Spatial distribution and influencing factors of soil organic carbon among different climate types in Sichuan, China
WANG Li-Hua,1,2, XUE Jing-Yue2,3, XIE Yu2, WU Yan,2,*1 Aba Teachers University, Wenchuan, Sichuan 623002, China 2 Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China 3University of Chinese Academy of Sciences, Beijing 100049, China
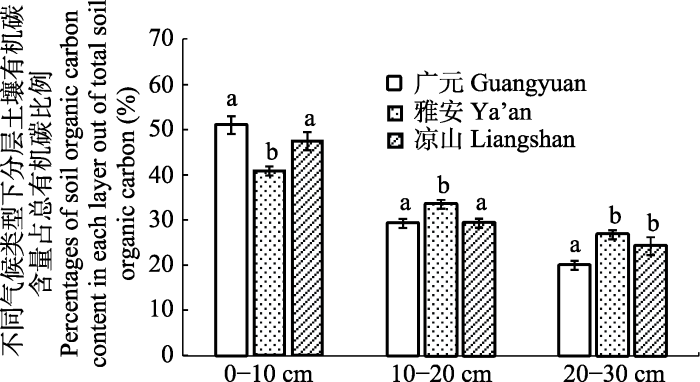
Abstract Aims Spatial distribution and influencing factors of soil organic carbon (SOC) content among different climate types were studied to gain new insights into the estimation and dynamics of SOC.Methods The study areas are located in the mountain meadow in Guanyuan City, Ya’an City, Liangshan Prefecture of Sichuan Province, China. Plant populations were recorded according to species, number, coverage, meantime, and soil samples (0-10, 10-20, 20-30 cm) were collected and analyzed for SOC content, the carbon content of root, soil pH value, and soil total nitrogen. The diversity indices for plant community diversity (Shannon-? Wiener index) was also calculated to analyze their relationships with SOC content among different climate types.Important findings Results showed that: (1) SOC in three sites was significantly different and was in the order of Ya’an > Liangshan Prefecture > Guanyuan, and decreased with soil depth. (2) The percentage of SOC content in each vertical layer out of total SOC was lower in the 0-10 cm layer at Ya’an site than at the other two sites, but was higher than the other two sites in the 10-20 cm layer. (3) At Guanyuan site, SOC content in 10-20 cm was significantly positively correlated with plant species richness index and SOC content in 20-30 cm was negatively correlated with plant Shannon-Winner index, while soil pH value and soil total nitrogen were significantly positively correlated with SOC content in each layer. At Ya’an site, SOC content in each layer only had positive correlation with soil pH value, but not with other examined factors. At Liangshan Prefecture site, SOC content in each layer only had positive correlation with soil nitrogen content, but not with other examined factors. (4) Principal component analysis showed that, at Guanyuan, Ya’an and Liangshan Prefecture sites, the total contents of SOC were dominantly affected by climate, followed by vegetation type. Keywords:climate factor;grassland;soil organic carbon;distribution characteristics;correlation analysis
PDF (912KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王丽华, 薛晶月, 谢雨, 吴彦. 不同气候类型下四川草地土壤有机碳空间分布及影响因素. 植物生态学报, 2018, 42(3): 297-306 doi:10.17521/cjpe.2017.0061 WANG Li-Hua, XUE Jing-Yue, XIE Yu, WU Yan. Spatial distribution and influencing factors of soil organic carbon among different climate types in Sichuan, China. Chinese Journal of Plant Ecology, 2018, 42(3): 297-306 doi:10.17521/cjpe.2017.0061
Table 2 表2 表2不同地区0-30 cm土壤有机碳含量之间的比较(平均值±标准误差, n = 7) Table 2Soil organic carbon content (0-30 cm) in different areas (mean ± SE, n = 7)
地区 Area
土壤有机碳含量 Soil organic carbon content (kg·m-2)
0-30 cm
0-10 cm
10-20 cm
20-30 cm
广元 Guangyuan
37.91 ± 3.76a
18.42 ± 1.61Aa
11.46 ± 1.40Ba
8.02 ± 1.21Ba
雅安 Ya’an
62.18 ± 4.40b
25.04 ± 1.66Ab
20.80 ± 1.61ABb
16.34 ± 1.56Bb
凉山 Liangshan
51.06 ± 3.54c
23.44 ± 1.34Ab
15.14 ± 1.31Ba
12.48 ± 1.48Bb
Different lowercase letters showed significant difference among different areas (p < 0.05). Different capital letters showed significant difference among different soil layers (p < 0.05). 不同小写字母表示各地区间的差异显著(p <0.05); 不同大写字母表示不同土壤层次间的差异显著(p <0.05)。
新窗口打开|下载原图ZIP|生成PPT 图1不同地区土壤分层有机碳含量占总有机碳含量比例(平均值±标准误差, n = 7)。不同小写字母表示不同土层间差异显著(p < 0.05)。
Fig. 1The percentages of soil organic carbon content in each layer out of total soil organic carbon in the three sites (mean ± SE, n = 7). Different lowercase letters showed significant difference among different soil layers (p < 0.05).
Table 4 表4 表4不同地区土壤分层有机碳含量与植被群落特征及土壤指标之间的相关系数 Table 4The relationship between soil carbon content in each layer and plant community characteristics and soil parameters in different areas
AlverzR, LavadoRS ( 1998). Climate organic matter and clay content relationship in the Pampa and Chaco, Argentina Geoderma, 83, 127-141. DOI:10.1016/S0016-7061(97)00141-9URL [本文引用: 2] Temperature and precipitation have strong effects on soil processes. The Pampa and the Chaco are vast plains with soils mainly derived from loess. Our objective was to study the effects of temperature and precipitation of both regions on selected soil properties. Using data from soil surveys for ca. 65 Mha, we defined 40 geographic units of ca. 1.6 Mha each. Organic carbon content (g cm), solum thickness and clay content were averaged for each unit. Temperature, precipitation and potential evapotranspiration were obtained from climatic records. Carbon inputs to the different soil layers were estimated by calculating net primary productivity of ecosystems and partitioning coefficients of above and belowground biomass. Inputs were affected by a retention factor taking into account temperature effects on organic matter decomposition. Soil organic carbon increases with higher precipitation and decreases with higher temperature. Consequently, the organic carbon content in the top 0–50 cm soil layer is positively correlated with the precipitation/temperature ratio (potential model=0.693,<0.001). Carbon inputs to the soil explained 72% of organic carbon variations. Mean turnover time of soil carbon was estimated to be 14 yr. Solum thickness and clay amount (g cm) are not associated with temperature but the ratio precipitation/potential evapotranspiration explained 85% (<0.001) of the variation in the former and 79% (<0.001) in the later. Soils are deeper and with higher clay contents where this ratio increases, a consequence of available water for pedogenic processes. Climate is a main soil forming factor in the Pampa and Chaco regions. Clay neoformation and possible future trends of organic carbon under different climatic global change scenarios were estimated.
