 ,
, ,1,2,**, 白永飞2
,1,2,**, 白永飞2Effects of litter removal and addition on ecosystem carbon fluxes in a typical steppe
ZHANG Su-Yan1,2,*, JIANG Hong-Zhi2,*, WANG Yang2, ZHANG Yan-Jie1, LU Shun-Bao,,1,2,**, BAI Yong-Fei2通讯作者:
第一联系人:
编委: 石培礼
责任编辑: 王葳
| 基金资助: |
Online:2018-03-20
| Supported by: | SupportedbytheNationalKeyTechnologyResearchandDevelopmentProgramoftheMinistryofScienceandTechnologyofChina( |

摘要
关键词:
Abstract
Keywords:
PDF (1460KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
张素彦, 蒋红志, 王扬, 张艳杰, 鲁顺保, 白永飞. 凋落物去除和添加处理对典型草原生态系统碳通量的影响. 植物生态学报, 2018, 42(3): 349-360 doi:10.17521/cjpe.2016.0083
ZHANG Su-Yan, JIANG Hong-Zhi, WANG Yang, ZHANG Yan-Jie, LU Shun-Bao, BAI Yong-Fei.
陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(Baldocchi, 2008), 是生态系统总初级生产力(GEP)与生态系统呼吸(ER)平衡的结果(Oberbauer et al., 2007)。在全球陆地生态系统中, 草地生态系统分布最广泛, 约占全球陆地总面积的30% (吴灏等, 2013)。在我国, 草地生态系统在碳平衡中扮演着重要角色(朴世龙等, 2004)。草地碳循环中CO2净交换及其对环境变化的响应是目前全球变化研究的热点, 各国研究异常活跃, 研究者们正尝试通过量化各种生态系统碳通量与环境因子之间的关系模型预测未来全球变化对陆地生态系统的影响(Li et al., 2005)。生态系统生物地球化学过程与全球变化相互作用, 促进或抑制植物净初级生产力(NPP)。例如氮沉降(Xia et al., 2009)、加水(Niu et al., 2008)和增温(Oberbauer et al., 2007)均增加了GEP, 促进了植物生产力。然而, 干旱(Zhao & Running, 2010)和增加对流层臭氧(Liu et al., 2005)一般会降低植物生产力。另外, 生物因子及人类活动干扰也通过改变物理环境因子直接或间接地影响着生态系统的碳通量(Dugas et al., 1999; Frank & Dugas, 2001; Li et al., 2005)。
凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(Sayer et al., 2006)。已有研究表明凋落物数量和质量发生变化, 会改变其对土壤C输入(Liu et al., 2005), 或者间接改变土壤温度(Sayer et al., 2006)和湿度等土壤微环境, 影响植物生长(Haeussler & Kabzems, 2005; Matsushima & Chang, 2006)。前人的研究结果表明, 凋落物去除会增加草原地表裸露面积、太阳辐射强度(Weaver & Rowland, 1952)和土壤水分蒸发量(Deutsch et al., 2010), 从而降低草地生产力(Heady, 1956); 而王静等(2013)研究认为凋落物去除对地上生物量无显著影响。因此, 在不同的草原生态系统中, 凋落物去除对草地生产力的影响不同; 凋落物添加加大了太阳辐射拦截和土壤表层热量阻隔作用(Matsushima & Chang, 2006), 同时增加了土壤渗透、减少了地表径流和土壤水分蒸发(Sayer et al., 2006), 对草地生产力有显著的促进作用(王静等, 2010)。目前, 凋落物研究主要集中在对群落结构组成、地上生物量以及土壤湿度的影响方面, 而有关凋落物去除和添加对碳通量的影响鲜见报道。在内蒙古典型草原成熟生态系统中, 长期围封会导致凋落物大量积累, 进而引起群落内物种多样性下降(Patrick et al., 2008), 同时也会影响草地生产力和植物种类组成(Knapp & Seastedt, 1986); 在退化生态系统中, 长期重度放牧使凋落物不能有效地积累, 导致生态系统微环境及土壤理化性质恶化, 加剧水土流失, 造成土壤中有机质含量降低及有机质在土壤中分布的变化, 影响地上植物生长, 从而影响植物群落的光合作用和呼吸作用。本实验运用静态箱法研究凋落物去除和添加处理下成熟群落和退化群落的碳通量, 提出两个科学问题: 1)在两种群落(成熟、退化)中, 生态系统碳通量对凋落物去除和添加是如何响应的?2)影响两种群落生态系统碳通量的主要因子有哪些?凋落物积累过多或者积累过少都会降低草地生产力, 为此本文以内蒙古典型草原成熟群落和退化群落为研究对象, 通过对成熟群落过多的凋落物去除, 添加到退化群落的处理中, 我们预期这样能够解除成熟群落凋落物过度积累和退化群落凋落物积累过少对草地生产力的限制, 从而增加生态系统固碳潜力, 研究结果可为合理利用凋落物、改善草地生态系统以及加速草地恢复提供理论依据。
1 材料和方法
1.1 研究地区概况
研究地区位于内蒙古自治区锡林郭勒盟锡林河流域中游白音锡勒牧场境内。样地设置在中国科学院内蒙古草原生态系统定位研究站(IMGER)的1999年围封样地西南侧, 海拔约1 224 m。成熟群落样地(43.93° N, 117.12° E)从1999年至2013年围封14年, 围封面积约25 hm2, 样方内物种有16种, 盖度90%; 退化群落样地(43.93° N, 117.12° E)从1999年至2013年处于围栏外, 自由放牧, 放牧强度约为2只羊·hm-2, 样方内物种有9种, 盖度为45%。该区域属于温带半干旱大陆性气候, 冬季寒冷干燥, 夏季温和湿润。年平均气温0.3 ℃, 年降水量296.8 mm。年际间与年内降水量变异很大, 一年当中降水主要集中在5-10月, 约占全年降水量的93%。土壤类型为典型栗钙土, 多为砂壤质地。样地内建群种为旱生根茎型禾草羊草(Leymus chinensis)和丛生禾草大针茅(Stipa grandis), 优势种为羽茅(Achnatherum sibiricum)、冰草(Agropyron cristatum)、黄囊薹草(Carex korshinskyi)和糙隐子草(Cleistogenes squarrosa)。2014和2015年连续两年进行凋落物去除和添加实验, 2014-2015年日平均气温与日降水量季节动态均为单峰曲线(图1), 年平均气温1.9 ℃, 7月最高气温25.1 ℃, 1月最低气温-24.1 ℃; 与1985-2015年年降水量(296.8 mm)相比, 2014年为较湿润年(年降水量333.6 mm), 而2015年则为过去30年中较干旱的一年(年降水量269.8 mm)。
图1
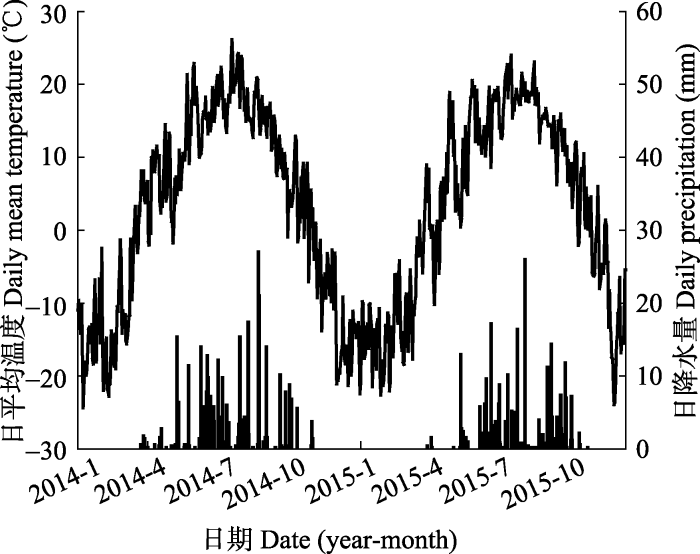 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12014-2015年日平均温度(曲线形)与日降水量(柱形) 动态。
Fig. 1Dynamics of daily mean temperature (curve) and daily precipitation (bars) in 2014 and 2015.
1.2 实验设计
成熟群落样地于1999年围封后一直处于无放牧自然恢复状态, 有大量凋落物积累。凋落物过度积累会影响种子萌发和幼苗更新, 导致群落结构和物种组成改变、物种多样性和草地生产力下降。凋落物去除实验于2013年和2014年的10月中旬进行, 在成熟群落样地内连续处理两年。群落组成以大针茅、羊草、羽茅和冰草为主要优势种。本实验采用裂区设计中的完全随机区组设计。在成熟群落样地中设置4水平随机区组, 包括对照、凋落物0去除、凋落物去除50%、凋落物去除100%; 每个水平设置6个重复, 共24个处理。实验小区面积3 m × 3 m, 各小区间设1 m缓冲带。凋落物0去除是对2013年和2014年围封群落同面积的所有凋落物进行剪碎但不去除处理(包括立枯), 无凋落物去除且无剪碎处理作为局部对照(表1)。Table 1
表1
表12013-2014年成熟群落不同处理下凋落物去除量(g·m-2)
Table 1
| 处理 Treatment | 2013 | 2014 |
|---|---|---|
| 对照 Control | 0 | 0 |
| 剪碎凋落物, 不移除0, litter cut but not removed | 0 | 0 |
| 凋落物去除50% 50% litter removal | 333 | 61 |
| 凋落物去除100% 100% litter removal | 667 | 122 |
新窗口打开|下载CSV
同时, 退化群落样地于1999年至2013年处于冬春放牧状态, 凋落物生产和积累较少。过少的凋落物导致地表裸露面积增大, 生态微环境改变、土壤理化性质恶化, 进而使物种组成改变、物种多样性和草地生产力下降(Bai et al., 2010)。凋落物添加实验于2013年和2014年的10月中旬进行, 在成熟群落样地内连续处理两年。群落组成以大针茅、羊草、黄囊薹草和糙隐子草为主要优势种。在该样地中设置3水平随机区组: 对照(control)、凋落物添加50%、凋落物添加100%; 每个水平设置6个重复, 共18个处理。实验小区面积3 m × 3 m, 各小区间设1 m缓冲带。把成熟群落去除50%小区的凋落物完全添加到退化群落同面积添加50%的小区中即为凋落物添加50%; 把成熟群落去除100%的小区凋落物完全添加到放牧群落同面积添加100%的小区中即为凋落物添加100% (表2)。各实验小区进行1次凋落物移除和添加处理。2014和2013年对凋落物的处理相同, 但是两年的凋落物量有差别。
Table 2
表2
表22013-2014年退化群落不同处理下每平方米凋落物的添加量(g·m-2)
Table 2
| 处理 Treatment | 2013 | 2014 |
|---|---|---|
| 对照 Control | 0 | 0 |
| 凋落物添加50% 50% litter addition | 333 | 61 |
| 凋落物添加100% 100% litter addition | 667 | 122 |
新窗口打开|下载CSV
1.3 实验方法
1.3.1 土壤温度和湿度的测定在测定生态系统碳通量的同时, 用智能数字测温仪(TH-212, 北京海创高科科技公司)测定土壤10 cm深度的温度。土壤水分使用TDR300土壤水分速测仪(MKY, Aurora, USA)测定。
1.3.2 地上生物量的测定
2014和2013年8月底进行地上生物量测定。本实验在每个实验小区内随机放置一个1 m × 0.5 m的样方, 估算样方内植物总物种分盖度、各物种的分盖度, 并记录各物种株丛数、营养高度、生殖高度, 以及植株长度和宽度。然后将样方内所有物种的地上部分按物种贴地面剪断并收获, 随后将所有物种放在烘箱烘干至恒质量后称量, 对于每个处理计算6个重复生物量干质量和物种数的平均值。
1.3.3 净生态系统CO2交换(NEE)测定
NEE利用LI-840红外线分析仪(LI-COR, Lincoln, USA)和密闭式箱法测定。在2014年8-9月和2015年5-9月, 挑选天气晴朗的日子, 在8:00-12:00进行NEE测定, 每个月测定2-3次。在每个样方永久设置一个0.5 m × 0.5 m的铝合金框, 一端砸入土壤 3 cm, 另一端向上保持平整。测定时将一个0.5 m × 0.5 m × 0.5 m的透明有机玻璃箱放置在铝合金框上, 保持整个空间密闭。箱内两个顶角有两个小风扇。箱内还装有一个测定箱内气体温度的数显温度计(DTM-280, 上海双桥仪器仪表厂)。对CO2浓度连续测定60 s, 为保证数据质量, 在CO2浓度开始稳定下降时记录并保存数据。完成NEE测定之后, 抬起箱子使其内部气体与外界一致, 再重新放在铝合金框上并在箱子上盖有内黑外白的不透明布以遮挡光照, 当CO2浓度开始稳定上升时, 开始记录数据且持续60 s, 此时测定生态系统呼吸(ER)。用NEE和ER的差值计算总生态系统生产力(GEP)。
1.4 数据分析方法
统计分析采用SPPS 17.0软件, 对不同处理下的GEP、ER、NEE、生物量、土壤水分、土壤温度进行单因素方差分析, 并对不同处理不同测定时间的GEP、ER、NEE进行重复测量并进行方差分析。用相关分析检验GEP、ER、NEE分别与土壤温度、土壤水分和地上生物量间的相关系数和显著性水平, 显著性水平为p < 0.05, 极显著水平为p < 0.001。通过SigmaPlot 12.5软件作图。2 结果和分析
2.1 凋落物去除和添加处理对土壤温度和湿度的影响
2.1.1 凋落物去除和添加处理对土壤温度的影响对凋落物去除的成熟群落中对照和0处理进行t检验, 结果表明对照处理和0处理之间无显著性差异。
2015年生长季土壤温度随季节变化明显, 最高值出现在7月13日(图1)。成熟群落中, 在不同的凋落物去除强度下(凋落物去除50%和100%)土壤温度总体呈上升趋势, 2015年不同强度的凋落物处理下, 土壤温度显著高于对照。连续两年凋落物去除100%处理的土壤温度季节均值与对照相比显著增加了 0.8 ℃ (图2A); 退化群落中, 在不同强度凋落物添加处理下(凋落物添加50%和100%), 土壤温度与凋落物添加量显著负相关, 连续两年的凋落物添加处理下土壤温度显著低于对照处理(p < 0.05)。从土壤温度的季节均值可以看出, 凋落物添加50%和100%均与对照处理有显著性差异(p < 0.05), 与对照相比分别降低了1.5和2.0 ℃ (图2B)。这些结果表明: 在成熟群落中, 凋落物去除可导致土壤温度升高, 第一年凋落物去除50%与对照有显著性差异, 凋落物去除50%和100%之间没有显著差异; 在退化群落中, 凋落物添加可导致土壤温度降低, 第一年和连续两年凋落物添加50%和100%与对照均有显著差异(p < 0.05)。
图2
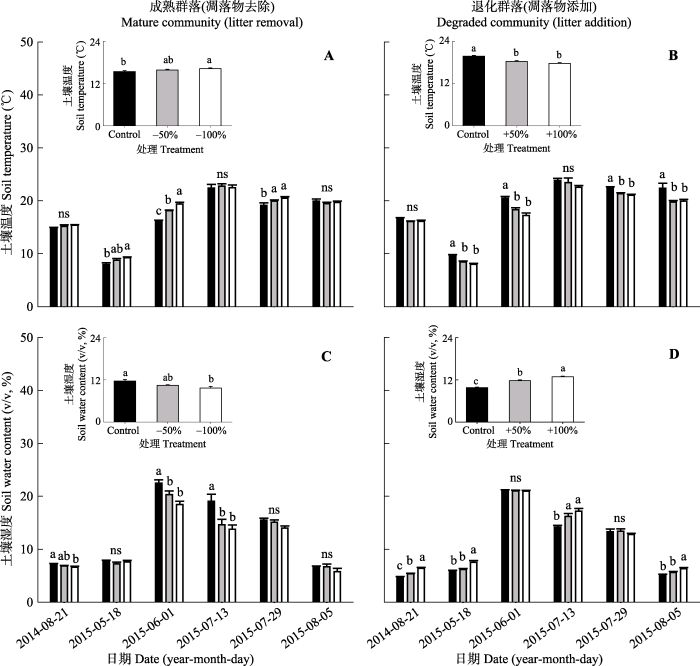 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图22014-2015年成熟群落凋落物去除和退化群落凋落物添加对土壤温度和土壤水分的影响(平均值±标准误差)。不同小写字母a、b、c表示同一群落不同处理间差异显著(p < 0.05)。ns表示处理之间差异不显著(p > 0.05)。Control, 对照; -50%, 凋落物去除50%; -100%, 凋落物去除100%; +50%, 凋落物添加50%; +100%, 凋落物添加100%。
Fig. 2Effects of litter removal in the mature community and litter addition in the degraded community on soil temperature and soil moisture in 2014 and 2015 (mean ± SE). Different small letters a, b and c indicate significant differences among treatments in the same community (p < 0.05). ns indicates no significant differences among treatments (p > 0.05). -50%, litter removal of 50%; -100%, litter removal of 100%; +50%, 50% litter addition; +100%, 100% litter addition.
