Vertical distribution pattern of mixed root systems of desert plants Reaumuria soongarica and Salsola passerina under different environmental gradients
SHAN Li-Shan*, SU Ming, ZHANG Zheng-Zhong, WANG Yang, WANG Shan, LI YiCollege of Forestry Sciences, Gansu Agricultural University, Lanzhou 730070, China编委: 王根轩
责任编辑: 王葳
| 基金资助: |
Online:2018-04-20
| Fund supported: |

摘要
植物种间相互作用及其对环境胁迫的响应一直是物种共存和生物多样性维持研究的一个热点, 从地下根系入手来探讨混生群落植物种间关系及其对环境胁迫响应的研究少见报道。该文以荒漠草原区(灵武)、典型荒漠区(张掖)和极端荒漠区(酒泉) 3个不同生境条件下单生与混生红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)为实验材料, 采用分层取样法对其垂直根系参数进行测定和分析, 探讨了两种植物根系分布对混生及荒漠环境梯度的响应。结果表明: 同一生境条件下, 混生红砂和珍珠猪毛菜比根长和比表面积均高于单生, 说明红砂、珍珠猪毛菜混生后其根系相互作用关系表现为互惠, 促进了植株对土壤养分和水分的吸收。不同生境条件下, 同一生长方式的红砂根系分布深度均大于珍珠猪毛菜, 且根系消弱系数也普遍高于珍珠猪毛菜, 说明二者在不同生境条件下占据不同生态位, 红砂表现为深根性, 根系位于土壤深层, 珍珠猪毛菜表现为浅根性, 根系分布于土壤浅层。随着荒漠环境胁迫增强, 单生和混生红砂与珍珠猪毛菜的比根长和比表面积均呈现出极端荒漠区>典型荒漠区>草原荒漠区的规律, 且生境越干旱, 混生群落根系分离越明显; 单生与混生红砂根系消弱系数也逐渐增大, 在极端干旱区达到最大值, 珍珠猪毛菜变化不大, 表明红砂-珍珠猪毛菜混生群落根系生态位分离随荒漠环境胁迫增强而加大, 验证了环境胁迫梯度假说。可见“地上聚生, 地下分离”的混生方式可能是红砂-珍珠猪毛菜混生群落适应干旱胁迫环境的生长策略。
关键词:
Abstract
Keywords:
PDF (1187KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
单立山, 苏铭, 张正中, 王洋, 王珊, 李毅. 不同生境下荒漠植物红砂-珍珠猪毛菜混生根系的垂直分布规律. 植物生态学报[J], 2018, 42(4): 475-486 DOI:10.17521/cjpe.2017.0300
SHAN Li-Shan, SU Ming, ZHANG Zheng-Zhong, WANG Yang, WANG Shan, LI Yi.
物种共存和多样性形成机制一直是生态学研究的核心问题(Debra et al., 2014), 植物间的正负相互作用对于物种分布(Xue et al., 2017)、植物生理生态特性(Su et al., 2012)、种群动态(Grime, 1979)、群落结构(Tilman, 1988)以及生态系统功能有重要影响(Callaway, 2007)。经典的生态位理论认为, 物种在某种(或多种)环境资源利用上存在差异, 即物种之间的生态位分化是物种共存的基本机制(Chesson, 2000)。其中, 相邻同种和异种个体根系竞争形成的生态位分离是物种共存和生物多样性维持的一条重要途径(February et al., 2011)。由于地下部分研究的困难, 人们对根系相互作用的关系了解甚少(Copely, 2000)。已有研究指出, 植物种间关系主要靠地下根系与土壤之间的物理、化学和生物过程表现出的竞争关系, 来实现对水资源进行合理分配利用(王政权等, 2003), 在判定竞争平衡、竞争强度和资源利用方面, 地下部分的竞争作用明显重要于地上部分, 从地下根系形态和分布特征来认识物种间竞争和互利共生现象, 才能了解种间关系的本质(陈伟和薛立, 2004)。已有研究表明, 植物会因相邻植物(同种或异种)的竞争来调整根系的空间分布格局(Hodge, 2004); 且这种变化在非生物胁迫环境(干旱/半干旱地区、高寒地区和盐沼地带)中更容易观测到(张炜平等, 2013)。在自然条件下, 植物同时与环境以及竞争者的根系产生联系, 但是我们不知道植物是如何整合与这些因子有关的信号, 并且如何做出响应(罗维成等, 2012)。 因此, 从地下根系入手, 开展植物种间相互作用及其对环境胁迫梯度响应的研究, 对胁迫环境下植被的恢复有重要指导意义, 同时对农林系统中混交林、纯林以及林木-草-农作物系统的构建具有实践意义。
植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(王政权和张彦东, 2000; Meyer et al., 2007; 王鹏等, 2012)。多数关于植物种间地下根系分布的研究表明, 为实现物种共存, 植物通过根系的生态位分离以避免或减少竞争(Holdo & Timberlake, 2008; Hajek et al., 2014)。早在20世纪30年代, Walter (1939)就对热带亚热带稀树草原中乔草共存机制进行了研究, 率先提出根系“双层分布”模型。林木混交因其根竞争也会诱发根系空间分布变化, 使林木在土壤水分和养分利用上存在差异(Leuschner et al., 2001; Curt & Prévosto, 2003; Bolte & Villanueva, 2006), 而发生根系生态位分离现象。然而, 植物根系生态位分离不仅与竞争者有关, 而且与环境的异质性关系紧密(Cahill, 2010)。Bertness和Callaway (1994)提出了环境胁迫梯度假说, 认为植物种间互利的强度或重要性随着环境胁迫度的增加而增加。Callaway等(2002)研究发现随着海拔的升高, 植物间互惠作用增强, 植物间邻体作用有利于其生态位的扩充, 也就是说, 正相互作用更多发生在较寒冷的环境中, 而竞争更多发生在较温暖的环境中。O’Connor和Roux (1995)也发现, 在草-灌共存的干旱生态系统中, 随着水分胁迫程度的增加, 植物可通过地下根系形态调整来减少种间竞争实现互惠。Schenk和Jackson (2002)、Padilla和Pugnaire (2007)研究指出, 随干旱胁迫进一步加剧, 在草本植物和豆科灌木混生的群落中, 豆科灌木通过根系深扎和根系进一步扩展形成地下根系生态位分离, 减少种间根系对土壤水分竞争。以上研究均表明, 随着干旱胁迫的增加, 根系竞争作用减
弱, 而促进作用相应增强, 当干旱胁迫增加到一定程度时, 根间的关系便会从竞争转变为促进作用。然而, 目前大多数对环境胁迫梯度假说的研究多集中在混交林、草-灌、草-草等植物群落, 而对荒漠区珍贵的木本植物混生群落, 特别是其地下根系相互作用关系及其对荒漠环境梯度的响应研究较缺乏。因此, 开展荒漠环境梯度下荒漠木本植物群落地下根系的分配格局及其相互关系的研究, 对揭示荒漠区物种共存和生物多样性维持具有重要意义。
红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是干旱荒漠和荒漠草原区分布最广的植物种, 两者常互为共建种组成混生群落, 混生群落适应极端环境的能力要高于单生群落, 这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用(Su et al., 2012)。在自然分布的混生群落中, 有时红砂和珍珠猪毛菜各自单独生长, 称之为单生, 有时它们紧密生长在一起, 称之为混生(Su et al., 2012)。近年来, 人工控制实验条件下红砂和珍珠猪毛菜混生后两者的生理变化特征正逐渐受到人们的重视(周海燕等, 2012), 就水分胁迫下红砂和珍珠猪毛菜混生后其垂直根系变化也有一些零星报道(严巧娣等, 2012)。然而, 红砂和珍珠猪毛菜组成的混生群落广泛分布于我国荒漠东部和荒漠草原区, 其生境类型多样, 由东向西其气候形成光-温-湿的明显梯度和多种多样的水热组合, 在我国形成了沿荒漠环境梯度从荒漠草原区、典型荒漠区到极端荒漠区的分布格局。这种沿荒漠环境梯度分布的木本植物自然混生群落是珍贵的植物混生生长类型, 为研究植物种间的相互作用关系提供了很好的研究对象和场所, 也为验证环境胁迫梯度假说提供了很好的实验材料。为此, 本研究按自然降雨梯度选取了单生和混生的红砂与珍珠猪毛菜为实验材料, 比较了其单生和混生时地下垂直根系分布差异, 试图回答以下几个问题: 1)同一生境条件下, 两种荒漠植物在单生和混生时其垂直根系分布规律是否有差异? 2)荒漠环境梯度下, 单生和混生红砂或珍珠猪毛菜其垂直根系分布格局如何变化, 是否支持环境胁迫梯度假说? 回答这些问题对深化理解环境胁迫驱动的植物相互作用负/正的转变和完善该理论具有重要的学术价值, 对指导胁迫环境下的植被恢复具有重要的意义。
1 材料和方法
1.1 研究区概况
在灵武、张掖、酒泉红砂-珍珠猪毛菜自然混生群落中设定采样点。其中, 灵武(38.10° N, 106.34° E)为荒漠草原区。气候干燥, 雨量少而集中, 蒸发强烈, 冬冷夏热, 气温日较差大; 日照较长, 光能丰富; 无霜期较短, 冬春季风沙天气较多。其地带性植被为沙生针茅(Stipa glareosa)、野蓟(Cirsium maackii)、甘草(Glycyrrhiza uralensis)、沙蒿(Artemisia sphaerocephala)等荒漠植被。张掖(38.55° N, 100.27° E)为典型荒漠区, 夏季炎热, 干旱少雨; 日照时间长, 光热资源充足, 冬寒夏暖, 四季分明, 降雨集中于夏季。其地带性植被为红砂、珍珠猪毛菜、甘草等荒漠植被。酒泉(39.53° N, 98.47° E)为极端荒漠区, 降水量少, 蒸发强烈, 日照时间长, 昼夜温差大, 冬季寒冷, 夏季炎热, 春季多风。其地带性植被有红砂、珍珠猪毛菜、盐爪爪(Kalidium foliatum)、合头草(Sysmpegma regelly)等。不同研究区的水热状况见表1。Table 1
表1
表1研究区水热条件比较
Table 1
| 采样点 Sample site | 平均海拔 Mean elevation (m) | 年平均气温 Mean annual temperature (°C) | 年降水量 Mean annual precipitation (mm) | 年蒸发量 Mean annual evaporation (mm) | 年日照时间 Mean annual sunshine times (h) |
|---|---|---|---|---|---|
