 ,*河北大学生命科学学院, 河北保定 071002
,*河北大学生命科学学院, 河北保定 071002Eco-geographical distribution of arbuscular mycorrhizal fungi associated with Hedysarum scoparium in the desert zone of northwestern China
LIU Hai-Yue, LI Xin-Mei, ZHANG Lin-Lin, WANG Jiao-Jiao, HE Xue-Li,*College of Life Sciences, Hebei University, Baoding, Hebei 071002, China通讯作者:
| 基金资助: |
Online:2018-04-19
| Supported by: | SupportedbytheNationalNaturalScienceFoundationofChina( |

摘要
关键词:
Abstract
Keywords:
PDF (1126KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
刘海跃, 李欣玫, 张琳琳, 王姣姣, 贺学礼. 西北荒漠带花棒根际丛枝菌根真菌生态地理分布. 植物生态学报, 2018, 42(2): 252-260 doi:10.17521/cjpe.2017.0138
LIU Hai-Yue, LI Xin-Mei, ZHANG Lin-Lin, WANG Jiao-Jiao, HE Xue-Li.
我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢。花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(徐朗然, 1985; 贺学礼等, 2006a)。花棒根系发达, 耐贫瘠, 具有抗热、抗旱、抗寒、耐盐碱、耐沙埋、抗风蚀等特性, 是荒漠地区防风固沙的优良灌木, 对荒漠生态系统维持和沙漠绿化有重要作用(段小圆和贺学礼, 2008)。
丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(Chen et al., 2012), 具有促进植物生长和植被恢复的功能(Feddermann et al., 2010; Gianinazzi et al., 2010)。AM真菌能在植物根细胞内形成泡囊、丛枝等结构(贺学礼等, 2010a), 可以增强宿主植物对水分和矿质营养的吸收(Li et al., 2010; 杨静等, 2011)。并且AM真菌能通过菌丝桥、菌根依赖性差异等机制, 对植物种群动态调控等生态学过程产生深远影响(Vogelsang et al., 2006; Wagg et al., 2011)。此外, AM真菌产生的球囊霉素是荒漠植被根际土壤碳、氮的重要来源(贺学礼等, 2011), 有利于改善土壤理化性质, 改良和稳定土壤结构。同时, 土壤因子也会影响AM真菌生态分布、侵染能力、孢子密度等(He et al., 2010)。郭清华等(2016)的研究表明, AM真菌孢子密度与土壤有机碳极显著负相关。目前, 在荒漠生境已发现AM真菌寄主植物69科389种, AM真菌7属89种, 且不断有新种发现(贺学礼等, 2012)。已有研究表明, AM真菌可与花棒形成良好的互惠共生体(段小圆和贺学礼, 2008), 由于花棒形态结构和生态适应性的大幅变化(徐朗然, 1985), 与之生长发育密切相关的AM真菌的群落组成、生物多样性及其地理分布格局也必然随之变化。
本试验沿西北荒漠带花棒天然分布区, 采集花棒根际土壤样品, 分析AM真菌群落结构、物种多样性及其与土壤因子的相关性, 阐明AM真菌生态地理分布规律, 为充分利用AM真菌资源促进荒漠植物生长和植被恢复提供材料和依据。
1 材料和方法
1.1 研究区域概况
采样区域位于我国西北荒漠带(73.15°- 111.17° E、31.32°-49.10° N), 属温带大陆性气候, 年降水量在东部为400 mm左右, 西部减少到200 mm甚至50 mm以下, 是一个从半干旱逐步过渡到极度干旱的完整荒漠系列, 自然景观由荒漠草原逐步演变为极旱荒漠(李旭谱等, 2012)。该区域受干旱影响, 生物多样性低, 群落结构简单, 主要以超旱生小乔木、灌木和半灌木为主, 生态系统极其脆弱(朱宗元等, 1999)。1.2 样品采集
2015年7月在内蒙古、宁夏、甘肃选取7个典型荒漠样地, 每个样地选取3个小样地, 在每个小样地选取5株长势良好的花棒植株, 去除表面枯枝落叶层, 采集0-30 cm土层根样, 将根样上的土轻轻抖落并进行收集, 放入已编号的自封袋中, 用温湿度计测量土壤温湿度并记录采样地基本信息(表1)。样品带回实验室, 土样过2 mm筛, 阴干后4 ℃冷藏, 用于AM真菌分离鉴定和土壤因子测定。Table 1
表1
表1采样地概况(平均值±标准误差)
Table 1
| 样地 Site | 海拔 Altitude (m) | 经纬度 Latitude and longitude | 土壤温度 Soil temperature (°C) | 土壤湿度 Soil moisture (%) |
|---|---|---|---|---|
| 鄂尔多斯沙地草地生态研究观测站 Ordos Sandland Ecological Research Station | 1 269.0 | 39.19° N, 110.11° E | 23.40 ± 0.21 | 9.81 ± 0.05 |
| 乌海海勃湾区 Wuhai Haibowan District | 1 150.0 | 39.49° N, 106.49° E | 30.54 ± 0.32 | 5.71 ± 0.07 |
| 磴口阿敦乌苏 Dengkou Aton Usu | 1 030.0 | 40.39° N, 106.74° E | 22.14 ± 0.23 | 4.67 ± 0.06 |
| 阿拉善木仁高勒苏木 Mu Ren Gao Le Su Mu, Alxa League | 1 295.0 | 39.10° N, 105.52° E | 28.06 ± 0.45 | 7.29 ± 0.04 |
| 沙坡头沙漠试验研究站 Shapotou Desert Experimental Research Station | 2 027.5 | 37.27° N, 104.59° E | 40.05 ± 0.51 | 6.34 ± 0.09 |
| 民勤连古城国家级自然保护区 Minqin Liangucheng National Nature Reserve | 1 400.0 | 39.00° N, 102.37° E | 29.35 ± 0.22 | 4.48 ± 0.05 |
| 安西极旱荒漠自然保护区 Anxi Extreme-Arid Desert National Nature | 1 514.0 | 40.20° N, 096.50° E | 27.66 ± 0.37 | 1.37 ± 0.02 |
新窗口打开|下载CSV
1.3 研究方法
1.3.1 AM真菌分离鉴定准确称取20 g风干土, 用湿筛倾析-蔗糖离心法(Ianson & Allen, 1986)分离AM真菌孢子, 在体视显微镜下观察记录孢子数量。将孢子置于浮载剂(水、乳酸等)中压片处理, 在光学显微镜下观察孢子大小、形态、颜色、表面纹饰、孢壁结构、连孢菌丝、内含物等特征并拍照记录。根据Schenck和Perez (1990) 《VA菌根鉴定手册》和国际AM真菌保藏中心(INVAM)在http://invam.caf.wvu.edu上提供的最新分类描述及图片进行种属鉴定。
1.3.2 AM真菌物种多样性测定
参照赵金莉和贺学礼(2013)的方法计算AM真菌种丰度、孢子密度、相对多度及物种多样性指数等。
种丰度(SR) = 20 g风干土含有的AM真菌种数
孢子密度(SD) = 20 g风干土中的孢子数量
相对多度(RA) = (该采样点AM真菌某属或种的孢子数/该采样点AM真菌总孢子数) × 100%
Shannon-Wiener多样性指数: \[H=-\sum\limits_{i=1}^{S}{{{P}_{i}}\ln {{P}_{i}}}\]
Simpson多样性指数: \[{{D}_{S}}=1-\sum\limits_{i=1}^{S}{{{P}_{i}}^{2}}\]
式中, S为某采样点AM真菌种类数, Pi为种i所占比例。
1.3.3 土壤理化性质测定
土壤有机碳采用马弗炉烧失法(Bai et al., 2009)测定; pH值用雷磁PHS-3C pH计测定; 氨态氮使用SMARTCHEM 200全自动连续分析仪(Alliance, Frépillon, France)测定; 有效磷采用碳酸氢钠-钼锑抗比色法(Bai et al., 2009)测定; 根据Hoffmann和Teicher (1961)的方法测定土壤脲酶, 活性以每克土样培养1 h催化尿素分解产生NH4+-N的量(μg·g-1)表示。根据Tarafdar和Marschner (1994)的方法测定土壤酸性磷酸酶和碱性磷酸酶, 活性分别以每克土样培养1 h酸性磷酸酶和碱性磷酸酶转化对硝基苯磷酸二钠(PNPP)的量(μg·g-1)表示。球囊霉素通过Wright和Upadhyaya (1998)的方法测定。易提取球囊霉素(EEG)上清液的制备方法: 取1 g风干土于试管中, 加入0.02 mol·L-1 (pH 7.0)柠檬酸钠浸提剂8 mL, 在103 kPa、121 ℃下连续提取30 min后, 在4 000 r·min-1转速下离心15 min, 收集上清液。总球囊霉素(TEG)上清液的制备方法: 取1 g风干土于试管中, 加入0.05 mol·L-1 (pH 8.0)柠檬酸钠浸提剂8 mL, 在103 kPa、121 ℃条件下连续提取60 min, 再重复提取2次, 在4 000 r·min-1转速下离心15 min, 收集上清液; 分别吸取上清液0.5 mL, 加入5 mL考马斯亮蓝G-250染色剂, 盖塞, 倒转混匀, 2 min后用1 cm光径的比色杯在595 nm波长下比色。用牛血清蛋白配制标准溶液, 考马斯亮蓝法显色, 绘制标准曲线, 求出球囊霉素含量。
1.3.4 数据统计
试验结果经Excel 2013处理后, 用SPSS 19.0软件和Canoco 4.5软件对数据进行统计分析。
2 结果和分析
2.1 AM真菌群落组成
在7个样地共分离鉴定了6属42种AM真菌, 包括无梗囊霉属(Acaulospora) 18种, 其中未定种3种; 球囊霉属(Glomus) 15种, 其中未定种1种; 管柄囊霉属(Funneliformis) 2种; 缩管柄囊霉属(Septoglomus) 2种; 近明囊霉属(Claroideoglomus) 4种; 盾巨孢囊霉属(Scutellospara) 1种(表2)。由图1A可见, 在西北荒漠带内由东到西AM真菌种丰度表现出下降趋势。在图1B所示的共有属中, 球囊霉属和无梗囊霉属对AM真菌种丰度的贡献高于其他属, 所占比重超过80%。各样地种类构成中, 球囊霉属和近明囊霉属丰度所占比重稳定, 但球囊霉属比例远高于近明囊霉属; 无梗囊霉属所占比重在西北荒漠带由东到西逐渐减少, 与缩管柄囊霉属相反, 其他各属丰度对群落种类构成的贡献较小。
图1
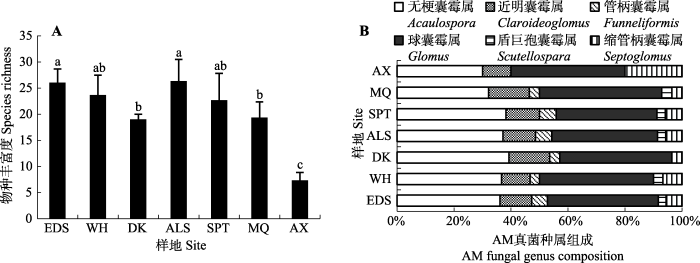 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同样地花棒根际AM真菌种丰富度(A)和组成(B)。不同小写字母表示不同样地间差异显著(p < 0.05)。ALS, 阿拉善; AX, 安西; DK, 磴口; EDS, 鄂尔多斯; MQ, 民勤; SPT, 沙坡头; WH, 乌海。
Fig. 1Species richness (A) and composition (B) of arbuscular mycorrhizae (AM) fungi associated with Hedysarum scoparium in different sites. Different lowercase letters indicate the significant difference among sites (p < 0.05). ALS, Alxa; AX, Anxi; DK, Dengkou; EDS, Ordos; MQ, Minqin; SPT, Shapotou; WH, Wuhai.