2.1.2 凋落物去除和添加处理对土壤湿度的影响
2015年生长季土壤湿度随季节变化明显, 土壤湿度最高值出现在6月(图2C、2D)。对两种群落和凋落物处理的结果进行单因素方差分析, 结果表明: 成熟群落中, 随着凋落物去除量的增加土壤湿度呈下降趋势(图2C)。连续两年凋落物去除处理的土壤湿度显著低于对照, 凋落物去除50%和100%处理的土壤湿度分别降低了10.3%和16.3% (图2C); 对于凋落物添加的退化群落, 随着凋落物添加量的增加土壤湿度呈上升趋势。连续两年凋落物添加的土壤湿度显著增加(p < 0.05), 凋落物添加100%处理与对照相比, 增加了9.2% (图2D)。
2.2 凋落物的去除和添加处理对地上生物量的影响
对两种群落凋落物处理的地上生物量进行方差分析, 结果表明: 对于凋落物去除的成熟群落, 一年去除和连续两年去除, 地上生物量均呈下降趋势, 且去除50%和去除100%凋落物与对照相比均显著下降(p < 0.05), 与对照相比分别下降33.91和44.43 g·m-2。对于凋落物添加的退化群落, 一年添加, 地上生物量无显著影响, 连续两年添加, 添加50%凋落物处理显著高于对照, 与对照相比提高25.80 g·m-2, 而添加100%凋落物处理对地上生物量无显著影响(图3)。图3
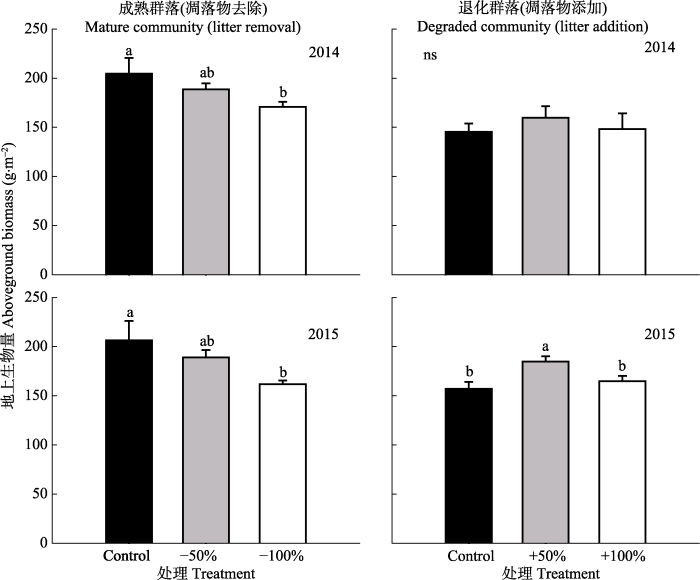 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3成熟群落凋落物去除和退化群落凋落物添加对地上生物量的影响(平均值±标准误差)。不同小写字母a、b表示同一群落不同处理间差异显著(p < 0.05)。ns表示处理之间差异不显著(p > 0.05)。Control, 对照; -50%, 凋落物去除50%; -100%, 凋落物去除100%; +50%, 凋落物添加50%; +100%, 凋落物添加100%。
Fig. 3Effects of litter removal in the mature community and litter addition in the degraded community on aboveground biomass (mean ± SE). Different small letters a and b indicate significant differences among treatments in the same community (p < 0.05). ns indicates no significant differences among treatments (p > 0.05). -50%, litter removal of 50%; -100%, litter removal of 100%; +50%, 50% litter addition; +100%, 100% litter addition.
2.3 凋落物去除和添加处理对GEP、ER、NEE的影响
对凋落物去除和添加的两种群落的GEP、ER、NEE进行方差分析的结果表明: 对于凋落物去除的成熟群落, 不同凋落物的去除强度对GEP、ER的影响均无显著差异, 对NEE有极显著性影响(p < 0.01) (表3); 对于添加凋落物的退化群落, 不同凋落物添加强度对ER的影响均无显著差异, 对NEE有显著性影响(p < 0.05)(表3)。不同测定时间, 两种群落的GEP、ER、NEE均具有极显著差异(p < 0.001)。同时, 监测时间与不同凋落物去除处理的交互作用对两种群落的ER无显著性影响, 对GEP、NEE有极显著性影响(p < 0.001)。Table 3
表3
表3凋落物去除和添加对生态系统总初级生产力(GEP)、生态系统呼吸(ER)和净生态系统CO2交换(NEE)影响的重复测量方差分析
Table 3
| 指标 Index | 因素 Factor | 自由度 Degree of freedom | p值 p value | |
|---|---|---|---|---|
| 成熟群落 Mature community | 退化群落 Degraded community | |||
| GEP | 时间 Time | 5 | 0.000**** | 0.000**** |
| 处理 Treatment | 2 | 0.132 | 0.037* | |
| 时间×处理 Time × treatment | 10 | 0.000**** | 0.000**** | |
| ER | 时间 time | 5 | 0.000**** | 0.000**** |
| 处理 Treatment | 2 | 0.605 | 0.709 | |
| 时间×处理 Time × treatment | 10 | 0.047* | 0.189 | |
| NEE | 时间 Time | 5 | 0.000**** | 0.000**** |
| 处理 Treatment | 2 | 0.008** | 0.018* | |
| 时间×处理 Time × treatment | 10 | 0.000**** | 0.013* | |
新窗口打开|下载CSV
从两种群落凋落物去除和添加处理下GEP、ER和NEE的季节动态(图4)可以看出, 对于凋落物去除的成熟群落, 总体来看, 在2015年生长季早期(4-5月), 凋落物去除显著增加了GEP和NEE。在生长季中期(6-7月)凋落物去除显著降低了GEP和NEE; 对于凋落物添加的退化群落, 在2015年生长季早期, 凋落物添加显著降低GEP和NEE。在生长季中期, 添加50%凋落物对GEP无影响, 而添加100%凋落物显著增加GEP。凋落物去除和添加处理对两种群落ER的影响均无显著差异。
图4
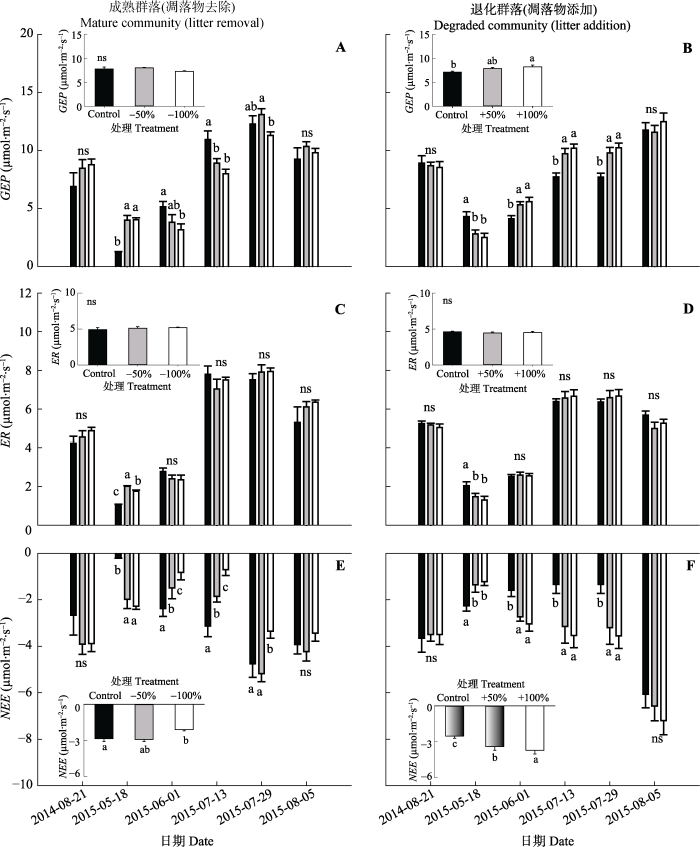 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4两种群落凋落物去除和添加处理下生态系统总初级生产力(GEP)、生态系统呼吸(ER)和CO2净交换(NEE)的季节动态。不同小写字母a、b、c表示相同测定时间下不同处理间差异显著(p < 0.05)。ns表示处理之间差异不显著(p > 0.05)。 Control, 对照; -50%, 凋落物去除50%; -100%, 凋落物去除100%; +50%, 凋落物添加50%; +100%, 凋落物添加100%。
Fig. 4Seasonal dynamics of net ecosystem CO2 exchange (NEE), ecosystem respiration (ER) and gross ecosystem productivity (GEP) under litter removal and addition treatments in the two communities. Different small letters a, b, and c indicate significant differences among treatments (p < 0.05). ns indicates no significant difference among treatments (p > 0.05). -50%, litter removal of 50%; -100%, litter removal of 100%; +50%, 50% litter addition; +100%, 100% litter addition.
