 ,1,2,*
,1,2,*Spatial patterns and drivers of root turnover in grassland ecosystems in China
SUN Yuan-Feng1,2, WAN Hong-Wei1, ZHAO Yu-Jin1, CHEN Shi-Ping1,2, BAI Yong-Fei,1,2,*通讯作者:
编委: 王德利
责任编辑: 李敏
| 基金资助: |
Online:2018-03-20
| Supported by: | SupportedbytheNationalKeypointResearchandInventionProgram( |

摘要
关键词:
Abstract
Keywords:
PDF (1755KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
孙元丰, 万宏伟, 赵玉金, 陈世苹, 白永飞. 中国草地生态系统根系周转的空间格局和驱动因子. 植物生态学报, 2018, 42(3): 337-348 doi:10.17521/cjpe.2017.0241
SUN Yuan-Feng, WAN Hong-Wei, ZHAO Yu-Jin, CHEN Shi-Ping, BAI Yong-Fei.
根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介。根系周转是指根系从产生到死亡并分解消失的过程。根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (Dahlman & Kucera, 1965), 周转速率越大表明周转越快, 植物利用营养元素的能力就越强(宇万太和于永强, 2001)。通过根系周转, 碳从植被流入土壤。研究表明, 通过该过程进入土壤的碳占土壤总输入碳量的14%-50% (Vogt et al., 1986), 因此根系周转是陆地生态系统土壤碳汇的重要途径。同时, 根系周转对全球变化关键驱动因子, 如气候变暖、降水格局改变、CO2浓度升高、土地覆盖变化等具有很强的敏感性(Gill & Jackson, 2000)。因此, 研究根系周转格局及其影响因子是认识生态系统碳分配和养分循环的关键(Jackson et al., 1997; Clark et al., 2001; Trumbore & Gaudinski, 2003), 也是正确理解陆地生态系统如何响应全球变化的基础(Tom et al., 1997; 贺金生等, 2004; 王娓等, 2008)。
草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(White et al., 2000), 在全球碳循环中发挥着重要作用。对草地生态系统而言, 地下生物量占很大比重(贺金生等, 2004)。对内蒙古羊草(Leymus chinensis)草原和大针茅(Stipa grandis)草原的生物量研究表明, 地下生物量分别占植物全部生物量的81%和73% (姜恕等, 1985)。因此, 研究根系周转对深入理解草原生态系统碳库动态和固碳速率与潜力具有重要意义(柴曦等, 2014)。
对于草地生态系统, 周转的概念最早是由Odum (1960)在研究弃耕地生态系统的演替时提出的。Odum (1960)对研究区的植物总生产力进行了系统分析, 而未对植物的地上部分和根系加以区分, 也没有提出周转速率的计算方法。直到1965年, 美国密苏里大学的Dahlman和Kucera (1965)在研究草地根系周转时, 提出了后来被广泛应用的根系周转速率计算方法, 即根系周转速率=净地下初级生产力(BNPP)/地下生物量最大值(BGBmax)。在此后的几十年间, Shaver和Billings (1975)对阿拉斯加湿润苔原的根系周转进行了测定。Gupta和Singh (1982)研究了物种组成对热带草原根系周转的影响。Milchunas等(1985)首次使用14C稀释技术测定了格兰马草(Bouteloua gracilis)的根系周转速率。Hayes和Seastedt (1987)研究了水分对北美高草草原的根系周转的影响。Fitter等(1997)研究了CO2浓度升高对根系周转的影响。然而, 由于植物地下部分取样难度大等原因, 关于根系周转的研究进展缓慢(Milchunas, 2009), 且多集中于局地尺度, 对根系周转在区域甚至全球尺度上的格局没有明确的认识。直到2000年, Gill和Jackson (2000)收集了在此之前发表的文献中的根系周转数据, 首次从全球尺度上分析了主要生态系统类型(森林、草地、灌丛、湿地)根系周转的格局和主要驱动因子。其中草地生态系统包含79组数据, 研究表明全球草地生态系统根系周转速率随年平均气温(MAT)呈指数增加, 而与平均年降水量(MAP)没有显著的相关关系。在局地尺度上, 通过对北美中部大平原14年的数据分析发现, 根系周转速率与生长季降水/最热月气温的比值呈显著正相关关系, 而与MAT及MAP均没有显著的相关关系(Gill & Jackson, 2000)。Gill和Jackson (2000)首次在全球尺度上整合已有的研究结果, 揭示根系周转的格局及主要驱动因子, 为之后的研究提供了重要思路。然而该研究也存在以下几方面的不足。首先, 在样点的分布上, 草地生态系统的样点主要分布在北美、非洲和印度, 而全球最大的草地分布区欧亚大陆草原的样点却非常稀少, 尤其是中国草地取样点数目为零。欧亚大陆草原独特的地理位置和气候条件决定了其异于北美和非洲草原的特性, 因而该区域数据的缺失必将严重影响全球数据的代表性和结果的可靠性。其次, Gill和Jackson (2000)收集的数据库中, 根系周转速率的计算除了传统的生物量法外, 还包含了微根管法、同位素法及模型模拟的方法。不同方法所依据的原理不同, 误差来源各异(Milchunas, 2009)。同一地点同一年份的数据, 因为选用的方法不同, 计算结果可能产生数量级的差异(Aber et al., 1985)。所以方法上的不统一, 也使得Gill和Jackson (2000)的研究结果存在很大的不确定性。另外, 在该研究中, 受数据提取途径的局限, 根系周转驱动因子只包含了气候因子, 而对于根系生长的直接载体——土壤的理化性质并未考虑, 所以土壤数据的缺失是Gill和Jackson (2000)的研究中另一个不足之处, 也是我们着重改善的方向。
近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多。如Schippers和Olff (2000)研究了温室盆栽条件下, 3种不同养分水平对6种草本植物根系周转的影响, 发现根系周转对养分水平的响应不明显。Allard等(2005)对新西兰北部草地进行连续4年的CO2气体施肥实验(free-air CO2 enrichment), 结果表明, CO2浓度升高可以加快根系周转。Volder等(2007)对澳大利亚水虉草(Phalaris aquatica)草地进行CO2倍增、增温及刈割处理, 发现CO2浓度升高对根系周转没有明显的影响, 而白天增温和刈割可以显著提高根系周转速率。Larreguy等(2012)对阿根廷Patagonian山地草原的研究发现, 放牧可以通过影响土壤水分加快根系周转。Leifeld等(2015)利用放射性同位素(14C)法对欧洲山地草原25个样地的研究表明, 根系周转随年平均气温的增加呈指数增长, 与Gill和Jackson (2000)的研究结果一致。Peng等(2017)对全球氮添加实验进行meta分析, 表明氮添加降低了草地生态系统根系周转速率。
关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(Dahlman & Kucera, 1965)。地下净初级生产力和地下生物量最大值主要通过对生长季群落地下生物量的月动态进行测定获取, 取样方法主要采用土柱法。生长季群落地下生物量最大值即为地下生物量最大值, 群落地下生物量最大值和最小值之差即为地下净初级生产力。基于上述方法, 陈佐忠和黄徳华(1988)对内蒙古锡林河流域羊草草原与大针茅草原地下部分生产力和根系周转速率进行了测定, 发现不同年份周转速率变化很大, 主要与降水有关。冯雨峰(1990)对内蒙古灌丛化石生针茅荒漠草原地下生产力和周转速率进行研究, 结果表明群落地下部分周转率在不同层次间存在很大差异, 水分条件是影响周转率的关键因素。李英年(1998)研究了海北高寒草甸生态系统的根系周转气候的关系, 发现在降水和气温较高的年份植物地下生物量周转速率也较高。除了直接针对根系周转速率的研究外, 一些关于地下生物量及地下净初级生产力的研究也为根系周转速率的计算提供了数据支持。Ni (2004)整合了2000年以前发表的有关中国北方温性草地生物量或净初级生产力的文献, 从中提取到72组包含地下生物量或地下净初级生产力的数据, 发现群落净地下初级生产力与夏季(5-9月)平均气温呈显著的负相关关系。该项工作为进一步研究根系周转提供了基础, 同时Ni (2004)也指出, 2000年以前中国****对于草地的研究成果大多发表在中文期刊上, 西方****由于语言上的障碍, 无法将这部分优秀成果整合到全球尺度的研究中去。进入2000年以后, 关于中国草地生态系统根系周转的研究逐渐增多, 方法也呈现多样化, 且逐渐由自然条件下的取样监测转向野外控制实验研究。王长庭等(2003)对高寒矮生嵩草(Kobresia humilis)草甸净初级生产力沿降雨及海拔梯度的分布规律进行了一系列研究。Bai等(2008)利用微根管法研究了氮添加对内蒙古典型草原羊草群落根系寿命的影响, 结果表明, 氮添加可以明显缩短夏季出生的羊草根系寿命。Bai等(2010)进一步研究了增温和增雨对内蒙古温性草地根系周转速率的影响, 发现增温对根系周转没有明显影响, 而增雨可以显著提高根系周转速率。吴伊波等(2014)探讨了不同取样方法(根钻法、内生长袋法及微根管法)对根系周转速率测定结果的影响, 发现不同方法测定结果间存在较大差异。应当指出, 随着不同草地类型相关研究的进展, 整合已有的结果, 进行根系周转速率的大尺度空间格局和驱动因子的分析, 以期获得更宏观的认识显得尤为迫切。
中国是世界草地资源大国, 拥有草地总面积近4亿hm2, 草地面积约占国土总面积的41.7% (中华人民共和国农业部畜牧兽医司, 全国畜牧兽医总站, 1996)。因此, 开展中国草地根系周转的整合分析, 对于揭示我国草地生态系统的碳循环特征, 准确评估草地的固碳速率和潜力具有十分重要的理论和现实意义。为此, 本文以中国草地生态系统为研究对象, 通过对已发表的研究论文和专著中有关根系周转的数据收集、整理和系统分析, 结合实测数据, 研究根系周转的空间格局及其与气候因子和土壤属性之间的关系。拟回答以下两方面的科学问题: 1)中国草地根系周转呈现出什么样的空间分布格局?该格局与全球尺度的草地根系周转格局有何异同?2)影响中国草地根系周转的主要气候因子和土壤属性有哪些?影响机制是什么?各个因子的相对贡献有多大?
1 材料和方法
1.1 数据筛选标准
为了避免不同测定方法的差异带来的影响, 本研究统一使用生物量法来计算根系周转速率, 对使用微根管法、同位素法或模型模拟方法获得的数据予以剔除。具体计算公式为: 根系周转速率=地下净初级生产力(BNPP)/地下生物量最大值(BGBmax), 单位为a-1 (Dahlman & Kucera, 1965)。在数据收集过程中, 如果文献直接提供根系周转速率, 且计算方法与前述方法一致, 则予以采用; 反之, 如果文献中没有直接给出根系周转速率, 则需要包含完整的BNPP及BGBmax数据, 或者根据BGB的季节动态从中提取出BGBmax进而代入公式计算。另外, 同一地点不同年份的数据视作独立的数据参与计算。根系取样方法为土柱法、壕沟法、根钻法或内生袋法等传统取样方法, BNPP的计算方法为极差法、矩阵法或积分法(吴伊波等, 2014)。1.2 数据获取途径及收集结果
根系周转数据主要来源于已发表的期刊论文、专著等文献提取数据和实测数据。前者主要通过ISI Web of Science (http://www.isiknowledge.com/)和中国知网数据库(http://www.cnki.net/), 输入“草地(grassland)”、“地下净初级生产力(BNPP)”及“根系周转(root turnover)”为关键词进行检索, 基于前述筛选标准, 从1985-2016年间发表的45篇文献(附录I)中获得107组符合条件的数据。若文献中的数据以图表的形式呈现, 则通过Engauge_?Digitizer_4.1 (http://engauge-?digitizer.soft?112.com/)图形数字化软件提取。具体的数据提取过程如下: 1)导入目标曲线图, 或者直接拖动曲线图将其拷贝到工作区界面中; 2)定坐标轴, 分别在坐标轴原点、x轴最大值和y轴最大值处点击, 在弹出的对话框中输入该点的坐标值; 3)在目标曲线上逐个描点; 4)把描出的点数据导出并保存。实测部分来源于中国科学院战略性先导科技专项第三课题“草地生态系统固碳现状、速率、潜力与机制” 2011-2013年对中国主要草地分布区的调查数据, 地下生物量和地下净初级生产力分别采用根钻法和内生长芯法测定, 该部分共47组数据。气候因子(MAT、MAP)数据如果文献中未提供, 则基于所在样地的经纬度信息从世界气象数据库(www.worldclim.org)中提取, 精度为30角秒(arc-seconds)。土壤理化性质数据全部来自于表层土壤(0-30 cm), 包含土壤质地、排水等级、砾石含量、黏粒含量、砂粒含量、有机碳(SOC)含量、容重(BD)、酸碱度(pH值)、阳离子交换量(CEC)。土壤数据全部基于样地经纬度从联合国粮食及农业组织(FAO)的归一化土壤数据库(Harmonized World Soil Database Version1.1) (http://www.iiasa.ac.At/Research/LUC/luc?07/External-World-soil-database/HTML/index.html? sb=1)中提取。
1.3 数据分析
所有数据分析均在R语言软件中进行(R version 3.3.3 by R Development Core Team 2017)。数据分布的正态性检验采用Kolmogorov-Smirnov检验, 符合正态分布(D > 0.05, p > 0.05), 无需对数转换。不同草地类型、土壤质地及排水等级下根系周转速率的差异采用非平衡单因素方差分析和最小显著差异(LSD)多重比较方法。根系周转速率与气候因子及土壤理化性质的相关性分析采用Pearson相关分析方法, 通过R语言中psych包中的corr.test函数实现。气候因子与根系周转速率之间的关系采用一元线性回归分析, 气候因子和土壤理化性质对根系周转速率的综合影响采用全子集回归法筛选最佳多元回归模型, 通过leaps包中的regsubsets函数实现。因子间的多重共线性采用方差膨胀因子(vif)进行检验, 各因子的相对贡献率用R语言软件提供的自定义函数relweights计算。图形绘制通过SigmaPlot 11.0完成。2 结果和分析
2.1 根系周转在空间上的分布
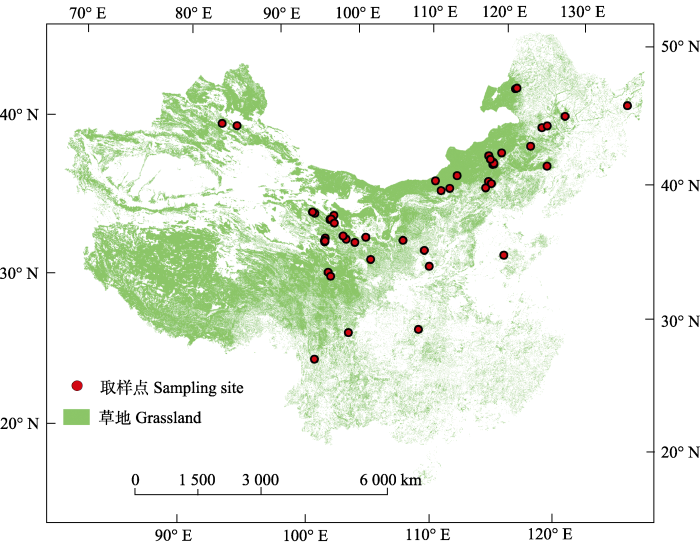
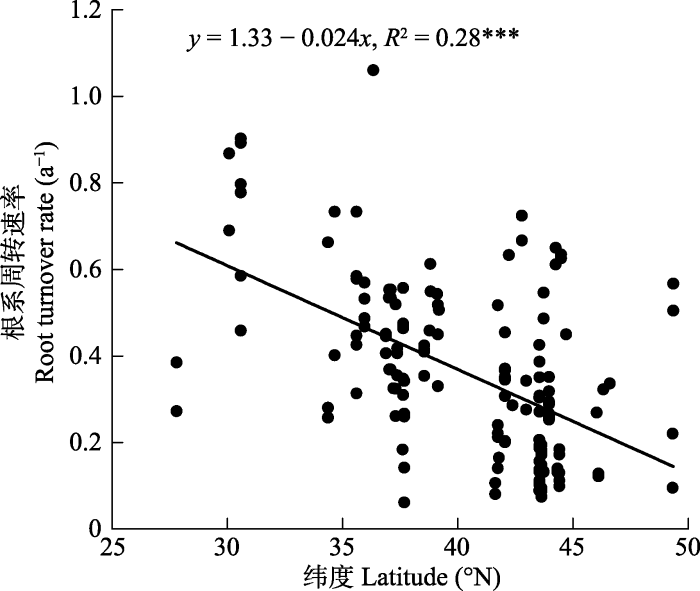
调查样地介于27.79° N与49.35° N之间, 取样点基本涵盖了中国草地的主要分布区(图1)。沿纬度梯度, 根系周转速率与纬度高低呈显著负相关关系(p < 0.001)(图2)。低纬度地区即南方热性草丛区, 根系周转较快, 相反, 北方干旱半干旱典型草原区根系周转相对较慢。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1取样点分布。
Fig. 1Distribution of the study sites.