CarsonWP, PickettSTA ( 1990). Role of resources and disturbance in the organization of an old-field plant community Ecology, 71, 226-238. DOI:10.2307/1940262URL [本文引用: 1] The roles of disturbance and resource availability in influencing species richness and plant abundance were examined by conducting a factorial experiment for 2 yr in an old field in New Jersey, USA, dominated by goldenrods (Solidago spp.). Replicate plots were treated by (1) adding macronutrients, (2) adding water, (3) tying back tall herbs to increase light to ground layer species, and (4) creating gaps in the vegetation by disturbing the soil with hand trowel early in the growing season. Nutrient additions increased the cover of the herbaceous canopy and enhanced the dominance of Solidago spp. Water additions also increased canopy during the second growing season, a year with low seasonal rainfall. Light enrichment as a consequence of canopy manipulation resulted in the competitive release of subcanopy species, primarily Fragaria virginiana. Other subcanopy dominants were limited by different resources; Hieracium pratense responded most strongly to water and Rumex acetosella to nutrients. These differences in limiting resources may be responsible for the continued coexistence of these three subcanopy species. Additionally, we observed marked seasonal and yearly fluctuations in the severity of resource limitation which might serve to favor different species in this community at different times. Species richness was decreased by light enrichment alone, whereas light with irrigation interacted to increase species richness. Nutrient additions had little direct effect on species richness A single small disturbance at the growing season had little impact on species richness or abundance. Relative to the influence of resources, small-scale disturbance plays only a minor role in this community. Our results suggest that nonequilibrium organization of old-field communities can occur via a shifting resource base in the absence of natural disturbances.
ChenFR, ChengJM, LiuW, LiY, ChenA, ZhaoXY ( 2012). Effect of disturbances on organic soil carbon in the typical grassland of Loess Plateau Acta Agrestia Sinica, 20, 298-304, 311. [本文引用: 1]
ChengXF, ShiXZ, YuDS, FanXZ ( 2004). Spatial variance and distribution of total nitrogen and organic matter of soil in Xingguo County of Jiangxi, China Chinese Journal of Applied and Environmental Biology, 10, 64-67. [本文引用: 2]
DavidsonEA, TrumboreSE, AmundsonR ( 2000). Soil warming and organic carbon content Nature, 408, 789-790. DOI:10.1038/35048672URLPMID:11130707 [本文引用: 1] Soils store two or three times more carbon than exists in the atmosphere as CO, and it is thought that the temperature sensitivity of decomposing organic matter in soil partly determines how much carbon will be transferred to the atmosphere as a result of global warming. Giardina and Ryan have questioned whether turnover times of soil carbon depend on temperature, however, on the basis of experiments involving isotope analysis and laboratory incubation of soils. We believe that their conclusions are undermined by methodological factors and also by their turnover times being estimated on the assumption that soil carbon exists as a single homogeneous pool, which can mask the dynamics of a smaller, temperature-dependent soil-carbon fraction. The real issue about release of carbon from soils to the atmosphere, however, is how temperature, soil water content and other factors interact to influence decomposition of soil organic matter. And, contrary to one interpretation of Giardina and Ryan's results, we believe that positive feedback to global warming is still a concern.
EL-ghamryAM, HuangCY, XuJM, XieZM ( 2000). Changes in soil biological properties with the addition of metsulfuron-?methyl herbicide Journal of Zhejiang University (Science), 1, 442-447. DOI:10.1631/jzus.2000.0442URL [本文引用: 1] An incubation study was conducted to investigate the effects of metsulfuron-methyl herbicide on the soil microbial biomass in loamy sand soil. The herbicide was applied to the soil at four concentrations: control, 0.01, 0.10, and 1.00 μg·g 611 soil. Determinations of microbial biomass-C and microbial biomass-N contents were carried out 1, 3, 5, 7, 10, 15, 25, and 45 days after herbicide application. In comparison to untreated soil, the microbial biomass-C and biomass-N decreased significantly in soils treated with herbicide at concentrations of 0.1 and 1.0 μg·g 611 soil within the first 7 days of incubation. The application of metsulfuron-methyl herbicide to the soil reduced the C mic /C org and N mic /N total percentages, which decreased with increasing application rate of metsulfuron-methyl herbicide. Compared to the untreated control, a marked increase in the microbial biomass C: N ratio was observed in the herbicide treated soil. This effect was transitory and was significant only at the higher rates of metsulfuron-methyl.
FangJY, ChenAP ( 2001). Dynamic forest biomass carbon pools in China and their significance Acta Botanica Sinica, 43, 967-973. DOI:10.1614/0890-037X(2001)015[0892:SSPCBP]2.0.CO;2URL [本文引用: 1] An improved biomass estimation method that converts timber volume to total forest biomass was used to estimate forest biomass carbon storage and its changes over to the past 50 years in China, by using inventory data of seven periods from 1949 to 1998 and forest biomass data obtained from direct field measurements. By the mid-1970s, forest carbon decreased by 0.62 Pg C (Pg=10 15 g), with a decreasing rate of 0.024 Pg C per year. Since then the biomass carbon showed a significant increase, from 4.38 Pg C by the end of 1970s to 4.75 Pg C by 1998, with an increasing rate of 0.022 Pg C per year, due mainly to the increased reforestation and afforestation. During the last two decades, planted forests have sequestered 0.45 Pg of carbon, with an annual rate of 0.021 Pg C. The average carbon density of plantations showed a significant increase from 15.3 Mg/hm 2 in the mid-1970 to 31.1 Mg/hm 2 in 1998, suggesting that the rising global temperature and CO 2 fertilization may enhance this increasing uptake as well as expansion of planted forests.