| 灵武 Lingwu | 1 278 | 8.9 | 213 | 1 762 | 3 011 |
| 张掖 Zhangye | 1 590 | 6.0 | 110 | 2 003 | 3 075 |
| 酒泉 Jiuquan | 1 500 | 7.5 | 58 | 2 038 | 3 228 |
新窗口打开|下载CSV
1.2 试验方法
本研究于植物生长季末2016年9月下旬至10月上旬开展。在灵武、张掖、酒泉3个红砂和珍珠猪毛菜混生群落自然分布区设定样地, 在各样地分别选取有代表性、大小均一的单生红砂与单生珍珠猪毛菜以及混生红砂和珍珠猪毛菜, 重复数为3。选择植株时, 尽量使所选取的灌丛微环境相同, 以减少环境不同而造成的实验误差。所选植物周围1 m的范围内无其他植物生长, 避免来自周围植物的种间或种内作用的影响。采用分层取样法(高杰, 2014; 孙文泰等, 2016)挖取根系, 具体方法为: 在离植株根茎为1 m处挖壕沟(长×宽×深= 50 cm × 70 cm × 120 cm), 沿沟壁将根系(主根与侧根)慢慢从上到下按10 cm一层分层挖出, 直至把根挖完, 用土筛将根系筛出, 然后分别装于不同的信封标记, 带回实验室。将这些根样用水洗净杂质和泥沙, 用Epson扫描仪进行扫描, 然后用Win-RhIZO根系分析软件分析出根系垂直分布指标, 如根长、根表面积。将扫描过的根系重新装入信封, 105 ℃杀青2 h后, 在65 ℃恒温烘箱中烘48 h至恒质量, 用天平称量得到红砂、珍珠猪毛菜单生和混生地下各层生物量(马晓东等, 2012)。根据Win-RhIZO根系分析软件测定的根系形态指标以及根系生物量(RB), 计算出两种荒漠植物根系的比根长(SRL)、比表面积(SRA)等参数。计算公式如下:
比根长(cm·g-1) =根长/生物量
比表面积(cm2·g-1) =表面积/生物量
利用Gale和Grigal (1987)植物根系垂直分布的非线性模型计算了单生和混生条件下红砂和珍珠猪毛菜的根系消弱系数β, 具体计算公式如下:
$\Upsilon =\text{1}-{{\beta }^{d}}$
式中, Y代表从土壤表面至某土层深度d (cm)的根系生物量的累积百分比, β为根系消弱系数。其中, β值越大(越接近1), 表明植物根系在较深土壤层次中分布的比例越多; 反之, 则表示植物根系在浅层土壤中分布的比例越多。
1.3 数据统计分析
所有的数据采用SPSS 22.0进行统计分析, 对同一生境不同生长方式下同种植物根系生态指标进行了独立样本t检验, 对同一生长方式不同生境条件下同种植物根系形态指标进行了单因素方差分析; 用一般线性模型对环境梯度和生长方式的交互效应进行双因素方差分析; 使用Matlab拟合红砂和珍珠猪毛菜根系从地表至d cm深度的根系累积消弱曲线。做图采用Microsoft Excel 2010, 图中数值均为平均值±标准误差。2 结果和分析
2.1 不同生境下单生和混生红砂与珍珠猪毛菜根系比根长(SRL)的垂直分布特征
从表2可以看出, 环境梯度和生长方式及两者的交互作用对SRL均无显著影响(p > 0.05)。然而, 各生境条件下, 单生或混生珍珠猪毛菜同层SRL均大于单生或混生红砂(图1)。同一生境下, 单生和混生红砂SRL随土壤深度加深均呈现出先增加后减少的趋势, 其根系垂直分布深度相同; 而单生和混生珍珠猪毛菜SRL随土壤深度加深呈现出增加的变化趋势, 单生珍珠猪毛菜的根系垂直分布深度比混生珍珠猪毛菜少10-20 cm。与单生相比, 混生红砂和珍珠猪毛菜各层SRL均有较大增幅, 最大可达89.65%和89.92%, 其中红砂在灵武的40-50 cm和张掖的0-10 cm土壤深度差异性达到显著水平(p < 0.05), 珍珠猪毛菜在酒泉的0-10 cm土壤深度差异性达到极显著水平(p < 0.001)。从图1还可看出, 随着荒漠环境梯度的增强, 单生和混生红砂与珍珠猪毛菜SRL大小排序均为: 酒泉>张掖>灵武; 在灵武与张掖、张掖与酒泉比较中发现, 同一生长方式红砂SRL于0-10 cm土壤深度的差异分别达到显著水平, 单生珍珠猪毛菜SRL则在灵武和酒泉地区10-20土壤深度的差异达到显著水平(p < 0.05)。图1
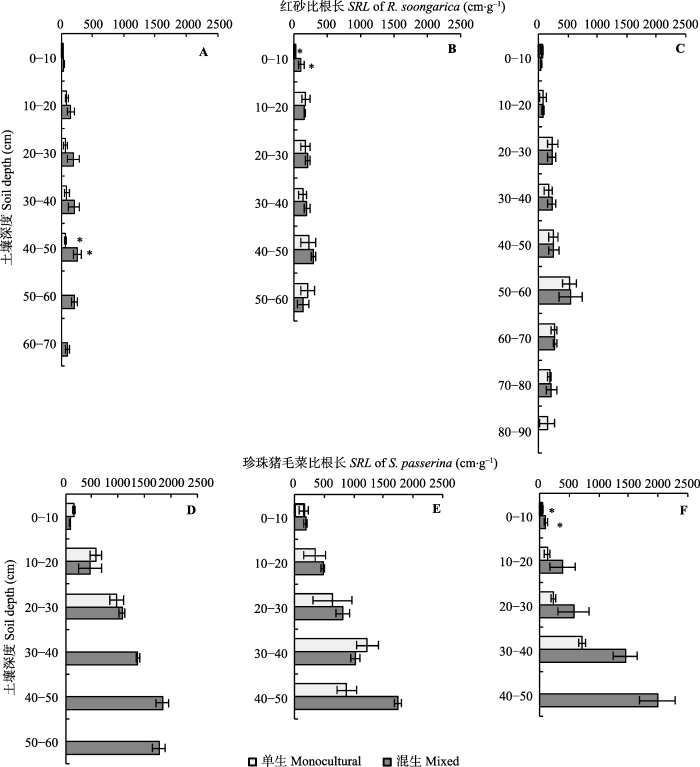 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同生境下单生和混生的红砂与珍珠猪毛菜根系比根长的垂直分布特征(平均值±标准误差)。A, 灵武红砂。B, 张掖红砂。C, 酒泉红砂。D, 灵武珍珠猪毛菜。E, 张掖珍珠猪毛菜。F, 酒泉珍珠猪毛菜。*表示同一生境下同层单生和混生红砂或珍珠猪毛菜的差异显著(p < 0.05)。
Fig. 1The vertical distributions of specific root length (SRL) in monocultural and mixed communities of Reaumuria soongarica and Salsola passerina in different habitats (mean ± SE). A, R. soongarica in Lingwu. B, R. soongarica in Zhangye. C, R. soongarica in Jiuquan. D, S. passerina in Lingwu. E, S. passerina in Zhangye. F, S. passerina in Jiuquan. * indicates the significant difference between monocultural and mixed communities of R. soongarica and S. passerina within the same habitat (p < 0.05).
Table 2
表2
表2环境胁迫梯度和生长方式对红砂和珍珠猪毛菜比根长(SRL)、比表面积(SRA)、根系生物量(RB)和根系消弱系数(β)影响的双因素方差分析(F值)
Table 2
| 变异来源 Source of variation | 红砂 R. soongarica | 珍珠猪毛菜 S. passerina | ||||||
|---|---|---|---|---|---|---|---|---|
| SRL | SRA | RB | β | SRL | SRA | RB | β | |
| 环境梯度 Environment gradient | 0.444 | 1.237 | 121.887** | 2.104 | 1.554 | 1.363 | 28.556** | 0.844 |
| 生长方式 Community type | 0.224 | 1.212 | 84.367** | 2.194 | 0.284 | 1.083 | 106.824** | 3.265 |
| 环境梯度×生长方式 Environment gradient × community type | 2.797 | 0.080 | 14.944** | 2.303 | 1.978 | 0.088 | 18.258** | 1.398 |
新窗口打开|下载CSV
2.2 不同生境下单生和混生红砂与珍珠猪毛菜根比表面积(SRA)的垂直分布特征
从表2可以看出, 环境梯度和生长方式及两者的交互作用对SRA均无显著影响(p > 0.05)。然而, 不同生境条件下, 单生或混生珍珠猪毛菜同层SRA均大于单生或混生红砂(图2)。相同生境下, 单生和混生红砂SRA基本随土壤深度增加呈现出先增加后减少, 珍珠猪毛菜则随土壤深度增加呈现出增加的趋势。与单生相比, 混生红砂和珍珠猪毛菜各层SRA均有较大增幅, 最大可达78.70%和86.76%, 其中, 红砂在灵武的50-60 cm土壤深度差异性达到极显著水平(p < 0.001), 而珍珠猪毛菜在张掖和酒泉的0- 10 cm土壤深度差异性达到显著水平(p < 0.05)。图2还显示, 随着环境胁迫梯度的增强, 单生和混生红砂和珍珠猪毛菜SRA基本呈现出灵武<张掖<酒泉的趋势。相同生长方式不同生境条件下, 在0-10 cm和20-30 cm土壤深度红砂SRA差异达到极显著水平 (p < 0.001), 单生珍珠猪毛菜在不同生境条件下10- 20 cm土壤深度的SRA差异达到显著水平(p < 0.05)。
图2
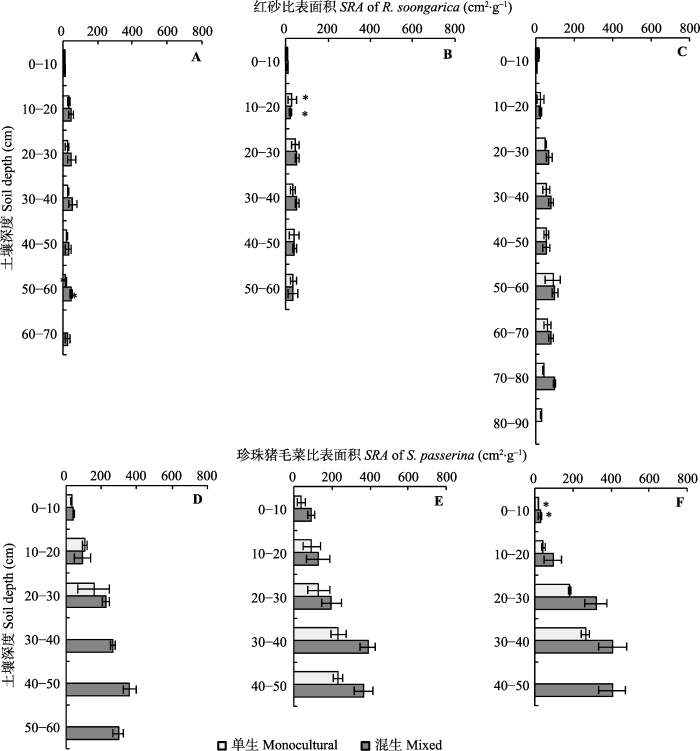 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同生境下单生和混生的红砂与珍珠猪毛菜根系比表面积的垂直分布特征(平均值±标准误差)。
Fig. 2The vertical distributions of specific root surface area (SRA) in monocultural and mixed communities of Reaumuria soongarica and Salsola passerina in different habitats (mean ± SE).
2.3 不同生境下单生和混生红砂与珍珠猪毛菜根系生物量垂直分布特征
从表2可以看出, 环境梯度和生长方式及两者的交互作用对两种荒漠植物根系生物量均有极显著的影响(p < 0.001)。不同生境条件下, 单生或混生红砂和珍珠猪毛菜根系生物量均随着土壤深度增加呈现出递减的趋势。同一生境条件下, 混生红砂根系生物量大于同层单生红砂, 珍珠猪毛菜则刚好相反; 其中, 红砂在张掖的0-10 cm和酒泉的10-20 cm土壤深度差异性达到显著水平(p < 0.05), 珍珠猪毛菜在酒泉的0-10 cm和30-40 cm土壤深度差异性达到极显著水平(p < 0.01)。图3还显示, 同一生长方式下, 极端荒漠区(酒泉)两种荒漠植物同层根系生物量均大于典型荒漠区(张掖)和半干旱区(灵武)。不同生境条件下, 在0-20 cm土壤深度, 单生与混生红砂根系生物量差异达到极显著水平(p < 0.01); 而单生与混生珍珠猪毛菜在0-10 cm和30-40 cm达到极显著水平(p < 0.01)。同时, 单生和混生红砂根系垂直分布深度随着环境胁迫的加剧而增加, 酒泉根深可达90 cm, 而单生和混生的珍珠猪毛菜根系垂直分布深度基本维持在30-50 cm。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同生境下单生和混生的红砂与珍珠猪毛菜根系生物量的垂直分布特征(平均值±标准误差)。
Fig. 3The vertical distributions of biomass between in monocultural and mixed communities of Reaumuria soongarica and Salsola passerina in different habitats (mean ± SE).