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2根系周转速率随纬度的分布格局。
Fig. 2Patterns of root turnover rate with increasing latitude.
2.2 不同草地类型根系周转
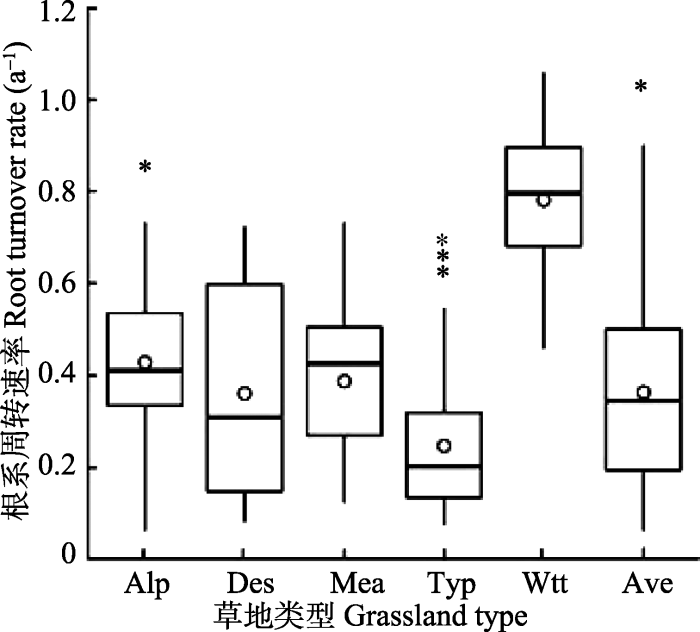
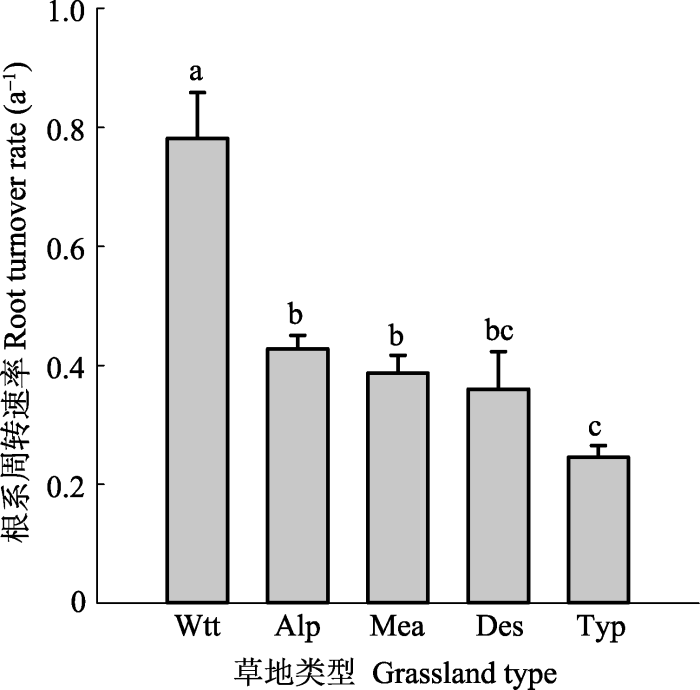
基于Zhu (1993)对中国草地的分类标准, 将研究区草原分为5类, 分别为高寒草甸、典型草原、草甸草原、荒漠草原和热性草丛。各草地类型的根系周转速率分布情况如图3, 除了高寒草甸、典型草原的根系周转速率存在离散点外, 其他类别数据分布相对集中。热性草丛的平均根系周转速率最高, 其次是高寒草甸、草甸草原、荒漠草原、最低的是典型草原, 平均值为0.246·a-1, 即典型草原群落水平上根系平均每5年左右周转一次。不同草地类型之间根系周转速率存在显著差异(p < 0.001)(图4)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同草地类型根系周转速率分布情况。矩形的上端和下端分别代表第四分位数和第一分位数, 横线代表中位数, 空心圆点代表均值, 实心点代表离散点。Alp, 高寒草甸; Ave, 平均值; Des, 荒漠草原; Mea, 草甸草原; Typ, 典型草原; Wtt, 热性草丛。
Fig. 3Changes in root turnover rate among grassland types in China. The boxes show the 25% and 75% percentiles, and the lines, hollow dots and solid dots represent the mean, median and outlier (95% confidence interval) values, respectively. Alp, alpine meadow; Ave, average value; Des, desert steppe; Mea, meadow steppe; Typ, typical steppe; Wtt, warm-temperate tussock;.
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同草地类型根系周转速率(平均值+标准误差)。不同小写字母表示类型间差异显著(p < 0.05)。Alp, 高寒草甸; Des, 荒漠草原; Mea, 草甸草原; Typ, 典型草原; Wtt, 热性草丛。
Fig. 4Root turnover rate for different grassland types (mean + SE). Different lower-case letters indicate significant differences among grassland types (p < 0.05). Alp, alpine meadow; Des, desert steppe; Mea, meadow steppe; Typ, typical steppe; Wtt, warm-temperate tussock.
2.3 根系周转与气候因子及土壤理化性质之间的关系
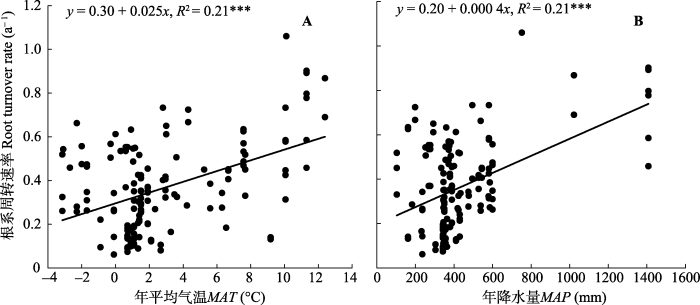
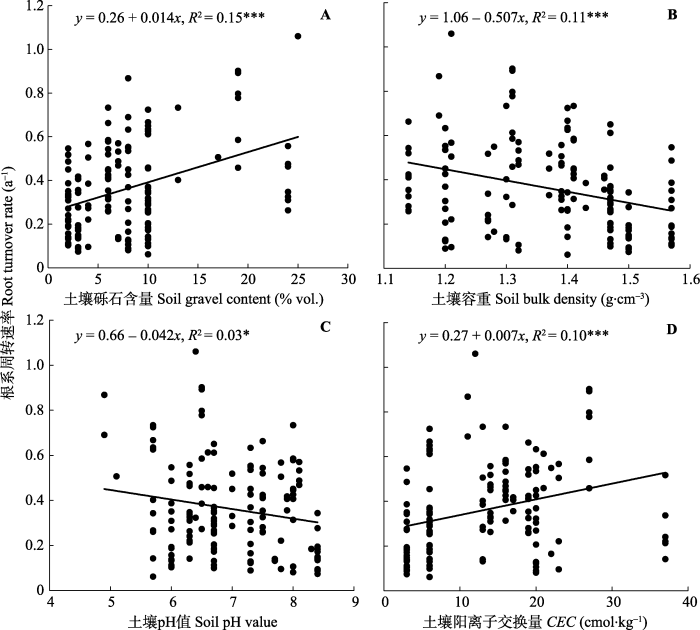
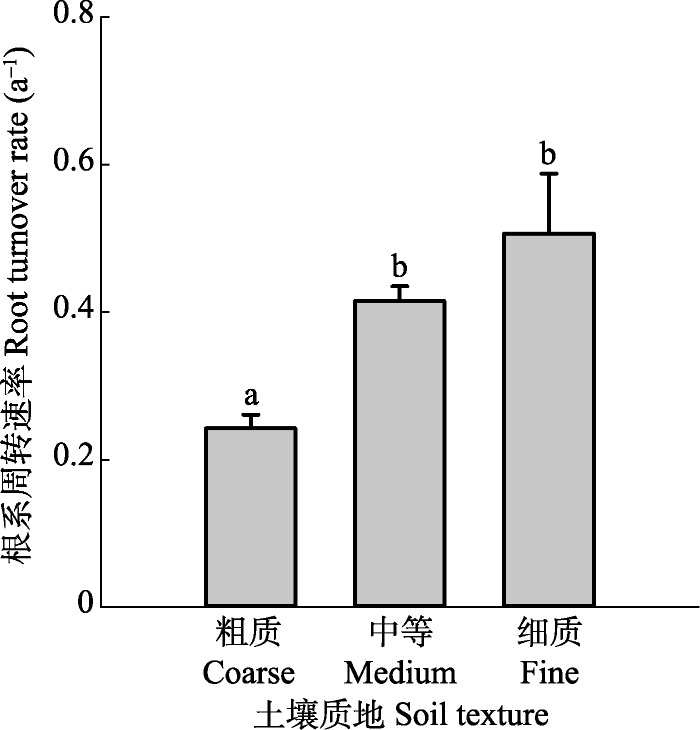
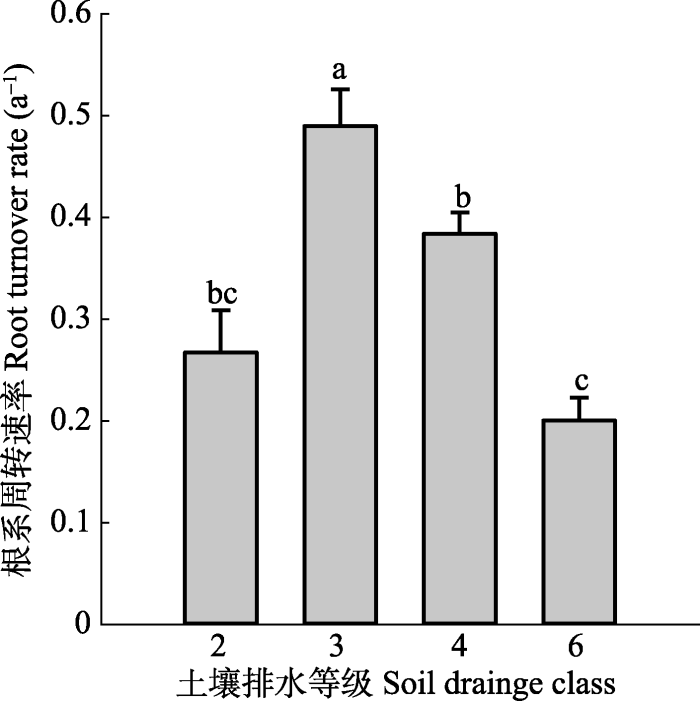
根系周转受多个因子影响, 与年平均气温(MAT)、年降水量(MAP)及土壤砾石含量呈显著正相关关系, 而与土壤容重(BD)、pH值及阳离子交换量(CEC)呈负相关关系(表1)。研究区覆盖的MAT范围为-3.2-12.4 ℃, 在该温度范围内, 根系周转速率随MAT的增加呈线性增加(R2 = 0.21, p < 0.001; 图5A), MAP则介于104-1408 mm之间, 根系周转速率随MAP的增加也呈线性增加(R2 = 0.21, p < 0.001; 图5B)。在土壤理化性质方面, 根系周转速率随砾石含量的增加呈线性增加(R2 = 0.15, p < 0.001; 图6A), 随土壤容重的增加呈线性减小(R2 = 0.11, p < 0.001; 图6B), 随阳离子交换量的增加呈线性增加(R2 = 0.10, p < 0.001; 图6D), 根系周转速率与土壤pH值间存在弱的线性负相关关系(R2 = 0.03, p < 0.05; 图6C)。在土壤质地方面, 根据不同土壤粒径所占百分比大小, 土壤质地分为3类: 粗质(砂粒>65%, 黏粒<18%)、中等(砂粒≤65%, 18% ≤黏粒≤35%), 细质(黏粒>35%), 不同土壤质地间根系周转速率有显著差异(图7), 细质土和中等土根系周转速率较高, 粗质土根系周转速率最低。在土壤排水性方面, 随着排水能力提高, 根系周转速率呈现出先增大后减小的趋势(图8)。Table 1
表1
表1不同草地类型根系周转速率统计
Table 1
| 草地类型 Grassland type | 样本数 N | 平均值 Mean | 标准误差 SE |
|---|---|---|---|
| 高寒草甸 Alpine meadow | 47 | 0.428 | 0.022 |
| 荒漠草原 Desert steppe | 14 | 0.360 | 0.063 |
| 草甸草原 Meadow steppe | 29 | 0.387 | 0.030 |
| 典型草原 Typical steppe | 57 | 0.246 | 0.019 |
| 热性草丛 Warm-temperate tussock | 7 | 0.782 | 0.077 |
新窗口打开|下载CSV
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5根系周转速率和年平均气温(MAT) (A)及平均年降水量(MAP) (B)之间的关系。
Fig. 5Relationship between root turnover rate and mean annual temperature (MAT) (A) and mean annual precipitation (MAP) (B).
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6根系周转速率和土壤砾石含量(A)、容重(B)、pH值(C)及阳离子交换量(D)之间的关系。
Fig. 6Relationships between root turnover rate and soil gravel content (A), bulk density (B), pH value (C) and cation exchange capacity (CEC) (D).
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7根系周转速率和土壤质地之间的关系(平均值+标准误差)。
Fig. 7Relationship between root turnover rate and soil texture (mean + SE).
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8根系周转速率和土壤排水等级之间的关系(平均值+标准误差)。
Fig. 8Relationship between root turnover rate and soil drainage class (mean + SE).
