 ,*, 柴强*, 李瑞霞*, 赵丹*新疆农业大学草业与环境科学学院, 新疆土壤与植物生态过程重点实验室, 乌鲁木齐 830052
,*, 柴强*, 李瑞霞*, 赵丹*新疆农业大学草业与环境科学学院, 新疆土壤与植物生态过程重点实验室, 乌鲁木齐 830052Carbon and nitrogen traits of typical shrubs in grassland of northern Xinjiang, China
ZHANG Jing, LIU Yun-Hua, SHENG Jian-Dong,*, CHAI Qiang*, LI Rui-Xia*, ZHAO Dan*Xinjiang Key Laboratory of Soil and Plant Ecological Processes, College of Grassland and Environmental Sciences, Xinjiang Agricultural University, ürümqi 830052, China通讯作者:
编委: 白永飞
责任编辑: 王葳
| 基金资助: |
Online:2018-03-20
| Supported by: | SupportedbytheStrategicPriorityResearchProgramoftheChineseAcademyofSciences( |

摘要
关键词:
Abstract
Keywords:
PDF (1426KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
张静, 刘耘华, 盛建东, 柴强, 李瑞霞, 赵丹. 新疆北部草地典型灌木的碳氮特征. 植物生态学报, 2018, 42(3): 307-316 doi:10.17521/cjpe.2015.1086
ZHANG Jing, LIU Yun-Hua, SHENG Jian-Dong, CHAI Qiang, LI Rui-Xia, ZHAO Dan.
灌木在维持荒漠生态系统稳定中发挥着重要的作用(侯学煜, 1982), 它能扩大物种生产力来源, 增强生态稳定性(李新荣, 2000), 一般生长在干旱荒漠区、草原牧场地、盐碱地、森林区, 在这些生境中, 灌木的生长还存在着交叉和重叠(李清河等, 2006)。生长在草地的灌木多样性丰富, 且草地具有生态过渡的特殊性, 已有很多研究报道了在干旱半干旱地区草地生态系统逐渐被灌木生态系统取代的现象(Barth & Klemmedson, 1978; Woodell et al., 1986; Grover & Musick, 1990; Archer et al., 1995; van Auken, 2000), 这种现象主要是由全球气候变化和人为活动(过度放牧、火灾等)引起的(Knapp et al., 2008)。
灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(Nepstad et al., 1994; Boutton et al., 1998; van Auken, 2000; Jackson et al., 2000), 从而引起生态系统碳氮循环及其储量的变化(Brantley & Young, 2010), 有研究表明, 灌木能使生物量和土壤碳氮储量增加(Goodale & Davidson, 2002), 因此灌木具有碳増汇的作用(Eldridge et al., 2011)。以往在国家和区域尺度上计算草地固碳储量的研究中, 忽视了灌木的固碳贡献(Ni, 2002, 2004)。而且, 在对生物量碳库的研究中, 大多以45%作为植物的含碳量。但实际上, 灌木的C、N含量在物种之间和同一物种的不同器官之间是有显著差异的(Northup et al., 2005)。因此, 不同灌木及其不同器官的C、N含量是精确估算陆地生态系统碳和氮储量的基本参数。
新疆是典型的干旱半干旱地区, 是我国仅次于西藏和内蒙古的第三大草原省区, 新疆可利用草地面积约为4800.68万hm2 (许鹏等, 1993), 占新疆总面积的21.4%, 是维系新疆生态环境稳定的重要生态系统。灌木在新疆草地中具有防风固沙、水土保持的作用。本研究以新疆北部(简称北疆)为研究区域, 以主要的草地类型为基本单元, 调查了不同草地类中典型灌木物种, 并分析测定了每一物种不同器官(叶、枝和茎)的碳、氮含量以及碳氮比值, 以期为新疆草地的碳库和氮库的准确估算及其碳氮循环提供参数。
1 材料和方法
1.1 研究区域概况
研究区位于新疆维吾尔自治区天山北部, 包括准噶尔盆地、伊犁河谷、塔额盆地、阿勒泰山南麓及天山北坡等灌木分布区, 地理坐标为80.73°- 91.12° E , 42.85°-48.48° N, 海拔230-2456 m。北疆属于温带大陆性干旱半干旱气候区, 光照充足, 无霜期短, 年降水量较少, 为150-200 mm, 年蒸发量大, 年平均气温-4-9 ℃ (许鹏等, 1993)。草地类是具有相同水热大气候带特征和植被特征、具有独特地带性的草地, 依据热量和水分条件, 将草地划分为不同草地类(中华人民共和国农业部畜牧兽医司, 全国畜牧兽医总站, 1996), 北疆的草地类分别是低地草甸、山地草甸、温性草甸草原、温性草原化荒漠、温性草原、温性荒漠草原、温性荒漠, 以草地类为基本单元在北疆设置102个具有灌木的样地进行实地调查。通过调查发现, 北疆草地典型灌木有白刺(Nitraria spp.)、刺旋花(Convol- vulus tragacanthoides)、红砂(Reaumuria songarica)、假木贼(Anabasis spp.)、锦鸡儿(Caragana spp.)、麻黄(Ephedra spp.)、沙拐枣(Calligonum mongolicum)、梭梭(Haloxylon spp.)、金丝桃叶绣线菊(Spiraea hypericifolia)、驼绒藜(Krascheninnikovia spp.)、小蓬(Nanophyton erinaceum)、盐爪爪(Kalidium spp.)、猪毛菜(Salsola spp.)。将梭梭的同化枝作为叶进行统计分析, 麻黄的器官只有枝和茎。
1.2 样地设置
累计调查了102个样地。在每个样地选择100 m × 100 m的区域进行取样调查, 在其对角线上设置一条100 m样线, 在样线旁设置5个面积为5 m × 5 m的灌木样方。1.3 样品采集和处理
2011-2012年的8月(植物生长高峰期)进行调查采样, 记录样地的经纬度、海拔、植物群落等基本信息, 在灌木样方内, 将每一个灌木物种用剪刀齐地面剪下, 将每一株灌木的地上部分按叶、枝和茎分开, 装入信封, 并在信封上注明物种名称及器官类型, 将植物清洗后及时放入105 ℃烘箱杀青30 min后带回室内, 放进65 ℃烘箱烘至恒质量, 记录各器官烘干后的质量。将同一样地、同一样方、同一物种的植物叶、枝、茎混合后粉碎, 然后取一定量粗粉植物碎屑, 经冷冻混合球磨仪(Retsch MM400, RETSCH, Dusseldorf, Germany)粉碎后, 使用元素分析仪(Euro EA3000-Single, EuroVector, Milan, Italy)测定植物样品全碳和全氮含量。
1.4 数据处理
灌木地上部分碳(C)、氮(N)含量由生物量加权平均算出, 计算公式为:C灌木地上部分 (%) = ( m叶 × C叶 + m枝 × C枝 + m茎 × C茎) / (m叶 + m枝 + m茎);
N灌木地上部分 (%) = ( m叶 × N叶 + m枝 × N枝 + m茎 × N茎) / (m叶 + m枝 + m茎)。
其中, C代表C含量(%), N代表N含量(%), m代表不同器官的质量(g)。
数据使用SPSS 20.0进行单因素方差分析, 使用Origin 8.0制作频数分布图。
2 结果
2.1 北疆典型灌木各器官碳氮含量特征
将在北疆调查的灌木按照叶、枝、茎器官的碳氮含量统计结果可知(表1), 典型灌木各器官的C含量大小为茎(45.81 ± 3.18)% >枝(44.27 ± 4.51)% >叶(39.15 ± 5.91)%; 而N含量与C含量相反, 即叶(2.21 ± 0.59)% >枝(1.55 ± 0.44)% >茎(1.34 ± 0.35)%, 进而导致C:N在各器官的大小顺序与C含量的分配相同, 为茎(36.77 ± 10.78) >枝(31.07 ± 10.43) >叶 (18.94 ± 5.82)。C含量、N含量以及C:N在各器官之间的差异均极显著(p < 0.01)。Table 1
表1
表1C、N在灌木叶、枝、茎器官中的含量(平均值±标准偏差)
Table 1
| 器官 Organ | 碳含量 C content (%) | 氮含量 N content (%) | C:N |
|---|---|---|---|
| 叶 Leaves | 39.15 ± 5.91c | 2.21 ± 0.59a | 18.94 ± 5.82c |
| 枝 Branches | 44.27 ± 4.51b | 1.55 ± 0.44b | 31.07 ± 10.43b |
| 茎 Stems | 45.81 ± 3.18a | 1.34 ± 0.35c | 36.77 ± 10.78a |
新窗口打开|下载CSV
北疆草地典型灌木叶的C、N含量和C:N变化范围分别为21.21%-51.00%、0.92%-3.75%和8.41- 40.91, 平均值为39.15%、2.21%和18.94, 变异系数为15%、27%和31%, 枝的C、N含量和C:N普遍比叶的高, 其变化范围为21.25%-57.25%、0.58%-3.35%和12.44-79.98, 平均值为44.27%、1.55%和31.07, 变异系数为10%、28%和34%; 茎C、N含量和C:N与枝相似, 其变化范围为31.88%-62.24%、0.53%-3.13% 和13.16-91.67, 平均值为45.81%、1.34%和36.77, 变异系数为7%、26%和29%。
由频数分布图(图1)还可看出, 灌木叶的C含量多集中在40%以下(占叶总样本数的56.6%), 枝和茎的C含量多集中在45%-50%之间(分别占枝、茎总样本数的48.5%和59.8%); 灌木叶的N含量多集中在2.0%以上(占叶总样本数的62%), 枝和茎的N含量多集中在1.0%-1.5%之间(分别占枝、茎总样本数的40.1%和57.2%); 叶的C:N多集中在20以下(占叶总样本数的64%), 枝和茎的C:N多集中在35以上(分别占枝、茎总样本数的44.1%和49.4%)。
图1
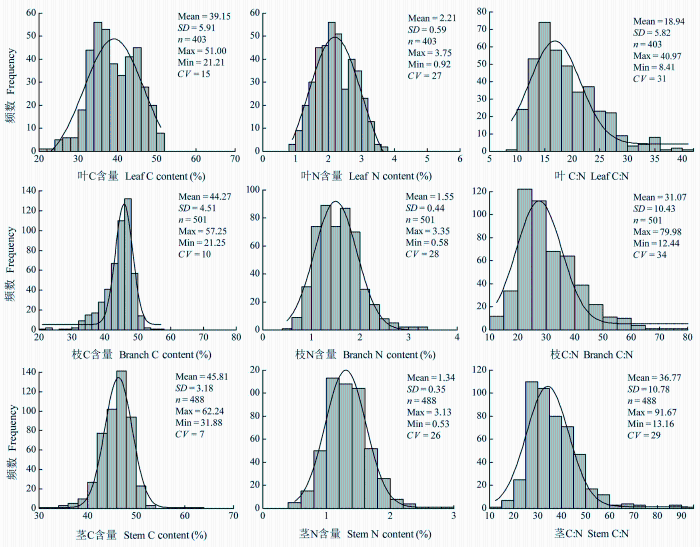 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1叶、枝、茎中C、N含量及C:N的频数分布图。
Fig. 1Frequency distributions of the C and N contents and C:N in leaves, branches and stem. CV, coefficient of variation; SD, standard deviation.