2.4 不同生境下单生和混生红砂与珍珠猪毛菜根系消弱系数的比较
双因子方差分析表明, 环境梯度和生长方式及两者的交互作用对根系消弱系数均无显著影响(p > 0.05)(表2)。从表3可以看出, 同一生境条件下, 单生或混生红砂根系削弱系数均大于单生或混生珍珠猪毛菜; 不同生境条件下两种荒漠植物混生后其根系消弱系数均大于或等于单生, 其中, 在灵武地区混生珍珠猪毛菜根系消弱系数显著大于单生珍珠猪毛菜(p < 0.01)。在相同生长方式下, 红砂根系消弱系数随着环境胁迫梯度增加呈现上升变化趋势, 在极端干旱区酒泉达到最大值; 不同生境条件下珍珠猪毛菜根系消弱系数无明显变化规律, 各生境条件下差异不大。Table 3
表3
表3不同生境下单生和混生红砂与珍珠猪毛菜的根系消弱系数(β)与决定系数(R2)(平均值±标准误差)
Table 3
| 物种 Species | 生长方式 Interspecific relationship | 灵武 Lingwu | 张掖 Zhangye | 酒泉 Jiuquan | |||
|---|---|---|---|---|---|---|---|
| β | R2 | β | R2 | β | R2 | ||
| 红砂 R. soongarica | 单生 Monocultural | 0.934β9 ± 0.002β4 | 0.981β8 | 0.941β2 ± 0.007β8 | 0.929β1 | 0.942β4 ± 0.011β2 | 0.936β7 |
| 混生 Mixed | 0.934β9 ± 0.011β2 | 0.962β0 | 0.942β1 ± 0.025β6 | 0.897β1 | 0.943β7 ± 0.004β5 | 0.968β6 | |
| 珍珠猪毛菜 S. passerina | 单生 Monocultural | 0.875β8 ± 0.007β8a | 0.949β9 | 0.921β7 ± 0.026β1 | 0.947β6 | 0.896β5 ± 0.004β9 | 0.902β3 |
| 混生 Mixed | 0.925β3 ± 0.006β0b | 0.986β1 | 0.919β3 ± 0.025β1 | 0.870β8 | 0.918β1 ± 0.003β9 | 0.923β4 | |
新窗口打开|下载CSV
3 讨论
3.1 混生对红砂和珍珠猪毛菜根系形态特征的影响
一般认为, 植物可通过根系形态结构的变化实现对资源的竞争(班芷桦和王琼, 2015)。SRL和SRA作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(Bauhus et al., 2000)。已有研究表明, 相邻植物可通过增加SRL和SRA来适应邻体根竞争(Curt & Prévosto, 2003; Fujii & Kasuya, 2008; Gao et al., 2010)。本研究发现, 同一生境条件下, 混生红砂和珍珠猪毛菜SRL和SRA均远远大于单生红砂和珍珠猪毛菜, 说明混生能促进两种植物SRL和SRA的增加, 以此提高对资源的获取能力, 以适应种间竞争。这与Zhang等(2014b)的张掖龙首山山前戈壁红砂-珍珠猪毛菜混生促进了两种荒漠植物细根SRL显著增加的结果一致; 也与王政权和张彦东(2000)对水曲柳(Fraxinus mandshurica)和落叶松(Larix gmelinii)混栽导致水曲柳根系密度和根长度的增加, Bolte和Villanueva (2006)对欧洲山毛榉(Fagus sylvatica)和挪威云杉(Picea abies)混交改变欧洲山毛榉根系形态, 提高其细根的SRL和SRA结果一致。Gao等(2010)和Zhang等(2013)在农作物间作研究中也发现了相似的研究结果, 间作显著增加作物的SRL、SRA、根长密度。表明相邻同种或异种植物可通过增加SRL和SRA促进根系对土壤水分和养分的吸收以适应竞争环境。然而, Beyer等(2013)和Hajek等(2014)基于根序理论发现植物种内和种间根竞争对细根形态影响不显著。由此可见, 由植物种间竞争引起的根系形态变化并不是一个普遍现象。本研究还发现, 不同生境条件下, 同一生长方式珍珠猪毛菜的SRL和SRA均大于红砂, 这可能是由于珍珠猪毛菜为非深根型植物, 其主要水源为大气降水(许皓和李彦, 2005), 因强烈蒸发, 其水分来源少, 为适应西北干旱环境, 通过增加根系次级分支来增大比根长和比表面积, 以获取维持水源适应干旱胁迫环境; 而红砂为深根型植物, 其主要维持水源为地下水, 主根发达根系分支少(单立山等, 2012), 其SRL和SRA小。
3.2 混生对红砂和珍珠猪毛菜根系分布的影响
根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(陈伟和薛立, 2004)。其中, 植物的根竞争对根系最为明显的效果就是影响根系的空间分布。大量研究表明, 植物种间根竞争导致根系空间分布发生变化而发生生态位分离现象(Holdo & Timberlake, 2008; Hajek et al., 2014)。本研究发现, 各生境条件下, 混生红砂根系分布深度均大于混生珍珠猪毛菜, 说明红砂比珍珠猪毛菜分布于更深的土壤深度, 珍珠猪毛菜则是在较浅的土壤深度分布, 这与杨昊天等(2013)的研究结果类似, 即珍珠猪毛菜主要分布在0-50 cm土层, 而红砂主要分布于0-70 cm土层。Bolte和Villanueva (2006)研究发现欧洲山毛榉和挪威云杉混交促进欧洲山毛榉根系深扎, 避免了根系间的过度竞争, 为充分利用不同层次土壤中的水分和养分奠定了基础。同样, 在农作物间作(Zhang et al., 2013, 2014a)、农林复合系统(Dawson et al., 2001)、混生草本(Fargione & Tilman, 2005; Mommer et al., 2011)、草灌混生群落(February et al., 2011)和入侵物种之间(Daneshgar & Jose, 2009)也发现了根系生态位分离现象, 表明植物根系的分布会在相邻植物根系格局的影响下做出相应调整(Cahill, 2010; 陈斌等, 2017), 以减少对资源的竞争(Schmid et al., 2015)。红砂-珍珠猪毛菜混生根系分离现象也说明两种植物虽地上部分以聚集丛生的混生群落生长, 而其地下根系垂直分布则存在分层现象, 即以空间生态位分离的方式减少种间对土壤资源的竞争(晨乐木格等, 2009), 实现物种共存, 这也是维持植物多样性的一种重要机制。为了描述根系的垂直分布规律, Gale和Grigal (1987)通过不同演替阶段不同树种根系分布特征提出了植物根系垂直分布非线性模型, 该模型为研究土壤垂直梯度中植物根系的总体分布提供了简洁标准的办法(Zhang et al., 2014)。目前, 该方法主要运用于比较不同群落根系垂直分布的差异性, 以及分析植物根系垂直分布特征对非生物环境的响应(陈斌等, 2017)。陈斌等(2017)研究发现, 在不同微生境分区内芨芨草(Achnatherum splendens)与白刺(Nitraria tangutorum)的根系消弱系数差异具有显著性, 说明两种植物根系的地下垂直分布在总体上存在分层现象。李鹏等(2005)研究表明, 在渭北黄土区的刺槐(Robinia pseudoacacia), 其细根根系消弱系数在阳坡要小于阴坡, 说明阳坡的生长环境对树木吸收深层土壤水分和养分有一定的限制作用。本研究发现, 同一生境条件下, 单生或混生红砂根系消弱系数大于单生或混生珍珠猪毛菜, 且同一生境条件下, 混生红砂和珍珠猪毛菜根系消弱系数均大于单生红砂和珍珠猪毛菜, 在灵武地区混生珍珠猪毛菜根系消弱系数显著大于单生珍珠猪毛菜, 说明两种植物混生后可以增大根系在深层土壤中分布的百分比。
3.3 环境胁迫梯度对根系相互作用的影响
植物间的互作类型及互作强度与其所处的环境条件之间存在着密切的关联。胁迫梯度假说认为, 当环境条件较好时, 物种间的负相互作用占据主导地位; 而在恶劣的环境条件下, 正相互作用的重要性上升, 并且其作用强度随着环境的恶劣程度而增加(Bertness & Callaway, 1994)。本研究发现, 随着荒漠环境胁迫梯度增强, 单生和混生红砂与珍珠猪毛菜SRL与SRA呈现出增加的趋势, 不同生境间的差异接近或达到显著水平, 表明两种荒漠植物可通过增大SRL和SRA获取更多维持水源适应干旱胁迫环境。两种荒漠植物的地下根系占有不同的生态位, 并随环境胁迫梯度增加表现更明显, 红砂根系在极端荒漠区可至地下90 cm, 而珍珠猪毛菜根系则是维持在地下30-50 cm的深度, 这与He等(2012)及He和Bertness (2014)的研究结果一致, 即随着环境胁迫度的增加物种间互利的强度或重要性加强, 从而支持了环境胁迫梯度假说。本研究也发现, 随着生境干旱胁迫程度的增强, 相同生长方式下, 红砂根系消弱系数呈上升趋势, 在酒泉到最大值, 而珍珠猪毛菜根系消弱系数无明显变化规律, 各生境条件下差异不大, 说明干旱胁迫的增强进一步促进了两种植物在地下的根系生态位分离现象。这与Zhang等(2014b)关于不同水分条件下混生红砂、珍珠猪毛菜根系对环境的适应策略相一致, 也与前面得出的结论一致, 进一步验证了Bertness和Callaway(1994)的环境胁迫梯度假说在我国西北荒漠地区适用。4 结论
本研究发现, 相同生境条件下, 混生红砂和珍珠猪毛菜SRL和SRA均高于单生, 表明红砂、珍珠猪毛菜混生时两者表现为互惠关系, 促进了根系对土壤养分和水分的吸收能力。不同生境条件下, 单生和混生红砂根系分布深度均大于珍珠猪毛菜, 且根系消弱系数也普遍高于珍珠猪毛菜, 说明两种植物虽然地上部分以聚集丛生的混生群落生长, 但地下根系形成了分层现象, 即生态位分离, 这是其共存的重要机制。随着荒漠胁迫环境梯度增加, 单生和混生红砂与珍珠猪毛菜SRL与SRA均呈现出增加的趋势, 且混生红砂和珍珠猪毛菜根系分离更明显; 单生与混生红砂根系消弱系数也逐渐增大, 在极端干旱区达到最大值, 珍珠猪毛菜变化不大, 表明红砂-珍珠猪毛菜混生群落的垂直根系发生生态位分离现象, 并随着干旱胁迫增强而加剧, 支持环境胁迫梯度假说。可见, 在西北干旱区红砂-珍珠猪毛菜混生群落地上部分的“成丛性”分布, 即通过聚集分布以降低蒸腾失水等生理胁迫; 而地下部分通过根系分布变化呈现出空间生态位分离, 即通过根系错位来增强对土壤不同深度的养分吸收, 减少种间竞争。这种“地上聚生、地下分离”的空间格局是其适应干旱胁迫环境的重要策略, 对揭示物种多样性及植物共存机制具有重要的意义。致谢 甘肃农业大学青年导师基金(GAU-QNDS- 201605)资助。

扫码加入读者圈
听语音, 看问答
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]

ABSTRACT This study investigated whether increased productivity in mixed plantations of Eucalyptus globulus Labill. and Acacia mearnsii de Wild when compared with monocultures could be explained by niche separation of the fine-root systems. For this purpose fine-root architecture, nutrient concentration, and fine-root distribution were examined in two horizons (0-15 and 15-30 cm) of mixed and pure stands. Investigations were carried out in 6.5-year-old plantations consisting of 100% eucalypts, 75% eucalypts + 25% acacia, 50% eucalypts + 50% acacia, 25% eucalypts + 75% acacia, and 100% acacia. Aboveground the two species interacted synergistically. Stem volume and tree height was highest in the 50:50 mixtures. For acacias, intraspecific competition was stronger than interspecific competition with eucalypts. Fine-root biomass and length density were similar for all species combinations, and there was no synergistic effect. The vertical distribution of fine roots and fine-root architecture were similar for acacias and eucalypts. This indicated that soil exploitation strategies may be similar, which can result in strong competition for soil resources. Fine-root nitrogen concentrations of eucalypts were highest in the 50:50 mixture. Improved productivity of mixtures appears to be a result of both canopy stratification and improved N nutrition of eucalypts through N fixation by acacias.
DOIURL [本文引用: 1]
DOIURL
The mechanisms of belowground competition are not well understood. Addressing literature reports on competition-induced changes in tree fine root morphology, we conducted a growth experiment with tree saplings to investigate competition effects on important root morphological and functional traits in a root order-focused analysis.European beech and European ash saplings were grown for 34 months in containers under greenhouse conditions in monoculture (2 conspecific plants), in mixture (1 beech and 1 ash) or as single plants. The root system was fractionated according to root orders and eight morphological and functional properties were determined.Root order was the most influential factor affecting the fine root traits (except for root diameter and delta C-13); a significant species identity effect was found for root diameter, tissue density, N concentration and delta C-13. Ash fine roots were thicker, but had lower tissue densities, contained more N and had systematically higher delta C-13 values than beech roots. The competition treatments had no significant effect on morphological root traits but altered delta C-13 in the 2nd root order.Neither intra- nor interspecific root competition affected fine root morphology significantly suggesting that competition-induced root modification may not be a universal phenomenon in temperate trees.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
First page of article
[本文引用: 1]
DOIURL
DOIURL [本文引用: 2]
本文以宁夏沙湖一个典型的干旱区灌草型白刺(Nitraria tangutorum)—芨芨草(Achnatherum splendens)群落为研究对象,将该群落自植物聚生丛冠层覆盖区至丛间裸地依次划分为冠盖区、过渡区和空旷区3个微生境分区,通过比较两优势种的根系生物量密度以及根系消弱系数(反映根系总体埋深)在这3个分区中的分布特征,研究了干旱胁迫环境下两物种的根系分布与共存机制。结果表明:两物种的总根与细根均集中分布于冠盖区,但芨芨草根系生物量密度自冠盖区至空旷区的下降幅度更大;白刺总根及细根生物量密度重心自冠盖区至空旷区逐渐下移,而芨芨草的总根和细根生物量密度重心在空旷区较过渡区有所抬升;另外,白刺细根生物量密度在冠盖区10~30 cm土层中最高,而芨芨草在0~10cm层最高;各微生境分区内两物种的根系消弱系数均具有显著性(或接近于显著性)差异。这暗示了白刺与芨芨草的根系分布存在明显的生态位分离现象。“地上聚生,地下分离”的共生方式可能是白刺与芨芨草共同适应干旱环境的生长策略。
DOIURL [本文引用: 2]
本文以宁夏沙湖一个典型的干旱区灌草型白刺(Nitraria tangutorum)—芨芨草(Achnatherum splendens)群落为研究对象,将该群落自植物聚生丛冠层覆盖区至丛间裸地依次划分为冠盖区、过渡区和空旷区3个微生境分区,通过比较两优势种的根系生物量密度以及根系消弱系数(反映根系总体埋深)在这3个分区中的分布特征,研究了干旱胁迫环境下两物种的根系分布与共存机制。结果表明:两物种的总根与细根均集中分布于冠盖区,但芨芨草根系生物量密度自冠盖区至空旷区的下降幅度更大;白刺总根及细根生物量密度重心自冠盖区至空旷区逐渐下移,而芨芨草的总根和细根生物量密度重心在空旷区较过渡区有所抬升;另外,白刺细根生物量密度在冠盖区10~30 cm土层中最高,而芨芨草在0~10cm层最高;各微生境分区内两物种的根系消弱系数均具有显著性(或接近于显著性)差异。这暗示了白刺与芨芨草的根系分布存在明显的生态位分离现象。“地上聚生,地下分离”的共生方式可能是白刺与芨芨草共同适应干旱环境的生长策略。
DOIURL [本文引用: 2]
植物根系间的相互作用分为竞争和互利两种形式,它是决定植物群落动态变化和群落结构的重要因素.根系间的竞争包括植株个体自身根系的竞争以及个体与个体根系间(同种或异种)的竞争两方面,前者的发生在农林系统中是不可避免的并且很难调控,后者可以分为种间植物根系的竞争和种内植物根系的竞争.还阐述了根系的竞争能力和与其密切相关的根系生长率、根组织的新陈代谢、植物的生长形式和根系的空间结构等植物特性,同时介绍了根系对水和养分的竞争机理、形式、影响以及竞争强度计算方法.接着具体分析包括根系错位在内的各种根系互利现象和相关机理.影响根系间相互作用的限制性因子有土壤营养的异质性、大气CO2浓度、地下草食生物、根系生产力和生物量、根系结构、形态和生理调节、土壤养分的扩散性以及植物间距等.随着科技的进步和各门学科的发展,未来根系的研究方向主要体现在结合实践优化农林系统中不同物种间的作用关系、预测根系竞争在全球气候变化下的发展规律、更新实验研究方法及手段研究作用机理等3个方面.