影响根系周转的部分因子之间存在自相关, 如MAT与MAP (r = 0.5, p < 0.001; 表2), 为此, 我们在进行多元线性回归分析之前, 对各因子间的多重共线性进行检验(附录II), 方差膨胀因子vif < 4, 表明各变量之间不存在多重共线性。进一步对根系周转速率6类影响因子(MAT、MAP、Gravel、BD、pH和CEC)进行全子集回归(附录III), 结果表明包含全部5个变量(MAT、MAP、BD、Gravel、pH和CEC)的回归模型为最佳模型, 对根系周转速率的解释度为44% (R2 = 0.44), 各因子对根系周转速率解释度的相对贡献(%)由高到低依次是MAT (34.4)、MAP (22.2)、BD (19.4)、Gravel (17.1)、pH (6.9) (图9)。
Table 2
表2
表2根系周转速率与气候因子和土壤理化性质之间的相关系数
Table 2
| 气候因子 Climatic factor | 土壤理化性质 Soil properties | |||||
|---|---|---|---|---|---|---|
| MAT | MAP | Gravel | BD | pH | CEC | |
| MAP | 0.50*** | |||||
| Gravel | 0.09 | 0.46*** | ||||
| BD | 0.01 | -0.13 | -0.16 | |||
| pH | 0.04 | -0.12 | -0.31*** | -0.17* | ||
| CEC | 0.14 | 0.35*** | 0.11 | -0.76*** | 0.11 | |
| RTR | 0.45*** | 0.46*** | 0.39*** | -0.33*** | -0.18* | 0.30*** |
新窗口打开|下载CSV
BD, bulk density (g.cm-3); CEC, cation exchange capacity (cmol.kg-1); MAT, mean annual temperature (℃); MAP, mean annual precipitation (mm); Gravel, gravel content (% vol.); pH, pH value; RTR, root turnover rate (a-1). *, p < 0.05; ***, p < 0.001.
图9
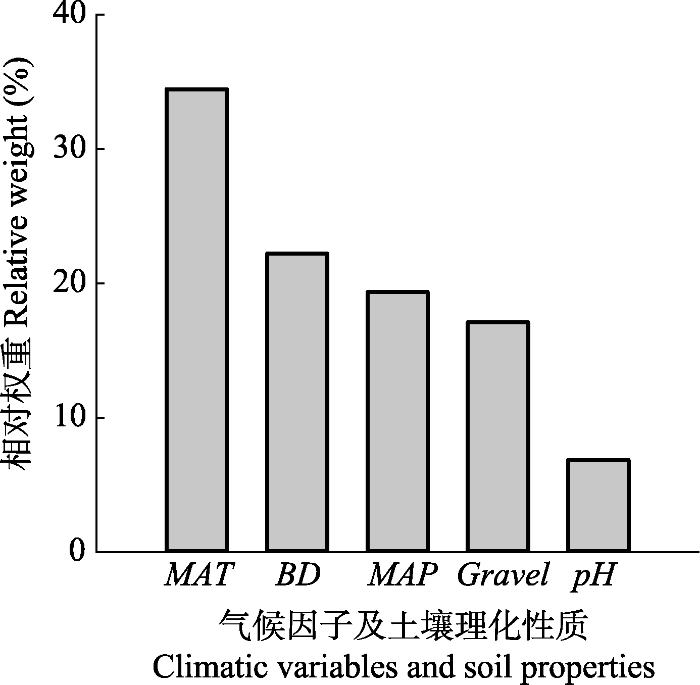 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9气候和土壤因子对根系周转速率解释度的相对贡献。BD, 土壤容重(kg.dm-3); MAT, 年平均气温(℃); MAP, 平均年降水量(mm); Gravel, 土壤砾石含量(% vol.); pH, 酸碱性。
Fig. 9The relative contributions of climatic variables and soil properties to root turnover rate. BD, bulk density (g.cm-3); MAT, mean annual temperature (℃); MAP, mean annual precipitation (mm); Gravel, gravel content (% vol.); pH, pH value.
3 讨论
3.1 气候因子对根系周转的影响机制
根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(de Vries, 1975), 使得根系生物量的消耗速率提高, 因而加快了根系周转; (2)在一定的温度范围内, 土壤有机质的矿化速率随温度的升高而增大(王常慧等, 2004; Curtin et al., 2012)。温度越高, 根系凋落物被微生物分解的速率就越快, 土壤中可利用的无机氮含量就越高, 使根系氮含量增加, 根系呼吸速率加快, 消耗根系中的碳水化合物, 进而加快根系周转(Burton et al., 2000; Eissenstat et al., 2000; Bai et al., 2008); (3)温暖地区全年无霜期较长, 延长了放牧时间(Ryan, 1991; Eissenstat & Yanai, 1997)。放牧过程中草食动物的采食和践踏会降低植物盖度, 使地表裸露面积增加, 接受更多的阳光照射, 提高土壤温度, 进而加快根系周转(马涛等, 2009; 高英志等, 2017)。在全球气候变化背景下, 随着气温上升, 中国草地生态系统的根系周转可能随之加快。沿平均年降水梯度, 根系周转呈现出线性增加的趋势。和温度升高带来的影响类似, 降水的增加提高了土壤湿度, 也会提高维持呼吸速率和土壤有机物的矿化速率, 进而加快根系周转。局地尺度上的控制实验也证实, 增水可以提高草地生态系统的根系周转速率(Bai et al., 2010; Larreguy et al., 2012)。另外, 降水增加可以提高地上生物量(Ni, 2004), 从而引起草食动物的增加及放牧强度的增加, 间接影响根系周转。
3.2 土壤理化性质对根系周转的影响机制
在土壤物理性质方面, 砾石含量、容重、土壤质地和排水性都是通过影响土壤水分来间接影响根系周转。已有研究表明, 砾石含量越高的土壤, 其导水率越大(潘永洁等, 2015)。增大砾石含量可以提高入渗量及有效贮水量(Wilcox & Wood, 1988; 时忠杰等, 2008)。而且砾石的存在可以避免由于降雨直接打击土壤而形成地表结皮(Wilcox & Wood, 1988)。另外, 增加砾石含量可以降低因为蒸发而造成的水分损失(Kosmas & Danalatos, 1993; 吕国安等, 2000; 陈士辉等, 2005)。
一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(李卓等, 2010), 随着土壤容重的增加, 土壤的团粒结构丧失, 土壤变得更加紧密, 导致入渗能力、滞留贮水量、吸持贮水量和饱和贮水量降低(王辉等, 2007; 李卓等, 2010; 吴军虎等, 2013), 土壤对水分的调节能力下降, 不利于植物根系的生长(吕殿青等, 2009), 进而减缓根系周转。
土壤质地是土壤不同粒径矿物颗粒组成的综合反映, 与土壤通气状况、保水保肥能力密切相关。细质土中根系周转速率最高, 可能是由于细质土中黏粒所占比重较大, 土壤保水能力较好, 而降水(尤其是生长季降水)是影响草地生态系统生产力的主要限制因子(Bai et al., 2004)。因此, 保水能力高的细质土更有利于植物根系的生长和周转(周文昌等, 2016)。然而, 随着排水等级提高(保水能力降低), 根系周转速率呈现先升高后下降的趋势, 可能是因为保水能力过高的土壤其透气性较差, 抑制了根系及土壤微生物呼吸, 从而降低根系周转速率。所以, 在适度的保水和透气性条件下根系周转速率才能达到最高。
在土壤的化学性质方面, pH值及CEC主要通过影响酶的活性及养分的可利用性来影响根系周转。
本研究中土壤pH值介于4.9-8.4, 涵盖了弱酸性土、中性土和弱碱性土。研究认为, 弱酸性的土壤环境有利于土壤有机物矿化相关酶的活性(Leifeld et al., 2013)。另外, 有研究表明, 对内蒙古典型草原弱碱性土壤进行酸化处理, 可以提高土壤水分及磷的可利用性(Chen et al., 2013)。因而弱酸性的土壤环境有利于植物根系对水分和养分的获取, 促进根系周转。
CEC是反映土壤保肥供肥性能的重要指标, 土壤速效磷含量与CEC呈显著正相关关系(姜林等, 2012)。因而CEC越大, 土壤肥力越高, 根系周转也就越快。
3.3 不同草地类型根系周转存在差异
本研究中涉及的样点南北跨越22个纬度, 东西跨越46个经度, 覆盖范围较广, 5种不同草地类型是其所在地区植被、气候、地形、土壤等特性的综合反映。不同草地类型之间根系周转存在显著的差异, 说明气候、土壤等因子的不同组合对根系周转的影响结果不同。对于温带地区的荒漠草原而言, 降水是影响根系周转的主要限制因素(冯雨峰, 1990), 而对于降水丰沛的高寒草甸而言, 温度就成了限制根系周转的主要因素(Leifeld et al., 2015)。这就造成了在全球气候变化背景下, 不同草地类型的根系周转对气候变化的响应有所不同。比如随着全球气温上升, 高寒草甸根系周转加快, 而荒漠草原的根系周转将可能受到更加严重的限制。3.4 中国草地根系周转的格局和驱动因子与全球及其他区域研究结果的差异
本研究中根系周转随温度升高而加快, 该结果与全球尺度(Gill & Jackson, 2000)及欧洲山地草原(Leifeld et al., 2015)的研究结果一致, 但是函数关系却有明显差异。前者是线性增长的关系, 后两者皆是指数增长的关系, 而一些局地尺度上的控制实验则表明年平均气温对根系周转没有明显的影响(Bai et al., 2010; Gill & Jackson, 2000), 甚至是负的影响(Edwards et al., 2004)。由此可见, 温度对根系周转的影响具有很强的尺度依赖性。对降水而言, 本研究中根系周转速率和降水之间呈现显著的正相关关系, 与Bai (2010)在局地尺度上的控制实验结果一致。中国草地根系周转沿温度和降水梯度上的分布格局与全球尺度的研究结果存在较大的差异, 分析原因主要包括以下两方面: 一方面缘于尺度效应, 另一方面是Gill和Jackson (2000)的研究中取样点较少, 且分布不均以及周转速率计算方法上的不统一带来的不确定性引起的。气候因子和土壤理化性质通过影响土壤酶的活性、呼吸、矿化、土壤水分及养分的可利用性等途径共同影响根系周转。不同草地类型之间根系周转速率存在显著差异。根系周转的空间格局具有尺度依赖性, 中国草地的研究结果与其他区域存在差异, 在对全球尺度根系周转的研究中需要将中国草地的数据纳入其中, 多尺度、多视角进行整合分析, 为草地生态系统生产力及固碳潜力估算提供更加可靠的参数, 具有重要的现实意义。
致谢 中国科学院战略性先导科技专项草地课题的李永强、刘伟、孙庚、唐立孙、刘耘华、吴彦、郝彦宾为本研究提供了部分野外实测数据, 在此表示感谢。
附录I 数据提取文献来源
Appendix I List of papers from which data are extracted for this database附录II 气候及土壤因子多重共线性检验
Appendix II Multicollinearity test of climatic variables and soil properties附录III 气候因子及土壤理化性质
Appendix III Climatic variables and soil properties
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/BF00378292URLPMID:28310856 [本文引用: 1]

Two methods of estimating fine root production and turnover are compared for 13 forest ecosystems exhibiting a wide range in form (NH 4 + vs. NO 3 - ) and quantity of available nitrogen. The two methods are by comparison of seasonal maxima and minima in biomess and by nitrogen budgeting. Both methods give similar results for stands with low rates of nitrification. The budgeting method predicts higher fine root turnover and productivity than the max-min method for systems with significant rates of nitrification.
DOI:10.1007/s11104-005-5675-9URL [本文引用: 1]
The aims of this study were to determine whether elevated atmospheric CO 2 concentration modifies plant organic matter (OM) fluxes to the soil and whether any change in the fluxes can modify soil OM accumulation. Measurements were made in a grazed temperate grassland after almost 402years exposure to elevated atmospheric CO 2 (47502μl02l -1 ) using a Free Air CO 2 Enrichment (FACE) facility located in the North Island of New Zealand. Aboveground herbage biomass and leaf litter production were not altered by elevated CO 2 but root growth rate, as measured with the ingrowth core method, and root turnover were strongly stimulated by elevated CO 2 particularly at low soil moisture contents during summer. Consequently, significantly more plant material was returned to the soil under elevated CO 2 leading to an accumulation of coarse (> 102mm) particulate organic matter (POM) but not of finer POM fractions. The accumulating POM exhibited a lower C/N ratio, which was attributed to the higher proportion of legumes in the pasture under elevated CO 2 . Only small changes were detected in the size and activity of the soil microbial biomass in response to the POM accumulation, suggesting that higher organic substrate availability did not stimulate microbial growth and activity despite the apparent lower C/N ratio of accumulating POM. As a result, elevated CO 2 may well lead to an accumulation of OM in grazed grassland soil in the long term.
DOI:10.1111/gcb.2010.16.issue-4URL [本文引用: 6]
[本文引用: 1]
DOI:10.1038/nature02850URLPMID:202020202020202020202020 [本文引用: 1]
Numerous studies have suggested that biodiversity reduces variability in ecosystem productivity through compensatory effects109“6;that is, a species increases in its abundance in response to the reduction of another in a fluctuating environment1,7. But this view has been challenged on several grounds809“10. Because most studies have been based on artificially constructed grass- lands with short duration, long-term studies of natural ecosystems are needed. On the basis of a 24-year study of the Inner Mongolia grassland, here we present three key findings. First, that January-July precipitation is the primary climatic factor causing fluctuations in community biomass production; second, that ecosystem stability (conversely related to variability in community biomass production) increases progressively along the hierarchy of organizational levels (that is, from species to functional group to whole community); and finally, that the community-level stability seems to arise from compensatory interactions among major components at both species and functional group levels. From a hierarchical perspective, our results corroborate some previous findings of compensatory effects1,4,7,11. Undisturbed mature steppe ecosystems seem to culminate with high biodiversity, productivity and ecosystem stability concurrently. Because these relationships are correlational, further studies are necessary to verify the causation among these factors. Our study provides new insights for better management and restoration of the rapidly degrading Inner Mongolia grassland.