对2015年凋落物去除的成熟群落和凋落物添加的退化群落的不同季节GEP、ER、NEE均值进行单因素方差分析的结果表明: 在成熟群落中, 连续两年凋落物去除的NEE随凋落物去除量的变化先增后减, 凋落物去除100%和50%处理间有显著差异 (p < 0.05)(图4)。与对照相比, 凋落物去除50%处理下NEE增加了24.1%。凋落物去除100%处理下NEE降低了26.4%。与NEE相似, GEP随着凋落物去除量的变化先增后减, 但两个水平凋落物去除处理间无显著差异; 在凋落物添加的退化群落中, 连续两年添加凋落物处理下, 不同季节的NEE均值呈上升趋势, 凋落物添加50%和100%与对照有显著性差异, 与对照相比分别提高了35%和47.3%。GEP与NEE的变化规律相似, 添加凋落物增加了GEP, 凋落物添加100%处理下GEP显著增加了15.2%。凋落物去除和添加处理下两种群落在不同季节的ER的平均值有显著性差异。这些结果表明凋落物去除和添加处理对NEE的影响主要是通过影响两种群落的GEP而产生的。凋落物去除100%处理降低了GEP和NEE; 随着凋落物量的增加, GEP和NEE也相应增加。
2.4 生态系统碳交换与土壤温度、土壤湿度的相关性分析
如图5所示, 对两种群落凋落物去除和添加处理下GEP、ER、NEE与土壤温度进行相关性分析的结果表明: 在成熟群落中, 对照和不同强度的凋落物去除处理下GEP、ER均与土壤温度呈线性正相关关系(p < 0.001), NEE则随土壤温度的升高先增后减, NEE拐点约在17 ℃。在生长季早期和末期, 土壤温度小于17 ℃, NEE与土壤温度呈线性正相关关系; 在生长季中期, 土壤温度大于17 ℃, 这时NEE与土壤温度呈线性负相关关系。在凋落物添加的退化群落中, GEP和ER均与土壤温度呈线性正相关关系(p < 0.001), NEE则随着土壤温度升高先增大后减小, 拐点约在20 ℃。在生长季中期, 土壤温度一般大于20 ℃, 即在生长季中期NEE与土壤温度呈线性负相关关系。对凋落物去除的成熟群落和凋落物添加的退化群落的GEP、ER、NEE和土壤湿度进行方差分析, 结果均无显著差异。图5
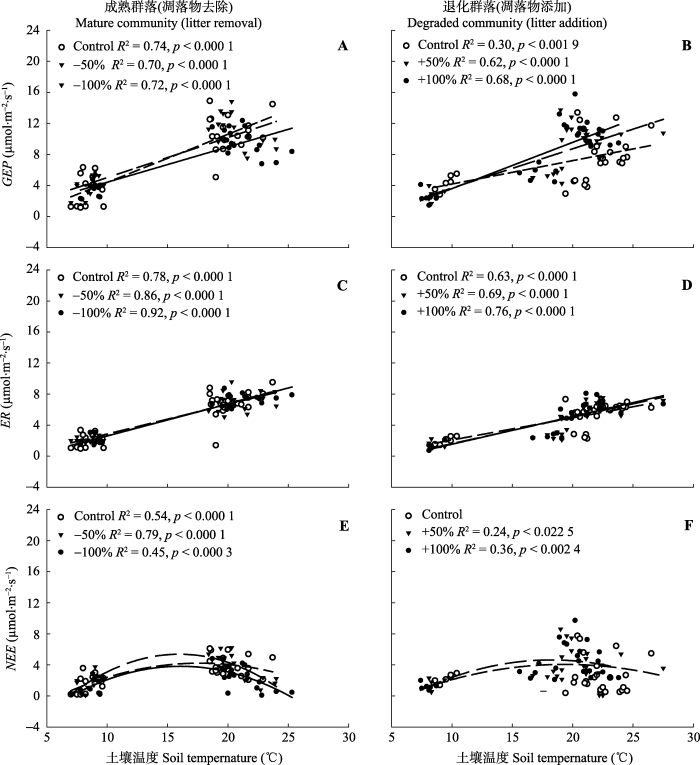 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5凋落物去除和添加两种群落生态系统总初级生产力(GEP)、生态系统呼吸(ER)和净生态系统CO2交换(NEE)与土壤温度的相关性分析。Control, 对照; -50%, 凋落物去除50%; -100%, 凋落物去除100%。+50%, 凋落物添加50%; +100%, 凋落物添加100%。
Fig. 5Correlation analysis of the two communitiess of net ecosystem CO2 exchange (NEE), ecosystem respiration (ER) and gross ecosystem productivity (GEP) and soil temperature under litter removal and addition treatments. -50%, litter removal of 50%; -100%, litter removal of 100%; +50%, 50% litter addition; +100%, 100% litter addition.
3 讨论
凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(Waring & Schlesinger, 1985)。凋落物对生态系统碳通量的影响是一个复杂的生态过程, 凋落物通过多种直接或间接途径实现对生态系统碳通量的影响。目前, 凋落物去除和添加对不同功能群生产力、土壤呼吸、土壤氮矿化以及土壤微生物等方面的影响已有报道, 更多研究集中在森林生态系统, 如张灏等(2014)基于亚热带森林地表CO2通量测定数据发现, 凋落物去除处理显著降低了森林地表CO2通量, 而凋落物加倍处理可以增加森林地表CO2通量。Norby等(2001)则发现凋落物的数量和质量变化可引起地表CO2通量变化, 而碳通量对典型草原凋落物去除和添加的响应规律报道很少。在本实验中, 成熟群落凋落物去除处理显著降低了NEE; 退化群落凋落物添加处理显著增加了NEE, 这与张灏等(2014)的研究结果一致。在成熟群落中, 凋落物去除50%对GEP和NEE是正效应, 去除100%对GEP和NEE是负效应。凋落物去除和添加是通过直接影响两种群落的GEP, 进而间接影响NEE, 这与NEE的变化受ER影响的观点(初小静和韩广轩, 2015)不符, 其原因可能是: 不同生态系统和不同土地利用方式, 凋落物去除和添加所引起的土壤环境变化和有机物分解不同。3.1 凋落物去除和添加处理对土壤温度和土壤湿度的影响
本研究的成熟群落中, 凋落物去除100%、凋落物去除50%处理和对照相比, 土壤温度有显著差异。季节温度均值也能证实: 凋落物去除100% >凋落物去除50%>对照, 这与Xu等(2013)的研究结果相一致, 凋落物去除会增加草原地表温度; 对于凋落物添加的退化群落, 凋落物添加100%和凋落物添加50%的土壤温度均显著低于对照, 季节动态同样也可以证实: 对照>凋落物去除50% >凋落物去除100%。由于凋落物主要是通过对土壤表层保温和热量阻隔缓冲大气温度的作用(李化山等, 2014), 以及太阳辐射拦截影响土壤温度, 凋落物移除增加了土壤表层的有效光辐射, 使地表温度升高。凋落物作为铺在地表的绝缘层起到遮阴作用, 可以减少阳光对土壤表面直接照射, 使地表温度降低, Weaver和Rowland (1952)发现有凋落物覆盖的地表温度要比裸露地表温度低8 ℃。NEE在生长季早期和末期与土壤温度呈线性正相关关系, 在生长季中期与土壤温度呈线性负相关关系。凋落物通过影响渗透、蒸发和径流来改变土壤湿度。在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与Xu等(2013)研究报道的凋落物去除使土壤湿度降低的结果相一致。凋落物连续两年去除间的土壤湿度有显著性差异, 且对照>凋落物去除50%>凋落物去除100%, 凋落物去除50%和凋落物去除100%分别比对照组降低了10.3%和16.3% (图2C)。凋落物连续两年添加的土壤湿度表现出: 凋落物添加100%>凋落物添加50%>对照, 凋落物添加100%与对照有显著差异, 比对照提高了9.2% (图2D)。本研究结果与王光军等(2009)发现的凋落物去除和添加可以相应减少和增加土壤湿度的结论相吻合。导致上述结果的原因是, 成熟群落围封13年, 已有大量的凋落物积累, 凋落物去除增加了地表裸露面积。在有少量降雨时, 较大的地表裸露面积有助于水分入渗到土壤, 但也加速了土壤水分蒸发。较大的降雨量会对缺乏凋落物层保护的地表产生击溅和冲刷水力侵蚀, 从而引起地表径流(薛立等, 2009), 使水分不能有效保持, 最终导致土壤湿度降低。添加凋落物增加了退化群落的土壤湿度。由于凋落物的存在, 有效地减少了地表径流, 同时减少了雨水对土壤的滴溅作用, 阻止了土壤颗粒分散, 使土壤保持良好的团聚结构(Dyksterhuis & Schmutz, 1947), 有利于土壤水分渗透。但是凋落物对土壤水分影响具有双重性, 对较大量的降水主要表现为蒸发限制(Deutsch et al., 2010), 而对少量降雨, 凋落物通过截流、吸水和持水作用来减少可利用水分, 导致土壤湿度降低(Pierson et al., 2002)。从图1可知, 2015年月平均降水量不大, 但较大量降雨次数较多, 凋落物主要表现为蒸发限制, 增加了土壤湿度。
3.2 凋落物去除对GEP、ER和NEE的影响
大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(Xu et al., 2004; Li et al., 2005)。在本研究中, 凋落物连续去除的成熟群落气体交换变化规律为: 从整个生长季来看, 对GEP而言, 凋落物去除50% >对照>凋落物去除100%; 对NEE而言, 凋落物去除50%>对照>凋落物去除100%, 凋落物去除100%和去除50%之间差异显著。对NEE而言, 凋落物去除显著改变了GEP, 但对ER没有显著影响, 从而达到改变NEE的效果。凋落物去除显著增加了土壤温度(图2A), 因GEP与土壤温度呈显著正相关关系(p < 0.001), 随着土壤温度升高而增大, 但是GEP的变化规律是先增后减, 说明土壤温度不是影响GEP的主要因子。这一结论与前人研究结果(Oberbauer et al., 2007)不一致, 是因为增温同时增加了GEP和NEE。凋落物去除使土壤湿度显著降低。有****指出干旱导致生产力和生态系统NEE降低, 而且还可能使生态系统由碳汇变为碳源(Hunt et al., 2002), 本研究结果与之相一致(图4A、4E)。地上生物量因土壤湿度降低而显著降低。Xu等(2004)发现NEE与地上生物量呈正相关关系, 生物量越小, NEE的值越大, 本研究结果与之相吻合(图3, 图4A、图4E)。此外, 凋落物去除带走了部分养分, 降低了土壤中总氮、无机氮和铵态氮含量(Xu et al., 2013), 也可能降低GEP和NEE。凋落物去除50%显著增加了NEE, 这一结论与先前报道(Niu et al., 2009)一致。其原因可能是凋落物去除促进了种子萌发和幼苗更新(Ruprecht & Szabo, 2012)、解除了光限制, 增加了光合作用(Xu et al., 2004)。ER主要受土壤温度影响(Falge et al., 2002), ER与土壤温度呈显著线性正相关关系(p < 0.001), 随着土壤温度升高而增加(图5C)。由于凋落物去除导致土壤湿度降低, 在土壤湿度较低的情况下, ER明显降低(Xu et al., 2004)。ER对去除凋落物响应的影响不显著, 可能是因为土壤温度对ER的正作用抵消了负作用(张文丽等, 2008)。这些结果意味着凋落物去除主要改变土壤湿度, 进而影响GEP和NEE。凋落物去除使养分丧失, 从而对NEE和GEP有一定的影响。3.3 凋落物添加对GEP、ER和NEE的影响
从整个生长季来看, 凋落物添加的退化群落气体交换变化规律是: 对GEP而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物添加100%与对照有显著差异; 对NEE而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物去除100%、凋落物去除50%与对照有显著差异。添加凋落物显著降低了土壤温度。NEE在生长季中期与土壤温度呈线性负相关关系(图5F), 土壤温度越低, NEE越大。因为内蒙古典型草原属于干旱半干旱地区, 水分是影响该地区生态系统结构和功能的关键因素(Bai et al., 2008), 所以土壤温度并不是影响GEP和NEE的驱动因子。凋落物添加可以使土壤湿度增加(图2D), 从而增加了NEE (Niu et al., 2008), 本研究得到类似结果(图4B、图4F)。由于土壤湿度增加, 地上生物量也显著增加(图3)。地上生物量与NEE呈正相关关系, 即NEE随着地上生物量增加而增加(图4F)。此外, 凋落物添加可能促进了凋落物分解, 使土壤无机氮含量增加(Sayer & Tanner, 2010), 添加氮能显著提高GEP, 但对ER无显著影响, 使得NEE显著增加(Xia et al., 2009)。对ER而言, 凋落物添加增加了土壤温度(图2B), 且ER与土壤温度呈正相关关系(图5D)。凋落物添加增加了土壤湿度(图2D), 且ER与土壤湿度为线性正相关关系(Xia et al., 2009)。ER对去除凋落物的响应不显著, 可能是由于土壤含水量对ER的正作用抵消了土壤温度对ER的负作用。这些结果说明, 凋落物添加主要影响土壤湿度, 进而影响GEP和NEE。凋落物添加所带来的养分对NEE和GEP也有一定的促进作用。对内蒙古典型草原凋落物去除和添加的研究表明: 成熟群落的凋落物去除解除了凋落物过度积累对草地生产力的限制, 同时, 把去除的凋落物添加到退化群落中, 有助于快速恢复退化群落的植被, 并使草地植被正向演替。合理管理地上凋落物对陆地生态系统功能和服务调控有重大意义。在成熟群落中保留适量的凋落物、在退化群落中添加适量的凋落物, 对维持典型草原草地结构和功能、加速草地的恢复以及防止草地退化都具有重要的意义。
4 结论
4.1 在成熟群落中, 在两个生长季早期和末期, 凋落物去除处理增加了GEP和NEE, 在生长季中期则降低了GEP和NEE。在退化群落中, 在两个生长季早期和末期, 凋落物添加处理降低了GEP和NEE, 在生长季中期则增加了GEP和NEE。但两种群落ER的季节动态无显著性差异。在两种群落中不同处理间, GEP、ER均与土壤温度呈显著性正相关关系, 但NEE随着土壤温度升高先增后减, 而且在两种群落中有不同的阈值与拐点。两种群落GEP、ER、NEE与土壤湿度均无显著性相关关系。这些结果表明凋落物去除和添加是通过直接影响两种群落的GEP, 进而间接影响NEE。4.2 在凋落物过度积累的成熟群落中, 凋落物在秋季去除对GEP和NEE的影响是正效应。在凋落物匮乏的退化群落中, 凋落物在春季添加对GEP和NEE的影响也是正效应。凋落物在春季的添加避免了对生长季早期光和土壤温度的影响, 同时解除了土壤水分及养分限制, 为植物提供充足的养分及水分, 使其能够快速生长。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/j.1365-2486.2009.02142.xURL [本文引用: 1]

Nitrogen (N) deposition is widely considered an environmental problem that leads to biodiversity loss and reduced ecosystem resilience; but, N fertilization has also been used as a management tool for enhancing primary production and ground cover, thereby promoting the restoration of degraded lands. However, empirical evaluation of these contrasting impacts is lacking. We tested the dual effects of N enrichment on biodiversity and ecosystem functioning at different organizational levels (i.e., plant species, functional groups, and community) by adding N at 0, 1.75, 5.25, 10.5, 17.5, and 28.0 g N m612 yr611 for four years in two contrasting field sites in Inner Mongolia: an undisturbed mature grassland and a nearby degraded grassland of the same type. N addition had both quantitatively and qualitatively different effects on the two communities. In the mature community, N addition led to a large reduction in species richness, accompanied by increased dominance of early successional annuals and loss of perennial grasses and forbs at all N input rates. In the degraded community, however, N addition increased the productivity and dominance of perennial rhizomatous grasses, with only a slight reduction in species richness and no significant change in annual abundance. The mature grassland was much more sensitive to N-induced changes in community structure, likely as a result of higher soil moisture accentuating limitation by N alone. Our findings suggest that the critical threshold for N-induced species loss to mature Eurasian grasslands is below 1.75 g N m612 yr611, and that changes in aboveground biomass, species richness, and plant functional group composition to both mature and degraded ecosystems saturate at N addition rates of approximately 10.5 g N m612 yr611. This work highlights the tradeoffs that exist in assessing the total impact of N deposition on ecosystem function.