DOIURL [本文引用: 2]
植物根系间的相互作用分为竞争和互利两种形式,它是决定植物群落动态变化和群落结构的重要因素.根系间的竞争包括植株个体自身根系的竞争以及个体与个体根系间(同种或异种)的竞争两方面,前者的发生在农林系统中是不可避免的并且很难调控,后者可以分为种间植物根系的竞争和种内植物根系的竞争.还阐述了根系的竞争能力和与其密切相关的根系生长率、根组织的新陈代谢、植物的生长形式和根系的空间结构等植物特性,同时介绍了根系对水和养分的竞争机理、形式、影响以及竞争强度计算方法.接着具体分析包括根系错位在内的各种根系互利现象和相关机理.影响根系间相互作用的限制性因子有土壤营养的异质性、大气CO2浓度、地下草食生物、根系生产力和生物量、根系结构、形态和生理调节、土壤养分的扩散性以及植物间距等.随着科技的进步和各门学科的发展,未来根系的研究方向主要体现在结合实践优化农林系统中不同物种间的作用关系、预测根系竞争在全球气候变化下的发展规律、更新实验研究方法及手段研究作用机理等3个方面.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
This study investigated the belowground development and strategy of late-successional European beech ( Fagus sylvatica L.) in ageing natural Scots pine ( Pinus sylvestris L.) and Silver birch ( Betula pendula Roth.) woodlands in a French volcanic mid-elevation area. For this purpose root biomass, root profile and fine-root architecture of competitor trees were examined in 53 mixed pine eech and 42 birch eech woodlands along a stand maturation gradient, using the root auger technique (0.75-cm). The total beech fine-root biomass highly correlated with aerial dimensions such as stem height and girth, whereas it moderately correlated with its age, thus indicating the effects of competition. Basic stand biometric data such as stand density and basal area had no significant effect on beech root biomass. Conversely, competition indices taking into account the vertical dimensions of competitor trees were efficient, probably due to redundancy with beech height. At similar age and height, beeches under birch had a greater belowground development than beeches under pine. Each species exhibited specific rooting pattern and plasticity of fine-root architecture along the gradients of stand maturation and competition. Beech had a heart-shaped rooting habit in both mixings, which strongly increased along stand maturation. Its fine-root system adopted a foraging strategy to respond to increasing stand competition. The Scots pine fine-root system was plate-like and showed a low morphological plasticity, thus presumably a conservative strategy. Silver birch exhibited a high biomass and a foraging capacity in the topsoil but a loose root system in the subsoil. The coexistence of pine and beech roots in the upper soil presumably leads to a high belowground competition. Beech root system becomes predominant throughout the soil profile and it adopts an efficient foraging strategy, but at the expense of its belowground development. Conversely, the niche partitioning strategy between beech and birch may explain why beech develops strongly belowground in spite of the fact that birch has a dense rooting and a competitive fine-root architecture. As a consequence, beech mid-term regeneration and development may be facilitated under birch as compared with pine.
DOIURL [本文引用: 1]
The role of species richness, functional diversity and species identity of native Florida sandhill understory species were tested with Imperata cylindrica, an exotic rhizomatous grass, in mesocosms . I. cylindrica was introduced 102year after the following treatments were established: a control with no native species, five monocultures, a grass mix treatment, a forb mix treatment, and a 3-species treatment and a 5-species treatment. Monthly cover, final biomass, root length, root length density (RLD) and specific root length (SRL) of all species were determined for one full growing season. There was a significant negative linear relationship between the cover of native species and I. cylindrica ( r 2 02=020.59, P 02=020.01) and a negative logarithmic relationship between the biomass of native species and I. cylindrica ( r 2 02=020.70, P 02=020.003). There was no diversity–invasibility relationship. Grasses proved to be the most resistant functional group providing resistance alone and in mixed functional communities. Repeated measures analysis demonstrated that treatments including Andropogon virginicus were the most resistant to invasion over time ( P 02<020.001). Significantly greater root length ( P 02=020.002), RLD ( P 02=020.011) and SRL ( P 02<020.001) than all of the native species and I. cylindrica in monocultures and in mixed communities made A. virginicus successful. The root morphology characteristics allowed it to be a great competitor belowground where I. cylindrica was most aggressive. The results suggest that species identity could be more important than species or functional richness in determining community resistance to invasion.
DOIURL [本文引用: 1]
We investigated the effect of competition from grass roots (as controlled by herbicide application) on the depth distribution of white roots in cherry trees, grown with varying rates and frequency of application of N in an agroforestry system. Statistical summaries of distribution, namely mean and skewness, produced a concise and interpretable analysis of the data. There was a large increase in tree root numbers in the surface horizons after the herbicide had reduced grass root competition. Where the surrounding grass had not been reduced by herbicide, the average depth of tree roots increased with time, contrasting with a marked shift in the mode of distribution of root numbers to shallow depths when grass competition was removed. These findings are important in the understanding of plant root competition and for prescribing best practise for tree establishment in agroforestry systems.
DOIURLPMID [本文引用: 1]
In experimental plant communities, relationships between biodiversity and ecosystem functioning have been found to strengthen over time, a fact often attributed to increased resource complementarity between species in mixtures and negative plant-soil feedbacks in monocultures. Here we show that selection for niche differentiation between species can drive this increasing biodiversity effect. Growing 12 grassland species in test monocultures and mixtures, we found character displacement between species and increased biodiversity effects when plants had been selected over 8 years in species mixtures rather than in monocultures. When grown in mixtures, relative differences in height and specific leaf area between plant species selected in mixtures (mixture types) were greater than between species selected in monocultures (monoculture types). Furthermore, net biodiversity and complementarity effects were greater in mixtures of mixture types than in mixtures of monoculture types. Our study demonstrates a novel mechanism for the increase in biodiversity effects: selection for increased niche differentiation through character displacement. Selection in diverse mixtures may therefore increase species coexistence and ecosystem functioning in natural communities and may also allow increased mixture yields in agriculture or forestry. However, loss of biodiversity and prolonged selection of crops in monoculture may compromise this potential for selection in the longer term.
DOIURLPMID [本文引用: 1]
Spatial and temporal niche differentiation are potential mechanisms of plant species' coexistence in many communities, including many grasslands. In a 6-year field experiment, a dominant prairie bunchgrass, Schizachyrium scoparium, excluded species sharing similar phenology and rooting depth, but coexisted with species differing in phenology and rooting depth. We used a series of experimental plots that differed in S. scoparium abundance to measure the effect of S. scoparium on abundances of other species and on soil nitrate concentrations across time and depth. Because we found that S. scoparium most strongly reduced soil nitrate levels at mid-season and at shallow depths, we hypothesized that at higher S. scoparium abundance, species with mid-season phenology and shallow roots would be excluded and that deep-rooted and early season species would be more likely to persist. As predicted, the proportional number and cover of species exploiting different niches than S. scoparium (early season and deep-rooted species) increased across the S. scoparium abundance gradient. This work provides novel empirical evidence for the factors that contribute to species coexistence in the field. Our study suggests that spatial and temporal niche differentiation promote species coexistence in these grasslands.
DOIURL [本文引用: 2]
Here we aim to demonstrate that in arid environments the competitive balance between species can be determined by niche separation with either nitrogen or water as the relevant niche axis. To do this we sampled roots <2/mm in diameter for 5 soil pits equidistant between two coexisting species, a shrub and a grass. Using stable carbon and nitrogen isotope ratios of fine roots we determine both photosynthetic pathway and rooting depth. We also examine the distribution of soil moisture and nitrogen relative to root biomass. Our results for root biomass and stable isotope ratios of fine roots demonstrate both niche separation and competition for resources. Root biomass is highest at the top of the profile where soil nitrogen is highest and soil moisture is lowest. We conclude that while there is competition for resources in the middle of the profile, competition is mitigated by photosynthetic pathway. The facultative CAM shrub grows whenever the soil at the surface is wet enough. The C 4 photosynthetic pathway of the grass is more nitrogen and water use efficient making it better adapted to the low nitrogen in the middle of the profile and low summer rainfall.
DOIURL [本文引用: 1]
The fine root (diameter 0909¤2.0 mm) biomass and morphology of Japanese red pines (Pinus densiflora) grown under different aboveground conditions (i.e., high and low competitive environments) were examined in a pine090009cypress mixed forest. All P. densiflora subject trees were about 40 years old, and the aboveground condition (i.e., size) of red pines appeared to be influenced by the surrounding Japanese cypress (Chamaecyparis obtusa). Smaller P. densiflora exhibited lower fine root biomasses, shorter root lengths, and lower root tip densities, but longer specific root lengths and higher specific root tip densities relative to larger pines. These results suggest that P. densiflora may adjust the morphological traits of fine roots to the different conditions in biomass allocation to fine roots of individuals with different aboveground growth.