DOI:10.1007/s004420000455URL [本文引用: 1]
DOI:10.5846/stxb201301190118URL [本文引用: 1]
地下根系是草原生态系统的重要组成部分,其生物量及其净生产力对地下碳库具有直接与间接作用,分析地下生物量季节动态与周转对深入揭示草原生态系统碳库动态及其固碳速率与潜力具有重要意义。应用钻土芯法对不同利用方式或管理措施下内蒙古草甸草原、典型草原地下生物量动态及其与温度、降水的相关性研究表明:草甸草原和典型草原地上生物量季节动态均为单峰型曲线,与上月降水显著正相关(P0.05),但地下生物量季节动态表现为草甸草原呈"S"型曲线,典型草原则是双峰型曲线,与温度、降水相关性均不显著(P0.05);两种草原根冠比和地下生物量垂直分布均为指数函数曲线,根茎型草原地下生物量集中在土壤0—5 cm,丛生型草原地下生物量集中于土壤5—10 cm,根冠比值在生长旺季(7—8月份)最小。草甸草原地下净生产力及碳储量范围分别为2167—2953 g m-2a-1和975—1329 gC m-2a-1,典型草原为2342—3333 g m-2a-1和1054—1450 gC m-2a-1,地下净生产力及其碳储量约为地上净生产力及其碳储量的10倍,具有较大的年固碳能力,且相对稳定;地下净生产力与地上净生产力呈显著负相关性(P0.05);地下生物量碳库是地上生物量碳库的10倍左右,适度放牧可增加地下生产力,但长期过度放牧显著降低其地下生物量与生产力,并使其垂直分布趋向于浅层化。
DOI:10.5846/stxb201301190118URL [本文引用: 1]
地下根系是草原生态系统的重要组成部分,其生物量及其净生产力对地下碳库具有直接与间接作用,分析地下生物量季节动态与周转对深入揭示草原生态系统碳库动态及其固碳速率与潜力具有重要意义。应用钻土芯法对不同利用方式或管理措施下内蒙古草甸草原、典型草原地下生物量动态及其与温度、降水的相关性研究表明:草甸草原和典型草原地上生物量季节动态均为单峰型曲线,与上月降水显著正相关(P0.05),但地下生物量季节动态表现为草甸草原呈"S"型曲线,典型草原则是双峰型曲线,与温度、降水相关性均不显著(P0.05);两种草原根冠比和地下生物量垂直分布均为指数函数曲线,根茎型草原地下生物量集中在土壤0—5 cm,丛生型草原地下生物量集中于土壤5—10 cm,根冠比值在生长旺季(7—8月份)最小。草甸草原地下净生产力及碳储量范围分别为2167—2953 g m-2a-1和975—1329 gC m-2a-1,典型草原为2342—3333 g m-2a-1和1054—1450 gC m-2a-1,地下净生产力及其碳储量约为地上净生产力及其碳储量的10倍,具有较大的年固碳能力,且相对稳定;地下净生产力与地上净生产力呈显著负相关性(P0.05);地下生物量碳库是地上生物量碳库的10倍左右,适度放牧可增加地下生产力,但长期过度放牧显著降低其地下生物量与生产力,并使其垂直分布趋向于浅层化。
DOI:10.1111/1365-2745.12119URL [本文引用: 1]
Anthropogenic acid deposition鈥搃nduced soil acidification is one of the major threats to biodiversity, ecosystem functioning and services. Few studies, however, have explored in detail how above-ground changes in plant species richness and productivity resulting from soil acidification are mediated by effects on below-ground biota and soil properties.To increase our understanding of this linkage, we collected data on below- and above-ground communities and soil properties in a 3-year field experiment with seven levels of acid addition rate to build-up broad intensities of soil acidification in the semi-arid Inner Mongolian grassland.Acid addition directly elevated concentrations of soil Al3+ ions, decreased the base cations Ca2+, Mg2+ and Na+, and increased soil moisture and available phosphorus. Acid addition also appears to have altered the soil microbial community via changes in H+ and Al3+ ions and altered the nematode community via changes in H+ ions and soil moisture.The observed changes in soil N availability (i.e. net N mineralization, -N and -N) could be explained by mediating changes in the H+ and Al3+ ions, microbial community (i.e. community structure, bacteria and fungi/bacteria as indicated by phospholipid fatty acids analysis) and the nematode community (i.e. total abundance, taxa richness and maturity index).Declines in plant species richness and productivity were greater at high intensities of soil acidification in the second sampling year than in the first sampling year. The changes in plant community observed were mostly explained by soil nutrient pathways (e.g. N availability or base mineral cations), which were in turn regulated by the soil microbial or nematode communities as well as by the direct effects of the increase in H+ or Al3+ ions.Synthesis. Our results suggest that the below-ground microbial and nematode communities are more sensitive to soil acidification than the plant communities are, and further that soil acidification nduced changes in plants are mediated by changes in below-ground communities and soil nutrients. These findings improve our understanding of the links between below- and above-ground communities in the Inner Mongolia grassland, especially in the context of anthropogenic acid enrichment.
DOI:10.3321/j.issn:1000-694X.2005.03.023URL [本文引用: 1]
为分析不同粒径砂砾石对砂田西瓜蒸散量和土壤蒸发的影响,2004年在位于黄土高原西北部的皋兰县境内进行了不同粒径砂砾石覆盖的水分效应研究,结果证明:砂砾石粒径大小对砂田西瓜蒸散量和土壤蒸发有显著影响,粒径2~5 mm覆盖处理的蒸散量显著低于粒径5~20 mm和20~60 mm处理,但与不覆砂的对照没有显著差异.粒径愈大,砂田土壤蒸发愈多,土壤蒸发在西瓜田总蒸散中占的比例愈高.覆砂能够有效减少土壤蒸发,未覆砂处理全生育期土壤蒸发耗水占西瓜蒸散的40.7%,而覆砂处理仅占总蒸散的17.8%~25%.西瓜田覆砂加覆膜,土壤蒸发比不覆盖田减少78~93.7 mm,比仅覆砂不覆膜田减少16.9~26.3 mm.不同粒径砂砾石处理之间的产量差异不显著,但水分生产率有显著差异,2~5 mm粒径砂砾石覆盖处理的水分生产率显著高于20~60mm粒径处理.但砂砾石粒径减小,砂田西瓜的含糖量降低.研究结果还证明,西瓜田覆砂能有效地提高其产量,含糖量和水分生产率,适合砂田覆盖的砂砾石粒径以5~20 mm为宜.
DOI:10.3321/j.issn:1000-694X.2005.03.023URL [本文引用: 1]
为分析不同粒径砂砾石对砂田西瓜蒸散量和土壤蒸发的影响,2004年在位于黄土高原西北部的皋兰县境内进行了不同粒径砂砾石覆盖的水分效应研究,结果证明:砂砾石粒径大小对砂田西瓜蒸散量和土壤蒸发有显著影响,粒径2~5 mm覆盖处理的蒸散量显著低于粒径5~20 mm和20~60 mm处理,但与不覆砂的对照没有显著差异.粒径愈大,砂田土壤蒸发愈多,土壤蒸发在西瓜田总蒸散中占的比例愈高.覆砂能够有效减少土壤蒸发,未覆砂处理全生育期土壤蒸发耗水占西瓜蒸散的40.7%,而覆砂处理仅占总蒸散的17.8%~25%.西瓜田覆砂加覆膜,土壤蒸发比不覆盖田减少78~93.7 mm,比仅覆砂不覆膜田减少16.9~26.3 mm.不同粒径砂砾石处理之间的产量差异不显著,但水分生产率有显著差异,2~5 mm粒径砂砾石覆盖处理的水分生产率显著高于20~60mm粒径处理.但砂砾石粒径减小,砂田西瓜的含糖量降低.研究结果还证明,西瓜田覆砂能有效地提高其产量,含糖量和水分生产率,适合砂田覆盖的砂砾石粒径以5~20 mm为宜.
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/1051-0761(2001)011[0371:NPPITF]2.0.CO;2URL [本文引用: 1]
DOI:10.2136/sssaj2012.0011URL [本文引用: 1]
Concern over climate change has stimulated interest in the temperature and moisture dependence of soil organic matter decomposition. In particular, there has been intense debate in relation to the factors that determine the temperature dependence of C mineralization. We examined temperature and moisture responses of C and N mineralization in an 85-d laboratory incubation (factorial combination of four temperatures [5, 12, 18, 25 C] and five moisture treatments [matric potential from -5 to -1200 kPa]) using three agriculturally important New Zealand soils (soils with a history of pasture, arable, or vegetable cropping). Mineralization was linearly related to gravimetric moisture content, except in the high-C pasture soil where O
DOI:10.2307/1935260URL [本文引用: 4]
Quantitative measurements of root systems on a seasonal basis for a 34-inch profile in Central Missouri Prairie ranged from 1,449 g/m^2 prior to resumption of growth in the spring to 1,901 g/m^2 at the end of the growing season. Underground parts in the upper 2 in. of the profile varied from 48% of the total quantity in April to 60% in July. Over 80% of the root mass occurred in the A_1 horizon, or the top 10 in. of the profile. Annual increment in the A_1 horizon or surface 10 in. was 429 g/m^2. In the A_2 horizon, 10-18 in., the measurement was 40 g/m^2 and in the B"2 horizon, 18-30 in., 41 g/m^2. These values were approximately 25% of the maximum dry matter measurement for each profile level, suggesting a turnover for the root system as a whole every 4 years. Variations in turnover for different parts of the root system were indicated, however, as shown by an annual loss of 22.8% for rhizomes, and 40.8% for roots in the 0- to 2-in. zone. Decomposition constants showed a marked decrease with depth, ranging from 4.3% in the A_1 horizon to 0.8% in the B^2 horizon. Estimated time required to reach 99% organic matter equilibrium under present conditions of production and breakdown was 110, 420, and 590 years for the A_1, A_2, and B"2 horizons respectively. Increasing time values would be expected with increasing depth as the reduction in annual increment was proportionately greater at lower levels in the profile than that for the total root-humus product.
DOI:10.1105/tpc.111.087312URL
ABSTRACT To what extent does plant clonality contribute to the assemblage of species in communities? Two apparently contrasting, and largely untested, hypotheses envisage the potential role of plant clonal traits in community assembly: (i) environmental filters constrain coexisting species to have functionally similar traits (i.e. trait convergence); (ii) niche differentiation selects for functionally dissimilar species (i.e. trait divergence) allowing them to exploit different spatial and temporal niches. These hypotheses are assessed using a large dataset of 369 plots (100 m2) covering altitudes between 4100 and 5800 m a.s.l. and including the major vegetation types found in Ladakh, NW Himalaya. Patterns of clonal traits, coexistence and turnover were assessed using a functional diversity partitioning framework in the context of different null-models. Functional diversity was expressed both for morphologically delimited clonal growth forms (17 categorical growth forms) and for functionally delimited clonal characters (combining 16 different traits differentiating the 17 growth forms). PERMANOVA revealed that both 8289(within-plots) and 8389(between-plots) functional diversity varied across environmental conditions and vegetation types highlighting a filtering effect on clonal traits. Alpha diversity, however, was more stable across habitats than 8389diversity. Despite the significant turnover of clonal traits across habitats, most of the diversity of clonal traits was found within plots, with a higher trait divergence than expected by chance, which suggests niche differences determine species coexistence. While both trait convergence and trait divergence were detected, convergence was stronger when using null-models that shuffled all species in the regional pool across plots and functional diversity expressed in terms of different clonal growth forms. Divergence, in contrast, was detected mostly when using null-models that shuffled species cover across species co-occuring in given plots and considering functional diversity in terms of clonal traits. By detecting both trait convergence and trait divergence this study supports both initial hypotheses and brings new evidence on the relevance of clonal traits as a function of species that both inhabit different environments and coexist.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-2486.2004.00729.xURL [本文引用: 1]
Abstract Accurate knowledge of the response of root turnover to a changing climate is needed to predict growth and produce carbon cycle models. A soil warming system and shading were used to vary soil temperature and received radiation independently in a temperate grassland dominated by Holcus lanatus L. Minirhizotrons allowed root growth and turnover to be examined non-destructively. In two short-term (8 week) experiments, root responses to temperature were seasonally distinct. Root number increased when heating was applied during spring, but root death increased during autumnal heating. An experiment lasting 12 months demonstrated that any positive response to temperature was short-lived and that over a full growing season, soil warming led to a reduction in root number and mass due to increased root death during autumn and winter. Root respiration was also insensitive to soil temperature over much of the year. In contrast, root growth was strongly affected by incident radiation. Root biomass, length, birth rate, number and turnover were all reduced by shading. Photosynthesis in H. lanatus exhibited some acclimation to shading, but assimilation rates at growth irradiance were still lower in shaded plants. The negative effects of shading and soil warming on roots were additive. Comparison of root data with environmental measurements demonstrated a number of positive relationships with photosynthetically active radiation, but not with soil temperature. This was true both across the entire data set and within a shade treatment. These results demonstrate that root growth is unlikely to be directly affected by increased soil temperatures as a result of global warming, at least in temperate areas, and that predictions of net primary productivity should not be based on a positive root growth response to temperature.
DOI:10.1046/j.1469-8137.2000.00686.xURL [本文引用: 1]
Root turnover is important to the global carbon budget as well as to nutrient cycling in ecosystems and to the success of individual plants. Our ability to predict the effects of environmental change on root turnover is limited by the difficulty of measuring root dynamics, but emerging evidence suggests that roots, like leaves, possess suites of interrelated traits that are linked to their life span. In graminoids, high tissue density has been linked to increased root longevity. Other studies have found root longevity to be positively correlated with mycorrhizal colonization and negatively correlated with nitrogen concentration, root maintenance respiration and specific root length. Among fruit trees, apple roots (which are of relatively small diameter, low tissue density and have little lignification of the exodermis) have much shorter life spans than the roots of citrus, which have opposite traits. Likewise, within the branched network of the fine root system, the finest roots with no daughter roots tend to have higher N concentrations, faster maintenance respiration, higher specific root length and shorter life spans than secondary and tertiary roots that bear daughter roots. Mycorrhizal colonization can enhance root longevity by diverse mechanisms, including enhanced tolerance of drying soil and enhanced defence against root pathogens. Many variables involved in building roots might affect root longevity, including root diameter, tissue density, N concentration, mycorrhizal fungal colonization and accumulation of secondary phenolic compounds. These root traits are highly plastic and are strongly affected by resource supply (CO 2 , N, P and water). Therefore the response of root longevity to altered resource availability associated with climate change can be estimated by considering how changes in resource availability affect root construction and physiology. A cost benefit approach to predicting root longevity assumes that a plant maintains a root only until the efficiency of resource acquisition is maximized. Using an efficiency model, we show that reduced tissue Nconcentration and reduced root maintenance respiration, both of which are predicted to result from elevated CO 2 , should lead to slightly longer root life spans. Complex interactions with soil biota and shifts in plant defences against root herbivory and parasitism, which are not included in the present efficiency model, might alter the effects of future climate change on root longevity in unpredicted ways.
DOI:10.1016/S0065-2504(08)60005-7URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1046/j.1469-8137.1997.00804.xURL [本文引用: 1]
summary Monoliths of two contrasting vegetation types, a species-rich grassland on a brown earth soil over limestone and species-poor community on a peaty gley, were transferred to solardomes and grown under ambient (350 μ 1 611 ) and elevated (600 μ11 611 ) CO 2 for 2 yr. Shoot biomass was unaltered but root biomass increased by 40–50% under elevated CO 2 . Root production was increased by elevated CO 2 in the peat soil, measured both as instantaneous and cumulative rates, but only the latter measure was increased in the limestone soil. Root growth was stimulated more at 6 cm depth than at 10 cm in the limestone soil. Turnover was faster under elevated CO 2 in the peat soil, but there was only a small effect on turnover in the limestone soil. Elevated CO 2 reduced nitrogen concentration in roots and might have increased mycorrhizal colonization. Respiration rate was correlated with N concentration, and was therefore lower in roots grown at elevated CO 2 . Estimates of the C budget of the two communities, based upon root production and on net C uptake, suggest that C sequestration in the peat soil increases by c. 0.2 kg C m 612 yr 611 (= 2 t ha yr 611 ) under elevated CO 2 .
DOI:10.11920/xnmdzk.2017.02.001URL [本文引用: 1]
放牧作为一种人类活动的干扰因子,主要通过动物的采食、践踏及其排泄物的输入对草原生态系统产生影响.放牧直接作用于草原生态系统的地上部分和土壤,从而改变草原生态系统的物质生产和碳分配,进而影响到地下净生产力和根系周转.本文综述了放牧及刈割对地下生物量、地下净生产力、碳分配和根系周转的影响,得到如下结论:北方干旱、半干旱草原在重度放牧下,地下净生产力、地下生物量会显著下降,地下碳分配减少.放牧也通常会加快根系周转.研究不同放牧方式(羊单牧、牛单牧、牛羊混牧和无牧)对根系周转的影响,发现牛单牧条件下细根的年周转率最高.文章也对今后根系生态学的研究方向提出建议和展望.
