子消化道传播(seed endozoochore dispersal)是指植物种子被食草(果)动物采食, 经过消化道作用以后, 通过反刍或粪便排出体外从而实现传播的现象(Oudtshoorn & Rooyen, 1998), 动物对植物种子的消化道传播是种子实现远距离传播的重要方式.种子由于动物采食、经消化道携带和排泄而产生空间上的大尺度位移, 一方面为植物种群开拓新栖息地创造了有利条件、避免了同胞间激烈的竞争, 另一方面大大增加了空间尺度上的景观多样性(Heidrun & Oliver, 2008).目前关于草食动物对植物种子消化道传播的研究, 所涉及的动物种类有牛(Willms et al., 1995)、绵羊(Ramos et al., 2006)、山羊(Mancilla-Leytón et al., 2012)、马(Miloti? & Hoffmann, 2016)和驴(Couvreur et al., 2005)等放牧家畜.研究的生态类型包括森林(Jaroszewicz, 2013)、草原(Myers et al., 2004)和沼泽(Brochet et al., 2010)等.具体研究内容包括: 种子自身形态学特征与种子被消化道作用后的回收率、活力以及在消化道内的滞留时间之间的关系(Blackshaw & Rode, 1991), 排出后的种子萌发所形成的幼苗的生长状况以及消化道传播对植被更新过程的影响(Plumptre, 2003), 并且基于上述研究内容可进一步探讨植物与草食动物二者之间是否存在协同进化(D’Hondt & Hoffmann, 2011). ...
Endozoochory of Mediterranean aquatic plant seeds by teal after a period of desiccation: Determinants of seed survival and influence of retention time on germinability and viability 1 2010
... id="C5">种子消化道传播(seed endozoochore dispersal)是指植物种子被食草(果)动物采食, 经过消化道作用以后, 通过反刍或粪便排出体外从而实现传播的现象(Oudtshoorn & Rooyen, 1998), 动物对植物种子的消化道传播是种子实现远距离传播的重要方式.种子由于动物采食、经消化道携带和排泄而产生空间上的大尺度位移, 一方面为植物种群开拓新栖息地创造了有利条件、避免了同胞间激烈的竞争, 另一方面大大增加了空间尺度上的景观多样性(Heidrun & Oliver, 2008).目前关于草食动物对植物种子消化道传播的研究, 所涉及的动物种类有牛(Willms et al., 1995)、绵羊(Ramos et al., 2006)、山羊(Mancilla-Leytón et al., 2012)、马(Miloti? & Hoffmann, 2016)和驴(Couvreur et al., 2005)等放牧家畜.研究的生态类型包括森林(Jaroszewicz, 2013)、草原(Myers et al., 2004)和沼泽(Brochet et al., 2010)等.具体研究内容包括: 种子自身形态学特征与种子被消化道作用后的回收率、活力以及在消化道内的滞留时间之间的关系(Blackshaw & Rode, 1991), 排出后的种子萌发所形成的幼苗的生长状况以及消化道传播对植被更新过程的影响(Plumptre, 2003), 并且基于上述研究内容可进一步探讨植物与草食动物二者之间是否存在协同进化(D’Hondt & Hoffmann, 2011). ...