DOIURL [本文引用: 1]
URL [本文引用: 1]
自工业革命以来,大气中CO2浓度已从1850年的285 ppmv上升到2012年的约398 ppmv。与20世纪初相比,全球气温已经上升了约0.78。C,据预测到本世纪末,全球温度还将增加1.1-6.4℃。大量研究表明,全球碳循环的动态变化与气候变化及人类活动影响有着密切关系,尤其是化石燃料的燃烧和土地利用方式的变化。据估算,1960-2008年,陆地生态系统吸收了人类活动释放碳的30%。因此,陆地生态系统碳循环是全球碳循环中的重要环节,在调节全球气候变化中扮演着重要角色。然而,由于陆地生态系统对全球气候变化反馈的复杂性,陆地生态系统未来吸收碳的能力是否会持续仍未可知。因此,为了能够更好的维持全球CO2浓度稳定,必须了解陆地生态系统对气候变化的响应和反馈机制。土壤呼吸是陆地生态系统碳循环中仅次于总初级生产力的第二大碳通量,是陆地生态系统碳循环的重要组成部分。因此准确测量土壤呼吸对于估算生态系统碳储量非常重要。由于土壤呼吸速率在自然生态系统中具有高度的空间异质性,因此在野外实验中很难准确估算出生态系统的土壤呼吸。探讨一套合适的野外采样方法对于评估整个陆地碳循环具有重要而又迫切的意义。为了评估取样策略(分层抽样和随机抽样法)和取样个数对实际土壤呼吸估算的影响,我们采用蒙特卡罗模拟法来检测随机和分层取样法的有效性,以浙江天童亚热带常绿阔叶林为研究对象,于2013年8月至2014年1月使用LI-8100土壤呼吸仪测定了不同呼吸环内的土壤呼吸速率及地下5厘米处的温度和水分。结果显示:土壤呼吸平均值不受取样数量和取样策略的影响,但其变异系数随着取样个数的增加而逐渐减少;在生长季,分层取样的精确度比随机取样法提高了约16.7%,工作量减少了约36.4%,在非生长季,这两个数值分别为8.7%和22.2%。运用模型估算出10℃下土壤呼吸速率(Rio)约为1.0μmol CO2/m2 s,温度敏感系数(Q10)约为2.0。二者的平均值虽不受取样策略和个数的影响,但其变异系数均随着取样个数的增加而减少,且两种取样策略间没有明显差异。我们于2013年6月至2014年5月使用LI-8100土壤呼吸仪测定了干旱、干扰和空白处理方式下的呼吸环内的土壤呼吸速率。结果显示:短期干旱对陆地生态系统土壤呼吸速率及其空间异质性没有明显影响,干扰对土壤呼吸的变异系数有较大影响,在非生长季时,变异系数增加了100%。干扰使Rio的值增加了33%,变异系数增加了109%,短期干旱对Rio的值没有明显影响,使变异系数降低了约48%。干扰使Q1o降低了约20%,变异系数增加了100%;短期干旱对Q10没有明显影响,使变异系数减少了约45%。
URL [本文引用: 1]
自工业革命以来,大气中CO2浓度已从1850年的285 ppmv上升到2012年的约398 ppmv。与20世纪初相比,全球气温已经上升了约0.78。C,据预测到本世纪末,全球温度还将增加1.1-6.4℃。大量研究表明,全球碳循环的动态变化与气候变化及人类活动影响有着密切关系,尤其是化石燃料的燃烧和土地利用方式的变化。据估算,1960-2008年,陆地生态系统吸收了人类活动释放碳的30%。因此,陆地生态系统碳循环是全球碳循环中的重要环节,在调节全球气候变化中扮演着重要角色。然而,由于陆地生态系统对全球气候变化反馈的复杂性,陆地生态系统未来吸收碳的能力是否会持续仍未可知。因此,为了能够更好的维持全球CO2浓度稳定,必须了解陆地生态系统对气候变化的响应和反馈机制。土壤呼吸是陆地生态系统碳循环中仅次于总初级生产力的第二大碳通量,是陆地生态系统碳循环的重要组成部分。因此准确测量土壤呼吸对于估算生态系统碳储量非常重要。由于土壤呼吸速率在自然生态系统中具有高度的空间异质性,因此在野外实验中很难准确估算出生态系统的土壤呼吸。探讨一套合适的野外采样方法对于评估整个陆地碳循环具有重要而又迫切的意义。为了评估取样策略(分层抽样和随机抽样法)和取样个数对实际土壤呼吸估算的影响,我们采用蒙特卡罗模拟法来检测随机和分层取样法的有效性,以浙江天童亚热带常绿阔叶林为研究对象,于2013年8月至2014年1月使用LI-8100土壤呼吸仪测定了不同呼吸环内的土壤呼吸速率及地下5厘米处的温度和水分。结果显示:土壤呼吸平均值不受取样数量和取样策略的影响,但其变异系数随着取样个数的增加而逐渐减少;在生长季,分层取样的精确度比随机取样法提高了约16.7%,工作量减少了约36.4%,在非生长季,这两个数值分别为8.7%和22.2%。运用模型估算出10℃下土壤呼吸速率(Rio)约为1.0μmol CO2/m2 s,温度敏感系数(Q10)约为2.0。二者的平均值虽不受取样策略和个数的影响,但其变异系数均随着取样个数的增加而减少,且两种取样策略间没有明显差异。我们于2013年6月至2014年5月使用LI-8100土壤呼吸仪测定了干旱、干扰和空白处理方式下的呼吸环内的土壤呼吸速率。结果显示:短期干旱对陆地生态系统土壤呼吸速率及其空间异质性没有明显影响,干扰对土壤呼吸的变异系数有较大影响,在非生长季时,变异系数增加了100%。干扰使Rio的值增加了33%,变异系数增加了109%,短期干旱对Rio的值没有明显影响,使变异系数降低了约48%。干扰使Q1o降低了约20%,变异系数增加了100%;短期干旱对Q10没有明显影响,使变异系数减少了约45%。
URL [本文引用: 1]
采用盆栽试验,分别在刺槐(Robinia pseudoacacia)幼树生长初期、生长盛期和生长后期进行了5种不同土壤水分含量试验,以研究不同生长阶段内干旱胁迫对该树种干物质分配的影响.结果表明:各生长阶段内刺槐幼树茎、枝分配比率并不受土壤水分含量变化的影响,短期(15 d)及中轻度干旱(70%和87.8%相对土壤含水量)胁迫对刺槐叶、地上部分干物质分配比率及根冠比的影响亦不明显;而长期(45~60 d)严重干旱(40%相对土壤含水量)胁迫则显著降低了刺槐叶、地上部分配比率,提高了粗根干物质分配比率及根冠比.受树木本身生长特性和气候因素的季节性变化影响,不同生长阶段之间干旱胁迫对刺槐干物质分配的影响存在明显差异,其影响程度大小为生长初期>生长盛期>生长后期.
URL [本文引用: 1]
采用盆栽试验,分别在刺槐(Robinia pseudoacacia)幼树生长初期、生长盛期和生长后期进行了5种不同土壤水分含量试验,以研究不同生长阶段内干旱胁迫对该树种干物质分配的影响.结果表明:各生长阶段内刺槐幼树茎、枝分配比率并不受土壤水分含量变化的影响,短期(15 d)及中轻度干旱(70%和87.8%相对土壤含水量)胁迫对刺槐叶、地上部分干物质分配比率及根冠比的影响亦不明显;而长期(45~60 d)严重干旱(40%相对土壤含水量)胁迫则显著降低了刺槐叶、地上部分配比率,提高了粗根干物质分配比率及根冠比.受树木本身生长特性和气候因素的季节性变化影响,不同生长阶段之间干旱胁迫对刺槐干物质分配的影响存在明显差异,其影响程度大小为生长初期>生长盛期>生长后期.
DOIURL
In a field experiment in the Yellow River Basin conducted in 2007 and 2008, it was found that, under full irrigation, the roots of maize not only penetrated deeper than those of soybean but also extended into soybean stands underneath the space between inner rows of soybean. The roots of soybean, however, were confined mainly to the zone near the plants. Horizontal growth of the roots of both the crops was confined mainly to the soil layer 16–22 cm below the surface, a layer that lay above an existing plough pan. Root length density (RLD) was much higher in the top layer (0–30 cm deep) and in the zone closer to the plants. The exponential model proved suitable to describe the RLD vertically and horizontally in both sole cropping and in intercropping.
[本文引用: 1]
DOIURL [本文引用: 2]
Background and aims Morphology and functioning of fine roots vary with distance from the distal root ending reflecting influences of root order and root age, but changes in tree fine root systems as a...
DOIURLPMID
Abstract Since proposed two decades ago, the stress-gradient hypothesis (SGH), suggesting that species interactions shift from competition to facilitation with stress, has been widely examined. Despite broad support across species and ecosystems, ecologists debate whether the SGH applies to extreme environments, arguing that species interactions switch to competition or collapse under extreme stress. We show that facilitation often expands distributions on species borders. SGH exceptions occur when weak stress gradients or stresses outside of species' niches are examined, multiple stresses co-occur canceling out their effects, temporally dependent effects are involved, or results are improperly analyzed. We suggest that ecologists resolve debates by standardizing key SGH terms, such as fundamental and realized niche, stress gradients vs. environmental gradients, by quantitatively defining extreme stress, and by critically evaluating the functionality of stress gradients. We also suggest that new research examine the breadth and relevance of the SGH. More rigor needs to be applied to SGH tests to identify actual exceptions rather than those due to failures to meet its underlying assumptions, so that the general principles of the SGH and its exceptions can be incorporated into ecological theory, conservation strategies, and environmental change predictions.
DOIURLPMID
Much is known about how environmental stress mediates the strength of facilitation, but less is known about how different plant traits affect facilitation. We examined interactions between the shrub Tamarix chinensis and two congeneric forbs (Suaeda salsa and S. glauca) on the Chinese coast. Although S. salsa and S. glauca are both annuals, morphologically similar, and have synchronous phenologies, they have contrasting adaptive strategies. S. glauca is salt intolerant but competitively superior, and S. salsa is salt tolerant but competitively inferior. Field surveys showed that S. glauca was associated with T. chinensis canopies while S. salsa was more abundant in open areas. A T. chinensis removal experiment showed that S. glauca cover was lower and soil salinity higher after two years in removal than in control plots. Transplant experiments showed that S. salsa performance under T. chinensis canopies was reduced by competition from S. glauca and T. chinensis, while in open areas S. glauca was not affected by S. salsa competition. Thus, contrasting competitive abilities and stress tolerances of S. glauca and S. salsa underlie their facultative and competitive interactions with T. chinensis, suggesting that plant strategies are critical to the outcome of species interactions.
DOIURL [本文引用: 1]
When roots encounter a nutrient-rich zone or patch they often proliferate within it. Roots experiencing nutrient-rich patches can also enhance their physiological ion-uptake capacities compared with roots of the same plant outside the patch zone. These plastic responses by the root system have been proposed as the major mechanism by which plants cope with the naturally occurring heterogeneous supplies of nutrients in soil. Various attempts to predict how contrasting species will respond to patches have been made based on specific root length (SRL), root demography and biomass allocation within the patch zone. No one criterion has proved definitive. Actually demonstrating that root proliferation is beneficial to the plant, especially in terms of nitrogen capture from patches, has also proved troublesome. Yet by growing plants under more realistic conditions, such as in interspecific plant competition, and with a complex organic patch, a direct benefit can be demonstrated. Thus, as highlighted in this review, the environmental context in which the root response is expressed is as important as the magnitude of the response itself.
DOIURL [本文引用: 2]
ABSTRACT p1 Present address: Department of Zoology, University of Florida, Gainesville, FL, USA. Email: RHOLDO@zoo.ufl.edu
DOIURLPMID [本文引用: 1]
Little is known about below-ground competition between different tree species in mixed forests. We investigated the evidence for asymmetric competition between fine roots (<2 mm) of adult European beech (Fagus sylvatica) and sessile oak (Quercus petraea) trees in a mixed temperate beech-oak forest by (1) conducting fine-root growth experiments in the field (root chamber technique), (2) comparing the fine-root mass of two-species and monospecific plots, and (3) analysing the density and overlap of beech and oak root systems in shared soil volumes. Field experiments with root chambers, which allow fine-root endings to grow under controlled conditions for several months, showed that beech grew more rapidly than oak roots when both species were grown together. In the mixed beech-oak wood, where stem densities and leaf areas of the two species were similar, beech outnumbered oak three- to five-fold in fine-root biomass, and root tip and ectomycorrhiza numbers, which led to a much greater root:shoot ratio (root area index:leaf area index, RAI:LAI) for beech (3.9) than oak (1.7). The remarkably small fine-root biomass of oak was attributed to competitive replacement by beech roots as indicated by comparison with monospecific oak wood. Although oak had much less fine-root mass than beech, oak outnumbered beech in the coarse root fraction (2<d<5 mm) resulting in a greater fine root:coarse root ratio in beech than oak. The fine-root systems of the two species completely overlapped in the nitrogen-rich organic topsoil but differed with respect to their horizontal distribution patterns: beech root biomass increased with increasing distance from a beech stem whereas oak root mass was not dependent on stem distance. Beech (but not oak) root mass was correlated with the thickness of the organic topsoil horizons which, at average, were thicker at greater distances from beech stems. We conclude that beech fine roots colonise nitrogen-rich patches in the organic horizons of this forest more successfully than do oak roots. Our results (root chamber experiments, oak root abundance in mixed vs. monospecific wood, beech root dominance in the organic topsoil) suggest that asymmetric interspecific root competition in favour of beech exists in this stand.
DOIURL [本文引用: 1]
AimsMethodsImportant findings A. sparsifolia roots needs both nutrients and soil space, and space is more important than nutrients. If neighbors are present, plant roots first occupy the space where competitors exist.
DOIURL [本文引用: 1]
AimsMethodsImportant findings A. sparsifolia roots needs both nutrients and soil space, and space is more important than nutrients. If neighbors are present, plant roots first occupy the space where competitors exist.
DOIURL [本文引用: 1]
多枝柽柳(Tamarix ramosissima)是塔里木河下游荒漠河岸林中的优势灌木,对荒漠河岸植被群落的稳定起着重要作用。该文通过研究多枝柽柳幼苗根系形态对不同灌溉处理的响应,分析人工水分干扰对多枝柽柳幼苗根系生长的影响。实验设计了侧渗分层和地表灌溉两种给水方式和高灌(50L·株–1)、中灌(25L·株–1)、低灌(12.5L·株–1)三个给水水平,并在整个生长季节监测每个植株的生物量及根系形态参数。结果显示:与地表灌溉比较,侧渗分层的灌溉方式显著提高了细根(0.5mmd2mm)长、细根表面积和根系生物量,并使根系生长至160cm深度的土层,大于地表灌溉深度(80–100cm);侧渗分层灌溉+高灌的组合促进根系生长的效果最显著(p0.05);侧渗分层灌溉方式下总细根(d2mm)的比根长随着给水量的增加显著增大,而地表灌溉下比根长无显著变化;侧渗分层灌溉方式下根冠比总体小于地表灌溉方式,即侧渗分层灌溉使多枝柽柳地上部分发育较好。因此,侧渗分层灌溉方式有显著促进多枝柽柳幼苗在生长早期快速发育的效果。
DOIURL [本文引用: 1]
多枝柽柳(Tamarix ramosissima)是塔里木河下游荒漠河岸林中的优势灌木,对荒漠河岸植被群落的稳定起着重要作用。该文通过研究多枝柽柳幼苗根系形态对不同灌溉处理的响应,分析人工水分干扰对多枝柽柳幼苗根系生长的影响。实验设计了侧渗分层和地表灌溉两种给水方式和高灌(50L·株–1)、中灌(25L·株–1)、低灌(12.5L·株–1)三个给水水平,并在整个生长季节监测每个植株的生物量及根系形态参数。结果显示:与地表灌溉比较,侧渗分层的灌溉方式显著提高了细根(0.5mmd2mm)长、细根表面积和根系生物量,并使根系生长至160cm深度的土层,大于地表灌溉深度(80–100cm);侧渗分层灌溉+高灌的组合促进根系生长的效果最显著(p0.05);侧渗分层灌溉方式下总细根(d2mm)的比根长随着给水量的增加显著增大,而地表灌溉下比根长无显著变化;侧渗分层灌溉方式下根冠比总体小于地表灌溉方式,即侧渗分层灌溉使多枝柽柳地上部分发育较好。因此,侧渗分层灌溉方式有显著促进多枝柽柳幼苗在生长早期快速发育的效果。
[本文引用: 1]
DOIURL [本文引用: 1]
Root systems are highly plastic as they express a range of responses to acquire patchily distributed nutrients. However, the ecological significance of placing roots selectively in nutrient hotspots is still unclear. Here, we investigate under what conditions selective root placement may be a significant functional trait that determines belowground competitive ability. We studied two grasses differing in root foraging behaviour, Festuca rubra and Anthoxanthum odoratum . The plants were grown in stable and more dynamic heterogeneous environments, by switching nutrient patches halfway through the experiment. A. odoratum was a factor of two less selective in placing its roots into nutrient-rich patches than F. rubra . A. odoratum produced overall higher root length densities with higher specific root length than F. rubra and acquired more nutrients. A. odoratum appeared to be the superior competitor, irrespective of the nutrient dynamics. Our results suggest that root behaviour consisting of producing high root length densities at relatively low biomass investments can be a more effective foraging strategy than placing roots selectively in nutrient hotspots. When understanding the functionality of root traits among different species, specific root length may play a key role.