FengJM ( 2008). Spatial patterns of species diversity of seed plants in China and their climatic explanation Biodiversity Science, 16, 470-476. [本文引用: 2]
FranzluebbersAJ, StuedemannJA, SchombergHH, WilkinsonSR ( 2000). Soil organic C and N pools under long term pasture management in the Southern Piedmont USA Soil Biology & Biochemistry, 32, 469-478. DOI:10.1016/S0038-0717(99)00176-5URL [本文引用: 1] Soil organic matter pools under contrasting long-term management systems provide insight into potentials for sequestering soil C, sustaining soil fertility and functioning of the soil–atmospheric interface. We compared soil C and N pools (total, particulate and microbial) under pastures (1) varying due to harvest technique (grazing or haying), species composition (cool- or warm-season), stand age and previous land use and (2) in comparison with other land uses. Grazed tall fescue-common bermudagrass pasture (20 yr old) had greater soil organic C (31%), total N (34%), particulate organic C (66%), particulate organic N (2.4 fold) and soil microbial biomass C (28%) at a depth of 0–200 mm than adjacent land in conservation-tillage cropland (24 yr old). Soil organic C and total N at a depth of 0–200 mm averaged 3800 and 294 g m 612 , respectively, under grazed bermudagrass and 3112 and 219 g m 612 , respectively, under hayed bermudagrass. A chronosequence of grazed tall fescue suggested soil organic N sequestration rates of 7.3, 4.4 and 0.6 g m 612 yr 611 to a depth of 200 mm during 0–10, 10–30 and 30–50 yr, respectively. Soil C storage under long-term grazed tall fescue was 85 to 88% of that under forest, whereas soil N storage was 77 to 90% greater under grazed tall fescue than under forest. Properly grazed pastures in the Southern Piedmont USA have great potential to restore natural soil fertility, sequester soil organic C and N and increase soil biological activity.
GuangGY ( 2015). The Study of Community Diversity and Soil Active Organic Carbon Pool Characteristics under Different Utilizations in Alpine Grassland. XingJiang Agricultural University ürmüqi. URL [本文引用: 1] 草地土壤有机碳库是陆地生态系统最大的碳库之一,草地利用方式对物种多样性及土壤中有机碳及其活性组分有重要影响。本研究以巴音布鲁克高寒草地不同封育时间(长期封育LF、短期封育SF)及不同放牧方式的草地(中度放牧MG、农作+放牧A+G)作为研究对象,对四种利用方式草地群落多样性特征、土壤有机碳及其活性组分(颗粒碳、易氧化碳、微生物碳)及其分配比例和剖面分布特征进行分析,并以碳库管理指数作为评价草地土壤质量变化的量化指标,得出以下结论:1.该区域草地群落多样性特征(多样性指数、均匀度指数、优势度指数)对利用方式具有不同响应,三种指数整体表现为:MGA+GLFSF。中度放牧提高了草地物种多样性和物种优势度;封育后多样性指数和优势度指数均下降。四种利用方式对草地均匀度没有显著影响。群落多样性特征与0~10 cm土层的易氧化碳、全氮呈显著正相关;与10~30 cm的多数土壤养分呈显著负相关。2.该区域土壤有机碳对利用方式具有不同响应,大体表现为:SFLFMGA+G,短期封育对有机碳的积累效果显著;农作放牧方式下有机碳损失显著。3.该区域不同利用方式土壤颗粒有机碳及其分配比例与有机碳的变化特征相一致,表现为:SFLFMGA+G。表明:短期封育有助于颗粒有机碳的积累,提高土壤碳库活性;放牧及农作放牧方式降低了土壤有机碳活性。不同利用方式颗粒碳及其分配比例及颗粒碳含量土层间变化幅度均大于有机碳的变化,表明其对外界变化反应更敏感。4.该区域土壤易氧化碳对不同利用方式同样具有不同响应,表现为:SFLFA+GMG,封育可以有效提高土壤易氧化碳含量,尤其是15~50 cm土层,效果显著。而易氧化碳分配比例则表现为:A+GLFSFMG,与易氧化碳变化趋势存在差异。各利用方式易氧化碳及其分配比例变化幅度均大于有机碳的变化,表明其可以作为指示不同利用方式对草地碳库稳定性影响的敏感性指标。5.该区域土壤微生物碳及微生物商对不同利用方式的响应表现为:LFMGSFA+G,与有机碳及其它两种活性组分和分配比例的变化特征有很大差异。表明其对碳库稳定性的表征除受自身活性的影响外,还受到其他环境因子的制约。6.该区域不同利用方式土壤碳库管理指数(CPMI)表现为:SF(112.30)A+G(100.00)LF(90.65)MG(87.44)。表明短期封育对草地土壤质量恢复有积极意义,并且易氧化碳含量的增加对碳库管理指数的提高效果更显著。碳库管理指数与草地群落多样性特征在0-30 cm土层存在显著相关性,表明其可以作为指示草地群落特征的指标。 管光玉 ( 2015). 不同利用方式高寒草地群落多样性及土壤活性有机碳库特征研究 新疆农业大学, 乌鲁木齐. URL [本文引用: 1] 草地土壤有机碳库是陆地生态系统最大的碳库之一,草地利用方式对物种多样性及土壤中有机碳及其活性组分有重要影响。本研究以巴音布鲁克高寒草地不同封育时间(长期封育LF、短期封育SF)及不同放牧方式的草地(中度放牧MG、农作+放牧A+G)作为研究对象,对四种利用方式草地群落多样性特征、土壤有机碳及其活性组分(颗粒碳、易氧化碳、微生物碳)及其分配比例和剖面分布特征进行分析,并以碳库管理指数作为评价草地土壤质量变化的量化指标,得出以下结论:1.该区域草地群落多样性特征(多样性指数、均匀度指数、优势度指数)对利用方式具有不同响应,三种指数整体表现为:MGA+GLFSF。中度放牧提高了草地物种多样性和物种优势度;封育后多样性指数和优势度指数均下降。四种利用方式对草地均匀度没有显著影响。群落多样性特征与0~10 cm土层的易氧化碳、全氮呈显著正相关;与10~30 cm的多数土壤养分呈显著负相关。2.该区域土壤有机碳对利用方式具有不同响应,大体表现为:SFLFMGA+G,短期封育对有机碳的积累效果显著;农作放牧方式下有机碳损失显著。3.该区域不同利用方式土壤颗粒有机碳及其分配比例与有机碳的变化特征相一致,表现为:SFLFMGA+G。表明:短期封育有助于颗粒有机碳的积累,提高土壤碳库活性;放牧及农作放牧方式降低了土壤有机碳活性。不同利用方式颗粒碳及其分配比例及颗粒碳含量土层间变化幅度均大于有机碳的变化,表明其对外界变化反应更敏感。4.该区域土壤易氧化碳对不同利用方式同样具有不同响应,表现为:SFLFA+GMG,封育可以有效提高土壤易氧化碳含量,尤其是15~50 cm土层,效果显著。而易氧化碳分配比例则表现为:A+GLFSFMG,与易氧化碳变化趋势存在差异。各利用方式易氧化碳及其分配比例变化幅度均大于有机碳的变化,表明其可以作为指示不同利用方式对草地碳库稳定性影响的敏感性指标。5.该区域土壤微生物碳及微生物商对不同利用方式的响应表现为:LFMGSFA+G,与有机碳及其它两种活性组分和分配比例的变化特征有很大差异。表明其对碳库稳定性的表征除受自身活性的影响外,还受到其他环境因子的制约。6.该区域不同利用方式土壤碳库管理指数(CPMI)表现为:SF(112.30)A+G(100.00)LF(90.65)MG(87.44)。表明短期封育对草地土壤质量恢复有积极意义,并且易氧化碳含量的增加对碳库管理指数的提高效果更显著。碳库管理指数与草地群落多样性特征在0-30 cm土层存在显著相关性,表明其可以作为指示草地群落特征的指标。
HoltJA ( 1997). Grazing pressure and soil carbon, microbial biomass and enzyme activities in semiarid Northeastern Australia Applied Soil Ecology, 5, 143-149. DOI:10.1016/S0929-1393(96)00145-XURL [本文引用: 1] ABSTRACT The effects of cattle grazing pressure on total carbon, microbial biomass carbon levels and selected enzyme activities of the soils at several sites in northeastern Australia have been examined. No changes in organic carbon levels were detected, but a significant reduction in soil microbial biomass carbon levels (approximately 24% and 51%) occurred at two tropical semi-arid woodland sites after 6 and 8 years of an experimentally imposed heavy grazing strategy. Peptidase and amidase activities were also significantly reduced in the soils of the heavily grazed treatments at one of these sites. A comparison of soils in areas where poor grazing management over a period of at least 10 years had given rise to pastures in poor condition with similar soils from nearby well managed areas supporting pasture in good condition was also made. Both total carbon and microbial biomass carbon were lower in the soils of the area that had been subjected to poor grazing management. Under poor management regimes, soil microbial biomass may provide an earlier indication of changes in organic matter status than total carbon.