不同样地间土壤理化性质存在差异(附件I), 导致AM真菌种属具有地域性分布。分析结果中两轴(RDA1和RDA2)能够解释变量方差的51.2%和20.2%, 累积方差贡献率达71.4%。图2中样地间得分距离越小, 表示样地间AM真菌群落构成相似程度越高。7个样地位于不同象限内, 明显分为3个类群, 阿拉善和鄂尔多斯各成一个类群, 其余5个样地组成一个类群, 说明3个类群间AM真菌群落构成差异明显。同时, 由图2可以看出, 造成样地间AM真菌群落构成差异的土壤因子各不相同。球囊霉素对鄂尔多斯样地的影响明显, 酸性磷酸酶对阿拉善样地影响显著, 而其余样地受土壤pH值以及有机碳的影响明显。
图2
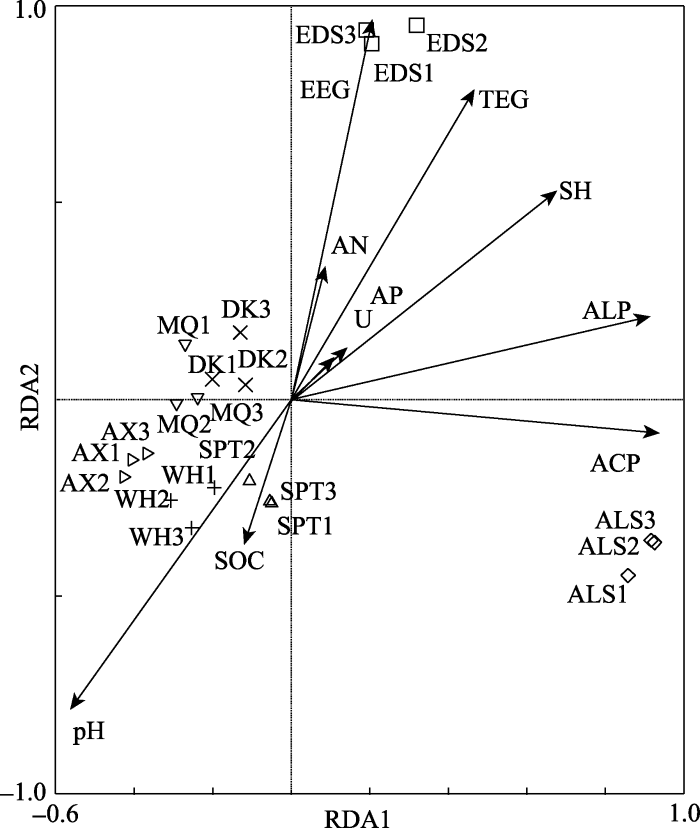 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同样地花棒根际AM真菌物种组成的RDA分析。ALS, 阿拉善; AX, 安西; DK, 磴口; EDS, 鄂尔多斯; MQ, 民勤; SPT, 沙坡头; WH, 乌海。ACP, 酸性磷酸酶; ALP, 碱性磷酸酶; AN, 氨态氮; AP, 有效磷; EEG, 易提取球囊霉素; pH, pH值; SH, 土壤湿度; SOC, 有机碳; ST, 土壤温度; TEG, 总体提取球囊霉素; UA, 脲酶。
Fig. 2RDA analysis of species composition of arbuscular mycorrhizae (AM) fungi associated with Hedysarum scoparium in different sites. ALS, Alxa; AX, Anxi; DK, Dengkou; EDS, Ordos; MQ, Minqin; SPT, Shapotou; WH, Wuhai. ACP, acid phosphatase; ALP, alkaline phosphatase; AN, ammonia nitrogen; AP, available phosphorus; EEG, easily extractable glomalin; pH, pH value; SH, soil moisture; SOC, organic carbon; ST, soil temperature; TEG, total extractable glomalin; UA, urease.
2.2 AM真菌孢子密度与球囊霉素
孢子密度作为表征AM真菌生存状态的重要指标, 其大小代表了AM真菌生存与繁殖能力的强弱。由图3A可以看出, 荒漠条件下各样地孢子密度显著不同, 除阿拉善外, 孢子密度在西北荒漠带内由东到西逐渐降低。球囊霉素作为AM真菌的代谢产物, 在一定程度上展现了AM真菌的生存状态。在图3B中可以看出, 西北荒漠带内由东到西球囊霉素含量逐渐降低。图3
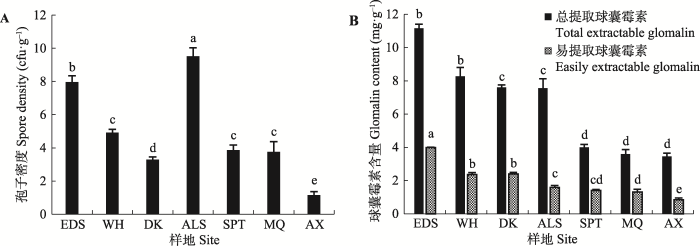 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同样地花棒根际AM真菌孢子密度(A)和球囊霉素含量(B) (平均值±标准误差)。不同小写字母表示不同样地间差异显著(p < 0.05)。ALS, 阿拉善; AX, 安西; DK, 磴口; EDS, 鄂尔多斯; MQ, 民勤; SPT, 沙坡头; WH, 乌海。
Fig. 3Spore density (A) and glomalin content (B) of arbuscular mycorrhizae (AM) fungi associated with Hedysarum scoparium in different sites (mean ± SE). Different lowercase letters indicate the significant difference among sites (p < 0.05). ALS, Alxa; AX, Anxi; DK, Dengkou; EDS, Ordos; MQ, Minqin; SPT, Shapotou; WH, Wuhai.
2.3 AM真菌共有种
由表2可知, 双网无梗囊霉、卷曲球囊霉、网状球囊霉和沙荒缩管柄囊霉是7个样地的共有种。图4A中共有种的孢子密度由东到西呈波动式下降, 与土壤湿度变化趋势相同, 表明干旱程度对AM真菌有明显影响。但图4B所示的共有种孢子密度所占比例中, 在研究区域内由东到西表现出上升趋势。Table 2
表2
表2丛枝菌根真菌空间分布
Table 2
| AM真菌 AM fungi | 鄂尔多斯 Ordos | 乌海 Wuhai | 磴口 Dengkou | 阿拉善 Alxa | 沙坡头 Shapotou | 民勤 Minqin | 安西 Anxi |
|---|---|---|---|---|---|---|---|
| 凹坑无梗囊霉 Acaulospora excavata | 1.00 | 5.33 | 1.67 | 2.33 | 0.33 | 0.33 | - |
| 刺无梗囊霉 Acaulospora spinosa | - | - | 0.33 | 0.33 | - | - | - |
| 附柄无梗囊霉 Acaulospora appendicola | - | 1.33 | - | - | 0.33 | - | - |
| 光壁无梗囊霉 Acaulospora laevis | 2.33 | 0.33 | 3.67 | 1.33 | 0.67 | 2.33 | - |
| 孔窝无梗囊霉 Acaulospora foveata | 2.33 | 3.00 | 0.67 | 6.33 | 4.33 | 2.00 | - |
| 毛氏无梗囊霉 Acaulospora morrowae | 7.33 | - | 3.67 | 1.67 | 3.67 | 5.00 | 1.33 |
| 蜜色无梗囊霉 Acaulospora mellea | 5.67 | 9.00 | 1.67 | 2.00 | 3.00 | 4.33 | - |
| 膨胀无梗囊霉 Acaulospora dilatata | 0.67 | 4.00 | - | 0.33 | 0.67 | - | - |
| 浅窝无梗囊霉 Acaulospora lacunosa | 14.33 | 4.67 | 1.33 | 1.00 | 0.33 | - | - |
| 疣状无梗囊霉 Acaulospora tuberculata | 0.67 | - | - | 0.33 | - | - | - |
| 瑞氏无梗囊霉 Acaulospora rehmii | 17.00 | - | 2.33 | 8.33 | 3.00 | 4.33 | 7.67 |
| 双网无梗囊霉 Acaulospora bireticulata | 3.33 | 9.67 | 2.67 | 3.33 | 0.67 | 7.67 | 0.33 |
| 细凹无梗囊霉 Acaulospora scrobiculata | 0.67 | 13.67 | - | 0.33 | 0.67 | 0.67 | - |
| 细齿无梗囊霉 Acaulospora denticulate | 1.00 | 0.33 | - | 2.00 | 0.33 | - | - |
| 皱壁无梗囊霉 Acaulospora rugosa | - | - | 0.33 | - | 0.33 | 0.33 | - |
| Acaulospora sp. 1 | 1.00 | - | 0.67 | - | - | - | - |
| Acaulospora sp. 2 | - | 0.33 | - | - | - | - | - |
| Acaulospora sp. 3 | 3.00 | - | 0.67 | 15.33 | 4.33 | 0.33 | 0.67 |
| 层状近明囊霉 Claroideoglomus lamellosum | 17.00 | 7.33 | 6.67 | 28.33 | 8.00 | 2.67 | - |
| 近明囊霉 Claroideoglomus claroideum | 7.00 | 3.33 | 1.33 | 0.67 | 3.67 | 0.67 | - |
| 黄近明囊霉 Claroideoglomus luteum | 2.00 | 0.33 | 0.33 | 1.00 | 0.67 | 1.33 | - |
| 幼套近明囊霉 Claroideoglomus etunicatum | 0.33 | 1.00 | - | 8.33 | 1.33 | 0.67 | - |
| 地管柄囊霉 Funneliformis geosporum | 0.67 | - | 0.67 | 1.33 | 0.33 | - | - |
| 摩西管柄囊霉 Funneliformis mosseae | 6.33 | 0.67 | - | 0.67 | 0.33 | 1.00 | - |
| 道氏球囊霉 Glomus dominikii | 2.33 | - | 0.33 | 8.00 | 4.33 | 7.67 | _ |
| 地表球囊霉 Glomus versiforme | 1.67 | - | - | 0.33 | 1.33 | 1.00 | - |
| 多梗球囊霉 Glomus multicaule | 5.33 | 6.00 | 2.67 | 29.67 | 7.00 | 2.00 | - |
| 黑球囊霉 Glomus melanosporum | 1.00 | 3.33 | - | - | 3.00 | 0.67 | - |
| 聚丛球囊霉 Glomus aggregatum | 0.33 | - | 0.33 | - | - | - | - |
| 聚集球囊霉 Glomus glomorulatum | 1.67 | 4.67 | - | 0.67 | 1.33 | 0.67 | - |
| 卷曲球囊霉 Glomus convolutum | 3.67 | 1.33 | 0.33 | 1.67 | 1.00 | 0.33 | 0.67 |
| 宽柄球囊霉 Glomus magnicaule | - | 1.00 | - | - | 0.33 | 6.67 | 1.00 |
| 膨果球囊霉 Glomus pansihalos | 3.00 | 0.33 | 5.33 | 21.67 | 6.00 | 0.33 | - |
| 透光球囊霉 Glomus diaphanum | 0.33 | 0.67 | 4.67 | 0.67 | 0.33 | - | - |
| 网状球囊霉 Glomus reticulatum | 31.33 | 6.00 | 15.67 | 19.00 | 9.00 | 13.00 | 4.00 |
| 微丛球囊霉 Glomus microaggregatum | 2.00 | 1.67 | 1.67 | 0.67 | - | 0.33 | - |
| 隐球囊霉 Glomus occultum | 2.33 | - | 3.00 | 1.33 | - | 1.33 | 0.33 |
| 粘质球囊霉 Glomus viscosum | 3.33 | 1.00 | 1.00 | 8.33 | 1.67 | - | - |
| Glomus sp. 1 | - | 1.00 | 0.33 | 1.00 | - | - | - |
| 美丽盾巨孢囊霉 Scutellospara calospora | 0.33 | 0.33 | - | 0.33 | 0.33 | 0.33 | - |
| 沙荒缩管柄囊霉 Septoglomus deserticola | 6.67 | 3.33 | 2.00 | 8.00 | 4.00 | 7.00 | 5.67 |
| 缩管柄囊霉 Septoglomus constrictum | 0.33 | 3.33 | - | 3.67 | 0.67 | - | 1.33 |
| 种数 Number of species | 36 | 30 | 28 | 35 | 34 | 28 | 10 |
新窗口打开|下载CSV
图4
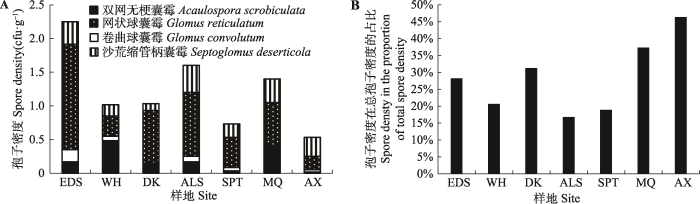 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4花棒根际AM真菌共有种的孢子密度(A)及其占总孢子密度的比例(B)。ALS, 阿拉善; AX, 安西; DK, 磴口; EDS, 鄂尔多斯; MQ, 民勤; SPT, 沙坡头; WH, 乌海。
Fig. 4The spore density of common species associated with Hedysarum scoparium (A) and its proportion among all arbuscular mycorrhizae (AM) fungal species (B) in different sites. ALS, Alxa; AX, Anxi; DK, Dengkou; EDS, Ordos; MQ, Minqin; SPT, Shapotou; WH, Wuhai.