Effectiveness of a varied assemblage of seed dispersers of a fleshy-fruited plant 1 2009
Incorporating dispersal distance into the disperser effectiveness framework: Frugivorous birds provide complementary dispersal to plants in a patchy environment 1 2007
... id="C8">供试种子采集于天山北坡中段山地(84.97°- 86.40° E, 43.43°-45.33° N, 海拔1β935-2β252 m)、塔城地区的巴尔鲁克山天然割草场(82.35°-82.41° E, 45.57°-45.58° N, 海拔1β100-1β600 m)和伊犁沙尔套山的山前倾斜平原(山麓带)(80.15°-80.54° E, 42.54°- 43.11° N, 海拔2β200-2β300 m), 共计15种, 分别是洋甘草(Glycyrrhiza glabra)、草木犀(Melilotus officinalis)、苦马豆(Sphaerophysa salsula)、蒙古黄耆(Astragalus mongholicus)、木黄耆(Astragalus arbuscula)、新疆棘豆(Oxytropis sinkiangensis)、多叶锦鸡儿(Caragana pleiophylla)、野火球(Trifolium lupinaster)、骆驼刺(Alhagi sparsifolia)、山羊豆(Galega officinalis)、铃铛刺(Halimodendron halodendron)、野苜蓿(Medicago falcata)、细叶野豌豆(Vicia tenuifolia)、狐尾黄耆(Astragalus alopecurus)和弯果胡卢巴(Trigonella arcuata).按适口性等级划分, 这15种豆科植物均属于优级或良级牧草, 即适口性极好, 一般情况下各类家畜均喜食或从草地中优先挑选采吃(于磊等, 2014).根据不同植物种子的成熟时期, 于2015年6-10月间进行种子采集.种子采集后放在室内通风处自然风干后, 装入信封袋内于4 ℃冰箱中保存备用. ...
Recovery and germinability of seeds ingested by yaks and Tibetan sheep could have important effects on the population dynamics of alpine meadow plants on the Qinghai-Tibetan Plateau 1 2012
 ,, 鲁为华
,, 鲁为华 ,*, 陈乙实*, 景鹏成*石河子大学动物科技学院, 新疆石河子 832003
,*, 陈乙实*, 景鹏成*石河子大学动物科技学院, 新疆石河子 832003
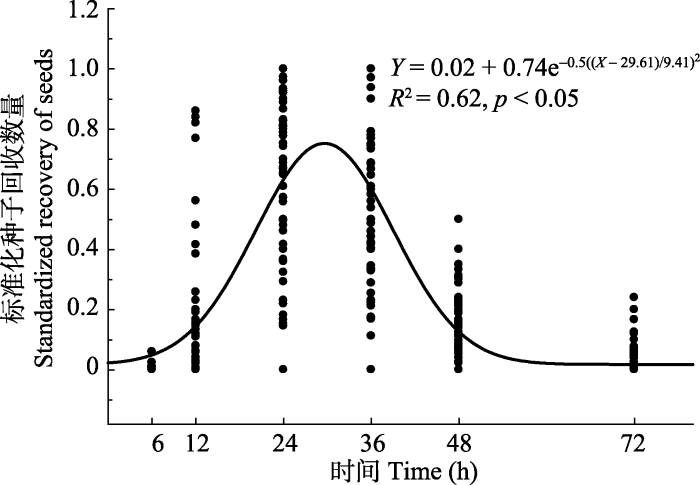 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT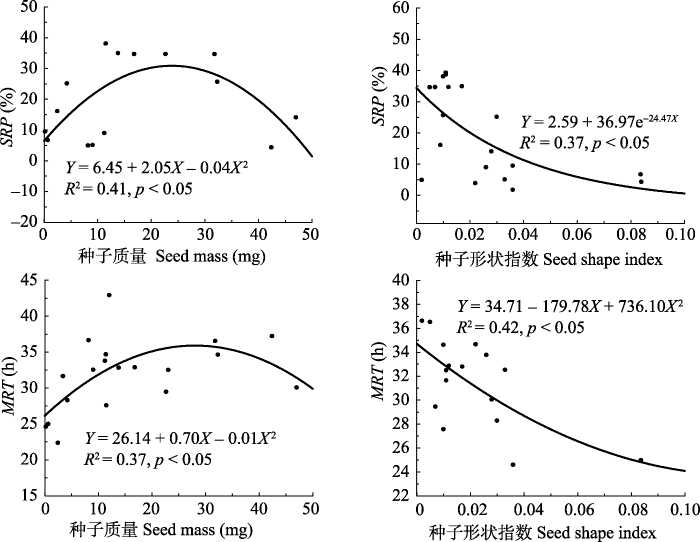 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}