DOI:10.11920/xnmdzk.2017.02.001URL [本文引用: 1]
放牧作为一种人类活动的干扰因子,主要通过动物的采食、践踏及其排泄物的输入对草原生态系统产生影响.放牧直接作用于草原生态系统的地上部分和土壤,从而改变草原生态系统的物质生产和碳分配,进而影响到地下净生产力和根系周转.本文综述了放牧及刈割对地下生物量、地下净生产力、碳分配和根系周转的影响,得到如下结论:北方干旱、半干旱草原在重度放牧下,地下净生产力、地下生物量会显著下降,地下碳分配减少.放牧也通常会加快根系周转.研究不同放牧方式(羊单牧、牛单牧、牛羊混牧和无牧)对根系周转的影响,发现牛单牧条件下细根的年周转率最高.文章也对今后根系生态学的研究方向提出建议和展望.
DOI:10.1046/j.1469-8137.2000.00681.xURL [本文引用: 11]
Root turnover is a critical component of ecosystem nutrient dynamics and carbon sequestration and is also an important sink for plant primary productivity. We tested global controls on root turnover across climatic gradients and for plant functional groups by using a database of 190 published studies. Root turnover rates increased exponentially with mean annual temperature for fine roots of grasslands ( r 2 = 0.48) and forests ( r 2 = 0.17) and for total root biomass in shrublands ( r 2 = 0.55). On the basis of the best-fit exponential model, the Q 10 for root turnover was 1.4 for forest small diameter roots (5 mm or less), 1.6 for grassland fine roots, and 1.9 for shrublands. Surprisingly, after accounting for temperature, there was no such global relationship between precipitation and root turnover. The slowest average turnover rates were observed for entire tree root systems (10% annually), followed by 34% for shrubland total roots, 53% for grassland fine roots, 55% for wetland fine roots, and 56% for forest fine roots. Root turnover decreased from tropical to high-latitude systems for all plant functional groups. To test whether global relationships can be used to predict interannual variability in root turnover, we evaluated 14 yr of published root turnover data from a shortgrass steppe site in northeastern Colorado, USA. At this site there was no correlation between interannual variability in mean annual temperature and root turnover. Rather, turnover was positively correlated with the ratio of growing season precipitation and maximum monthly temperature ( r 2 = 0.61). We conclude that there are global patterns in rates of root turnover between plant groups and across climatic gradients but that these patterns cannot always be used for the successful prediction of the relationship of root turnover to climate change at a particular site.
DOI:10.1111/j.1442-9993.1982.tb01311.xURL [本文引用: 1]
Plant biomass, net primary productivity and dry matter turnover were studied in a grassland situated in a tropical monsoonal climate at Kurukshetra, India (29°58'N, 76°51'E). Based on differences in vegetation in response to microrelief, three stands were distinguished on the study site. The stand I was dominated by Sesbania bispinosa , stand II represented mixed grasses and stand III was dominated by Desmostachya bipinnata. Floristic composition of the three stands revealed the greatest number of species on stand II (75). The study of life form classes indicated a thero-cryptophytic flora. The biomass of live shoots in all the three stands attained a maximum value in September (424–1921 g m -2 ) and below ground plant biomass in November (749–1868 g m -2 ). The annual above ground net primary production was greatest on stand I (2143 g m -2 ) and lowest on stand II (617 g m -2 ). The rate of production was highest during the rainy season (15.34 to 3.18 g m -2 day -2 ). Below ground net production ranged from 1592 to 785 g m -2 y -2 and the rates were high in winter and summer seasons. Total annual net primary production was estimated to be 3141, 1403, 2493 and 2134 g m -2 on stands I, II, III and on the grassland as a whole, respectively. The turnover of total plant biomass plus below ground biomass indicated almost a complete replacement of phytomass within the year. The system transfer functions showed greater transfer of material from total net primary production to the shoot compartment during rainy season and to the root compartment during winter and summer seasons.
DOI:10.1139/b87-105URL [本文引用: 1]
Abstract Root dynamics were studied using root windows at Konza Prairie, a tallgrass prairie in north central Kansas, during dry (1984) and wet (1985) years. Amounts, production, and disappearance of root length decreased during drought but increased when rains resumed; however, standing crop remained low. The 1985 root lengths increased throughout the growing season, while production and disappearance remained constant. Yearly summaries of amounts, productivity, and decomposition by 10-cm increments in soil depth show that the effect of drought on these variables decreased with increasing soil depth. Turnover rates of root length averaged 564 in the dry year versus 389% in the wet year, with the largest difference noted in the 0- to 10-cm depth (800 in 1984 versus 540% in 1985). Production and decay patterns observed using root windows were also noted in root biomass data (obtained from soil cores). The average total root biomass turnover rate was 31%. Failure to sort below-ground materials into tissue types (rhizomes, roots) and live versus dead status results in reduced estimates of biomass turnover rates. The greatest possible separation of plant components presents the most comprehensive picture of (belowground) growth dynamics.
DOI:10.1360/csb2004-49-13-1226URL [本文引用: 2]
生态学100多年的探索和发展主要集中在地上部分.然而,当今的生态学家已经越来越强烈地认识到,鲜为人知的地下部分已成为生态系统结构、功能与过程研究中最不确定的因素,因而严重制约着生态系统与全球变化研究的理论拓展.自1990年代后期以来,伴随着全球生态学研究的深入,一个新兴的生态学领域——地下生态学(belowgroundecology)开始形成,并得到了快速发展.地下生态学从不同学科层次探索地下部分的结构、功能、过程以及与地上部分的关系,并特别关注其对全球变化的响应.它的研究对象包括植物根系、地下动物和土壤微生物.分析了生态系统地上和地下部分的关联、根系生态、根系生物地理,以及地下生物多样性等方面的主要研究进展和亟待解决的问题,着重评述地下过程对全球变化响应的若干理论问题,指出地下生态学将是21世纪生态学的重要发展方向.
DOI:10.1360/csb2004-49-13-1226URL [本文引用: 2]
生态学100多年的探索和发展主要集中在地上部分.然而,当今的生态学家已经越来越强烈地认识到,鲜为人知的地下部分已成为生态系统结构、功能与过程研究中最不确定的因素,因而严重制约着生态系统与全球变化研究的理论拓展.自1990年代后期以来,伴随着全球生态学研究的深入,一个新兴的生态学领域——地下生态学(belowgroundecology)开始形成,并得到了快速发展.地下生态学从不同学科层次探索地下部分的结构、功能、过程以及与地上部分的关系,并特别关注其对全球变化的响应.它的研究对象包括植物根系、地下动物和土壤微生物.分析了生态系统地上和地下部分的关联、根系生态、根系生物地理,以及地下生物多样性等方面的主要研究进展和亟待解决的问题,着重评述地下过程对全球变化响应的若干理论问题,指出地下生态学将是21世纪生态学的重要发展方向.
DOI:10.1073/pnas.94.14.7362URL [本文引用: 1]
DOI:10.5846/stxb201104280563URL [本文引用: 1]
以祁连山西水林区分布的棕钙土、灰褐土、栗钙土和高山草甸土为对象,研究 了阳离子交换量和盐基离子(K+、Na+、Ca2+、Mg2+)的剖面分布规律及其与土壤理化因子的关系.结果表明:土壤阳离子交换量(CEC,介于 4.80-48.10 cmol/kg)和盐基总量(TEB,介于4.67-21.34 cmol/kg)随剖面深度的增加逐渐减小,不同土壤类型的大小顺序为:灰褐土>高山草甸土>栗钙土>棕钙土;土壤盐基组成以Ca2+、Mg2+为主(占 TEB的比例平均为71.6%、22.9%),K+、Na+所占比例较低(占TEB的比例平均为3.3%、2.2%);棕钙土、灰褐土和栗钙土盐基离子的 剖面分布由浅至深呈现:K+≈ Ca2+> Na+≈Mg2+,高山草甸土盐基离子则呈现:K+>Na+>Mg2+ >Ca2+.不同土壤类型间盐基离子的含量及饱和度随发生层次不同存在较大差异.土壤有机质是CEC的主要贡献因素,粉粒对CEC也有显著的促进作用,而 砂粒、CaCO3对CEC有显著抑制作用.土壤生物复盐基作用弱于淋溶作用,造成盐基饱和度较大(BSP,介于44.4%-97.2%),并随剖面深度的 增加逐渐增大.相关性分析表明,土壤交换性Na+、Mg2+的含量及饱和度均呈极显著正相关,交换性Na+、Mg2+饱和度与CaCO3含量呈极显著正相 关;pH值与BSP呈极显著正相关;土壤速效P含量与CEC呈极显著正相关,速效K含量与交换性K+含量呈极显著正相关.
DOI:10.5846/stxb201104280563URL [本文引用: 1]
以祁连山西水林区分布的棕钙土、灰褐土、栗钙土和高山草甸土为对象,研究 了阳离子交换量和盐基离子(K+、Na+、Ca2+、Mg2+)的剖面分布规律及其与土壤理化因子的关系.结果表明:土壤阳离子交换量(CEC,介于 4.80-48.10 cmol/kg)和盐基总量(TEB,介于4.67-21.34 cmol/kg)随剖面深度的增加逐渐减小,不同土壤类型的大小顺序为:灰褐土>高山草甸土>栗钙土>棕钙土;土壤盐基组成以Ca2+、Mg2+为主(占 TEB的比例平均为71.6%、22.9%),K+、Na+所占比例较低(占TEB的比例平均为3.3%、2.2%);棕钙土、灰褐土和栗钙土盐基离子的 剖面分布由浅至深呈现:K+≈ Ca2+> Na+≈Mg2+,高山草甸土盐基离子则呈现:K+>Na+>Mg2+ >Ca2+.不同土壤类型间盐基离子的含量及饱和度随发生层次不同存在较大差异.土壤有机质是CEC的主要贡献因素,粉粒对CEC也有显著的促进作用,而 砂粒、CaCO3对CEC有显著抑制作用.土壤生物复盐基作用弱于淋溶作用,造成盐基饱和度较大(BSP,介于44.4%-97.2%),并随剖面深度的 增加逐渐增大.相关性分析表明,土壤交换性Na+、Mg2+的含量及饱和度均呈极显著正相关,交换性Na+、Mg2+饱和度与CaCO3含量呈极显著正相 关;pH值与BSP呈极显著正相关;土壤速效P含量与CEC呈极显著正相关,速效K含量与交换性K+含量呈极显著正相关.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/0933-3630(93)90024-9URL [本文引用: 1]
ABSTRACT The effect of drought on the biomass production of rainfed wheat was investigated along catenas in the semi-arid climatic zone of Greece. These catenas are located in hilly areas with rolling topography and soils formed on Tertiary and Quaternary deposits of marl, conglomerates and shale-sandstones. Total above ground biomass production was measured on specific hillslope components (shoulder, backslope and footslope) in three successive growing periods and was related to the soil properties, landscape position and climatic data. Crop-water use, calculated according to a broadly used simplified methodology, was logarithmically correlated with biomass production. Gravel and stones on the soil surface appeared to be extremely important in dry years by conserving appreciable amounts of soil water from evaporation through surface mulching and preventing large areas from desertification. Stony soils along slope catenas of conglomerates and shale-sandstones, despite their normally low productivity, may supply appreciable amounts of previously stored water to the stressed plants and to secure a not negligible biomass production even in extremely dry years. Soils formed on marl are free of coarse fragments and despite their considerable depth and high productivity in normal and wet years, they are very susceptible to desertification, being unable to support any vegetation in particularly dry years due to adverse soil physical properties and the absence of gravel and stone mulching.
DOI:10.1007/s11284-011-0869-5URL [本文引用: 2]
Selective sheep grazing in arid rangelands induces a decrease in total cover and grass cover and an increase in the dominance of shrubs. Both life forms differ in aboveground and belowground traits. We hypothesized that grazing disturbance leads to the replacement of grass by shrub fine roots in the upper soil, and this is reflected in changes in the seasonal dynamics of shallow fine roots at the community level. In two sites representative of non-grazed and grazed vegetation states in the Patagonian Monte, we assessed the canopy structure, and the fine root biomass, N concentration, production, and turnover during two consecutive years. The non-grazed site exhibited higher total, grass, and shrub cover than the grazed site. The grazed site had larger or equal fine root biomass than the non-grazed site except for late spring of the second year. This could be associated with the ability of shrubs to develop dimorphic-root systems occupying the soil freed by grasses at the grazed site, and with the larger contribution of grass than shrub fine roots in relation to an extraordinary precipitation event at the non-grazed site. This was consistent with the N concentration in fine roots. Fine root production was positively correlated to temperature at the grazed site and with precipitation at the non-grazed site. Fine root turnover did not differ between sites. Our results indicate that grazing leads to a shifting in the seasonality and main climatic controls of fine root production, while fine root turnover is mostly affected by changes in soil water conditions.
DOI:10.1007/s10533-011-9689-5URL [本文引用: 1]
Grasslands store substantial amounts of carbon in the form of organic matter in soil and roots. At high latitudes and elevation, turnover of these materials is slow due to various interacting biotic and abiotic constraints. Reliable estimates on the future of belowground carbon storage in cold grassland soils thus require quantitative understanding of these factors. We studied carbon turnover of roots, labile coarse particulate organic matter (cPOM) and older non-cPOM along a natural pH gradient (3.9鈥5.9) in a subalpine grassland by utilizing soil fractionation and radiocarbon dating. Soil carbon stocks and root biomass, turnover, and decomposability did not scale with soil pH whereas mean residence times of both soil organic matter fractions significantly increased with declining pH. The effect was twice as strong for non-cPOM, which was also stronger enriched in 15 N at low pH. Considering roots as important precursors for cPOM, the weaker soil pH effect on cPOM turnover may have been driven by comparably high root pH values. At pH < 5, long non-cPOM mean residence times were probably related to pH dependent changes in substrate availability. Differences in turnover along the pH gradient were not reflected in soil carbon stocks because aboveground productivity was lower under acidic conditions and, in turn, higher inputs from aboveground plant residues compensated for faster soil carbon turnover at less acidic pH. In summary, the study provides evidence for a strong and differential regulatory role of pH on the turnover of soil organic matter that needs consideration in studies aiming to quantify effects of changing environmental conditions on belowground carbon storage.
DOI:10.1371/journal.pone.0119184URLPMID:25734640 [本文引用: 3]
Abstract Root turnover is an important carbon flux component in grassland ecosystems because it replenishes substantial parts of carbon lost from soil via heterotrophic respiration and leaching. Among the various methods to estimate root turnover, the root's radiocarbon signature has rarely been applied to grassland soils previously, although the value of this approach is known from studies in forest soils. In this paper, we utilize the root's radiocarbon signatures, at 25 plots, in mountain grasslands of the montane to alpine zone of Europe. We place the results in context of a global data base on root turnover and discuss driving factors. Root turnover rates were similar to those of a subsample of the global data, comprising a similar temperature range, but measured with different approaches, indicating that the radiocarbon method gives reliable, plausible and comparable results. Root turnover rates (0.06-1.0 y-1) scaled significantly and exponentially with mean annual temperatures. Root turnover rates indicated no trend with soil depth. The temperature sensitivity was significantly higher in mountain grassland, compared to the global data set, suggesting additional factors influencing root turnover. Information on management intensity from the 25 plots reveals that root turnover may be accelerated under intensive and moderate management compared to low intensity or semi-natural conditions. Because management intensity, in the studied ecosystems, co-varied with temperature, estimates on root turnover, based on mean annual temperature alone, may be biased. A greater recognition of management as a driver for root dynamics is warranted when effects of climatic change on belowground carbon dynamics are studied in mountain grasslands.