DOI:10.1890/07-0992.1URLPMID:18724724 [本文引用: 1]
Understanding how the aboveground net primary production (ANPP) of arid and semiarid ecosystems of the world responds to variations in precipitation is crucial for assessing the impacts of climate change on terrestrial ecosystems. Rain-use efficiency (RUE) is an important measure for acquiring this understanding. However, little is known about the response pattern of RUE for the largest contiguous natural grassland region of the world, the Eurasian Steppe. Here we investigated the spatial and temporal patterns of ANPP and RUE and their key driving factors based on a long-term data set from 21 natural arid and semiarid ecosystem sites across the Inner Mongolia steppe region in northern China. Our results showed that, with increasing mean annual precipitation (MAP), (1) ANPP increased while the interannual variability of ANPP declined, (2) plant species richness increased and the relative abundance of key functional groups shifted predictably, and (3) RUE increased in space across different ecosystems but decreased with increasing annual precipitation within a given ecosystem. These results clearly indicate that the patterns of both ANPP and RUE are scale dependent, and the seemingly conflicting patterns of RUE in space vs. time suggest distinctive underlying mechanisms, involving interactions among precipitation, soil N, and biotic factors. Also, while our results supported the existence of a common maximum RUE, they also indicated that its value could be substantially increased by altering resource availability, such as adding nitrogen. Our findings have important implications for understanding and predicting ecological impacts of global climate change and for management practices in arid and semiarid ecosystems in the Inner Mongolia steppe region and beyond.
DOI:10.1071/BT07151URL [本文引用: 1]
URL [本文引用: 1]
湿地由于具有较高的初级生产力以及较低的有机质降解速率而成为缓解全球变暖的潜在有效碳汇.虽然近年来中国湿地生态系统CO2交换过程及其影响机制研究取得了一系列进展,但尚缺乏对数据进行系统性整合分析.基于29篇文献的数据,对中国21个典型湿地植被净生态系统CO2交换(NEE)、生态系统呼吸(Reco)、总初级生产力(GPP)、NEE的光响应参数以及Reco的温度响应参数进行整合分析,并探讨了这些指标对温度与降雨的响应.结果表明:年尺度上,气温和降雨量对NEE(R2=50%,R2=57%)、GPP(R2=60%,R2=50%)和Reco(R2=44%,R2=50%)均有显著影响(P0.05).生长季尺度上,NEE(R2=50%)、GPP(R2=36%)和Reco(R2=19%)与气温呈显著相关(P0.05);同时NEE(R2=33%)和GPP(R2=25%)也与降雨量呈显著相关(P0.05),但Reco与降雨量的相关关系不显著(P0.05).生长季降雨量与最大光合速率(Amax)之间呈显著相关(P0.01),但与表观量子产率(α)、白天生态系统呼吸速率(Reco,day)无显著相关(P0.05).生长季气温对α、Amax和Reco,day均无显著影响(P0.05).生态系统基础呼吸速率(Rref)与降雨量无显著相关(P0.05),但是生态系统呼吸的温度敏感系数(Q10)与降雨量呈显著的线性负相关(P0.05),同时气温对Q10(R2=0.35)、Rref(R2=0.46)均产生显著影响(P0.05).
URL [本文引用: 1]
湿地由于具有较高的初级生产力以及较低的有机质降解速率而成为缓解全球变暖的潜在有效碳汇.虽然近年来中国湿地生态系统CO2交换过程及其影响机制研究取得了一系列进展,但尚缺乏对数据进行系统性整合分析.基于29篇文献的数据,对中国21个典型湿地植被净生态系统CO2交换(NEE)、生态系统呼吸(Reco)、总初级生产力(GPP)、NEE的光响应参数以及Reco的温度响应参数进行整合分析,并探讨了这些指标对温度与降雨的响应.结果表明:年尺度上,气温和降雨量对NEE(R2=50%,R2=57%)、GPP(R2=60%,R2=50%)和Reco(R2=44%,R2=50%)均有显著影响(P0.05).生长季尺度上,NEE(R2=50%)、GPP(R2=36%)和Reco(R2=19%)与气温呈显著相关(P0.05);同时NEE(R2=33%)和GPP(R2=25%)也与降雨量呈显著相关(P0.05),但Reco与降雨量的相关关系不显著(P0.05).生长季降雨量与最大光合速率(Amax)之间呈显著相关(P0.01),但与表观量子产率(α)、白天生态系统呼吸速率(Reco,day)无显著相关(P0.05).生长季气温对α、Amax和Reco,day均无显著影响(P0.05).生态系统基础呼吸速率(Rref)与降雨量无显著相关(P0.05),但是生态系统呼吸的温度敏感系数(Q10)与降雨量呈显著的线性负相关(P0.05),同时气温对Q10(R2=0.35)、Rref(R2=0.46)均产生显著影响(P0.05).
DOI:10.1016/j.agee.2009.08.002URL [本文引用: 2]
Soil moisture can limit plant production in cool-temperate grasslands, particularly under recent increases in drought severity and predictions of future climate change. This necessitates research that examines moisture mitigation strategies under the dominant land use of grazing. We examined the effectiveness of plant litter in regulating soil moisture dynamics and associated plant production in a native grassland and tame pasture in the Aspen Parkland of Alberta, Canada during 2007/2008, specifically testing (1) how litter and defoliation (as a surrogate for grazing) alter the micro-environment at the soil surface, (2) whether litter, in combination with defoliation, alters forage production, and (3) whether production changes can be linked to observed micro-environmental changes created by litter modification. The full factorial randomized complete block design consisted of three litter treatments (litter removal, in situ control, and double litter) and three defoliation treatments (undefoliated control, light defoliation to 6.5 cm stubble height, and heavy defoliation to 2.5 cm). Soil moisture levels were assessed over four periods after summer precipitation to examine soil moisture depletion. Plant phenological development and canopy cover were also assessed throughout 2007, and biomass was measured in August 2007 and 2008. Abundant litter (up to 14,000 kg ha 1 ) improved soil moisture levels in both native and tame grasslands by either maintaining higher absolute moisture levels or reducing total moisture loss after rainfall. Both litter removal and double litter reduced total growing season production compared to in situ levels at the Native Site, despite highest vegetation cover in litter removal plots. High litter levels also improved growth of individual grass tillers at this site, but negatively impacted sexual production. Conversely litter removal improved overall production at the Tame Site, although litter did enhance grass tiller recovery following mid-season defoliation at both sites, in part due to positive influences on soil moisture. Treatment effects on plant production were less pronounced in 2008 than in 2007. Overall, high litter levels improve soil moisture retention and can improve or stabilize plant production particularly in native prairie sites, and can provide an important drought management strategy in the Aspen Parkland.
DOI:10.1016/S0168-1923(98)00118-XURL [本文引用: 1]
The Bowen ratio/energy balance (BREB) method was used to measure 3002min water vapor and carbon dioxide (CO 2 ) fluxes over three fields dominated by different C 4 grasses (bermudagrass, tallgrass native prairie, and sorghum) at the Blackland Research Center, Temple, TX. Fluxes were related to biotic and abiotic phenomena. Carbon accumulation rates calculated from BREB measurements were compared with those determined from plant biomass measurements. Thirty min CO 2 fluxes were measured continuously from 22 February to 22 November (days 53–326) in 1993 and from 3 March to 22 November (days 62–326) in 1994. Soil CO 2 fluxes were measured periodically in each field and from a bare soil. In all fields, average daily evapotranspiration varied from less than 102mm day 611 in March and November to about 502mm day 611 in June and July. The CO 2 fluxes, which were affected by leaf area, radiation, and soil water content, were near zero in each field in the fall and spring and were maximum in the summer (positive toward surface). Total annual CO 2 flux in the bermudagrass, which was sprigged (planted with grass stolons) in April 1993, was 610.4 and 2.802kg of CO 2 m 612 year 611 (which are equivalent to 610.1 and 0.802kg of carbon (C) m 612 year 611 ) in 1993 and 1994, respectively. The larger flux in the second year was due to greater bermudagrass leaf area. Corresponding annual CO 2 fluxes were 0.2 and 0.302kg02m 612 year 611 (0.05 and 0.0802kg C02m 612 year 611 ) for the prairie, and 0.9 and 610.302kg02m 612 year 611 (0.2 and 610.0902kg C02m 612 year 611 ) for the sorghum. Bare soil CO 2 flux was a small fraction of soil CO 2 flux from all fields, suggesting that a considerable amount of soil CO 2 flux was due to root respiration. The dry matter accumulation rate calculated for each year and field from CO 2 flux measurements was within 20% of the rate calculated from gravimetric biomass measurements. The prairie and sorghum fields were in approximate equilibrium for C storage because estimated annual CO 2 fluxes were near zero. By contrast, the bermudagrass field was a large C sink, primarily in the roots, during the second year. This substantiates other evidence that conversion from continuously cultivated cropland to grassland could create a short-term C sink.
DOI:10.2307/1930949URL [本文引用: 1]
See full-text article at JSTOR
DOI:10.1016/S0168-1923(02)00102-8URL [本文引用: 1]
Differences in the seasonal pattern of assimilatory and respiratory processes are responsible for divergences in seasonal net carbon exchange among ecosystems. Using FLUXNET data ( http://www.eosdis.ornl.gov/FLUXNET) we have analyzed seasonal patterns of gross primary productivity ( F GPP), and ecosystem respiration ( F RE) of boreal and temperate, deciduous and coniferous forests, Mediterranean evergreen systems, a rainforest, temperate grasslands, and C 3 and C 4 crops. Based on generalized seasonal patterns classifications of ecosystems into vegetation functional types can be evaluated for use in global productivity and climate change models. The results of this study contribute to our understanding of respiratory costs of assimilated carbon in various ecosystems. Seasonal variability of F GPP and F RE of the investigated sites increased in the order tropical< Mediterranean< temperate coniferous< temperate deciduous< boreal forests. Together with the boreal forest sites, the managed grasslands and crops show the largest seasonal variability. In the temperate coniferous forests, seasonal patterns of F GPP and F RE are in phase, in the temperate deciduous and boreal coniferous forests F RE was delayed compared to F GPP, resulting in the greatest imbalance between respiratory and assimilatory fluxes early in the growing season. F GPP adjusted for the length of the carbon uptake period decreased at the sampling sites across functional types in the order C 4 crops, temperate and boreal deciduous forests (7.5–8.3 g C m 612 per day)> temperate conifers, C 3 grassland and crops (5.7–6.9 g C m 612 per day)> boreal conifers (4.6 g C m 612 per day). Annual F GPP and net ecosystem productivity ( F NEP) decreased across climate zones in the order tropical>temperate>boreal. However, the decrease in F NEP with latitude was greater than the decrease in F GPP, indicating a larger contribution of respiratory (especially heterotrophic) processes in boreal systems.
DOI:10.1016/S0168-1923(01)00238-6URL [本文引用: 1]
Temperate grassland ecosystems are an important component of the global carbon (C) cycle. The Bowen ratio/energy balance (BREB) technique was used to measure CO 2 fluxes over a mixed-grass prairie at Mandan, ND from 24 April to 26 October in 1996–1999. Above-ground biomass and leaf area index (LAI) were measured about every 21 days throughout the season. Root biomass and soil organic C and nitrogen (N) contents to 1.102m depth were measured in mid-July each year. Peak above-ground biomass typically occurred between mid-July to early-August and ranged from 78202kg02ha 611 in 1998 to 169202kg02ha 611 in 1996. Maximum LAI ranged from 0.3 in 1998 to 0.5 in 1999. Root biomass ranged from 12.702mg02ha 611 in 1996 to 16.202mg02ha 611 in 1997. Maximum daily CO 2 fluxes generally coincided with periods of maximum above-ground biomass. Total CO 2 flux for the 24 April to 26 October period ranged from 18102g02CO 2 02m 612 in 1998 to 47402g02CO 2 02m 612 in 1999 (positive flux is CO 2 movement from atmosphere to plants or soil). The 4-year average flux for this period was 34502g02CO 2 02m 612 . Fluxes during the 1999–2000 dormant season were estimated from BREB (6134902g02CO 2 02m 612 ) and soil flux (6126502g02CO 2 02m 612 ) measurements and when combined with the growing season fluxes, resulted in estimated annual fluxes in 1999 of 125 and 20902g02CO 2 02m 612 for BREB and soil flux dormant season measurements, respectively. These results suggest that the C budget of Northern Great Plains mixed-grass prairie grasslands may be near equilibrium.