2.2 不同物种(属)灌木碳氮特征
从表2可看出, 北疆草地13种典型灌木中, 叶的C含量最高的是金丝桃叶绣线菊, 为(46.60 ± 2.86)%, 最低的是小蓬, 为(31.97 ± 5.37)%; 叶的N含量最高的是锦鸡儿, 为(2.66 ± 0.48)%, 最低的是刺旋花, 为(1.57 ± 0.48)% ; 叶的C:N最高的是刺旋花, 为(27.82 ± 8.63), 最低的是红砂, 为(14.92 ± 3.24)。Table 2
表2
表2北疆草地典型灌木叶、枝、茎C、N含量(平均值±标准偏差)
Table 2
| 物种 Species | 科 Family | 器官 Organ | n | 碳含量 C content (%) | CV | 氮含量 N content (%) | CV | C:N | CV |
|---|---|---|---|---|---|---|---|---|---|
| 白刺 | 蒺藜科 | 叶 Leaves | 12 | 38.40 ± 4.07 | 11 | 2.42 ± 0.47 | 20 | 16.40 ± 3.57 | 22 |
| Nitraria spp. | Zygophyllaceae | 枝 Branches | 12 | 44.29 ± 2.97 | 7 | 1.56 ± 0.28 | 18 | 29.59 ± 7.34 | 25 |
| 茎 Stems | 14 | 45.82 ± 2.07 | 5 | 1.36 ± 0.22 | 17 | 34.66 ± 6.29 | 18 | ||
| 灌木地上部分 Shrub aboveground | 43.95 ± 3.49 | 8 | 1.57 ± 0.40 | 19 | 30.85 ± 7.86 | 22 | |||
| 刺旋花 | 旋花科 | 叶 Leaves | 11 | 40.48 ± 6.52 | 16 | 1.57 ± 0.48 | 30 | 27.82 ± 8.63 | 31 |
| Convolvulus | Convolvulaceae | 枝 Branches | 9 | 43.22 ± 4.16 | 10 | 1.14 ± 0.30 | 26 | 39.99 ± 10.00 | 25 |
| ragacanthoides | 茎 Stems | 13 | 44.10 ± 2.24 | 5 | 1.27 ± 0.17 | 14 | 35.31 ± 5.54 | 16 | |
| 灌木地上部分 Shrub aboveground | 42.18 ± 5.85 | 14 | 1.33 ± 0.31 | 23 | 33.55 ± 8.56 | 26 | |||
| 红砂 | 柽柳科 | 叶 Leaves | 79 | 34.67 ± 3.39 | 10 | 2.42 ± 0.52 | 22 | 14.92 ± 3.24 | 22 |
| Reaumuria | Tamaricaceae | 枝 Branches | 79 | 44.61 ± 1.80 | 4 | 1.74 ± 0.36 | 20 | 26.90 ± 6.65 | 25 |
| songarica | 茎 Stems | 76 | 45.76 ± 2.40 | 5 | 1.37 ± 0.20 | 15 | 34.35 ± 6.43 | 19 | |
| 灌木地上部分 Shrub aboveground | 42.84 ± 2.97 | 7 | 1.73 ± 0.38 | 21 | 27.60 ± 6.48 | 23 | |||
| 假木贼 | 藜科 | 叶 Leaves | 15 | 35.57 ± 2.78 | 8 | 1.76 ± 0.33 | 19 | 20.76 ± 3.92 | 19 |
| Anabasis spp. | Chenopodiaceae | 枝 Branches | 37 | 38.08 ± 4.19 | 11 | 1.61 ± 0.42 | 26 | 25.16 ± 6.70 | 27 |
| 茎 Stems | 25 | 40.15 ± 2.70 | 7 | 1.54 ± 0.42 | 27 | 27.50 ± 6.19 | 23 | ||
| 灌木地上部分 Shrub aboveground | 38.05 ± 2.48 | 7 | 1.63 ± 0.39 | 24 | 24.72 ± 4.66 | 19 | |||
| 锦鸡儿 | 豆科 | 叶 Leaves | 70 | 44.34 ± 2.99 | 7 | 2.66 ± 0.48 | 18 | 17.24 ± 3.77 | 22 |
| Caragana spp. | Leguminosae | 枝 Branches | 71 | 46.71 ± 2.64 | 6 | 1.80 ± 0.48 | 27 | 27.72 ± 7.17 | 26 |
| 茎 Stems | 66 | 47.14 ± 2.48 | 5 | 1.62 ± 0.29 | 18 | 30.26 ± 6.58 | 22 | ||
| 灌木地上部分 Shrub aboveground | 46.44 ± 2.63 | 6 | 1.91 ± 0.45 | 23 | 26.76 ± 6.96 | 26 | |||
| 麻黄 | 麻黄科 | 枝 Branches | 29 | 45.68 ± 3.43 | 8 | 1.89 ± 0.48 | 25 | 25.81 ± 7.26 | 28 |
| Ephedra spp. | Ephedraceae | 茎 Stems | 28 | 44.99 ± 4.33 | 10 | 1.52 ± 0.44 | 29 | 32.20 ± 9.40 | 29 |
| 灌木地上部分 Shrub aboveground | 45.11 ± 2.94 | 7 | 1.75 ± 0.41 | 23 | 27.84 ± 7.16 | 26 | |||
| 沙拐枣 | 蓼科 | 枝 Branches | 11 | 43.56 ± 4.53 | 10 | 1.34 ± 0.16 | 12 | 33.17 ± 6.79 | 20 |
| Calligonum | Polygonaceae | 茎 Stems | 17 | 46.76 ± 2.60 | 6 | 1.18 ± 0.18 | 15 | 40.41 ± 5.28 | 13 |
| mongolicum | 灌木地上部分 Shrub aboveground | 46.46 ± 2.75 | 6 | 1.21 ± 0.16 | 13 | 39.19 ± 4.38 | 17 | ||
| 梭梭 | 藜科 | 叶 Leaves | 63 | 35.73 ± 4.09 | 11 | 2.17 ± 0.50 | 23 | 17.66 ± 5.76 | 33 |
| Haloxylon spp. | Chenopodiaceae | 枝 Branches | 95 | 42.80 ± 3.81 | 9 | 1.58 ± 0.43 | 27 | 29.52 ± 9.57 | 32 |
| 茎 Stems | 95 | 45.57 ± 2.71 | 6 | 1.31 ± 0.36 | 27 | 36.96 ± 9.40 | 25 | ||
| 灌木地上部分 Shrub aboveground | 42.95 ± 3.02 | 7 | 1.54 ± 0.40 | 26 | 28.05 ± 8.24 | 27 | |||
| 金丝桃叶绣线菊 | 蔷薇科 | 叶 Leaves | 58 | 46.60 ± 2.86 | 6 | 2.07 ± 0.40 | 19 | 23.24 ± 4.41 | 19 |
| Spiraea | Rosaceae | 枝 Branches | 62 | 47.66 ± 2.91 | 6 | 1.25 ± 0.23 | 18 | 39.27 ± 6.52 | 17 |
| hypericifolia | 茎 Stems | 62 | 47.57 ± 2.37 | 5 | 1.11 ± 0.19 | 17 | 43.56 ± 5.26 | 12 | |
| 灌木地上部分 Shrub aboveground | 47.43 ± 2.31 | 5 | 1.21 ± 0.17 | 14 | 35.97 ± 4.48 | 15 | |||
| 驼绒藜 | 藜科 | 叶 Leaves | 60 | 38.89 ± 2.93 | 8 | 1.97 ± 0.66 | 33 | 21.72 ± 6.70 | 31 |
| Krascheninnikovia spp. | Chenopodiaceae | 枝 Branches | 61 | 46.14 ± 2.17 | 5 | 1.24 ± 0.4 | 32 | 41.48 ± 14.56 | 35 |
| 茎 Stems | 61 | 46.32 ± 4.50 | 10 | 1.10 ± 0.37 | 34 | 47.14 ± 17.06 | 36 | ||
| 灌木地上部分 Shrub aboveground | 45.55 ± 2.40 | 5 | 1.32 ± 0.49 | 37 | 41.11 ± 14.45 | 35 | |||
| 小蓬 | 藜科 | 叶 Leaves | 19 | 31.97 ± 5.37 | 17 | 1.64 ± 0.41 | 25 | 20.12 ± 3.89 | 19 |
| Nanophyton | Chenopodiaceae | 枝 Branches | 19 | 35.00 ± 7.40 | 21 | 1.40 ± 0.28 | 20 | 25.90 ± 8.40 | 32 |
| erinaceum | 茎 Stems | 20 | 43.09 ± 4.02 | 9 | 1.44 ± 0.30 | 21 | 31.77 ± 10.56 | 33 | |
| 灌木地上部分 Shrub aboveground | 35.82 ± 6.75 | 19 | 1.42 ± 0.30 | 21 | 27.14 ± 10.71 | 29 | |||
| 盐爪爪 | 藜科 | 叶 Leaves | 7 | 33.36 ± 5.48 | 16 | 2.42 ± 0.78 | 32 | 15.46 ± 6.81 | 44 |
| Kalidium spp. | Chenopodiaceae | 枝 Branches | 7 | 42.11 ± 4.10 | 10 | 1.58 ± 0.28 | 17 | 27.28 ± 5.34 | 20 |
| 茎 Stems | 6 | 44.23 ± 2.66 | 6 | 1.37 ± 0.10 | 7 | 32.33 ± 2.22 | 7 | ||
| 灌木地上部分 Shrub aboveground | 37.90 ± 7.91 | 11 | 1.83 ± 0.59 | 19 | 23.88 ± 8.75 | 37 | |||
| 猪毛菜 | 藜科 | 叶 Leaves | 9 | 40.66 ± 5.29 | 13 | 2.05 ± 0.13 | 7 | 19.99 ± 3.71 | 19 |
| Salsola spp. | Chenopodiaceae | 枝 Branches | 9 | 45.72 ± 2.70 | 6 | 1.62 ± 0.24 | 15 | 28.92 ± 6.08 | 21 |
| 茎 Stems | 5 | 46.84 ± 2.49 | 5 | 1.36 ± 0.03 | 2 | 34.38 ± 2.39 | 7 | ||