Table 3
表3
表3AM真菌物种多样性和环境因子相关性分析
Table 3
| 环境因子Environmental factor | 种丰度 Species richness | 孢子密度 Spore density | Shannon-Wiener指数 Shannon-Wiener index | 辛普森指数 Simpson index | 易提取球囊霉素 Easily extractable glomalin | 总提取球囊霉素 Total extractable glomalin |
|---|---|---|---|---|---|---|
| SOC | 0.271 | -0.511* | 0.347 | 0.338. | -0.512* | -0.435* |
| AP | 0.386 | 0.051 | 0.537* | 0.532* | 0.283 | 0.302 |
| AN | 0.275 | 0.637** | 0.215 | 0.170 | 0.309 | 0.256 |
| pH | 0.411 | -0.725** | 0.303 | 0.275 | -0.764** | -0.751** |
| ST | 0.030 | -0.305 | 0.158 | 0.188 | -0.090 | 0.055 |
| SH | 0.787** | 0.581** | 0.688** | 0.608** | 0.563** | 0.664** |
| LA | 0.762** | -0.381 | 0.772** | 0.718** | 0.504* | 0.518* |
新窗口打开|下载CSV
2.4 AM真菌多样性与土壤因子的相关性
相关性分析表明, AM真菌种丰度与土壤湿度极显著正相关; 孢子密度与氨态氮、pH值和湿度极显著正相关, 与有机碳显著负相关; Shannon-Wiener指数和Simpson指数与土壤湿度极显著正相关, 与有效磷显著正相关; 球囊霉素与土壤湿度极显著正相关, 与pH值极显著负相关, 与有机碳显著负相关。同时, 种丰度、多样性指数以及球囊霉素含量与经度梯度表现出显著正相关。3 讨论
研究发现, 花棒根际AM真菌各属呈现球囊霉属>无梗囊霉属>近明囊霉属>缩管柄囊霉属>管柄囊霉属>盾巨孢囊霉属, 球囊霉属和无梗囊霉属是各样地的优势属, 这与张美庆等(1994)认为球囊霉属和无梗囊霉属是广谱生态型的结论一致。本试验共分离到无梗囊霉属18种, 多于球囊霉属, 与以往报道(贺学礼等, 2006b; 冀春花等, 2007)不同。Pringle和Bever (2002)发现, 不同AM真菌具有不同的物候特性, 无梗囊霉属主要在暖季产孢。本试验采样时间在7月, 西北地区正处于暖季, 是无梗囊霉属的产孢旺季, 这可能是无梗囊霉属种类高于球囊霉属的原因。本试验中, 双网无梗囊霉、卷曲球囊霉、网状球囊霉和沙荒缩管柄囊霉是7个样地的共有种, 说明这些AM真菌具有极强的适应性, 这为筛选花棒高效AM菌种提供了依据。生物物种组成、群落结构及多样性是反映生物地理分布的重要指标(Odland & Moral, 2002)。本试验中, 7个样地的AM真菌群落组成表现出明显的差异。Davison等(2015)研究表明, 在全球尺度下, AM真菌未形成明显的空间格局, 但空间距离对AM真菌群落组成有重要影响, 即扩散限制是影响AM真菌地理分布的一个原因。AM真菌群落形成受菌丝扩散和自然扩散的严重限制, 人为因素是推动AM真菌孢子扩散的主要原因(Rosendahl, 2008)。本次试验样品采自保护区及野外观测站周围, 降低了人为干扰。Hazard等(2013)发现在景观尺度上, AM真菌群落组成由局部非生物环境决定, 如土壤pH值、降雨、土壤类型等。
西北荒漠带东西跨度极大, 受海陆位置影响, 由东到西干旱程度逐渐增加, 表现出荒漠草原、荒漠和极旱荒漠的景观变化。本试验中, 由东到西沿经度变化降雨量和土壤湿度逐渐减少, 受降雨影响, 花棒植株表现出对环境的强烈适应性, 这为AM真菌侵染和共生提供了适宜的生存环境。Bever等(2009)研究证实, 宿主植物种类和根系特性是AM真菌发生和分布的重要因子。豆科植物不仅能与AM真菌形成良好的共生联合体, 而且能够通过固氮、调节根系碳水化合物分配、产生次生代谢物质、改变根际土壤微环境等途径影响AM真菌的活动和分布(白春明等, 2009), 这也可能是造成西北地区AM真菌分布差异的原因。
AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响。土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(Aliasgharzadeh et al., 2001)。在影响AM真菌群落结构的土壤因子中, 土壤pH值、有机碳、有效磷的影响最为显著(Rillig, 2004)。土壤中性至微酸性时有利于AM真菌孢子形成和发育(van Aarle et al., 2002)。Fitzsimons等(2008)发现, 土壤pH值能够很好地预测AM真菌群落变化。西北荒漠土壤为弱碱性, 由东向西沿经度梯度降雨量逐渐减少, 土壤pH值随之升高(丁小慧等, 2012), 不利于AM真菌孢子的形成和发育, 孢子密度和球囊霉素含量与土壤pH值表现出极显著负相关关系。有机碳含量变化无明显规律, 但有机碳含量与孢子密度和球囊霉素含量显著负相关, 这与之前的研究结果(贺学礼等, 2010b)一致。同时, AM真菌分泌的球囊霉素又是土壤有机碳库的重要来源和组成部分, 有利于改善土壤肥力, 促进植物生长(郭清华等, 2016)。土壤pH值升高, 使土壤中金属离子对磷的吸附作用上升, 使土壤中有效磷含量降低(丁小慧等, 2012)。本次试验中, 有效磷含量与AM真菌多样性显著正相关, 与Alguacil等(2010)的磷含量增加导致AM真菌多样性降低的结论不一致, 这可能是因为研究区域为荒漠, 磷含量较低的缘故。Landis等(2004)发现, AM真菌多样性与土壤养分含量显著相关, 氮含量高的土壤, AM真菌物种丰富度较高。AM真菌侵染宿主植物后, 能够吸收大量的无机氮, 这些无机氮可参与孢子形成过程。由此可见, 养分对AM真菌发育的促进作用是有一定范围的, 养分过高或过低都不利于AM真菌生长发育(Santos et al., 2006)。同时, 各个土壤因子对AM真菌群落组成的影响并不是孤立的, 而是相互关联的整体。当研究某一生态因子对AM真菌的影响时, 不能忽略其他因子的作用。
综上所述, 由于经度梯度变化引起的水热条件变化和土壤因子的作用, AM真菌群落组成和分布具有明显的空间异质性。
致谢 本研究受到中国科学院鄂尔多斯沙地草地生态研究观测站、中国科学院沙坡头沙漠试验研究站、民勤连古城国家级自然保护区、安西极旱荒漠国家自然保护区等单位及河北省研究生创新项目(CXZZBS2017019)支持,特此致谢!
Appendix I
附件I
附件I不同样地土壤因子(平均值±标准误差, n = 3)
Appendix I
| 样地 Site | SOC (mg·g-1) | AP (µg·g -1) | AN (μg·g-1) | pH | ACP (µg·g-1·h-1) | ALP (µg·g-1·h-1) | UA (μg·g-1·h-1) | EEG (mg·g-1) | TEG (mg·g-1) |
|---|---|---|---|---|---|---|---|---|---|
| 鄂尔多斯 Ordos | 10.00 ± 1.00c | 6.14 ± 0.50b | 64.24 ± 2.13b | 7.62 ± 0.27c | 51.97 ± 1.62c | 59.82 ± 0.92b | 7.33 ± 0.23e | 3.98 ± 0.03a | 11.17 ± 0.24a |
| 乌海 Wuhai | 36.78 ± 2.01a | 5.16 ± 0.46c | 53.22 ± 2.70c | 8.57 ± 0.04a | 32.01 ± 1.41f | 17.33 ± 2.00e | 1.58 ± 0.05g | 2.36 ± 0.12b | 8.28 ± 0.53b |
| 磴口 Dengkou | 9.92 ± 1.61c | 8.57 ± 1.19a | 93.34 ± 3.92a | 8.51 ± 0.06a | 55.70 ± 0.93b | 56.50 ± 0.94c | 10.64 ± 0.38b | 2.39 ± 0.10b | 7.61 ± 0.15c |
| 阿拉善 Alxa | 14.42 ± 1.63b | 5.97 ± 0.16bc | 50.20 ± 0.52cd | 8.25 ± 0.17b | 85.19 ± 1.19a | 82.72 ± 2.40a | 7.94 ± 0.35d | 1.59 ± 0.12c | 7.56 ± 0.56c |
| 沙坡头 Shapotou | 10.92 ± 0.52c | 7.83 ± 0.07a | 40.27 ± 1.02e | 8.54 ± 0.08a | 44.46 ± 2.96d | 30.92 ± 0.83d | 11.41 ± 0.23a | 1.40 ± 0.06cd | 4.01 ± 0.17d |
| 民勤 Minqin | 11.67 ± 0.58c | 6.40 ± 0.22b | 26.29 ± 0.82f | 8.56 ± 0.13a | 29.04 ± 1.45f | 17.62 ± 0.80e | 10.08 ± 0.39c | 1.31 ± 0.16d | 3.60 ± 0.26d |
| 安西 Anxi | 9.58 ± 1.18c | 3.81 ± 0.10d | 46.58 ± 1.90d | 8.68 ± 0.12a | 37.58 ± 2.07e | 17.15 ± 1.67e | 6.35 ± 0.30f | 0.84 ± 0.11e | 3.46 ± 0.19d |
新窗口打开|下载CSV

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
.