DOI:10.1139/x05-133URL [本文引用: 1]
Organic matter removal and reduced soil aeration porosity during logging are important factors influencing the sustained productivity of managed forest ecosystems. We studied the 4-year effect of these factors on diversity and composition of a trembling aspen (Michx.) plant community in northeastern British Columbia, Canada, in a completely randomized experiment with three levels of organic matter removal (tree stems; stems and slash; stems, slash, and forest floor) and three levels of soil compaction (none; intermediate (2-cm impression); heavy (5-cm impression)). Tree stem removal caused the greatest change in species diversity (30% of variance; ANOVA 02≤020.01), increasing the dominance of aspen and (Michx.) Beauv. over other species. Slash removal had little effect. Forest floor removal caused the greatest compositional change (37% of variance; MANOVA 02=020.001), favoring ruderal over bud-banking species. Presence or absence of forest floor better explained these changes than any soil physical or chemical parameter. Although dominance of aspen over was positively correlated with soil aeration porosity (02=020.50, 02=0227, 02<020.001), there were few differences between intermediate and heavy compaction. In this ecosystem, disturbances that reduce forest floor thickness without compacting soils will likely optimize plant species diversity and enhance aspen regrowth.L'enlèvement de la matière organique et la réduction de la macroporosité du sol associés à l'exploitation forestière sont des facteurs importants qui influencent le rendement soutenu des écosystèmes forestiers sous aménagement. Nous avons étudié leurs effets après quatre ans sur la diversité et la composition d'une communauté végétale de peuplier faux-tremble (Michx.) dans le nord-est de la Colombie-Britannique, au Canada. Le dispositif expérimental était complètement aléatoire et comprenait trois niveaux d'enlèvement de la matière organique (seulement la tige des arbres, la tige et les déchets de coupe, la tige, les déchets de coupe et la couverture morte) et trois niveaux de compaction du sol (aucune, intermédiaire (empreinte de 202cm) et forte (empreinte de 502cm). L'enlèvement de la tige des arbres a causé le changement le plus important dans la diversité des espèces (variance de 3002%; valeur de d'ANOVA02≤020,01), entra06nant la dominance du peuplier et de (Michx.) Beauv. sur les autres espèces. L'enlèvement des déchets de coupe a eu peu d'effet. L'enlèvement de la couverture morte a entra06né le changement de composition le plus important (variance de 3702%; valeur de de MANOVA02=020,001), ce qui a favorisé les espèces rudérales plut00t que celles qui ont des bourgeons latents. La présence ou l'absence de couverture morte expliquait ces changements mieux que n'importe quel paramètre physique ou chimique du sol. Bien que la dominance du peuplier sur le soit positivement corrélée avec la macroporosité du sol (02=020,50, 02=0227, 02<020,001), il y avait peu de différences entre la compaction intermédiaire et forte. Dans cet écosystème, les perturbations qui réduisent l'épaisseur de la couverture morte sans compacter le sol devraient optimiser la diversité des espèces végétales et favoriser la régénération de peuplier.[Traduit par la Rédaction]
DOI:10.2307/1933071URL [本文引用: 1]
DOI:10.1016/S0168-1923(02)00006-0URL [本文引用: 1]
For 212 days, including a 117-day-long summer drought with only 45 mm rainfall, fluxes of latent and sensible heat ( λE and H, respectively) and carbon dioxide (CO 2) were measured by eddy covariance over a tussock grassland in New Zealand. The measurement site was grazed and now has a low leaf area index (650.5), dominated by the invasive weed Hieracium, and has a high proportion of bare soil (43%). Energy balance closure was excellent (0.97H+λE)619 ( W m 612)=R a, r 2=0.93, n=8407 half-hourly averages and R a is the available energy. As the drought progressed, the daily average Bowen ratio ( H/ λE) increased from 1 to 15 which corresponded with increasing air and surface temperatures and vapour pressure deficit. Maximum evaporation rate was 3.8 mm per day and it averaged 1.1 mm per day, seasonally equivalent to 93% of the rainfall (241 mm) in accordance with mass conservation, but only 47% of the equilibrium evaporation rate. It was estimated that 50% of soil water was utilised before water deficit began to limit λE and, on average, the evaporative fraction λE/ R a declined exponentially with time after rainfall from 0.43 to 0.05 in 15 days. Maximum net carbon dioxide uptake (denoted negative) and efflux (respiration) rates ( R e) were 615 and 2 μmol m 612 s 611, respectively. These rates are much less than those for grasslands elsewhere. Soil respiration rate and temperature were exponentially related when water was plentiful, but at the same temperature, respiration declined by a factor of 4 as water content ranged from 0.21 (field capacity) to 0.04 m 3 m 613. Soil respiration responded to drought-breaking rainfall and irrigation with a return to maximum rates within 1 day. During spring, when soil water content was >0.10 m 3 m 613 (October and November), the daily CO 2 flux density ( F CO 2) was relatively conservative at 611.2 g C m 612 per day. During drought (January–February), the ecosystem became a carbon source with a mean emission flux of 0.70 g C m 612 per day. Twenty days after a drought-breaking rainfall, the ecosystem returned to being a net sink, but by this time the mean daily air temperature was falling and the first frost of the winter (9 April; 611.8 °C) had occurred thus reducing the ability of the ecosystem to respond to the increased soil water content. Water availability severely limits the carbon acquisition of this ecosystem with the result that during summer, net productivity is closely related to both the frequency and the timing of rainfall events. During summer droughts the inter-montane-basin tussock grasslands may be carbon neutral.
DOI:10.2307/1310387URL [本文引用: 1]
URL [本文引用: 1]
凋落物是土壤呼吸的重要碳源,氮沉降将改变其输入数量和质量,进而影响土壤呼吸。为揭示氮沉降和去除凋落物对土壤呼吸的影响,以太岳山油松林为研究对象,对林地分别作2种凋落物处理:去除凋落物(LR)、对照(CK1),设计4个施氮水平:不施氮(CK2,0 kg N·hm-2·a-1),低氮(LN,50 kg N·hm-2·a-1),中氮(MN,100 kg N·hm-2·a-1)和高氮(HN,150 kg N·hm-2·a-1),于2010—2012年生长季测定土壤呼吸速率的动态变化,并分析土壤呼吸速率与土壤温度、土壤湿度、土壤微生物生物量C、N的关系。结果表明:随着观测年限的推移,模拟氮沉降对对照处理的土壤呼吸速率、去凋处理的土壤呼吸速率、凋落物层呼吸速率的促进作用逐渐减弱。去除凋落物使土壤呼吸速率降低了29.0%,施氮减小了去除凋落物后土壤呼吸速率的变化幅度。土壤呼吸速率与土壤温度均呈显著指数相关(P0.05),土壤温度解释了土壤呼吸速率变异的37.3%~62.2%,去除凋落物降低了模型决定系数R2;以土壤温度和土壤水分构建的复合关系方程拟合效果均好于单因子模型,土壤温度和水分共同解释了土壤呼吸季节变化的67.6%~85.6%,并且施氮降低了去凋处理的复合模型决定系数R2,而对对照处理没有显著影响。施氮提高了土壤微生物生物量C、N,并且土壤微生物生物量C、N与土壤呼吸速率呈显著正相关(P0.05)。说明氮沉降、凋落物是影响油松林土壤CO2通量的两个重要因子。
URL [本文引用: 1]
凋落物是土壤呼吸的重要碳源,氮沉降将改变其输入数量和质量,进而影响土壤呼吸。为揭示氮沉降和去除凋落物对土壤呼吸的影响,以太岳山油松林为研究对象,对林地分别作2种凋落物处理:去除凋落物(LR)、对照(CK1),设计4个施氮水平:不施氮(CK2,0 kg N·hm-2·a-1),低氮(LN,50 kg N·hm-2·a-1),中氮(MN,100 kg N·hm-2·a-1)和高氮(HN,150 kg N·hm-2·a-1),于2010—2012年生长季测定土壤呼吸速率的动态变化,并分析土壤呼吸速率与土壤温度、土壤湿度、土壤微生物生物量C、N的关系。结果表明:随着观测年限的推移,模拟氮沉降对对照处理的土壤呼吸速率、去凋处理的土壤呼吸速率、凋落物层呼吸速率的促进作用逐渐减弱。去除凋落物使土壤呼吸速率降低了29.0%,施氮减小了去除凋落物后土壤呼吸速率的变化幅度。土壤呼吸速率与土壤温度均呈显著指数相关(P0.05),土壤温度解释了土壤呼吸速率变异的37.3%~62.2%,去除凋落物降低了模型决定系数R2;以土壤温度和土壤水分构建的复合关系方程拟合效果均好于单因子模型,土壤温度和水分共同解释了土壤呼吸季节变化的67.6%~85.6%,并且施氮降低了去凋处理的复合模型决定系数R2,而对对照处理没有显著影响。施氮提高了土壤微生物生物量C、N,并且土壤微生物生物量C、N与土壤呼吸速率呈显著正相关(P0.05)。说明氮沉降、凋落物是影响油松林土壤CO2通量的两个重要因子。
DOI:10.1111/j.1365-2486.2005.01047.xURL [本文引用: 3]
Abstract This paper presents results of 1 year (from March 25, 2003 to March 24, 2004, 366 days) of continuous measurements of net ecosystem CO 2 exchange (NEE) above a steppe in Mongolia using the eddy covariance technique. The steppe, typical of central Mongolia, is dominated by C 3 plants adapted to the continental climate. The following two questions are addressed: (1) how do NEE and its components: gross ecosystem production (GEP) and total ecosystem respiration ( R eco ) vary seasonally? (2) how do NEE, GEP, and R eco respond to biotic and abiotic factors? The hourly minimal NEE and the hourly maximal R eco were 613.6 and 1.2μmolm 612 s 611 , respectively (negative values denoting net carbon uptake by the canopy from the atmosphere). Peak daily sums of NEE, GEP, and R eco were 612.3, 3.5, and 1.5gCm 612 day 611 , respectively. The annual sums of GEP, R eco , and NEE were 179, 138, and 6141gCm 612 , respectively. The carbon removal by sheep was estimated to range between 10 and 82gCm 612 yr 611 using four different approaches. Including these estimates in the overall carbon budget yielded net ecosystem productivity of 6123 to +20gCm 612 yr 611 . Thus, within the remaining experimental uncertainty the carbon budget at this steppe site can be considered to be balanced. For the growing period (from April 23 to October 21, 2003), 26% and 53% of the variation in daily NEE and GEP, respectively, could be explained by the changes in leaf area index. Seasonality of GEP, R eco , and NEE was closely associated with precipitation, especially in the peak growing season when GEP and R eco were largest. Water stress was observed in late July to early August, which switched the steppe from a carbon sink to a carbon source. For the entire growing period, the light response curves of daytime NEE showed a rather low apparent quantum yield ( α =610.0047μmolCO 2 μmol 611 photons of photosynthetically active radiation). However, the α values varied with air temperature ( T a ), vapor pressure deficit, and soil water content.
DOI:10.1093/treephys/25.12.1511URLPMID:16137937 [本文引用: 2]
Human activities are increasing the concentrations of atmospheric carbon dioxide ([CO2]) and tropospheric ozone ([O3]), potentially leading to changes in the quantity and chemical quality of leaf litter inputs to forest soils. Because the quality and quantity of labile and recalcitrant carbon (C) compounds influence forest productivity through changes in soil organic matter content, characterizing changes in leaf litter in response to environmental change is critical to understanding the effects of global change on forests. We assessed the independent and combined effects of elevated [CO2] and elevated [O3] on foliar litter production and chemistry in aspen (Populus tremuloides Michx.) and birch-(Betula papyrifera Marsh.) aspen communities at the Aspen free-air CO2 enrichment (FACE) experiment in Rhinelander, WI. Litter was analyzed for concentrations of C, nitrogen (N), soluble sugars, lipids, lignin, cellulose, hemicellulose and C-based defensive compounds (soluble phenolics and condensed tannins). Concentrations of these chemical compounds in naturally senesced litter were similar in aspen and birch-aspen communities among treatments, except for N, the C:N ratio and lipids. Elevated [CO2] significantly increased C:N (+8.7%), lowered mean litter N concentration (-10.7%) but had no effect on the concentrations of soluble sugars, soluble phenolics and condensed tannins. Elevated [CO2] significantly increased litter biomass production (+33.3%), resulting in significant increases in fluxes of N, soluble sugars, soluble phenolics and condensed tannins to the soil. Elevated [O3] significantly increased litter concentrations of soluble sugars (+78.1%), soluble phenolics (+53.1%) and condensed tannins (+77.2%). There were no significant effects of elevated [CO2] or elevated [O3] on the concentrations of individual C structural carbohydrates (cellulose, hemicellulose and lignin). Elevated [CO2] significantly increased cellulose (+37.4%) input to soil, whereas elevated [O3] significantly reduced hemicellulose and lignin inputs to soil (-22.3 and -31.5%, respectively). The small changes in litter chemistry in response to elevated [CO2] and tropospheric [O3] that we observed, combined with changes in litter biomass production, could significantly alter the inputs of N, soluble sugars, condensed tannins, soluble phenolics, cellulose and lignin to forest soils in the future.