| 灌木地上部分 Shrub aboveground | 43.86 ± 4.38 | 10 | 1.58 ± 0.21 | 13 | 29.84 ± 5.54 | 19 | |||
| 物种 Species | 科 Family | 器官 Organ | n | 碳含量 C content (%) | CV | 氮含量 N content (%) | CV | C:N | CV |
| 平均值 | 叶 Leaves | 39.15 ± 5.91 | 15 | 2.21 ± 0.59 | 27 | 18.94 ± 5.82 | 31 | ||
| Mean | 枝 Branches | 44.27 ± 4.51 | 10 | 1.55 ± 0.44 | 28 | 31.07 ± 10.43 | 34 | ||
| 茎 Stems | 45.81 ± 3.43 | 7 | 1.34 ± 0.35 | 26 | 36.77 ± 10.78 | 29 | |||
| 灌木地上部分 Shrub aboveground | 43.77 ± 4.43 | 10 | 1.56 ± 0.45 | 27 | 31.78 ± 10.12 | 32 |
新窗口打开|下载CSV
13种典型灌木中枝的C含量最高的是金丝桃叶绣线菊, 为(47.66 ± 2.91)%, 最低的是小蓬, 为(35.00 ± 7.40)%; 枝的N含量最高的是麻黄, 为(1.89 ± 0.48)%, 最低的是刺旋花, 为(1.14 ± 0.30)%; 枝的C:N最高的是驼绒藜, 为41.48 ± 14.56, 最低的是假木贼, 为25.16 ± 6.70。
13种典型灌木中茎的C含量最高的金丝桃叶绣线菊, 为(47.57 ± 2.37)%, 最低的是假木贼, 为(40.15 ± 2.70)%; 茎的N含量最高的是锦鸡儿, 为(1.62 ± 0.29)%, 最低的是驼绒藜, 为(1.10 ± 0.37)%; 茎的C:N最高的是驼绒藜, 为47.14 ± 17.06, 最低的是假木贼, 为27.50 ± 6.19。
13种典型灌木中地上部分C含量最高的是金丝桃叶绣线菊, 为(47.43 ± 2.31)%, 最低的是小蓬, 为(35.82 ± 6.75)%; 灌木地上部分N含量最高的锦鸡儿, 为(1.91 ± 0.45)%, 最低的是沙拐枣, 为(1.21 ± 0.16)%; 灌木地上部分C:N最高的是驼绒藜, 为(41.11 ± 14.45)%, 最低的是盐爪爪, 为(23.88 ± 8.75)%。
C含量的变异系数的平均值(10%)低于N含量的(27%)。另外, 同一属不同器官的N含量的变异系数均大于C含量的变异系数, 不同属同一器官的C、N含量的变异系数也有一定差异。
2.3 不同科灌木C、N含量特征
对于不同科的植物, 将具有确定植物功能特性的一系列植物进行归类组合(Smith et al., 1996)。与物种水平不同的是, 不同科的植物可以将具有相同或相似的生理、形态、生活史等一些生物学特性的植物进行系统分类(孙国钧等, 2003), 本研究将13种灌木归类为8个科进行统计分析。2.3.1 不同科灌木叶C、N含量特征
北疆典型灌木物种(属)归类到不同科后, 叶的C、N含量见表3。由表3可知, 灌木叶的C、N含量及C:N在不同科间有显著差异(p < 0.05)。其中, 蔷薇科C含量显著大于豆科、旋花科、蒺藜科、藜科、柽柳科(p < 0.05); 豆科的N含量显著大于蒺藜科、柽柳科、蔷薇科、藜科、旋花科(p < 0.05); 旋花科的C:N显著大于蔷薇科、藜科科、蒺藜科、柽柳科(p < 0.05)。旋花科的C、N含量及C:N的变异系数较高, 分别为16%、30%和31%。
Table 3
表3
表3不同科灌木叶C、N含量(平均值±标准偏差)
Table 3
| 科 Family | n | 碳含量 C content (%) | CV | 氮含量 N content (%) | CV | C:N | CV |
|---|---|---|---|---|---|---|---|
| 柽柳科 Tamaricaceae | 79 | 34.67 ± 3.39e | 10 | 2.42 ± 0.52b | 22 | 14.92 ± 3.24d | 22 |
| 豆科 Leguminosae | 70 | 44.34 ± 2.99b | 7 | 2.66 ± 0.48a | 18 | 17.24 ± 3.77cd | 22 |
| 蒺藜科 Zygophyllaceae | 12 | 38.40 ± 4.07cd | 11 | 2.42 ± 0.47b | 20 | 16.40 ± 3.57d | 22 |
| 藜科 Chenopodiaceae | 173 | 36.56 ± 4.56d | 12 | 2.01 ± 0.57c | 28 | 19.64 ± 6.02c | 31 |
| 蔷薇科 Rosaceae | 58 | 46.60 ± 2.86a | 6 | 2.07 ± 0.40c | 19 | 23.24 ± 4.41b | 19 |
| 旋花科 Convolvulaceae | 11 | 40.48 ± 6.52c | 16 | 1.57 ± 0.48d | 30 | 27.82 ± 8.63a | 31 |
| 平均值 Average | 39.15 ± 5.91 | 15 | 2.21 ± 0.59 | 27 | 18.94 ± 5.82 | 31 |
新窗口打开|下载CSV
2.3.2 不同科灌木枝C、N含量特征
北疆典型灌木物种(属)归类到不同科后, 枝的C、N含量见表4。由表4可知: 灌木枝的C、N含量及C:N在不同科间有显著差异(p < 0.05), 其中, 蔷薇科的C含量显著大于柽柳科、蒺藜科、蓼科、旋花科、藜科(p < 0.05); 麻黄科的N含量显著大于蒺藜科、藜科、蓼科、蔷薇科、旋花科(p < 0.05); 旋花科的C:N显著大于蓼科、藜科、蒺藜科、豆科、柽柳科、麻黄科(p < 0.05)。藜科的C含量、N含量及C:N的变异系数最高, 分别为12%、29%和38%。
Table 4
表4
表4不同科灌木枝C、N含量(平均值±标准偏差)
Table 4
| 科 Family | n | 碳含量 C content (%) | CV | 氮含量 N content (%) | CV | C:N | CV |
|---|---|---|---|---|---|---|---|
| 柽柳科 Tamaricaceae | 79 | 44.61 ± 1.80bcd | 4 | 1.74 ± 0.36ab | 20 | 26.90 ± 6.65bc | 25 |
| 豆科 Leguminosae | 71 | 46.71 ± 2.64ab | 6 | 1.80 ± 0.48ab | 27 | 27.72 ± 7.17bc | 26 |
| 蒺藜科 Zygophyllaceae | 12 | 44.29 ± 2.97bcd | 7 | 1.56 ± 0.28bc | 18 | 29.59 ± 7.34bc | 25 |
| 藜科 Chenopodiaceae | 228 | 42.37 ± 5.19d | 12 | 1.48 ± 0.42cd | 29 | 31.62 ± 12.13bc | 38 |
| 蓼科 Polygonaceae | 11 | 43.56 ± 4.53cd | 10 | 1.34 ± 0.16cde | 12 | 33.17 ± 6.79b | 20 |
| 麻黄科 Ephedraceae | 29 | 45.68 ± 3.43abc | 8 | 1.89 ± 0.48a | 25 | 25.81 ± 7.26c | 28 |
| 蔷薇科 Rosaceae | 62 | 47.66 ± 2.91a | 6 | 1.25 ± 0.23de | 18 | 39.27 ± 6.52a | 17 |
| 旋花科 Convolvulaceae | 9 | 43.22 ± 4.16cd | 10 | 1.14 ± 0.30e | 26 | 39.99 ± 10.00a | 25 |
| 平均值 Average | 44.27 ± 4.51 | 10 | 1.34 ± 0.35 | 28 | 31.07 ± 10.43 | 34 |
新窗口打开|下载CSV
2.3.3 不同科灌木茎C、N含量特征
北疆典型灌木物种(属)归类到不同科后, 茎的C、N含量见表5。由表5可知: 灌木茎的C、N含量及C:N在不同科间有显著差异(p < 0.05), 其中, 蔷薇科的C含量显著大于柽柳科、麻黄科、藜科、旋花科(p < 0.05); 豆科的N含量显著大于柽柳科、蒺藜科、藜科、旋花科、蓼科、蔷薇科(p < 0.05); 蔷薇科的C:N显著大于旋花科、蒺藜科、柽柳科、麻黄科、豆科(p < 0.05)。麻黄科的C含量变异系数最高, 为10%, 藜科N含量和C:N的变异系数最高, 分别为30%和35%。
Table 5
表5
表5不同科灌木茎C、N含量(平均值±标准偏差)
Table 5
| 科 Family | n | 碳含量 C content (%) | CV | 氮含量 N content (%) | CV | C:N | CV |
|---|---|---|---|---|---|---|---|
| 柽柳科 Tamaricaceae | 76 | 45.76 ± 2.4bcd | 5 | 1.37 ± 0.2bc | 15 | 34.35 ± 6.43cd | 19 |
| 豆科 Leguminosae | 66 | 47.14 ± 2.48ab | 5 | 1.62 ± 0.29a | 18 | 30.26 ± 6.58d | 22 |
| 蒺藜科 Zygophyllaceae | 14 | 45.82 ± 2.07abcd | 5 | 1.36 ± 0.22bcd | 17 | 34.66 ± 6.29bcd | 18 |
| 藜科 Chenopodiaceae | 212 | 44.91 ± 3.94cd | 9 | 1.29 ± 0.38cde | 30 | 38.13 ± 13.43abc | 35 |
| 蓼科 Polygonaceae | 17 | 46.76 ± 2.60abc | 6 | 1.18 ± 0.18de | 15 | 40.41 ± 5.28ab | 13 |
| 麻黄科 Ephedraceae | 28 | 44.99 ± 4.33cd | 10 | 1.52 ± 0.44ab | 29 | 32.20 ± 9.40d | 29 |
| 蔷薇科 Rosaceae | 62 | 47.57 ± 2.37a | 5 | 1.11 ± 0.19e | 17 | 43.56 ± 5.26a | 12 |
| 旋花科 Convolvulaceae | 13 | 44.10 ± 2.24d | 5 | 1.27 ± 0.17cde | 14 | 35.31 ± 5.54bcd | 16 |
| 平均值 Average | 45.81 ± 3.43 | 7 | 1.34 ± 0.35 | 26 | 36.77 ± 10.78 | 29 |
新窗口打开|下载CSV
2.3.4 不同科灌木地上部分C、N含量特征
将北疆典型灌木物种(属)归类到不同科后, 统计分析灌木地上部分C、N含量(表6)。由表6可知: 灌木地上部分C、N含量及C:N在不同科间也有显著性差异(p < 0.05), 其中, 蔷薇科的C含量显著大于麻黄科、蒺藜科、柽柳科、旋花科、藜科(p < 0.05); 豆科的N含量显著大于蒺藜科、藜科、旋花科、蔷薇科、蓼科(p < 0.05); 蔷薇科C:N显著大于旋花科、藜科、蒺藜科、麻黄科、柽柳科、豆科(p < 0.05); 旋花科的C含量变异系数较高, 为14%, 藜科N含量和C:N的变异系数较高, 分别为29%和36%。