DOI:10.11766/trxb200811190404URL [本文引用: 2]
通过人工改变土壤颗粒级配,并设置不同容重水平,测定土壤水分特征参数,研究了容重对土壤水分蓄持能力的定量影响。结果表明:(1)容重对土壤水分特征曲线、比水容量有较大影响,试验土壤各吸力段水分蓄持能力均随容重增大递减,比水容量也随容重增大递减。(2)容重对试验土壤饱和含水量、田间持水量、凋萎系数有较大影响,此3个水分参数均随容重增大递减。饱和含水量与容重呈幂函数负相关关系,田间持水量及凋萎系数均与容重呈指数负相关关系。(3)容重对试验土壤有效水、易效水、迟效水含量有较大影响,此3水分参数均随容重增大递减,分别与容重呈指数、幂函数、对数负相关关系。
DOI:10.11766/trxb200811190404URL [本文引用: 2]
通过人工改变土壤颗粒级配,并设置不同容重水平,测定土壤水分特征参数,研究了容重对土壤水分蓄持能力的定量影响。结果表明:(1)容重对土壤水分特征曲线、比水容量有较大影响,试验土壤各吸力段水分蓄持能力均随容重增大递减,比水容量也随容重增大递减。(2)容重对试验土壤饱和含水量、田间持水量、凋萎系数有较大影响,此3个水分参数均随容重增大递减。饱和含水量与容重呈幂函数负相关关系,田间持水量及凋萎系数均与容重呈指数负相关关系。(3)容重对试验土壤有效水、易效水、迟效水含量有较大影响,此3水分参数均随容重增大递减,分别与容重呈指数、幂函数、对数负相关关系。
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1000-2421.2000.04.010URL [本文引用: 1]
研究了湖北省郧县石渣土三种不同土石比土壤水分运动特性。结果表明:土壤蒸发、持水、导水能 力均与土石比大小有关。三种土壤的持水能力,以石砾含量较少质地偏粘的土壤最强,以石砾含量较多质地偏壤的土壤最低。而土壤水分蒸发能力与砾石含量呈反相 关,土壤水分扩散率则与砾石含量呈正相关。
DOI:10.3321/j.issn:1000-2421.2000.04.010URL [本文引用: 1]
研究了湖北省郧县石渣土三种不同土石比土壤水分运动特性。结果表明:土壤蒸发、持水、导水能 力均与土石比大小有关。三种土壤的持水能力,以石砾含量较少质地偏粘的土壤最强,以石砾含量较多质地偏壤的土壤最低。而土壤水分蒸发能力与砾石含量呈反相 关,土壤水分扩散率则与砾石含量呈正相关。
DOI:10.11821/yj2009040018URL [本文引用: 1]
采用静态暗箱法,比较测定了放牧对内蒙古锡林河流域羊草群落土壤呼吸的影响以及水热等相关环 境因子与土壤呼吸的关系。结果表明:放牧没有改变羊草群落土壤呼吸的季节性变化特征,但降低了土壤呼吸速率的年幅度;生长季放牧样地土壤呼吸速率显著低于 封育样地,非生长季两样地土壤呼吸强度均处于较低水平,而且出现负通量的现象,放牧使羊草群落土壤呼吸年总量下降了约33.95%;从全年来看,无论是围 栏还是放牧样地,封育样地和放牧样地土壤呼吸与温度因子均显著正相关(p〈0.01,n=15),其中与10cm处地温相关性最好,但放牧降低了土壤呼吸 对温度变化的敏感性;生长季水分影响作用高于温度,围栏封育样地0~10cm土壤含水量的变化可以解释土壤呼吸变异的87.4%,放牧样地10~20cm 和20~30cm土壤含水量的变化共同可以解释土壤呼吸变异的74.9%。
DOI:10.11821/yj2009040018URL [本文引用: 1]
采用静态暗箱法,比较测定了放牧对内蒙古锡林河流域羊草群落土壤呼吸的影响以及水热等相关环 境因子与土壤呼吸的关系。结果表明:放牧没有改变羊草群落土壤呼吸的季节性变化特征,但降低了土壤呼吸速率的年幅度;生长季放牧样地土壤呼吸速率显著低于 封育样地,非生长季两样地土壤呼吸强度均处于较低水平,而且出现负通量的现象,放牧使羊草群落土壤呼吸年总量下降了约33.95%;从全年来看,无论是围 栏还是放牧样地,封育样地和放牧样地土壤呼吸与温度因子均显著正相关(p〈0.01,n=15),其中与10cm处地温相关性最好,但放牧降低了土壤呼吸 对温度变化的敏感性;生长季水分影响作用高于温度,围栏封育样地0~10cm土壤含水量的变化可以解释土壤呼吸变异的87.4%,放牧样地10~20cm 和20~30cm土壤含水量的变化共同可以解释土壤呼吸变异的74.9%。
DOI:10.1007/s10021-009-9295-8URL [本文引用: 2]
Estimating root production has been difficult due to multiple potential biases associated with both old and new methods. This shortgrass steppe site is the only place we are aware of that can compare most methods including sequential coring, ingrowth cores, and ingrowth donuts, 14 C pulse-isotope dilution, 14 C pulse-isotope turnover, rhizotron windows, and minirhizotron, and indirect methods including nitrogen budget, carbon flux, simulation carbon flow model, and regression model. We used the studies at this site, other comparisons, a summary of potential directional biases, and different ways of calculating estimates in a logical, comparative approach of evaluating methods. Much of the literature for root production is based on sequential biomass coring, a method resulting in erroneous estimates. Root ingrowth estimates of production are generally conservative compared to minirhizotron and isotope turnover methods. The size of the ingrowth area may be the most important determinant of the underestimation. Estimates based on pulse-isotope dilution are also erroneous due to non-uniform labeling of tissues. Uniform labeling is not an assumption of the pulse-isotope turnover method, and this method has the least severe potential biases. Root production estimates from pulse-isotope turnover were lower than those using minirhizotron when the most common method of calculation was used. This agrees with literature concerning bomb 14 C continuous-isotope labeling comparisons with minirhizotron, although some potential biases between isotope methods are different. However, good agreement between pulse-isotope turnover and minirhizotron were obtained when minirhizotron estimates were calculated from regression of decomposition versus production to equilibrium and when pulse-isotope turnover estimates were calculated from two-phase life-span regressions. This minirhizotron method bypasses biases associated with the artificial surface similar to root-cohort methods that may be practical only in mesic systems, and takes into account both short- and long-lived roots and corrects for soil-isotope contamination that the continuous-isotope labeling bomb 14 C method is not able to account for. Comparisons of these direct methods are also made with four indirect methods.
DOI:10.1007/BF02197492URL [本文引用: 1]
DOI:10.1023/B:VEGE.0000049097.85960.10URL [本文引用: 3]
Data on field biomass measurements in temperate grasslands of northern China (141 samples from 74 sites) were obtained from 23 Chinese journals, reports and books. Net primary productivity (NPP) of grasslands was estimated using three algorithms (peak live biomass, peak standing crop and maximum minus minimum live biomass), respectively, based on availability of biomass data in sites. 135 samples which have aboveground biomass (AGB) measurements, have peak AGB ranges from 20 to 2021 g m –2 (mean = 325.3) and the aboveground NPP (ANPP) ranges from 15 to 1647.1 g m –2 per year (mean = 295.7). 72 samples which have belowground biomass (BGB) measurements, have peak BGB ranges from 226.5 to 12827.5 g m –2 (mean = 3116) and the belowground NPP (BNPP) ranges from 15.8 to 12827.5 g m –2 per year (mean = 2425.6). In total 66 samples have the total NPP (TNPP), ranging from 55.3 to 13347.8 g m –2 per year (mean = 2980.3). Mean peak biomass and NPP varied from different geographical sampling locations, but they had a general rough regularity in ten grasslands. Meadow, mountain and alpine grasslands had high biomass and NPP (sometimes including saline grassland). Forested steppe, saline grassland and desert had median values. Meadowed and typical steppes had low biomass and NPP (sometimes including desert). The lowest biomass and NPP occurred in deserted steppe and stepped desert. Grassland ANPP has significant positive relationships with annual and summer precipitation as well as summer temperature (all p<0.01). However, grassland BNPP and TNPP have more significant negative relationships with summer temperature (p<0.01) than with annual temperature (p<0.05). The analysis of climate – productivity correlations implied that aboveground productivity is more controlled by rainfall, whereas belowground and total productivity is more influenced by temperature in the temperate grasslands of northern China. The present study might underestimate grassland NPP in northern China due to limitation of biomass measurements. Data on relative long-term aboveground and belowground biomass dynamics, as well as data of standing dead matter, litterfall, decomposition and turnover, are required if grassland NPP is to be more accurately estimated and the role of temperate grasslands in the regional to global carbon cycles is to be fully appreciated.
DOI:10.2307/1931937URL [本文引用: 2]
Net primary production during the first 7 years of secondary succession was studied on abandoned farmland on the Savannah River Plant reservation. Annual and biennial forbs produced most of the organic matter during the first 5 years, with Leptilon canariense, Haplopappus divaricatus and Heterotheca subaxillaris as major dominants on well-drained soil. The perennial grass, Andropogon spp., did ...
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/geb.12508URL [本文引用: 1]
Abstract Root production and turnover play a key role in regulating carbon (C) flow in terrestrial ecosystems. However, a general pattern reflecting the responses of roots to increasing nitrogen (N) input has yet to be described. Global terrestrial ecosystems. We conducted a meta-analysis to assess the central tendencies of root production, turnover rate and standing crop with respect to the experimental addition of N. We evaluated the effect of the form of N, root diameter and climatic (mean annual temperature, MAT; mean annual precipitation, MAP), biotic (ecosystem type, plant type and forest stand age) and forcing factors (experimental duration, N addition rate and cumulative amount of N) on the variations in root response. Globally, the addition of N significantly decreased root production and turnover rate but had only a minor impact on root standing crop. In different ecosystems, the three root variables exhibited heterogeneous responses to N enrichment. Additionally, root production and turnover rate responded distinctly to diverse forms of N. The responses of root production and turnover rate to the addition of N were generally positively correlated with MAT and MAP but negatively related to forest stand age and experimental duration. The response pattern of root standing crop was negatively affected by MAT, MAP and forest stand age. However, none of the three root parameters had any obvious correlations with N addition rate or cumulative amount of N. Our results demonstrate that, on aggregate, the addition of N decreased root production and turnover rate at the global scale. These root response patterns and the regulatory factors can be incorporated into earth system models to improve the prediction of belowground C dynamics.
DOI:10.1093/oxfordjournals.aob.a084919URL [本文引用: 1]
DOI:10.2307/1941808URLPMID:27755662 [本文引用: 1]
Abstract Plant respiration is a large, environmentally sensitive component of the ecosystem carbon balance, and net ecosystem carbon flux will change as the balance between photosynthesis and respiration changes. Partitioning respiration into the functional components of construction, maintenance, and ion uptake will aid the estimation of plant respiration for ecosystems. Maintenance respiration is the component most sensitive to changes in temperature, CO 2 , protein concentration and turnover, water stress, and atmospheric pollutants. For a wide variety of plant tissues, maintenance respiration, corrected for temperature, appears to be linearly related to Kjeldahl nitrogen content of live tissue. Total and maintenance respiration may decline under CO 2 enrichment, but the mechanism, independence from changes in protein content, and acclimation are unknown. Response of respiration to temperature can be modelled as a Q 1 0 relationship, if corrections for bias arising from daily and annual temperature amplitude are applied. Occurrence and control of the cyanide-resistant respiratory pathway and acclimation of respiration rates to different climates are poorly understood, but may substantially affect the reliability of model estimates of plant respiration. 1991 by the Ecological Society of America.
DOI:10.1023/A:1026531420580URL [本文引用: 2]
Three grasses ( Holcus lanatus, Anthoxanthum odoratum and Festuca ovina ) and three herbs ( Rumex obtusifolius, Plantago lanceolata and Hieracium pilosella ) were grown in a greenhouse at 3 nutrient levels in order to evaluate plant allocation, architecture and biomass turnover in relation to fertility level of their habitats.Four harvests were done at intervals of 4 weeks. Various plant traits related to biomass partitioning, plant architecture, biomass turnover and performance were determined. Differences in nutrient supply induced a strong functional response in the species shoot:root allocation, but architecture and turnover showed little or no response. Architectural parameters like specific leaf area and specific root length, however, in general decreased during plant development.Species from more nutrient-rich successional stages were characterized by a larger specific leaf area and longer specific shoot height (height/shoot biomass), resulting in a higher RGR and total biomass in all nutrient conditions. There was no evidence that species from nutrient-poor environments had a longer specific root length or any other superior growth characteristic. The only advantage displayed by these species was a lower leaf turnover when expressed as the fraction of dead leaves and a shorter specific shoot height (SSH) which might prevent herbivory and mowing losses.The dead leaf fraction, which is a good indicator for biomass and nutrient loss, appeared to be not only determined by the leaf longevity, but was also found to be directly related to the RGR of the species. This new fact might explain the slow relative growth rates in species from a nutrient-poor habitat and should be considered in future discussions about turnover.
DOI:10.2307/1934970URL [本文引用: 1]
The wet tundra near Barrow, Alaska, is dominated by three species of graminoids: Dupontia fischeri, Carex aquatilis, and Eriophorum angustifolium. Root production, root turnover, and root distribution patterns of these three species were studied by direct observations of growing roots and by analysis of whole, interconnected tiller systems dug from the soil. Root weight per unit length and density of individual tillers were also measured in the field. Production of new roots was found to be strongly correlated with age of individual tillers, each species having a distinctive pattern and phenology. Root turnover rates also varied considerably; the range is from an annual turnover in E. angustifolium to 6-8(10) yr in C. aquatis. An estimated of root turnover on an ecosystem basis is about 100 g @? m^-^2 @? yr^-^2, or 25% of the live root biomass. Species with the shallowest and longest lived roots have the greatest weight per unit length of root, and vice versa. Each species has a characteristic root distribution pattern with depth and in relation to the progress of soil thaw.