DOI:10.1016/j.soilbio.2010.03.012URL

In the present study we investigated how the community of arbuscular mycorrhizal fungi (AMF) in roots of Centrosema macrocarpum responded to different doses and sources of phosphorus (40kgha 611 of P as rock phosphate, 150kgha 611 of P as rock phosphate and 75kgha 611 of P as diammonium phosphate together with 75kgha 611 of P as rock phosphate) in a Venezuelan savanna ecosystem. We also related AMF diversity to soil parameters (total N, total P, available P, extractable K, total Ca, total Mg, total Fe, total Cu, total Zn, total Mn, glomalin-related soil protein, microbial biomass C, dehydrogenase, urease and acid phosphatase activities, water-soluble carbon and carbohydrates and aggregate stability) at different doses of P. The AM fungal small-subunit (SSU) rRNA genes were subjected to PCR, cloning, SSCP, sequencing and phylogenetic analyses. Nine fungal types were identified: six belonged to the genus Glomus and three to Acaulospora . The majority of fungal types showed high similarity to sequences of known glomalean isolates: Aca 1 to Acaulospora mellea , Aca 2 to Acaulospora rugosa , Aca 3 to Acaulospora spinosa , Glo 1 to Glomus intraradices and Glo 3 to Glomus fasciculatum . The control treatment was dominated by species belonging to the genus Acaulospora . However, when the soil was fertilised with low doses of P, the colonisation of roots increased and there was a change in the AMF diversity, the genus Glomus dominating. The AM development and the abundance of AM fungal types in roots were decreased dramatically by the fertilisation with high doses of P, without differences between the sources of P used. The available P in soil was negatively correlated with the AMF diversity. In conclusion, the application of low doses of P as rock phosphate stimulated mycorrhization and enhanced the soil quality parameters except water-soluble carbohydrates, helping to offset a loss of fertility in P-poor tropical savanna soils.
.
DOI:10.1007/s005720100113URLPMID:28401349 [本文引用: 1]
The abundance and distribution of arbuscular mycorrhizal fungi (AMF) were evaluated in the Tabriz Plain, where soil salinity levels range from 7.3 to 92.0聽dS/m. Soil and root samples were collected from the rhizosphere of several glycophytes ( Allium cepa L., Medicago sativa L., Triticum aestivum L. and Hordeum vulgare L.) and halophytes ( Salicornia sp. and Salsola sp.) and were analysed for spore number in soil, root colonization with AMF and some physical and chemical soil properties. The number of AMF spores was not correlated significantly with soil salinity but suffered adverse effects from the accumulation of some anions and cations. Cluster analysis of correlation matrices showed that root colonization, soil pH, sand and clay percent, and soil-available P, rather than soil salinity and ion concentrations, were closely related with spore number. The percentage of root length colonized in glycophytes significantly decreased with increasing soil salinity. Barley roots showed 5% mycorrhizal colonization in high soil salinity (~20聽dS/m). Halophyte roots were not mycorrhizal but more spores were found in rhizosphere than in non-rhizosphere soil.
.
URL [本文引用: 3]
【目的】研究漠境AM真菌对宿主沙打旺(Astragalus adsurgens)的作用机理及对根部土壤酶活性的影响。【方法】分别采集陕西靖边、宁夏盐池、内蒙古包头和集宁等4个样地沙打旺根围0~10,10~20,20~30,30~40和40~50 cm等5个土层的土壤样品,研究AM真菌在土壤中的空间分布及其与土壤脲酶、蛋白酶、酸性磷酸酶活性和碱解N、速效P含量的关系。【结果】AM真菌定殖率及孢子密度与样地生态条件密切相关,其中集宁样地孢子密度最高,靖边样地泡囊定殖率最高,包头样地丛枝、菌丝和总定殖率均高于其他样地;孢子密度与AM真菌不同结构定殖率间无显著相关性。土壤酸性磷酸酶活性对孢子密度有极显著正的效应;脲酶活性对孢子密度和丛枝定殖率有极显著的正效应;蛋白酶活性对丛枝定殖率、菌丝定殖率、总定殖率均有极显著正效应。土壤蛋白酶与泡囊定殖率呈显著负相关。碱解氮、速效磷对与以上3种土壤酶活性均呈极显著正相关。【结论】沙漠生境下AM真菌与土壤酶活性显著相关。AM真菌孢子密度和菌根不同结构的定殖程度可作为荒漠土壤生态系统环境状况检测的有效指标。
URL [本文引用: 3]
【目的】研究漠境AM真菌对宿主沙打旺(Astragalus adsurgens)的作用机理及对根部土壤酶活性的影响。【方法】分别采集陕西靖边、宁夏盐池、内蒙古包头和集宁等4个样地沙打旺根围0~10,10~20,20~30,30~40和40~50 cm等5个土层的土壤样品,研究AM真菌在土壤中的空间分布及其与土壤脲酶、蛋白酶、酸性磷酸酶活性和碱解N、速效P含量的关系。【结果】AM真菌定殖率及孢子密度与样地生态条件密切相关,其中集宁样地孢子密度最高,靖边样地泡囊定殖率最高,包头样地丛枝、菌丝和总定殖率均高于其他样地;孢子密度与AM真菌不同结构定殖率间无显著相关性。土壤酸性磷酸酶活性对孢子密度有极显著正的效应;脲酶活性对孢子密度和丛枝定殖率有极显著的正效应;蛋白酶活性对丛枝定殖率、菌丝定殖率、总定殖率均有极显著正效应。土壤蛋白酶与泡囊定殖率呈显著负相关。碱解氮、速效磷对与以上3种土壤酶活性均呈极显著正相关。【结论】沙漠生境下AM真菌与土壤酶活性显著相关。AM真菌孢子密度和菌根不同结构的定殖程度可作为荒漠土壤生态系统环境状况检测的有效指标。
DOI:10.1016/j.soilbio.2009.02.010URL
To measure and manage plant growth in arid and semi-arid sandlands, improved understanding of the spatial patterns of desert soil resources and the role of arbuscular mycorrhizal (AM) fungi is needed. Spatial patterns of AM fungi, glomalin and soil enzyme activities were investigated in five plots located in the Mu Us sandland, northwestern China. Soils to 5002cm depth in the rhizosphere of Astragalus adsurgens Pall. were sampled. The study demonstrated that A. adsurgens Pall. could form strong symbiotic relationships with AM fungi. Arbuscular mycorrhizal fungal status and distributions were significantly different among the five studied plots. Correlation coefficient analysis demonstrated that spore density was significantly and positively correlated with soil organic carbon (SOC), soil acid phosphatase and to two Bradford-reactive soil protein (BRSP) fractions ( P02<020.01). Colonization of arbuscules and vesicles were positively correlated with protease activity. The BRSP fractions were also significantly and positively correlated to edaphic factors (e.g. SOC, available nitrogen, and Olsen phosphorus) and soil enzymes (e.g. soil urease and acid phosphatase). The means of total BRSP and easily extractable BRSP were 0.9502mg02g 611 and 0.502mg02g 611 in all data, respectively. The levels of BRSP in the desert soil were little lower than those in native and arable soils, but the ratios of BRSP to SOC were much higher than farmland soils. The results of this study support the conclusion that glomalin could be an appropriate index related to the level of soil fertility, especially in desert soil. Moreover, AM fungal colonizations and glomalin might be useful to monitor desertification and soil degradation.
.
DOI:10.1111/j.1461-0248.2008.01254.xURLPMID:19019195
Abstract Mutualisms, beneficial interactions between species, are expected to be unstable because delivery of benefit likely involves fitness costs and selection should favour partners that deliver less benefit. Yet, mutualisms are common and persistent, even in the largely promiscuous associations between plants and soil microorganisms such as arbuscular mycorrhizal fungi. In two different systems, we demonstrate preferential allocation of photosynthate by host plants to the more beneficial of two AM fungal symbionts. This preferential allocation could allow the persistence of the mutualism if it confers sufficient advantage to the beneficial symbiont that it overcomes the cost of mutualism. We find that the beneficial fungus does increase in biomass when the fungi are spatially separated within the root system. However, in well-mixed fungal communities, non-beneficial fungi proliferate as expected from their reduced cost of mutualism. Our findings suggest that preferential allocation within spatially structured microbial communities can stabilize mutualisms between plants and root symbionts.
.
DOI:10.1007/s13199-012-0186-yURL [本文引用: 1]
The spatial diversity and distribution of AM fungi were investigated in three plots located in farming–pastoral zone, north China. The rhizospheres of Caragana korshinskii , Artemisia sphaerocephala and Salix psammophila were sampled and thirty AM fungal species belonging to five genera were isolated. The study demonstrated that AM fungal diversity and distribution differed significantly among the three host plants and the three studied plots. Spore density of AM fungi ranged between 2 and 22 spores per g 611 of soil and species richness between 8 and 23. Correlation coefficient analysis demonstrated that spore density was significantly and positively correlated with soil organic matter and available N ( P 65<650.01). Species richness was significantly and positively correlated with soil organic matter and available P ( P 65<650.01), but significantly and negatively correlated with soil pH ( P 65<650.01). Finally, the Shannon–Weiner index was significantly and positively correlated with soil organic matter ( P 65<650.05). In this farming–pastoral zone, Glomus reticulatum and G . melanosporum may be more adaptable to the arid conditions than other AM fungal species. This research into AM fungal diversity may lead to exploitation of AM fungi for the mitigation of soil erosion and desertification using mycorrhizal plants, such as C.korshinskii , A . sphaerocephala and S . psammophila . The results of this study support the conclusion that diversity and distribution of AM fungi might be useful to monitor desertification and soil degradation.
.
DOI:10.1126/science.aab1161URLPMID:26315436
Abstract The global biogeography of microorganisms remains largely unknown, in contrast to the well-studied diversity patterns of macroorganisms. We used arbuscular mycorrhizal (AM) fungus DNA from 1014 plant-root samples collected worldwide to determine the global distribution of these plant symbionts. We found that AM fungal communities reflected local environmental conditions and the spatial distance between sites. However, despite AM fungi apparently possessing limited dispersal ability, we found 93% of taxa on multiple continents and 34% on all six continents surveyed. This contrasts with the high spatial turnover of other fungal taxa and with the endemism displayed by plants at the global scale. We suggest that the biogeography of AM fungi is driven by unexpectedly efficient dispersal, probably via both abiotic and biotic vectors, including humans. Copyright 脗漏 2015, American Association for the Advancement of Science.
.