HuZL, PanGX, LiLQ, DuYX, WangXZ ( 2009). Changes in pool sand heterogeneity of soil organic carbon, nitrogen and phosphorus under different vegetation types in karst mountainous area of central Guizhou Province, China Acta Ecologica Sinica, 29, 4187-4193. DOI:10.3321/j.issn:1000-0933.2009.08.021URL [本文引用: 1] 西南喀斯特地区是中国四大生态脆弱地区之一,为了解喀斯特生态环境下植被演替对土壤养分循环的影响,选取贵州中部喀斯特地区由乔木林、灌木林和灌草丛等不同植被类型下的土壤为研究对象,采用样块法采集了表层土壤样品,测定C、N、P全量及有效态含量,分析了这些元素的空间异质性特点。结果表明,各养分元素无论全量和有效态含量均在乔木林下最高,灌木林下有机碳及N、P全量的下降并不显著,但养分有效态含量显著下降,而灌草丛下土壤养分无论是全量还是有效态含量均较灌木林下显著降低下降;土壤养分空间异质性在灌木林下最高,土壤全磷素特别是速效磷的空间异质性高于有机碳和N。这显示,不仅是土壤养分含量,而且土壤养分的空间异质性都随植被演替而改变。这种变化中,土壤养分有效态较全量更为剧烈。植物类型和结构变化下凋落物返还及土壤生物化学转化环节的变化可能是引起喀斯特生态系统退化下土壤养分库降低而空间异质化提高的主要原因,这最终也会影响土壤养分在生态系统内的循环和分布。 胡忠良, 潘根兴, 李恋卿, 杜有新, 王新洲 ( 2009). 贵州喀斯特山区不同植被下土壤C、N、P含量和空间异质性 生态学报, 29, 4187-4193. DOI:10.3321/j.issn:1000-0933.2009.08.021URL [本文引用: 1] 西南喀斯特地区是中国四大生态脆弱地区之一,为了解喀斯特生态环境下植被演替对土壤养分循环的影响,选取贵州中部喀斯特地区由乔木林、灌木林和灌草丛等不同植被类型下的土壤为研究对象,采用样块法采集了表层土壤样品,测定C、N、P全量及有效态含量,分析了这些元素的空间异质性特点。结果表明,各养分元素无论全量和有效态含量均在乔木林下最高,灌木林下有机碳及N、P全量的下降并不显著,但养分有效态含量显著下降,而灌草丛下土壤养分无论是全量还是有效态含量均较灌木林下显著降低下降;土壤养分空间异质性在灌木林下最高,土壤全磷素特别是速效磷的空间异质性高于有机碳和N。这显示,不仅是土壤养分含量,而且土壤养分的空间异质性都随植被演替而改变。这种变化中,土壤养分有效态较全量更为剧烈。植物类型和结构变化下凋落物返还及土壤生物化学转化环节的变化可能是引起喀斯特生态系统退化下土壤养分库降低而空间异质化提高的主要原因,这最终也会影响土壤养分在生态系统内的循环和分布。
JenyH ( 1980). The Soil Resource: Origin and Behavior. Springer-Verlag New York.377. [本文引用: 1]
LiL, ZhouXY, HuangZL, WeiSG, ShiJH ( 2006). Study on the relationship between α diversity of plant community and environment on Dinghushan Acta Ecologica Sinica, 26, 2301-2307. DOI:10.3321/j.issn:1000-0933.2006.07.033URL [本文引用: 1] 用样带取样法,来研究不同取样尺度和不同取样尺度条件下的多个环境因子与鼎湖山植物群落α多样性的关系.取样尺度分别为10、20、40、80m和160m.涵盖鼎湖山主要的植被类型:季风常绿阔叶林、针阔混交林、沟谷常绿阔叶林和针叶林.相关分析和主成分分析结果表明,环境因子对各层次的α多样性的影响程度各不相同,达到显著相关性的取样尺度也不一样,表现出较大的复杂性,同时也表明样带上的环境异质性较高.因此,用海拔梯度作为主要的环境梯度来研究鼎湖山植物群落多样性具有合理性.海拔高度与乔木层多样性的关系在所有取样尺度上都较密切,这说明海拔高度可能是影响乔木层α多样性的最重要环境因子. 李林, 周小勇, 黄忠良, 魏识广, 史军辉 ( 2006). 鼎湖山植物群落α多样性与环境的关系 生态学报, 26, 2301-2307. DOI:10.3321/j.issn:1000-0933.2006.07.033URL [本文引用: 1] 用样带取样法,来研究不同取样尺度和不同取样尺度条件下的多个环境因子与鼎湖山植物群落α多样性的关系.取样尺度分别为10、20、40、80m和160m.涵盖鼎湖山主要的植被类型:季风常绿阔叶林、针阔混交林、沟谷常绿阔叶林和针叶林.相关分析和主成分分析结果表明,环境因子对各层次的α多样性的影响程度各不相同,达到显著相关性的取样尺度也不一样,表现出较大的复杂性,同时也表明样带上的环境异质性较高.因此,用海拔梯度作为主要的环境梯度来研究鼎湖山植物群落多样性具有合理性.海拔高度与乔木层多样性的关系在所有取样尺度上都较密切,这说明海拔高度可能是影响乔木层α多样性的最重要环境因子.