Table 6
表6
表6不同科灌木地上部分C、N含量(平均值±标准偏差)
Table 6
| 科 Family | n | 碳含量 C content (%) | CV | 氮含量 N content (%) | CV | C:N | CV |
|---|---|---|---|---|---|---|---|
| 柽柳科 Tamaricaceae | 76 | 42.84 ± 2.97d | 7 | 1.73 ± 0.38ab | 22 | 27.60 ± 6.48cd | 23 |
| 豆科 Leguminosae | 66 | 46.44 ± 2.63ab | 6 | 1.91 ± 0.45a | 23 | 26.76 ± 6.96d | 26 |
| 蒺藜科 Zygophyllaceae | 14 | 43.95 ± 3.49cd | 8 | 1.57 ± 0.40bc | 25 | 30.85 ± 7.86bcd | 25 |
| 藜科 Chenopodiaceae | 212 | 42.03 ± 4.80d | 11 | 1.50 ± 0.44cd | 29 | 32.24 ± 11.71bc | 36 |
| 蓼科 Polygonaceae | 17 | 46.46 ± 2.75ab | 6 | 1.21 ± 0.16e | 13 | 39.19 ± 4.38a | 11 |
| 麻黄科 Ephedraceae | 28 | 45.11 ± 2.94bc | 7 | 1.75 ± 0.41ab | 23 | 27.84 ± 7.16cd | 26 |
| 蔷薇科 Rosaceae | 62 | 47.43 ± 2.31a | 5 | 1.21 ± 0.17e | 14 | 40.97 ± 4.48a | 11 |
| 旋花科 Convolvulaceae | 13 | 42.18 ± 5.85d | 14 | 1.33 ± 0.31de | 23 | 33.55 ± 8.56b | 26 |
| 平均值 Average | 43.77 ± 4.43 | 10 | 1.56 ± 0.45 | 27 | 31.78 ± 10.12 | 32 |
新窗口打开|下载CSV
3 讨论
本研究通过大量的野外调查采样与室内分析, 获取了北疆13种典型灌木的C、N特征参数: 叶的C含量为39.15%, N含量为2.21%, C:N为18.94; 枝的C含量为44.27%, N含量为1.55%, C:N为31.07; 茎的C含量为45.81%, N含量为1.34%, C:N为36.77; 灌木地上部分C含量为43.77%, N含量为1.56%, C:N为31.78。植物枝和茎的主要功能是起疏导作用, 将根吸收的水分和无机盐及根合成的营养物质运输至生殖器官, 同时将叶制造的营养物质运输至根, 对于多年生植物来说, 枝和茎有较强的储藏功能(许鸿川, 2008), 研究区灌木各器官的C含量大小为茎>枝>叶, 灌木地上部分C含量比较接近器官中枝的C含量, 且研究区中灌木多为多年生灌木, 枝和茎具有较高的储C能力, 前人也有相似的研究(马钦彦等, 2002)。一般植物在干物质中C含量约占40% (许鸿川, 2008), 然而本研究灌木叶的C含量集中在40% 以下(平均值为39.15%), 枝和茎的C含量大多集中在45%-50%之间(平均值分别为44.27%和45.76%), 而且不同物种同一器官及同一物种不同器官之间的C、N含量之间都有差异, 因此不能简单地用同一个参数来估算整株灌木的C、N含量。
叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(Field & Mooney, 1986), N素在叶中占很大的比重, 其中在C3植物中, 叶绿体中的N含量占整个叶N含量的75% (Chapin et al., 1987), 因此叶片的N含量远比其他器官的高(Trewavas, 1985), 北疆草地典型灌木各器官N含量大小为叶>枝>茎, 灌木地上部分N含量比较接近枝的N含量, 研究区灌木叶的N含量大多集中在2%以上(平均值2.21%), 而枝和茎的N含量较低, 集中在1.0%-1.5% (平均值分别为1.55%和1.34%), 叶片N含量高于全球尺度内植物叶片N平均含量(1.83%和1.77%)(Elser et al., 2000a; Reich & Oleksyn, 2004)和中国陆地植物叶片平均N含量(1.86%)(He et al., 2008), 可能是因为研究区降水量低, 可以被灌木吸收的N多为可溶性铵态氮, 土壤淋溶过程中N水平升高, 导致可供植物吸收的有效氮增加, 故灌木叶N含量升高(Han et al., 2005)。
C、N在植物个体水平上是相互影响和相互制约的(Tingey et al., 2003), 它们的相互作用及其与生境的关系影响着植物的生长发育和营养状况(贺金生和韩兴国, 2010)。植物的C:N反映了植物生长速率(许振柱等, 2004), 体现植物的营养利用效率, 是植物生命活动的重要维持者和调节者, 同时也是枯枝落叶分解速率的调节因素之一(Field & Mooney, 1986), 植物C:N也代表植物吸收N时同化C的能力, 反映了植物对C和N的相对协调能力(羊留冬等, 2011), 也反映了N的利用效率和固C效率的高低, 因此, 植物C:N是C储存能力和积累速率与限制植物生长的N供应偶联的结果(Herbert et al., 2003), 北疆草地典型灌木各器官的C:N大小为茎>枝>叶, 灌木地上部分C:N比较接近枝的C:N水平。叶片的C:N低于全球尺度植物叶片的C:N (22.50)(Elser et al., 2000b); C:N高, 说明土壤中微生物的同化量超过矿化作用时提供的有效氮量, 植物会出现缺氮的现象; C:N低, 说明土壤在进行矿化作用的时候植物能得到所需的有效氮量(Sabik et al., 2000)。研究区灌木的C、N含量及C:N在不同器官的分布差异极显著(p < 0.01), 与秦岭亚高山草甸30种草本植物的研究结论趋势一致(何亚婷等, 2008), 进一步说明植物不同器官的营养元素分配是有差异的。
由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(Aerts & Chapin, 1999), 北疆灌木中蔷薇科叶、枝和茎的C含量均高于其他科灌木, 说明蔷薇科灌木有较高的储C能力。豆科植物可与根瘤菌共生固氮, 改良土壤的结构, 提高土壤的肥力, 干旱区的豆科植物更是生态系统中有效氮的存储中心(Glasener et al., 2002), 因此豆科叶和茎的N含量高于其他科, 枝的N含量最高的是麻黄科, 是因为麻黄科的枝是进行光合作用的器官(许鸿川, 2008), 叶和枝的C:N最高的是旋花科, 最低的是豆科植物, 有研究表明, 种植豆科灌木对严重退化地恢复和重建有显著的效果(杨玉盛等, 1999)。本研究区各科灌木C的变异系数较低, 是因为C的主要来源是植物叶片进行光合作用将大气中的CO2转变为植物的有机碳, 所以植物体内的C含量相对稳定(高伟, 2010), 植物养分含量的变异大多可以通过物种的形成与发育来解释(Broadley et al., 2004; He et al., 2008), N的变异系数相对较高, 研究区灌木各器官N含量的变异系数均大于C含量的, 因此C:N的变异更多的来自于N含量的变异。
4 结论
本研究以新疆北部草地13种典型灌木为对象进行调查研究, 测定其C、N含量及确定其C:N, 统计分析了本区灌木C、N特征, 得出以下结论:(1)确定了北疆草地13种典型灌木不同器官及灌木地上部分C、N参数, 可为不同尺度估算灌木碳氮储量提供基础数据;
(2)叶是植物重要的生产器官, 枝和茎是植物的疏导和储藏器官, 各器官的C、N元素分配有差异, 本区灌木C含量大小为茎>枝>叶, N含量大小为叶>枝>茎, C:N大小为茎>枝>叶; 组成灌木主要成分的N含量变异比C含量的变异强。
(3)北疆草地典型灌木不同器官的C、N含量及C:N在不同科间有显著差异性(p < 0.05)。蔷薇科的C含量最高; 豆科叶和茎的N含量最高, 由于麻黄的光合器官是枝, 因此麻黄科枝的N含量最高; 旋花科叶和枝的C:N最高, 蔷薇科枝和茎的C:N最高。
致谢 感谢新疆农业大学草业与环境科学学院的张文太、武红旗、贾宏涛、孙宗玖、李志强等教师以及刘秋雨等同学在野外调查采样中付出的艰辛劳动。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/S0065-2504(08)60016-1URL [本文引用: 1]
DOI:10.1007/BF01091640URL [本文引用: 1]
DOI:10.2136/sssaj1978.03615995004200050031xURL [本文引用: 1]

Spatial patterns of dry matter, nitrogen, and organic carbon were investigated for velvet mesquite [(Swartz)] DC and for palo verde [Benth.] ecosystems in the Upper Sonoran Desert of Arizona. Standing understory vegetation and litter originating from the overstory shrub decreased in weight as distance from the shrub center increased. Litter originating from understory vegetation displayed weak spatial patterns in dry weight. Gradients in percentage N and percentage carbon of understory vegetation, understory litter, and shrub litter were generally lacking. However, percentage soil N and organic carbon decreased as horizontal distance from the shrub center and depth to 60 cm from the surface layer increased; soil pH under both shrubs changed with depth and horizontal distance. Quadratic response surfaces for these three soil properties differed significantly between velvet mesquite and palo verde. Limited sampling of honey mesquite (var Torr.) ecosystems in the Chichuahuan Desert in New Mexico showed spatial patterns of dry matter and soil properties that differed somewhat from those of Sonoran Desert shrubs.