DOI:10.5846/stxb201105020571URL [本文引用: 2]
植物化学计量学特征在大尺度上主要受纬度和经度两个因素影响。纬度梯度上温度因子变化对植物化学计量特征的影响已有大量研究,但是关于经度梯度上降雨因子变化对植物化学计量特征影响的研究却较少。选取呼伦贝尔草原,研究经度梯度上植物化学计量特征和土壤养分指标的变化规律,从经度梯度和养分供给两方面分析植物群落化学计量特征的变化规律,研究结果如下:1)植物群落叶片C含量变化范围为440.76—452.72 mg/g,N含量变化范围为17.79—30.88 mg/g,P含量变化范围为1.31—1.71 mg/g;群落叶片C含量、C/N随经度升高显著增加;群落叶片N含量随经度升高显著下降;植物群落P含量也呈下降趋势,但是关系不显著;植物群落C、N和P元素总量随着经度升高而显著增加。2)0—10 cm土壤全碳、全氮、全磷、有机碳受降雨量变化和植物群落元素总量影响,随着经度梯度升高而增加;但土壤铵态氮、硝态氮在经度梯度上没有表现出规律性的变化趋势。3)土壤全碳、有机碳、全氮、全磷和速效磷与植物群落叶片C、N和P含量没有显著相关关系,但与植物群落C、N和P元素总量呈显著正相关关系。该区土壤有效磷含量(8.13 mg/kg)高于全球平均值(7.65 mg/kg),但植物群落叶片磷含量平均值(1.5 mg/g)低于全球平均值(1.77 mg/g)。通过研究结果推测:植物通过对气候条件的长期适应,群落水平C、N和P含量沿经度梯度形成一定的分布格局;降雨量影响植物群落元素积累的总量,从而与土壤养分含量呈显著正相关关系;植物叶片P含量低的原因并非是由于土壤中磷的供给不足所致,而是植物对环境长期适应形成的策略。
DOI:10.5846/stxb201105020571URL [本文引用: 2]
植物化学计量学特征在大尺度上主要受纬度和经度两个因素影响。纬度梯度上温度因子变化对植物化学计量特征的影响已有大量研究,但是关于经度梯度上降雨因子变化对植物化学计量特征影响的研究却较少。选取呼伦贝尔草原,研究经度梯度上植物化学计量特征和土壤养分指标的变化规律,从经度梯度和养分供给两方面分析植物群落化学计量特征的变化规律,研究结果如下:1)植物群落叶片C含量变化范围为440.76—452.72 mg/g,N含量变化范围为17.79—30.88 mg/g,P含量变化范围为1.31—1.71 mg/g;群落叶片C含量、C/N随经度升高显著增加;群落叶片N含量随经度升高显著下降;植物群落P含量也呈下降趋势,但是关系不显著;植物群落C、N和P元素总量随着经度升高而显著增加。2)0—10 cm土壤全碳、全氮、全磷、有机碳受降雨量变化和植物群落元素总量影响,随着经度梯度升高而增加;但土壤铵态氮、硝态氮在经度梯度上没有表现出规律性的变化趋势。3)土壤全碳、有机碳、全氮、全磷和速效磷与植物群落叶片C、N和P含量没有显著相关关系,但与植物群落C、N和P元素总量呈显著正相关关系。该区土壤有效磷含量(8.13 mg/kg)高于全球平均值(7.65 mg/kg),但植物群落叶片磷含量平均值(1.5 mg/g)低于全球平均值(1.77 mg/g)。通过研究结果推测:植物通过对气候条件的长期适应,群落水平C、N和P含量沿经度梯度形成一定的分布格局;降雨量影响植物群落元素积累的总量,从而与土壤养分含量呈显著正相关关系;植物叶片P含量低的原因并非是由于土壤中磷的供给不足所致,而是植物对环境长期适应形成的策略。
[本文引用: 2]
[本文引用: 2]
.
DOI:10.1016/j.funeco.2009.07.003URL [本文引用: 1]
.
DOI:10.1007/s00442-008-1117-8URLPMID:18690479
Arbuscular mycorrhizal fungi (AMF) are mutualistic with most species of plants and are known to influence plant community diversity and composition. To better understand natural plant communities and the ecological processes. they control it is important to understand what determines the distribution and diversity of AMF. We tested three putative niche axes: plant species composition, disturbance history, and soil chemistry against AMF species composition to determine which axis correlated most strongly with a changing AMF community. Due to a scale dependency we were not able to absolutely rank their importance, but we did find that each correlated significantly with AMF community change at our site. Among soil properties, pH and NO鈧 were found to be especially good predictors of AMF community change. In a similar analysis of the plant community we found that time since disturbance had by far the largest impact on community composition.
.
DOI:10.1007/s00572-010-0333-3URLPMID:20697748 [本文引用: 1]
The beneficial effects of arbuscular mycorrhizal (AM) fungi on plant performance and soil health are essential for the sustainable management of agricultural ecosystems. Nevertheless, since the ‘first green revolution’, less attention has been given to beneficial soil microorganisms in general and to AM fungi in particular. Human society benefits from a multitude of resources and processes from natural and managed ecosystems, to which AM make a crucial contribution. These resources and processes, which are called ecosystem services, include products like food and processes like nutrient transfer. Many people have been under the illusion that these ecosystem services are free, invulnerable and infinitely available; taken for granted as public benefits, they lack a formal market and are traditionally absent from society’s balance sheet. In 1997, a team of researchers from the USA, Argentina and the Netherlands put an average price tag of US $33 trillion a year on these fundamental ecosystem services. The present review highlights the key role that the AM symbiosis can play as an ecosystem service provider to guarantee plant productivity and quality in emerging systems of sustainable agriculture. The appropriate management of ecosystem services rendered by AM will impact on natural resource conservation and utilisation with an obvious net gain for human society.
DOI:10.5846/stxb201503300609URL [本文引用: 1]
进一步探究荒漠植物与AM(arbuscular mycorrhiza)真菌共生关系及其生态适应性,为以蒙古沙冬青为建群种适生区的植被恢复与生态改良提供依据.于2013年6月在内蒙古荒漠带选取以蒙古沙冬青为建群种的3个样地乌海、磴口和阿拉善,从每个样地选择2种主要伴生植物,按0-10、10-20、20-30、30-40、40-50 cm共5个土层采集土样和根样,研究了蒙古沙冬青伴生植物AM真菌空间分布及其与土壤因子的关系.从梭梭(Haloxylon ammodendron)、油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和蒙古扁桃(Amygdalus mongolica)4种伴生植物根围土壤共分离鉴定4属25种AM真菌,其中球囊霉属(Glomus)14种,无梗囊霉属(Acaulospora)7种,管柄囊霉属(Funneliformis)3种,盾巨孢囊霉属(Scutellospora)1种,优势菌种为网状球囊霉(Glomus reticulatum),AM真菌属种分布具有不均衡性和地域性.4种伴生植物根系均能与AM真菌形成Ⅰ-型(intermediate type)丛枝菌根,其共生程度和定殖规律具有明显空间异质性.AM真菌种数随土层深度增加而下降.AM真菌最大定殖率在10-30 cm土层,最大孢子密度在10-20 cm土层.相关性分析表明,AM真菌菌丝与土壤有机C极显著正相关(P<0.01),与易提取球囊霉素(EEG)显著负相关(P<0.05);孢子密度与有机C、碱性磷酸酶极显著负相关(P<0.01),与碱解N极显著正相关(P<0.01).主成分分析表明,土壤有效P、酸性磷酸酶、碱性磷酸酶和总球囊霉素(TEG)等土壤因子能综合反映内蒙古荒漠带营养状况.TEG和EEG平均含量分别为4.76 mg/g和1.62 mg/g,占土壤有机C平均含量为61.26%和20.8%,说明在贫瘠荒漠环境中球囊霉素是土壤有机碳库重要来源和组成部分.
DOI:10.5846/stxb201503300609URL [本文引用: 1]
进一步探究荒漠植物与AM(arbuscular mycorrhiza)真菌共生关系及其生态适应性,为以蒙古沙冬青为建群种适生区的植被恢复与生态改良提供依据.于2013年6月在内蒙古荒漠带选取以蒙古沙冬青为建群种的3个样地乌海、磴口和阿拉善,从每个样地选择2种主要伴生植物,按0-10、10-20、20-30、30-40、40-50 cm共5个土层采集土样和根样,研究了蒙古沙冬青伴生植物AM真菌空间分布及其与土壤因子的关系.从梭梭(Haloxylon ammodendron)、油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和蒙古扁桃(Amygdalus mongolica)4种伴生植物根围土壤共分离鉴定4属25种AM真菌,其中球囊霉属(Glomus)14种,无梗囊霉属(Acaulospora)7种,管柄囊霉属(Funneliformis)3种,盾巨孢囊霉属(Scutellospora)1种,优势菌种为网状球囊霉(Glomus reticulatum),AM真菌属种分布具有不均衡性和地域性.4种伴生植物根系均能与AM真菌形成Ⅰ-型(intermediate type)丛枝菌根,其共生程度和定殖规律具有明显空间异质性.AM真菌种数随土层深度增加而下降.AM真菌最大定殖率在10-30 cm土层,最大孢子密度在10-20 cm土层.相关性分析表明,AM真菌菌丝与土壤有机C极显著正相关(P<0.01),与易提取球囊霉素(EEG)显著负相关(P<0.05);孢子密度与有机C、碱性磷酸酶极显著负相关(P<0.01),与碱解N极显著正相关(P<0.01).主成分分析表明,土壤有效P、酸性磷酸酶、碱性磷酸酶和总球囊霉素(TEG)等土壤因子能综合反映内蒙古荒漠带营养状况.TEG和EEG平均含量分别为4.76 mg/g和1.62 mg/g,占土壤有机C平均含量为61.26%和20.8%,说明在贫瘠荒漠环境中球囊霉素是土壤有机碳库重要来源和组成部分.
.