MenMX, PengZP, LiuYH, YuZR ( 2005). Spatial distribution of soil organic carbon and nitrogen in Hebei Province based on SOTER Chinese Journal of Soil Science, 36, 469-473. [本文引用: 1]
ParkerJL, FernandezIJ, RustadLE, NortonSA ( 2001). Effects of nitrogen enrichment, wildfire, and harvesting on forest soil carbon and nitrogen Soil Science Society of America Journal, 65, 1248-1255. DOI:10.2136/sssaj2001.6541248xURL [本文引用: 1]
PenningsSC, GrantMB, BertnessMD ( 2005). Plant zonation in low-latitude salt marshes: Disentangling the roles of flooding salinity and competition Journal of Ecology, 93, 159-167. DOI:10.1111/j.1365-2745.2004.00959.xURL [本文引用: 1] Summary 1 We investigated the factors producing zonation patterns of the dominant plants in south-eastern USA salt marshes where Juncus roemerianus dominates the high marsh, and Spartina alterniflora the middle and low marsh. 2 Juncus did not occur naturally in the Spartina zone and performed poorly when transplanted there, irrespective of whether neighbours were present or removed, indicating that its lower limit was set by physical stress. 3 In contrast, although Spartina occurred naturally at low densities in the Juncus zone, it performed well if transplanted there only if neighbours were removed, indicating that its upper limit was set by competition. 4 Parallel laboratory and field manipulations of flooding, salinity and competition indicated that the lower limit of Juncus was mediated by both flooding and salinity, but not by competition. 5 The general mechanisms producing zonation patterns of vegetation in coastal salt marshes may be universal, as suggested by previous studies, but the importance of particular factors is likely to vary geographically. In particular, salinity stress probably plays a much more important role in mediating plant zonation patterns at lower latitudes. 6 Our results suggest that the nature of ecological interactions is likely to vary geographically because of variation in the physical environment, and this variation must be taken into account in order to successfully generalize the results of field studies across geographical scales.
PostWM, KingAM, WullschlegerSD ( 1996). Soil organic matter models and global estimates of soil organic carbon. In: Powlson DS, Smith P, Smith JU eds Evaluation of Soil Organic Matter Models: Using Existing Long-Term Datasets (Nato ASI Subseries I). Springer-Verlag, Heidelberg. 38, 201-222. [本文引用: 1]
RitchieJC, McCartyGW, VenterisER ( 2007). Soil and soil organic carbon redistribution on the lands cap Geomorpholosry, 89, 163-171. DOI:10.1016/j.geomorph.2006.07.021URL [本文引用: 1] Patterns of soil organic carbon (SOC) vary widely across the landscape leading to large uncertainties in the SOC budget especially for agricultural landscapes where water, tillage and wind erosion redistributes soil and SOC across the landscape. It is often assumed that soil erosion results in a loss of SOC from the agricultural ecosystem but recent studies indicate that soil erosion and its subsequent redistribution within fields can stimulate carbon sequestration in agricultural ecosystems. This study investigates the relationship between SOC and soil redistribution patterns on agricultural landscapes. Soil redistribution (erosion and deposition) patterns were estimated in three tilled agricultural fields using the fallout 137 Cesium technique. 137 Cs and SOC concentrations of upland soils are significantly correlated in our study areas. Upland areas (eroding) have significantly less SOC than soils in deposition areas. SOC decreased as gradient slope increases and soils on concave slopes had higher SOC than soils on convex slopes. These data suggest that soil redistribution patterns and topographic patterns may be used to help understand SOC dynamics on the landscape. Different productivity and oxidation rates of SOC of eroded versus deposited soils also contribute to SOC spatial patterns. However, the strong significant relationships between soil redistribution and SOC concentrations in the upland soil suggest that they are moving along similar physical pathways in these systems. Our study also indicates that geomorphic position is important for understanding soil movement and redistribution patterns within a field or watershed. Such information can help develop or implement management systems to increase SOC in agricultural ecosystems.
SmolanderA, PrihaO, PaavolainenL, SteerJ, M?lk?nenE ( 1998). Nitrogen and carbon transformations before and after clear cutting in repeatedly N fertilized and limed forest soil Soil Biology & Biochemistry, 30, 477-490. DOI:10.1016/S0038-0717(97)00141-7URL [本文引用: 1] Nitrogen and carbon transformations were monitored in a Norway spruce ( Picea abies L.) stand in the summer before clear-cutting, and for the following three summers. During 30 y before the clear-cutting the stand had been repeatedly limed (total 6 t limestone ha 611 ), fertilized with N (total about 900 kg N ha 611 ), and both treatments were combined. Aerobic incubation experiments in the laboratory showed that, before clear-cutting, nitrification took place only in the soil that had been both limed and N-fertilized. Clear-cutting increased soil pH and net formation of mineral N, and initiated nitrification in all soils. These effects were observed throughout the study period. The only exception was the soil that had been both limed and N-fertilized, where the effect of clear-cutting on these N transformations was negligible or even suppressive. Generally, the greatest response in N transformations to clear-cutting was observed in the control soil. There was a small increase in microbial biomass C and N, and C mineralization in the first summer after clear-cutting. Net formation of mineral N correlated positively with pH at a lower pH range (pH 3.9–4.9) and negatively at a higher pH range (pH 4.9–6.9). C mineralization correlated positively with microbial biomass C, but there was no linear relationship between net formation of mineral N and microbial biomass N. C mineralization and net N mineralization were not correlated.