DOIURL
DOIURL
1. Seedling survival is one of the most critical stages in a plant's life history, and is often reduced by drought and soil desiccation. It has been hypothesized that root systems accessing moist soil layers are critical for establishment, but very little is known about seedling root growth and traits in the field. 2. We related seedling mortality to the presence of deep roots in a field experiment in which we monitored soil moisture, root growth and seedling survival in five Mediterranean woody species from the beginning of the growing season until the end of the drought season. 3. We found strong positive relationships between survival and maximum rooting depth, as well as between survival and soil moisture. Species with roots in moist soil layers withstood prolonged drought better, whereas species with shallow roots died more frequently. In contrast, biomass allocation to roots was not related to establishment success. 4. Access to moist soil horizons accounted for species-specific survival rates, whereas large root: shoot (R:S) ratios did not. The existence of soil moisture thresholds that control establishment provides insights into plant population dynamics in dry environments.
DOIURL
1 In water-limited environments, the availability of water and nutrients to plants depends on environmental conditions, sizes and shapes of their root systems, and root competition. The goal of this study was to predict root system sizes and shapes for different plant growth forms using data on above-ground plant sizes, climate and soil texture. 2 A new data set of > 1300 records of root system sizes for individual plants was collected from the literature for deserts, scrublands, grasslands and savannas with 鈮 1000 mm mean annual precipitation (MAP). Maximum rooting depths, maximum lateral root spreads and their ratios were measured. 3 Root system sizes differed among growth forms and increased with above-ground size: annuals < perennial forbs = grasses < semi-shrubs < shrubs < trees. Stem succulents were as shallowly rooted as annuals but had lateral root spreads similar to shrubs. 4 Absolute rooting depths increased with MAP in all growth forms except shrubs and trees, but were not strongly related to potential evapotranspiration (PET). Except in trees, root systems tended to be shallower and wider in dry and hot climates and deeper and narrower in cold and wet climates. Shrubs were more shallowly rooted under climates with summer than winter precipitation regimes. 5 Relative to above-ground plant sizes, root system sizes decreased with increasing PET for all growth forms, but decreased with increasing MAP only for herbaceous plants. Thus relative rooting depths tended to increase with aridity, although absolute rooting depths decreased with aridity. 6 Using an independent data set of 20 test locations, rooting depths were predicted from MAP using regression models for three broad growth forms. The models succeeded in explaining 62% of the observed variance in median rooting depths. 7 Based on the data analysed here, Walter's two-layer model of soil depth partitioning between woody and herbaceous plants appears to be most appropriate in drier regimes (< 500 mm MAP) and in systems with substantial winter precipitation.
DOIURL [本文引用: 1]
Background and Aims Root segregation has been observed in a variety of species. Though it is usually attributed to a plant’s attempt to avoid below ground competition, a direct correlation of root...
URL [本文引用: 1]
The architecture parameters of Reaumuria soongorica root system at different habitats in Gansu Province, China were analyzed to examine its ecological adaptability to arid environment. Results show that: (1) Topological indices of R. soongorica root system are small at all the habitats, and root branching pattern tends to be dichotomous. The topological indices of R. soongorica root system gradually increase in the Minqin windblown sand region and the Zhangye gobi region in Hexi Corridor, which indicates that drought makes root branching pattern tend to be herringbone-like. (2) Fractal dimension values of R. soongorica root system all are small in the Minqin windblown sand region and the Zhangye gobi region in Hexi Corridor, and the values are 1.1778 and 1.1169, respectively, which indicates that the fractal characteristics are not obvious in these areas. While the fractal dimension values are relative large in Jiuzhoutai semi-arid hilly and gully region of the Loess Plateau, which indicates that the R. soongorica root system has better fractal characteristics in this region than in the other regions. (3) Total branching ratios of the R. soongorica root system in arid region of Hexi Corridor are smaller than that in the Jiuzhoutai semi-arid hilly and gully region of the Loess Plateau. It shows that the root branching ability in the semi-arid region is stronger, and it decreases at some degree with drought increase. (4) The root link lengths of R. soongorica root system are long at all the habitats, but there are significant differences between the different habitats. The root link length at the Minqin windblown sand region is the longest. It is concluded that R. soongorica adapts to arid environment by decreasing root branching, decreasing root overlap and increasing the root link length, which makes its root branching pattern tend to be herringbone-like to reduce compete degree in root internal environment for nutrients and to enhance root absorption rate to nutrients, and ensure the effective nutrition space, thus it can absorb enough water and nutrients in resource-poor settings to ensure its normal physiological requirements.
URL [本文引用: 1]
The architecture parameters of Reaumuria soongorica root system at different habitats in Gansu Province, China were analyzed to examine its ecological adaptability to arid environment. Results show that: (1) Topological indices of R. soongorica root system are small at all the habitats, and root branching pattern tends to be dichotomous. The topological indices of R. soongorica root system gradually increase in the Minqin windblown sand region and the Zhangye gobi region in Hexi Corridor, which indicates that drought makes root branching pattern tend to be herringbone-like. (2) Fractal dimension values of R. soongorica root system all are small in the Minqin windblown sand region and the Zhangye gobi region in Hexi Corridor, and the values are 1.1778 and 1.1169, respectively, which indicates that the fractal characteristics are not obvious in these areas. While the fractal dimension values are relative large in Jiuzhoutai semi-arid hilly and gully region of the Loess Plateau, which indicates that the R. soongorica root system has better fractal characteristics in this region than in the other regions. (3) Total branching ratios of the R. soongorica root system in arid region of Hexi Corridor are smaller than that in the Jiuzhoutai semi-arid hilly and gully region of the Loess Plateau. It shows that the root branching ability in the semi-arid region is stronger, and it decreases at some degree with drought increase. (4) The root link lengths of R. soongorica root system are long at all the habitats, but there are significant differences between the different habitats. The root link length at the Minqin windblown sand region is the longest. It is concluded that R. soongorica adapts to arid environment by decreasing root branching, decreasing root overlap and increasing the root link length, which makes its root branching pattern tend to be herringbone-like to reduce compete degree in root internal environment for nutrients and to enhance root absorption rate to nutrients, and ensure the effective nutrition space, thus it can absorb enough water and nutrients in resource-poor settings to ensure its normal physiological requirements.
DOIURL [本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL
react-text: 487 A vast body of literature demonstrated that anthropogenic disturbances such as overgrazing and fire are key drivers of abrupt transition between vegetation types in ecosystems. In this study, we propose that the hydrological context (described in terms of rainfall, evapotranspiration and water yield) is a first-order, primordial determinant of the propensity of ecosystems to undergo... /react-text react-text: 488 /react-text [Show full abstract]
DOIURL [本文引用: 1]
为了更有效地从土壤中获取养分,植物根系在长期的进化与适应中产生了一系列塑性反应,以响应自然界中广泛存在的时空异质性.同时,植物根系的养分吸收也要面对来自种内和种间的竞争.多种因素都会影响植物根竞争的结果,包括养分条件、养分异质性的程度、根系塑性的表达等.竞争会改变植物根系的塑性反应,比如影响植物根系的空间分布;植物根系塑性程度差异也会影响竞争.已有研究发现根系具有高形态塑性和高生理塑性的植物在长期竞争过程中会占据优势.由于不同物种根系塑性的差异,固定的对待竞争的反应模式在植物根系中可能并不存在,其响应随竞争物种以及土壤环境因素的变化而变化.此外,随着时间变化,根系塑性的反应及其重要性也会随之改变.植物对竞争的反应可能与竞争个体之间的亲缘关系有关,有研究表明亲缘关系近的植物可能倾向于减小彼此之间的竞争.根竞争对植物的生存非常重要,但目前还没有研究综合考虑植物的各种塑性在根竞争中的作用.另外根竞争对群落结构的影响尚待深入的研究.
DOIURL [本文引用: 1]
为了更有效地从土壤中获取养分,植物根系在长期的进化与适应中产生了一系列塑性反应,以响应自然界中广泛存在的时空异质性.同时,植物根系的养分吸收也要面对来自种内和种间的竞争.多种因素都会影响植物根竞争的结果,包括养分条件、养分异质性的程度、根系塑性的表达等.竞争会改变植物根系的塑性反应,比如影响植物根系的空间分布;植物根系塑性程度差异也会影响竞争.已有研究发现根系具有高形态塑性和高生理塑性的植物在长期竞争过程中会占据优势.由于不同物种根系塑性的差异,固定的对待竞争的反应模式在植物根系中可能并不存在,其响应随竞争物种以及土壤环境因素的变化而变化.此外,随着时间变化,根系塑性的反应及其重要性也会随之改变.植物对竞争的反应可能与竞争个体之间的亲缘关系有关,有研究表明亲缘关系近的植物可能倾向于减小彼此之间的竞争.根竞争对植物的生存非常重要,但目前还没有研究综合考虑植物的各种塑性在根竞争中的作用.另外根竞争对群落结构的影响尚待深入的研究.
DOIURL [本文引用: 1]
对3种密度进行栽培试验,利用通径分析的方法,研究了水曲柳地下竞争和地上竞争的关系及对总竞争的影响.结果表明,水曲柳苗木的地下部分生物量、地上部分生物量和总生物量与营养空间有密切关系.随着苗木空间距离增加,由生物量计算的竞争指数下降.在同一密度条件下,地下竞争指数明显大于地上竞争指数.由于地下生长与地上生长的相互作用,各竞争指数之间具有明显的相关性.但是地下竞争和地上竞争对总竞争的影响是不同的,通径分析可以定量的区分地下竞争和地上竞争的相对大小.地下竞争对总竞争的直接作用范围在 0.5543~0.7426之间,明显大于地上竞争对总竞争的直接作用(0.2851~0.5282).随着距离的增加,单株苗木的生长空间加大,地上部分的竞争作用增加,地下部分的竞争程度减弱.但是,地下根系的竞争在水曲柳苗木总的竞争中占有重要地位.
DOIURL [本文引用: 1]
对3种密度进行栽培试验,利用通径分析的方法,研究了水曲柳地下竞争和地上竞争的关系及对总竞争的影响.结果表明,水曲柳苗木的地下部分生物量、地上部分生物量和总生物量与营养空间有密切关系.随着苗木空间距离增加,由生物量计算的竞争指数下降.在同一密度条件下,地下竞争指数明显大于地上竞争指数.由于地下生长与地上生长的相互作用,各竞争指数之间具有明显的相关性.但是地下竞争和地上竞争对总竞争的影响是不同的,通径分析可以定量的区分地下竞争和地上竞争的相对大小.地下竞争对总竞争的直接作用范围在 0.5543~0.7426之间,明显大于地上竞争对总竞争的直接作用(0.2851~0.5282).随着距离的增加,单株苗木的生长空间加大,地上部分的竞争作用增加,地下部分的竞争程度减弱.但是,地下根系的竞争在水曲柳苗木总的竞争中占有重要地位.
DOIURL [本文引用: 1]
Root-root interactions were studied in green-house potted Manchurian ash (Fraxinus mandshurica) and Dahurian larch (Larix gmelinii) seedlings. Above and below ground growth of ash mixed was influenced significantly by the interaction with larch in mixture. The root system of ash was more sensitive to soil conditions than that of the larch, and its competitive effectiveness was enhanced by impoved nutrition. Relative to their mass in unmixed pots the fine and coarse root mass was increased by 47% and 46% respectively, and leaf and stem mass was increased about 30% and 48% respectively in mixed pots. Root-root interactions affected the root/shoot ratio, especially the fine root mass/leaf mass ratio, in ash. In contrast, biomass components and root/shoot ratio of larch showed no significant response in mixture. Through morphological adaptations ash altered its root distribution and morphology in mixture: fine root density and specific root length increased by 47 and 34% over pure planted pots. With relatively larger root mass and thinner root diameter, ash is a superior competitor for soil nutrients and moisture. Larch, on the other hand, is capable of improving soil conditions through its growth.