DOI:10.1016/S0016-7061(97)00095-5URL [本文引用: 1]
ABSTRACT Plants with C3, C4, and CAM photosynthesis have unique δ13C values which are not altered significantly during decomposition and soil organic matter formation. Consequently, δ13C values of soil organic carbon reflect the relative contribution of plant species with C3, C4, and CAM photosynthetic pathways to community net primary productivity, and have been utilized to document vegetation change, to quantify soil organic matter turnover, and to refine our understanding of earth–atmosphere–biosphere interactions. Here, we review the basis of this methodology, and illustrate its use as a tool for studying grass–woody plant dynamics in a savanna ecosystem. In the Rio Grande Plains of southern Texas, C4 grasslands and savannas have been largely replaced by C3 subtropical thorn woodlands dominated by Prosopis glandulosa. We used δ13C values of soil organic matter, above- and belowground plant biomass, and litter in conjunction with radiocarbon dating and dendrochronology to test the hypotheses that: (1) C3Prosopis groves in uplands and C3Prosopis woodlands in low-lying drainages have been long-term components of the landscape; and (2) Prosopis woodlands of low-lying drainages have expanded up-slope since Anglo-European settlement. Current organic matter inputs were not in isotopic equilibrium with soil organic carbon in any of the patch types sampled. In upland grasslands, δ13C values of vegetation (6120‰) were lower than those of soil organic matter (6117‰), suggesting increased C3 forb abundance in response to long-term, heavy grazing (herbaceous retrogression). In wooded landscape elements, δ13C values of current organic matter inputs were characteristic of C3 plants (6128 to 6125‰), while those of the associated soil organic matter were typically 6120 to 6115‰. These δ13C values indicate that woodlands, groves, and shrub clusters dominated almost exclusively by C3 plants now occupy sites once dominated by C4 grasses. A particularly strong memory of the C4 grasslands that once occupied these sites was recorded in the δ13C values of organic carbon associated with fine and coarse clay fractions (6118 to 6114‰), probably a consequence of the slow organic carbon turnover rates in those soil fractions. When δ13C values of soil organic carbon were evaluated in conjunction with radiocarbon measurements of that same carbon, it appeared that herbaceous retrogression and a shift from C4 grassland to C3 woodland occurred recently, probably within the last 50–100 years. Demographic characteristics of the dominant tree species corroborated the δ13C and 14C evidence, and indicated widespread establishment of P. glandulosa and associated shrubs over the past 100 years. Together, these data provide direct, spatially explicit evidence that vegetation change has occurred recently across the entire landscape at this site. Environmental conditions where C3, C4, and CAM plants coexist (e.g., dry, alkaline soils) generally do not favor the preservation of pollen and phytoliths, and these same areas usually lack historical records of vegetation change. Consequently, vegetation dynamics have been difficult to quantify in grasslands, savannas, and woodlands. However, our results demonstrate clearly that δ13C values of soil organic matter afford a direct and powerful technique for reconstructing vegetation change in these areas.
DOI:10.1111/j.1365-2486.2009.02129.xURL [本文引用: 1]
Abstract Expansion of woody vegetation in grasslands is a worldwide phenomenon with implications for C and N cycling at local, regional and global scales. Although woody encroachment is often accompanied by increased annual net primary production (ANPP) and increased inputs of litter, mesic ecosystems may become sources for C after woody encroachment because stimulation of soil CO 2 efflux releases stored soil carbon. Our objective was to determine if young, sandy soils on a barrier island became a sink for C after encroachment of the nitrogen-fixing shrub Morella cerifera , or if associated stimulation of soil CO 2 efflux mitigated increased litterfall. We monitored variations in litterfall in shrub thickets across a chronosequence of shrub expansion and compared those data to previous measurements of ANPP in adjacent grasslands. In the final year, we quantified standing litter C and N pools in shrub thickets and soil organic matter (SOM), soil organic carbon (SOC), soil total nitrogen (TN) and soil CO 2 efflux in shrub thickets and adjacent grasslands. Heavy litterfall resulted in a dense litter layer storing an average of 809gCm 612 and 36gNm 612 . Although soil CO 2 efflux was stimulated by shrub encroachment in younger soils, soil CO 2 efflux did not vary between shrub thickets and grasslands in the oldest soils and increases in CO 2 efflux in shrub thickets did not offset contributions of increased litterfall to SOC. SOC was 3.6–9.8 times higher beneath shrub thickets than in grassland soils and soil TN was 2.5–7.7 times higher under shrub thickets. Accumulation rates of soil and litter C were highest in the youngest thicket at 101gm 612 yr 611 and declined with increasing thicket age. Expansion of shrubs on barrier islands, which have low levels of soil carbon and high potential for ANPP, has the potential to significantly increase ecosystem C sequestration.
DOI:10.1093/jxb/erh002URL [本文引用: 1]
DOI:10.2307/1310177URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1461-0248.2011.01630.xURLPMID:21592276 [本文引用: 1]
Abstract Ecology Letters (2011) 14 : 709 722 Abstract Encroachment of woody plants into grasslands has generated considerable interest among ecologists. Syntheses of encroachment effects on ecosystem processes have been limited in extent and confined largely to pastoral land uses or particular geographical regions. We used univariate analyses, meta-analysis and structural equation modelling to test the propositions that (1) shrub encroachment does not necessarily lead to declines in ecosystem functions and (2) shrub traits influence the functional outcome of encroachment. Analyses of 43 ecosystem attributes from 244 case studies worldwide showed that some attributes consistently increased with encroachment (e.g. soil C, N), and others declined (e.g. grass cover, pH), but most exhibited variable responses. Traits of shrubs were associated with significant, though weak, structural and functional outcomes of encroachment. Our review revealed that encroachment had mixed effects on ecosystem structure and functioning at global scales, and that shrub traits influence the functional outcome of encroachment. Thus, a simple designation of encroachment as a process leading to functionally, structurally or contextually degraded ecosystems is not supported by a critical analysis of existing literature. Our results highlight that the commonly established link between shrub encroachment and degradation is not universal.
DOI:10.1038/35046058URL [本文引用: 1]
DOI:10.1111/j.1461-0248.2000.00185.xURL [本文引用: 1]
Ecological stoichiometry is the study of the balance of multiple chemical elements in ecological interactions. This paper reviews recent findings in this area and seeks to broaden the stoichiometric concept for use in evolutionary studies, in integrating ecological dynamics with cellular and genetic mechanisms, and in developing a unified means for studying diverse organisms in diverse habitats. This broader approach would then be considered “biological stoichiometry”. Evidence supporting a hypothesised connection between the C:N:P stoichiometry of an organism and its growth rate (the “growth rate hypothesis”) is reviewed. Various data indicate that rapidly growing organisms commonly have low biomass C:P and N:P ratios. Evidence is then discussed suggesting that low C:P and N:P ratios in rapidly growing organisms reflect increased allocation to P-rich ribosomal RNA (rRNA), as rapid protein synthesis by ribosomes is required to support fast growth. Indeed, diverse organisms (bacteria, copepods, fishes, others) exhibit increased RNA levels when growing actively. This implies that evolutionary processes that generate, directly or indirectly, variation in a major life history trait (specific growth rate) have consequences for ecological dynamics due to their effects on organismal elemental composition. Genetic mechanisms by which organisms generate high RNA, high growth rate phenotypes are discussed next, focusing on the structure and organisation of the ribosomal RNA genes (the “rDNA”). In particular, published studies of a variety of taxa suggest an association between growth rate and variation in the length and content of the intergenic spacer (IGS) region of the rDNA tandem repeat unit. In particular, under conditions favouring increased growth or yield, the number of repeat units (“enhancers”) increases (and the IGS increases in length), and transcription rates of rRNA increase. In addition, there is evidence in the literature that increased numbers of copies of rDNA genes are associated with increased growth and production. Thus, a combination of genetic mechanisms may be responsible for establishing the growth potential, and thus the RNA allocation and C:N:P composition, of an organism. Furthermore, various processes, during both sexual and asexual reproduction, can generate variation in the rDNA to provide the raw material for selection and to generate ecologically significant variation in C:N:P stoichiometry. This leads us to hypothesize that the continuous generation of such variation may also play a role in how species interactions develop in ecosystems under different conditions of energy input and nutrient supply.
[本文引用: 2]
DOI:10.7666/d.y1735647URL [本文引用: 1]
放牧、施肥作为草地经营管理的重要措施,会对草地生态系统产生深 远的影响。本文选取内蒙古锡林郭勒盟生态梯度上三种草原类型(草甸草原、典型草原和荒漠草原)的原生群落和退化群落,对其植被、根系和土壤进行了综合研 究,探讨生态环境条件、放牧退化对各草原类型群落、能量、碳氮特征影响的异同规律。并且对内蒙古典型草原不同利用方式下的草地生态系统进行氮素添加实验, 揭示其土壤与植被对氮素添加的响应规律。 本文的主要结论如下: (1)放牧退化使草地生态系统生物量(地上、地下)减少,根冠比增加,群落生物量分配比例发生了一定变化;根系生...
DOI:10.7666/d.y1735647URL [本文引用: 1]
放牧、施肥作为草地经营管理的重要措施,会对草地生态系统产生深 远的影响。本文选取内蒙古锡林郭勒盟生态梯度上三种草原类型(草甸草原、典型草原和荒漠草原)的原生群落和退化群落,对其植被、根系和土壤进行了综合研 究,探讨生态环境条件、放牧退化对各草原类型群落、能量、碳氮特征影响的异同规律。并且对内蒙古典型草原不同利用方式下的草地生态系统进行氮素添加实验, 揭示其土壤与植被对氮素添加的响应规律。 本文的主要结论如下: (1)放牧退化使草地生态系统生物量(地上、地下)减少,根冠比增加,群落生物量分配比例发生了一定变化;根系生...