DOI:10.1038/ismej.2012.127URLPMID:23096401
Arbuscular fungi have a major role in directing the functioning of terrestrial ecosystems yet little is known about their biogeographical distribution. The Baas-Becking hypothesis ('everything is everywhere, but, the environment selects') was tested by investigating the distribution of arbuscular mycorrhizal fungi (AMF) at the landscape scale and the influence of environmental factors and geographical distance in determining community composition. AMF communities in Trifolium repens and Lolium perenne roots were assessed in 40 geographically dispersed sites in Ireland representing different land uses and soil types. Field sampling and laboratory bioassays were used, with AMF communities characterised using 18S rRNA terminal-restriction fragment length polymorphism. Landscape-scale distribution of AMF was driven by the local environment. AMF community composition was influenced by abiotic variables (pH, rainfall and soil type), but not land use or geographical distance. Trifolium repens and L. perenne supported contrasting communities of AMF, and the communities colonising each plant species were consistent across pasture habitats and over distance. Furthermore, L. perenne AMF communities grouped by soil type within pasture habitats. This is the largest and most comprehensive study that has investigated the landscape-scale distribution of AMF. Our findings support the Baas-Becking hypothesis at the landscape scale and demonstrate the strong influence the local environment has on determining AMF community composition. The ISME Journal (2013) 7, 498-508; doi:10.1038/ismej.2012.127; published online 25 October 2012
DOI:10.5846/stxb201104270557URL [本文引用: 1]
为了阐明荒漠柠条锦鸡儿(Caragana korshinskii Kom.)根围AM真菌群落组成及其分布特点,2010年7月从河北省与内蒙古交界荒漠带选择多伦湖、黑城子和二羊点3个样地,按0—10、10—20、20—30、30—40、40—50 cm 5个土层采集柠条根围土壤样品,研究了柠条根围AM真菌物种组成、生态分布及与土壤因子的相关性。共分离鉴定出4属24种AM真菌,其中球囊霉属(Glomus)16种,是3个样地共同优势属;无梗囊霉属(Acaulospora)5种,是3个样地共有属;盾巨孢囊霉属(Scutellospora)2种;多孢囊霉属(Diversispora)1种;网状球囊霉(G.reticulatum)是黑城子和多伦湖样地优势种,二羊点亚优势种;黑球囊霉(G.melanosporum)是黑城子和二羊点样地优势种,多伦湖亚优势种;二羊点的AM真菌种类和孢子密度最高;黑城子和多伦湖样地无明显差异。AM真菌孢子密度与土壤碱解N、有机质和速效P含量极显著正相关,其中土壤碱解N影响最大。结果表明,柠条根围AM真菌物种多样性丰富,具有明显空间异质性,并与土壤因子关系密切,这为进一步分离筛选优势AM真菌菌种,充分利用AM真菌资源促进荒漠柠条生长提供了依据。
DOI:10.5846/stxb201104270557URL [本文引用: 1]
为了阐明荒漠柠条锦鸡儿(Caragana korshinskii Kom.)根围AM真菌群落组成及其分布特点,2010年7月从河北省与内蒙古交界荒漠带选择多伦湖、黑城子和二羊点3个样地,按0—10、10—20、20—30、30—40、40—50 cm 5个土层采集柠条根围土壤样品,研究了柠条根围AM真菌物种组成、生态分布及与土壤因子的相关性。共分离鉴定出4属24种AM真菌,其中球囊霉属(Glomus)16种,是3个样地共同优势属;无梗囊霉属(Acaulospora)5种,是3个样地共有属;盾巨孢囊霉属(Scutellospora)2种;多孢囊霉属(Diversispora)1种;网状球囊霉(G.reticulatum)是黑城子和多伦湖样地优势种,二羊点亚优势种;黑球囊霉(G.melanosporum)是黑城子和二羊点样地优势种,多伦湖亚优势种;二羊点的AM真菌种类和孢子密度最高;黑城子和多伦湖样地无明显差异。AM真菌孢子密度与土壤碱解N、有机质和速效P含量极显著正相关,其中土壤碱解N影响最大。结果表明,柠条根围AM真菌物种多样性丰富,具有明显空间异质性,并与土壤因子关系密切,这为进一步分离筛选优势AM真菌菌种,充分利用AM真菌资源促进荒漠柠条生长提供了依据。
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 2]
URL
2007年5月、8月和10月分别从宁夏沙坡头地区豆科植物沙冬青(Ammopiptanthus mongolicus)根围分0-10cm,10-20cm,20-30cm,30-40cm和40-50cm5个土层采集土壤样品。研究了AM真菌时空分布及与土壤因子的相关性,在分离的3属12种AM真菌中,球囊霉属(Glomus)6种,无梗囊霉属(Acaulospora)4种,盾巨孢囊霉属(Scutellospora)2种,3属12种AM真菌在3个时期均有分布。AM真菌定殖率与孢子密度有明显的时空分布规律,并与土壤因子关系密切。孢子密度和泡囊定殖率在8月份最高,丛枝、菌丝和总定殖率10月份最高。土壤采样深度对孢子密度和定殖率有显著影响,AM真菌最高定殖率和最大孢子密度均出现在0-30cm浅土层。孢子密度与土壤有机质、碱解N极显著正相关,与pH显著正相关;菌丝、总定殖率与pH、碱解N和速效P极显著正相关。在评估荒漠生态系统和沙冬青形成菌根的能力时,AM真菌孢子密度、菌根不同结构的定殖程度是十分有用的指标。
URL
2007年5月、8月和10月分别从宁夏沙坡头地区豆科植物沙冬青(Ammopiptanthus mongolicus)根围分0-10cm,10-20cm,20-30cm,30-40cm和40-50cm5个土层采集土壤样品。研究了AM真菌时空分布及与土壤因子的相关性,在分离的3属12种AM真菌中,球囊霉属(Glomus)6种,无梗囊霉属(Acaulospora)4种,盾巨孢囊霉属(Scutellospora)2种,3属12种AM真菌在3个时期均有分布。AM真菌定殖率与孢子密度有明显的时空分布规律,并与土壤因子关系密切。孢子密度和泡囊定殖率在8月份最高,丛枝、菌丝和总定殖率10月份最高。土壤采样深度对孢子密度和定殖率有显著影响,AM真菌最高定殖率和最大孢子密度均出现在0-30cm浅土层。孢子密度与土壤有机质、碱解N极显著正相关,与pH显著正相关;菌丝、总定殖率与pH、碱解N和速效P极显著正相关。在评估荒漠生态系统和沙冬青形成菌根的能力时,AM真菌孢子密度、菌根不同结构的定殖程度是十分有用的指标。
.
DOI:10.3969/j.issn.1000-1565.2006.06.017URL [本文引用: 1]
依据种的现代分布、替代关系、 地质历史、古气候和化石资料,探讨了中国岩黄耆属植物区系的发生、演变及形成机制.结果表明:岩黄耆属可能是由古地中海旱生植物区系中一类中生的原始灌木 类群,在晚始新世到早渐新世或比这个时期稍晚一些,经过第三纪以来的环境分异变化,逐渐形成适应高寒山地生长的扁荚组种类和适应干旱环境生长的多茎组、短 茎组种类,短茎组种类是多茎组对干旱强化进一步适应的结果,与此同时,原始的灌木类群自身也在继续演化,形成适应荒漠半荒漠生长的种类.这种演化过程揭示 出生存环境中水热条件的剧烈变化是植物演化的主要动力.
DOI:10.3969/j.issn.1000-1565.2006.06.017URL [本文引用: 1]
依据种的现代分布、替代关系、 地质历史、古气候和化石资料,探讨了中国岩黄耆属植物区系的发生、演变及形成机制.结果表明:岩黄耆属可能是由古地中海旱生植物区系中一类中生的原始灌木 类群,在晚始新世到早渐新世或比这个时期稍晚一些,经过第三纪以来的环境分异变化,逐渐形成适应高寒山地生长的扁荚组种类和适应干旱环境生长的多茎组、短 茎组种类,短茎组种类是多茎组对干旱强化进一步适应的结果,与此同时,原始的灌木类群自身也在继续演化,形成适应荒漠半荒漠生长的种类.这种演化过程揭示 出生存环境中水热条件的剧烈变化是植物演化的主要动力.
URL
URL
URL [本文引用: 2]
URL [本文引用: 2]
.
DOI:10.1002/jpln.19610950107URL
First page of article
.
DOI:10.2307/3793161URL [本文引用: 1]
Extraction methods for vesicular-arbuscular mycorrhizal fungal spores were compared using soils from arid habitats. The methods were wet-sieving and decanting, flotation-adhesion, two density gradient centrifugation techniques and differential water/sucrose centrifugation. In clay soils, all methods except differential water/sucrose centrifugation were statistically similar. Differential water/sucrose centrifugation extracted 185% more spores than the other methods. When comparing methods using soils of differing textures, the efficiency of extraction of Gigaspora spores was similar, but more Glomus spores were extracted with differential centrifugation. Exposure to a 2 M solution dehydrated 52% of exposed spores in 24 h. Following immersion of those spores in distilled water, 98% were hydrated after 24 h. The concentration of sucrose used in the differential centrifugation method appeared to cause little permanent damage to spores. We conclude that several methods should be compared initially when spores are extracted from new habitats to ascertain the optimum technique for the given soils.
.
URL [本文引用: 1]
URL [本文引用: 1]
.
DOI:10.1111/j.1469-8137.2004.01202.xURL
DOI:10.3969/j.issn.1000-7601.2012.05.034URL [本文引用: 1]
基于SPOTVGT—NDVI影像数据,通过最大值合成法、均值法、线性回归以及最小二乘法 等研究方法分析西北五省1999-2007年9a问的植被覆盖时空演变与动态变化规律。结果表明:9a间西北地区的植被覆盖有明显改善趋势,改善面积百分 比为26.92%;部分地区植被覆盖仍有退化现象,面积百分比为1.22%,但其明显小于植被改善比例;除陕西省外,西北地区植被覆盖整体不高;7、8月 份该区域植被覆盖达到一年中最大值。
DOI:10.3969/j.issn.1000-7601.2012.05.034URL [本文引用: 1]
基于SPOTVGT—NDVI影像数据,通过最大值合成法、均值法、线性回归以及最小二乘法 等研究方法分析西北五省1999-2007年9a问的植被覆盖时空演变与动态变化规律。结果表明:9a间西北地区的植被覆盖有明显改善趋势,改善面积百分 比为26.92%;部分地区植被覆盖仍有退化现象,面积百分比为1.22%,但其明显小于植被改善比例;除陕西省外,西北地区植被覆盖整体不高;7、8月 份该区域植被覆盖达到一年中最大值。
[本文引用: 1]
.
DOI:10.1023/A:1020388910724URL [本文引用: 1]
Myrkdalen Lake in Western Norway was subjected to a 1.4 m drawdown in June 1987. Plant establishment and vegetation succession on the exposed sediments of a fluvial delta plain was monitored through 2000. The investigated area extended from the original Equisetum fluviatile zone to the new lake edge. The substrate was homogeneous and consisted mainly of minerogenous fluvial sediments. Vegetation data were sampled within continuous quadrats along transects perpendicular to the shore, and they dropped 93 cm in elevation. Detrended Correspondence Analysis and Canonical Correspondence Analysis confirmed that "time since drawdown" and "elevation of the quadrats" appeared to be of nearly equal importance in explaining succession. Plant establishment was rapid on the exposed sediments due to a seed bank and to rapid invasion of plants. The succession includes both floristic change as a function of time and a spatial separation in relation to the water level. The species succession was marked by a growth from progression: mosses and annuals 鈫 non-clonal perennials 鈫 clonal perennials. After one month, the annual Subularia aquatica and small acrocarpous mosses dominated the site. Dominance then shifted to Deschampsia cespitosa, Juncus filiformis, Blasia pusilla and Polytrichum commune. Subsequently there was an increase of Carex vesicaria in the lower zone and Calamagrostis purpurea in the upper zone, while Phalaris arundinacea was common over most of the elevational gradient. Equisetum declined where it had dominated before drawdown, but it expanded gradually towards the new shoreline. The vegetation remained dynamic after 13 years and it is not considered to be in equilibrium with the new environmental gradient yet. However, annual changes measured by DCA scores have slowed and two vegetation zones have developed. The major vegetational differences along the elevational gradient can be explained by the height of the mean June water limit. This example of species turnover in space and time may be a model for other successions that occur along a strong gradient.
.
DOI:10.3732/ajb.89.9.1439URLPMID:21665745
Abstract Interest in the diversity of arbuscular mycorrhizal (AM) fungal communities has been stimulated by recent data that demonstrate that fungal communities influence the competitive hierarchies, productivity, diversity, and successional patterns of plant communities. Although natural communities of AM fungi are diverse, we have a poor understanding of the mechanisms that promote and maintain that diversity. Plants may coexist by inhabiting disparate temporal niches; plants of many grasslands are either warm or cool season specialists. We hypothesized that AM fungi might be similarly seasonal. To test our hypothesis, we tracked the sporulation of individual AM fungal species growing within a North Carolina grassland. Data were collected in 1996 and 1997; in 1997, sampling focused on two common species. We found that AM fungi, especially Acaulospora colossica and Gigaspora gigantea , maintained different and contrasting seasonalities. Acaulospora colossica sporulated more frequently in the warm season, but Gi. gigantea sporulated more frequently in the cool season. Moreover, AM fungal species were spatially aggregated at a fine scale. Contrasting seasonal and spatial niches may facilitate the maintenance of a diverse community of AM fungi. Furthermore, these data may illuminate our understanding of the AM fungal influence on plant communities: various fungal species may preferentially associate with different plant species and thereby promote diversity in the plant community.
.