DOI:10.1016/j.foreco.2006.09.018URL [本文引用: 2]
Canada bluejoint grass ( Calamagrostis canadensis (Michx.) Beauv.) is a very competitive understory species and can create a thick litter layer that may decrease tree growth in white spruce ( Picea glauca (Moench.) Voss) stands in the boreal region. We examined the effects of Canada bluejoint grass dominated understory competition, N fertilization, and litter layer removal on the nutrition and growth of white spruce, in relation to soil N availability in a 13-year-old plantation near Whitecourt, Alberta, Canada, in 2003 and 2004. We hypothesized that: (1) understory competition reduces white spruce growth as a result of competition for available N; (2) litter layer removal increases white spruce growth by increasing soil temperature and N availability; (3) N fertilization improves white spruce growth by increasing N availability. Soil gravimetric moisture content in LFH was reduced by understory competition and litter layer removal in 2003, which had a very dry summer. Understory removal increased white spruce diameter growth, 100-needle biomass, and needle N concentration and content. In 2003, N fertilization consistently increased 100-needle weight and needle N content when litter layer was removed, but decreased or did not affect those values without litter layer removal. Nitrogen fertilization increased needle N concentration in 2004. The inconsistent effects of N fertilization may be due to the short-term nature of fertilization effects and immobilization of N by organic matter. Rates of white spruce diameter growth and soil mineral N supply in 2004 were related ( R 2 = 0.65, P < 0.001). Vector analysis showed that N fertilization or litter layer removal with intact understory vegetation either increased or did not affect needle N concentration but decreased tree biomass. We conclude that understory control will benefit tree growth in young white spruce stands in the boreal region that have passed the free-to-grow stage. However, N fertilization or litter layer removal alone in bluejoint infested sites may not benefit the trees.
DOI:10.1111/j.1469-8137.2007.02237.xURLPMID:17944829 [本文引用: 2]
090004 Global warming and a changing precipitation regime could have a profound impact on ecosystem carbon fluxes, especially in arid and semiarid grasslands where water is limited. A field experiment manipulating temperature and precipitation has been conducted in a temperate steppe in northern China since 2005. 090004 A paired, nested experimental design was used, with increased precipitation as the primary factor and warming simulated by infrared radiators as the secondary factor. 090004 The results for the first 2 yr showed that gross ecosystem productivity (GEP) was higher than ecosystem respiration, leading to net C sink (measured by net ecosystem CO 2 exchange, NEE) over the growing season in the study site. The interannual variation of NEE resulted from the difference in mean annual precipitation. Experimental warming reduced GEP and NEE, whereas increased precipitation stimulated ecosystem C and water fluxes in both years. Increased precipitation also alleviated the negative effect of experimental warming on NEE. 090004 The results demonstrate that water availability plays a dominant role in regulating ecosystem C and water fluxes and their responses to climatic change in the temperate steppe of northern China.
DOI:10.1890/0012-9658(2001)082[1387:EOFOSP]2.0.CO;2URL [本文引用: 1]
Changes in precipitation and nitrogen (N) deposition can influence ecosystem carbon (C) cycling and budget in terrestrial biomes, with consequent feedbacks to climate change. However, little is known about the main and interactive effects of water and N additions on net ecosystem C exchange (NEE). In a temperate steppe of northern China, a field-manipulated experiment was conducted to evaluate the responses of NEE and its components to improve N and water availability from 2005 to 2008. The results showed that both water and N additions stimulated gross ecosystem productivity (GEP), ecosystem respiration (ER), and NEE. Water addition increased GEP by 17%, ER by 24%, and NEE by 11% during the experimental period, whereas N addition increased GEP by 17%, ER by 16%, and NEE by 19%. The main effects of both water and N additions changed with time, with the strongest water stimulation in the dry year and a diminishing N stimulation over time. When water and N were added in combination, there were non-additive effects of water and N on ecosystem C fluxes, which could be explained by the changes in species composition and the shifts of limiting resources from belowground (water or N) to aboveground (light). The positive water and N additions effects indicate that increasing precipitation and N deposition in the future will favor C sequestration in the temperate steppe. The non-additive effects of water and N on ecosystem C fluxes suggest that multifactor experiments are better able to capture complex interactive processes, thus improving model simulations and projections.
DOI:10.1007/s004420000615URLPMID:24577644 [本文引用: 1]
The results of published and unpublished experiments investigating the impacts of elevated [CO 2 ] on the chemistry of leaf litter and decomposition of plant tissues are summarized. The data do not support the hypothesis that changes in leaf litter chemistry often associated with growing plants under elevated [CO 2 ] have an impact on decomposition processes. A meta-analysis of data from naturally senesced leaves in field experiments showed that the nitrogen (N) concentration in leaf litter was 7.1% lower in elevated [CO 2 ] compared to that in ambient [CO 2 ]. This statistically significant difference was: (1) usually not significant in individual experiments, (2) much less than that often observed in green leaves, and (3) less in leaves with an N concentration indicative of complete N resorption. Under ideal conditions, the efficiency with which N is resorbed during leaf senescence was found not to be altered by CO 2 enrichment, but other environmental influences on resorption inevitably increase the variability in litter N concentration. Nevertheless, the small but consistent decline in leaf litter N concentration in many experiments, coupled with a 6.5% increase in lignin concentration, would be predicted to result in a slower decomposition rate in CO 2 -enriched litter. However, across the assembled data base, neither mass loss nor respiration rates from litter produced in elevated [CO 2 ] showed any consistent pattern or differences from litter grown in ambient [CO 2 ]. The effects of [CO 2 ] on litter chemistry or decomposition were usually smallest under experimental conditions similar to natural field conditions, including open-field exposure, plants free-rooted in the ground, and complete senescence. It is concluded that any changes in decomposition rates resulting from exposure of plants to elevated [CO 2 ] are small when compared to other potential impacts of elevated [CO 2 ] on carbon and N cycling. Reasons for experimental differences are considered, and recommendations for the design and execution of decomposition experiments using materials from CO 2 -enrichment experiments are outlined.
DOI:10.1890/06-0649URL [本文引用: 2]
Climate warming is expected to differentially affect CO2 exchange of the diverse ecosystems in the Arctic. Quantifying responses of M exchange to warming in these ecosystems will require coordinated experimentation using standard temperature manipulations and measurements. Here, we used the International Tundra Experiment (ITEX) standard warming treatment to determine CO2 flux responses to growing-season warming for ecosystems spanning natural temperature and moisture ranges across the Arctic biome. We used the four North American Arctic ITEX sites (Toolik Lake, Atqasuk, and Barrow [USA] and Alexandra Fiord [Canada]) that span 10 degrees of latitude. At each site, we investigated the CO2 responses to warming in both dry and wet or moist ecosystems. Net ecosystem CO2 exchange (NEE), ecosystem respiration (ER), and gross ecosystem photosynthesis (GEP) were assessed using chamber techniques conducted over 24-h periods sampled regularly throughout the summers of two years at all sites. At Toolik Lake, warming increased net CO2 losses in both moist and dry ecosystems. In contrast, at Atqasuk and Barrow, warming increased net CO2 uptake in wet ecosystems but increased losses from dry ecosystems. At Alexandra Fiord, warming improved net carbon uptake in the moist ecosystem in both years, but in the wet and dry ecosystems uptake increased in one year and decreased the other. Warming generally increased ER, with the largest increases in dry ecosystems. In wet ecosystems, high soil moisture limited increases in respiration relative to increases in photosynthesis. Warming generally increased GEP, with the notable exception of the Toolik Lake moist ecosystem, where warming unexpectedly decreased GEP >25%. Overall, the respiration response determined the effect of warming on ecosystem CO2 balance. Our results provide the first multiple-site comparison of arctic tundra CO2 flux responses to standard warming treatments across a large climate gradient. These results indicate tha
DOI:10.1007/s11258-007-9369-7URL [本文引用: 1]
Plant litter may play an important role in herbaceous plant communities by limiting primary production and influencing plant species richness. However, it is not known how the effect of litter interacts with fertilization. We tested for the role of litter and fertilization in a large-scale experiment to investigate effects on diversity and biomass of plant species, growth forms, native vs. non-native groups, and abiotic ecosystem components (e.g., soil moisture, PAR). We manipulated plant litter (removed vs. left in situ) and nutrient availability (NPK-fertilized vs. unfertilized) for 4 years in 314-m 2 plots, replicated six times, in an old-field grassland. While many of our species-level results supported previously published studies and theory, our plant group results generally did not. Specifically, grass species richness and forb biomass was not affected by either fertilization or plant litter. Moreover, plant litter removal significantly increased non-native plant species richness. Relative to native plant species, all of our experimental manipulations significantly increased both the biomass and the species richness of non-native plant species. Thus, this grassland system was sensitive to management treatments through the facilitation of non-native plant species. We coupled biotic and abiotic components within a nonmetric multidimensional scaling (NMS) analysis to investigate treatment effects, which revealed that specific treatments altered ecosystem development. These results suggest that fertilization and plant litter may have larger impacts on plant communities and on ecosystem properties than previously understood, underscoring the need for larger-scale and longer-term experiments.
[本文引用: 1]
[本文引用: 1]
DOI:10.2307/4003999URL [本文引用: 1]
There are several generalizations or assumptions concerning rangeland hydrology and erosion relationships found in the literature and in the management arena. These generalizations have found their way into rangeland models, where modelers have assumed that diverse rangeland types can be lumped or averaged together in some way to develop one algorithm or equation to describe a process or relationship across the entire spectrum of rangeland types. These assumptions and modeling approaches based on the universal concept may not be appropriate for diverse rangeland types. This paper presents a comprehensive data set of vegetation, soils, hydrology, and erosion relationships of diverse western rangelands, and utilizes the data to assess the validity of the various assumptions/generalizations for rangelands. The data set emphasizes the difficulty in understanding hydrologic responses on semiarid rangelands, where the relationship between plant/soil characteristics and infiltration/erosion is not well established. When all sites were pooled together, infiltration and sediment production were not correlated with any measured vegetation or soil characteristic. A myriad group of factors determine infiltration and erosion, and is dependent on rangeland type and site conditions. The infiltration and erosion responses and correlation/regression analyses presented highlight the risk of using generalized assumptions about rangeland hydrologic response and emphasize the need to change the current modeling approach. Universal algorithms to represent the response of all rangeland types, such as the pooled multiple regression equations presented, will not provide sufficient accuracy for prediction or assessment of management. We need to develop a rationale to organize rangeland types/vegetation states according to similarities in relationships and responses. These functional rangeland units would assist in the development of more accurate predictive equations to enhance model performance and management of rangelands. DOI:10.2458/azu_jrm_v55i6_pierson
DOI:10.1111/j.1654-1103.2011.01376.xURL [本文引用: 1]
QuestionsLitter quantity in grasslands is highly affected by disturbance regime and influences seed recruitment of constituent species through different mechanisms. Does litter act as a mechanical barrier to burial of freshly shed seeds, and does seed morphology affect this? How is the fate and mobility of seeds affected by litter compared to that of seeds in the seed bank?LocationTransylvanian Lowland, Romania.MethodsWe analysed the seed content of litter and underlying soil collected from six dry grassland sites having different disturbance histories, ranging from sites that are currently grazed to those that have been long abandoned. We related seed content of litter to the litter quantity, seed morphology and the seed content of soil, and the estimated seed production of the above-ground vegetation.ResultsGrass litter represented a natural trap for seeds; we identified significant quantities of seeds of a large number of species (37) within litter samples, and there was a mass effect in the seed trapping by litter higher litter quantities trapped more seeds. As a long-term consequence, we expected that seed bank stores would be gradually depleted in abandoned grassland due to the elimination of seeds by litter, but this was not the case. The higher seed production of the above-ground vegetation in undisturbed sites very probably compensated for seed losses through litter. Seeds retained in the litter were larger, more rounded and had appendages than those in soil. Especially for seeds meeting any of these criteria, litter represents a hazardous medium, since the seeds may fail or have delayed germination.ConclusionsThe role of litter as a seed trap is more relevant in grasslands that have not been disturbed for a long time, where litter seed entrapment can exert a selective pressure on certain species, and thus drive community assembly in grasslands.
DOI:10.1016/j.soilbio.2009.11.014URL [本文引用: 1]
Soil respiration in tropical forests is a major source of atmospheric CO 2 . The ability to partition soil respiration into its individual components is becoming increasingly important to predict the effects of disturbance on CO 2 efflux from the soil as the responses of heterotrophic and autotrophic respiration to change are likely to differ. However, current field methods to partition respiration suffer from various methodological artefacts; root–rhizosphere respiration is particularly difficult to estimate. We used trenched subplots to estimate root–rhizosphere respiration in large-scale litter addition (L+), litter removal (L61) and control (CT)plots in a lowland tropical semi-evergreen forest in Panama. We took a new approach to trenching bymaking measurements immediately before-and-after trenching and comparing them to biweekly measurements made over one year. Root–rhizosphere respiration was estimated to be 38%, 17% and 27% in the CT, L+, and L61 plots, respectively, from the measurements taken immediately before and one day after trenching in May–June 2007. Biweekly measurements over the following year provided no estimates of root–rhizosphere respiration for the first seven months due to decomposition of decaying roots. We were also unable to estimate root–rhizosphere respiration during the dry season due to differences in soil water content between trenched and untrenched soil. However, biweekly measurements taken during the early rainy season one year after trenching (May–June 2008) provided estimates of root–rhizosphere respiration of 39%, 24% and 36% in the CT, L+, and L61 plots, respectively, which are very similar to those obtained during the first day after trenching. We suggest that measurements taken immediately before and one day after root excision are a viable method for a rapid estimation of root–rhizosphere respiration without the methodological artefacts usually associated with trenching experiments.