DOI:10.2136/sssaj2002.0523URL [本文引用: 1]
Legume mulches are important sources of N for cereal crop production, particularly for organic and resource-poor producers. A field study was conducted using a direct method to determine if the amount of N in cereal crops derived from either the shoots or roots of preceding tropical legume cover crops was affected by their chemical composition and mineralization potential. Desmodium ovalifolium Guill. & Perr. [= D. adscendens (Sw.) DC. and Pueraria phaseoloides (Roxb.) Benth.], were grown in 6.0-m2 microplots and foliar-labeled with 99 atom % 15N urea. A cereal sequence of maize (Zea mays L.)-rice (Oryza sativa L.)-maize followed the legumes. Cereal accumulation of legume N from either the shoot (shoot + leaf litter) or the root-soil sources was evaluated by spatially separating the legume N sources. This was achieved by interchanging surface applications of nonlabeled and 15N-labeled legume shoots with in situ 15N-labeled and nonlabeled legume roots. Initially the Desmodium shoot N source contained 316 kg N ha(-1) and roots contained 12.5 kg N ha(-1). Pueraria shoots and root N sources initially contained 262 and 14.8 kg N ha(-1), respectively. About 90 g kg(-1) of the initial N of each legume shoot was recovered in the total aboveground tissues from the three cereal crops, while 490 g kg(-1) of Desmodium and 280 g kg(-1) of Pueraria root-soil N sources were recovered. Of the 181 kg N ha(-1) accumulated aboveground by the cereal sequence, the contribution of shoot plus root-soil N sources was 200 g kg(-1) from Desmodium and 150 g kg(-1) from Pueraria. Cereal N was derived primarily from mineralization of soil organic matter present before the legumes and possibly from N deposition (precipitation and dry) occurring during the cereal crop sequence. After harvest of the last cereal crop, 13 and 180 g kg(-1) of the initial legume N was present as inorganic and organic N fractions, respectively, in the top 75 cm of soil. Even though Pueraria shoots had a lower C:N ratio and concentration of polyphenols than Desmodium shoots, the relative contributions of the shoot N source were similar for both legumes. Decomposition of legume residues, particularly legume shoots, make a meaningful contribution to the N economy of cereal crops grown in the tropics. The legume cover crops (root + shoot) contributed nearly 280 g kg(-1) of the aboveground N in the first cereal crop and as much as 110 g kg(-1) of the N in the third crop during the 15-mo sequence of cereals.
DOI:10.1038/418593aURL [本文引用: 1]
Replacement of grassland by shrubland, which is occurring on a large scale in the United States, is thought to lock up considerable amounts of carbon. This 'carbon sink' may be much smaller than previously estimated.
DOI:10.1007/BF00138373URL [本文引用: 1]
DOI:10.1111/j.1469-8137.2005.01530.xURLPMID:16219077 [本文引用: 1]
Abstract Leaf nitrogen and phosphorus stoichiometry of Chinese terrestrial plants was studied based on a national data set including 753 species across the country. Geometric means were calculated for functional groups based on life form, phylogeny and photosynthetic pathway, as well as for all 753 species. The relationships between leaf N and P stoichiometric traits and latitude (and temperature) were analysed. The geometric means of leaf N, P, and N : P ratio for the 753 species were 18.6 and 1.21 mg g(-1) and 14.4, respectively. With increasing latitude (decreasing mean annual temperature, MAT), leaf N and P increased, but the N : P ratio did not show significant changes. Although patterns of leaf N, P and N : P ratios across the functional groups were generally consistent with those reported previously, the overall N : P ratio of China's flora was considerably higher than the global averages, probably caused by a greater shortage of soil P in China than elsewhere. The relationships between leaf N, P and N : P ratio and latitude (and MAT) also suggested the existence of broad biogeographical patterns of these leaf traits in Chinese flora.
DOI:10.3773/j.issn.1005-264x.2010.01.002URL [本文引用: 1]
正从1909年丹麦哥本哈根大学Johannes Warming(1841-1924)出版第一本生态学教科书《植物生态学》到现在,生态学经历了100年的发展。作为研究
DOI:10.3773/j.issn.1005-264x.2010.01.002URL [本文引用: 1]
正从1909年丹麦哥本哈根大学Johannes Warming(1841-1924)出版第一本生态学教科书《植物生态学》到现在,生态学经历了100年的发展。作为研究
DOI:10.1007/s00442-007-0912-yURL [本文引用: 2]
URLMagsci [本文引用: 1]
以秦岭佛坪国家级自然保护区内亚高山草甸30种草本植物为对象,研究了碳、氮在亚高山草甸植物中的分布情况。结果表明:碳、氮在植物组织中总的分布趋势为叶茎根,碳/氮在植物组织中的分布为茎根叶,方差分析显示碳、氮及碳/氮在植物根、茎、叶中的分布差异极显著(P0.01)。在单个植物的根、茎、叶中,碳、氮及碳/氮的分布具有种的特异性。在以其光合特征进行聚类分析得到的4个聚类组群中,碳、氮及碳/氮在每个组群根、茎、叶中的分布差异不显著(P0.05)。
URLMagsci [本文引用: 1]
以秦岭佛坪国家级自然保护区内亚高山草甸30种草本植物为对象,研究了碳、氮在亚高山草甸植物中的分布情况。结果表明:碳、氮在植物组织中总的分布趋势为叶茎根,碳/氮在植物组织中的分布为茎根叶,方差分析显示碳、氮及碳/氮在植物根、茎、叶中的分布差异极显著(P0.01)。在单个植物的根、茎、叶中,碳、氮及碳/氮的分布具有种的特异性。在以其光合特征进行聚类分析得到的4个聚类组群中,碳、氮及碳/氮在每个组群根、茎、叶中的分布差异不显著(P0.05)。
DOI:10.1023/A:1026020210887URL [本文引用: 1]
Productivity and carbon (C) storage in many mature tropical forests are considered phosphorus (P) limited because of advanced soil weathering. However, disturbance can shift limitation away from P and toward nitrogen (N) because of disproportionately large N losses associated with its mobility relative to P in ecosystems. This shift was illustrated by model analyses in which large disturbances including timber extraction and slash-burn were simulated in a P-limited tropical forest. Re-accumulation of ecosystem C during secondary forest growth was initially N-limited, but long term limitation reverted to P. Mechanisms controlling shifts between N and P limitation included: (1) N volatility during slash combustion produced ash that increased soil solution P more than N, (2) a wide N:P ratio in residual fuel and belowground necromass relative to soil organic matter (SOM) N:P produced a simultaneous P sink and N source during decomposition, (3) a supplemental (to aerosol deposition) external N source via biological N fixation. Redistribution of N and P from low C:nutrient SOM to high C:nutrient vegetation was the most important factor contributing to the resilience of ecosystem C accumulation during secondary growth. Resilience was diminished when multiple harvest and re-growth cycles depleted SOM. Phosphorus losses in particular resulted in long-term reductions of C storage capacity because of slow re-supply rates via deposition and the absence of other external sources. Sensitivity analyses limiting the depth of microbially active SOM in soil profiles further illustrated the importance of elements stored in SOM to ecosystem resilience, pointing to a need for better knowledge on the functioning of deeply buried SOM.
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/1051-0761(2000)010[0470:BCOVCA]2.0.CO;2URL [本文引用: 1]
The extent and consequences of global land-cover and land-use change are increasingly apparent. One consequence not so apparent is the altered structure of plants belowground. This paper examines such belowground changes, emphasizing the interaction of altered root distributions with other factors and their treatment in models. Shifts of woody and herbaceous vegetation with deforestation, afforestation, and woody plant encroachment typically alter the depth and distribution of plant roots, influencing soil nutrients, the water balance, and net primary productivity (NPP). For example, our analysis of global soil data sets shows that the major plant nutrients C, N, P, and K are more shallowly distributed than are Ca, Mg, and Na, but patterns for each element vary with the dominant vegetation type. After controlling for climate, soil C and N are distributed more deeply in arid shrublands than in arid grasslands, and subhumid forests have shallower nutrient distributions than do subhumid grasslands. Consequently, changes in vegetation may influence the distribution of soil carbon and nutrients over time (perhaps decades to centuries). Shifts in the water balance are typically much more rapid. Catchment studies indicate that the water yield decreases 25-40 mm for each 10% increase in tree cover, and increases in transpiration of water taken up by deep roots may account for as much as 50% of observed responses. Because models are increasingly important for predicting the consequences of vegetation change, we discuss the treatment of belowground processes and how different treatments affect model outputs. Whether models are parameterized by biome or plant life form (or neither), use single or multiple soil layers, or include N and water limitation will all affect predicted outcomes. Acknowledging and understanding such differences should help constrain predictions of vegetation change.
DOI:10.1111/j.1365-2486.2007.01512.xURL [本文引用: 1]
Abstract Shrub encroachment into grass-dominated biomes is occurring globally due to a variety of anthropogenic activities, but the consequences for carbon (C) inputs, storage and cycling remain unclear. We studied eight North American graminoid-dominated ecosystems invaded by shrubs, from arctic tundra to Atlantic coastal dunes, to quantify patterns and controls of C inputs via aboveground net primary production (ANPP). Across a fourfold range in mean annual precipitation (MAP), a key regulator of ecosystem C input at the continental scale, shrub invasion decreased ANPP in xeric sites, but dramatically increased ANPP (>1000gm 2 ) at high MAP, where shrub patches maintained extraordinarily high leaf area. Concurrently, the relationship between MAP and ANPP shifted from being nonlinear in grasslands to linear in shrublands. Thus, relatively abrupt (<50 years) shifts in growth form dominance, without changes in resource quantity, can fundamentally alter continental-scale pattern of C inputs and their control by MAP in ways that exceed the direct effects of climate change alone.
DOI:10.3969/j.issn.1003-7578.2006.02.031URL [本文引用: 1]
文章论述了国内外在灌木的生态性和生态效能方面的研究概况,介绍了灌木的主要特征及其对生长环境和干扰响应的生态学论述,并分别就干旱荒漠区灌木、草牧场地灌木、盐碱地灌木和森林区次生灌木四种主要灌木类型进行了说明.另外主要从生物多样性、水土保持、防风固沙、土壤改良、改善气候等五个方面阐述了灌木林的生态效能研究现状.最后指出灌木今后的重点研究方向.
DOI:10.3969/j.issn.1003-7578.2006.02.031URL [本文引用: 1]
文章论述了国内外在灌木的生态性和生态效能方面的研究概况,介绍了灌木的主要特征及其对生长环境和干扰响应的生态学论述,并分别就干旱荒漠区灌木、草牧场地灌木、盐碱地灌木和森林区次生灌木四种主要灌木类型进行了说明.另外主要从生物多样性、水土保持、防风固沙、土壤改良、改善气候等五个方面阐述了灌木林的生态效能研究现状.最后指出灌木今后的重点研究方向.