DOIURL [本文引用: 1]
Root-root interactions were studied in green-house potted Manchurian ash (Fraxinus mandshurica) and Dahurian larch (Larix gmelinii) seedlings. Above and below ground growth of ash mixed was influenced significantly by the interaction with larch in mixture. The root system of ash was more sensitive to soil conditions than that of the larch, and its competitive effectiveness was enhanced by impoved nutrition. Relative to their mass in unmixed pots the fine and coarse root mass was increased by 47% and 46% respectively, and leaf and stem mass was increased about 30% and 48% respectively in mixed pots. Root-root interactions affected the root/shoot ratio, especially the fine root mass/leaf mass ratio, in ash. In contrast, biomass components and root/shoot ratio of larch showed no significant response in mixture. Through morphological adaptations ash altered its root distribution and morphology in mixture: fine root density and specific root length increased by 47 and 34% over pure planted pots. With relatively larger root mass and thinner root diameter, ash is a superior competitor for soil nutrients and moisture. Larch, on the other hand, is capable of improving soil conditions through its growth.
DOIURL [本文引用: 1]
以新疆古尔班通古特沙漠南缘原始盐生旱生荒漠的3种建群灌木多枝柽柳(Tamarixramosissima)、梭梭(Haloxylonammodendron)和琵琶柴(Reaumuriasoongorica)为对象,跟踪自然降雨过程,利用LI-6400光合作用系统和Model3005植物水分压力室测定光合作用和叶水势的变化,以研究浅层土壤水分条件改变对荒漠灌木主要叶片生理特征的影响;并在原始生境中将植株根系完整地深挖取出,进行根系形态结构调查,以确定此3种灌木根系功能型与用水策略。当浅层土壤分别处在水分充足及匮缺的条件下时,测定3种灌木的光合作用响应曲线和日过程曲线,以及黎明前和正午叶水势,结果表明:浅层土壤水分状况变化时,3种灌木的光合能力均没有显著改变;多枝柽柳的叶水势亦没有明显波动;而梭梭和琵琶柴的叶水势却表现出显著差异。在两种功能型根中,多枝柽柳为深根型,生存和生理活动的维持主要依赖于地下水;而梭梭和琵琶柴为非深根型植物,主要水源是降水形成的浅层土壤水,其用水策略是根据水分条件有效调节根系和冠层生长,从而维持正常的光合作用。即荒漠灌木在长期适应的过程中,已形成不同的根系功能型和用水策略;叶水势对浅层土壤水分状况变化的种间差异性响应在一定程度上反映了这一点。同时,此3种荒漠灌木通过不同的个体适应策略都能够实现水分平衡和碳收支的有效调节,这主要体现为浅层土壤水分条件变化时光合响应的种间一致性。
DOIURL [本文引用: 1]
以新疆古尔班通古特沙漠南缘原始盐生旱生荒漠的3种建群灌木多枝柽柳(Tamarixramosissima)、梭梭(Haloxylonammodendron)和琵琶柴(Reaumuriasoongorica)为对象,跟踪自然降雨过程,利用LI-6400光合作用系统和Model3005植物水分压力室测定光合作用和叶水势的变化,以研究浅层土壤水分条件改变对荒漠灌木主要叶片生理特征的影响;并在原始生境中将植株根系完整地深挖取出,进行根系形态结构调查,以确定此3种灌木根系功能型与用水策略。当浅层土壤分别处在水分充足及匮缺的条件下时,测定3种灌木的光合作用响应曲线和日过程曲线,以及黎明前和正午叶水势,结果表明:浅层土壤水分状况变化时,3种灌木的光合能力均没有显著改变;多枝柽柳的叶水势亦没有明显波动;而梭梭和琵琶柴的叶水势却表现出显著差异。在两种功能型根中,多枝柽柳为深根型,生存和生理活动的维持主要依赖于地下水;而梭梭和琵琶柴为非深根型植物,主要水源是降水形成的浅层土壤水,其用水策略是根据水分条件有效调节根系和冠层生长,从而维持正常的光合作用。即荒漠灌木在长期适应的过程中,已形成不同的根系功能型和用水策略;叶水势对浅层土壤水分状况变化的种间差异性响应在一定程度上反映了这一点。同时,此3种荒漠灌木通过不同的个体适应策略都能够实现水分平衡和碳收支的有效调节,这主要体现为浅层土壤水分条件变化时光合响应的种间一致性。
DOIURL [本文引用: 1]
Spatial pattern and spatial association analysis are important in the study of population characteristics and interspecies relationships. To investigate the spatial distribution pattern and association in different developmental stages in a Quercus acutissima population,the natural secondary Q. acutissima forest of the Qiaoshan Mountains in the southern area of the Loess Plateau was used as our study area. One forest plot( 50 m×50 m) was established in each Q. acutissima natural secondary pure forest( plot A) and Q. acutissima-Pinus tabuliformis natural secondary mixed forest( plot B). The coordinate of each Q. acutissima was mapped,the diameter at breast height and height of each individual were measured by plot to investigate means. We divided the individuals into four growth stages using the method of diameter class. The spatial distribution pattern and associations were analyzed using Ripley's L( r) Function. The results showed the followings:( 1)The quantity structure of each developmental stage of the Q. acutissima population in the Qiaoshan Mountains exhibited an irregular inverted " J" type,indicating a stable population structure with persistent regeneration capability.( 2) The spatial distribution of the two populations was greatly influenced by habitat heterogeneity. Spatial distribution exhibited high aggregation at the scale of 0—10 m in plot A,whereas it showed low aggregation at the scale of 5—35 m. In plot B,the degree of aggregation changed quickly in response to habitat heterogeneity. It exhibited high aggregation at the 0—5 m scaleand low aggregation at the 5—35 m scale.( 3) With the development of populations and increase in scale,the spatial aggregation of the Q. acutissima population was reduced. In plot A,the seedling stage showed an aggregated distribution at the small scale of 0—6 m and a random distribution at the larger scale of 6—12 m. The sapling stage showed an aggregated distribution at all scales. The mid-mature stage and mature stage showed a random distribution at all scales. In plot B,all stages showed a random distribution at all the scales,except for the aggregated distribution of the seedling stage at the small scale of 0—6 m.( 4) The spatial correlation between different growth stages and scales was discriminating in the two plots.In plot A,we found a negative correlation between the seedling and sapling stage at the 1—2 m scale. Positive correlation was found between the sapling and mid-mature stage at the 8 m scale. However,the relationship among other stages was not significant at any scale. In plot B,there was a significant positive correlation between the sapling and mid-mature stage at the 2—12 m scale. There was almost no correlation among other stages at any scales. Population distribution patterns were influenced by developmental stages,biological characteristics,and ecological factors. In future forest management,different managerial measures need to be taken to promote the regeneration of the population according to the different developmental stages and different habitats of the population.
URL [本文引用: 1]
The mixed community of C3 and C4 plants is easily seen in grassland ecosystem, but hardly seen in desert ecosystem. In desert regions of China, some C3 and C4 shrubs coexist in particular manners in some specific habitats and adapt to the extreme arid environment, which is associated with high temperature and intensive radiation. The mixed communities of C3 undershrub Reaumuria soongorica and C4 subshrub Salsola passerina in the Heihe River Basin in Hexi Corridor were investigated. The physiological characteristics of gas exchange of R. soongorica and S. passerina in the mixed community were determined under different drought degrees. Results showed that net photosynthetic rate (Pn), transpiration rate (E), stomatal conductance (Gs) of R. soongorica were all higher than S. passerina, while water use efficiency of S. passerina was higher than R. soongorica. The differential responses of the two species to severe habitat shows that R. soongorica and S. passerina take different strategies for survival when suffering water scarcity in the same desert habitats: R. soongorica uses the means of maintaining a higher net photosynthetic rate and a higher transpiration rate to survive, while S. passerina survives here through the higher water use efficiency.
URL [本文引用: 1]
The mixed community of C3 and C4 plants is easily seen in grassland ecosystem, but hardly seen in desert ecosystem. In desert regions of China, some C3 and C4 shrubs coexist in particular manners in some specific habitats and adapt to the extreme arid environment, which is associated with high temperature and intensive radiation. The mixed communities of C3 undershrub Reaumuria soongorica and C4 subshrub Salsola passerina in the Heihe River Basin in Hexi Corridor were investigated. The physiological characteristics of gas exchange of R. soongorica and S. passerina in the mixed community were determined under different drought degrees. Results showed that net photosynthetic rate (Pn), transpiration rate (E), stomatal conductance (Gs) of R. soongorica were all higher than S. passerina, while water use efficiency of S. passerina was higher than R. soongorica. The differential responses of the two species to severe habitat shows that R. soongorica and S. passerina take different strategies for survival when suffering water scarcity in the same desert habitats: R. soongorica uses the means of maintaining a higher net photosynthetic rate and a higher transpiration rate to survive, while S. passerina survives here through the higher water use efficiency.
DOIURL
Biomass mainly reflected the accumulated energy by plant. Plant species have evolved specialized strategies to regulate biomass allocation among various organs or between above- and belowground biomass(R/S). Biomass allocation can affect carbon turnover among different pools and the terrestrial ecosystem carbon cycling. We investigated biomass allocation patterns and characters of root distribution of four common shrubs on individual levels in desert area, southeast of the Tengger Desert, China. Our results indicated that the biomass allocation strategy in above ground organs were different among four shrubs, the R/S were also different. These results reflect every shrub have evolved their unique growth strategy. The canopy and biomass increased and R/S decreased with the growth of shrubs. Shrubs may have higher competition needs for limited resources at the young than older stage. Root biomass distribution are different among Kalidium foliatum (in the uppermost 0-30 cm of the soil), Salsola passerina (in the uppermost 0-50 cm of the soil), Ceratoides latens and Reaumuria soongorica (in the uppermost 0-70 cm of the soil). Reduced major axis analysis showed that the slopes of the relationship between above- and belowground biomass of Ceratoides lateens, Salsola passerine and Reaumuria soongorica close to 1, which support isometric scaling hypothesis. However, the slope of the relationship between above- and belowground biomass of Kalidium foliatum was significantly different from 1.0, which don support this hypothesis. There is a significant linear relationship between above- and belowground biomass after logarithmic transformation. The relationship can be used to estimate belowground biomass effectively.
DOIURL
Biomass mainly reflected the accumulated energy by plant. Plant species have evolved specialized strategies to regulate biomass allocation among various organs or between above- and belowground biomass(R/S). Biomass allocation can affect carbon turnover among different pools and the terrestrial ecosystem carbon cycling. We investigated biomass allocation patterns and characters of root distribution of four common shrubs on individual levels in desert area, southeast of the Tengger Desert, China. Our results indicated that the biomass allocation strategy in above ground organs were different among four shrubs, the R/S were also different. These results reflect every shrub have evolved their unique growth strategy. The canopy and biomass increased and R/S decreased with the growth of shrubs. Shrubs may have higher competition needs for limited resources at the young than older stage. Root biomass distribution are different among Kalidium foliatum (in the uppermost 0-30 cm of the soil), Salsola passerina (in the uppermost 0-50 cm of the soil), Ceratoides latens and Reaumuria soongorica (in the uppermost 0-70 cm of the soil). Reduced major axis analysis showed that the slopes of the relationship between above- and belowground biomass of Ceratoides lateens, Salsola passerine and Reaumuria soongorica close to 1, which support isometric scaling hypothesis. However, the slope of the relationship between above- and belowground biomass of Kalidium foliatum was significantly different from 1.0, which don support this hypothesis. There is a significant linear relationship between above- and belowground biomass after logarithmic transformation. The relationship can be used to estimate belowground biomass effectively.
DOIURL [本文引用: 2]
Black locust (Robinia pseudoacacia L.) and Chinese pine (Pinus tabulaeformis Carr.) are two woody plants that are widely planted on the Loess Plateau for controlling soil erosion and land desertification. In this study, we conducted an excavation experiment in 2008 to investigate the overall vertical root distribution characteristics of black locust and Chinese pine. We also performed triaxial compression tests to evaluate the root cohesion (additional soil cohesion increased by roots) of black locust. Two types of root distribution, namely, vertical root (VR) and horizontal root (HR), were used as samples and tested under four soil water content (SWC) conditions (12.7%, 15.0%, 18.0% and 20.0%, respectively). Results showed that the root lengths of the two species were mainly concentrated in the root diameter of 5–20 mm. A comparison of root distribution between the two species indicated that the root length of black locust was significantly greater than that of Chinese pine in nearly all root diameters, although the black locust used in the comparison was 10 years younger than the Chinese pine. Root biomass was also significantly greater in black locust than in Chinese pine, particularly in the root diameters of 3–5 and 5–10 mm. These two species were both found to be deep-rooted. The triaxial compression tests showed that root cohesion was greater in the VR samples than in the HR samples. SWC was negatively related to both soil shear strength and root cohesion. These results could provide useful information on the architectural characteristics of woody root system and expand the knowledge on shallow slope stabilization and soil erosion control by plant roots on the Loess Plateau.