TangGY, LiK, SunYY, ZhangCH ( 2010). Soil labile organic carbon contents and their allocation characteristics under different land uses at dry-hot valley Environmental Science, 31, 1365-1371. URL [本文引用: 1] 对比研究了干热河谷新银合欢林地、大叶相思林地、旱耕地和荒地土壤有机碳(SOC)、易氧化有机碳(ROC)、微生物生物量碳(MBC)和可溶性有机碳(DOC)含量及其分配比例.结果表明,4类利用方式下SOC含量在4.22~5.19g·kg-1之间,其差异不显著.新银合欢(2.14g·kg-1)和大叶相思林地ROC含量(2.03g·kg-1)显著高于旱耕地(1.38g·kg-1)和荒地(1.34g·kg-1);4类利用方式下,旱耕地MBC和DOC含量均最高,荒地最低.林地ROC分配比例是荒地和旱耕地的1.3~1.6倍;旱耕地MBC和DOC的分配比例均高于其他3类利用方式,林地和荒地MBC、DOC分配比例接近.植被凋落量和管理措施是不同利用方式下ROC含量差异的主要原因,而土壤含水量和植被凋落性质是4类利用方式下MBC、DOC含量变异的主要影响因素.干热河谷ROC含量变化可以敏感地指示SOC动态,但MBC、DOC含量变化则不能反映SOC动态. 唐国勇, 李昆, 孙永玉, 张春华 ( 2010). 干热河谷不同利用方式下土壤活性有机碳含量及其分配特征 环境科学, 31, 1365-1371. URL [本文引用: 1] 对比研究了干热河谷新银合欢林地、大叶相思林地、旱耕地和荒地土壤有机碳(SOC)、易氧化有机碳(ROC)、微生物生物量碳(MBC)和可溶性有机碳(DOC)含量及其分配比例.结果表明,4类利用方式下SOC含量在4.22~5.19g·kg-1之间,其差异不显著.新银合欢(2.14g·kg-1)和大叶相思林地ROC含量(2.03g·kg-1)显著高于旱耕地(1.38g·kg-1)和荒地(1.34g·kg-1);4类利用方式下,旱耕地MBC和DOC含量均最高,荒地最低.林地ROC分配比例是荒地和旱耕地的1.3~1.6倍;旱耕地MBC和DOC的分配比例均高于其他3类利用方式,林地和荒地MBC、DOC分配比例接近.植被凋落量和管理措施是不同利用方式下ROC含量差异的主要原因,而土壤含水量和植被凋落性质是4类利用方式下MBC、DOC含量变异的主要影响因素.干热河谷ROC含量变化可以敏感地指示SOC动态,但MBC、DOC含量变化则不能反映SOC动态.
TilmanD ( 1993). Species richness of experimental productivity gradients: How important is colonization? Ecology, 74, 2179-2191. DOI:10.2307/1939572URL [本文引用: 1] The biodiversity of a site should depend on the interplay of local colonization (gain) and extinction (loss) rates, but few theoretical explanations of diversity patterns on productivity gradients have included effects of productivity on colonization. In an 11-yr study, experimental increases in productivity via nitrogen addition generally led to decreased species richness in four grassland fields. Decreased diversity in productive plots was caused as much by lower rates of species gain as by greater rates of loss of existing species. Annual grasses and forbs had high gain and loss rates, but these were independent of productivity. In contrast, the rates of gain of perennial grass and forb species declined with productivity, and their rates of loss increased. Species richness was dependent on litter mass and on light penetration, but not on aboveground living plant mass, suggesting that there was no direct effect of productivity on diversity. Diversity in periodically burned prairie, which had low litter mass, was independent of productivity by the 10th and 11th yr of the study. Results suggest that diversity is lower in productive grasslands because accumulated litter, and possibly lower light penetration, inhibit germination and/or survival of seedlings, and thus decrease rates of establishment by new species. Higher productivity also leads to higher rates of loss, presumably via competitive displacement, of existing species. Results do not support the hypothesis that soil N heterogeneity controls diversity on the temporal and spatial scales of this study.
WangCT, LongRJ, LiuW, WangQL, ZhangL, WuPF ( 2010). Relationships between soil carbon distribution and species diversity and community biomass at different alpine meadows Resources Science, 32, 2022-2099. URL [本文引用: 1] 利用2003年-2005年中国科学院海北站不同群落类型草地和 土壤的实测资料,研究了高寒草甸不同群落类型土壤有机碳、微生物量碳的分布特征与物种多样性、生物量的关系.结果显示:不同群落类型物种多样性随土壤有机 碳、微生物量碳的增加而降低;群落生物量随着土壤有机碳、微生物量碳含量的增加而增加;不同类型草地土壤中,藏嵩草沼泽化草甸不同土层土壤微生物量碳和土 壤有机全碳均显著高于其它三个类型草地各层的土壤微生物量碳、土壤有机全碳(P<0.01),4种群落类型土壤剖面上土壤有机碳和微生物量碳随土层 深度的增加而降低,同一土层、不同群落类型土壤有机碳和微生物量碳存在明显差异;相关分析表明,土壤有机碳和土壤微生物量碳与有机质、全氮呈显著相关,土 壤有机碳和土壤微生物量碳含量可作为衡量土壤肥力和土壤质量变化的重要指标.不同类型草地其最初的植物群落物种组成、生产力水平和土壤养分条件影响着土壤 微生物生物量、组成和活性. 王长庭, 龙瑞军, 刘尉, 王启兰, 张莉, 吴鹏飞 ( 2010). 高寒草甸不同群落类型土壤碳分布于物种多样性、生物量关系 资源科学, 32, 2022-2099. URL [本文引用: 1] 利用2003年-2005年中国科学院海北站不同群落类型草地和 土壤的实测资料,研究了高寒草甸不同群落类型土壤有机碳、微生物量碳的分布特征与物种多样性、生物量的关系.结果显示:不同群落类型物种多样性随土壤有机 碳、微生物量碳的增加而降低;群落生物量随着土壤有机碳、微生物量碳含量的增加而增加;不同类型草地土壤中,藏嵩草沼泽化草甸不同土层土壤微生物量碳和土 壤有机全碳均显著高于其它三个类型草地各层的土壤微生物量碳、土壤有机全碳(P<0.01),4种群落类型土壤剖面上土壤有机碳和微生物量碳随土层 深度的增加而降低,同一土层、不同群落类型土壤有机碳和微生物量碳存在明显差异;相关分析表明,土壤有机碳和土壤微生物量碳与有机质、全氮呈显著相关,土 壤有机碳和土壤微生物量碳含量可作为衡量土壤肥力和土壤质量变化的重要指标.不同类型草地其最初的植物群落物种组成、生产力水平和土壤养分条件影响着土壤 微生物生物量、组成和活性.