DOI:10.1016/j.foreco.2006.04.007URL [本文引用: 2]
Differences in forest productivity due to climate change may result in permanently altered levels of litterfall and litter on the forest floor. Using experimental litter removal and litter addition treatments, we investigated the effects of increased and decreased litterfall on early-stage litter decomposition and the abundance of meso-arthropods in a moist tropical forest. Litterbags containing freshly fallen leaves of Cecropia insignis (above and below the litter on the forest floor, and with and without fungicide) and Simarouba amara , or untreated birch wood ( Betula sp.) were placed in either (1) plots where all litterfall was removed monthly (L61); (2) plots where litterfall was doubled monthly (L+), or (3) control plots (CT). Litter removal significantly slowed decomposition of both species and reduced the abundance of meso-arthropods on Simarouba litter. The fungicide treatment did not reduce apparent mass loss of Cecropia leaves. The litter addition treatment accelerated the decay of birch wood, probably because of increased nutrient availability from the extra litter; but there was no change in leaf-litter decomposition or meso-arthropod abundance in the L+ treatment. After 68 days, the concentrations of nitrogen, phosphorus, potassium, and magnesium in partially decomposed Cecropia litter were higher in the L+ treatment and lower in the L61 treatment. The accumulation of phosphorus and nitrogen was greater in the litter in L+ plots and lower in the L61 plots while the release of potassium and magnesium from decomposing litter was lower in the L+ treatment and greater in the L61 plots. Thus, differences in the quantity of litterfall affect decomposition with consequences for carbon and nutrient storage and cycling.
[本文引用: 1]
[本文引用: 1]
DOI:10.11686/cyxb20130604URL
通过对内蒙古典型草原围封23 年的样地进行凋落物去除处理后,连续5年的植被调查研究,探讨凋落物对内蒙古典型草原地上生物量的影响及对群落各功能群生物量的影响。研究结果表明,凋落 物的不同去除处理对同年植被地上生物量没有显著影响(P0.05);相同处理不同年际间地上生物量差异显著(P0.05)。但在处理后的第2年起,草原地 上生物量随着凋落物去除程度的增加而增加。凋落物去除对各个功能群生物量的影响表现在处理后的第5年,中度去除条件下的多年生杂类草生物量明显多于重度去 除处理和对照区(P0.05);多年生丛生型禾草在重度处理区的生物量明显高于中度处理区(P0.05),但二者与对照区均无显著差别(P0.05);其 余各年影响均不显著(P0.05)。通过对各功能群生物量与总的地上生物量的相关分析得出:在重度去除处理下,地上生物量与禾草的地上生物量相关性较高, 多年生杂类草其次。多年生根茎型禾草在中度处理区与地上生物量的相关性最高且显著(P0.05),其次是一、二年生草本。对照区的多年生丛生型禾草与总地 上生物量有显著相关性(P0.05)。
DOI:10.11686/cyxb20130604URL
通过对内蒙古典型草原围封23 年的样地进行凋落物去除处理后,连续5年的植被调查研究,探讨凋落物对内蒙古典型草原地上生物量的影响及对群落各功能群生物量的影响。研究结果表明,凋落 物的不同去除处理对同年植被地上生物量没有显著影响(P0.05);相同处理不同年际间地上生物量差异显著(P0.05)。但在处理后的第2年起,草原地 上生物量随着凋落物去除程度的增加而增加。凋落物去除对各个功能群生物量的影响表现在处理后的第5年,中度去除条件下的多年生杂类草生物量明显多于重度去 除处理和对照区(P0.05);多年生丛生型禾草在重度处理区的生物量明显高于中度处理区(P0.05),但二者与对照区均无显著差别(P0.05);其 余各年影响均不显著(P0.05)。通过对各功能群生物量与总的地上生物量的相关分析得出:在重度去除处理下,地上生物量与禾草的地上生物量相关性较高, 多年生杂类草其次。多年生根茎型禾草在中度处理区与地上生物量的相关性最高且显著(P0.05),其次是一、二年生草本。对照区的多年生丛生型禾草与总地 上生物量有显著相关性(P0.05)。
DOI:10.3773/j.issn.1005-264x.2010.08.003URL [本文引用: 1]
通过连续5年对退化草地进行凋 落物添加处理,分析不同处理强度植物群落动态,探讨凋落物对草地地上和不同功能群生产力的影响。结果表明:凋落物添加对草地地上生产力有显著的促进作用, 草地地上生产力随着凋落物添加强度的增加而变化不大。处理效应主要表现在处理后第一年,重度处理与对照有极显著差异(p0.05),以后几年差异不显著 (p0.05),年度间差异极显著(p0.001)。凋落物添加对植物群落各功能群生产力无显著影响。年度间不同处理各功能群相对生物量随年际变化的主成 分分析(PCA)结果说明:草地生产力主要是由多年生丛生型禾草与多年生杂类草在生态系统中的竞争补偿作用决定的,随着凋落物添加强度的增加,二者的竞争 作用减弱,相关性下降,多年生丛生型禾草占更大优势,预示着多年生杂类草与一二年生草本、灌木、多年生根茎型禾草的竞争补偿作用增大。
DOI:10.3773/j.issn.1005-264x.2010.08.003URL [本文引用: 1]
通过连续5年对退化草地进行凋 落物添加处理,分析不同处理强度植物群落动态,探讨凋落物对草地地上和不同功能群生产力的影响。结果表明:凋落物添加对草地地上生产力有显著的促进作用, 草地地上生产力随着凋落物添加强度的增加而变化不大。处理效应主要表现在处理后第一年,重度处理与对照有极显著差异(p0.05),以后几年差异不显著 (p0.05),年度间差异极显著(p0.001)。凋落物添加对植物群落各功能群生产力无显著影响。年度间不同处理各功能群相对生物量随年际变化的主成 分分析(PCA)结果说明:草地生产力主要是由多年生丛生型禾草与多年生杂类草在生态系统中的竞争补偿作用决定的,随着凋落物添加强度的增加,二者的竞争 作用减弱,相关性下降,多年生丛生型禾草占更大优势,预示着多年生杂类草与一二年生草本、灌木、多年生根茎型禾草的竞争补偿作用增大。
[本文引用: 1]
DOI:10.1086/335741URL [本文引用: 2]
1. An upland prairie, undisturbed by mowing, grazing, or burning for 15 years, was studied. About 80% was covered with a nearly pure stand of Andropogon gerardi. Similar stands of Panicum virgatum and Sporobolus heterolepis each occupied about 9%. 2. A dense natural mulch, 4.5-8 inches deep and weighing 6-9 tons per acre, covered the deep loam soil. The mulch intercepted much precipitation but promoted more rapid infiltration of water which reached the soil and greatly retarded evaporation. It increased the organic matter of the surface soil 1.5-2% and nitrogen content 0.1% of its oven-dry weight. 3. Roots and rhizomes grew thickly below the mulch and mellowed the surface soil. Pore space was 61.7%, and volume-weight was 0.97. 4. Soil temperatures 22090601-28090601 F. lower under the mulch in May delayed growth about 3 weeks compared with plants where the mulch had been removed. Production of flower stalks was considerably later in all mulched grasses and also less in Andropogon. Flower stalks were abundant on all unmulched grasses. 5. Yields in June, July, and August were 53, 26, and 29% less from mulched stands of Andropogon, and 57, 55, and 26% less in Panicum than those from unmulched stands. 6. Consequences of the effects of the mulch upon the environment were the production of a nearly pure, but some-what thinner than normal, stand of Andropogon. The understory characteristic of upland prairie had all but disappeared. The usual mid grasses of upland were few or none. Only a few of the taller forbs remained. 7. Dense stands of Panicum with forbs characteristic of very moist sites were typical. Between the large bunches of prairie dropseed the deeply mulched soil was usually free of vegetation.
DOI:10.11686/cyxb20130402URLMagsci [本文引用: 1]
<p>采用涡动相关技术对2009,2010和2011年疏勒河上游高寒草甸生态系统CO2 通量观测和分析表明,疏勒河上游高寒草甸生态系统CO<sub>2</sub> 通量具有明显的日变化和年变化特征,6月、7月和8月为CO<sub>2</sub> 的强吸收期,4月、5月和10月为CO<sub>2</sub> 的强释放期。计算得到3年的CO<sub>2</sub> 净吸收量分别为134.5,151.3和194.4gCO<sub>2</sub>/m<sup>2</sup>,平均吸收量为160.0gCO<sub>2</sub>/m<sup>2</sup>,在区域起着碳汇的作用。生长季节,净生态系统交换量(netecosystem CO<sub>2</sub>exchange,NEE)与温度、降水量、相对湿度以及地表长波辐射呈负相关,气温在0~7℃范围内NEE 随气温增加线性减小,当温度大于7℃时,NEE 随温度的增加而增大;非生长季节,NEE 与温度、降水量、相对湿度以及地表长波辐射呈正相关。当地表反射率在0.2左右,NEE 呈现快速增长趋势,当反射率超过0.3时,NEE 接近最大值,并保持稳定。</p>
DOI:10.11686/cyxb20130402URLMagsci [本文引用: 1]
<p>采用涡动相关技术对2009,2010和2011年疏勒河上游高寒草甸生态系统CO2 通量观测和分析表明,疏勒河上游高寒草甸生态系统CO<sub>2</sub> 通量具有明显的日变化和年变化特征,6月、7月和8月为CO<sub>2</sub> 的强吸收期,4月、5月和10月为CO<sub>2</sub> 的强释放期。计算得到3年的CO<sub>2</sub> 净吸收量分别为134.5,151.3和194.4gCO<sub>2</sub>/m<sup>2</sup>,平均吸收量为160.0gCO<sub>2</sub>/m<sup>2</sup>,在区域起着碳汇的作用。生长季节,净生态系统交换量(netecosystem CO<sub>2</sub>exchange,NEE)与温度、降水量、相对湿度以及地表长波辐射呈负相关,气温在0~7℃范围内NEE 随气温增加线性减小,当温度大于7℃时,NEE 随温度的增加而增大;非生长季节,NEE 与温度、降水量、相对湿度以及地表长波辐射呈正相关。当地表反射率在0.2左右,NEE 呈现快速增长趋势,当反射率超过0.3时,NEE 接近最大值,并保持稳定。</p>
DOI:10.1111/gcb.2009.15.issue-6URL [本文引用: 3]
DOI:10.1029/2004GB002281URL [本文引用: 3]
[1] In this paper, we analyzed 3 years of carbon flux data from continuous eddy covariance measurements to investigate how soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. The data were acquired over an annual grassland and from the grass understory of an oak/grass savanna ecosystem in California. We observed that ecosystem respiration was an exponential function of soil temperature during the winter wet season and a jump in ecosystem respiration occurred, at comparable temperatures, during the spring growth period. The depletion of the moisture from the soil reservoir, during spring, limited ecosystem respiration after its volumetric water content dropped below a threshold of 0.15 m3 m0908083. The senescence of grass during the summer switched the source of ecosystem respiration to heterotrophic bacteria and fungi. During the summer, respiration proceeded at a low basal rate (about 0.10 to 0.3 g C m0908082 d0908081), except when summer rain events stimulated large dynamic pulses in heterotrophic respiration. Peak respiratory pulses were on the order of 6009000980 times baseline and could not be explained by functions that depend on mean soil moisture and temperature. We found that the magnitude of the respiratory pulses was inversely related to its prerain value and that the time constant, describing the exponential decay of the respiratory pulses after the rain event, was a function of the amount of rainfall. The amount of carbon lost, in association with a few summer rain events, was greater at the site with higher primary productivity and soil carbon content.
DOI:10.5194/bg-10-7423-2013URL [本文引用: 3]
Global change has been shown to alter the amount of above-ground litter inputs to soil greatly, which could cause substantial cascading effects on below-ground biogeochemical cycling. Despite extensive study, there is uncertainty about how changes in above-ground litter inputs affect soil carbon and nutrient turnover and transformation. Here, we conducted a meta-analysis on 70 litter-manipulation experiments in order to assess how changes in above-ground litter inputs alter soil physicochemical properties, carbon dynamics and nutrient cycles. Our results demonstrated that litter removal decreased soil respiration by 34%, microbial biomass carbon in the mineral soil by 39% and total carbon in the mineral soil by 10%, whereas litter addition increased them by 31, 26 and 10%, respectively. This suggests that greater litter inputs increase the soil carbon sink despite higher rates of carbon release and transformation. Total nitrogen and extractable inorganic nitrogen in the mineral soil decreased by 17 and 30%, respectively, under litter removal, but were not altered by litter addition. Overall, litter manipulation had a significant impact upon soil temperature and moisture, but not soil pH; litter inputs were more crucial in buffering soil temperature and moisture fluctuations in grassland than in forest. Compared to other ecosystems, tropical and subtropical forests were more sensitive to variation in litter inputs, as altered litter inputs affected the turnover and accumulation of soil carbon and nutrients more substantially over a shorter time period. Our study demonstrates that although the magnitude of responses differed greatly among ecosystems, the direction of the responses was very similar across different ecosystems. Interactions between plant productivity and below-ground biogeochemical cycling need to be taken into account to predict ecosystem responses to environmental change.