DOI:10.3321/j.issn:1000-0933.2008.12.037URL [本文引用: 1]
为了解山地土壤中的砾石对土壤水文循环的影响,测定了砾石土壤的贮水能力、渗透速率和蒸发速率。结果表明,砾石含量与土壤有效贮水量呈正相关关系,但与土壤最大贮水量无明显相关。在0-40cm土层,当砾石体积含量小于15%~20%时,稳渗速率随砾石含量增加而增大,当砾石体积含量大于15%-20%时,土壤稳渗速率随砾石含量增加而减小;在40cm以下土层,稳渗速率随砾石含量增加而提高。在土壤砾石含量为0~20%时,土壤蒸发速率随砾石含量增加而降低,但在砾石含量超过20%时,土壤蒸发速率基本保持稳定。土壤蒸发速率随砾石粒径增大有升高的趋势。
DOI:10.3321/j.issn:1000-0933.2008.12.037URL [本文引用: 1]
为了解山地土壤中的砾石对土壤水文循环的影响,测定了砾石土壤的贮水能力、渗透速率和蒸发速率。结果表明,砾石含量与土壤有效贮水量呈正相关关系,但与土壤最大贮水量无明显相关。在0-40cm土层,当砾石体积含量小于15%~20%时,稳渗速率随砾石含量增加而增大,当砾石体积含量大于15%-20%时,土壤稳渗速率随砾石含量增加而减小;在40cm以下土层,稳渗速率随砾石含量增加而提高。在土壤砾石含量为0~20%时,土壤蒸发速率随砾石含量增加而降低,但在砾石含量超过20%时,土壤蒸发速率基本保持稳定。土壤蒸发速率随砾石粒径增大有升高的趋势。
DOI:10.1038/38260URL [本文引用: 1]
A large source of uncertainty in present understanding of the global carbon cycle is the distribution and dynamics of the soil organic carbon reservoir. Most of the organic carbon in soils is degraded to inorganic forms slowly, on timescales from centuries to millennia. Soil minerals are known to play a stabilizing role, but how spatial and temporal variation in soil mineralogy controls the quantity and turnover of long-residence-time organic carbon is not well known. Here we use radiocarbon analyses to explore interactions between soil mineralogy and soil organic carbon along two natural gradients-of soil-age and of climate-in volcanic soil environments. During the first ~150,000 years of soil development, the volcanic parent material weathered to metastable, non-crystalline minerals. Thereafter, the amount of non-crystalline minerals declined, and more stable crystalline minerals accumulated. Soil organic carbon content followed a similar trend, accumulating to a maximum after 150,000 years, and then decreasing by 50% over the next four million years. A positive relationship between non-crystalline minerals and organic carbon was also observed in soils through the climate gradient, indicating that the accumulation and subsequent loss of organic matter were largely driven by changes in the millennial scale cycling of mineral-stabilized carbon, rather than by changes in the amount of fast-cycling organic matter or in net primary productivity. Soil mineralogy is therefore important in determining the quantity of organic carbon stored in soil, its turnover time, and atmosphere-ecosystem carbon fluxes during long-term soil development; this conclusion should be generalizable at least to other humid environments.
DOI:10.1126/science.1091841URL [本文引用: 1]
DOI:10.1016/S0065-2504(08)60122-1URL [本文引用: 1]
DOI:10.1111/j.1365-2486.2007.01321.xURL [本文引用: 1]
Abstract We investigated seasonal root production and root turnover of fertilized and well-watered monocultures of Phalaris for 2 years using minirhizotrons installed in six newly designed temperature gradient tunnels, combined with sequential soil coring. Elevated atmospheric CO 2 treatments were combined with two cutting frequencies and three warming scenarios: no warming, +3.0/+3.0 and +2.2/+4.0 C (day/night) atmospheric warming. The elevated CO 2 treatment increased both new and net root length production primarily when combined with atmospheric warming, where the constant warming treatment had a greater positive effect than the increased night-time warming treatment. Responses to elevated CO 2 were greater when the swards were cut more frequently and responsiveness varied with season. For Phalaris swards, 17% of total net primary productivity went belowground. On account of root turnover, only one-third of the new roots produced in the year following establishment could be expected, on average, to be recovered from soil cores. The interaction between the effects of CO 2 and warming, combined with the differential effects of the two warming treatments, has important implications for modelling belowground responses to projected climate change.
DOI:10.3321/j.issn:1000-0933.2004.11.018URL [本文引用: 1]
土壤氮素的矿化是反映土壤供氮能力的重要因素之一,也是目前国内外研究的热点.通过测定内蒙古典型羊草草 原自由放牧地土壤净氮矿化量和净氮矿化速率,揭示影响草地生态系统土壤氮循环过程的有关机理,为草地生态系统建模提供理论依据.在实验室条件下,运用恒温 恒湿培养箱控制土壤的温度与湿度,测定羊草草原长期自由放牧地土壤氮素矿化量的积累.将不同水分含量的土柱分别放在温度为-10℃、0℃、5℃、15℃、 25℃和35℃的恒温恒湿培养箱中培养,培养1、2、3、5周后取出,分析培养前后的NH+4-N和 NO-3-N含量,以确定土壤净氮矿化(NH+4-N+NO-3-N)的累积和不同时间段内的矿化速率.结果表明:不同处理温度和水分之间的差异均达到显 著水平(p<0.0001).温度和水分之间具有显著的交互作用(p<0.0001).随着培养时间的延长,矿化氮累积量增加,但是矿化速率 下降.
DOI:10.3321/j.issn:1000-0933.2004.11.018URL [本文引用: 1]
土壤氮素的矿化是反映土壤供氮能力的重要因素之一,也是目前国内外研究的热点.通过测定内蒙古典型羊草草 原自由放牧地土壤净氮矿化量和净氮矿化速率,揭示影响草地生态系统土壤氮循环过程的有关机理,为草地生态系统建模提供理论依据.在实验室条件下,运用恒温 恒湿培养箱控制土壤的温度与湿度,测定羊草草原长期自由放牧地土壤氮素矿化量的积累.将不同水分含量的土柱分别放在温度为-10℃、0℃、5℃、15℃、 25℃和35℃的恒温恒湿培养箱中培养,培养1、2、3、5周后取出,分析培养前后的NH+4-N和 NO-3-N含量,以确定土壤净氮矿化(NH+4-N+NO-3-N)的累积和不同时间段内的矿化速率.结果表明:不同处理温度和水分之间的差异均达到显 著水平(p<0.0001).温度和水分之间具有显著的交互作用(p<0.0001).随着培养时间的延长,矿化氮累积量增加,但是矿化速率 下降.
DOI:10.1300/J079v30n03_01URL
对不同海拔梯度高寒草甸群落植物多样性和初级生产力关系的研究结果表明 :1 )不同海拔梯度上 ,中间海拔梯度群落植物多样性最高 ,即物种丰富度、均匀度和多样性最大 ;2 )不同海拔梯度上 ,群落生产力水平和物种丰富度中等时 ,物种多样性最高 ;3)随着海拔的逐渐升高 ,地上生物量逐渐减少 ;4 )地下生物量具有“V”字形季节变化规律 ,在牧草返青期和枯黄期地下生物量最大 ,7月最小 ,且地下生物量主要分布在 0~ 1 0cm的土层中。地下生物量垂直分布呈明显的倒金字塔特征
DOI:10.1300/J079v30n03_01URL
对不同海拔梯度高寒草甸群落植物多样性和初级生产力关系的研究结果表明 :1 )不同海拔梯度上 ,中间海拔梯度群落植物多样性最高 ,即物种丰富度、均匀度和多样性最大 ;2 )不同海拔梯度上 ,群落生产力水平和物种丰富度中等时 ,物种多样性最高 ;3)随着海拔的逐渐升高 ,地上生物量逐渐减少 ;4 )地下生物量具有“V”字形季节变化规律 ,在牧草返青期和枯黄期地下生物量最大 ,7月最小 ,且地下生物量主要分布在 0~ 1 0cm的土层中。地下生物量垂直分布呈明显的倒金字塔特征
DOI:10.3321/j.issn:1009-2242.2007.03.003URL [本文引用: 1]
农田化学物质随地表径流迁移的问题,既是农业问题,又是水环境问题。地表土壤容重是影响土壤溶质随径流迁移的重要因素之一。通过室内模拟试验,研究了降雨条件下土壤容重对黄土区坡面土壤氮、磷和钾随径流迁移过程的影响。结果表明,随着土壤容重增大,初始产流时间提早,径流系数增大,土壤流失量增多,土壤磷和钾流失量也随之增大。径流养分浓度变化对土壤容重响应程度的大小次序是:NO3--N>PO43--P>K+。径流养分流失率过程曲线存在“峰值点”,它是土壤容重、溶质理化特性以及土壤水文参数综合作用的结果。通过数学模拟,进一步证实了幂函数是模拟黄土区非饱和水流条件下养分流失过程的最佳模型。本研究为深入理解黄土坡面径流溶质迁移机理提供了参考。
DOI:10.3321/j.issn:1009-2242.2007.03.003URL [本文引用: 1]
农田化学物质随地表径流迁移的问题,既是农业问题,又是水环境问题。地表土壤容重是影响土壤溶质随径流迁移的重要因素之一。通过室内模拟试验,研究了降雨条件下土壤容重对黄土区坡面土壤氮、磷和钾随径流迁移过程的影响。结果表明,随着土壤容重增大,初始产流时间提早,径流系数增大,土壤流失量增多,土壤磷和钾流失量也随之增大。径流养分浓度变化对土壤容重响应程度的大小次序是:NO3--N>PO43--P>K+。径流养分流失率过程曲线存在“峰值点”,它是土壤容重、溶质理化特性以及土壤水文参数综合作用的结果。通过数学模拟,进一步证实了幂函数是模拟黄土区非饱和水流条件下养分流失过程的最佳模型。本研究为深入理解黄土坡面径流溶质迁移机理提供了参考。
URL [本文引用: 1]
通过收集我国近20年来草地生物量的有关文献,估算了我国北方天然草地根冠比(R/S)及地下生产力占总生产力比例(fBNPP)的大小及其对气候变化的响应。结果表明:不同草地类型R/S比及fBNPP变异较大,根冠比为1.66~15.21,fBNPP为0.29~0.98;荒漠草原的R/S比及fBNPP较大,但变幅较小;森林草原R/S比和fBNPP较小,但变幅较大;R/S比及fBNPP随年降水的增加而显著降低,随年平均气温增加而降低的趋势不明显。采用生长季最大生物量估算的方法可能高估了R/S比fBNPP。长期、高质量的生物量观测数据,尤其是地下生物量的数据以及开展不同研究方法对生产力估算结果的影响,对于准确评价草地在区域及全球碳循环中的作用是十分必要的。
URL [本文引用: 1]
通过收集我国近20年来草地生物量的有关文献,估算了我国北方天然草地根冠比(R/S)及地下生产力占总生产力比例(fBNPP)的大小及其对气候变化的响应。结果表明:不同草地类型R/S比及fBNPP变异较大,根冠比为1.66~15.21,fBNPP为0.29~0.98;荒漠草原的R/S比及fBNPP较大,但变幅较小;森林草原R/S比和fBNPP较小,但变幅较大;R/S比及fBNPP随年降水的增加而显著降低,随年平均气温增加而降低的趋势不明显。采用生长季最大生物量估算的方法可能高估了R/S比fBNPP。长期、高质量的生物量观测数据,尤其是地下生物量的数据以及开展不同研究方法对生产力估算结果的影响,对于准确评价草地在区域及全球碳循环中的作用是十分必要的。
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.5846/stxb201307031831URL [本文引用: 2]
植物根系是陆地生态系统重要的碳汇和养分库,细根周转过程是陆地生态系统地下部分碳氮循环的核心环节,在陆地生态系统如何响应全球变化中起着关键作用。在全球变化敏感地区之一的青藏高原,对该地区的主要植被类型矮嵩草草甸同时采用根钻法、内生长袋法和微根管法3种观测方法研究细根生产和周转速率,并探讨了极差法、积分法、矩阵法和Kaplan-Meier法等数据处理方法对计算值的影响。研究结果显示:在估算细根净初级生产力时,根钻法宜采用积分法,内生长袋法宜选用矩阵法;由此进一步以最大细根生物量为基础,根钻法和内生长袋法估测的细根年周转速率分别为0.36 a-1和0.52 a-1,内生长袋法的估算结果是根钻法的1.44倍。对于微根管法,将其观测得到的细根长度转换为单位面积的生物量值后,采用积分法计算出细根周转速率为0.84 a-1,远高于传统方法的估算结果;若采用Kaplan-Meier生存分析方法,则计算出的细根周转速率更高达3.41 a-1。
DOI:10.5846/stxb201307031831URL [本文引用: 2]
植物根系是陆地生态系统重要的碳汇和养分库,细根周转过程是陆地生态系统地下部分碳氮循环的核心环节,在陆地生态系统如何响应全球变化中起着关键作用。在全球变化敏感地区之一的青藏高原,对该地区的主要植被类型矮嵩草草甸同时采用根钻法、内生长袋法和微根管法3种观测方法研究细根生产和周转速率,并探讨了极差法、积分法、矩阵法和Kaplan-Meier法等数据处理方法对计算值的影响。研究结果显示:在估算细根净初级生产力时,根钻法宜采用积分法,内生长袋法宜选用矩阵法;由此进一步以最大细根生物量为基础,根钻法和内生长袋法估测的细根年周转速率分别为0.36 a-1和0.52 a-1,内生长袋法的估算结果是根钻法的1.44倍。对于微根管法,将其观测得到的细根长度转换为单位面积的生物量值后,采用积分法计算出细根周转速率为0.84 a-1,远高于传统方法的估算结果;若采用Kaplan-Meier生存分析方法,则计算出的细根周转速率更高达3.41 a-1。
URL [本文引用: 1]
在生物量的研究过程中,地下部分生物量十分重要却又多被忽视本文首先比较了当前地下生物量研 究中较为常用的4种研究方法:挖土块法、钻土芯法、内生长土芯法和微根区管法.然后总结了地下生物量在空间和时间上变化规律的研究成果.最后又讨论了环境 因素诸如水分、温度和开垦、放牧对地下生物量的影响以及地下生物量的周转.
URL [本文引用: 1]
在生物量的研究过程中,地下部分生物量十分重要却又多被忽视本文首先比较了当前地下生物量研 究中较为常用的4种研究方法:挖土块法、钻土芯法、内生长土芯法和微根区管法.然后总结了地下生物量在空间和时间上变化规律的研究成果.最后又讨论了环境 因素诸如水分、温度和开垦、放牧对地下生物量的影响以及地下生物量的周转.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Fine root turnover in forest ecosystems in relation to quantity and form of nitrogen availability: A comparison of two methods
1
1985
... id="C8">对于草地生态系统, 周转的概念最早是由
Increased quantity and quality of coarse soil organic matter fraction at elevated CO2 in a grazed grassland are a consequence of enhanced root growth rate and turnover
1
2005
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling
6
2010
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... )草甸净初级生产力沿降雨及海拔梯度的分布规律进行了一系列研究.
... 利用微根管法研究了氮添加对内蒙古典型草原羊草群落根系寿命的影响, 结果表明, 氮添加可以明显缩短夏季出生的羊草根系寿命.