DOI:10.3321/j.issn:1007-7588.2000.03.013URLMagsci [本文引用: 1]
鄂尔多斯高原是我国北方一个相对独立的自然单元,从大气环流、地质地貌、土壤、水文、植被及人文和社会背景等方面讲,它是一个多层次的,十分复杂的生态过渡带。特殊的自然生态背景使之成为灌木种类分布的“天然天国”。该地区的西北部植物区系在古地中海的海浸区,且未直接
DOI:10.3321/j.issn:1007-7588.2000.03.013URLMagsci [本文引用: 1]
鄂尔多斯高原是我国北方一个相对独立的自然单元,从大气环流、地质地貌、土壤、水文、植被及人文和社会背景等方面讲,它是一个多层次的,十分复杂的生态过渡带。特殊的自然生态背景使之成为灌木种类分布的“天然天国”。该地区的西北部植物区系在古地中海的海浸区,且未直接
DOI:10.3321/j.issn:1000-1522.2002.05.019URL [本文引用: 1]
应用干烧法对华北地区主要森林类型的 8个乔木建群种和 1 0个灌木树种不同器官的有机含碳率进行了测定 ,同时利用生物量标准地资料对其中的 7个乔木树种的林分平均含碳率进行了分析 .8种乔木及 1 0种灌木的器官平均含碳率分别为辽东栎 0 475 0、白桦 0 51 2 5、山杨 0 4880、椴树 0 476 4、油松 0 51 0 5、侧柏 0 50 1 0、华北落叶松 0 51 5 8、红皮云杉 0 51 1 8,1 0种灌木 0 4897;7个乔木树种林分的平均含碳率分别为 ,辽东栎 0 476 1、白桦 0 50 0 8、山杨0 485 9、油松 0 50 3 0、侧柏 0 50 5 3、华北落叶松 0 50 97、红皮云杉 0 51 1 1 .各树种器官含碳率的种内变动系数在1 49%~ 6 32 %之间 ;器官含碳率的种间变动系数在 2 1 5 %~ 7 48%之间 .针叶树种器官的平均含碳率普遍比阔叶树种平均高 1 6 %~ 3 4% ,相应的针叶林分的平均含碳率也高于阔叶林
DOI:10.3321/j.issn:1000-1522.2002.05.019URL [本文引用: 1]
应用干烧法对华北地区主要森林类型的 8个乔木建群种和 1 0个灌木树种不同器官的有机含碳率进行了测定 ,同时利用生物量标准地资料对其中的 7个乔木树种的林分平均含碳率进行了分析 .8种乔木及 1 0种灌木的器官平均含碳率分别为辽东栎 0 475 0、白桦 0 51 2 5、山杨 0 4880、椴树 0 476 4、油松 0 51 0 5、侧柏 0 50 1 0、华北落叶松 0 51 5 8、红皮云杉 0 51 1 8,1 0种灌木 0 4897;7个乔木树种林分的平均含碳率分别为 ,辽东栎 0 476 1、白桦 0 50 0 8、山杨0 485 9、油松 0 50 3 0、侧柏 0 50 5 3、华北落叶松 0 50 97、红皮云杉 0 51 1 1 .各树种器官含碳率的种内变动系数在1 49%~ 6 32 %之间 ;器官含碳率的种间变动系数在 2 1 5 %~ 7 48%之间 .针叶树种器官的平均含碳率普遍比阔叶树种平均高 1 6 %~ 3 4% ,相应的针叶林分的平均含碳率也高于阔叶林
DOI:10.1038/372666a0URL [本文引用: 1]
DOI:10.1006/jare.2001.0902URL [本文引用: 1]
Carbon storage in grasslands of China was estimated by the carbon density method and based on a nationwide grassland resource survey finished by 1991. The grasslands in China were classified into 18 types, which are distributed mostly in the temperate region and on the Tibetan Plateau, and scattered in the warm-temperate and tropical regions. Based on the median estimate, vegetation, soil and total carbon storage of grasslands in China were 3·06, 41·03 and 44·09 Pg C, respectively. Vegetation had low carbon storage and most carbon was stored in soils. Of the four types of regions that have grasslands, alpine region (54·5%) and temperate region (31·6%) hold more than 85% of the total grassland carbon (in both vegetation and soils) in China. Considering specific types within these two regions, three grassland types, alpine meadow (25·6%), alpine steppe (14·5%) and temperate steppe (11%) constituted more than half of all carbon stored in China's grasslands. In general and regardless of regional vegetation types, steppes (38·6%) and meadows (38·2%) made up more than 2/3 of total grassland carbon. The carbon storage in alpine grasslands may have a significant and long-lived effect on global C cycles. This study estimated more carbon storage in vegetation and less in soils than previous studies. The differences of grassland carbon between this study and two previous studies were due probably to four reasons, i.e. different estimation methods, different classification systems of grasslands, different areas of grasslands, and different carbon densities. China's grasslands cover only 6–8% of total world grassland area and have 9–16% of total carbon in the world grasslands. They make a big contribution to the world carbon storage and may have significant effects on carbon cycles, both in global and in arid lands.
DOI:10.1007/s10584-004-0070-8URL [本文引用: 1]
Forage yield-based carbon storage in 18 grasslands of China was estimated according to the detailed investigation of grassland area and forage yield (standing crop), which were derived from a 10-year national grassland survey. The total forage yield carbon in Chinese grasslands is 134.09 Tg C for ca. 299 10 6 ha of grassland area and 1232 kg/ha of mean forage yield. The carbon storage is different depending on grassland types and climatic regions. Meadow, steppe and tussock occupy 93.3% (125.14 Tg C), and desert and swamp only accounts for 6.7% (8.95 Tg C) of total forage yield carbon. Forage yield carbon is stored largely in temperate (38.4%, 51.54 Tg C) and alpine regions (30.4%, 40.78 Tg C), and to less extent in tropical regions (22.1%, 29.66 Tg C). These three regions take 91% of the forage yield carbon in grasslands of China. The warm-temperate region accounts for only 9% (12.1 Tg C) of forage yields carbon. The forage yield-based carbon in grasslands of China is more accurate than the site biomass-based carbon estimate and the carbon density-based estimate. Although, forage yield carbon storage is small compared with the total carbon storage in China, carbon budgets of grasslands are often a dominant component in many regions and provide an important management opportunity to enhance terrestrial carbon sinks in vast areas of China.
DOI:10.1016/j.jaridenv.2004.09.019URL [本文引用: 1]
DOI:10.1073/pnas.0403588101URLPMID:15213326 [本文引用: 1]
A global data set including 5,087 observations of leaf nitrogen (N) and phosphorus (P) for 1,280 plant species at 452 sites and of associated mean climate indices demonstrates broad biogeographic patterns. In general, leaf N and P decline and the N/P ratio increases toward the equator as average temperature and growing season length increase. These patterns are similar for five dominant plant groups, coniferous trees and four angiosperm groups (grasses, herbs, shrubs, and trees). These results support the hypotheses that (i) leaf N and P increase from the tropics to the cooler and drier midlatitudes because of temperature-related plant physiological stoichiometry and biogeographical gradients in soil substrate age and then plateau or decrease at high latitudes because of cold temperature effects on biogeochemistry and (ii) the N/P ratio increases with mean temperature and toward the equator, because P is a major limiting nutrient in older tropical soils and N is the major limiting nutrient in younger temperate and high-latitude soils.
DOI:10.1016/S0021-9673(99)01084-5URLPMID:10941674 [本文引用: 1]
The review describes the use of solid-phase extraction (SPE) techniques for monitoring priority pesticides in ground and surface waters. The focus is on triazine herbicides and their degradation products. Data concerning the fate, occurrence, properties and extraction of triazines and their degradation products using different SPE techniques are tabulated and discussed.
[本文引用: 1]
DOI:10.3321/j.issn:1000-0933.2003.07.023URL [本文引用: 1]
综述了植物功能多样性与功能群研究的最新进展.介绍了植物功能群的定义及 植物功能群的划分方法.在功能多样性与生态系统资源动态关系方面,抽样效应和生态位互补效应用来解释植物多样性在生态系统资源动态中的作用.功能多样性与 生态系统的稳定性间的关系可以用生态冗余或生态保险概念来解释,这两个概念是一个问题的两个侧面,是多样性与生态系统功能争论的焦点.
DOI:10.3321/j.issn:1000-0933.2003.07.023URL [本文引用: 1]
综述了植物功能多样性与功能群研究的最新进展.介绍了植物功能群的定义及 植物功能群的划分方法.在功能多样性与生态系统资源动态关系方面,抽样效应和生态位互补效应用来解释植物多样性在生态系统资源动态中的作用.功能多样性与 生态系统的稳定性间的关系可以用生态冗余或生态保险概念来解释,这两个概念是一个问题的两个侧面,是多样性与生态系统功能争论的焦点.
DOI:10.1046/j.1365-2486.2003.00646.xURL [本文引用: 1]
Abstract The effects of elevated CO 2 and temperature on principal carbon constituents (PCC) and C and N allocation between needle, woody (stem and branches) and root tissue of Pseudotsuga menziesii Mirb. Franco seedlings were determined. The seedlings were grown in sun-lit controlled-environment chambers that contained a native soil. Chambers were controlled to reproduce ambient or ambient +180ppm CO 2 and either ambient temperature or ambient +3.5 C for 4years. There were no significant CO 2 temperature interactions; consequently the data are presented for the CO 2 and temperature effects. At the final harvest, elevated CO 2 decreased the nonpolar fraction of the PCC and increased the polar fraction and amount of sugars in the needles. In contrast, elevated temperature increased the nonpolar fraction of the PCC and decreased sugars in needles. There were no CO 2 or temperature effects on the PCC fractions in the woody tissue or root tissue. Elevated CO 2 and temperature had no significant effects on the C content of any of the plant tissues or fractions. In contrast, the foliar N content declined under elevated CO 2 and increased under elevated temperature; there were no significant effects in other tissues. The changes in the foliar N concentrations were in the cellulose and lignin fractions, the fractions, which contain protein, and are the consequences of changes in N allocation under the treatments. These results indicate reallocation of N among plant organs to optimize C assimilation, which is mediated via changes in the selectivity of Rubisco and carbohydrate modulation of gene expression.
[本文引用: 1]
DOI:10.1146/annurev.ecolsys.31.1.197URL [本文引用: 2]
DOI:10.2307/2260247URL [本文引用: 1]
This is the second part of a two-volume work which presents an authoritative world-wide view of our knowledge about, and understanding of, hot-desert ecosystems. This includes some semi-arid and arid areas, as well as deserts in the strict sense. The hot deserts are distinguished from the temperate deserts (which form the subject of another volume in the series) by the virtual absence of snowfall, even though frosts may occur. For each major hot-desert region, expert authors have summarized existing knowledge according to a general outline. This includes descriptions of the ecosystem components (climate, soil, flora and fauna), and discussion of interaction between components and overall ecosystem functioning. The information from the regional chapters has been integrated into a world-wide view in the ``synthesis'' chapters. Because of its length, the volume is published in two parts. The first part included the general synthesis chapters, and regional descriptions of the hot deserts of America and Australia. The present volume covers hot deserts of Asia and Africa.