DOI:10.3773/j.issn.1005-264x.2009.05.006URL [本文引用: 1]
以佛山的加勒比松林(Pinus caribaea)为对象,研究了其地表径流及N和P流失量特点。结果表明,有凋落物加勒比松林和无凋落物加勒比松林的年径流量分别为12.6和51.8mm,其中夏季占90%以上。2种林分的地表径流量和降雨量的关系遵循线性方程。除2007年7月和2008年2月外,各月地表径流N浓度为有凋落物加勒比松林大于无凋落物加勒比松林,浓度的差异主要是因为年径流量不同引起的。总体来看,有凋落物加勒比松林的地表径流P浓度高,无凋落物加勒比松林低。地表径流N浓度与径流量之间存在着负指数关系。有凋落物加勒比松林和无凋落物加勒比松林的地表径流总N输出量分别为341和983g·hm–2,夏季分别占其年N流失量的73%和64%。无凋落物加勒比松林的地表径流P流失量高达21.5g·hm–2,而有凋落物加勒比松林为6.5g·hm–2。2008年6月为有凋落物加勒比松林和无凋落物加勒比松林的地表径流P流失高峰,分别占其年流失量的46%和60%。因此,保留林下凋落物可以降低加勒比松林的N和P的流失。
DOI:10.3773/j.issn.1005-264x.2009.05.006URL [本文引用: 1]
以佛山的加勒比松林(Pinus caribaea)为对象,研究了其地表径流及N和P流失量特点。结果表明,有凋落物加勒比松林和无凋落物加勒比松林的年径流量分别为12.6和51.8mm,其中夏季占90%以上。2种林分的地表径流量和降雨量的关系遵循线性方程。除2007年7月和2008年2月外,各月地表径流N浓度为有凋落物加勒比松林大于无凋落物加勒比松林,浓度的差异主要是因为年径流量不同引起的。总体来看,有凋落物加勒比松林的地表径流P浓度高,无凋落物加勒比松林低。地表径流N浓度与径流量之间存在着负指数关系。有凋落物加勒比松林和无凋落物加勒比松林的地表径流总N输出量分别为341和983g·hm–2,夏季分别占其年N流失量的73%和64%。无凋落物加勒比松林的地表径流P流失量高达21.5g·hm–2,而有凋落物加勒比松林为6.5g·hm–2。2008年6月为有凋落物加勒比松林和无凋落物加勒比松林的地表径流P流失高峰,分别占其年流失量的46%和60%。因此,保留林下凋落物可以降低加勒比松林的N和P的流失。
[本文引用: 2]
[本文引用: 2]
DOI:10.3773/j.issn.1005-264x.2008.06.011URL [本文引用: 1]
植被–大气间CO2净交换及其对环境变化的响应是目前全球变化研究的热点问题。该研究通过同化箱式法,在内蒙古农牧交错带对比研究生长季草地生态系统和耕种多年的小麦田生态系统碳通量的变化,以探讨该地区碳通量的变化规律及影响碳通量主要因子,并揭示农田开垦对草原碳通量的影响。结果显示:两个生态系统的群落净气体交换(Net ecosystem exchange,NEE)有明显的季节变化。整个测定期间,草地生态系统的净气体交换NEE的最高值为–11.26μmolCO2·m–2·s–1,平均群落净气体交换为–5.33μmolCO2·m–2·s–1;小麦田群落NEE最大值为–12.29μmolCO2·m–2·s–1,平均群落净气体交换为–7.66μmolCO2·m–2·s–1。分析发现,叶面积指数LAI是影响该地区生态系统NEE的主要因子,相对贫瘠的土壤也是限制该地区生态系统碳固定的一个重要因子。因小麦的生长特性,在生长中后期,小麦田生态系统NEE随LAI的变化没有草地生态系统的敏感。此外,较低的土壤含水量限制了小麦田群落呼吸,使得小麦田群落呼吸对温度的敏感性降低。
DOI:10.3773/j.issn.1005-264x.2008.06.011URL [本文引用: 1]
植被–大气间CO2净交换及其对环境变化的响应是目前全球变化研究的热点问题。该研究通过同化箱式法,在内蒙古农牧交错带对比研究生长季草地生态系统和耕种多年的小麦田生态系统碳通量的变化,以探讨该地区碳通量的变化规律及影响碳通量主要因子,并揭示农田开垦对草原碳通量的影响。结果显示:两个生态系统的群落净气体交换(Net ecosystem exchange,NEE)有明显的季节变化。整个测定期间,草地生态系统的净气体交换NEE的最高值为–11.26μmolCO2·m–2·s–1,平均群落净气体交换为–5.33μmolCO2·m–2·s–1;小麦田群落NEE最大值为–12.29μmolCO2·m–2·s–1,平均群落净气体交换为–7.66μmolCO2·m–2·s–1。分析发现,叶面积指数LAI是影响该地区生态系统NEE的主要因子,相对贫瘠的土壤也是限制该地区生态系统碳固定的一个重要因子。因小麦的生长特性,在生长中后期,小麦田生态系统NEE随LAI的变化没有草地生态系统的敏感。此外,较低的土壤含水量限制了小麦田群落呼吸,使得小麦田群落呼吸对温度的敏感性降低。
DOI:10.1126/science.1192666URLPMID:20724633 [本文引用: 1]
Terrestrial net primary production (NPP) quantifies the amount of atmospheric carbon fixed by plants and accumulated as biomass. Previous studies have shown that climate constraints were relaxing with increasing temperature and solar radiation, allowing an upward trend in NPP from 1982 through 1999. The past decade (2000 to 2009) has been the warmest since instrumental measurements began, which could imply continued increases in NPP; however, our estimates suggest a reduction in the global NPP of 0.55 petagrams of carbon. Large-scale droughts have reduced regional NPP, and a drying trend in the Southern Hemisphere has decreased NPP in that area, counteracting the increased NPP over the Northern Hemisphere. A continued decline in NPP would not only weaken the terrestrial carbon sink, but it would also intensify future competition between food demand and proposed biofuel production.
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia Grasslands
1
2010
... id="C13">同时, 退化群落样地于1999年至2013年处于冬春放牧状态, 凋落物生产和积累较少.过少的凋落物导致地表裸露面积增大, 生态微环境改变、土壤理化性质恶化, 进而使物种组成改变、物种多样性和草地生产力下降(
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau
1
2008
... id="C40">从整个生长季来看, 凋落物添加的退化群落气体交换变化规律是: 对GEP而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物添加100%与对照有显著差异; 对NEE而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物去除100%、凋落物去除50%与对照有显著差异.添加凋落物显著降低了土壤温度.NEE在生长季中期与土壤温度呈线性负相关关系(
Breathing of the terrestrial biosphere: Lessons learned from a global network of carbon dioxide flux measurement systems
1
2008
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
气温和降雨量对中国湿地生态系统CO2交换的影响
1
2015
... id="C36">凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(
气温和降雨量对中国湿地生态系统CO2交换的影响
1
2015
... id="C36">凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(
Soil moisture and plant growth responses to litter and defoliation impacts in parkland grasslands
2
2010
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
Carbon dioxide fluxes over bermudagrass, native prairie, and sorghum
1
1999
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
Natural mulches or litter of grasslands―With kinds and amounts on a southern prairie
1
1947
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements
1
2002
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
Carbon dioxide fluxes over a northern, semiarid, mixed-grass prairie
1
2001
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
Aspen plant community response to organic matter removal and soil compaction
1
2005
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
Changes in a California annual plant community induced by manipulation of natural mulch
1
1956
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
Evaporation and carbon dioxide exchange between the atmosphere and a tussock grassland during a summer drought
1
2002
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
Detritus accumulation limits productivity of tallgrass prairie
1
1986
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
模拟氮沉降下去除凋落物对太岳山油松林土壤呼吸的影响
1
2014
... id="C37">本研究的成熟群落中, 凋落物去除100%、凋落物去除50%处理和对照相比, 土壤温度有显著差异.季节温度均值也能证实: 凋落物去除100% >凋落物去除50%>对照, 这与
模拟氮沉降下去除凋落物对太岳山油松林土壤呼吸的影响
1
2014
... id="C37">本研究的成熟群落中, 凋落物去除100%、凋落物去除50%处理和对照相比, 土壤温度有显著差异.季节温度均值也能证实: 凋落物去除100% >凋落物去除50%>对照, 这与
Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia
3
2005
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
... ;
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
Effects of elevated concentrations of atmospheric CO2 and tropospheric O3 on leaf litter production and chemistry in trembling aspen and paper birch communities
2
2005
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
Vector analysis of understory competition, N fertilization, and litter layer removal effects on white spruce growth and nutrition in a 13-year- old plantation
2
2006
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
... ); 而王静等(2013)研究认为凋落物去除对地上生物量无显著影响.因此, 在不同的草原生态系统中, 凋落物去除对草地生产力的影响不同; 凋落物添加加大了太阳辐射拦截和土壤表层热量阻隔作用(
Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe
2
2008
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
... id="C40">从整个生长季来看, 凋落物添加的退化群落气体交换变化规律是: 对GEP而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物添加100%与对照有显著差异; 对NEE而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物去除100%、凋落物去除50%与对照有显著差异.添加凋落物显著降低了土壤温度.NEE在生长季中期与土壤温度呈线性负相关关系(
Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe
1
2009
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
Elevated CO2, litter chemistry, and decomposition: A synthesis
1
2001
... id="C36">凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(
Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients
2
2007
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
Large-scale manipulation of plant litter and fertilizer in a managed successional temperate grassland
1
2008
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
中国草地植被生物量及其空间分布格局
1
2004
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
中国草地植被生物量及其空间分布格局
1
2004
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
Hydrologic response of diverse western rangelands
1
2002
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
Grass litter is a natural seed trap in long-term undisturbed grassland
1
2012
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
A new approach to trenching experiments for measuring root-rhizosphere respiration in a lowland tropical forest
1
2010
... id="C40">从整个生长季来看, 凋落物添加的退化群落气体交换变化规律是: 对GEP而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物添加100%与对照有显著差异; 对NEE而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物去除100%、凋落物去除50%与对照有显著差异.添加凋落物显著降低了土壤温度.NEE在生长季中期与土壤温度呈线性负相关关系(
Effects of litter manipulation on early-stage decomposition and meso-arthropod abundance in a tropical moist forest
2
2006
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
... ), 或者间接改变土壤温度(
改变凋落物输入对杉木人工林土壤呼吸的短期影响
1
2009
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
改变凋落物输入对杉木人工林土壤呼吸的短期影响
1
2009
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
内蒙古典型草原不同功能群生产力对凋落物去除的响应
2013
内蒙古典型草原不同功能群生产力对凋落物去除的响应
2013
内蒙古典型草原不同功能群生产力对凋落物添加的响应
1
2010
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
内蒙古典型草原不同功能群生产力对凋落物添加的响应
1
2010
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
1
1992
... id="C36">凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(
Effects of excessive natural mulch on development, yield, and structure of native grassland
2
1952
... id="C7">凋落物是草地生态系统的重要组成部分, 它通过形成缓冲界面来调节土壤表面和大气之间的土壤微环境(
... id="C37">本研究的成熟群落中, 凋落物去除100%、凋落物去除50%处理和对照相比, 土壤温度有显著差异.季节温度均值也能证实: 凋落物去除100% >凋落物去除50%>对照, 这与
疏勒河上游高寒草甸生态系统CO2通量观测研究
1
2013
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
疏勒河上游高寒草甸生态系统CO2通量观测研究
1
2013
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe
3
2009
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
... id="C40">从整个生长季来看, 凋落物添加的退化群落气体交换变化规律是: 对GEP而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物添加100%与对照有显著差异; 对NEE而言, 凋落物添加100% >凋落物添加50% >对照, 凋落物去除100%、凋落物去除50%与对照有显著差异.添加凋落物显著降低了土壤温度.NEE在生长季中期与土壤温度呈线性负相关关系(
... 与土壤湿度为线性正相关关系(
How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature
3
2004
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
... )、解除了光限制, 增加了光合作用(
... 明显降低(
Variability of above-ground litter inputs alters soil physicochemical and biological processes: A meta-analysis of litterfall-manipulation experiments
3
2013
... id="C37">本研究的成熟群落中, 凋落物去除100%、凋落物去除50%处理和对照相比, 土壤温度有显著差异.季节温度均值也能证实: 凋落物去除100% >凋落物去除50%>对照, 这与
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
加勒比松林凋落物对地表径流和氮、磷流失的影响
1
2009
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
加勒比松林凋落物对地表径流和氮、磷流失的影响
1
2009
... id="C38">凋落物通过影响渗透、蒸发和径流来改变土壤湿度.在本研究中, 凋落物去除的成熟群落土壤湿度降低, 这与
凋落物处理对森林地表CO2通量的影响及其调控机理
2
2014
... id="C36">凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(
... 这与
凋落物处理对森林地表CO2通量的影响及其调控机理
2
2014
... id="C36">凋落物作为草原生态系统的重要组成部分, 在维持土壤肥力、影响初级生产力、改善土壤理化性质、调节生态系统能量流动和养分循环过程中起着重要作用(
... 这与
开垦对克氏针茅草地生态系统碳通量的影响
1
2008
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
开垦对克氏针茅草地生态系统碳通量的影响
1
2008
... id="C39">大量研究表明, 温度和水分是控制生态系统碳通量的重要环境因子(
Drought-induced reduction in global terrestrial net primary production from 2000 through 2009
1
2010
... id="C6">陆地生态系统与大气之间的CO2净交换(NEE)也称为生态系统碳收支(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}