DOIURL [本文引用: 1]
Little attention has been paid to root distribution in combined perennial legume and annual cereal crops. The main objectives of this study were to explore the regularity of root distribution and the influence on nitrogen (N) acquisition in different alfalfa (Medicago sativa L.) and corn (Zea mays L.) intercropping patterns. A three year (2007-2009) field experiment studied four intercropping patterns of alternating alfalfa and corn rows with alfalfa:corn rows sown at 2:2, 3:2, 4:2, and 5:2. Sole crops of corn and alfalfa were used as controls. Roots were sampled over 3 consecutive years by auger sampling method in the prime filling stage of corn from different soil depth, and the root length density (RLD) was used to describe the root spatial distribution of intercropped alfalfa and corn. The results showed that the alfalfa/corn intercropping system had the greater RLD values compared to the sole cropped alfalfa or corn. In addition, the root mass centre of intercropped alfalfa descended into deeper soil layers with advancing stand age, and likewise proliferated laterally towards associated corn rows. Whereas the corn root mass spread at relative shallow soil profile, and also more deployed laterally to neighboring alfalfa root zone. The gap between alfalfa and corn rows was the highest colonized area with greater intermingling and the highest RLD of both species root. The complementary and compatibility of the root spatial distribution of component crops in alfalfa/corn community were the essential cause for better biomass yield.
DOIURL
DOIURL [本文引用: 1]
植物间的相互作用对种群动态和群落结构有着重要的影响。大量的野外实验已经揭示了正相互作用(互利)在群落中的普遍存在及其重要性。为了弥补野外实验方法的不足,模型方法被越来越多地应用于正相互作用及其生态学效应的研究中。该文基于个体模型研究,探讨了植物间正相互作用对种群动态和群落结构的影响。介绍了植物间正相互作用的定义和发生机制、植物间相互作用与环境梯度的关系。正相互作用是指发生在相邻的植物个体之间,至少对其中一个个体有益的相互作用。植物通过直接(生境改善或资源富集)或间接(协同防御等)作用使局部环境有利于邻体而发生正相互作用。胁迫梯度假说认为互利的强度或重要性随着环境胁迫度的增加而增加,但是越来越多的经验研究认为胁迫梯度假说需要改进。以网格模型和影响域模型为例,介绍了基于个体的植物间相互作用模型方法。基于个体模型,对近年来国内外正相互作用对种群时间动态(如生物量-密度关系)、空间分布格局和群落结构(如群落生物量-物种丰富度关系)影响的研究进行了总结。指出未来的研究应集中在对正相互作用概念和机制的理解,新的模型,新的种群、群落,甚至生态系统问题,以及在全球变化背景下进行相关的研究。
DOIURL [本文引用: 1]
植物间的相互作用对种群动态和群落结构有着重要的影响。大量的野外实验已经揭示了正相互作用(互利)在群落中的普遍存在及其重要性。为了弥补野外实验方法的不足,模型方法被越来越多地应用于正相互作用及其生态学效应的研究中。该文基于个体模型研究,探讨了植物间正相互作用对种群动态和群落结构的影响。介绍了植物间正相互作用的定义和发生机制、植物间相互作用与环境梯度的关系。正相互作用是指发生在相邻的植物个体之间,至少对其中一个个体有益的相互作用。植物通过直接(生境改善或资源富集)或间接(协同防御等)作用使局部环境有利于邻体而发生正相互作用。胁迫梯度假说认为互利的强度或重要性随着环境胁迫度的增加而增加,但是越来越多的经验研究认为胁迫梯度假说需要改进。以网格模型和影响域模型为例,介绍了基于个体的植物间相互作用模型方法。基于个体模型,对近年来国内外正相互作用对种群时间动态(如生物量-密度关系)、空间分布格局和群落结构(如群落生物量-物种丰富度关系)影响的研究进行了总结。指出未来的研究应集中在对正相互作用概念和机制的理解,新的模型,新的种群、群落,甚至生态系统问题,以及在全球变化背景下进行相关的研究。
[本文引用: 1]
[本文引用: 1]
喜旱莲子草和接骨草竞争对模拟增温的响应
1
2015
... 一般认为, 植物可通过根系形态结构的变化实现对资源的竞争(
喜旱莲子草和接骨草竞争对模拟增温的响应
1
2015
... 一般认为, 植物可通过根系形态结构的变化实现对资源的竞争(
Aboveground and belowground interactions in mixed plantations of Eucalyptus globules and Acacia mearnsii.
1
2000
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
Positive interactions in communities.
1
1994
... 植物间的互作类型及互作强度与其所处的环境条件之间存在着密切的关联.胁迫梯度假说认为, 当环境条件较好时, 物种间的负相互作用占据主导地位; 而在恶劣的环境条件下, 正相互作用的重要性上升, 并且其作用强度随着环境的恶劣程度而增加(
Fine root morphological and functional traits in Fagus sylvatica and Fraxinus excelsior saplings as dependent on species, root order and competition.
2013
Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.).
1
2006
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
The multifunctionality of agriculture.
2
2010
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Positive Interactions and Interdependence in Plant Communities. Springer-Verlag, Dordrecht
1
2007
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
Positive interactions among alpine plants increase with stress.
2002
西北干旱区灌草型白刺——芨芨草群落根系分布与互作
2
2017
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
... 为了描述根系的垂直分布规律, Gale和Grigal (1987)通过不同演替阶段不同树种根系分布特征提出了植物根系垂直分布非线性模型, 该模型为研究土壤垂直梯度中植物根系的总体分布提供了简洁标准的办法(
西北干旱区灌草型白刺——芨芨草群落根系分布与互作
2
2017
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
... 为了描述根系的垂直分布规律, Gale和Grigal (1987)通过不同演替阶段不同树种根系分布特征提出了植物根系垂直分布非线性模型, 该模型为研究土壤垂直梯度中植物根系的总体分布提供了简洁标准的办法(
根系间的相互作用——竞争与互利
2
2004
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
根系间的相互作用——竞争与互利
2
2004
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Mechanisms of maintenance of species diversity.
1
2000
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
Ecology goes underground.
1
2000
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
Rooting strategy of naturally regenerated beech in silver birch and scots pine woodlands.
2
2003
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
Role of species identity in plant invasions: Experimental test using Imperata cylindrical.
1
2009
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Depth distribution of cherry (Prunus avium L.) tree roots as influenced by grass root competition.
1
2001
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Selection for niche differentiation in plant communities increases biodiversity effects.
1
2014
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
Niche differences in phenology and rooting depth promote coexistence with a dominant C4 bunchgrass.
1
2005
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Coexistence of a C4 grass and a leaf succulent shrub in an arid ecosystem. The relationship between rooting depth, water and nitrogen.
2
2011
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Fine root biomass and morphology of Pinus densiflora under competitive stress by Chamaecyparis obtusa.
1
2008
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
Vertical root distributions of northern tree species in relation to successional status.
1
1987
... 利用
浙江天童亚热带常绿阔叶林土壤呼吸空间异质性研究
1
2014
... 境不同而造成的实验误差.所选植物周围1 m的范围内无其他植物生长, 避免来自周围植物的种间或种内作用的影响.采用分层取样法(
浙江天童亚热带常绿阔叶林土壤呼吸空间异质性研究
1
2014
... 境不同而造成的实验误差.所选植物周围1 m的范围内无其他植物生长, 避免来自周围植物的种间或种内作用的影响.采用分层取样法(
不同生长期干旱胁迫对刺槐幼树干物质分配的影响
1
2010
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
不同生长期干旱胁迫对刺槐幼树干物质分配的影响
1
2010
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
Distribution of roots and root length density in a maize/soybean strip intercropping system.
2010
Plant Strategies and Vegetation Processes. Wiley, Chichester
1
1979
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
Root order- and root age-dependent response of two poplar species to belowground competition.
2
2014
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Extreme stresses, niches, and positive species interactions along stress gradients.
2014
Testing the importance of plant strategies on facilitation using congeners in a coastal community.
2012
The plastic plant: Root responses to heterogeneous supplies of nutrients.
1
2004
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
Rooting depth and above- ground community composition in Kalahari sand woodlands in western Zimbabwe.
2
2008
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Root competition between beech and oak: A hypothesis.
1
2001
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
渭北黄土区刺槐根系空间分布特征研究
2005
渭北黄土区刺槐根系空间分布特征研究
2005
疏叶骆驼刺根系对土壤异质性和种间竞争的响应
1
2012
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
疏叶骆驼刺根系对土壤异质性和种间竞争的响应
1
2012
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
多枝柽柳幼苗根系形态及生物量对不同灌溉处理的响应
1
2012
... 境不同而造成的实验误差.所选植物周围1 m的范围内无其他植物生长, 避免来自周围植物的种间或种内作用的影响.采用分层取样法(
多枝柽柳幼苗根系形态及生物量对不同灌溉处理的响应
1
2012
... 境不同而造成的实验误差.所选植物周围1 m的范围内无其他植物生长, 避免来自周围植物的种间或种内作用的影响.采用分层取样法(
Multi-proxy evidence for competition between savanna woody species. Perspectives in Plant Ecology,
1
2007
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
Contrasting root behaviour in two grass species: A test of functionality in dynamic heterogeneous conditions.
1
2011
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Vegetation changes in a semi-arid, grassy dwarf shrub land in the Karoo, South Africa: Influence of rainfall variability and grazing by sheep.
1995
Rooting depth and soil moisture control Mediterranean woody seedling survival during drought.
2007
Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems.
2002
Should I stay or should I go? Roots segregate in response to competition intensity.
1
2015
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
红砂根系构型对干旱的生态适应
1
2012
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
红砂根系构型对干旱的生态适应
1
2012
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
Associated growth of C3 and C4 desert plants helps the C3 species at the cost of the C4 species.
4
2012
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
... 红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是干旱荒漠和荒漠草原区分布最广的植物种, 两者常互为共建种组成混生群落, 混生群落适应极端环境的能力要高于单生群落, 这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用(
... ).在自然分布的混生群落中, 有时红砂和珍珠猪毛菜各自单独生长, 称之为单生, 有时它们紧密生长在一起, 称之为混生(
... 境不同而造成的实验误差.所选植物周围1 m的范围内无其他植物生长, 避免来自周围植物的种间或种内作用的影响.采用分层取样法(
1
1988
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
我国西北地区芨芨草-苦豆子群落根系分布与种间关系
1
2009
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
我国西北地区芨芨草-苦豆子群落根系分布与种间关系
1
2009
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Grasland: Savanne und Busch der arideren Teile Afrikas in ihrer ?kologischen Bedingtheit.
1939
植物根系养分捕获塑性与根竞争
1
2012
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
植物根系养分捕获塑性与根竞争
1
2012
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
水曲柳苗木地下竞争与地上竞争的定量研究
1
2003
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
水曲柳苗木地下竞争与地上竞争的定量研究
1
2003
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
水曲柳落叶松根系之间的相互作用研究
1
2000
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
水曲柳落叶松根系之间的相互作用研究
1
2000
... 植物根系之间的相互作用是一种复杂的生态学过程, 在此过程中, 为了适应竞争环境, 提高竞争效率, 吸收较多的养分和水分, 根系表现出明显的可塑性(
3种荒漠灌木的用水策略及相关的叶片生理表现
1
2005
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
3种荒漠灌木的用水策略及相关的叶片生理表现
1
2005
... 作为根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值越大, 表明投入相同的成本有较高的养分利用效率(
Spatial pattern and spatial association of Quercus acutissima at different developmental stages in the Qiaoshan Mountains.
1
2017
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
干旱程度对C3植物红砂和C4植物珍珠光合生理参数的影响
1
2012
... 红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是干旱荒漠和荒漠草原区分布最广的植物种, 两者常互为共建种组成混生群落, 混生群落适应极端环境的能力要高于单生群落, 这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用(
干旱程度对C3植物红砂和C4植物珍珠光合生理参数的影响
1
2012
... 红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是干旱荒漠和荒漠草原区分布最广的植物种, 两者常互为共建种组成混生群落, 混生群落适应极端环境的能力要高于单生群落, 这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用(
荒漠草地4种灌木生物量分配特征
2013
荒漠草地4种灌木生物量分配特征
2013
Vertical root distribution and root cohesion of typical tree species on the Loess Plateau, China.
2
2014a
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
... 为了描述根系的垂直分布规律, Gale和Grigal (1987)通过不同演替阶段不同树种根系分布特征提出了植物根系垂直分布非线性模型, 该模型为研究土壤垂直梯度中植物根系的总体分布提供了简洁标准的办法(
Root distribution and N acquisition in an Alfalfa and corn intercropping system.
1
2013
... 根系之间的相互作用包括竞争和互利, 它们可以同时对植物生长产生影响(
Response of root traits of Reaumuria soongarica and Salsola passerina to facilitation.
2014b
植物间正相互作用对种群动态和群落结构的影响: 基于个体模型的研究进展
1
2013
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
植物间正相互作用对种群动态和群落结构的影响: 基于个体模型的研究进展
1
2013
... 物种共存和多样性形成机制一直是生态学研究的核心问题(
红砂和珍珠对极端环境的生理响应与调节机制
1
2012
... 红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是干旱荒漠和荒漠草原区分布最广的植物种, 两者常互为共建种组成混生群落, 混生群落适应极端环境的能力要高于单生群落, 这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用(
红砂和珍珠对极端环境的生理响应与调节机制
1
2012
... 红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是干旱荒漠和荒漠草原区分布最广的植物种, 两者常互为共建种组成混生群落, 混生群落适应极端环境的能力要高于单生群落, 这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}