WangDD, ShiXZ, LuXX, WangHJ, YuDS, SunWX, ZhaoYC ( 2010). Response of soil organic carbon spatial variability to the expansion of scale in the uplands of Northeast China Geoderma, 154, 302-310. DOI:10.1016/j.geoderma.2009.10.018URL Soil organic carbon (SOC) plays an important role in maintaining and improving soil fertility and quality as well as mitigating climate change. Understanding SOC density spatial variability is fundamental for describing soil resources and predicting SOC. Three categories were used to create spatial scales: administrative category (county, city, province and region scale), soil taxonomic category (family, sub-group, great group and order scale) and soil type (zonal soil and azonal soil)-administrative category. Soil organic carbon density variability and its response to the expansion of scales in the topsoil (0 20 cm) and soil profile (a depth of 1 m) layers in the uplands of Northeast China were examined based on coefficient of variation (CV) values using data of 1041 profiles obtained from the Second National Soil Survey of China. The results depicted that SOC density variability increased not only in the topsoil layer but also in the soil profile layer with the expansion of scales in all categories. In the administrative category, there was a strong logarithmic relationship between upland areas or administrative areas and mean SOC density CV. Though mean SOC density CV within each soil order increased from family to order, the trend and range of increase varied greatly. Soil organic carbon density variability for zonal and azonal soils was similar in terms of trends but different in terms of rate with increasing scale from county to region. A strong logarithmic relationship between upland area and mean SOC density CV was also observed. These relationships indicated that reducing upland area by five orders of magnitude would halve the CV. Therefore, when estimating the SOC pool in uplands, both administrative and soil type scales should be considered in the sampling design, especially for azonal soil.
WangSX, WangXA, LiGQ, GuoH, ZhuZH ( 2010). Species diversity and environmental interpretation in the process of community succession in the Ziwu Mountain of Shaanxi Province Acta Ecologica Sinica, 30, 1638-1647. DOI:10.3969/j.issn.1674-697X.2014.08.318URL [本文引用: 1] 采用空间代替时间的方法研究了黄土高原子午岭植物群落自然演替过程中不同层α物种多样性的变化规律,并通过CCA分析了物种多样性与环境间的关系。结果表明:(1)随演替的进展,草本层物种丰富度、均匀度及物种多样性均逐渐降低(P〈0.05);灌木层物种的丰富度与物种多样性均呈现相似的"J"形变化趋势,而均匀度(虎榛子(Ostryopsis davidiana)群落除外)差异不显著(P〉0.05);乔木层物种丰富度、均匀度以及物种多样性差异均不显著(P〉0.05);群落总体物种丰富度随演替进展呈现明显的单峰模型,与中期物种丰富度假说一致。(2)林冠郁闭度是影响草本层和灌木层物种组成的主要环境因子(P〈0.01),其次为土壤养分和坡位;而坡位对乔木层物种组成的影响极显著(P〈0.01),这可能与乔木树种的更新差异有关。(3)群落演替是物种扩散和环境筛综合作用的过程,物种对演替过程中变化光照资源的响应差异可能是该区不同层物种多样性变化的主要原因。 王世雄, 王孝安, 李国庆, 郭华, 朱志红 ( 2010). 陕西子午岭植物群落演替过程中物种多样性变化与环境解释 生态学报, 30, 1638-1647. DOI:10.3969/j.issn.1674-697X.2014.08.318URL [本文引用: 1] 采用空间代替时间的方法研究了黄土高原子午岭植物群落自然演替过程中不同层α物种多样性的变化规律,并通过CCA分析了物种多样性与环境间的关系。结果表明:(1)随演替的进展,草本层物种丰富度、均匀度及物种多样性均逐渐降低(P〈0.05);灌木层物种的丰富度与物种多样性均呈现相似的"J"形变化趋势,而均匀度(虎榛子(Ostryopsis davidiana)群落除外)差异不显著(P〉0.05);乔木层物种丰富度、均匀度以及物种多样性差异均不显著(P〉0.05);群落总体物种丰富度随演替进展呈现明显的单峰模型,与中期物种丰富度假说一致。(2)林冠郁闭度是影响草本层和灌木层物种组成的主要环境因子(P〈0.01),其次为土壤养分和坡位;而坡位对乔木层物种组成的影响极显著(P〈0.01),这可能与乔木树种的更新差异有关。(3)群落演替是物种扩散和环境筛综合作用的过程,物种对演替过程中变化光照资源的响应差异可能是该区不同层物种多样性变化的主要原因。
ZhangHZ, ZhuQK, WangJ, KuangGM, XieJ, ZhaoWJ ( 2011). Soil physical properties of micro-topography on loess slope in North Shaanxi Province Bulletin of Soil and Water Conservation, 31(6), 55-58. [本文引用: 1]