DOI:10.1111/j.1461-0248.2004.00620.xURL [本文引用: 1]
Abstract Arbuscular mycorrhizal fungi (AMF; phylum Glomeromycota) are ubiquitous in terrestrial ecosystems. Despite their acknowledged importance in ecology, most research on AMF has focused on effects on individual plant hosts, with more recent efforts aimed at the level of the plant community. Research at the ecosystem level is less prominent, but potentially very promising. Numerous human-induced disturbances (including global change and agro-ecosystem management) impinge on AMF functioning; hence study of this symbiosis from the ecosystem perspective seems timely and crucial. In this paper, I discuss four (interacting) routes via which AMF can influence ecosystem processes. These include indirect pathways (through changes in plant and soil microbial community composition), and direct pathways (effects on host physiology and resource capture, and direct mycelium effects). I use the case study of carbon cycling to illustrate the potentially pervasive influence of AMF on ecosystem processes. A limited amount of published research on AMF ecology is suited for direct integration into ecosystem studies (because of scale mismatch or ill-adaptation to the 鈥榩ools and flux鈥 paradigm of ecosystem ecology); I finish with an assessment of the tools (experimental designs, response variables) available for studying mycorrhizae at the ecosystem scale.
.
DOI:10.1111/j.1469-8137.2008.02378.xURLPMID:18248587 [本文引用: 1]
Abstract Contents Summary 1 I. Introduction 1 II.鈥 Taxonomy and species recognition 2 III. Communities of arbuscular mycorrhizal fungi 4 IV. Populations of arbuscular mycorrhizal fungi 6 V. Individuals, genets or clones? 8 VI.鈥 Speciation in Glomeromycota 9 VII. Conclusion 10 Acknowledgements 11 References 11 Summary Arbuscular mycorrhizal fungi in the phylum Glomeromycota are found globally in most vegetation types, where they form a mutualistic symbiosis with plant roots. Despite their wide distribution, only relatively few species are described. The taxonomy is based on morphological characters of the asexual resting spores, but molecular approaches to community ecology have revealed a considerable unknown diversity from colonized roots. Although the lack of genetic recombination is not unique in the fungal kingdom, arbuscular mycorrhizal fungi are probably ancient asexuals. The long asexual evolution of the fungi has resulted in considerable genetic diversity within morphologically recognizable species, and challenges our concepts of individuals and populations. This review critically examines the concepts of species, communities, populations and individuals of arbuscular mycorrhizal fungi.
.
DOI:10.1111/j.1469-8137.2006.01799.xURLPMID:16945098 [本文引用: 1]
The community of arbuscular mycorrhizal fungi (AMF) colonizing the roots of Festuca pratensis and Achillea millefolium was characterized in a Swedish pasture at different times, along a gradient of fertilization. The small subunit ribosomal RNA gene was subjected to PCR and denaturing gradient gel electrophoresis (DGGE), sequencing and phylogenetic analysis. The sequences found in this study clustered in 10 discrete sequence groups, seven belonging to Glomus, two to Scutellospora and one to Diversispora. A negative correlation was observed between soil mineral nitrogen and the number of AMF sequence groups in the roots. The frequency of occurrence of AMF in roots decreased dramatically between June and September. No plant-host specificity could be detected.
.
DOI:10.1016/0038-0717(94)90288-7URL
Mycorrhizal inoculation increased plant dry weight. P content and total P uptake irrespectively of P sources and soils. Of the total P uptake mycorrhizal contribution accounted for 24–33% with P supplied in inorganic form, and 48–59% with P supplied in organic form. The results demonstrate a stimulatory effect of organic P in the form of phytate on mycorrhizal infection and hyphal growth as well as the efficient use of phytate-P by phosphatase of mycorrhizal hyphae.
.
DOI:10.1046/j.1469-8137.2002.00439.xURL [本文引用: 1]
61 To test the response of arbuscular mycorrhizal (AM) fungi to a difference in soil pH, the extraradical mycelium of Scutellospora calospora or Glomus intraradices, in association with Plantago lanceolata, was exposed to two different pH treatments, while the root substrate pH was left unchanged. 61 Seedlings of P. lanceolata, colonized by one or other of the fungal symbionts, and nonmycorrhizal controls, were grown in mesh bags placed in pots containing pH-buffered sand (pH around 5 or 6). The systems were harvested at approximately 2-wk intervals between 20 and 80 d. 61 Both fungi formed more extraradical mycelium at the higher pH. Glomus intraradices formed almost no detectable extraradical mycelium at lower pH. The extraradical mycelium of S. calospora had higher acid phosphatase activity than that of G. intraradices. Total AM root colonization decreased for both fungi at the higher pH, and high pH also reduced arbuscule and vesicle formation in G. intraradices. 61 In conclusion, soil pH influences AM root colonization as well as the growth and phosphatase activities of extraradical mycelium, although the two fungi responded differently.
.
DOI:10.1111/j.1469-8137.2006.01854.xURLPMID:17083685 [本文引用: 1]
Summary 6168 We investigated the effects of arbuscular mycorrhizal fungal (AMF) species richness and composition on plant community productivity and diversity, and whether AMF mediate plant species coexistence by promoting niche differentiation in phosphorus use. 6168 Our experiment manipulated AMF species richness and identity across a range of P conditions in tallgrass prairie mesocosms. 6168 We showed that increasing AMF richness promoted plant diversity and productivity, but that this AMF richness effect was small relative to the effects of individual AMF species. We found little support for AMF-facilitated complementarity in P use. Rather, the AMF richness effect appeared to be caused by the inclusion of particular diversity- and productivity-promoting AMF (a sampling effect). Furthermore, the identity of the diversity-promoting fungi changed with P environment, as did the relationship between the diversity-promoting and productivity-promoting benefits of AMF. 6168 Our results suggest that plant diversity and productivity are more responsive to AMF identity than to AMF diversity per se , and that AMF identity and P environment can interact in complex ways to alter community-level properties.
.
DOI:10.1111/j.1461-0248.2011.01666.xURLPMID:21790936 [本文引用: 1]
Soil microbes play key roles in ecosystems, yet the impact of their diversity on plant communities is still poorly understood. Here we demonstrate that the diversity of belowground plant-associated soil fungi promotes plant productivity and plant coexistence. Using additive partitioning of biodiversity effects developed in plant biodiversity studies, we demonstrate that this positive relationship can be driven by complementarity effects among soil fungi in one soil type and by a selection effect resulting from the fungal species that stimulated plant productivity the most in another soil type. Selection and complementarity effects among fungal species contributed to improving plant productivity up to 82% and 85%, respectively, above the average of the respective fungal species monocultures depending on the soil in which they were grown. These results also indicate that belowground diversity may act as insurance for maintaining plant productivity under differing environmental conditions.
.
DOI:10.1023/A:1004347701584URL
Understanding the contributions of soil microorganisms to soil stabilization at the molecular level will lead to ways to enhance inputs for sustainable agricultural systems. Recent discoveries of copious production of glycoprotein (glomalin) by arbuscular mycorrhizal (AM) fungi and the apparent recalcitrance of this material in soils led to the comparison between concentration of glomalin and aggregate stability. Stability was measured on air-dried aggregates rewetted by capillary action and then subjected to wet sieving for 10 min. Thirty-seven samples from four geographic areas of the U.S. and one area of Scotland were tested. The monoclonal antibody used to discover glomalin on AM hyphae was employed to assess immunoreactive glomalin on aggregate surfaces by immunofluorescence and in extracts from aggregates by enzyme-linked immunosorbent assay (ELISA). Immunofluorescence was observed on at least some surfaces of aggregates from all soils examined, but was most evident on aggregates with high glomalin concentrations. Easily extractable glomalin (EEG) was solubilized by 20 mM citrate, pH 7.0 at 121 C for 30 min, and total glomalin (TG) was solubilized with 50 mM citrate, pH 8.0 at 121 C for 90 to 450 min. Some soils required up to seven sequential extractions to remove all of the glomalin. Aggregate stability was linearly correlated (p < 0.001) with all measures of glomalin (mg/g of aggregates) in these soils. The best predictor of aggregate stability (AS) was immunoreactive easily extractable glomalin (IREEG) according to the following relationship: AS = 42.7 +61.3 log10 IREEG (r2 = 0.86; p <0.001, n = 37).
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
.
URL
虽然VA真菌遍布全球,但其属和种的分布是不均衡的,常有程度不同的地域性。以新疆、北京和 吉林三地为代表的我国北方土壤中的VA真菌大都为球囊霉属Glomns。对摩西球囊霉G.mosseae、地表球囊霉G.versiforme及未发表种 Glomussp.9等菌种和某些生态因子的统计分析表明:土壤pH、土壤有机质含量和采集地海拔高度对三菌种的分布影响显著。在pH5-9.5范围内, 球囊霉属所占比例随土壤pH
URL
虽然VA真菌遍布全球,但其属和种的分布是不均衡的,常有程度不同的地域性。以新疆、北京和 吉林三地为代表的我国北方土壤中的VA真菌大都为球囊霉属Glomns。对摩西球囊霉G.mosseae、地表球囊霉G.versiforme及未发表种 Glomussp.9等菌种和某些生态因子的统计分析表明:土壤pH、土壤有机质含量和采集地海拔高度对三菌种的分布影响显著。在pH5-9.5范围内, 球囊霉属所占比例随土壤pH
.
DOI:10.3724/SP.J.1011.2013.00199URL [本文引用: 1]
中国科学院机构知识库(中国科学院机构知识库网格(CAS IR GRID))以发展机构知识能力和知识管理能力为目标,快速实现对本机构知识资产的收集、长期保存、合理传播利用,积极建设对知识内容进行捕获、转化、传播、利用和审计的能力,逐步建设包括知识内容分析、关系分析和能力审计在内的知识服务能力,开展综合知识管理。
DOI:10.3724/SP.J.1011.2013.00199URL [本文引用: 1]
中国科学院机构知识库(中国科学院机构知识库网格(CAS IR GRID))以发展机构知识能力和知识管理能力为目标,快速实现对本机构知识资产的收集、长期保存、合理传播利用,积极建设对知识内容进行捕获、转化、传播、利用和审计的能力,逐步建设包括知识内容分析、关系分析和能力审计在内的知识服务能力,开展综合知识管理。
.