... id="C27">沿平均年降水梯度, 根系周转呈现出线性增加的趋势.和温度升高带来的影响类似, 降水的增加提高了土壤湿度, 也会提高维持呼吸速率和土壤有机物的矿化速率, 进而加快根系周转.局地尺度上的控制实验也证实, 增水可以提高草地生态系统的根系周转速率(
... id="C36">本研究中根系周转随温度升高而加快, 该结果与全球尺度(
... ).由此可见, 温度对根系周转的影响具有很强的尺度依赖性.对降水而言, 本研究中根系周转速率和降水之间呈现显著的正相关关系, 与
Spatial and temporal effects of nitrogen addition on root life span of
1
2008
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
Ecosystem stability and compensatory effects in the Inner Mongolia grassland
1
2004
... id="C31">土壤质地是土壤不同粒径矿物颗粒组成的综合反映, 与土壤通气状况、保水保肥能力密切相关.细质土中根系周转速率最高, 可能是由于细质土中黏粒所占比重较大, 土壤保水能力较好, 而降水(尤其是生长季降水)是影响草地生态系统生产力的主要限制因子(
Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests
1
2000
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
内蒙古草甸草原与典型草原地下生物量与生产力季节动态及其碳库潜力
1
2014
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
内蒙古草甸草原与典型草原地下生物量与生产力季节动态及其碳库潜力
1
2014
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe
1
2013
... id="C33">本研究中土壤pH值介于4.9-8.4, 涵盖了弱酸性土、中性土和弱碱性土.研究认为, 弱酸性的土壤环境有利于土壤有机物矿化相关酶的活性(
砂田西瓜不同粒径砂砾石覆盖的水分效应研究
1
2005
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
砂田西瓜不同粒径砂砾石覆盖的水分效应研究
1
2005
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
内蒙古锡林河流域羊草草原与大针茅草原地下部分生产力和周转值的研究. 见: 中国科学院内蒙草原生态系统研究站主编
1
1988
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
内蒙古锡林河流域羊草草原与大针茅草原地下部分生产力和周转值的研究. 见: 中国科学院内蒙草原生态系统研究站主编
1
1988
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
Net primary production in tropical forests: An evaluation and synthesis of existing field data
1
2001
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
Temperature and moisture effects on microbial biomass and soil organic matter mineralization
1
2012
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
Root productivity and turnover in native prairie
4
1965
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
... id="C8">对于草地生态系统, 周转的概念最早是由
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... id="C12">为了避免不同测定方法的差异带来的影响, 本研究统一使用生物量法来计算根系周转速率, 对使用微根管法、同位素法或模型模拟方法获得的数据予以剔除.具体计算公式为: 根系周转速率=地下净初级生产力(BNPP)/地下生物量最大值(BGBmax), 单位为a-1 (
Plant clonal traits, coexistence and turnover in East Ladakh, Trans-Himalaya
2011
1
1996
... id="C11">中国是世界草地资源大国, 拥有草地总面积近4亿hm2, 草地面积约占国土总面积的41.7% (
1
1996
... id="C11">中国是世界草地资源大国, 拥有草地总面积近4亿hm2, 草地面积约占国土总面积的41.7% (
Root production is determined by radiation flux in a temperate grassland community
1
2004
... id="C36">本研究中根系周转随温度升高而加快, 该结果与全球尺度(
Building roots in a changing environment: Implications for root longevity
1
2000
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
The ecology of root lifespan
1
1997
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
内蒙古灌丛化石生针茅荒漠草原地下生物量与周转值的测定. 见: 中国科学院内蒙草原生态系统研究站主编
2
1990
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... id="C35">本研究中涉及的样点南北跨越22个纬度, 东西跨越46个经度, 覆盖范围较广, 5种不同草地类型是其所在地区植被、气候、地形、土壤等特性的综合反映.不同草地类型之间根系周转存在显著的差异, 说明气候、土壤等因子的不同组合对根系周转的影响结果不同.对于温带地区的荒漠草原而言, 降水是影响根系周转的主要限制因素(
内蒙古灌丛化石生针茅荒漠草原地下生物量与周转值的测定. 见: 中国科学院内蒙草原生态系统研究站主编
2
1990
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... id="C35">本研究中涉及的样点南北跨越22个纬度, 东西跨越46个经度, 覆盖范围较广, 5种不同草地类型是其所在地区植被、气候、地形、土壤等特性的综合反映.不同草地类型之间根系周转存在显著的差异, 说明气候、土壤等因子的不同组合对根系周转的影响结果不同.对于温带地区的荒漠草原而言, 降水是影响根系周转的主要限制因素(
Root production and turnover and carbon budgets of two contrasting grasslands under ambient and elevated atmospheric carbon dioxide concentrations
1
1997
... id="C8">对于草地生态系统, 周转的概念最早是由
放牧和刈割对草原地下净生产力和根系周转的影响
1
2017
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
放牧和刈割对草原地下净生产力和根系周转的影响
1
2017
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
Global patterns of root turnover for terrestrial ecosystems
11
2000
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
... id="C8">对于草地生态系统, 周转的概念最早是由
... 均没有显著的相关关系(
... ).
... 首次在全球尺度上整合已有的研究结果, 揭示根系周转的格局及主要驱动因子, 为之后的研究提供了重要思路.然而该研究也存在以下几方面的不足.首先, 在样点的分布上, 草地生态系统的样点主要分布在北美、非洲和印度, 而全球最大的草地分布区欧亚大陆草原的样点却非常稀少, 尤其是中国草地取样点数目为零.欧亚大陆草原独特的地理位置和气候条件决定了其异于北美和非洲草原的特性, 因而该区域数据的缺失必将严重影响全球数据的代表性和结果的可靠性.其次,
... ).所以方法上的不统一, 也使得
... 的研究结果存在很大的不确定性.另外, 在该研究中, 受数据提取途径的局限, 根系周转驱动因子只包含了气候因子, 而对于根系生长的直接载体——土壤的理化性质并未考虑, 所以土壤数据的缺失是
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
... id="C36">本研究中根系周转随温度升高而加快, 该结果与全球尺度(
... ;
... 在局地尺度上的控制实验结果一致.中国草地根系周转沿温度和降水梯度上的分布格局与全球尺度的研究结果存在较大的差异, 分析原因主要包括以下两方面: 一方面缘于尺度效应, 另一方面是
Influence of floristic composition on the net primary production and dry matter turnover in a tropical grassland
1
1982
... id="C8">对于草地生态系统, 周转的概念最早是由
Root dynamics of tallgrass prairie in wet and dry years
1
1987
... id="C8">对于草地生态系统, 周转的概念最早是由
全球变化下的地下生态学: 问题与展望
2
2004
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
全球变化下的地下生态学: 问题与展望
2
2004
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
A global budget for fine root biomass, surface area, and nutrient contents
1
1997
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
祁连山西水林区土壤阳离子交换量及盐基离子的剖面分布
1
2012
... id="C34">CEC是反映土壤保肥供肥性能的重要指标, 土壤速效磷含量与CEC呈显著正相关关系(
祁连山西水林区土壤阳离子交换量及盐基离子的剖面分布
1
2012
... id="C34">CEC是反映土壤保肥供肥性能的重要指标, 土壤速效磷含量与CEC呈显著正相关关系(
羊草草原群落和大叶针茅草原群落生物量的初步比较研究. 见: 中国科学院内蒙草原生态系统研究站主编
1
1985
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
羊草草原群落和大叶针茅草原群落生物量的初步比较研究. 见: 中国科学院内蒙草原生态系统研究站主编
1
1985
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
The impacts of parent material and landscape poison on drought and biomass production of wheat under semiarid conditions
1
1993
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
Production and turnover rates of shallow fine roots in rangelands of the Patagonian Monte, Argentina
2
2012
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
... id="C27">沿平均年降水梯度, 根系周转呈现出线性增加的趋势.和温度升高带来的影响类似, 降水的增加提高了土壤湿度, 也会提高维持呼吸速率和土壤有机物的矿化速率, 进而加快根系周转.局地尺度上的控制实验也证实, 增水可以提高草地生态系统的根系周转速率(
Control of soil pH on turnover of belowground organic matter in subalpine grassland
1
2013
... id="C33">本研究中土壤pH值介于4.9-8.4, 涵盖了弱酸性土、中性土和弱碱性土.研究认为, 弱酸性的土壤环境有利于土壤有机物矿化相关酶的活性(
Turnover of grassland roots in mountain ecosystems revealed by their radiocarbon signature: Role of temperature and management
3
2015
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
... id="C35">本研究中涉及的样点南北跨越22个纬度, 东西跨越46个经度, 覆盖范围较广, 5种不同草地类型是其所在地区植被、气候、地形、土壤等特性的综合反映.不同草地类型之间根系周转存在显著的差异, 说明气候、土壤等因子的不同组合对根系周转的影响结果不同.对于温带地区的荒漠草原而言, 降水是影响根系周转的主要限制因素(
... id="C36">本研究中根系周转随温度升高而加快, 该结果与全球尺度(
高寒草甸植物地下生物量与气象条件的关系及周转值分析
1998
高寒草甸植物地下生物量与气象条件的关系及周转值分析
1998
容重对土壤水分蓄持能力影响模拟试验研究
2
2010
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
... ;
容重对土壤水分蓄持能力影响模拟试验研究
2
2010
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
... ;
容重变化与土壤水分特征的依赖关系研究
1
2009
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
容重变化与土壤水分特征的依赖关系研究
1
2009
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
丹江口库区石渣土土壤水分特性研究
1
2000
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
丹江口库区石渣土土壤水分特性研究
1
2000
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
放牧对内蒙古羊草群落土壤呼吸的影响
1
2009
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
放牧对内蒙古羊草群落土壤呼吸的影响
1
2009
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
Estimating root production: Comparison of 11 methods in shortgrass steppe and review of biases
2
2009
... id="C8">对于草地生态系统, 周转的概念最早是由
... 收集的数据库中, 根系周转速率的计算除了传统的生物量法外, 还包含了微根管法、同位素法及模型模拟的方法.不同方法所依据的原理不同, 误差来源各异(
Root turnover and production by 14C dilution: Implications of carbon partitioning in plants
1
1985
... id="C8">对于草地生态系统, 周转的概念最早是由
Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China
3
2004
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... 整合了2000年以前发表的有关中国北方温性草地生物量或净初级生产力的文献, 从中提取到72组包含地下生物量或地下净初级生产力的数据, 发现群落净地下初级生产力与夏季(5-9月)平均气温呈显著的负相关关系.该项工作为进一步研究根系周转提供了基础, 同时
... id="C27">沿平均年降水梯度, 根系周转呈现出线性增加的趋势.和温度升高带来的影响类似, 降水的增加提高了土壤湿度, 也会提高维持呼吸速率和土壤有机物的矿化速率, 进而加快根系周转.局地尺度上的控制实验也证实, 增水可以提高草地生态系统的根系周转速率(
Organic production and turnover in old field succession
2
1960
... id="C8">对于草地生态系统, 周转的概念最早是由
... 在研究弃耕地生态系统的演替时提出的.
砾石对青藏高原土壤水热特性影响的数值模拟
1
2015
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
砾石对青藏高原土壤水热特性影响的数值模拟
1
2015
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
Global patterns of root dynamics under nitrogen enrichment
1
2017
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
The cost of maintenance process in plant cells
1
1975
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
Effects of climate change on plant respiration
1
1991
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
Biomass partitioning, architecture and turnover of six herbaceous species from habitats with different nutrient supply
2
2000
... id="C7">草地占地球陆地总面积的40.5% (不包格陵兰岛和南极)(
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
Root production and root turnover in a wet tundra ecosystem, Barrow, Alaska
1
1975
... id="C8">对于草地生态系统, 周转的概念最早是由
六盘山森林土壤中的砾石对渗透性和蒸发的影响
1
2008
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
六盘山森林土壤中的砾石对渗透性和蒸发的影响
1
2008
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
Mineral control of soil organic carbon storage and turnover
1
1997
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
The secret lives of roots
1
2003
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests
1
1986
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
Effects of elevated atmospheric CO2, cutting frequency, and differential day/night atmospheric warming on root growth and turnover of Phalaris swards
1
2007
... id="C9">近年来, 随着技术的发展及研究草地生态系统根系周转意义的凸显, 相关研究逐渐增多.如
温度和湿度对我国内蒙古羊草草原土壤净氮矿化的影响
1
2004
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
温度和湿度对我国内蒙古羊草草原土壤净氮矿化的影响
1
2004
... id="C26">根系周转速率随MAT的增加呈线性增加, 即南方温暖地区根系周转要比北方寒冷地区快, 可能有以下三方面的原因: (1)正常活细胞的细胞膜内外需要维持一定的离子浓度差, 并且更换已降解的蛋白, 而这些支持和修复功能需要借助维持呼吸来提供ATP, 随着温度的升高, 蛋白质及细胞膜脂的更新加快, 需要更强的呼吸作用来维持(
高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究
2004
高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究
2004
高寒矮嵩草草甸群落植物多样性和初级生产力对模拟降水的响应
2003
高寒矮嵩草草甸群落植物多样性和初级生产力对模拟降水的响应
2003
表层土壤容重对黄土坡面养分随径流迁移的影响
1
2007
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
表层土壤容重对黄土坡面养分随径流迁移的影响
1
2007
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
中国北方天然草地的生物量分配及其对气候的响应
1
2008
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
中国北方天然草地的生物量分配及其对气候的响应
1
2008
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
Factors influencing inter rill erosion from semiarid slopes in New Mexico
2
1988
... id="C29">已有研究表明, 砾石含量越高的土壤, 其导水率越大(
... ).而且砾石的存在可以避免由于降雨直接打击土壤而形成地表结皮(
容重对不同有机质含量土壤水分入渗特性的影响
1
2013
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
容重对不同有机质含量土壤水分入渗特性的影响
1
2013
... id="C30">一般认为, 土壤容重越大, 非毛管孔隙度、毛管孔隙度和总孔隙度就越小(
高寒草甸植被细根生产和周转的比较研究
2
2014
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... id="C12">为了避免不同测定方法的差异带来的影响, 本研究统一使用生物量法来计算根系周转速率, 对使用微根管法、同位素法或模型模拟方法获得的数据予以剔除.具体计算公式为: 根系周转速率=地下净初级生产力(BNPP)/地下生物量最大值(BGBmax), 单位为a-1 (
高寒草甸植被细根生产和周转的比较研究
2
2014
... id="C10">关于中国草地根系周转的研究, 早在20世纪80年代已经开展, 根系周转速率的计算普遍采用根系周转速率=净地下初级生产力/地下生物量最大值(
... id="C12">为了避免不同测定方法的差异带来的影响, 本研究统一使用生物量法来计算根系周转速率, 对使用微根管法、同位素法或模型模拟方法获得的数据予以剔除.具体计算公式为: 根系周转速率=地下净初级生产力(BNPP)/地下生物量最大值(BGBmax), 单位为a-1 (
植物地下生物量研究进展
1
2001
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
植物地下生物量研究进展
1
2001
... id="C6">根系作为植物体的重要器官, 能够吸收水分和营养物质并储存光合产物, 同时根系的凋亡也是土壤有机质的主要来源, 因此根系是联系植物地上部分和土壤的重要媒介.根系周转是指根系从产生到死亡并分解消失的过程.根系周转的快慢用根系周转速率来表征, 它等于一年中生物量的增加值与最大值之比, 单位为a-1 (
排水对若尔盖高原泥炭地土壤有机碳储量的影响
1
2016
... id="C31">土壤质地是土壤不同粒径矿物颗粒组成的综合反映, 与土壤通气状况、保水保肥能力密切相关.细质土中根系周转速率最高, 可能是由于细质土中黏粒所占比重较大, 土壤保水能力较好, 而降水(尤其是生长季降水)是影响草地生态系统生产力的主要限制因子(
排水对若尔盖高原泥炭地土壤有机碳储量的影响
1
2016
... id="C31">土壤质地是土壤不同粒径矿物颗粒组成的综合反映, 与土壤通气状况、保水保肥能力密切相关.细质土中根系周转速率最高, 可能是由于细质土中黏粒所占比重较大, 土壤保水能力较好, 而降水(尤其是生长季降水)是影响草地生态系统生产力的主要限制因子(
Grasslands of China. In: Coupland RT ed
1
1993
... id="C17">基于
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}