ZhangL, WangCY ( 2014). Relationship between species diversity and environmental factors of plant community at different succession stages in ecotone of Great Xing’an and Xiaoxing’an Mountains Forest Engineering, 30(5), 1-5. DOI:10.3969/j.issn.1001-005X.2014.05.001URL [本文引用: 1] 为揭示阔叶红松林次生演替阶段群落多样性状况,研究生物多样性指数与环境因子的关系,为该区植被保护、植被恢复提供理论支持。运用物多样性指数、典范对应分析和次生演替的空间序列代替时间序列方法,以大小兴安岭过渡区阔叶红松林不同演替阶段群落为研究对象,设置33个网格样地数据,记录每个样地的物种多度、盖度、频度及土壤环境数据,测定12个环境指标。结果表明:1原始阔叶红松林群落经过顺行演替向逆行演替发展,演替趋势为:原始阔叶红松林→天然次生阔叶林→天然次生针叶林→次生裸地;2保存较好的原始阔叶红松林物种多样性指数最高,物种最丰富,其次是天然次生阔叶林,物种多样性指数最低的是次生裸地,群落结构简单。3物种多样性指数与纬度、土壤含水率、土壤全氮含量相关性最大。 张玲, 王承义 ( 2014). 大小兴安岭过渡区阔叶红松林次生演替阶段群落多样性指数与环境因子关系 森林工程, 30(5), 1-5. DOI:10.3969/j.issn.1001-005X.2014.05.001URL [本文引用: 1] 为揭示阔叶红松林次生演替阶段群落多样性状况,研究生物多样性指数与环境因子的关系,为该区植被保护、植被恢复提供理论支持。运用物多样性指数、典范对应分析和次生演替的空间序列代替时间序列方法,以大小兴安岭过渡区阔叶红松林不同演替阶段群落为研究对象,设置33个网格样地数据,记录每个样地的物种多度、盖度、频度及土壤环境数据,测定12个环境指标。结果表明:1原始阔叶红松林群落经过顺行演替向逆行演替发展,演替趋势为:原始阔叶红松林→天然次生阔叶林→天然次生针叶林→次生裸地;2保存较好的原始阔叶红松林物种多样性指数最高,物种最丰富,其次是天然次生阔叶林,物种多样性指数最低的是次生裸地,群落结构简单。3物种多样性指数与纬度、土壤含水率、土壤全氮含量相关性最大。
ZhangWJ, ZengLJ, WangWQ, ZengCS ( 2011). Soil carbon stock of alligator weed in Min River Estuarine wetland Wetland Science and Management, 7(1), 52-56. DOI:10.3969/j.issn.1673-3290.2011.01.13URL [本文引用: 1] 以闽江河口区最大的洲滩湿地——鳝鱼滩湿地为研究区域,选取空心莲子草湿地土壤为研究对象,对研究区内18个土壤剖面以10cm为间隔分层取样,对其土壤有机碳含量、储量和垂直变化特征及其影响因子进行分析。研究结果表明,空心莲子草湿地土壤有机碳含量和储量最大值均出现在0~10cm土壤剖面,含量和储量分别为32.77g/kg和2817.96t/km2,并与其它土壤剖面的含量和储量存在显著差异,0~60cm土层平均有机碳含量为9.70g/kg,平均有机碳储量为1002.86t/km2,且各个土壤理化因子在表层0~10cm的含量均与其它土层存在显著差异。空心莲子草湿地土壤有机碳含量与影响因子相关性较显著,其中,土壤含水量和土壤盐度与土壤有机碳含量呈极显著正相关(r=0.990,p=0.000〈0.01,n=6;r=0.922,p=0.004〈0.01,n=6),土壤容重与土壤有机碳含量呈极显著负相关(r=-0.982,p=0.000〈0.01,n=6),土壤灰分和土壤pH值与土壤有机碳含量呈显著负相关(r=-0.857,p=0.015〈0.05,n=6;r=-0.838,p=0.019〈0.05,n=6)。 张文娟, 曾路金, 王维奇, 曾从盛 ( 2011). 闽江河口空心莲子草土壤碳库研究 湿地科学与管理, 7(1), 52-56. DOI:10.3969/j.issn.1673-3290.2011.01.13URL [本文引用: 1] 以闽江河口区最大的洲滩湿地——鳝鱼滩湿地为研究区域,选取空心莲子草湿地土壤为研究对象,对研究区内18个土壤剖面以10cm为间隔分层取样,对其土壤有机碳含量、储量和垂直变化特征及其影响因子进行分析。研究结果表明,空心莲子草湿地土壤有机碳含量和储量最大值均出现在0~10cm土壤剖面,含量和储量分别为32.77g/kg和2817.96t/km2,并与其它土壤剖面的含量和储量存在显著差异,0~60cm土层平均有机碳含量为9.70g/kg,平均有机碳储量为1002.86t/km2,且各个土壤理化因子在表层0~10cm的含量均与其它土层存在显著差异。空心莲子草湿地土壤有机碳含量与影响因子相关性较显著,其中,土壤含水量和土壤盐度与土壤有机碳含量呈极显著正相关(r=0.990,p=0.000〈0.01,n=6;r=0.922,p=0.004〈0.01,n=6),土壤容重与土壤有机碳含量呈极显著负相关(r=-0.982,p=0.000〈0.01,n=6),土壤灰分和土壤pH值与土壤有机碳含量呈显著负相关(r=-0.857,p=0.015〈0.05,n=6;r=-0.838,p=0.019〈0.05,n=6)。
ZhangY, ShiXZ, YuDS, LiuXM ( 2009). Factors affecting variation of soil organic carbon density in Yunnan-?Guizhou-Guangxi Region Acta Pedologica Sinica, 46, 526-531. DOI:10.3321/j.issn:0564-3929.2009.03.022URL [本文引用: 1] 正土壤有机碳是土壤肥力的重要指标之一,也是陆地碳库的主要部分。揭示影响土壤有机碳储量的主要因素,对合理地利用土壤资源,减少土壤温室气体的排放,增加土壤碳存储等方面都具有重要的意义。土壤有机碳储量受气候、植被、土壤属 张勇, 史学正, 于东升, 刘秀明 ( 2009). 滇黔桂地区土壤有机碳密度变异的影响因素研究 土壤学报, 46, 526-531. DOI:10.3321/j.issn:0564-3929.2009.03.022URL [本文引用: 1] 正土壤有机碳是土壤肥力的重要指标之一,也是陆地碳库的主要部分。揭示影响土壤有机碳储量的主要因素,对合理地利用土壤资源,减少土壤温室气体的排放,增加土壤碳存储等方面都具有重要的意义。土壤有机碳储量受气候、植被、土壤属
ZhouL, LiBG, ZhouGS ( 2005). Advances in controlling factors of soil organic carbon Advances in Earth Science, 20, 99-105. DOI:10.3321/j.issn:1001-8166.2005.01.016URL [本文引用: 1] 土壤有机碳库是全球碳循环的重要组成部分,其积累和分解的变化直接影响全球的碳平衡。理解土壤有机碳蓄积过程对生物、物理和人为因素的响应,把握关键的控制因子是准确预测土壤有机碳在全球变化情景下对大气CO2的源/汇方向及准确评估碳收支的关键。综述了土壤有机碳主导影响因子的研究进展,并针对陆地碳循环特点,提出未来土壤有机碳研究应加强土壤有机碳过程与状态的定量化、土壤有机碳分解对环境因子的敏感性、氮沉降对土壤有机碳的影响、土壤有机碳对气候变率的响应及其反馈作用,以及土壤有机碳动态的综合模拟5个方面的研究,为准确评估陆地碳收支提供依据。 周莉, 李保国, 周广胜 ( 2005). 土壤有机碳的主导影响因子及其研究进展 地球科学进展, 20, 99-105. DOI:10.3321/j.issn:1001-8166.2005.01.016URL [本文引用: 1] 土壤有机碳库是全球碳循环的重要组成部分,其积累和分解的变化直接影响全球的碳平衡。理解土壤有机碳蓄积过程对生物、物理和人为因素的响应,把握关键的控制因子是准确预测土壤有机碳在全球变化情景下对大气CO2的源/汇方向及准确评估碳收支的关键。综述了土壤有机碳主导影响因子的研究进展,并针对陆地碳循环特点,提出未来土壤有机碳研究应加强土壤有机碳过程与状态的定量化、土壤有机碳分解对环境因子的敏感性、氮沉降对土壤有机碳的影响、土壤有机碳对气候变率的响应及其反馈作用,以及土壤有机碳动态的综合模拟5个方面的研究,为准确评估陆地碳收支提供依据。 Climate organic matter and clay content relationship in the Pampa and Chaco, Argentina 2 1998
 ,1,2, 薛晶月2,3, 谢雨2, 吴彦
,1,2, 薛晶月2,3, 谢雨2, 吴彦 ,2,*
,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}