DOI:10.1088/0256-307X/15/12/025URL [本文引用: 1]
阿拉善—鄂尔多斯是我国西北干旱区特有植物的集中分布区,有中国种子植物特有属15个。其中,当地特有属5个、戈壁荒漠特有属2个、蒙古高原特有属1个。有当地种子植物特有种128个,其中贺兰山等山地特有种62个、平原荒漠特有种66个。该中心特有植物的性质,表现为古地中海旱生植物后裔的古老性和温带干旱荒漠性质,以及山地森林植物与华北或东亚植物的亲缘联系
DOI:10.1088/0256-307X/15/12/025URL [本文引用: 1]
阿拉善—鄂尔多斯是我国西北干旱区特有植物的集中分布区,有中国种子植物特有属15个。其中,当地特有属5个、戈壁荒漠特有属2个、蒙古高原特有属1个。有当地种子植物特有种128个,其中贺兰山等山地特有种62个、平原荒漠特有种66个。该中心特有植物的性质,表现为古地中海旱生植物后裔的古老性和温带干旱荒漠性质,以及山地森林植物与华北或东亚植物的亲缘联系
Phosphorus fertilization management modifies the biodiversity of AM fungi in a tropical savanna forage system
2010
Occurrence of arbuscular mycorrhizal fungi in saline soils of the tabriz plain of iran in relation to some physical and chemical properties of soil
1
2001
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
荒漠境沙打旺根围AM真菌与土壤酶活性的关系
3
2009
... 土壤有机碳采用马弗炉烧失法(
... )测定; pH值用雷磁PHS-3C pH计测定; 氨态氮使用SMARTCHEM 200全自动连续分析仪(Alliance, Frépillon, France)测定; 有效磷采用碳酸氢钠-钼锑抗比色法(
... 西北荒漠带东西跨度极大, 受海陆位置影响, 由东到西干旱程度逐渐增加, 表现出荒漠草原、荒漠和极旱荒漠的景观变化.本试验中, 由东到西沿经度变化降雨量和土壤湿度逐渐减少, 受降雨影响, 花棒植株表现出对环境的强烈适应性, 这为AM真菌侵染和共生提供了适宜的生存环境.Bever等(2009)研究证实, 宿主植物种类和根系特性是AM真菌发生和分布的重要因子.豆科植物不仅能与AM真菌形成良好的共生联合体, 而且能够通过固氮、调节根系碳水化合物分配、产生次生代谢物质、改变根际土壤微环境等途径影响AM真菌的活动和分布(
荒漠境沙打旺根围AM真菌与土壤酶活性的关系
3
2009
... 土壤有机碳采用马弗炉烧失法(
... )测定; pH值用雷磁PHS-3C pH计测定; 氨态氮使用SMARTCHEM 200全自动连续分析仪(Alliance, Frépillon, France)测定; 有效磷采用碳酸氢钠-钼锑抗比色法(
... 西北荒漠带东西跨度极大, 受海陆位置影响, 由东到西干旱程度逐渐增加, 表现出荒漠草原、荒漠和极旱荒漠的景观变化.本试验中, 由东到西沿经度变化降雨量和土壤湿度逐渐减少, 受降雨影响, 花棒植株表现出对环境的强烈适应性, 这为AM真菌侵染和共生提供了适宜的生存环境.Bever等(2009)研究证实, 宿主植物种类和根系特性是AM真菌发生和分布的重要因子.豆科植物不仅能与AM真菌形成良好的共生联合体, 而且能够通过固氮、调节根系碳水化合物分配、产生次生代谢物质、改变根际土壤微环境等途径影响AM真菌的活动和分布(
Spatial distribution of arbuscular mycorrhizal fungi, glomalin and soil enzymes under the canopy of Astragalus adsurgens Pall. in the Mu Us sandland, China.
2009
Preferential allocation to beneficial symbiont with spatial structure maintains mycorrhizal mutualism
2009
Diversity of arbuscular mycorrhizal fungi in the rhizosphere of three host plants in the farming-pastoral zone, North China
1
2012
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism
2015
呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化
2
2012
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
... ).土壤pH值升高, 使土壤中金属离子对磷的吸附作用上升, 使土壤中有效磷含量降低(
呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化
2
2012
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
... ).土壤pH值升高, 使土壤中金属离子对磷的吸附作用上升, 使土壤中有效磷含量降低(
毛乌素沙地花棒(Hedysarum scoparium)根围AM真菌生态学研究
2
2008
... 我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢.花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
毛乌素沙地花棒(Hedysarum scoparium)根围AM真菌生态学研究
2
2008
... 我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢.花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
Functional diversity in arbuscular mycorrhiza—The role of gene expression, phosphorous nutrition and symbiotic efficiency
1
2010
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
Scale-dependent niche axes of arbuscular mycorrhizal fungi
2008
Agroecology: The key role of arbuscular mycorrhizas in ecosystem services
1
2010
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
蒙古沙冬青伴生植物AM真菌的空间分布
1
2016
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
蒙古沙冬青伴生植物AM真菌的空间分布
1
2016
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
The role of local environment and geographical distance in determining community composition of arbuscular mycorrhizal fungi at the landscape scale
2013
荒漠柠条锦鸡儿AM真菌多样性
1
2012
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
荒漠柠条锦鸡儿AM真菌多样性
1
2012
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
北方两省农牧交错带沙棘根围AM真菌与球囊霉素空间分布
1
2011
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
北方两省农牧交错带沙棘根围AM真菌与球囊霉素空间分布
1
2011
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
Dynamics of arbuscular mycorrhizal fungi and glomalin in the rhizosphere of Artemisia ordosica Krasch.
2
2010
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
... ), 有利于改善土壤理化性质, 改良和稳定土壤结构.同时, 土壤因子也会影响AM真菌生态分布、侵染能力、孢子密度等(
(
2010
(
2010
(
1
2006
... 我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢.花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(
(
1
2006
... 我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢.花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(
b). 河北省安国地区丹参根围AM真菌多样性
2010
b). 河北省安国地区丹参根围AM真菌多样性
2010
(
2
2006
... 研究发现, 花棒根际AM真菌各属呈现球囊霉属>无梗囊霉属>近明囊霉属>缩管柄囊霉属>管柄囊霉属>盾巨孢囊霉属, 球囊霉属和无梗囊霉属是各样地的优势属, 这与张美庆等(1994)认为球囊霉属和无梗囊霉属是广谱生态型的结论一致.本试验共分离到无梗囊霉属18种, 多于球囊霉属, 与以往报道(
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
(
2
2006
... 研究发现, 花棒根际AM真菌各属呈现球囊霉属>无梗囊霉属>近明囊霉属>缩管柄囊霉属>管柄囊霉属>盾巨孢囊霉属, 球囊霉属和无梗囊霉属是各样地的优势属, 这与张美庆等(1994)认为球囊霉属和无梗囊霉属是广谱生态型的结论一致.本试验共分离到无梗囊霉属18种, 多于球囊霉属, 与以往报道(
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
A colorimetric technique for determining urease activity in soil
1961
The effects of soil texture on extraction of vesicular-arbuscular mycorrhizal fungal spores from arid sites
1
1986
... 准确称取20 g风干土, 用湿筛倾析-蔗糖离心法(
西北干旱区AM真菌多样性研究
1
2007
... 研究发现, 花棒根际AM真菌各属呈现球囊霉属>无梗囊霉属>近明囊霉属>缩管柄囊霉属>管柄囊霉属>盾巨孢囊霉属, 球囊霉属和无梗囊霉属是各样地的优势属, 这与张美庆等(1994)认为球囊霉属和无梗囊霉属是广谱生态型的结论一致.本试验共分离到无梗囊霉属18种, 多于球囊霉属, 与以往报道(
西北干旱区AM真菌多样性研究
1
2007
... 研究发现, 花棒根际AM真菌各属呈现球囊霉属>无梗囊霉属>近明囊霉属>缩管柄囊霉属>管柄囊霉属>盾巨孢囊霉属, 球囊霉属和无梗囊霉属是各样地的优势属, 这与张美庆等(1994)认为球囊霉属和无梗囊霉属是广谱生态型的结论一致.本试验共分离到无梗囊霉属18种, 多于球囊霉属, 与以往报道(
Relationships among arbuscular mycorrhizal fungi, vascular plants and environmental conditions in oak savannas
2004
基于SPOT NDVI的植被覆盖时空演变规律分析——以西北五省为例
1
2012
... 采样区域位于我国西北荒漠带(73.15°- 111.17° E、31.32°-49.10° N), 属温带大陆性气候, 年降水量在东部为400 mm左右, 西部减少到200 mm甚至50 mm以下, 是一个从半干旱逐步过渡到极度干旱的完整荒漠系列, 自然景观由荒漠草原逐步演变为极旱荒漠(
基于SPOT NDVI的植被覆盖时空演变规律分析——以西北五省为例
1
2012
... 采样区域位于我国西北荒漠带(73.15°- 111.17° E、31.32°-49.10° N), 属温带大陆性气候, 年降水量在东部为400 mm左右, 西部减少到200 mm甚至50 mm以下, 是一个从半干旱逐步过渡到极度干旱的完整荒漠系列, 自然景观由荒漠草原逐步演变为极旱荒漠(
Tempo-spatial dynamics of arbuscular mycorrhizal fungi under clonal plant
1
2010
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
Thirteen years of wetland vegetation succession following a permanent drawdown, Myrkdalen Lake, Norway
1
2002
... 生物物种组成、群落结构及多样性是反映生物地理分布的重要指标(
Divergent phenologies may facilitate the coexistence of arbuscular mycorrhizal fungi in a north carolina grassland
2002
Arbuscular mycorrhizae and terrestrial ecosystem processes
1
2004
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
Communities, populations and individuals of arbuscular mycorrhizal fungi
1
2008
... 生物物种组成、群落结构及多样性是反映生物地理分布的重要指标(
Molecular analysis of arbuscular mycorrhizal fungi colonising a semi-natural grassland along a fertilisation gradient
1
2006
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
Manual for the identification of VA Mycorrhizal Fungi. 3rd edn. Synergistic, Gainesville
1990
Phosphatase activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus
1994
Arbuscular mycorrhizal fungi respond to the substrate pH of their extraradical mycelium by altered growth and root colonization
1
2002
... AM真菌作为联系宿主植物根系与土壤环境之间的桥梁, 其分布必然受到宿主植物和根际土壤环境的直接影响.土壤作为最适合AM真菌生存的天然培养基, 其成分变化直接影响着AM真菌的生态分布(
Mycorrhizal fungal identity and richness determine the diversity and productivity of a tallgrass prairie system
1
2006
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
Belowground biodiversity effects of plant symbionts support aboveground productivity
1
2011
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi
1998
中国岩黄耆属植物的生态分化及地理分布
2
1985
... 我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢.花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
中国岩黄耆属植物的生态分化及地理分布
2
1985
... 我国荒漠带主要分布在西北地区, 这里气候条件恶劣, 植物种类组成与群落结构简单, 生态系统极其脆弱, 极易受到破坏并且恢复缓慢.花棒(Hedysarum scoparium)是豆科岩黄耆属植物, 是亚洲内陆沙地的特有种, 广泛分布于我国西北的干旱和半干旱荒漠地区(
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
内蒙古荒漠沙柳AM真菌物种多样性
1
2011
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
内蒙古荒漠沙柳AM真菌物种多样性
1
2011
... 丛枝菌根(AM)真菌是一类分布广泛的土壤微生物, 能与绝大多数维管植物的根系形成互惠共生体(
我国北方VA菌根真菌某些属和种的生态分布
1994
我国北方VA菌根真菌某些属和种的生态分布
1994
毛乌素沙地典型克隆植物根际AM真菌多样性研究
1
2013
...
毛乌素沙地典型克隆植物根际AM真菌多样性研究
1
2013
...
阿拉善—鄂尔多斯生物多样性中心的特有植物和植物区系的性质
1
1999
... 采样区域位于我国西北荒漠带(73.15°- 111.17° E、31.32°-49.10° N), 属温带大陆性气候, 年降水量在东部为400 mm左右, 西部减少到200 mm甚至50 mm以下, 是一个从半干旱逐步过渡到极度干旱的完整荒漠系列, 自然景观由荒漠草原逐步演变为极旱荒漠(
阿拉善—鄂尔多斯生物多样性中心的特有植物和植物区系的性质
1
1999
... 采样区域位于我国西北荒漠带(73.15°- 111.17° E、31.32°-49.10° N), 属温带大陆性气候, 年降水量在东部为400 mm左右, 西部减少到200 mm甚至50 mm以下, 是一个从半干旱逐步过渡到极度干旱的完整荒漠系列, 自然景观由荒漠草原逐步演变为极旱荒漠(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}