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
DOI:10.3321/j.issn:1000-0933.2004.10.014URL [本文引用: 1]
研究利用大型环境生长箱模拟了两种沙地优势灌木柠条和羊柴对 CO2 浓度倍增和土壤干旱交互作用的响应。 CO2 浓度倍增使柠条和羊柴的生物量分别增加了 6 2 .90 %和 5 0 .0 0 % ,使植株叶面积分别增加了 4 1.86 %和 4 5 .84 %。 CO2 浓度的倍增效应随着土壤干旱的增加而下降。 CO2 浓度倍增和土壤干旱都增加单位叶面积质量 (L MA) ,但 CO2 浓度倍增主要增加了水分充足时的 L MA。 CO2 倍增使柠条和羊柴叶片含氮量分别降低了 10 .4 0 %和 5 .0 6 %。柠条叶片含氮量在所有土壤干旱条件下均呈现出增加的趋势 ,而羊柴叶片的含氮量仅在严重干旱条件下增加。 CO2 倍增使叶片的碳氮比显著增加 ,但土壤干旱使之降低。CO2 浓度倍增降低叶肉细胞质膜的过氧化产物丙二醛 (MDA )的含量 ,干旱使之增加。叶片含氮量与 MDA呈显著正相关。研究表明 CO2 倍增有保护叶片免受严重土壤干旱的作用 ,但干旱的负面影响是 CO2 倍增效应所难以弥补的
DOI:10.3321/j.issn:1000-0933.2004.10.014URL [本文引用: 1]
研究利用大型环境生长箱模拟了两种沙地优势灌木柠条和羊柴对 CO2 浓度倍增和土壤干旱交互作用的响应。 CO2 浓度倍增使柠条和羊柴的生物量分别增加了 6 2 .90 %和 5 0 .0 0 % ,使植株叶面积分别增加了 4 1.86 %和 4 5 .84 %。 CO2 浓度的倍增效应随着土壤干旱的增加而下降。 CO2 浓度倍增和土壤干旱都增加单位叶面积质量 (L MA) ,但 CO2 浓度倍增主要增加了水分充足时的 L MA。 CO2 倍增使柠条和羊柴叶片含氮量分别降低了 10 .4 0 %和 5 .0 6 %。柠条叶片含氮量在所有土壤干旱条件下均呈现出增加的趋势 ,而羊柴叶片的含氮量仅在严重干旱条件下增加。 CO2 倍增使叶片的碳氮比显著增加 ,但土壤干旱使之降低。CO2 浓度倍增降低叶肉细胞质膜的过氧化产物丙二醛 (MDA )的含量 ,干旱使之增加。叶片含氮量与 MDA呈显著正相关。研究表明 CO2 倍增有保护叶片免受严重土壤干旱的作用 ,但干旱的负面影响是 CO2 倍增效应所难以弥补的
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
1
1999
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
Mechanisms of shrubland expansion: Land use, climate, or CO2
1
1995
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
Shrub-induced spatial patterns of dry matter, nitrogen, and organic carbon
1
1978
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
δ 13C values of soil organic carbon and their use in documenting vegetation change in a subtropical savanna ecosystem
1
1998
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Shrub expansion stimulates soil C and N storage along a coastal soil chronosequence
1
2010
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Phylogenetic variation in the shoot mineral concentration of angiosperms
1
2004
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
Plant responses to multiple environmental factors
1
1987
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
1
1996
... id="C10">草地类是具有相同水热大气候带特征和植被特征、具有独特地带性的草地, 依据热量和水分条件, 将草地划分为不同草地类(
1
1996
... id="C10">草地类是具有相同水热大气候带特征和植被特征、具有独特地带性的草地, 依据热量和水分条件, 将草地划分为不同草地类(
Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis
1
2011
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Nutritional constraints in terrestrial land freshwater food webs
1
2000a
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
Biological stoichiometry from genes to ecosystems
1
2000b
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
2
1992
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
退化草地生态系统碳氮变化规律及氮添加的生态效应研究
1
2010
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
退化草地生态系统碳氮变化规律及氮添加的生态效应研究
1
2010
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
Contributions of shoot and root nitrogen-15 labeled legume nitrogen sources to a sequence of three cereal crops
1
2002
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
Carbon cycle: Uncertain sinks in the shrubs
1
2002
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Shrubland encroachment in southern New Mexico, U.S.A.: An analysis of desertification processes in the American southwest
1
1990
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
Leaf nitrogen: Phosphorus stoichiometry across Chinese grassland Biomes
2
2008
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
秦岭亚高山草甸30种草本植物的碳、氮分布研究
1
2008
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
秦岭亚高山草甸30种草本植物的碳、氮分布研究
1
2008
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
A model analysis of N and P limitation on carbon accumulation in Amazonian secondary forest after alternate land-use abandonment
1
2003
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
1
1982
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
1
1982
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
Belowground consequences of vegetation change and their treatment in models
1
2000
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Shrub encroachment in North American grasslands: Shifts in growth form dominance rapidly alters control of ecosystem carbon inputs
1
2008
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
灌木的生态特性与生态效能的研究与进展
1
2006
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
灌木的生态特性与生态效能的研究与进展
1
2006
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
试论鄂尔多斯高原灌木多样性的若干特点
1
2000
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
试论鄂尔多斯高原灌木多样性的若干特点
1
2000
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
华北主要森林类型建群种的含碳率分析
1
2002
... id="C61">植物枝和茎的主要功能是起疏导作用, 将根吸收的水分和无机盐及根合成的营养物质运输至生殖器官, 同时将叶制造的营养物质运输至根, 对于多年生植物来说, 枝和茎有较强的储藏功能(
华北主要森林类型建群种的含碳率分析
1
2002
... id="C61">植物枝和茎的主要功能是起疏导作用, 将根吸收的水分和无机盐及根合成的营养物质运输至生殖器官, 同时将叶制造的营养物质运输至根, 对于多年生植物来说, 枝和茎有较强的储藏功能(
The role of deep roots in the hydrological and carbon cycles of Amazonian forests and pastures
1
1994
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Carbon storage in grasslands of China
1
2002
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Forage yield-based carbon storage in grasslands of China
1
2004
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Above-ground biomass and carbon and nitrogen content of woody species in a subtropical thornscrub parkland
1
2005
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Global patterns of plant leaf N and P in relation to temperature and latitude
1
2004
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
Multiresidue methods using solid-phase extraction techniques for monitoring priority pesticides, including triazines and degradation products, in ground and surface waters
1
2000
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
1
1992
... id="C35">对于不同科的植物, 将具有确定植物功能特性的一系列植物进行归类组合(
植物功能多样性与功能群研究进展
1
2003
... id="C35">对于不同科的植物, 将具有确定植物功能特性的一系列植物进行归类组合(
植物功能多样性与功能群研究进展
1
2003
... id="C35">对于不同科的植物, 将具有确定植物功能特性的一系列植物进行归类组合(
Elevated CO2 and temperature alter nitrogen allocation in Douglas-fir
1
2003
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
1
1992
... id="C62">叶片进行光合作用的N来源于植物根系对N的吸收及茎向叶片的运输(
Shrub invasions of North American semiarid grasslands
2
2000
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
... id="C7">灌木能使草地生态系统的生物量及生物量分配、根系深度、凋落物数量和质量等发生改变(
Hot deserts and arid shrublands A
1
1986
... id="C6">灌木在维持荒漠生态系统稳定中发挥着重要的作用(
3
2008
... id="C61">植物枝和茎的主要功能是起疏导作用, 将根吸收的水分和无机盐及根合成的营养物质运输至生殖器官, 同时将叶制造的营养物质运输至根, 对于多年生植物来说, 枝和茎有较强的储藏功能(
... ).一般植物在干物质中C含量约占40% (
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
3
2008
... id="C61">植物枝和茎的主要功能是起疏导作用, 将根吸收的水分和无机盐及根合成的营养物质运输至生殖器官, 同时将叶制造的营养物质运输至根, 对于多年生植物来说, 枝和茎有较强的储藏功能(
... ).一般植物在干物质中C含量约占40% (
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
2
1993
... id="C8">新疆是典型的干旱半干旱地区, 是我国仅次于西藏和内蒙古的第三大草原省区, 新疆可利用草地面积约为4800.68万hm2 (
... id="C9">研究区位于新疆维吾尔自治区天山北部, 包括准噶尔盆地、伊犁河谷、塔额盆地、阿勒泰山南麓及天山北坡等灌木分布区, 地理坐标为80.73°- 91.12° E , 42.85°-48.48° N, 海拔230-2456 m.北疆属于温带大陆性干旱半干旱气候区, 光照充足, 无霜期短, 年降水量较少, 为150-200 mm, 年蒸发量大, 年平均气温-4-9 ℃ (
2
1993
... id="C8">新疆是典型的干旱半干旱地区, 是我国仅次于西藏和内蒙古的第三大草原省区, 新疆可利用草地面积约为4800.68万hm2 (
... id="C9">研究区位于新疆维吾尔自治区天山北部, 包括准噶尔盆地、伊犁河谷、塔额盆地、阿勒泰山南麓及天山北坡等灌木分布区, 地理坐标为80.73°- 91.12° E , 42.85°-48.48° N, 海拔230-2456 m.北疆属于温带大陆性干旱半干旱气候区, 光照充足, 无霜期短, 年降水量较少, 为150-200 mm, 年蒸发量大, 年平均气温-4-9 ℃ (
CO2浓度倍增条件下土壤干旱对两种沙生灌木碳氮含量及其适应性的影响
1
2004
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
CO2浓度倍增条件下土壤干旱对两种沙生灌木碳氮含量及其适应性的影响
1
2004
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
短期增温对贡嘎山峨眉冷杉幼苗生长季及其CNP化学计量学特征的影响
1
2011
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
短期增温对贡嘎山峨眉冷杉幼苗生长季及其CNP化学计量学特征的影响
1
2011
... id="C63">C、N在植物个体水平上是相互影响和相互制约的(
严重退化生态系统不同恢复和重建措施的植物多样性与地力差异研究
1
1999
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
严重退化生态系统不同恢复和重建措施的植物多样性与地力差异研究
1
1999
... id="C64">由于植物养分在各个器官的含量及分配不仅受生长环境养分有效性的制约, 而且也与植物自身的生理特征及生长型有关(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}